Embed Size (px)
Citation preview

Calliandra (Leguminosae) in continental Africa
Mats Thulin, Philippe Guinet and Asfaw Hunde
/7 f / P
Thulin, M., Guinet, P., and Hunde, A. 1981. Calliandra (Leguminosae) in continen- tal Africa. - Nord. J . Bot. 1: 27-34. Copenhagen. ISSN 0107-055X.
The mainly neotropical genus Calliandrn Benth. (Leguminosae-Mimosoideae- Ingeae) is reported for the first time as native in continental Africa. There are two species known, C. gilberrii Thulin & Hunde sp. nov. in E Kenya and Somalia and C. redoctn (J. H. Ross) Thulin & Hunde comb. nov. in South Africa near the Namibia border. On pollen morphological grounds they are supposed to be most related to certain neotropical species but the pollen does not agree entirely with any previously known group of the genus. The distribution of C. gilbertii and C. redacfa stresses the phytogeographical link between the arid zones of northern and southern Africa.
M . Thuliri and A. Hunde, Inst. of Systemnric Botiiny, Univ. of Uppsnln, Bow 541 , S- 751 2 I Uppsnln, Sweden. P. Guinet, Laboratoire de Pnlynologie, Univ. des Sciences et Techniques du Lnnguedoc, F-340 60 Montpellier Cedew, Frnnre.
Introduction
A curious shrubby species of Leguminosae- Mimosoideae, known from a single fruiting collection from the arid plains of NE Kenya, was provisionally described as Dichrostnchys sp. B. by Brenan (1959). It was then not possible to settle its generic position with any certainty, but recently collected flowering material shows that the plant is not at all a Dichrostachys but instead agrees with the genus Cnllinndra Benth. in the tribe Ingeae. This is a large, mainly neotropical genus with outliers in tropical Asia (India, Burma), the Al- dabra Islands and Madagascar, but with no truly native species previously known from continental Africa.
On phytogeographical grounds one would assume the Kenyan species to have its nearest affinities among the Old World Callinndrns, but pollen morphological data (see below) indicate rather an affinity with certain neotropical species. The closest relative is, however, ‘Acacia” redncta J. H. Ross, a recently described species from near the South Africa/Namibia border, which has very similar pollen, and also agrees in impor- tant gross morphological features. Accordingly the Ke- nyan species is here described as new and A. rednctn is transferred to Cnllinndrn.
Ph. Guinet has contributed the data on pollen mor- phology and the pollen illustrations. A. Hunde has as- sisted with the description of the new species and with
the preparation of some of the illustrations and M. Thu- lin has written most of the text and has been responsible for the preparation of the final manuscript.
Methods
The pollen samples were acetolyzed and mounted in part in glycerol for LM observation, in part on a metal holder in a few drops of distilled water for SEM observation. Mucilage coating on “tail cells” was observed on non-acetolyzed samples cleared with lac- tophenol.
Calliandra gilbertii Thulin & Hunde sp. nov.
Type: Gilbert & Thulin 1288, Kenya, Mandera District, War Gedud, ca. 3”11‘N, 40”52’E, 1. May 1978 (UPS holotype, BR, C , EA, FT, K, MO, P, PRE, WAG, isotypes).
Species nova a C. redoctn (J. H. Ross) Thulin & Hunde stipulis herbaceis parvis, folioliis 4-9-jugis usque ad 3 x 1 mm supra glabris, corolla 3-3.5 mm longa lobis tantum apice pubescentibus, filamentis 8-10.5 mm lon- gis, legumine 40-70 mm longo 24-seminali glabro differt. Figs 1-2, 3A-G.
