Embed Size (px)
Citation preview

(CANCER RESEARCH (SLJPPL.) 55, 57I4s-5720s, December I, 1995(
Abstract
The in vivo fate of various “In-labeled polypeptides has been thesubject of many investigations. Intracellular metabolism has been StUdiedthrough the use of “In-labeled glycoproteins that are concentrated in thelysosome by receptor-mediated endocytosis. These studies have Indicatedthat the main lysosomal metabolite is “In-chelate-e-lysine, both in vitroand in vivo (Y. Arano et aL, J. NucI. Med., 35: 890—898,1994; F. N.Franano et aL, Nucl. Med. Biol., 21: 1023—1034,1994). Since the vastmajority of radiolabeled antibodies do not localize within the target tissue,an understanding of the metabolism of “In-labeledantibodies in nontarget tissues Is important for the rational design of future radiolabeledantibodies.
We investigated the in vivo metabolism of “11n.DTPA3-conjugatedantibody in female Sprague-Dawley rats using the anticolorectal cardnoma monoclonal antibody (MAb) 1A3 and MAb 1A3-F(ab')2.Livers andkidneys were harvested from rats injected with either intact MAb or MAbfragments and analyzed by gel filtration chromatography. Thirty-five %ofthe radioactivity from “‘In-DTPA-1A3MAb present in the liver was inthe form of a low molecular weight species at 1 through 5 days. Incontrast, @In-DTPA-lA3-F(ab')2 was >98% degraded to a low molecularweight species in the kidney after 1 day. In each case, the low molecularweight metabolites were collected and further analyzed by sifica gelthin-layer chromatography, reversed phase high-performance liquid
chromatography, and ion-exchange chromatography and compared to“In-DTPAand “‘In-DTPA-e-lysinestandards. In each system, the major metabolite co-eluted with “In-DTPA-€-lysine,similar to the resultsobtained with WIn.labeled glycoproteins that are delivered to lysosomes
by receptor-mediated endocytosis. A minor metabolite that was morehighly charged than “In-DTPA was also observed. Analysis of urine andfeces demonstrated that the main excretory product of both 1111n-labeledintact 1A3 and 1A3-F(ab')2 was ‘@In-DTPA-e-lysine. Based on this data,we propose that @In-DTPA-antibodiesare degraded within lysosomes ofnontarget organs such as the liver and kidneys.
Introduction
The identification of in vivo metabolites of antibodies, proteins, andpolypeptides radiolabeled with metal radionuclides is important forunderstanding the mechanisms involved in the uptake and retention ofactivity in nontarget organs. These molecules were initially labeled
with isotopes of iodine (1231, 1251 or 1311);however, conventionallabeling with these isotopes results in poor target retention (1, 2). Thislack of retention most likely results from the rapid egress of theradiolabeled metabolites (3, 4). Antibodies labeled with ‘‘‘In(throughthe use of bifunctional chelates) frequently demonstrate higher uptakeand slower clearance of radioactivity at target sites when compared toradioiodine-labeled molecules. However, I1‘InMAbs3 demonstrate alarge amount of nontarget uptake in the liver and kidneys (5—7).
I Presented at the “Fifth Conference on Radioimmunodetection and Radioimmuno
therapy of Cancer,―October 6—8,1994, Princeton, NJ. This work was funded byDepartment of Energy Grant DE-FGO2-87ER-6O512 (to M. J. W.) and the NIH (GrantCA-42925) with the National Cancer Institute (to M. J. W.).
2 To whom requests for reprints should be addressed, at Division of Radiation Sci
ences, Mallinckrodt Institute of Radiology. 510 South Kingshighway Boulevard, Box8225, St. Louis, MO 631 10.
3 The abbreviations used are: MAb, monoclonal antibody; DTPA, diethylenetriamine
pentaacetic acid; cDTPAA, diethylenetriaminepentaacetic dianhydride; TLC, thin-layerchromatography; RP-HPLC, reverse phase high pressure liquid chromatography.
Different investigators have varied the bifunctional chelates andchemical linkers between the radiometal and protein in attempts todecrease radiometal accumulation in nontarget tissues. The success ofthese efforts has been limited (8—11). These results reflect, in part, ouruncertainty as to what metabolites are produced in vivo. By determining the structures of the various metabolites, we hope to learn themechanisms of radiolabel retention as well as develop possible methods of circumventing the high hepatic and renal background of ‘@ ‘Inlabeled molecules.
Much data implicate lysosomes in the metabolism of MAbs at both
target and nontarget sites. Several studies have shown that radiolabeled antibodies bound to cell surface antigens are internalized anddelivered to the lysosome for degradation (4, 12—14).For nontargetedantibodies, the evidence is more indirect. Intact antibodies and theirfragments could be internalized and delivered to lysosomes via threedifferent endocytic mechanisms: absorptive endocytosis, fluid phaseendocytosis, and receptor-mediated endocytosis. Absorptive endocytosis is exemplified by the renal tubular reabsorption of proteins (15).Fluid phase endocytosis is well studied in the liver, where hepatocytesinternalize 20% of their volume per hour, and 18% of that volume istransported to intracellular storage compartments such as lysosomes(16). Receptor-mediated endocytosis of antibody-antibody and antibody-antigen complexes by the Fc receptor could also deliver radiolabeled antibodies to hepatic lysosomes (17—19).
Recent studies with ‘‘‘In-chelate-glycoproteinhave shown thatthese molecules are delivered to lysosomes by receptor-mediatedendocytosis (20, 21). Once in lysosomes, the glycoprotein polypeptidebackbone is rapidly degraded, yielding low molecular weight radiolabeled metabolites, and these metabolites are retained within thelysosome (21). Studies with several different bifunctional chelateshave found that the predominant metabolites are of the general structure, ‘‘‘In-chelate-lysine (20, 22). This result indicates that the chelate-lysine bond is inefficiently hydrolyzed within the lysosome, andthe ‘‘‘In-chelatebond is also stable in the acidic environment of thelysosome.
Prior studies on the metabolism of ‘@ ‘In-chelate-antibodies havefound that they were degraded to low molecular weight metabolites(23, 24). This result, together with the above data implicating lysosomes in antibody metabolism and the identification of metabolitesproduced by lysosomal metabolism, suggested that ‘‘‘In-DTPA-antibodies would be degraded to ‘‘‘In-DTPA-€-lysine.Therefore, westudied the metabolism of the lA3 MAb and its F(ab')2 fragments.This antibody is specific for a human colorectal carcinoma antigenand thus should behave as a nontargeted antibody in the rat. Wecompared the radiolabeled metabolites prepared from urine, feces,liver extracts, and renal homogenates to a series of standards including1 1 ‘In-DTPA-€-lysine.
Prior studies on the metabolism of nontargeted antibodies havefound that the radiolabel was recovered in a low molecular weight
form. Metabolism studies of antibodies labeled with ‘‘‘In have mdicated that they are catabolized to small molecular weight compoundsin the liver and kidneys (25—27).Studies with other radiometals have
57l4s
Identification of Metabolites of “In-Diethylenetriaminepentaacetic Acid..Monoclonal Antibodies and Antibody Fragments in Vivo'
Buck E. Rogers, F. Nicholas Franano, James R. Duncan, W. Barry Edwards, Carolyn J. Anderson,Judith M. Connett, and Michael J. Welch2
The Mallinckrodz Institute of Radiology [B. E. R., F. N. F., J. R. D.. W. B. E., C. J. A., M. I. W.J and Department of Surgery (J. M. C.], Washington University, School ofMedicine, St. Louis, Missouri 63110
on May 28, 2020. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

METABOLISM OF ‘In-LABELEDMAbs
identified radiometal-chelate-lysine metabolites in the urine after administration of whole antibody and antibody fragments (28—30).
