Embed Size (px)
Citation preview

J. Physiol. (1971), 215, pp. 119-137 119With 12 text-figuresPrinted in Great Britain
CONTRACTIONS OF THE CAUDAL REGION OF THEOESOPHAGUS OF THE CAT
By D. A. TITCHEN* AND J. S. WHEELERFrom the Department of Physiology and Anatomy,
Massey University, and Plant Chemistry Division, D.S.I.R.,Palmerston North, New Zealand
(Received 15 December 1970)
SUMMARY
1. The motility of the most caudal part of the oesophagus has beeninvestigated in cats which were decerebrated, had the whole of the centralnervous system pithed or were anaesthetized with chloralose.
2. Contractions of the most caudal region of the oesophagus wereevoked by its stretch and the presence in it of HC1. These reactions wereobtained in all types of preparations used.
3. In decerebrate preparations oesophageal contractions occurred inresponse to the introduction of acid into the stomach. These were notobtained after the vagus nerves had been cut.
4. Slight degrees of gastric distension stimulated oesophageal con-tractions, greater distension was inhibitory.
5. Both reductions in oesophageal activity and contractions of theoesophagus were obtained with different parameters of stimulation ofefferent vagal fibres.
6. Excitatory and inhibitory effects on the oesophagus were obtainedfrom stimulation of the central end of the vagus nerves cut in the abdomenor in the neck.
7. In totally pithed preparations of cats stretch of one region of theoesophagus was found to modify the activity of a distant region of theoesophagus cranial or caudal to the site of distension.
8. The reactions of the caudal oesophagus were abolished by atropine.9. The importance of vago-vagal reflexes in the modification of basically
direct oesophageal responses has been discussed.* Present address: Department of Veterinary Preclinical Sciences, University of
Melbourne, Parkville, 3052, Australia.

D. A. TITCHEN AND J. S. WHEELER
INTRODUCTION
Contractions of the oesophagus may be elicited by distension of theoesophagus itself and although it has long since been suggested that suchoesophageal responses contribute to the pattern of motility of the oeso-phagus during swallowing (see Meltzer, 1899) many aspects of the mechan-isms involved in the control of oesophageal motility remain obscure(Christensen & Daniel, 1966). The work reported here was concerned withthe reactions of the oesophagus of the cat, and particularly of its mostcaudal part, to different stimuli. This region of the cat's oesophagus hasbeen shown to function as a sphincter (Clark & Vane, 1961; Schenk &Fredrickson, 1961), due to the activity of the circular layer of the smoothmuscle in this most caudal 2 cm of the oesophagus (Clark & Vane, 1961).A number of studies have provided evidence on afferent stimuli which
are effective when delivered to the oesophagus and which stimulateoesophageal and other reactions. Studies on impulses recorded in afferentvagal nerve fibres during distension of the oesophagus have provided anindication of the presence of stretch sensitive receptors in the oesophagus(Andrew, 1957; Iggo, 1957a; Mei, 1965). A concentration of such receptorsin both more caudal and more cranial parts of the thoracic oesophagus hasbeen suggested from one of these experimental studies in the cat (Mei,1965). Judged by the salivary and gastric responses which can be evokedmore readily by distension of these same regions in the sheep than fromother parts of the oesophagus there may be, in this species as well as thecat, a concentration of receptors in the most caudal part of the thoracicoesophagus and also at about the level of the first rib (Sellers & Titchen,1959). A particular sensitivity of the most caudal regions of the thoracicoesophagus to tactile stimulation has been detected in ruminants (Clark& Weiss, 1952; Comline & Kay, 1955): these reports were concerned withthe reflex stimulation of salivary secretion from the oesophagus. In thecat and man infusion of acid into the most caudal segment of the oeso-phagus may evoke contractions of the region (Cannon, 1908; Clark & Vane,1961; Giles, Humphries, Mason & Clark, 1969) and in man leads to thesensation commonly called heartburn (Siegel & Hendrix, 1960).The experiments described below were concerned with an examination
of the responses of the most caudal region of the oesophagus to stretch ofitself, the introduction of acid into its lumen, gastric acid and stretchstimulation and stimulation of the vagus nerves in an afferent and efferentsense.
120