Much-branched spreading shrub, 1-2.5 m high;
0 NORDIC JOURNAL OF BOTANY NORD. J . BOT. 1: 27-34. TROP 001
Nord J Bot l ( 1 ) 1981 27

branches grey to dark grey-brown with minutely flaking bark, lateral branches diverging at wide angles and be- coming ? spinescent at the ends; young branches pubescent with f curled hairs and with some minute purplish glandular bodies particularly near the nodes. Leaves and flowers on cushion-shaped short-shoots which remain as regularly spaced black knobs on older wood; internodes very short, up to ca. 8 mm long. Stipules lanceolate-falcate, 2.5-3 mm long, connate at the base, ciliate when young, strongly veined, hardened, persistent. Leaves very small, 0.7-2 cm long in all; petiole up to 8 mm long, pubescent, with a prominent mucro at the apex; pinnae 1 pair; rhachillae 4-10 mm long, with a prominent mucro at the apex, pubescent to subglabrous and usually with minute purplish or whitish glandular bodies at the point of attachment of the leaf- lets; leaflets 4-9 pairs, oblong, up to 3 x 1 mm, oblique at the base, shortly acute at the apex; venation raised beneath. Flowers creamy white in (1-)2-3(-4)- flowered heads; peduncles up to 9 mm long, pubescent; bracts ? 1 mm long, ciliate, usually with minute purplish glandular bodies at their point of attachment; pedicels up to 0.5 mm long. Calyx 1.5-2 mm long, with 5 obtuse lobes, ? 0.5 mm long, pubescent on the lobes only. Corolla 3-3.5 mm long, 5-lobed, pubescent at the tips of the lobes only. Stamens ? 20, long-exserted; fi- laments 8-10.5 mm long, united into a tube for t 1.5 mm at the base; anthers dorsifixed, ca. 0.2 x 0.25 mm, glabrous and eglandular. Pollen in large, 8-celled polyads, 135 x 80 x 60 Fm, highly asymmetrical in shape with a single symmetry plane and t oblique as regards the longest axis, pointed at one extremity (a “tail cell”); cells strongly heteromorphic, the central ones 15-25 Fm, the peripheral ones 40-65 pm, all nearly entirely free from each other, except on the apertural areas; exine with a scabrous surface; tectum crossed by few and very small channels (perforations); columellae distinct; apertures of simple pores, both distal and proximal (internal) pores present, the distal ones faced by threes and by fours. Ovary glabrous; style filiform, 10-12 mm long; stigma capitate. Pods erect, compressed, oblanceolate, 4-7 x 0.8-1 cm, attenuate at the base, shortly acute at the apex, brown, glabrous, f transversely venulose and with the sutures much thickened, dehiscing from apex downwards, the valves elastically recurving, 24-seeded . Young seeds rounded, ca. 5 X 3 mm, compressed, flattened and pointed at the hilar end, areolate; rapheole ca. 0.6 mm long; funicle ca. 2 mm long, swollen at the placental end, tapering towards the hilum.
Distribution and habitat. Only known from north-east and east-central Kenya and Somalia (Fig. 5) in rich Acacia-Commiphora bush or woodland on sandy soil at 340-500 m altitude. The single specimen known from
only, but as far as Can be judged at present it is identical with the Kenyan plant.
Fig. 1, Cn[[jnndrG gi/ber,ij, from the type locality, A: Branches Somalia is very poor with loose withered jnflorescences with flowers and buds. B: Erect young pod with markedly thickened sutures. C: Mature pods with recurving valves.
