Embed Size (px)
Citation preview

“Cambio en la Población de Células de Paneth en Ratas Diabéticas”
RESUMEN
La diabetes mellitus es un padecimiento de diversa etiología, debido a insuficiencia completa o relativa de la secreción y/o actividad de la insulina y manifestada con desórdenes metabólicos múltiples que se caracteriza por alteraciones en el procesamiento de los carbohidratos, las grasa y las proteínas y se expresan a nivel bioquímico, fisiológico o morfológico. La diabetes experimental utilizando como modelo biológico la rata permite analizar los eventos de diferente índole que suceden en este estado patológico. La estreptozotocina es uno de los medios utilizados para inducir esta patología. La célula de Paneth forma parte del epitelio del intestino delgado; posee en su citoplasma apical numerosos gránulos bipartitas cuyo contenido le confiere sus múltiples funciones, siendo la principal, el control de la flora bacteriana normal que habita en el intestino, formada en su mayoría por anaerobios. En el contenido de dichos gránulos destacan alfa defensinas, lisozimas, IgA e IgG. Como todas las células del organismo, la célula de Paneth necesita de condiciones ideales para llevar a cabo su función, por lo que la llegada de O2 y de nutrientes es vital; de ahí que las alteraciones de la homeostasis tengan una repercusión directo sobre su microambiente, provocando cambios en su función y también en su morfología. Investigar si la hiperglicemia sostenida de un cuadro diabético, influye sobre la morfología de las células de Paneth. Se trabajo con ratas Wistar macho de 250g de peso fueron divididas en un grupo control, uno diabético y uno diabético con restitución hormonal; se mantuvieron vivas por dos meses y se sacrificaron por perfusión con paraformaldehido al 4%, extrayendo el intestino delgado, del que se obtuvieron los segmentos duodeno, yeyuno, ileon medio e íleon terminal; se procesaron por el método histológico convencional y se obtuvieron cortes de 5 micras de espesor que se colorearon, con el método tricrómico de Gomori, histoquímica e Inmunohistoquímica se examinaron en microscopio de campo claro. Se observaron los gránulos de las células, con su centro denso y halo claro pero de tamaño variable, clasificándolos como pequeños, medianos, grandes y gigantes. Se encontró que su frecuencia fue diferente en los grupos de animales, siendo los pequeños más abundantes en el grupo control y los gigantes en el grupo diabético; Estos cambios pueden indicar que la célula es estimulada en su función de síntesis, o inhibida en lo que respecta a la secreción de sus gránulos hacia la luz intestinal.
INTRODUCCIÓN El metabolismo de todo organismo viviente se encuentra regulado por una inmensidad de señales bioquímicas que tienen como objetivo común mantener un estado de equilibrio para realizar sus funciones de manera adecuada, tales como respirar, moverse, pensar, defenderse de organismos patógenos, responder a estímulos externos, producir energía, regular su temperatura, entre otras más. La célula, unidad anatomofuncional de cualquier organismo, es una estructura sumamente compleja y diferenciada, capaz de realizar una tarea en común en conjunto con las demás que conforman un organismo; para que ésta se lleve a cabo se necesita una regulación endocrina compuesta por sustancias biológicamente activas, teniendo así en primer lugar a las hormonas.