OESOPHAGEAL CONTRACTIONS 121
METHODS
Young and adult cats ofboth sexes weighing between 1-5 and 5 kg were used. Theywere fasted for 24 hr before an experiment, during which time they had wateravailable. Anaesthesia was induced with ethyl chloride and maintained with ether.Intravenous injections were made via femoral venous cannulae, blood pressure wasrecorded from a femoral or carotid artery. A tracheal cannula or endotracheal tubewas always inserted. Decerebrate preparations were made following the method usedby Comline & Titchen (1951). Chloralose (70 mg/kg) was injected intravenouslyafter being dissolved in saline at 700 C or in di-methyl sulphoxide (British DrugHouses); the injections were made in cats whilst anaesthetized with ether.Both in decerebrate preparations in which the spinal cord caudal to the 2nd
or 3rd cervical segment was pithed and in totally pithed preparations the pithingprocedure was similar to that described by Iorio & McIsaac (1965). Pithing was bythe introduction into the vertebral canal of a length of rubber tube attached to aplastic covered copper wire which when passed through the canal pushed the spinalcord before it, and on the release of tension completely filled the vertebral canal.Care taken to maintain continued positive pressure in the lungs (even during expira-tion) with artificial ventilation ensured that there was slight continual hemorrhagefrom the laminectomy wounds during the actual pithing: in this way air or fatembolism was avoided during pithing.A glass cannula 7 mm internal diameter tied in position by a purse string suture
placed through the mucosa at the region of the gastro-oesophageal mucosal junctionprovided for drainage of fluid from the oesophagus and allowed recording from themost caudal regions of the oesophagus without tubes leading through more cranialparts of the oesophagus. Fluid drained from the oesophagus through a latex tubeattached to the shaft of the cannula and passing through the stomach and out of itvia the gastrostomy. Isolation of the stomach was completed when a ligature wastied around the most cranial part of the duodenum. A glass cannula 12 mm externaldiameter, tied into the gastrostomy incision in the greater curvature,allowed changesto be made in conditions in the stomach. These arrangements are shown diagram-matically in Fig. 1.The balloon used to record the activity of the most caudal part of the oesophagus
was placed immediately craniad to the flange of the cannula tied into the gastro-oesophageal mucosal junction. Balloons used to record from more cranial regions ofthe oesophagus were introduced via the mouth or through incisions made in themid-cervical region of the oesophagus. Balloons placed in the thoracic regions of theoesophagus inflated when the tubes to which they were connected were opened tothe atmosphere. The volume of distension of the balloons (estimated from thedeflation possible with a syringe) was up to 0-5 ml. That it was not greater wasprobably due to tonic contractions of the oesophagus.The experiments were undertaken with two recording systems. Earlier experi-
ments involved the use of simple kymographic recording techniques, a stethograph-tambour system for recording respiration, a mercury manometer for recording bloodpressure and a water-filled balloon reservoir system associated with an air-filledtambour to provide a 'displacement' system to record oesophageal contractions, amethod akin to that described by Setekliev (1964a) and used to obtain recordingsof uterine contractions.In some experiments use was made of hot stylus recorders (Sanborn and Devices)
in conjunction with Statham and Elema Schonander transducers, and in theseeither the isometric or isotonic recording systems described by Setekliev (1964a)were used.

D. A. TITCHEN AND J. S. WHEELERAtropine sulphate (British Drug Houses) was injected intravenously as a 1 mg/
ml. solution in 0 9% (w/v) saline.The balloons used were made from latex finger stalls (London Rubber Co.) and
constructed so that they were between 12 and 17 mm long and 10-20 mm diameterat 20 cm H20 pressure. The balloons were attached to vinyl tubing (Portex Londonsize 7) about 4 9 mm external and 3-5 internal diameter.A Palmer (London) student's stimulator was used in conjunction with fluid
electrodes of the type described by Porter & Allamon (1936), for nerve stimulation,in earlier experiments. Later a Grass model S4E stimulator was used.
L <S
Fig. 1. Diagram of arrangements for isolation of the stomach and record-ing from the caudal oesophagus. A purse string suture, S, placed at thegastro-oesophageal mucosal junction was used to tie a cannula, C, into theoesophagus. A ligature, L, tied round the most cranial part of the duo-denum isolated the stomach caudally. The tube, I, was used to makeinfusions into the oesophagus; the balloon B was used to distend, andrecord from, the oesophagus. The tube, T, ensured isolation from thestomach of the cannulated oesophagus by being passed through a gastriccannula- G. A loose sleeve, SI, reduced the possibility of interference withthe oesophageal recording balloon when the stomach was distended.Another gastric cannula, GC, was in some experiments also tied in thegreater curvature and used to alter conditions in the stomach indepen-dently of those in the oesophagus.
RESULTS
Contractions of the most caudal part of the oesophagus were recordedwithin 10 min of decerebration, pithing and the injection of chloralose andwith distension of the oesophagus limited to that resulting from the balloonbeing open to the atmosphere before or during its connexion to the record-ing apparatus. Contractions of the oesophagus recorded under these circum-stances declined over periods of up to 1 hr after decerebration or the otherprocedures had been completed. Recurring contractions of the oesophagus
122

OESOPHAGEAL CONTRACTIONS 123
were detected when the Dieffenbach arterial (bulldog) clamps, placed onthe common carotid arteries before decerebration, were still in place: theydid not appear to be changed by the gross fall in blood pressure whichfollowed removal of these clamps. In both anaesthetized and decerebratepreparations, deeper inspiratory movements were frequently succeeded byone or a series of contractions of the oesophagus. It was found that greaterrespiratory efforts stimulated when the tracheal cannula was temporarily
Occ Occ Occ Occ- - - Sig
°° 30 -
E_A___i_ Caud oes
W.tJ~it ! X vbt~t~tWW!4t$ If ~tri t t t tortstrW.'~rt Resp
W 120 B.B.P.E 60 LE 0
a bFig. 2. Decerebrate cat. Oesophageal contractions following respiratorymovements made during occlusion of the tracheal cannula. Records fromabove downwards: Sig, signal, Caud oes, pressure recorded from a caudaloesophageal balloon distended to 0-6 ml. with an isometric recordingsystem, Sec, 1 sec time marker, Resp, thoracic wall movement, inspira-tion upwards, B.P., blood pressure recorded from the right femoral artery.a, Caudal oesophageal contractions following occlusion of the trachealcannula (Occ) during expiration for one respiration in each case. b, Theeffects of each of two periods of occlusion, during inspiration, of thetracheal cannula (Occ) first for one respiration and later for two res-pirations.
occluded might be succeeded by one or more oesophageal contractions;this is evident in Fig. 2. Accordingly care was taken to demonstrate all ofthe procedures described as modifying oesophageal contractions at least60-90 min after decerebration and when respiration was regular and quietand heart rate and blood pressure were also regular. In other preparationsused experimental observations usually commenced within 20 min of com-pletion of pithing or administration of the anaesthetic agent. These delaysresulted in the oesophageal responses to oesophageal, gastric, or vagalstimuli reported below being observed on a background of decliningspontaneous activity or when such activity had largely ceased.
Distension of the terminal thoracic oesophagus was frequently (but notinvariably) followed by one or a series of contractions of this region of theoesophagus. There were similarities between these responses and those