28 Nord. J . Boi. l ( 1 ) 1981
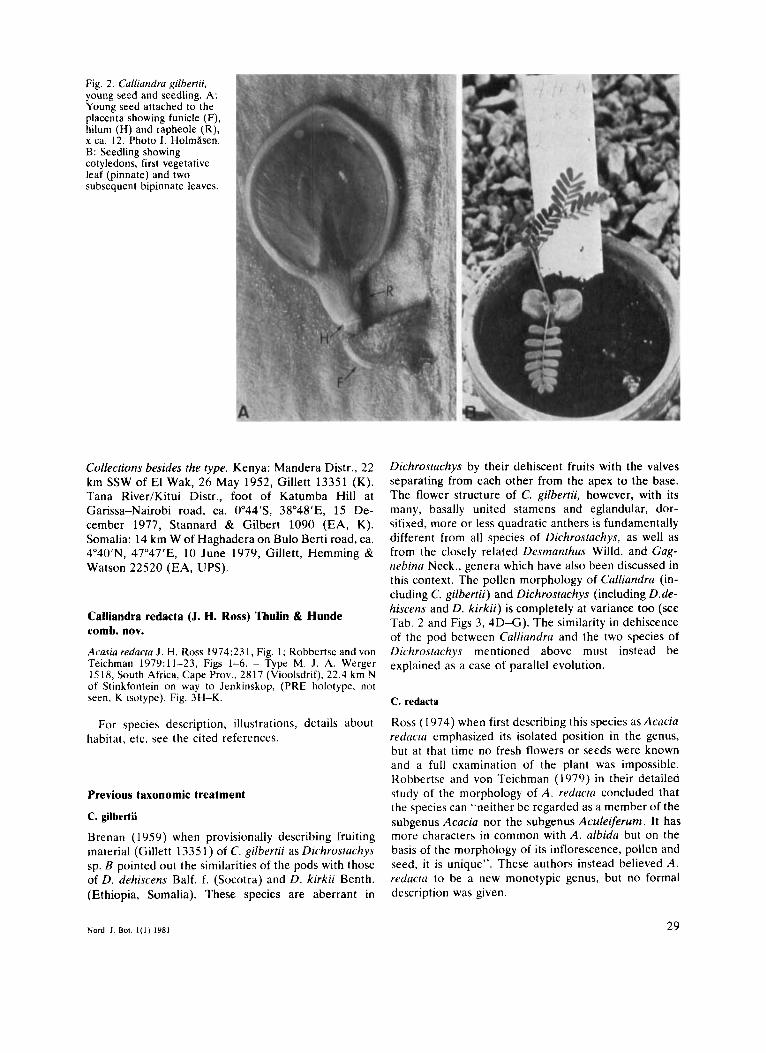
Fig. 2. Culliandru gilbertii, young seed and seedling. A: Young seed attached to the placenta showing funicle (F), hilum (H) and rapheole (R), x ca. 12. Photo I. Holmhen. B: Seedling showing cotyledons, first vegetative leaf (pinnate) and two subsequent bipinnate leaves.
Collections besides the type. Kenya: Mandera Distr., 22 km SSW of El Wak, 26 May 1952, Gillett 13351 (K) . Tana River/Kitui Distr., foot of Katumba Hill at Garissa-Nairobi road, ca. 0"44'S, 38"48'E, 15 De- cember 1977, Stannard & Gilbert 1090 (EA, K). Somalia: 14 km W of Haghadera on Bulo Berti road, ca. 4"40'N, 47"47'E, 10 June 1979, Gillett, Hemming & Watson 22520 (EA, UPS).
Calliandra redacta (J. H. Ross) Thulin & Hunde comb. nov.
Acasiu reducru J . H. Ross 1074:231, Fig. 1; Robbertse and von Teichman 1979:11-23, Figs 1-6. - Type M. J . A. Werger 1518, South Africa, Cape Prov., 2817 (Vioolsdrif), 22.4 km N of Stinkfontein on way to Jenkinskop, (PRE holotype, not seen, K isotype). Fig. 3H-K.
For species description, illustrations, details about habitat, etc. see the cited references.