La célula de Paneth, también llamada “La enigmática célula de Paneth” o “La multifacética célula de Paneth” se encuentra formando parte del epitelio de la mucosa del intestino delgado, situada en el fondo de las criptas intestinales o de Lieberkuhn; su ciclo vital es largo careciendo de mitosis ya que se origina a partir de una célula progenitora pluripotencial. La célula de Paneth se caracteriza por su morfología piramidal y sobre todo por la presencia de gránulos bipartitas acidófilos en su citoplasma apical, siendo un célula secretora de diversas moléculas entre las que destacan lisozimas, fosfolipasa A, IgA y péptidos antimicrobianos (defensinas o criptidinas), los cuales tienen actividad contra bacterias gram positivas, gram negativas, hongos y virus, de ahí que entre otras funciones sea considerada como elemento de importancia en el control de la flora bacteriana y la inmunidad innata. La extensión de la población de células de Paneth y el aspecto de sus gránulos son variables en diferentes especies animales así como su respuesta a factores diversos, tanto externos como internos entre ellos el tipo de dieta alimentaria, la administración de ciertos fármacos, distintos grados de estrés y estímulos nocivos. Su respuesta corresponde a estímulo o inhibición de sus funciones de síntesis y de secreción que se expresan por modificaciones de su contenido o por el cambio en el número y/o tamaño de sus gránulos. Las modificaciones en su población son, por otro lado, expresión del efecto sobre las células madre a partir de las cuales se diferencia, o de efecto directo del agente utilizado sobre las células de Paneth. Siendo la diabetes mellitus un padecimiento de diversa etiología, que se debe ya sea a un déficit completo o parcial de Insulina o un déficit en la presentación o eficiencia de sus receptores en las células blanco, que se manifiesta con desórdenes metabólicos múltiples caracterizados por alteraciones en el procesamiento de carbohidratos, grasas y proteínas, en este trabajo se buscó identificar respuesta de las células de Paneth a las señales metabólicas provocadas por este padecimiento que entre otras se encarguen de regular su actividad. Hipótesis La célula de Paneth responde a los cambios de la glicemia producidos por la diabetes. Objetivo Investigar mediante métodos histológicos e histoquímicos cambios morfofuncionales en las células de Paneth en rata, mediante un modelo experimental de Diabetes tipo 1 insulinodependiente producido por Estreptozotocina. Metodología Se utilizaron 30 ratas Wistar macho de 200g de peso, las cuales se dividieron en tres grupos de diez ratas cada uno, teniendo así un grupo control (C), uno diabético (D) y uno diabético bajo restitución hormonal (R). Para montar el modelo experimental de Diabetes insulinodependiente se hizo uso de un fármaco tóxico altamente afín a las células beta pancreáticas sintetizado por el Streptomyces achromogenes llamado Estreptozotocina (STZ). De acuerdo a la bibliografía consultada se aplicó un tratamiento de 80 mg/kg de peso de STZ vía intraperitoneal dividida en dos dosis de 40 mg/kg, aplicadas con intervalo de 24 horas para las ratas de los grupos diabéticos (D y R), mientras que al grupo control, para

simular el estrés de la inyección, se les aplico únicamente el vehículo de la STZ, un amortiguador de citratos a pH de 4.5. Durante toda la duración del experimento se realizaron tomas de peso y de glicemia cada semana con el objeto de observar los cambios sistémicos llevando así un control del estado hiperglicémico y posteriormente para la regulación en las dosis de insulina. El peso se midió mediante una balanza digital y la medida de la glicemia se realizó mediante un glucómetro de la marca Accutrend GC a base de tiras reactivas el cual da los datos en mg/dl ; la toma de sangre se hizo en ayuno mediante un corte en la porción distal de la cola de cada rata. Los grupos de ratas se mantuvieron durante 33 días, al término se sacrificaron las10 ratas diabéticas del grupo D y 5 de las ratas del grupo C. Enseguida se comenzó la aplicación de insulina NPH de la marca HUMULIN a las ratas diabéticas restantes (grupo R) para restituirlas hormonalmente. Este tipo de Insulina de acción intermedia, se administró por un periodo de 43 días por vía subcutánea a las 7 de la noche considerando que la aplicación debería