Embed Size (px)
Citation preview

CAPÍTULO 3 ECOLOGÍA DE LOS MONOGENEOS
0
1
2
12
34

Paula S. Marcotegui
75Capítulo 3. Ecología de Monogeneos
3.1 Introducción
El estudio del parasitismo desde una perspectiva ecológica implica el análisis de
organismos que viven a expensas de otros. De ese modo, los hábitats en los que
residen las poblaciones de parásitos son organismos vivos (hospedadores),
potencialmente capaces de desarrollar un sistema defensivo y sujetos a las influencias
ambientales. Debido a esto, la ecología de parásitos debe tener en cuenta
simultáneamente la ecología de los hospedadores (Luque & Poulin, 2008).
El estudio de las poblaciones y de las comunidades parasitarias posibilita el
conocimiento de los factores que las estructuran y brinda información acerca de las
relaciones ecológicas, geográficas y filogenéticas entre hospedadores y parásitos. La
causa de por qué algunas especies de peces presentan mayores abundancias de
parásitos que otras, representa un importante capítulo de la ecología de peces (Luque
& Poulin, 2008).
Los parásitos en general muestran una distribución de tipo agregada, donde
típicamente pocos hospedadores presentan una alta intensidad parasitaria, mientras
que numerosos hospedadores presentan pocos o ningún individuo parásito (Poulin,
1998).
El ensamble de individuos de una misma especie de parásitos presente en un
individuo hospedador en un tiempo dado se denomina infrapoblación, de este modo al
conjunto de infrapoblaciones presentes en un individuo hospedador en un tiempo
determinado se la denomina infracomunidad. La suprapoblación incluye a todas las
infrapoblaciones de una especie dada, en todas sus etapas de desarrollo, incluyendo
sus fases de vida libre, en las poblaciones de las especies hospedadoras en un espacio
y tiempo particulares (Bush et al., 1997).
Los monogeneos, debido a su ciclo de vida directo resultan especialmente útiles
para investigar patrones de diversidad de especies, evolución morfológica y
especificidad por el hospedador (Morand et al., 2002a).
En el contexto de las relaciones parásito-hospedador, Lambert (1990) define
dos niveles de análisis. El nivel del hospedador y las condiciones bióticas y abióticas

Paula S. Marcotegui
76Capítulo 3. Ecología de Monogeneos
que éste brinda a los monogeneos y un segundo nivel que corresponde al ambiente
donde se desarrolla el sistema parasito-hospedador. Estos dos niveles, no están
aislados, si no que interaccionan, ya que a menudo las condiciones ambientales
pueden producir cambios en el hospedador al variar las condiciones ecofisiológicas del
mismo (migración, períodos de reproducción, etc.).
De acuerdo con Lambert (1990), la heterogeneidad branquial puede influenciar
los niveles de infestación y de colonización branquial, lo cual podría explicar la
variación en los patrones de riqueza y de abundancia en las comunidades de parásitos
branquiales.
En un ambiente natural, las densidades de parásitos normalmente son altas,
sin embargo, numerosos autores han documentado la existencia de una gran cantidad
de nichos vacantes en el microambiente branquial (Gutiérrez & Martorelli, 1999 b;
Buckmann & Lindenstrom, 2002; Simková et al., 2006). Esto favorece a su vez, la
colonización simultánea por varias especies de monogeneos en el mismo hospedador
(Boungou et al., 2008).
Por otro lado, como pauta general, el tamaño del cuerpo del hospedador es un
buen indicador de la riqueza específica de monogeneos (Morand et al., 2002a). Sin
embargo, numerosos estudios revelan la inexistencia de relaciones entre el tamaño
del hospedador y la riqueza de especies, sugiriendo que los procesos que actúan en la
composición de las comunidades de ectoparásitos difieren profundamente de las
comunidades de endoparásitos (Poulin & Rohde, 1997; Sasal et al., 1997; Morand et
al., 2002b). Mientras que los endoparásitos se acumulan normalmente en
hospedadores que están en un alto nivel de la cadena trófica, los monogeneos
ectoparásitos se acumulan frecuentemente en peces de mayor tamaño que viven en
grupos, o en poblaciones de peces de diferentes tamaños pero con alta densidad
poblacional (Morand et al., 2002b).
La descripción de los patrones de variación de la ocurrencia estacional de los
parásitos, permite inferir procesos relacionados con su ciclo de vida. Del mismo modo,