Previous taxonomic treatment
C. gilbertii
Brenan ( 1 959) when provisionally describing fruiting material (Gillett 13351) of C. gilbertii as Dichroslachys sp. B pointed out the similarities of the pods with those of D . dehiscens Balf. f. (Socotra) and D. kirkii Benth. (Ethiopia, Somalia). These species are aberrant in
Dichrostachys by their dehiscent fruits with the valves separating from each other from the apex to the base. The flower structure of C. gilbertii, however, with its many, basally united stamens and eglandular, dor- sifixed, more or less quadratic anthers is fundamentally different from all species of Dichroslachys, as well as from the closely related Desmanthus Willd. and Gag- nehina Neck., genera which have also been discussed in this context. The pollen morphology of Calliandra (in- cluding C. gilbertii) and Dichrostachys (including D.de- hiscens and D . kirkii) is completely at variance too (see Tab. 2 and Figs 3, 4D-G). The similarity in dehiscence of the pod between Calliandra and the two species of Dichrostachys mentioned above must instead be explained as a case of parallel evolution.
C. redacta
Ross (1974) when first describing this species as Acacia redacta emphasized its isolated position in the genus, but at that time no fresh flowers or seeds were known and a full examination of the plant was impossible. Robbertse and von Teichman (1979) in their detailed study of the morphology of A . redacta concluded that the species can "neither be regarded as a member of the subgenus Acacia nor the subgenus Aculeiferum. It has more characters i n common with A. albida but on the basis of the morphology of its inflorescence, pollen and seed, i t is unique". These authors instead believed A. redocra to be a new monotypic genus, but no formal description was given.
Nord 1. Bot. l ( 1 ) 1981 29

Tab. I . Differences between Cdlirindrri gdberrzi and c‘. redacftr.
Characters C. gilberrii C. redacro
Stipules herbaceous, 2.5-3 mm long spinescent, 5-15.5 mm long
Leaflets
Sterile bract absent present on peduncle Flowers shortly pedicelled sessile
Calyx 1.5-2 mm long, glabrous except for tips of lobes Corolla 3-3.5 mm long, lobes pubescent at tips only Filaments 8-10.5 mm long 12-15 mm long
Pods 40-70 mm long, 24-seeded, glabrous
4-9 pairs per pinna, up to 3x 1 mm, pubescent beneath only
2 4 pairs per pinna, up to 5 x 3 mm, pubescent on both surfaces
1-1.5 mm long, pubescent 4-6 mm long, lobes pubescent
25-40 mm long, 1-2-seeded, densely pubescent
Comparison of C. gilbertii and C. redacta
All important diagnostic characters for the genus Cal- liandra are found in both C. gilbertii and C. redacia. Such characters include bipinnate leaves, capitate in- florescences, many long-exserted stamens basally un- ited into a tube and pods with the valves elastically opening from the apex to the base. In no gross mor- phological feature the two species fall outside the varia- tion found elsewhere in the genus (for discussion of pollen morphology see below). Nevertheless the two species differ inter se in many respects and the more prominent gross morphological differences are sum- marized in Tab. 1.
Seedlings of C. gilbertii were raised in greenhouses at Uppsala Botanical Garden from seeds collected on the type locality (Fig. 2B). The cotyledons are petiolate and slightly cordate at the base. The first vegetative leaf is pinnate with seven pairs of leaflets. The second and all subsequent leaves are bipinnate with one pair of pinnae, each with about six pairs of leaflets. Thus already in the second leaf all features of the mature leaves are present. There are some quantitative differences, however, The petiole of the second leaf is ca. 13 mm long and the leaflets are ca. 5 X 2 mm, which is longer respectively larger than corresponding dimensions on mature leaves. The seedling of C. gilbertii belongs to group 8 B in the classification proposed by Robbertse and van der Schijff (1971) for the seedlings of Acacia. This type of seedling was also reported for C. redacta (as Acacia r . ) by Rob- bertse and von Teichman (1979:12, Fig. 1B). N o in- formation on seedlings in neotropical Calliandra has been available to us.
Comments on pollen morphology
A comparison of some important pollen morphological characters in Calliandra gilbertii and C. redacta, the rest of the genus Calliandra, Acacia, and Dichrostachys is presented in Tab. 2.