The ‘‘‘In complex and the amide bond formed when the chelate islinked to the c-amine of lysine are both stable to the low pH of thelysosome and to lysosomal hydrolases (24, 26, 31—35).If ‘‘‘Indissociates from the complex, ‘‘‘Inreassociation with the chelate isunlikely because of the low concentrations occurring with carrier-freeradionuclides. If decomplexation occurred, most likely the@ 1‘Inwould bind some other protein. This other protein could be observedby size-exclusion chromatography and would likely have a biodistribution pattern which differed from chelate-associated ‘‘‘In.If theamide bond joining the chelate to the protein was not stable, ‘@ ‘Inlabeled chelate-amino acid(s) would not have been seen in previousstudies (24, 26, 27).
Intracellular metabolism of ‘‘‘In-labeledpolypeptides has beenlargely attributed to internalization and degradation within lysosomes.These polypeptides are delivered to the lysosome through receptormediated endocytosis (36, 37). A single receptor can deliver manypolypeptide molecules to the lysosome, where the polypeptide iscatabolized to amino acids in a low pH environment (33—35).Throughthe use of glycoproteins targeted to cell surface receptors in the liver,
it has been shown that ‘‘‘In-DTPA-labeled polypeptides are hydro
lyzed to small molecular weight compounds in the lysosome that areslowly released from the cell (21 ). Similar studies have identified
the small molecular weight compound as ‘‘‘In-chelate-€-lysine(20,22,38).
We were interested in identifying the exact structure of the majormetabolite produced when whole antibody and antibody fragmentslabeled with ‘‘‘Inwere administered in vivo and seeing if this structure was consistent with the metabolite identified in the glycoproteinstudies. MAb lA3, an anticolorectal carcinoma MAb (39), was labeled with ‘‘‘Inthrough the use of the bifunctional chelate DTPA.DTPA was conjugated to the c-amino groups of lysines available inboth MAb 1A3 and MAb 1A3-F(ab')2 to form amide linkages. Thelabeled conjugates were administered to mature, female SpragueDawley rats, and metabolites were isolated from the liver, kidneys,urine, and feces at various time points. Because ‘‘‘In-DTPA-€-lysinewas identified as the major metabolite in the glycoprotein studies, wesynthesized ‘‘‘In-DTPA-€-lysineas a standard. The isolated metabolites were then compared with ‘‘‘In-DTPA-€-lysine,‘‘‘In-DTPA,and ‘@@ In-acetate standards.
Materials and Methods
Materials. DTPA and the cyclic anhydride of cDTPAA were purchasedfrom Sigma Chemical Co. (St. Louis, MO). DTPA-€-lysine was synthesized asdescribed previously (22). RP-HPLC column (201 HS 104; 4.6 X 250 mm,C,8) was from Vydac (Hesperia, CA). Silica gel 60 F-254 TLC plates werefrom EM Science (Gibbstown, NJ). Chelex-100 was from Bio-Rad (Richmond,CA). Centricon-30 concentrators were purchased from Amicon, Inc. (Beverly,MA). Ultrafree-MC 0.22 @mfilter units were from Millipore (Bedford, MA).Bio-Spin 6 chromatography columns were from Bio-Rad Laboratories(Hercules, CA). A Tekmar tissue homogenizer (Cincinnati, OH), a Branson
Sonifier 185 cell disrupter, and a Sorvall RC2-B centrifuge were used for themetabolism experiments. ‘‘‘InCl3(4.2 X l0@Ci/g) was provided by Mallinckrodt, Inc. (St. Louis, MO). Strong cation exchange column (Mono Q HR 5/5)and Superose 12 size exclusion column were from Pharmacia Biotech, Inc.(Piscataway, NJ). MAbs lA3 and lA3-F(ab')2 were purified from serum-freemedium by Invitron (St. Louis, MO) using proprietary methods. Mature,female Sprague-Dawley rats were from Sasco (Omaha, NE). All animalexperiments were in compliance with guidelines specified by the WashingtonUniversity Animal Studies Committee.
The Mono Q column was used with a Spectra-Physics SP8700XR HPLCpump. All ion exchange chromatography used an isocratic eluant of 100%bufferA for5 mm, and then a gradientwas startedwhich went from 100%
buffer A to 0% buffer A over the next 20 mm (A = 0.0183 M NH4OAc,pH = 9.2; B = 1.0 MNH4OAc,pH = 9.2). Flow rate for this system was 1.0mi/mm, with each fraction collected for 24 s. RP-HPLC was performed witha Waters 600E system controller from Millipore. The RP system used anisocratic run of 99% 100 mM NaOAc, 150 mist NaCl, pH 5.5, with 1%
methanol at 1.0 mi/mm, collecting 2-drop fractions. TLC plates were elutedwith 1:1 methanol:lO% NH4OAc in water and analyzed using a BioscanSystem 200 imaging scanner for detecting radioactive peaks. Fast proteinliquid chromatography was performed on a Pharmacia/LKB system using aSuperose 12 gel filtration column with 20 mM HEPES-300 nmi NaC1, pH 7.3,at a flow rate of 0.4 mI/mm, and with fractions collected every minute. Allradioactive fractions were counted on a Beckman Gamma 8000 automatedwell-type gamma counter.
DTPA Conjugation to 1A3 and 1A3-(Fab')2. The intact MAb lA3 and its(Fab')2 fragments were diluted to 50—65mMin 0.1 MNH4OAc, pH 5.5, andthen added to a 10-fold molar excess of cDTPAA and mixed at room temperattire for one h. The reaction mixtures were then purified by gel filtrationchromatography, and the appropriate fractions were collected and concentrated
using a Centricon-30 concentrator. The DTPA-1A3 and DTPA-1A3-(Fab')2conjugates were stored at —80°Cuntil ready for use.
Radiolabeling. ‘‘‘In-acetate complexes were prepared by diluting ‘@ â€mCI3
in 0.4 MNaOAc, pH 5.5. ‘‘‘In-DTPA,‘‘‘In-DTPA-E-lysine,‘‘‘In-DTPA-lA3,and ‘‘‘In-DTPA-1A3-F(ab')2were prepared by incubating the standards orconjugates with@ ‘‘In-acetatefor 1 h at room temperature. Free metal wasremoved from ‘‘‘In-DTPAand ‘‘‘In-DTPA-€-lysinecomplexes by adding 200mg of Chelex-100 metal binding resin for 50 mm and filtering through a 0.22pm spin filter. Uncomplexed ‘‘‘In(ffl)was removed from ‘‘‘In-DTPA-1A3
and ‘‘‘In-DTPA-lA3-F(ab')2 by gel filtration using a Bio-Spin 6 column thathad been equilibrated with 0.4 M NaOAc, pH 5.5, according to the manufacturer's instructions. Radiochemical purity was determined using silica gelradio-TLC for the small molecular weight standards and gel filtration for theantibody conjugates. ‘‘‘In-DTPAand ‘‘‘In-DTPA-€-lysineshowed singleradioactive peaks at R,.s of 0.56 and 0.40, respectively, by TLC with 1:1methanol:10% NH4OAc in water as the eluant. Gel filtration of ‘‘‘In-DTPA1A3 and ‘‘‘In-DTPA-1A3-F(ab')2 demonstrated a single radioactive peak foreach sample.