Paula S. Marcotegui
77Capítulo 3. Ecología de Monogeneos
se pueden establecer relaciones entre el ciclo de vida del parásito y el comportamiento
del hospedador.
Los estudios en dinámica poblacional de los monogeneos y los factores que
afectan su ocurrencia indican que los patrones de prevalencia, abundancia e
intensidad de infestación están relacionados con factores abióticos como temperatura
y concentración de oxígeno y bióticos como comportamiento, edad e inmunidad del
hospedador (Chubb, 1977).
El propósito del presente capitulo es caracterizar las poblaciones y las
comunidades de monogeneos hallados en Micropogonias furnieri, Mugil liza,
Parapimelodus valenciennis y Odontesthes argentinensis en el área de estudio. Se
analiza entonces la distribución de las poblaciones, las características del nicho
ocupado y las relaciones inter e intraespecíficas, la relación de los parámetros
poblacionales parasitarios con las características del hospedador (longitud y peso) y
con las características ambientales (concentración de oxigeno disuelto, pH,
temperatura del agua, conductividad).
3.2 Métodos de análisis
Se calcularon los siguientes índices parasitológicos definidos según Bush et al. (1997):
Prevalencia (P): número de hospedadores infectados por una especie parásita
particular, dividido por el número de total de hospedadores examinados.
Intensidad: número de individuos de una especie parásita particular hallados en un
hospedador individual infestado.
Intensidad media (IM): intensidad promedio de una especie particular de parásito
entre los hospedadores infestados. Se calcula como número total de parásitos/
número de hospedadores infestados.

Paula S. Marcotegui
78Capítulo 3. Ecología de Monogeneos
Abundancia: número de parásitos de una especie particular hallados en un
hospedador sin tener en cuenta si el hospedador está o no infestado.
Debido a que los hospedadores fueron mantenidos vivos hasta su prospección,
para detectar posibles pérdidas de monogeneos, se analizó la relación de la
abundancia de los monogeneos en función del orden de procesamiento mediante un
Test de Kruskal-Wallis.
La distribución espacial de los parásitos en el seno de las poblaciones
hospedadoras se calculó empleando los siguientes índices:
- Coeficiente de dispersión (CD): se calcula como:
CD= S2
La agregación es definida por la magnitud de la diferencia entre la varianza (S2) y la
media ( ) de la población. Cuando la varianza es mayor al promedio de parásitos por
hospedador se trata de una distribución agregada (S2/ >1). En distribuciones
uniformes será igual a cero (S2 nula) e igual a 1 en distribuciones aleatorias (S2= ).
(Morales & Pino, 1987).
-Coeficiente de agregación k: permite evaluar el grado de contagio de una especie
parásita en la población hospedadora; se calcula como:
k = 2
(S2- )
Este coeficiente es negativo en poblaciones con distribución normal, cercano a
8 en poblaciones con disposición al azar y muy inferior a 8 en poblaciones
sobredispersas (Morales y Pino, 1987).
De acuerdo con Esch et al. (1990), los estudios a nivel comunitario en parásitos
deben llevarse a cabo con especies que presentan una prevalencia mayor al 10%. Por

Paula S. Marcotegui
79Capítulo 3. Ecología de Monogeneos
este motivo, el análisis de la estructura comunitaria solo se llevó a cabo para M.
furnieri. Éste fue el único hospedador que presentó dos especies de monogeneos con
prevalencias mayores al 10%.
Para evaluar la relación entre las abundancias de A. bychowskyi y M.
argentinensis n comb. se realizó un test de Chi cuadrado con corrección de
continuidad de Yates. Las diferencias entre las intensidades medias de A. bychowskyi
y M. argentinensis n. comb. en los distintos arcos branquiales en los casos de
infecciones monoespecíficas y concurrentes se evaluaron mediante un test de Chi
cuadrado. Las diferencias en la abundancia en cada hemibranquia se evaluaron
mediante un test de Kruskal- Wallis para cada especie. En el caso de hallar diferencias
significativas se realizó el test a posteriori (Procedimiento de Marascuilo) con el fin de
establecer entre que arcos branquiales las diferencias fueron significativas (Berenson
& Levine, 1996).
La amplitud específica del nicho se analizó mediante el Índice de Levin B
(Simková et al., 2000), el cual se calcula como:
1
∑p
Donde, pj es la proporción de especímenes de una especie encontrada en la
hemibranquia j.
Para estandarizar este índice y obtener valores en una escala de 0 a 1, se
aplicó la fórmula:
1
N 1
Donde N es el número de sectores branquiales.
Los valores cercanos a 1 indican una máxima amplitud de nicho, mientras que
los valores cercanos a 0 evidencian nichos más restringidos.