Comparison with Acacia
A columellar exine structure as in C. gilbertii and C. redacta does occur in the subgenus Acacia (= Gum- miferae Benth.), which is otherwise characterized by its colporate apertures. Such apertures do not occur in Calliandra. Eight-celled polyads with the cells in a single plane and with a “tail cell” occur in Acacia only in a few species of the subgenus Heterophyllum which is mainly Australian (but with one species in Madagascar and another on Reunion). However, these species all have polyads with a very poorly differentiated “tail cell”, a supratectal reticulum, a granular exine structure and a pseudo-furrow (see Fig. 4A-C).
The suggestion by Ross (1974) that “Acacia redacta” may be closely related to Acacia (Faidherbin) albida Del. can be ruled out by the entirely different pollen of this species (Guinet and Lugardon 1976)
Comparison with Calliandra
The genus Callinndra as presently understood is com- prised of three groups which correlate with geography at least in part. The first, previously called “Calliandra group I-A” in reference to its pollen characters (Guinet
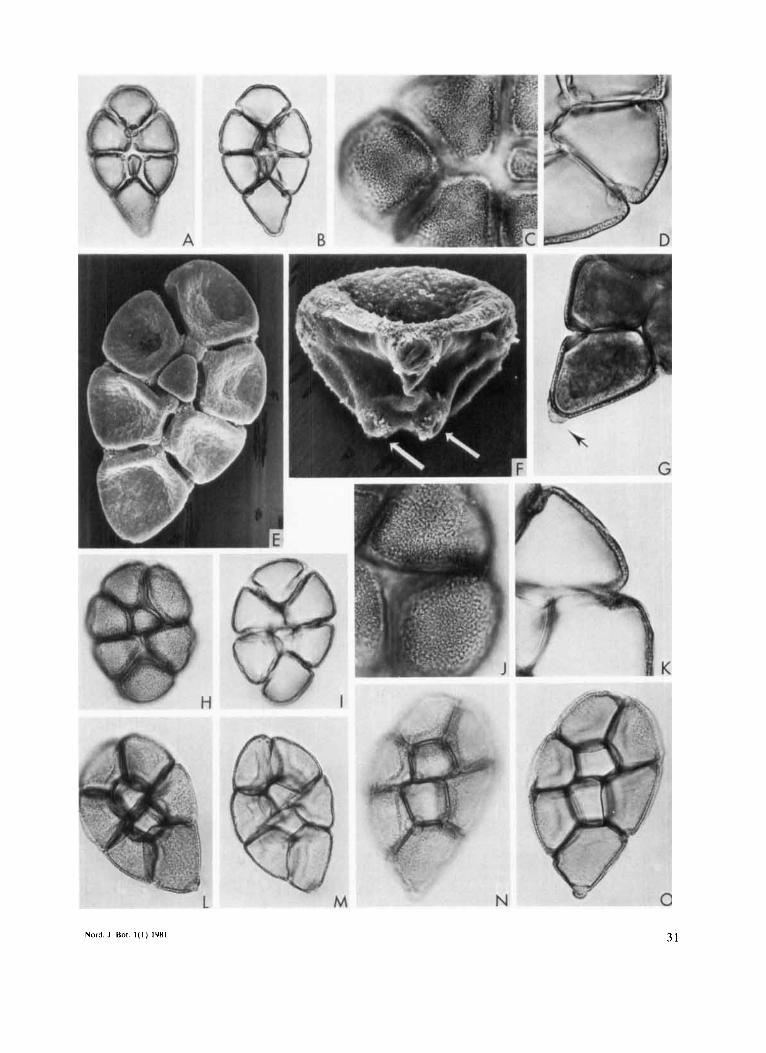
Fig. 3. Pollen of Culliundru. A-G: C. gilbertii, from the holotype. A: 8-celled polyad; note great difference in size between central and peripheral cells and presence of proximal pores, xca. 400. B: Ditto in optical section. C: Part of polyad showing exine ornamentation, xca. 800. D: Ditto showing columellar structure of the exine. E: 8-celled polyad; the cells are free from one another, except in the places where apertures are present, xca. 750 (SEM). F: Detail of an isolated cell; note presence of proximal pores (arrows), Xca. 2050 (SEM). G: Detail of polyad (fresh material) showing tail cell with mucilage coating (arrow), xca. 750. H-L: C. redacta, from the isotype (K). H: 8-celled polyad, xca. 400. 1: Ditto in optical section. J: Part of polyad showing exine ornamentation, xca. 800. K: Ditto showing columellar structure of the exine. L-M: C. carbonariu Benth., from Hartweg 964, Colombia (P). L: 8-celled polyad, xca. 400. M: Ditto in optical section. N-O: C. subnervosa Benth., from Camp 3177, Ecuador (NY). N: 8-celled polyad, xca. 400. 0: Ditto in optical section.