D. A. TITCHEN AND J. S. WHEELER
obtained by Setekliev (1964b) from the rabbit uterus. There was an initialrapid pressure rise associated with the distension corresponding to thatwhich Setekliev (1964b) attributed to a passive response of the myome-trium to stretch. In the case of the oesophagus this was usually succeededby a contraction starting as rapidly as 1 see but up to 30 sec after thedistension was delivered. The shorter latencies were observed in all of thepreparations used but the longer latencies were only seen in decerebrate oranaesthetized preparations. Up to about 4 ml. greater distension led to agreater response. In Fig. 3 which is a record obtained from a totally pithed
I ml. 3 ml. 1 ml. 3 ml._ - oSig
mm Hg.. Owlg *. AAVj; Thor oes
100 _
mm Hg 50F L@ W
IiA, Caud oes0 ........ . . . . ... . . . . . . . . . . . . . . M in
Fig. 3. Totally pithed preparation of a cat. Effect on the region dis-tended and a cranial region of the oesophagus of two levels of distensionof the caudal oesophagus. Records from above downwards: Sig, signal,Thor oes, mid-thoracic oesophagus, 0 5 ml. balloon distension, Caud oescaudal oesophagus, balloon distending to 0 5 ml. at atmospheric pressure,Min, 1 min time marker. Responses to four periods each of 3 min of dis-tension of the caudal oesophagus. Extent of distension in each successivesegment of the record was as indicated 1 ml., 3 ml., 1 ml. and 3 ml. Thecaudal oesophageal balloon was opened to the atmosphere after each dis-tension was relieved. Isometric recording.
preparation of a cat when the caudal region of the oesophagus was dis-tended with 1 ml. and 3 ml., the responses of the part of the oesophagusdistended to the two degrees of distension show features commonlydetected when different volumes were introduced into the balloon. Themain features to be noted are that with greater volumes of distension theinitial rapid response on inflation of the balloon was to a higher pressure,more contractions were stimulated and these occurred from a higher baseline (of pressure) than with lesser degrees of distension.
Distant excitatory effects of distension on the oesophaqgs were revealed asincreased activity or the initiation of activity in a part of the oesophaguscranial to the region distended (Fig. 3). The first an excitatory effect ofcaudal oesophageal distension on more cranial regions was observed waswithin 10 see after the distension of the caudal region of the oesophagus.
124

OESOPHAGEAL CONTRACTIONSThis excitatory effect on the more cranial region continued for up to 120see after the distension of the caudal oesophagus was withdrawn. No clearassociation between the individual contractions of the two parts of theoesophagus was established either in their rhythm or frequency.
Distant inhibitory effects of distension on the oesophagus were also seen inpithed preparations in which distension of a more cranial region of thethoracic oesophagus was shown to inhibit activity of the caudal part of theoesophagus. In this case also the response of the region distended was inthe form of a series of contractions and a sustained tone. During the periodof distension the amplitude of the recurring contractions of the caudaloesophagus was reduced (Fig. 4).
4 ml.in out
100 l pn Sig
E Sig, scaud oes
bo
isometi Thor oes*Min
Fig. 4. Totally pithed preparation of a cat. Effect of distension of themid-thoracic oesophagus on the caudal oesophagus. Records from abovedownwards: Sig, signal, Caud oes, caudal oesophagus, 1-5 ml. distension,isometric recording. Thor oes, mid thoracic oesophagus, initial distension05 ml., isometric recording, Min, 1 min time marker.The response of the mid-thoracic oesophagus to its distension with 4 ml.
is shown. This also produced a reduction in the height of the contractionsof the caudal oesophagus.
The responses of the oesophagus to stimulation of itself were notlimited to those elicited by its stretch.The infusion of HCt into the caudal thoracic oesophagus stimulated con-
tractions of this region of the oesophagus with latencies between the startof infusions and the responses to them of 15 see or more in both decere-brate and pithed preparations. The reaction in decerebrate preparationscharacteristically consisted of an initial small contraction followed within5 see by a rapidly developing intense oesophageal contraction which wasin turn succeeded by a series of less strong and progressively diminishingcontractions recurring at frequencies up to 6/min (Fig. 5).
125

D. A. TITCHEN AND J. S. WHEELER
In pithed preparations the responses of the caudal oesophagus to in-fusion of acid through it consisted most obviously of an increase in thetone of the oesophagus and superimposed on this was a series of rhythmi-cally recurring oesophageal contractions. As in decerebrate preparationseffects due to the volume of the infusion and its acidity could be distin-guished by comparing responses to the infusion through the oesophagus ofequal volumes of saline and 01 N-HCl, and by choosing infusion ratesbelow threshold for evoking stretch responses (Fig. 6).
Sal HCI Sal
100 Sigg 50 t 02.iS ~~~~~~~~~Caud oes-Min
200_
B.P.E 100 BP
Fig. 5. Decerebrate preparation of a cat. Effects of the introduction ofsaline and of acid into the caudal oesophagus on caudal oesophagealmotility. Records from above downwards: Sig, signal, Caud oes, caudaloesophagus, Min, 1 min time marker, B.P. carotid arterial blood pressure.Sal indicates the infusion of 1-5 ml. 0 9% NaCl into the caudal oesophagus.HC1, indicates the introduction of 1-5 ml. 0-1 N-HCl into the caudaloesophagus. Isotonic recording of the oesophagus with 3 0 ml. in theballoon. The calibration (0, 50, 100%) refers to the extent of emptying ofthe oesophageal balloon with an oesophageal contraction. Completeemptying occurred with an alteration of 1 mm Hg pressure. Recordingpressure 20 mm Hg.
In addition to the differences in the form of the reactions of the oeso-phagus in decerebrate and pithed preparations to the infusion of acidthrough its terminal region there appeared to be a difference in the sensi-tivity of the two types of preparation. Usually a more sensitive set ofrecording conditions was used to obtain records of the responses in pithedpreparations. These differences between pithed and decerebrate prepara-tions suggested the possibility of both direct and reflex oesophagealreactions to exposure of its mucosa to acid. It did not prove possible toimplicate the vagus nerves in any reflex responses of the oesophagus to its
126