Paula S. Marcotegui
80Capítulo 3. Ecología de Monogeneos
Se calculó el nivel de agregación intraespecífica (J) (Morand et al., 1999), el
cual se calcula como:
J11/ 1 1
m1
Donde, m1 y V1 representan la media y varianza de la especie 1.
Un valor de J=0 indica que los individuos están distribuidos al azar dentro del
hospedador, mientras que un valor de J mayor a 0 indica que existe una agregación
mayor que la esperada por azar.
Se calculó además, el nivel de agregación interespecífica (Morand et al., 1999),
el cual se calcula como:
C12 12
m1m2
Donde Cov12 es la covarianza entre la especie 1 y 2, m1 y m2 son las medias
de las especies 1 y 2 respectivamente.
Si el valor de C12 es mayor que 0, las dos especies están positivamente asociadas,
mientras que si el valor es menor a 0 se considera que existe una asociación negativa
entre las dos especies.
La reducción de competencia causada por la agregación interespecífica se
calculó como:
A121 1 2 1
12 1 2
Si este coeficiente tiene un valor mayor a 1, la agregación intraespecífica es
más fuerte que la agregación interespecífica (Morand et al., 1999).

Paula S. Marcotegui
81Capítulo 3. Ecología de Monogeneos
Para establecer la existencia de relaciones entre la longitud y el peso del hospedador
con la abundancia de cada especie de monogeneo se realizó el análisis de correlación
de Spearman (Siegel, 1979).
Con el fin de establecer si existen variaciones estacionales en la prevalencia y
la intensidad de los monogeneos hallados, se realizó un test de Kruskal-Wallis,
agrupando las estaciones de los 3 años de muestreo. La abundancia estacional fue
analizada mediante un test de Kruskal-Wallis, utilizando el método de Conover (1990)
para las comparaciones a posteriori.
Con el fin de verificar posibles cambios de la prevalencia, y la intensidad media
de los parásitos en función de los parámetros físico-químicos del ambiente, se
tomaron en cada muestreo la temperatura (con termómetro limnológico sumergible),
el pH (con peachímetro Hanna portátil), la concentración oxígeno disuelto y la
conductividad (con Analizador múltiple portátil Sper Scientific modelo 850081). La
posible correlación entre la prevalencia, la intensidad media y los parámetros
ambientales fue evaluada mediante correlación de Spearman. Adicionalmente la
relación entre la intensidad media y los parámetros físico-químicos fue analizado
mediante un test MLG (Modelo Lineal Generalizado). Las diferencias presentadas por
los parámetros físico-químicos analizados en relación a la estación del año fueron
evaluadas mediante un test de Kruskal-Wallis.
Para los análisis estadísticos se utilizó el programa XLSTAT Pro 7.5 (Addinsoft),
MedCalc y Statistica.
Los gráficos de Cajas y Bigotes realizados en el presente capítulo y en el
capítulo 5 representan la media y los desvíos estándar.
3.3. Resultados
3.3.1. Características de las poblaciones de hospedadores estudiados.
El número de ejemplares, los rangos de longitud y de peso de cada especie
hospedadora examinada se disponen en la Tabla 3.3.1.1.