preceder la ingesta de alimento y que las ratas tienen hábitos de alimentación nocturna. De la misma manera, a los controles (grupo C) se les aplicó una inyección de solución fisiológica al 9%. Al término del experimento se sacrificaron las ratas del grupo R y las 5 restantes del grupo C. El sacrificio se realizó por perfusión intracardíaca de paraformaldehido al 4%. Se extrajo el intestino delgado, del que se obtuvieron los segmentos duodeno, yeyuno, íleon medio e íleon terminal, se procesaron por el método histológico convencional y se obtuvieron cortes de 5 micras de espesor que se montaron en serie y se colorearon con el método topográfico tricrómico de Gomori, histoquímico con Lectina de Trigo WGA e inmunohistoquímico con anticuerpo contra la subunidad beta del receptor de insulina. El examen de las preparaciones se realizó en un microscopio de campo claro Carl Zeiss y de las coloreadas por el método topográfico y el histoquímico primero se leyeron todos los cortes de una preparación para seleccionar los dos mejores de cada una; sobre los cortes seleccionados se realizó la cuenta del número de criptas por corte, la descripción de la morfología de las células de Paneth y la cuenta de su número por cada corte. Los resultados fueron analizados estadísticamente comparando los tres grupos de animales y estableciendo correlaciones entre el estado metabólico y el número de células de Paneth
RESULTADOS Y DISCUSIÓN
A.-Condiciones metabólicas
300
350
400
450
Control Diab Diab + INS
orpo
ral (
g)
300
350
400
450
Control Diab Diab + INSControl Diab Diab + INS
orpo
ral (
g)

Figura 1. Cambios en el peso corporal de las ratas a lo largo del experimento. Se indujo diabetes con estreptozotocina el día 1, se mantuvieron hasta el día 33 (Diab) y a partir de éste se controló la diabetes con inyecciones diarias de insulina (3 UI/día) hasta el final del experimento (Diab+INS). El grupo control recibió inyecciones de solución salina en volumen equivalente. Los datos mostrados son medias ± e.e. (n=10 para grupos diabéticos; n=5 para controles). La administración de estreptozotocina induce la muerte de células beta del páncreas endócrino, lo que conduce a un estado de diabetes dependiente de insulina. En estas condiciones, se reduce en los animales la capacidad de asimilación de nutrimentos y se eleva la concentración de glucosa circulante. Los resultados de este experimento (figura 1) muestran que los animales tratados con estreptozotocina redujeron su tasa de ganancia de peso desde el día 10 postratamiento, y la tendencia a ganar peso prácticamente desapareció a partir del día 15. La administración diaria de insulina a partir del día 33 restableció la ganancia de peso con una tasa mayor que la ganancia normal de los controles, de manera que ambos grupos llegaron a tener promedios de peso no diferentes entre sí a partir del día 77. Los datos de glucemia (figura 2) complementan las observaciones sobre el peso: el tratamiento con estreptozotocina induce un aumento muy marcado de la glucosa sanguínea, que es evidente a partir del día 10 y que se mantiene prácticamente sin cambio hasta el día 60. A partir de este momento la glucemia se reduce y los animales y tratados retornan a valores semejantes al grupo control en el día 71. La restitución de insulina (iniciada el día 33) no tiene un efecto inmediato sobre la glucosa pero sí sobre el peso, lo que sugiere una recuperación gradual del estado metabólico que se inicia con la recuperación generalizada de las funciones celulares de asimilación y crecimiento (respuesta de umbral bajo) y continúa con la normalización de los procesos de suministro energético (respuesta de umbral alto). Es notable el hecho de que los animales diabéticos tratados con insulina mostraron tasas aceleradas de ganancia de peso hasta alcanzar finalmente el mismo peso corporal que los controles, lo que indica que la diabetes, aún implicando cambios metabólicos dramáticos, no modifica el punto de regulación del peso corporal. En conjunto, estos resultados demuestran que las condiciones experimentales de diabetes dependiente de insulina y la recuperación mediante inyecciones de insulina produjeron la situación metabólica que se esperaba; los cambios observados en la morfología intestinal pueden ser interpretados como consecuencia de las alteraciones asociadas a la diabetes.