OESOPHAGEAL CONTRACTIONS
acidification since in these experiments there~were frequently long-lastingchanges in excitability following their section which made it difficult toestablish comparable conditions before and after they were cut.
Stimulation of the vagus nerves in an efferent sense had both excitatoryand inhibitory effects on the caudal thoracic region of the oesophagus. Theeffects were examined in pithed, anaesthetized and decerebrate prepara-tions which had both vagus nerves cut in the neck.
Excitatory effects on the caudal thoracic oesophagus were seen as acontraction which commenced within 2 see of the application of thestimulus, reached a peak within 5 see and in which relaxation was not
Sal HCI Sal Sal10 ~~~~~~~~~~~~~~~Sig10[
E _£'
~ w~_Caudoes
IHi pqili 1w!;, rNs.I m Q 'I IINHIP ' Resp
100 -
so50 B.P.
Fig. 6. Totally pithed preparation of a cat. Stimulation of oesophagealcontractions by infusing 10 ml. 0.1 N-HCR through the caudal oesophagus.Records from above downwards: Sig, signal, Caud oes, caudal thoracicoesophagus, 0-6 ml. in balloon, isometric recording system, Resp, thoracicmovements of respiration, inspiration upwards. Min, 1 min time marker,B.P. Carotid arterial blood pressure. Successive infusions through the caudaloesophagus of 10 ml. saline (Sal) and 10 ml. 0-1 N-HC1 were indicated onthe record by the interruption of the signal. Note the sensitivity of oeso-phageal recording used.
complete until up to 8 see after stimulation of the vagus nerve had ceased.The reactions of the caudal, smooth muscle part of the oesophagus con-trasted markedly with those of the more cranial, striated muscle, regionsof the thoracic oesophagus. This was most obvious with low frequenciesof vagus nerve stimulation. A smooth contraction curve was obtained inthe case of the most caudal region ofthe thoracic oesophagus. The responseof more cranial parts of the oesophagus to low frequency stimulation of thevagus in an efferent sense consisted of individual contraction peaks corre-sponding to each of the individual stimuli applied. With the balloons usedin these experiments a region of the thoracic oesophagus caudal to theheart was characterized as one in which both slow (smooth muscle) and
5 PHY 2I5
127

D. A. TITCHEN AND J. S. WHEELER
fast (striated muscle) responses were obtained (Fig. 7). Bremner, Shorter &Ellis (1970) have shown that in thoracic regions of the cat's oesophagusthere may be a mixture of smooth and striated muscle over a distance of0*5-2 cm. In the case of the striated muscle of the oesophagus responses toefferent vagus stimulation were recorded within 0-2 see of the start ofstimulation, and tetanus of the cranial thoracic oesophagus was producedby efferent vagus nerve stimulation at 15 stimuli/sec as judged by recordsobtained from oesophageal balloons. The response of the striated muscle of
VAG
100 '' ^ Sig3:i
E50[ i\ Caud oes
0~~~~~~~~~~~~~~~~~~
E Thdr oes
0 .............. ... Sec
E LsNSNmStsS~S-0x\~N'E~EEysNS'N\\[email protected].
Fig. 7. Totally pithed preparation of a cat. Responses of the caudaloesophagus and the mid-thoracic oesophagus to stimulation of the peri-pheral end of the left vagus nerve cut in the mid-cervical region. Recordsfrom above downwards: Sig, signal, Caud oes, caudal oesophagus, Thoroes, mid-thoracic oesophagus. Sec, 1 see time marker. B.P., carotid arterialblood pressure. VAG, marks stimulation for 2 see of the peripheral end ofthe left vagus cut in jthe neck, frequency 5/sec. The mid-thoracic oeso-phageal record has been retouched to show more clearly the skeletalmuscle component of its response, ten rapid contractions which occurredduring the development of the smooth muscle contraction of this regionof the oesophagus. Isometric contraction from both regions of the oeso-phagus with 0-5 ml. in the balloons.
the oesophagus to efferent vagal nerve stimulation persisted after theadministration of atropine (in doses up to 1-0 mg/kg). In all types of pre-paration used atropine suppressed the caudal oesophageal contractions inresponse to efferent vagal stimulation, to stretch of itself and reduced therhythmic recurring contractions of the oesophagus (Fig. 8).An inhibitory effect on the caudal oesophagus of stimulation ofthe vagus
nerve was seen in some experiments with lower frequencies of stimulation.This inhibition was apparent within 5 see of the start of stimulation. It
128