Paula S. Marcotegui
82Capítulo 3. Ecología de Monogeneos
Tabla 3.3.1.1. Tabla resumen del número de hospedadores examinados, rangos de longitud total (en cm) y peso total (en gr) para cada especie.
Longitud Peso
Hospedador Nº hospedadores Mínimo Máximo Mínimo Máximo
M. furnieri 206 3,37 27,2 0,97 102,88
M. liza 149 2,8 32 0,22 331,41
P. valenciennis 118 5,29 22,3 1,07 111,7
O. argentinensis 103 1,25 19,99 0,39 68,24
3.3.2. Características de la infestación y dinámica de las poblaciones de
monogeneos
El análisis de la abundancia en función del orden de procesamiento de los
hospedadores no arrojó diferencias significativas en ninguna de las especies de peces.
En la tabla 3.3.2.1 se exponen en los resultados de los tests de Kruskall-Wallis
realizados para cada especie.
Tabla 3.3.2.1. Valores del Test de Kruskall-Wallis para cada especie hospedadora analizada.
Hospedador KW p M. furnieri 25,08 0,97M. liza 22,6 0,99
P. valenciennis 32,1 0,98O. argentinensis 27,3 0,99
A continuación se detallan para cada especie hospedadora los resultados obtenidos.
-Micropogonias furnieri
Un total de 65 (32%) hospedadores presentó de 1 a 6 ejemplares de A.
bychowskyi, mientras que la intensidad media fue de 1,8. Se halló un total de 117
ejemplares de A. bychowskyi.
En la Tabla 3.3.2.2 se muestran el número de hospedadores examinados, el
número de hospedadores parasitados por A. bychowskyi y el número de ejemplares
de esta especie hallados en cada clase de longitud total de M. furnieri.

Paula S. Marcotegui
83Capítulo 3. Ecología de Monogeneos
Tabla 3.3.2.2. Número de hospedadores examinados, parasitados por Absonifibula bychowskyi, número de especímenes de esta especie hallados.
Clases (cm) Nº hospedadores examinados Nº hospedadores parasitados Nº monogeneos
2,5‐7,5 88 31 59
7,6‐12,5 105 31 48
12,6‐17,5 11 2 6
17,6‐22,5 1 0 0
22,6‐27,5 1 1 4
Gráfico 3.3.2.1. Prevalencia e intensidad media por clase de longitud total de Micropogonias furnieri.
La población de A. bychowskyi presentó distribución agregada (CD=1,87;
K=0,64). El gráfico 3.3.2.2 expone el histograma de frecuencias de la abundancia de
A. bychowskyi.
Gráfico 3.3.2.2. Histograma de frecuencias de Absonifibula bychowskyi.
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
4,0
4,5
2,5‐7,5 7,6‐12,5 12,6‐17,5 17,6‐22,5 22,6‐27,5
Prevalen
cia e intensidad
med
ia
Clases (cm)
Intensidad media Prevalencia
0
20
40
60
80
100
120
140
160
0 1 2 3 4 5 6
Nº de
peces
Nº de parásitos

Paula S. Marcotegui
84Capítulo 3. Ecología de Monogeneos
Un total de 33 (16%) hospedadores presentó de 1 a 4 especímenes de M.
argentinensis n. comb., mientras que la intensidad media fue de 1,79. Se halló un
total de 33 ejemplares de M. argentinensis n. comb.
En la Tabla 3.3.2.3 se muestran el número de hospedadores examinados, parasitados
por M. argentinensis n. comb. y el número de monogeneos de esta especie hallados
en cada clase de longitud total del hospedador.
Tabla 3.3.2.3. Número de hospedadores examinados, parasitados por Macrovalvitrema. argentinensis n. comb. y número de especímenes hallados en Micropogonias furnieri.
Clases Nº hospedadores examinados Nº hospedadores parasitados Nº monogeneos
2,5‐7,5 88 11 19
7,6‐12,5 105 18 28
12,6‐17,5 11 4 12
17,6‐22,5 1 0 0
22,6‐27,5 1 0 0
Gráfico 3.3.2.3. Prevalencia e intensidad media de Macrovalvitrema argentinensis n. comb. por clase de longitud total de Micropogonias furnieri.
La población de M. argentinensis n comb. presentó distribución agregada
(CD=1,98; K=0,29). El gráfico 3.3.2.4 expone el histograma de frecuencias de
abundancias de M. argentinensis n comb.
0,0
0,5
1,0
1,5
2,0
2,5
3,0
3,5
2,5‐7,5 7,6‐12,5 12,6‐17,5 17,6‐22,5 22,6‐27,5
Prevalen
cia e intensidad
med
ia
Clases (cm)
Intensidad media Prevalencia