Figura 2. Concentración plasmática de glucosa medida en diferentes momentos a lo largo del experimento. Se indujo diabetes con estreptozotocina el día 1, se mantuvieron hasta el día 33 (Diab) y a partir de éste se controló la diabetes con inyecciones diarias de insulina (3 UI/día) hasta el final del experimento (Diab+INS). El grupo control recibió inyecciones de solución salina en volumen equivalente. Los datos mostrados son medias ± e.e. (n=10 para grupos diabéticos; n=5 para controles).
B.- Morfológicos Se observaron cambios en el aspecto de los gránulos en relación con su dimensión aparente. Los gránulos se clasificaron de acuerdo a su tamaño en pequeños, medianos, grandes y gigantes. Las ratas control presentaron gránulos pequeños (en su mayoría) y gránulos grandes; las ratas diabéticas fueron las que presentaron cambios importantes con la presencia de gránulos grandes y gigantes; y las ratas tratadas presentaron gránulos medianos y grandes, lo que sugiere que al tratar de regresar a su estado normal la glicemia, los gránulos poco a poco se vuelven a asemejar a los de as ratas control.
Grupo de ratas
Gránulos predominantes
Grupo Control
Pequeños y Grandes
0 20 40 60 800
100
200
300
400
Días
Glu
cem
ia (
mg/
dL)
Control Diab Diab + INS
0 20 40 60 800
100
200
300
400
Días
Glu
cem
ia (
mg/
dL)
Control Diab Diab + INS
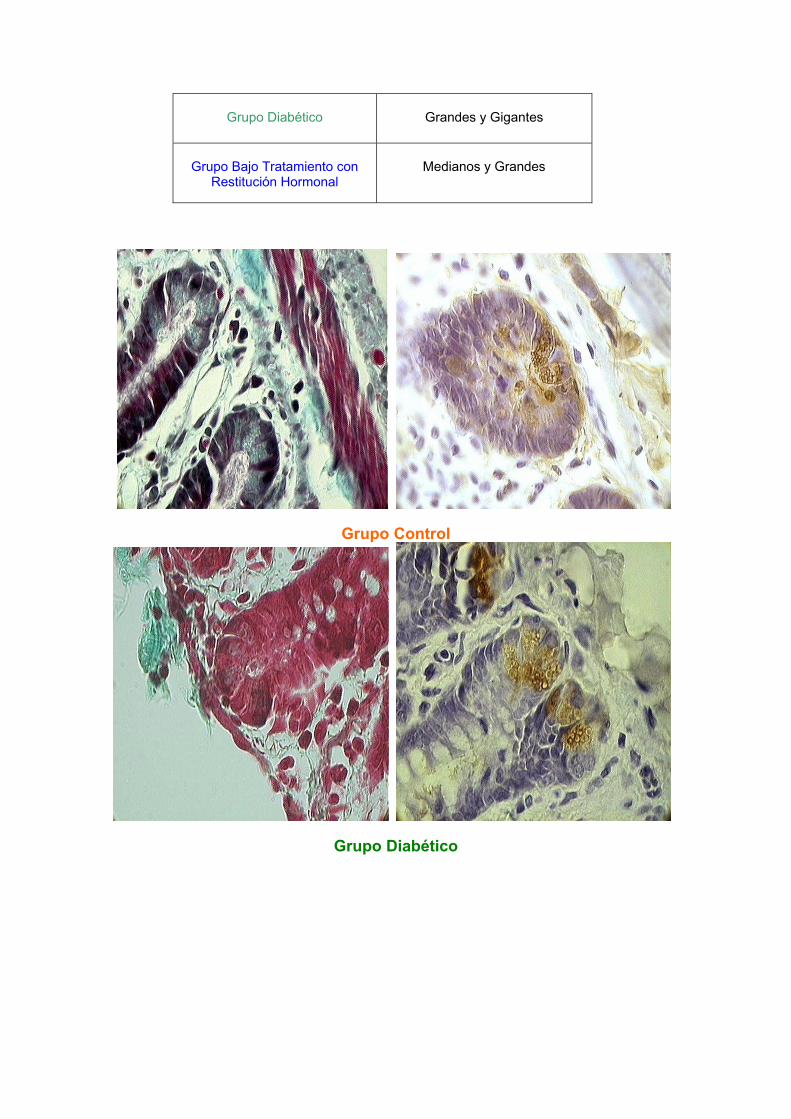
Grupo Diabético
Grandes y Gigantes
Grupo Bajo Tratamiento con
Restitución Hormonal
Medianos y Grandes
Grupo Control
Grupo Diabético

Grupo Diabético bajo Restitución Hormonal
C.- Inmunohistoquímica: Se observó positividad ligera a nivel de los capilares de la capa submucosa y no a nivel de las células de Paneth al anticuerpo utilizado en esta investigación contra receptor a insulina, lo cual sugiere que estas células no expresan este tipo de receptores, y que la vía de señalización y el mecanismo de la respuesta de esta población celular, es mediada por otros factores pendientes de investigar. D.- Número de criptas intestinales
Duodeno
33 810
100
200
Días
No.