OESOPHAGEAL CONTRACTIONSwas evident as a disappearance of the rhythmically recurring contractionsof the oesophagus and of the tone of the oesophagus. The duration of theinhibitory effect of efferent vagus nerve stimulation closely correspondedto the periods of stimulation of the vagus nerve (Fig. 8). On withdrawal ofa stimulus of the vagus nerve which was inhibitory, increases in the forceof one or more contractions of the caudal segment of the oesophagus werefrequently recorded.
Neither the excitatory nor the inhibitory effects of vagus nerve stimu-lation on the terminal thoracic oesophagus were demonstrated after theadministration of atropine (041-0-2 mg/kg) which led to a diminution or
VAG ATR VAG- - Sig
% so Caud oes
0
V 'JJJI_ _ _ _ _ WINK RespMin
mm Hg [ ~WB.P.
Fig. 8. Pithed preparation of cat. Inhibitory effects of vagus nervestimulation, and blocking by atropine, of the rhythmically recurring con-tractions of the caudal oesophagus. Records from above downwards: Sig,signal, Caud oes, caudal thoracic oesophagus, isotonic recording system,3 ml. in balloon, calibration as in Fig. 5. Resp, thoracic movements, in-spiration upwards, Min, 1 min time marker, B.P. carotid arterial blood pres-sure. VAG, stimulation of peripheral end of right vagus cut in the neck at5/sec, 12-5 V, 1-5 msec pulse duration for 15 sec. ATR, intravenous in-jection of atropine, 0-2 nmg/kg.
disappearance in the tone of the oesophagus. It was thus no longer possibleto demonstrate an inhibitory effect of vagal stimulation after atropine wasgiven.
Reflex responses of the oesophagus. Both direct and indirect indicationsof the oesophagus responding as part of a reflex reaction to stimuli wereobtained. Indirect evidence of the reflex modification of terminal thoracicoesophageal motility was obtained in experiments in which the effects ofgastric stimulation on the motility ofthe caudal oesophagus were examined.The experimental arrangements described in the section on Methods andshown diagrammatically in Fig. 1 facilitated the production of alterations
5-2
129

D. A. TITCHEN AND J. S. WHEELER
in the stomach without those changes directly affecting the caudal regionof the oesophagus.The activity of the caudal segment of the thoracic oesophagus was
inhibited by handling of the gastric cannula: repeated manipulation of itled to vomiting and a loss in the excitability or disappearance of contrac-tions of the caudal oesophagus. These reactions limited the nature of pro-cedures which could be performed in attempts to analyse possible modi-fication of oesophageal motility by changes in conditions in the stomach.
Sal Sal Sal-
100 ------- -Sig
0 - - > _ _ 0 ' Caud oes
100 - Min
E 50 L-B.P.TEEFig. 9. Decerebrate preparation of cat. The greater effects on caudaloesophageal contractions of successive periods of the introduction of 20 ml.saline into the stomach.
Records from above downwards: Sig, signal, Caud oes, caudal oesophagus,isotonic recording system 3 ml. in balloon scale as in Fig. 5. Resp, thoracicwall movements, inspiration upwards, Min, 1 min time marker, B.P.femoral arterial blood pressure. Sal, 20 ml. saline was infused into thestomach and 15 sec later allowed to start flowing out.
Distension of the stomach with 50-100 ml. saline stimulated vomiting insome decerebrate preparations, and in others, in which it did not, theselevels of gastric distension inhibited caudal thoracic oesophageal activity.In contrast the introduction into the stomach of lesser volumes of saline(20-30 ml.) initiated caudal oesophageal activity or increased pre-existingcontractions (Fig. 9).
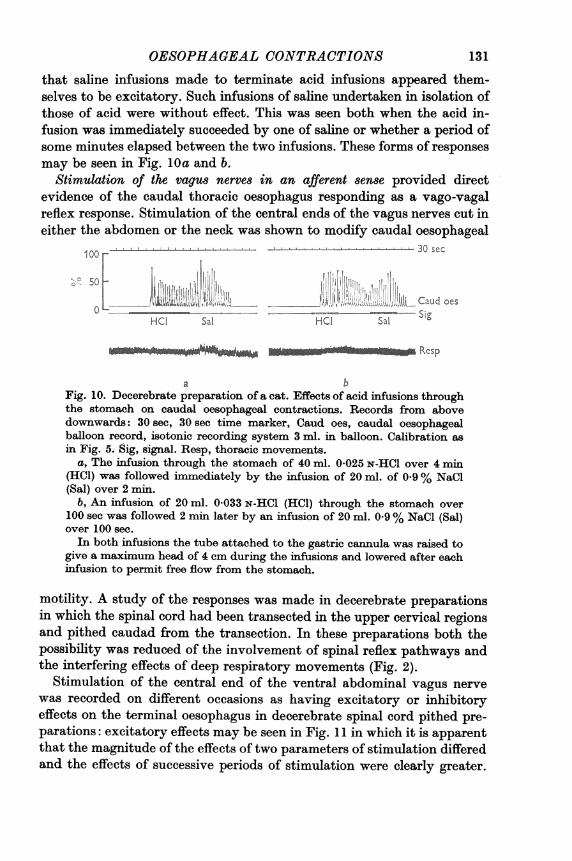
Infusions of acid into the stomach started contractions of the caudalthoracic oesophagus after latencies of 30-90 sec in decerebrate prepara-tions (Fig. 10) when distension of the stomach to a pressure greater than4-6 cm of water was avoided during the infusions. If pressures greaterthan this developed excitatory effects of acid infusions were delayed untilthe pressures were relieved. A feature of the responses to infusions of acidthrough the stomach noted in a number of decerebrate preparations was
130
OESOPHAGEAL CONTRACTIONS
that saline infusions made to terminate acid infusions appeared them-selves to be excitatory. Such infusions of saline undertaken in isolation ofthose of acid were without effect. This was seen both when the acid in-fusion was immediately succeeded by one of saline or whether a period ofsome minutes elapsed between the two infusions. These forms of responsesmay be seen in Fig. 10a and b.
Stimulation of the vagus nerves in an afferent sense provided directevidence of the caudal thoracic oesophagus responding as a vago-vagalreflex response. Stimulation of the central ends of the vagus nerves cut ineither the abdomen or the neck was shown to modify caudal oesophageal
00 30sec
(Se2I¢JI~i IIIIRS>IIIIZ~DF!IID2IIIHCaud oesHCI Sal HCI Sal Sig
Resp
a bFig. 10. Decerebrate preparation of a cat. Effects of acid infusions throughthe stomach on caudal oesophageal contractions. Records from abovedownwards: 30 sec, 30 sec time marker, Caud oes, caudal oesophagealballoon record, isotonic recording system 3 ml. in balloon. Calibration asin Fig. 5. Sig, signal. Resp, thoracic movements.
a, The infusion through the stomach of 40 ml. 0.025 N-HCl over 4 min(HC1) was followed immediately by the infusion of 20 ml. of 0 9% NaCl(Sal) over 2 min.
b, An infusion of 20 ml. 0*033 N-HCl (HCI) through the stomach over100 sec was followed 2 min later by an infusion of 20 ml. 0 9% NaCl (Sal)over 100 sec.In both infusions the tube attached to the gastric cannula was raised to
give a maximum head of 4 cm during the infusions and lowered after eachinfusion to permit free flow from the stomach.
motility. A study of the responses was made in decerebrate preparationsin which the spinal cord had been transacted in the upper cervical regionsand pithed caudad from the transaction. In these preparations both thepossibility was reduced of the involvement of spinal reflex pathways andthe interfering effects of deep respiratory movements (Fig. 2).
Stimulation of the central end of the ventral abdominal vagus nervewas recorded on different occasions as having excitatory or inhibitoryeffects on the terminal oesophagus in decerebrate spinal cord pithed pre-parations: excitatory effects may be seen in Fig. 11 in which it is apparentthat the magnitude of the effects of two parameters of stimulation differedand the effects of successive periods of stimulation were clearly greater.
131