Paula S. Marcotegui
85Capítulo 3. Ecología de Monogeneos
Gráfico 3.3.2.4. Histograma de frecuencias Macrovalvitrema argentinensis n. comb.
-Mugil liza
Un total de 118 (79%) hospedadores presentó de 1 a 368 especímenes de L.
saladensis, mientras que la intensidad media fue de 24,8. Se halló un total de 2.929
ejemplares de L. saladensis. En la Tabla 3.3.2.4 se muestran el número de
hospedadores examinados, parasitados por L. saladensis y el número de monogeneos
de esta especie hallados en cada clase de longitud total de Mugil liza.
Tabla 3.3.2.4. Número de hospedadores examinados, parasitados por Ligophorus saladensis, número de monogeneos, por clase de longitud total de Mugil liza.
Clases (cm) Nº hospedadores examinados Nº hospedadores parasitados Nº monogeneos
2,5‐5 9 4 9
5,1‐7,5 21 17 153
7,6‐10 14 11 147
10,1‐12,5 11 10 189
12,6‐15 23 16 202
15,1‐17,5 31 29 847
17,6‐20 19 14 774
20,1‐22,5 11 10 392
22,6‐25 7 5 135
25,1‐27,5 2 1 74
27,6‐30 0 0 0
30,1‐32,5 1 1 7
0
50
100
150
200
0 1 2 3 4
Nº de
peces
Nº de parásitos

Paula S. Marcotegui
86Capítulo 3. Ecología de Monogeneos
Gráfico 3.3.2.5. Prevalencia y la intensidad media de Ligophorus saladensis por clase de longitud total de Mugil liza.
La población de L. saladensis presentó distribución agregada (CD=83,08;
K=0,24). El gráfico 3.3.2.6 expone el histograma de frecuencias de abundancias de L.
saladensis.
Gráfico 3.3.2.6. Histograma de frecuencias de Ligophorus saladensis.
-Parapimelodus valenciennis
Un total de 67 (57%) hospedadores presentó de 1 a 77 individuos de
Demidospermus annulus n sp., mientras que la intensidad media fue de 17,84. Se
halló un total de 1.195 ejemplares de D. annulus n. sp.
0
20
40
60
80
100
120
2,5‐5 5,1‐7,5 7,6‐10 10,1‐12,5 12,6‐15 15,1‐17,5 17,6‐20 20,1‐22,5 22,6‐25 25,1‐27,5 27,6‐30 30,1‐32,5
Prevalen
cia e intensidad
med
ia
Clases (cm)
PrevalenciaIntensidad media
0
5
10
15
20
25
30
35
0 20 40 60 80 100 120 140 160 180 200 220 240 260 280 300 320 340 360
Nº de
peces
Nº de parásitos

Paula S. Marcotegui
87Capítulo 3. Ecología de Monogeneos
La población de D. annulus n sp. presentó distribución agregada (CD=26,27;
K=0,4). El gráfico 3.3.2.7 expone el histograma de frecuencias de abundancias de D.
annulus n sp.
En la Tabla 3.3.2.5 se muestran el número de hospedadores examinados, parasitados
por D. annulus n sp. y el número de monogeneos de esta especie hallados para cada
clase de longitud total del hospedador.
Tabla 3.3.2.5. Número de hospedadores examinados, parasitados por Demidospermus annulus n. sp. y número de monogeneos hallados en cada clase de longitud total de Parapimelodus valenciennis.
Clases (cm) Nº hospedadores examinados Nº hospedadores parasitados Nº monogeneos
5,1‐7,5 15 4 53
7,6‐10 33 18 325
10,1‐12,5 15 9 115
12,6‐15 17 11 242
15,1‐17,5 21 18 323
17,6‐20 12 2 5
20,1‐22,5 5 5 132
Gráfico 3.3.2.6. Prevalencia y la intensidad media por clase de longitud total de Parapimelodus valenciennis.
0,00
5,00
10,00
15,00
20,00
25,00
30,00
5,1‐7,5 7,6‐10 10,1‐12,5 12,6‐15 15,1‐17,5 17,6‐20 20,1‐22,5
Prevalen
cia e intensidad
med
ia
Clases (cm)
Intensidad media Prevalencia