Crip
tas
/ cor
te
Yeyuno
33 810
100
200
Días
No.
Crip
tas
/ cor
te
Íleon medio200
s / c
orte
Íleon terminal
100
150
s / c
orte
Criptas de Lieberkühn
**Duodeno
33 810
100
200
Días
No.
Crip
tas
/ cor
te
Yeyuno
33 810
100
200
Días
No.
Crip
tas
/ cor
te
Íleon medio200
s / c
orte
Íleon terminal
100
150
s / c
orte
Criptas de Lieberkühn
Duodeno
33 810
100
200
Días
No.
Crip
tas
/ cor
te
Yeyuno
33 810
100
200
Días
No.
Crip
tas
/ cor
te
Íleon medio200
s / c
orte
Íleon terminal
100
150
s / c
orte
Criptas de Lieberkühn
**

Figura 3. No. de criptas intestinales por corte en 4 diferentes regiones del intestino delgado de ratas con diabetes no compensada (sacrificadas el día 33) o controlada con inyecciones diarias de insulina (3 UI/día) a partir del día 33 y sacrificadas el día 81 después de la inducción de la diabetes, o animales control. Los datos mostrados son medias ± e.e. (n=10 para grupos diabéticos; n=5 para controles). **: P<0.01 entre los grupos indicados. Los datos fueron analizados mediante pruebas de análisis de varianza bifactorial. En los animales control, el número de criptas fue significativamente más alto en el duodeno y el íleon medio que en las demás regiones (P<0.001). Solamente en el duodeno se redujo el número de criptas del día 33 al día 81, quizás como consecuencia del aumento de peso de los animales o de las inyecciones repetidas de solución salina, aunque esto último es poco probable. Por lo demás, el número se criptas no se modificó significativamente por efecto de la diabetes ni de la inyección diaria de insulina. Estos datos muestran que el número de criptas intestinales no es afectado por cambios, aun importantes, en la concentración de insulina y en la disponibilidad de glucosa. E) Número de células de Paneth
Duodeno
33 810
50
100
150
200
250
Días
célu
las
Pane
th /
cort
e
Yeyuno
33 810
50
100
150
200
250
Días
célu
las
Pane
th /
cort
e
Íleon medio350
e
Íleon terminal300
e
Células de Paneth
***
*
*
Duodeno
33 810
50
100
150
200
250
Días
célu
las
Pane
th /
cort
e
Yeyuno
33 810
50
100
150
200
250
Días
célu
las
Pane
th /
cort
e
Íleon medio350
e
Íleon terminal300
e
Células de Paneth
Duodeno
33 810
50
100
150
200
250
Días
célu
las
Pane
th /
cort
e
Yeyuno
33 810
50
100
150
200
250
Días
célu
las
Pane
th /
cort
e
Íleon medio350
e
Íleon terminal300
e
Células de Paneth
***
*
*