In Vivo Metabolism of “In-DTPA-1A3 and “In-DTPA-1A3-F(ab')2.Mature, female Sprague-Dawley rats were injected with 500—1200 @Ciof1@ ‘In-DTPA-1A3 (specific activity, 4.9—6.7 @Ci4.@g) or â€â€â€˜In-DTPA-lA3-
F(ab')2 (specific activity, 2.3—5.4pCi4.tg). Urine and feces were collectedfrom 72—96h for intact antibody and 48—72h for F(ab')2 fragments. The urinewas then passed through a 0.2 @mfilter and either directly analyzed, orconcentrated and analyzed by TLC, RP-HPLC, and anion exchange chromatography. Feces were suspended in an equal volume ofO.l MNH4OAc, pH 5.5,homogenized, and sonicated with a Branson Sonifier 185 (3 mm at power4). The suspension was clarified by centrifugation (23,500 X g for 60 mm),and the supernatant was passed through a 0.2 @.tmfilter. The fecal sample wasconcentrated to about 500 pAusing a Labconco Centrivap Concentrator beforeanalysis by RP-HPLC. Livers were removed from rats injected with “In
DTPA-lA3, and kidneys were removed from rats injected with ‘‘‘In-DTPA1A3-F(ab')2at l, 3-, and 5-day time points. The organs were rinsed with Waterto remove as much blood as possible, homogenized in an equal volume oflabeling buffer using a Tekmar tissue homogenizer, and somcated (3 mmnatpower = 4). The samples were clarified by centrifugation (23,500 X g for 60minutes), and the supernatant was passed through a 0.22 @mspin filter. A
0.2-ml sample was analyzed by gel filtration. Fractions were collected and
counted on an automated gamma counter. Low molecular weight fractionswere further analyzed by TLC, RP-HPLC, and anion exchange HPLC, asdescribed above.
Results
Gel Filtration Chromatography of Liver and Kidney Metabolites. Since prior studies indicated that significant amounts of the1@ ‘In radiolabel from â€â€â€˜In-1A3 MAb accumulated within the liver
(23, 24, 27), livers from Sprague-Dawley rats injected with “InDTPA-1A3 were harvested at 1, 3, and 5 days after injection. Between
8—10%of the injected dose was recovered in the liver at these timepoints. The livers were then processed, and metabolites were analyzed
5715s
on May 28, 2020. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
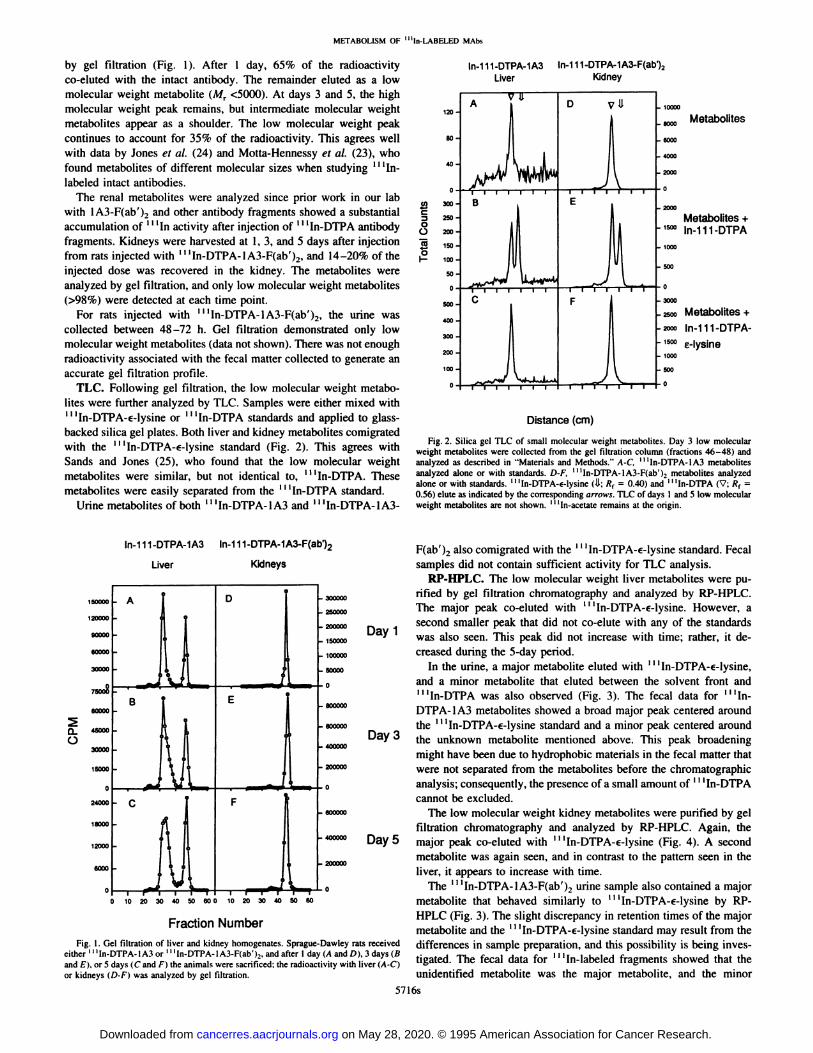
METABOLISM OF “In-LABELED MAbs
by gel filtration (Fig. 1). After 1 day, 65% of the radioactivityco-eluted with the intact antibody. The remainder eluted as a lowmolecular weight metabolite (Mr <5000). At days 3 and 5, the highmolecular weight peak remains, but intermediate molecular weightmetabolites appear as a shoulder. The low molecular weight peakcontinues to account for 35% of the radioactivity. This agrees wellwith data by Jones et a!. (24) and Motta-Hennessy et a!. (23), whofound metabolites of different molecular sizes when studying ‘‘‘Inlabeled intact antibodies.
The renal metabolites were analyzed since prior work in our labwith 1A3-F(ab')2 and other antibody fragments showed a substantialaccumulation of ‘‘‘Inactivity after injection of ‘‘‘In-DTPAantibodyfragments. Kidneys were harvested at 1, 3, and 5 days after injectionfrom rats injected with ‘‘‘In-DTPA-lA3-F(ab')2, and 14—20%of theinjected dose was recovered in the kidney. The metabolites wereanalyzed by gel filtration, and only low molecular weight metabolites(>98%) were detected at each time point.
For rats injected with ‘‘‘In-DTPA-1A3-F(ab')2, the urine wascollected between 48—72h. Gel filtration demonstrated only lowmolecular weight metabolites (data not shown). There was not enoughradioactivity associated with the fecal mauer collected to generate anaccurate gel filtration profile.
TLC. Following gel filtration,the low molecularweight metabolites were further analyzed by TLC. Samples were either mixed withI I ‘In-DTPA-€-lysine or â€â€â€˜In-DTPA standards and applied to glass
backed silica gel plates. Both liver and kidney metabolites comigratedwith the ‘‘‘In-DTPA-€-lysinestandard (Fig. 2). This agrees withSands and Jones (25), who found that the low molecular weightmetabolites were similar, but not identical to, ‘‘‘In-DTPA.Thesemetabolites were easily separated from the ‘‘‘In-DTPAstandard.
Urine metabolites of both ‘‘‘In-DTPA-1A3and ‘‘‘In-DTPA-1A3-
In-I I 1-DTPA-1A3 In-I I 1-DTPA-1A3-F(ab')2
Liver Kidneys
In-I I 1-DTPA-1A3 In-i I 1-DTPA-1A3-F(ab@2Liver Kidney
j@D
U I I I I ! I I
120
80
40
U) 3@
@2500C.) 2@
@ 1500I— 100
50
500
400
300
200
100
.10000
.e000 Metabobtes
.6000
.4000
.2000
.2000
Metabolites +.1500 ln-111-DTPA
.1000
.500
Metabolites +
In-i 11-DTPA
c-Iysine
.2000
.2500
.2000
. 1500
.1000
.500
Distance (cm)
Fig. 2. Silica gel TLC of small molecular weight metabolites. Day 3 low molecularweight metabolites were collected from the gel filtration column (fractions 46—48)andanalyzed as described in “Materialsand Methods.―A-C, ‘I‘In-DTPA-1A3metabolitesanalyzed alone or with standards. D-F, ‘‘‘In-DTPA-lA3-F(ab')2metabolites analyzedalone or with standards. ‘‘‘In-DTPA-€-lysine(U; R1 = 0.40) and ‘‘‘In-DTPA(V; R1 =0.56) elute as indicated by the corresponding arrows. TLC of days 1 and 5 low molecularweight metabolites are not shown. ‘‘‘In-acetateremains at the origin.