Paula S. Marcotegui
88Capítulo 3. Ecología de Monogeneos
Gráfico 3.3.2.7. Histograma de frecuencias de Demidospermus annulus n sp.
-Odontesthes argentinensis
Un total de 52 (50%) hospedadores presentó de 1 a 15 ejemplares de
Gyrodactylus mauri n sp., mientras que la intensidad media fue de 4,44. Se halló un
total de 231 ejemplares de Gyrodactylus mauri n sp. En la tabla 3.3.2.6 se muestran
el número de hospedadores examinados, parasitados por G. mauri n. sp. y el número
de monogeneos de esta especie hallados en cada clase de longitud total de O.
argentinensis.
La población de Gyrodactylus mauri n. sp. presentó distribución agregada
(CD=4,5; K=0,64). El gráfico 3.3.2.9 expone el histograma de frecuencias de
abundancias de Gyrodactylus mauri n. sp.
Tabla 3.3.2.6. Número de hospedadores examinados, parasitados por Gyrodactylus mauri n. sp.,
nº de monogeneos, por clase de longitud total de Odontesthes argentinensis.
Clases (cm)
Nº hospedadores examinados
Nº hospedadores parasitados
Nº monogeneos
1‐2 1 0 0
3‐5 15 9 48
6‐8 33 16 64
9‐11 10 8 30
12‐15 34 15 66
16‐18 7 4 23
19‐20 3 0 0
0
10
20
30
40
50
60
0 2 4 6 8 10 12 14 16 18 20 22 24 26 28 30 32 34 36 38 40 42 44 46 48 50 52 54 56 58 60 62 64 66 68 70 72 74 76
Nº de
peces
Nº de parásitos

Paula S. Marcotegui
89Capítulo 3. Ecología de Monogeneos
Gráfico 3.3.2.8. Prevalencia y la intensidad media de Gyrodactylus mauri n. sp. por clase de longitud total de Odontesthes argentinensis.
Gráfico 3.3.2.9. Histograma de frecuencias de Gyrodactylus mauri n sp.
3.3.3. Características de las infracomunidades de monogeneos en M. furnieri
El 60% de los ejemplares de M. furnieri parasitados presentó solo A.
bychowskyi, el 26 % solo M. argentinensis n. comb., mientras que el porcentaje de co-
ocurrencia fue del 13%. Si bien los valores de intensidad media y prevalencia
estacionales de A. bychowskyi y M. argentinensis n. comb. tienen una relación inversa
(Gráfico 3.3.3.1 y 3.3.3.2), por lo que cabría esperar una asociación negativa, los
resultados del test de Chi cuadrado no arrojaron valores significativos de asociación
(ver Tabla 3.3.3.1).
0,00
1,00
2,00
3,00
4,00
5,00
6,00
7,00
1‐2 3‐5 6‐8 9‐11 12‐15 16‐18 19‐20
Prevalen
cia e intensidad
med
ia
Clases (cm)
Intensidad media Prevalencia
0
10
20
30
40
50
60
0 1 2 3 4 5 6 7 8 9 10 11 12 13 14
Nº de
peces
Nº de parásitos

Paula S. Marcotegui
90Capítulo 3. Ecología de Monogeneos
Gráfico 3.3.3.1. Intensidad media de Absonifibula bychowskyi y Macrovalvitrema argentinensis n. comb. en las diferentes estaciones durante los tres años de muestreo.
Gráfico 3.3.3.2. Prevalencia de Absonifibula bychowskyi y Macrovalvitrema argentinensis n. comb. en las diferentes estaciones durante los tres años de muestreo.
0
0,5
1
1,5
2
2,5
3
3,5
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
Intensidad
med
ia
A. bychowskyi M. argentinensis n. comb,
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
Prevalen
cia
A. bychowskyi M. argentinensis n. comb,
2006 2007 2008
A. bychowskyi M. argentinensis n. comb.
2006 2007 2008
A. bychowskyi M. argentinensis n. comb.

Paula S. Marcotegui
91Capítulo 3. Ecología de Monogeneos
Gráfico 3.3.3.3. Prevalencia de Absonifibula bychowskyi y Macrovalvitrema argentinensis n. comb. en cada clase de longitud total de M. furnieri.
Tabla 3.3.3.1. Resultados de los test realizados entre las abundancias de Absonifibula bychowskyi y Macrovalvitrema argentinensis n. comb.
Valor observado
Valor critico
p
Chi‐cuadrado 0,196 3,841 0,658
G² de Wilks 0,193 3,841 0,661
Chi‐cuadrado con corrección de la continuidad de Yates 0,055 3,841 0,814
Monte Carlo I 0,196 ‐ 0,679
Los parámetros poblacionales de ambas especies de monogeneos no mostraron
correlación significativa. El gráfico 3.3.3.4 exhibe la relación entre las abundancias de
A. bychowskyi y M. argentinensis n. comb. Las abundancias de estas dos especies no
exhibieron correlación significativa (rs= 0,019, p= 0,789; n= 206).
El gráfico 3.3.3.5 exhibe la relación entre intensidades medias de A. bychowskyi y M.
argentinensis n. comb. Las intensidades medias de estas dos especies en las
diferentes clases de longitud total de los hospedadores no exhibieron correlación
significativa (rs= 0,204, p= 0,572; n= 5).
Las prevalencias de A. bychowskyi y M. argentinensis n comb. no presentaron
correlación significativa en los diferentes clases de longitud total (rs= -0,093, p=
0,797; n= 5). El gráfico 3.3.3.6. exhibe la relación entre las prevalencias de las dos
especies.
0,0
0,2
0,4
0,6
0,8
1,0
1,2
2,5‐7,5 7,6‐12,5 12,6‐17,5 17,6‐22,5 22,6‐27,5
Prevalen
cia
Clases (cm)
A. bychowskyi M. argentinensis n. comb.A. bychowskyi M. argentinensis n. comb.