30 Nord. J . Bot. l ( 1 ) 1981
Nord J BOI l(1) 1981 31
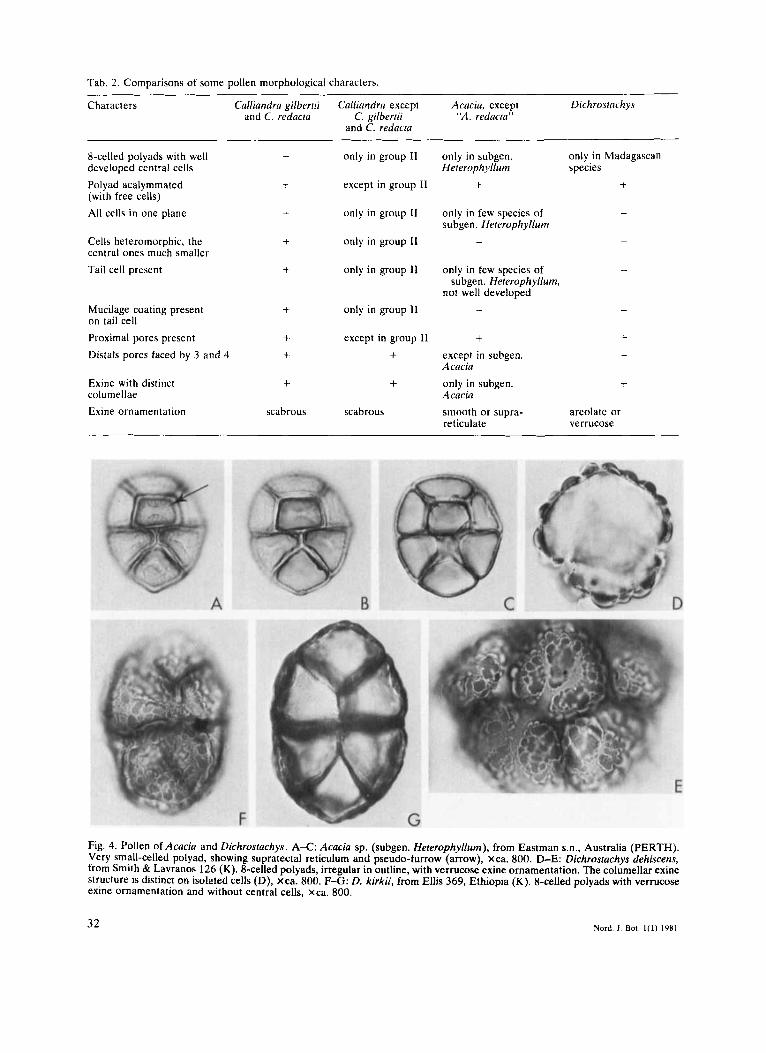
Tab. 2. Comparisons of some pollen morphological characters.