D. A. TITCHEN AND J. S. WHEELERThe latencies in all these responses were about the same, being 7-8 secwith both the initial stimuli and the later stimuli, to which the responseswere greater. Other indications of lasting changes in excitability werefound in experiments in which the first of two or more periods of afferentvagus nerve stimulation was without overt effect on the caudal thoracicoesophagus, but in which a second similar vagus nerve stimulation causeda strong oesophageal response. None of the excitatory effects on the caudalthoracic oesophagus of afferent vagus nerve stimulation persisted after
A B A B
100 ---Sig
so0 Caud oes
Thor
100 - -----y --Min.
Iso ------- B.P.
Fig. 11. Decerebrate spinal cord pithed preparation of a cat. The effecton caudal oesophageal contractions of stimulation of the central end of theventral abdominal vagus nerve trunk. Records from above downwards:Sig, signal, Caud oes, caudal thoracic oesophagus, isotonic recording system3 ml. in balloon. Calibration as in Fig. 5. Thor, thoracic wall movements,inspiration upwards, Min, 1 min time marker, B.P. femoral arterial bloodpressure. Effects of four successive periods each of 10 see of ventral vagusnerve trunk stimulation are shown: in each of those marked A the stimu-lator settings were for 10 pulses/sec, 5 V, 1-5 msec pulse duration: in theperiods marked B the parameters were 10 pulses/sec, 5 V, but pulseduration was altered to 0-5 msec. Note the similarity of the summationeffect to that shown in Fig. 9.
both vagus nerves had been cut in the neck or after atropine had beenadministered in doses of 0-1-0-2 mg/kg.
Inhibitory effects of afferent vaqus nerve stimulation on the caudalthoracic oesophagus were recorded. These were most obvious when stimu-lation of the vagus nerve led to vomiting. In this case there was a markedreduction in the tone of the terminal thoracic oesophagus (Fig. 12). Thisinhibitory effect was also observed to be succeeded by an increase in theintensity of the contractions of the oesophagus. These inhibitory reactionswere observed in decerebrate spinal cord pithed preparations in whichmuch of the somatic activity of vomiting was eliminated and also in suchpreparations and simple decerebrate preparations when afferent vagusnerve stimulation did not produce vomiting.
132

OESOPHAGEAL CONTRACTIONS 133
VAG
100 --- Sig
[oSAMJW Caud oes
RespMin
,o150-
E 100[ L.-Ie
E 50 .
Fig. 12. Decerebrate spinal cord pithed preparation of a cat. The effect onthe caudal oesophagus of afferent vagus nerve stimulation when it alsocaused vomiting. Records from above downwards, Sig, signal, Caud oes,caudal oesophagus, isotonic recording system 3 ml. in balloon. Calibra-tion as in Fig. 5. Resp, thoracic movements of respiration. Min, minutetime marker, B.P. femoral arterial blood pressure. The central end of thedorsal abdominal vagus nerve trunk was stimulated for 12 see with thestimulator set at 12-5 V, 20/sec and pulse duration of 0 5 msec (VAG).
DISCUSSION
The area of the oesophagus studied in detail in the present experimentswas its most caudal 1-2 cm which has been defined, in the cat, by Clark &Vane (1961) and by Botha (1962) as the cardiac or the inferior (caudal)thoracic oesophageal sphincter. The functions of the caudal thoracicoesophageal sphincter require a complex and varied set of reactions tobring them about. The nature of these reactions still remains to be defined,but from the present work a local mechanism can be seen to account formany of the responses of the caudal sphincter of the oesophagus of the cat,which after its dissociation (by denervation) from the central nervoussystem continued to react to distension, the introduction of acid into itand distension of more cranial parts of the oesophagus.The possibility of participation of both local oesophageal and gastro-
oesophageal mechanisms in the stimulation of caudal oesophageal con-tractions by acidification of the oesophagus or of the stomach has beenrecognized (Cannon, 1908; Clark & Vane, 1961; Giles, Humphries, Mason& Clark, 1969). In the three sets of observations referred to, the first two