Figura 4. No. de células de Paneth por corte en 4 diferentes regiones del intestino delgado de ratas con diabetes no compensada (sacrificadas el día 33) o controlada con inyecciones diarias de insulina (3 UI/día) a partir del día 33 y hasta el día 81 después de la inducción de la diabetes. Los datos mostrados son medias ± e.e. (n=10 para grupos diabéticos; n=5 para controles). *: P<0.05; **: P<0.01 entre los grupos indicados. Los datos sobre el número de células de Paneth (CP) en las distintas regiones intestinales de los animales control coinciden con observaciones reportadas previamente, que indican densidades mayores de estas células en las porciones distales del intestino (P<0.001). A diferencia del número de criptas, el número de CP aumentó ligeramente en los animales diabéticos (efecto no significativo) pero se redujo después del tratamiento con insulina (P<0.05). Un análisis de correlación aplicado a estos datos muestra este efecto más claramente (figura 5): el número de células de Paneth en los animales control tiende a aumentar ligeramente con la glucemia (o con la reducción de la insulina circulante), de modo que la recta que relaciona estas variables tiene una pendiente positiva (0.13). Los animales diabéticos muestran la tendencia opuesta, de manera que la pendiente es negativa (-0.06), pero al aplicarse a estos animales el tratamiento con insulina, la pendiente negativa se invierte hasta hacerse similar a la de los controles (0.12). Cambios parecidos se observan al relacionar el peso corporal con el número de células de Paneth (figura 6): la pendiente negativa (-0.15) que relaciona el número de células de Paneth con el peso del animal se torna positiva (0.18) en los animales diabéticos, y vuelve a ser negativa (-0.19) cuando éstos son tratados con insulina. El número de criptas intestinales no muestra los mismos cambios en las tendencias, porque las relaciones entre estas variables son muy parecidas entre las tres situaciones experimentales (figura 7). En conjunto, los resultados de este experimento indican que la hiperglucemia diabética, la insulina por sí misma o algún otro factor asociado tienen efecto sobre la densidad de CP, de modo independiente del número de criptas intestinales, y que este efecto se observa principalmente en las porciones proximales y medias del intestino delgado. E) Correlaciones