Characters Cnlliandra gilberiii Cullicmdra except Acacia, except Dichrostachys and C. redacra C. gilberrii "A. redacta"
and C. redncta
8-celled polyads with well developed central cells Polyad acalymmated (with free cells) All cells in one plane
Cells heteromorphic, the central ones much smaller Tail cell present
Mucilage coating present on tail cell Proximal pores present Distals pores faced by 3 and 4
Exine with distinct columellae Exine ornamentation
+ + + +
+
+ + +
+
scabrous
only in group I1
except in group I1
only in group I1
only in group I1
only in group I1
only in group I1
except in group I1 + +
scabrous
only in subgen. Heterophyllum
+
only in few species of subgen. Hererophyllum
-
only in few species of
not well developed subgen. Heterophyllum,
-
+ except in subgen. Acacia only in subgen. Acacia smooth or supra- reticulate
only in Madagascan species
+
+ -
areolate or verrucose
Fig. 4. Pollen of Acacia and Dichrostachys. A X : Acacia sp. (subgen. HeterophyNum), from Eastman s.n., Australia (PERTH). Very small-celled polyad, showing supratectal reticulum and pseudo-furrow (arrow), xca. 800. D-E: Dichrostachys dehiscens, from Smith & Lavranos 126 (K). 8-celled polyads, irregular in outline, with verrucose exine ornamentation. The columellar exine structure is distinct on isolated cells (D), xca. 800. F-G: D. kirkii, from Ellis 369, Ethiopia (K). %celled polyads with verrucose exine ornamentation and without central cells. xca. 800.
32 Nord. J . Bot. l ( 1 ) 1981

1965) unites the few previously known paleotropical species (from India, Burma, Aldabra Islands, and Madagascar), whose pollen characters show the generalized pollen type of the Ingeae, i.e. 16-celled symmetric polyads with an areolate surface and a granular exine structure. This group, possibly generi- cally distinct, needs further study (Nielsen and Guinet in preparation), but is quite distinct from the two species treated here. The second group, “Calliandra group I-B”, is restricted to the neotropics and cannot be compared with C. gilbertii or C . redacta, the pollen morphology is too different (Guinet 196.5). The third group, “Calliandrtl group II”, which includes the type of the genus, C. grandiflora (L’Herit.) Benth., is again en- tirely neotropical and is defined by its highly asymmet- rical eight-celled polyads provided with a “tail cell” (Fig. 3L-0) which in several species is smeared with mucilage. It is with this group, uniting most species classified by Bentham (187.5) in his series Nitidae and Pedicellatae, that C. gilbertii and C. redacta show af- finities. However, these two species differ from all the species of pollen group I1 by two characters: the cells of the polyad are free from one another (acalymmated polyad) and provided with proximal (internal) pores (Fig. 3E, F). Such pores, however, are present in groups I-A and I-B. Obviously the two African species cannot be included in any previously known group of Cal- liundra, although they are clearly congeneric, at least with the present broad circumscription of Culliandra. From a speculative point of view, accepting the idea that before being,fused within a polyad, the cells should be free from one another, these two species would repre- sent old remnants of an archaic group in this genus.
Comments on phytogeography
Despite the obvious similarity in flower and fruit struc- ture between the two species treated here and Cril- linndra no reference at all has been made to this genus by the previous authors dealing with these species. Certainly this owes to the fact that they are the first native species of Calliandra to be discovered on the African continent (C. xylocarpu Sprague 1908:290 de- scribed as a native species from Mozambique is now Bussea xylocarpa (Sprague) Sprague & Craib). Several New World species have been cultivated in various parts of Africa, however, and in West Africa at least one of them, C . portoricensis (Jacq.) Benth., has become naturalized.