D. A. TITCHEN AND J. S. WHEELER
in the cat, the last in man, the responses could have been to gastric oroesophageal stimulation. Giles et al. (1969) implicated 'local receptorsbelow the cardia' in the responses they recorded of the oesophagus togastric acidification. Gastric stimulation was avoided in the presentexperiments by having a cannula tied into the gastro-oesophageal junctionand thus limiting the acid and other forms of stimulation to the oesophagusor to the stomach. We suggest that in addition to the local gastro-oeso-phageal mechanisms which Clark & Vane (1961) and Giles et al. (1969)proposed there is a direct response of the oesophagus to its acidificationwhich we obtained when the oesophagus was both isolated from thestomach and freed of any control exercised from the central nervoussystem through its extrinsic innervation, as it was in pithed preparationsof cats.
It is also possible that the pH of the gastric contents contributes to theregulation of the activity of the cardiac sphincter by vago-vagal reflexes.Some indication of this is provided by the experiments shown in Fig. 10.Our failure to evoke such responses after the vagus nerves had been cut inthe abdomen is of interest. Iggo (1957 b) obtained evidence of acid sensitivereceptors in the cat's stomach in studies on vagal afferent fibres. Inruminants such receptors and vagal afferent fibres have been implicatedin the initiation or acceleration of reflex contractions of more cranialdivisions of the stomach on the introduction of acid into the most caudalpart of the ruminant stomach (Titchen, 1953, 1958; Iggo & Leek, 1967).
Excitatory and inhibitory effects of gastric distension on caudal oeso-phageal motility mediated by vagal afferents have been revealed in anumber of studies including the present experiments and those of Clark& Vane (1961). Excitatory effects of small degrees of stretch of thestomach have been interpreted in ruminants as due to vagal afferents,inhibitory effects to splanchnic afferents with still larger degrees of dis-tension exerting inhibitory effects through the effects of other vagalafferents (Dussardier, 1960; Titchen, 1958, 1960). It is of interest to notethat lesser degrees of gastric distension increased, whereas greater gastricdistension inhibited, the caudal oesophageal tone and motility in decere-brate preparations with the spinal cord pithed.
This reduction in caudal oesophageal tone in response to gastric dis-tension demonstrated both by Clark & Vane (1961) and ourselves is animportant part of vomiting. This was obvious in our experiments in whichvomiting was readily stimulated by gastric distension and characteristi-cally was accompanied by a reduction in caudal oesophageal tone andmotility. In both sets of experiments the possibility of distension of thestomach having more direct effects on the caudal region of the oesophaguswas reduced by the experimental procedures adopted, but in neither set of
134

OESOPHAGEAL CONTRACTIONS
experiments were all possible extra-vagal routes of gastro-oesophagealcommunications excluded; the routes referred to include intramuralnervous connexions, communications via abdominal and thoracic gangliaand, in those preparations in which the spinal cord was not pithed, com-munications in the form of gastro-oesophageal spinal reflexes. Nor can thepossibility of some of the responses to gastric stimulation being mediatedby hormonal agencies be excluded. Giles, Mason, Humphries & Clark (1969)have demonstrated effects ofgastrin on the cardiac sphincter in man whichexhibited greater tone and offered a greater resistance to the reflux ofgastriccontents during gastrin administration.The involvement in gastro-oesophageal reactions of vago-vagal reflexes
is further indicated by the demonstration of both excitatory and inhi-bitory responses to afferent vagal stimulation of the abdominal vagusnerve trunks. Since these demonstrations were made in decerebrate spinalcord pithed preparations the possibility that the reactions involvedafferents which leave the vagus in the thorax and enter the thoracic spinalcord (Harper, McSwiney & Suffolk, 1935) has been avoided.The oesophageal contractions evoked by deeper respiratory movements,
either when these occurred spontaneously or were produced by blockingthe tracheal cannula, would presumably be brought about by stimulationof the oesophageal mechanoreceptors both Iggo (1957a) and Mei (1965)identified in their studies on vagal afferent nerve fibres. These workersrecorded greater activity in fibres from the oesophagus at the height ofinspiration. The observation made by Mei (1965) that the receptorsappeared to behave as 'on-off' receptors exhibiting greater frequency ofaction potentials both during oesophageal distension and its relief mayhave some bearing on the dual respiratory effects recorded in the presentexperiments,.with exaggerated inspiratory and expiratory movements.Iggo (1957a) and Mei (1965) found that there were increases in the fre-quency of action potentials in vagal afferents during oesophageal con-traction as well as oesophageal distension and it seems reasonable tosuggest that initially local responses of the oesophagus may assume agreater significance if, following them, there are vagal reflex responses ofthe oesophagus. The temporal sequences of the reflex responses are ofinterest. Lengthy after-discharges and an enduring period over whichexcitability was altered (and summation of an initially inadequate stimuluscould be demonstrated) have been noted in the present experiments, asthey were in studies on the reflex stimulation of gastric movements inruminants (Titchen, 1958, 1960). In the present experiments reflex latencieswere shorter in some reactions than most of the responses encountered inthe ruminant stomach. This may be an important aspect of oesophagealreactions contributing even in the case of its smooth muscle regions to the
135