F(ab')2 also comigrated with the ‘‘‘In-DTPA-€-lysinestandard. Fecalsamples did not contain sufficient activity for TLC analysis.
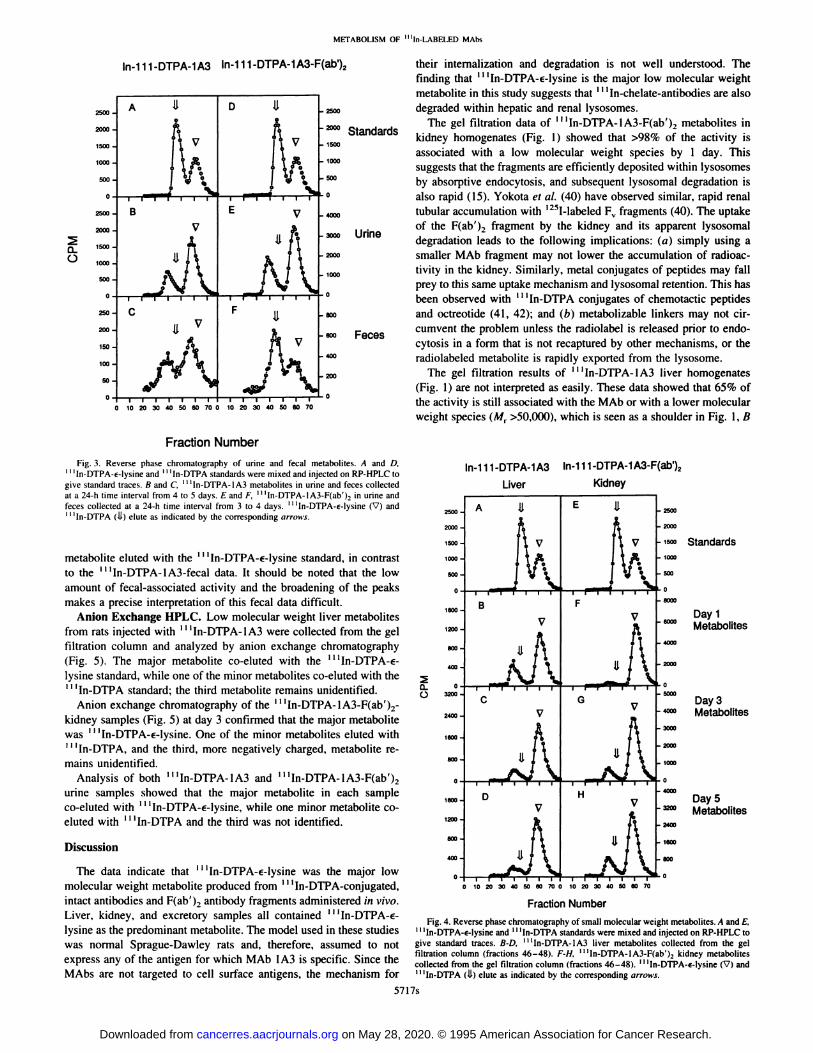
RP-HPLC. The low molecular weight liver metabolites were pu200000 rifled by gel filtration chromatography and analyzed by RP-HPLC.
250000 The major peak co-eluted with â€â€â€˜In-DTPA-€-lysine. However, a
200000 r@@ second smaller peak that did not co-elute with any of the standards
150000 @ay was also seen. This peak did not increase with time; rather, it de
100000 creased during the 5-day period.
50000 In the urine, a major metabolite eluted with â€â€â€˜In-DTPA-€-lysine,
and a minor metabolite that eluted between the solvent front andI I ‘In-DTPA was also observed (Fig. 3). The fecal data for â€â€œIn
@ 600000 DTPA-1A3 metabolites showed a broad major peak centered around
@ 600000 the â€â€â€˜In-DTPA-€-lysine standard and a minor peak centered around
0 400000 Day 3 the unknown metabolite mentioned above. This peak broadening
might have been due to hydrophobic materials in the fecal matter that200000 were not separated from the metabolites before the chromatographic
0 analysis; consequently, the presence of a small amount of ‘@ ‘In-DTPA
cannot be excluded.600000 The low molecular weight kidney metabolites were purified by gel
filtration chromatography and analyzed by RP-HPLC. Again, the400000 Day 5 major peak co-eluted with â€â€â€˜In-DTPA-€-lysine (Fig. 4). A second
metabolite was again seen and in contrast to the pattern seen in the200000
liver, @tappears to increase with time.0 The ‘@ ‘In-DTPA-lA3-F(ab')2 urine sample also contained a major
metabolite that behaved similarly to ‘‘‘In-DTPA-€-lysineby RPHPLC (Fig. 3). The slight discrepancy in retention times of the major
Fraction Number metaboliteandthe ‘‘‘In-DTPA-€-lysinestandardmayresultfrom theFig. I. Gel filtration of liver and kidney homogenates. Sprague-Dawley rats received differences in sample preparation, and this possibility is being inves
either ‘‘‘In-DTPA-1A3or ‘‘‘ln-DTPA-lA3-F(ab'),, and after I day (A and D), 3 days (B . j @‘and E), or 5 days (C and F) the animals were sacrificed; the radioactivity with liver (A-C) ligated. The fecal data for In-labeled fragments showed that theor kidneys (D-F) was analyzed by gel filtration. unidentified metabolite was the major metabolite, and the minor
5716s
on May 28, 2020. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
METABOLISM OF “11n-LABELEDMAbs
In-i 11-DTPA-1A3-F(ab')2 their internalization and degradation is not well understood. Thefinding that ‘‘‘In-DTPA-€-lysineis the major low molecular weightmetabolite in this study suggests that ‘‘‘In-chelate-antibodies are alsodegraded within hepatic and renal lysosomes.
The gel filtration data of ‘‘‘In-DTPA-lA3-F(ab')2 metabolites inkidney homogenates (Fig. 1) showed that >98% of the activity isassociated with a low molecular weight species by 1 day. Thissuggests that the fragments are efficiently deposited within lysosomesby absorptive endocytosis, and subsequent lysosomal degradation isalso rapid (15). Yokota et a!. (40) have observed similar, rapid renaltubular accumulation with ‘251-labeledF@fragments (40). The uptakeof the F(ab')2 fragment by the kidney and its apparent lysosomaldegradation leads to the following implications: (a) simply using asmaller MAb fragment may not lower the accumulation of radioactivity in the kidney. Similarly, metal conjugates of peptides may fallprey to this same uptake mechanism and lysosomal retention. This hasbeen observed with ‘‘‘In-DTPAconjugates of chemotactic peptidesand octreotide (41, 42); and (b) metabolizable linkers may not circumvent the problem unless the radiolabel is released prior to endocytosis in a form that is not recaptured by other mechanisms, or theradiolabeled metabolite is rapidly exported from the lysosome.
The gel filtration results of ‘‘‘In-DTPA-lA3 liver homogenates(Fig. 1) are not interpreted as easily. These data showed that 65% ofthe activity is still associated with the MAb or with a lower molecularweight species (Mr >50,000), which is seen as a shoulder in Fig. 1, B
In-I 11-DTPA-1A3
.2500
. 2000 Standards
.1500
.1000
.500
.4000
.2000 Urine
.2000
.1000
.600
.600 Feces
.400
.200
B
2500.
2000.
1500.
1000.
500.
2500.
0@
500.
250.
200.
150.
100.
50.