The distribution of C . gilberrii and C. reductti (Fig. 5 ) provides yet another example of the phytogeographical link between the arid zones of northern and southern Africa. Other examples on various taxonomic levels are given by Volk (19,65:44). de Winter (lY71). Lebrun (1971), Thulin (1975:54) and Goldblatt (1978). To find examples where this link is combined with a neotropical affinity as in the case of Calliandra is more difficult, but
Fig. 5. Known distributions of Cdliondm gilherrii ( 0 ) and C. rcdocro (.).
two excellent parallels are found by the genera Turnera L. (Turneraceae) and Parkinsoniu L. (Leguminosae- Caesalpinioideae). Turnera has some 60 species in tropical and subtropical America, one species in SW Africa and one in E and N E Africa (Story 1961). Par- kinsonia has about a dozen species in the drier parts of N and S America, one in S and SW Africa and three (one of which is not yet published) in E and NE Africa (Brenan 1963:203, 1967:43). The isolated, narrow dis- tributions found in Africa for the species in these exam- ples support the view that they are of a relic nature and indicate that a widespread extinction of these groups has taken place on this continent.
Acknowlrdgenicria - The specific epithet o f the new species is given in honour of Mr Michael G. Gilbert, Nairobi, from whose friendship and knowledge of the NE African flora we have much profited. We are indebted to the Director and Curators at Kew for providing polliniferous material and for the loan of an isotype of C‘czllirindro redrrc,tcr. We are also grateful to Dr B. Maslin. Perth, for providing material of the Acncirr species studied and to Prof. P. J . Robbertse, Pretoria, for supplementary flowering material of C. rrdwtm.
3 Nord J . Bot ! ( I ) 1Y81 33

References Bentham, G. 1875. Revision of the suborder Mimoseae. -
Trans. Linn. SOC. London 30: 335-668. Brenan, J. P. M. 1959. Leguminosae subfamily Mimosoideae.
- In: Hubbard, C. E. and Milne-Redhead, E. (eds.): Flora of Tropical East Africa. Crown Agents for Oversea Gov- ernments and Administrations, London and Tonbridge.
- 1963. Notes on African Caesalpiniofdeae. - Kew Bull. 17: 197-2 14.
- 1967. Leguminosae subfamily Caesalpinioideae. - In: Milne-Redhead, E. and Polhill, R. (eds.): Flora of Tropical East Africa. Crown Agents for Oversea Governments and Administrations, London and Tonbridge.
Goldblatt, P. 1978. An analysis of the flora of Southern Africa. its characteristics, relationships, and origins. - Ann. Mis- souri Bot. Gard. 65: 369-436.
Guinet, Ph. 1965. etude des charactem du pollen dans le genre Calliandra (Mimosaceae). - Pollen Spores 7:
- and Lugardon, B. 1976. Diversite des structures de l’exine dans le genre Acacia (Mimosaceae). - Pollen Spores 18:
157-173.
483-5 1 1.
Lebrun, J.-P. 1971. Quelques phanerogames africaines A aire disjointe. - Mitt. Bot. Staatssamml. Miinchen 10: 438448.
Robbertse, P. J. and van der Schijff, H. P. 1971. The genus Acacia Miller in South Africa. - Mitt. Bot. Staatssamml. Miinchen 10: 170-177.
- and von Teichman, I. 1979. The morphology of Acacia redacta J. H. Ross. - J.S. African Bot. 45: 11-23.
Ross, J. H. 1974. Notes on Acacia species in Southern Africa: IV. - Bothalia 11: 231-234.
Sprague, T. A. 1908. Culliandra xylocarpa. - Kew Bull. 1908: 290.
Story, R. 1961. Turneraceae. - Bothalia 7: 493-496. Thulin, M. 1975. The genus Wahlenbergiu slat. (Cam-
panulaceae) in tropical Africa and Madagascar. - Symb. Bot. Upsal. 21 (1).
Volk, 0. H. 1965. Die Florengebiete von Siidwestafrika. - J. South West Africa Sci. SOC. 20: 25-58.
Winter, B. de 1971. Floristic relationship between the north- em and southern arid areas in Africa. - Mitt. Bot. Staatssamml. Miinchen 10: 424-437.
34 Nord. J. Bot. l ( 1 ) 1981





