Figura 5. Correlación entre la concentración de glucosa en sangre el día del sacrificio y el promedio del número de células de Paneth en las 4 regiones intestinales estudiadas, de ratas con diabetes (Diab), con diabetes compensada por inyecciones diarias de insulina (Diab+INS) o controles.
0 100 200 300 400 500 600
Glucemia (mg/dL)
100
150
200
250
300
350
célu
las
Pan
eth
/ c
orte
Control Diab Diab + INS
0 100 200 300 400 500 600
Glucemia (mg/dL)
100
150
200
250
300
350
célu
las
Pan
eth
/ c
orte
Control Diab Diab + INS
0 100 200 300 400 500 6000
100
200
300
400
Peso (g)
célu
las
Pan
eth
/ c
orte
Control Diab Diab + INS
0 100 200 300 400 500 6000
100
200
300
400
Peso (g)
célu
las
Pan
eth
/ c
orte
Control Diab Diab + INS
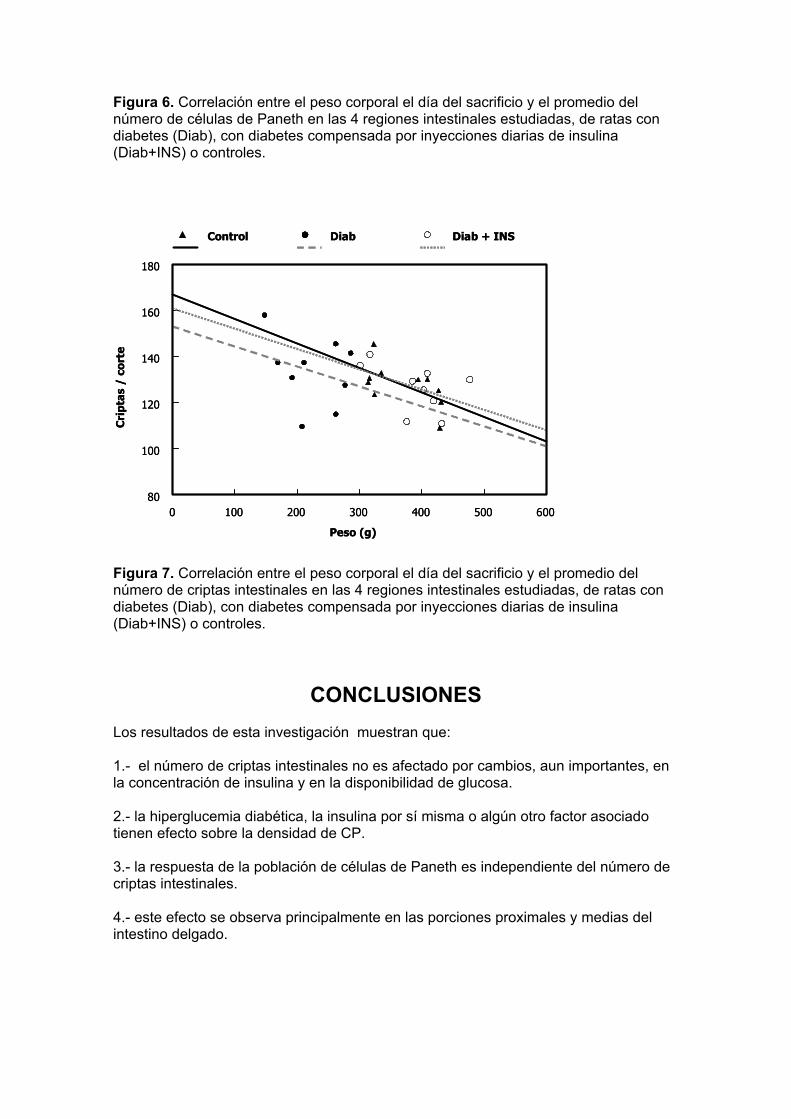
Figura 6. Correlación entre el peso corporal el día del sacrificio y el promedio del número de células de Paneth en las 4 regiones intestinales estudiadas, de ratas con diabetes (Diab), con diabetes compensada por inyecciones diarias de insulina (Diab+INS) o controles. Figura 7. Correlación entre el peso corporal el día del sacrificio y el promedio del número de criptas intestinales en las 4 regiones intestinales estudiadas, de ratas con diabetes (Diab), con diabetes compensada por inyecciones diarias de insulina (Diab+INS) o controles.
CONCLUSIONES Los resultados de esta investigación muestran que: 1.- el número de criptas intestinales no es afectado por cambios, aun importantes, en la concentración de insulina y en la disponibilidad de glucosa. 2.- la hiperglucemia diabética, la insulina por sí misma o algún otro factor asociado tienen efecto sobre la densidad de CP. 3.- la respuesta de la población de células de Paneth es independiente del número de criptas intestinales. 4.- este efecto se observa principalmente en las porciones proximales y medias del intestino delgado.
0 100 200 300 400 500 60080
100
120
140
160
180
Peso (g)
Cri
ptas
/ c
orte
Control Diab Diab + INS
0 100 200 300 400 500 60080
100
120
140
160
180
Peso (g)
Cri
ptas
/ c
orte
Control Diab Diab + INS




![Paneth Colloquium 2008 () 1€¦ · Paneth Colloquium 2008 () 1 [PC2008 #001] Glass spherules related to the El’gygytgyn impact crater (Siberia) Adolph, L.*, Deutsch, A](https://img.pdfslide.net/doc/110x75/5f06df747e708231d41a28bc/paneth-colloquium-2008-1-paneth-colloquium-2008-1-pc2008-001-glass-spherules.jpg)

