C
10 20 30 40 50 60 700 10 20 30 40 @060 70
FractionNumber
metabolite eluted with the ‘‘‘In-DTPA-€-lysinestandard, in contrastto the ‘‘‘In-DTPA-lA3-fecal data. It should be noted that the lowamount of fecal-associated activity and the broadening of the peaksmakes a precise interpretation of this fecal data difficult.
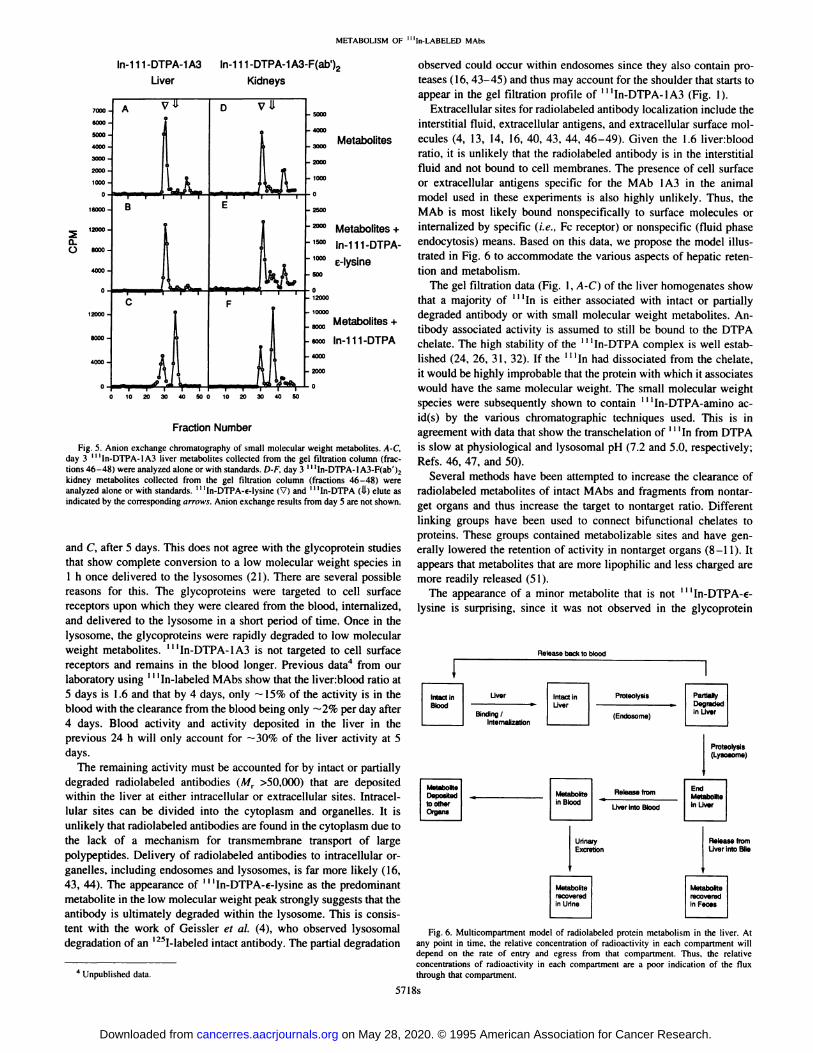
Anion Exchange HPLC. Low molecularweight liver metabolitesfrom rats injected with ‘‘‘In-DTPA-1A3were collected from the gelfiltration column and analyzed by anion exchange chromatography(Fig. 5). The major metabolite co-eluted with the ‘@ ‘In-DTPA-€lysine standard, while one of the minor metabolites co-eluted with the
I I ‘In-DTPA standard; the third metabolite remains unidentified.
Anion exchange chromatography of the ‘‘‘In-DTPA-1A3-F(ab')2-kidney samples (Fig. 5) at day 3 confirmed that the major metabolitewas ‘‘‘In-DTPA-€-lysine.One of the minor metabolites eluted withI I ‘ln-DTPA, and the third, more negatively charged, metabolite re
mains unidentified.Analysis of both ‘‘‘In-DTPA-lA3 and ‘‘‘In-DTPA-lA3-F(ab')2
urine samples showed that the major metabolite in each sampleco-eluted with ‘‘‘In-DTPA-€-lysine,while one minor metabolite coeluted with ‘‘‘In-DTPAand the third was not identified.
Discussion
Fig. 3. Reverse phase chromatography of urine and fecal metabolites. A and D,â€â€â€˜In-DTPA-€-lysine and â€â€â€˜In-DTPA standards were mixed and injected on RP-HPLC to
give standard traces. B and C, ‘‘‘In-DTPA-1A3metabolites in urine and feces collectedat a 24-h time interval from 4 to 5 days. E and F, “In-DTPA-IA3-F(ab')2in urine andfeces collected at a 24-h time interval from 3 to 4 days. ‘‘‘In-DTPA-€-lysine(V) andâ€â€â€˜In-DTPA (i.L) elute as indicated by the corresponding arrows.
In-i 11-DTPA-1A3
Liver
In-i 11-DTPA-1A3-F(ab')2
Kidney
Standards
Day 1Metabolites
Day3Metabolites
Day 5Metabolites
0@0
10203040506070010203040506070
The data indicate that ‘‘‘In-DTPA-€-lysinewas the major lowmolecular weight metabolite produced from ‘‘‘In-DTPA-conjugated,intact antibodies and F(ab')2 antibody fragments administered in vivo. Fraction Number
Liver, kidney, and excretory samples all contained@ ‘‘In-DTPA-€- Fig.4. Reversephasechromatographyofsmallmolecularweightmetabolites.AandE,lysine as the predominant metabolite. The model used in these studies ‘I‘In-DTPA-€-lysineand‘‘‘In-DTPAstandardsweremixedandinjectedonRP-HPLCtowas normal Sprague-Dawley rats and, therefore, assumed to not give standardtraces,B-D, ‘‘‘In-DTPA-lA3liver metabolitescollectedfrom the gel
filtration column (fractions 46—48). F-H, ‘‘‘In-DTPA-lA3-F(ab')2 kidney metabolitesexpress any of the antigen for which MAb lA3 is specific. Since the collectedfromthegel filtrationcolumn(fractions46—48).‘‘‘In-DTPA-s-lysine(V)andMAbs are not targeted to cell surface antigens, the mechanism for 1I‘In-D'rpA(ii) eluteas indicatedby thecorrespondingarrows.
5717s
A
tv
I@—I'
D U
0
I @i•@'@@T@ I
E
F
on May 28, 2020. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
observed could occur within endosomes since they also contain proteases (16, 43—45)and thus may account for the shoulder that starts toappear in the gel filtration profile of ‘‘‘In-DTPA-1A3(Fig. 1).
Extracellular sites for radiolabeled antibody localization include theinterstitial fluid, extracellular antigens, and extracellular surface mol
Metabolites ecules (4, 13, 14, 16, 40, 43, 44, 46—49). Given the 1.6 liver:bloodratio, it is unlikely that the radiolabeled antibody is in the interstitialfluid and not bound to cell membranes. The presence of cell surfaceor extracellular antigens specific for the MAb lA3 in the animalmodel used in these experiments is also highly unlikely. Thus, theMAb is most likely bound nonspecifically to surface molecules or
Metabolites + internalized by specific (i.e., Fc receptor) or nonspecific (fluid phaseIn-i11-DTPA- endocytosis) means. Based on this data, we propose the model illusc-lysine tratedin Fig. 6 to accommodatethe variousaspects of hepatic reten
tion and metabolism.The gel filtration data (Fig. 1, A-C) of the liver homogenates show
that a majority of ‘‘‘Inis either associated with intact or partiallydegraded antibody or with small molecular weight metabolites. Antibody associated activity is assumed to still be bound to the DTPAchelate. The high stability of the ‘‘‘In-DTPAcomplex is well established (24, 26, 3 1, 32). If the II‘Inhad dissociated from the chelate,it would be highly improbable that the protein with which it associates
would have the same molecular weight. The small molecular weightspecies were subsequently shown to contain ‘‘‘In-DTPA-amino acid(s) by the various chromatographic techniques used. This is in
agreement with data that show the transchelation of ‘‘‘Infrom DTPAis slow at physiological and lysosomal pH (7.2 and 5.0, respectively;Refs. 46, 47, and 50).