D. A. TITCHEN AND J. S. WHEELER
rapid removal from its lumen of ingesta which has entered either fromswallowing or reflux from the stomach.
All ofthe reactions of the caudal oesophagus recorded in the experimentsdescribed above contribute to its effectiveness as a sphincter. The reduc-tion of caudal oesophageal activity with distension of cranial parts of theoesophagus provides that an oesophageal relaxation will occur in advanceof the wave of contraction which propels a swallowed bolus caudad. Thedemonstration ofthis reaction in the totally pithed preparation emphasizesthe importance of local oesophageal reactions in the integration of caudaloesophageal sphincter activity. The effectiveness of a sphincter in separat-ing two regions of the alimentary tract depends on its capacity to keep itslumen clear of ingesta; the more direct reactions of the caudal region of theoesophagus to stretch assures its function in this regard, and the activityit displays with the presence in its lumen of acid adds to this effectiveness.Reflex vagal reactions such as described in the present work contribute tothe regulation of oesophageal motility by modifying local more directreactions and coordinating them with conditions in, and reactions of, thestomach and more distant parts of the oesophagus.
We especially wish to record our appreciation of the assistance received from DrC. S. W. Reid to whom our thanks are also due for the generous loan of apparatuswhich allowed the continuation of these experiments at a critical stage of theirdevelopment. The late Mr B. A. Reynolds did much to ensure the success of theseexperiments and his help also is gratefully recorded. The technical assistance receivedfrom Messrs B. Barnett and D. Dellow is acknowledged with gratitude.
REFERENCES
ANDREW, B. L. (1957). Activity in afferent nerve fibres from the oesophagus. J.Phy8iol. 135, 54-55P.
BOTHA, G. S. M. (1962). The Ga8tro-Oe8ophageat Junction, p. 100. London: J. A.Churchill.
BREMNER, C. G., SHORTER, R. G. & ELLIS, F. H. (1970). Anatomy of feline esophaguswith special reference to its muscular wall and phrenoesophageal membrane. J.8urg. Res. 10, 327-331.
CANNON, W. B. (1908). The acid closure of the cardia. Am. J. Phy8iol. 23,105-114.
CHRISTENSEN, J. & DANIEL, E. E. (1966). Effect of some autonomic drugs on circularesophageal smooth muscle. J. Pharmac. exp. Ther. 159, 243-249.
CLARK, C. G. & VANE, J. R. (1961). The cardiac sphincter in the cat. Gut 2, 252-262.CLARK, R. & WEISS, K. E. (1952). Reflex salivation in sheep and goats initiated by
mechanical stimulation of the cardiac area of the forestomachs. Ji S. Afr. vet.med. A88. 23, 163-165.
COMLINE, R. S. & KAY, R. N. B. (1955). Reflex secretion by the parotid gland of thesheep. J. Phy8iol. 129, 55-56P.
COMLINE, R. S. & TITCHEN, D. A. (1951). Reflex contractions of the oesophagealgroove in young ruminants. J. Physiol. 115, 210-226.
136

OESOPHAGEAL CONTRACTIONSDUSSARDIER, M. (1960). Recherches sur la contr6le bulbaire de la motricitA gastrique
chez les ruminants. These Doctorat es Sciences, Institut Nationale de la RechercheAgronomique, Paris.
GILES, G. R., HumPHRIES, C., MASON, M. C. & CLARK, C. G. (1969). Effect of pHchanges on the cardiac sphincter. Gut 10, 852-856.
GILES, G. R., MASON, M. C., HuMPiuRiEs, C. & CLARK, C. G. (1969). Action of gastrinon the lower oesophageal sphincter in man. Gut 10, 730-734.
HARPER, A. A., McSwINEY, B. A. & SUFFOLK, S. F. (1935). Afferent fibres from theabdomen in the vagus nerves. J. Phyaiol. 85, 267-276.
IGGO, A. (1957a). Gastro-intestinal tension receptors with unmyelinated afferentfibres in the vagus of the cat. Q. Jl exp. Phy8iol. 42, 130-143.
IGGo, A. (1957b). Gastric mucosal chemoreceptors with vagal afferent fibres in thecat. Q. JZ exp. Phy8iol. 42, 398-409.
IGGo, A. & LEEK, B. F. (1967). An electrophysiological study of some reticulo-ruminal and abomasal reflexes in sheep. J. Phy8iol. 193, 95-119.
IoRio, L. C. & McIsAAc, R. J. (1965). A simplified method for preparing a pithedcat. J. Pharm. Pharmac. 17, 667-668.
MEI, N. (1965). Etude electrophysiologique des r6cepteurs sensible de l'oesophagethoracique du Chat. C. r. hebd. &anc. Acad. Sci., Pari8 260, 302-305.
MELTZER, S. J. (1899). On the causes of the orderly progress of the peristaltic move-ments in the oesophagus. Am. J. Phy8iol. 2, 266-272.
PORTER, E. L. & ALLAMON, E. L. (1936). Barbiturate-strychnine antagonism in thespinal cat. A quantitative study. J. Pharmac. exp. Ther. 58, 178-191.
SCHENK, E. A. & FREDRICKSON, E. L. (1961). Pharmacologic evidence for a cardiacsphincter mechanism in the cat. Gastroenterology, 40, 75-80.
SELLERS, A. F. & TITCHEN, D. A. (1959). Responses to localized distension of theoesophagus in decerebrate sheep. Nature, Lond. 184, 645-646.
SETEKLIEV, J. (1964a). Uterine motility of the estrogenized rabbit I. Isotonic andisometric recording in vivo - influence of anaesthesia and temperature. Actaphyeiol. 8cand. 62, 68-78.
SETEKLIEV, J. (1964b). Uterine motility of the estrogenized rabbit II. Response todistension. Acta physiol. 8cand. 62, 79-93.
SIEGEL, C. I. & HENDRIX, T. R. (1960). Oesophageal motor disturbances induced byacid perfusion in patients with oesophagitis. Clin. Re8. 8, 369.
TITCHEN, D. A. (1953). Reflex contractions of the reticulum. J. Phyeiol. 122, 32P.TITCHEN, D. A. (1958). Reflex stimulation and inhibition of reticulum contractions
in the ruminant stomach. J. Physiol. 141, 1-21.TrrcIHN, D. A. (1960). The production of rumen and reticulum contractions in
decerebrate preparations of sheep and goats. J. Physiot. 151, 139-153.
137





