Several methods have been attempted to increase the clearance ofradiolabeled metabolites of intact MAbs and fragments from nontarget organs and thus increase the target to nontarget ratio. Differentlinking groups have been used to connect bifunctional chelates toproteins. These groups contained metabolizable sites and have generally lowered the retention of activity in nontarget organs (8—11). Itappears that metabolites that are more lipophilic and less charged aremore readily released (51).
The appearance of a minor metabolite that is not ‘‘‘In-DTPA-€lysine is surprising, since it was not observed in the glycoprotein
Proteotysis(Lysosome)
MetaboSe EndDeposIted@ Metabolte@ Release from MetabolIc@ InBbodLIverintoBlood@ Liver
Uñnaiy Release fromExcretion Uverinto Sic
MetabolIcmcovemdIn Feces
Fig. 6. Multicompartment model of radiolabeled protein metabolism in the liver. Atany point in time, the relative concentration of radioactivity in each compartment willdepend on the rate of entry and egress from that compartment. Thus, the relativeconcentrations of radioactivity in each compartment are a poor indication of the fluxthrough that compartment.
Metabolites +
In-i 11-DTPA
METABOLISM OF “In-LABELED MAbs
In-I 11-DTPA-1A3 In-i 11-DTPA-1A3-F(ab')2
Liver Kidneys
0@0
Fig. 5. Anion exchange chromatography of small molecular weight metabolites. A-C,day 3 ‘‘‘In-DTPA-lA3liver metabolites collected from the gel filtration column (fractions 46—48)were analyzed alone or with standards. D-F, day 3 ‘‘‘In-DTPA-1A3-F(ab')2kidney metabolites collected from the gel filtration column (fractions 46—48)wereanalyzed alone or with standards. ‘‘‘In-DTPA-€-lysine(V) and ‘‘‘In-DTPA(1.1)elute asindicated by the corresponding arrows. Anion exchange results from day 5 are not shown.
Fraction Number
and C, after S days. This does not agree with the glycoprotein studiesthat show complete conversion to a low molecular weight species in1 h once delivered to the lysosomes (21). There are several possiblereasons for this. The glycoproteins were targeted to cell surfacereceptors upon which they were cleared from the blood, internalized,and delivered to the lysosome in a short period of time. Once in thelysosome, the glycoproteins were rapidly degraded to low molecularweight metabolites. ‘‘‘In-DTPA-lA3 is not targeted to cell surfacereceptors and remains in the blood longer. Previous data4 from ourlaboratory using ‘‘‘In-labeledMAbs show that the liver:blood ratio at5 days is 1.6 and that by 4 days, only —15% of the activity is in the
blood with the clearance from the blood being only —2%per day after4 days. Blood activity and activity deposited in the liver in theprevious 24 h will only account for —30%of the liver activity at 5days.
The remaining activity must be accounted for by intact or partiallydegraded radiolabeled antibodies (Mr >50,000) that are depositedwithin the liver at either intracellular or extracellular sites. Intracellular sites can be divided into the cytoplasm and organelles. It isunlikely that radiolabeled antibodies are found in the cytoplasm due tothe lack of a mechanism for transmembrane transport of largepolypeptides. Delivery of radiolabeled antibodies to intracellular organelles, including endosomes and lysosomes, is far more likely (16,43, 44). The appearance of ‘‘‘In-DTPA-€-lysineas the predominantmetabolite in the low molecular weight peak strongly suggests that theantibody is ultimately degraded within the lysosome. This is consistent with the work of Geissler et a!. (4), who observed lysosomaldegradation of an ‘251-labeledintact antibody. The partial degradation
Releasebackto blood
Liver
Bfrdng/Internalization
Proteotyals
(Endosome)
4 Unpublished data.
57l8s
Intact InBlood
Intact inLiver
PartiallyDe@edIn Liver
Metabolite
in Urine
on May 28, 2020. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

METABOLISM OF ‘1'In-LABELEDMAIn
metabolism studies of Franano et al. (22). We have shown that the
minor metabolite is not ‘‘‘In-DTPA-€-lysine, I I ‘In-DTPA, or free
I I ‘In-acetate by comparison to known standards (Figs. 2—5). In addi
tion, anion exchange chromatography shows this minor metaboliteeluting later than ‘‘‘In-DTPA-€-lysine and ‘‘‘In-DTPA, which mdi
cates a compound with a greater negative charge (Fig. 5). Franano et
a!. (22) show that ‘‘‘In-DTPA-€-lysinewas retained in the kidney
more than ‘‘‘In-DTPAat 1 h after injection (22). This is explained bythe more positively charged ‘‘‘In-DTPA-€-lysinebeing bound to thelumenal surfaces of proximal tubular cells in the kidney. Therefore,the higher concentration of the more negative minor metabolite in thekidney over time appears to be a function of the metabolism within thekidney and not due to kidney uptake after metabolism at another site.
The structure of this more negatively charged minor metabolite hasnot been determined, but there are several possibilities that mayexplain it. It could be the product of further metabolism of “InDTPA-€-lysine; however, this is unlikely due to the decrease in itssize in the liver RP-HPLC traces (Fig. 4). The ‘‘‘Incould be released
from the chelate and then to some other small molecule. This isunlikely given the strength of the ‘‘‘In-chelatebond (24, 26, 31, 32).Finally, the unknown metabolite could be the result of@ IIIn-DTPAbound to an amino acid other than lysine. Under the conjugation
conditions, it is unlikely that available hydroxyl groups are reactivewith cDTPAA. Also, any available sulfhydryl groups on ‘‘‘In-DTPA
lA3-F(ab')2 were alkylated in the process of making lA3-F(ab')2from 1A3, leaving them unreactive towards cDTPAA. The formationof a sulfhydryl coordinated species is, therefore, excluded. It is mostlikely that cDTPAA reacted to some extent with an NFI2-terminal
amino acid. The results of Rana and Meares (52) showed thatNH,-termini of Lym-l were one possible site of attachment of
SCN-chelates. Some of the DTPA may have been conjugated to the
NH2-termini of lA3 and lA3-F(ab'),, which would account for another low molecular weight metabolite.
From this discussion, it is clear that radiolabeled antibody metabolism encompasses a wide variety of issues. Fundamentally, the
process is dynamic. The observed levels of various intermediates are
governed by the rate constants of each step (Fig. 6). To begin deciphering this complex biochemical pathway, we must identify the
intermediates and start fitting them into the known biochemical pathways. Then we can begin varying the radiolabel-antibody linkages toimprove target to background ratios.
Note Added in Proof
Since this paper was submitted, our group has shown by NH2-terminal sequencing of lA3 and 1A3-F(ab')2 that the NH2-terminal amino acid on lA3 and
lA3-F(ab'), is aspartic acid. This agrees with our hypothesis that a fraction of the
cDTPAA chelates were conjugated to the N1-12-terminalamine availableon theantibody. The unknown metabolite is less lipophilic and more negatively charged
than ‘‘‘In-DTPA-€-lysine. An ‘‘‘In-DTPA-aspartic acid metabolite is consistent
with both NW-terminal sequencing and the chromatographic results.
Acknowledgments
We thank Henry Lee and Elizabeth Sherman for their excellent technicalassistance. We also thank the Protein and Nucleic Acid Chemistry Laboratory
at Washington University in St. Louis for their sequence analysis on lA3 and
lA3-F(ab'),.
References
1. Scheinberg, D. A., and Strand, M. Leukemic cell targeting and therapy by monoclonal
antibody in a mouse model system. Cancer Res., 42: 44—49,1982.2. Fritzberg, A. R., Beminger, R. W., Hadley, S. W., and Wester, D. W. Approaches to
radiolabeling of antibodies for diagnosis and therapy of cancer. Pharmaceut. Res., 5:325—334,1988.
3. LaBadie, J. H., Chapman, K. P., and Aronson, N. N. Glycoprotein catabolism in rat
liver. Lysosomal digestion of iodinated asialo-fetuin. Biochem. J., 152: 27 1—279,1975.
4. Geissler, F., Anderson, S. K., and Press, 0. Intracellular catabolism of radiolabeledanti-CD3 antibodies by leukemic T cells. Cell. Immunol., 137: 96—110, 1991.
5. P1mm, M. V., Perkins, A. C., and Baldwin, R. W. Differences in tumour and normal
tissue concentrations of iodine- and indium-labelled monoclonal antibody. Eur. J.Nucl. Med., 11: 300—304, 1985.
6. Otsuka,F.L., Fleischman,J.B.,andWelch,M. J.Comparativestudiesusing1251and‘I ‘In-labeled monoclonal antibodies. Int. J. Radiat. AppI. Part B NucI. Med. Biol., 13:
325—334,1986.7. Andrew, S. M., Perkins, A. C., Pimm, M. V., and Baldwin, R. W. A comparison of
iodine and indium labelled anti CEA intact antibody, F(ab)2 and Fab fragments byimaging tumour xenografts. Eur. J. NucI. Med., 13: 598—604, 1988.
8. Meares, C. F., McCall, M. J., Deshpande, S. V., DeNardo, S. J., and Goodwin, D. A.Chelate radiochemist,'y:cleavable linkers lead to altered levels of radioactivity in theliver. Int. J. Cancer, 2: 99—102, 1988.
9. Paik, C. H., Yokoyama, K., Reynolds, J. C., Quadri, S. M., Mm, C. Y., Shin, S. Y.,Maloney, P. J., Larson, S. M., and Reba, R. C. Reduction of background activities byintroduction of a diester linkage between antibody and a chelate in radioimmunodetection of tumor. J. NucI. Med., 30: 1693—1701,1989.
10. Deshpande, S. V., DeNardo, S. J., Meares, C. F., McCall, M. J., Adams, G. P., andDeNardo, G. L. Effect of different linkages between chelates and monoclonal antibodies on levels of radioactivity in the liver. NucI. Med. Biol., 16: 587—597,1989.
11. Weber, R. W., Boutin, R. H., Nedelman, M. A., James, J. L., and Dean, R. T.Enhanced kidney clearance with ester-linked Tc99m-radiolabeled antibody Fab'chelator conjugate. Bioconj. Chem., 1: 431—437, 1990.
12. Hopkins, C, R., and Trowbridge, I. S. Intemalization and processing oftransferrin andthe transferrin receptor in human carcinoma A43l cells. J. Cell Biol., 97: 508—521,1983.
13. Press, 0. W., DeSantes, K., Anderson, S. K., and Geissler, F. Inhibition of catabolismof radiolabeled antibodies by tumor cells using lysosomotropic amines and carboxylicionophores. Cancer Res., 50: 1243—1250,1990.
14. Geissler, F., Anderson, S. K., Venkatesan, P., and Press, 0. Intracellular catabolismof radiolabeled anti-es antibodies by malignant B-cells. Cancer Res., 52: 2907—2915,1992.
15. Maack, T., Park, C. H., and Camargo, J. F. Renal filtration, transport, and metabolismof proteins. In: D. W. Seldin and G. Giebisch (eds.), The Kidney: Physiology andPathophysiology, Ed. 2, pp. 3005—3038.New York: Raven Press, Ltd., 1992.
16. Scharschmidt, B. F., Lake, J. R., Renner, E. L., Licko, V., and Van Dyke, R. W. Fluidphase endocytosis by cultured rat hepatocytes and perfused rat liver: implications forplasma membrane turnover and vesicular trafficking of fluid phase markers. Proc.NatI. Acad. Sci. USA, 83: 9488—9492, 1986.
17. Goodwin, D., Meares, C., Diamanti, C., McCall, M., La, C., Toni, F., McTigue, M.,and Martin, B. Use of specific antibody for rapid clearance of circulating bloodbackground from radiolabelled tumor imaging proteins. Eur. J. Nucl. Med., 9:209—215,1984.
18. Mellman, I. S., Plutner, H., Steinman, R. M., Unkeless, J. C., and Cohn, z. A.Internalization and degradation of macrophage Fc receptor during receptor mediatedphagocytosis. J. Cell Biol., 96: 887—895,1983.
19. Ukkonnen, P., Lewis, V., Marsh, M., Helenius, A., and Mellman, I. Transport ofmacrophage Fc receptors and Fc receptor-bound ligands to lysosomes. J. Exp. Med.,163:952—971,1986.
20. Arano, Y., Mukai, T., Uezono, T., Wakisaka, K., Motonari, H., Akizawa, H., Taoka,Y., and Yokoyama, A. A biological method to evaluate bifunctional chelating agentsto label antibodies with metallic radionuclides. J. NucI. Med., 35: 890—898, 1994.
21. Duncan, J. R., and Welch, M. J. Intracellular metabolism of Indium-l I l-DTPAlabeled receptor targeted proteins. J. Nucl. Med., 34: 1728—1738, 1993.
22. Franano,F. N., Edwards,W. B., Welch,M. J., and Duncan,J. R. Metabolismof receptortargeted ‘‘‘In-DTPA-glycoproteins:identification ‘‘‘In-DTPA-s-lysineas the primarymetabolic and excretory product. NucI. Med. Biol., 2!: 1023—1034,1994.
23. Motta-Hennessy, C., Sharkey, R. M., and Goldenberg, D. M. Metabolism of IndiumI 1I-labeled murine monoclonal antibody in tumor and normal tissue of the athymicmouse. J. Nucl. Med., 31: 1510—1519,1990.
24. Jones, P. L., Brown, B. A., and Sands, H. Uptake and metabolism of ‘‘‘In-labeledmonoclonal antibody B6.2 by the rat liver. Cancer Res., 50: 852s—856s, 1990.
25. Sands, H., and Jones, P. L. Methods for the study of the metabolism of radiolabeledmonoclonal antibodies by liver and tumor. J. Nucl. Med., 28: 390—398,1987.
26. Paik, C. H., Sood, V. K., Le, N., Cioloca, L., Carrasquillo, J. A., Reynolds, J. C.,Neumann, R. D., and Reba, R. C. Radiolabeled products in rat liver and serum afteradministration of antibody-amide-DTPA-Indium-lll. Nucl. Med. Biol., 19: 517—522, 1992.
27. Himmelsbach, M., and Wahl, R. L. Studies on the metabolic fate of ‘‘‘In-labeledantibodies. Int. J. Radiat. AppI. Part B Nucl. Med. Biol., 16: 839—845,1989.
28. Axworthy, D. B., Kasina, S., Rao, T. N., Srinivasan, A., and Fritzberg, A. R.Metabolite analysis, biliary and urinary excretion of Tc-99m-N2S2 preformed chelatelabeled anti-melanoma monoclonal antibody Fab fragment in mice and rats. J. NucI.Med., 32: 915, 1991.
29. Axworthy, D. B., Su, F. M., Vanderheyden, J. L., Srinivasan, A., Fitzner, J,, Galster,J., Beaumier, P., and Fritzberg, A. R. Excretion and biodistribution of rhenium-l86radiolabeled small molecule metabolites of radioimmunoconjugates. J. Nucl. Med.,32: 915, 1991.
30. Su, F. M., Axworthy, D., Galster, J., Weiden, P., Vanderheyden, J. L., and Fritzberg,A. R. Characterization of patients' urinary catabolites from Rh-l86 radiolabeledmonoclonal antibodies and fragments. J. Nucl. Med., 31: 823, 1990.
31. Deshpande, S. V., Subramanian, R., McCall, M. J., DeNardo, S. J., DeNardo, G. L.,and Meares, C. F. Metabolism of indium chelates attached to monoclonal antibody:
5719s
on May 28, 2020. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

METABOLISM OF ‘‘‘In-LABELEDMAbs
minimal transchelation of indium from Benzyl-EDTA chelate in vivo. J. Nucl. Med.,31: 218—224,1990.
32. Franssen, E. J. F., Koiter, J., Kuipers, C. A. M., Bruins, A. P., Moolenaar, F., deZeeuw, D., Kruizinga, W. H., Kellogg, R. M., and Meijer, D. K. F. Low molecularweight proteins as carriers for renal targeting. Preparation of drug-protein conjugatesand drug-spacer derivatives and their catabolism in renal cortex homogenates andlysosomal lysates. J. Med. Chem., 35: 1246—1259.1992.
33. Kirschke, H., and Barrett. A. J. Chemistry of lysosomal proteases, in lysosomes: theirrole in protein breakdown, pp. 103—240.London: Academic Press, 1987.
34. Ehrereich, B. A., and Cohn, Z. A. The fate of metabolites pinocytosed by macrophages in vitro. J. Exp. Med., 129: 227—243,1969.
35. de Duve, D., and Wattiaux, R. Functions of lysosomes. Annu. Rev. Physiol., 28:435—492, 1966.
36. Goldstein, J. L., Brown, M. S., Anderson, R. G. W., Russel, D. W., and Schneider, W.J. Receptor-mediated endocytosis: concepts emerging from the LDL receptor system.Annu. Rev. Cell Biol., 1: 1—39,1985.
37. Wileman, 1., Harding, C., and Stahl, P. Receptor-mediated endocytosis. Biochem. J.,232:1—14,1985.
38. Arano, Y., Mukai, 1., Uezono, 1., Taoka, Y., and Yokoyama, A. A novel biologicalmethod to evaluate chelating agents for labeling antibodies with metallic radionuclides. J. NucI. Med., 34: 59P, 1993.
39, Connett, J., Fenwick, J., Timmcke, A., and Philpott, G. Characterization of thebinding properties of murine monoclonal antibody (MAb) lA3, a newly describedantibody displaying anti-human colon cancer selectivity. Proc. Am. Assoc. CancerRes., 28: 352, 1987.
40. Yokota, T., Milenic, D. E., Whitlow, M., Wood, J. F., Hubert, S. L., and Schlom, J.Microautoradiographic analysis of the normal organ distribution of radioiodinatedsingle-chain Fv and other immunoglobulin forms. Cancer Res., 53: 3776—3783,1993.
41. Fischman, A. J., Pike, M. C., Kroon, D., Fucello, A. J., Rexinger, D., tenKate, C.,Wilkinson, R., Rubin, R. H., and Strauss, H. W. Imaging focal sites of bacterialinfection in rats with indium-I I I-labeled chemotactic peptide analogs. J. Nucl. Med.,32:483—491,1991.
42. Bakker, W. H., Krenning, E. P., Reubi, J. C., Breeman, W. A. P., Setyono-Han, B.,de Jong, M., Kooij, P. P. M., Bruns, C., van Hagen, P. M., Marbach, P., Visser, T. J.,
Pless, J., and Lamberts, S. W. J. In vivo application of lIn-1I l-DTPA-D-Phe]Octreotide for detection of somatostatin receptor-positive tumors in rats. Life Sci., 49:1593—1601,1991.
43. Yamazaki, K., and LaRusso, N. F. The Liver and Intracellular Digestion: How LiverCells Eat! Hepatology, 10: 877—886,1989.
44. Straus, W. Cytochemical observations on the relationship between lysosomes andphagosomes in the kidney and liver by combined staining for acid phosphatase andintravenously injected horseradish peroxidase. J. Cell Biol., 20: 497—507,1964.
45, Schneider, Y-J., Tulkens, P., de Duve, C., and Trouet, A. Fate of plasma membraneduring endocytosis. II. Evidence for recycling (shuttle) of plasma membrane constituents. J. Cell Biol., 82: 466—474,1979.
46. Kyriakos, R. J., Shih, L. B., Ong, G. L., Patel, K., Goldenberg, D. M., and Mattes, M.J, The fate of antibodies bound to the surface of tumor cells in vitro. Cancer Res., 52:835—842, 1992.
47, Shih, L. B., Thorpe. S. R., Griffiths, G. L., Diril, H., Ong, G. L., Hansen, H. J.,Goldenberg, D. M., and Mattes, M. J. The processing and fate of antibodies and theirradiolabels bound to the surface of tumor cells in vitro: a comparison of nineradiolabels. J. NucI. Med., 35: 899—907,1994.
48. Mattes, J. M., Griffiths, G. L., Diril, H., Goldenberg, D. M., Ong, G. L., and Shih, L.B. Processing of antibody-radioisotope conjugates after binding to the surface oftumor cells. Cancer (Phila), 73: 787—793,1994.
49, Naruki, Y., Carrasquillo, J. A., Reynolds, J. C., Maloney, P. J., Frincke, J. M.,Neumann, R. D., and Larson, S. M. Differential cellular catabolism of ‘‘‘In.40Yand
251 radiolabeled TiOl anti-CD5 monoclonal antibody. Int. J. Radiat. AppI. Part B
Nucl. Med. Biol., 17: 201—207,1990.50. Ohkuma, 5,, and Poole, B. Fluorescence probe measurement of the intralysosomal pH
in living cells and the perturbation of pH by various agents. Proc. Nati. Acad. Sci.USA, 75: 3327—3331,1978.
51. Gansow, 0. A. Newer approaches to the radiolabeling of monoclonal antibodies byuse of metal chelates. Int. J. Radiat. AppI. Part B NucI. Med. Biol., 18: 369—381,1991.
52. Rana, T. M., and Meares, C. F. N-terminal modification of immunoglobulin polypeptide chains tagged with isothiocyanato chelates. Bioconj. Chem.. 1: 357—362,1990.
5720s
on May 28, 2020. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1995;55:5714s-5720s. Cancer Res Buck E. Rogers, F. Nicholas Franano, James R. Duncan, et al.
in Vivoand Antibody Fragments In-Diethylenetriaminepentaacetic Acid-Monoclonal Antibodies
111Identification of Metabolites of
Updated version
http://cancerres.aacrjournals.org/content/55/23_Supplement/5714s
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/55/23_Supplement/5714sTo request permission to re-use all or part of this article, use this link
on May 28, 2020. © 1995 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
![Rapamycin Enhances Apoptosis and ... - Cancer Research · [CANCER RESEARCH 55, 1982-1988, May 1, 1995] Rapamycin Enhances Apoptosis and Increases Sensitivity to Cisplatin in Vitro1](https://img.pdfslide.net/doc/110x75/5fc155478e57503b59573a1a/rapamycin-enhances-apoptosis-and-cancer-research-cancer-research-55-1982-1988.jpg)







![(CANCER RESEARCH 55, 2615-2619, June 15, 1995] Inhibition ...cancerres.aacrjournals.org/content/canres/55/12/2615.full.pdf · suspended in 0.8 ml Buffer A containing 10 mM Tris/HCl](https://img.pdfslide.net/doc/110x75/5c877b8d09d3f2bc6b8bcd20/cancer-research-55-2615-2619-june-15-1995-inhibition-suspended-in-08.jpg)




