Paula S. Marcotegui
92Capítulo 3. Ecología de Monogeneos
Gráfico 3.3.3.4. Relación entre las abundancias de Absonifibula bychowskyi y Macrovalvitrema argentinensis n. comb.
Gráfico 3.3.3.5. Relación entre las intensidades medias de Absonifibula bychowskyi y Macrovalvitrema argentinensis n. comb.
Gráfico 3.3.3.6. Relación entre las prevalencias de Absonifibula bychowskyi y Macrovalvitrema argentinensis n. comb.
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
0 1 2 3 4 5 6 7
Nº de
M. a
rgen
tine
nsisn. sp.
Nº de A. bychowskyi
0
0,5
1
1,5
2
2,5
3
3,5
0 0,5 1 1,5 2 2,5 3 3,5 4 4,5
M. a
rgen
tine
nsisn. com
b.
A. bychowskyi
0
0,1
0,2
0,3
0,4
0,5
0,6
0 0,2 0,4 0,6 0,8 1 1,2
M. a
rgen
tine
nsisn. com
b.
A. bychowskyi

Paula S. Marcotegui
93Capítulo 3. Ecología de Monogeneos
3.3.4. Preferencia por hemibranquias y/o arco branquial
Los valores de abundancia de los monogeneos hallados en las hemibranquias
anteriores y las posteriores y en los arcos izquierdos y derechos no presentaron
diferencias significativas. En las tablas 3.3.4.1 y 3.3.4.2 se exhiben los valores de
Kruskal-Wallis obtenidos por especie, comparando las hemibranquias anteriores y
posteriores e izquierda y derecha respectivamente. Este análisis no se ha realizado
con los ejemplares de Gyrodactylus maurin n sp., debido a que solo se hallaron en la
superficie corporal. Los gráficos 3.3.4.1 y 3.3.4.2 exhiben la distribución de la
abundancia en los diferentes arcos branquiales para cada especie de monogeneo
analizada.
Tabla 3.3.4.1. Valores de Kruskal-Wallis obtenidos al comparar la abundancia de las especies de monogeneos analizadas en las hemibranquias anteriores y posteriores.
H (valor observado) H (valor crítico) p
A. bychowskyi 2,083 3,841 0,149 M. argentinensis n. comb. 0,759 3,841 0,384
L. saladensis 3 3,841 0,083 D. annulus n sp. 2,083 3,841 0,149
Tabla 3.3.4.2. Valores de Kruskal-Wallis obtenidos al comparar la abundancia de las especies de monogeneos analizadas en los arcos izquierdos y derechos.
H (valor observado) H (valor crítico) p
A. bychowskyi 0,333 3,841 0,564 M. argentinensis n comb. 2,551 3,841 0,11
L. saladensis 1,333 3,841 0,248 D. annulus n sp. 3 3,841 0,083

Paula S. Marcotegui
94Capítulo 3. Ecología de Monogeneos
Gráfico 3.3.4.1. Distribución de la abundancia de Absonifibula bychowskyi (a), de Macrovalvitrema argentinensis n comb. (b), de Ligophorus saladensis (c) y de Demidospermus annulus n sp. (d) en las hemibranquias anteriores y posteriores.
a)
b)
c)
d)
0
10
20
30
40
50
1 2 3 4
Abu
ndancia A. bycho
wskyi
Nº arco branquial
Anterior Posterior
0
2
4
6
8
10
12
14
16
1 2 3 4
Abu
ndancia M. argentin
ensis n.
comb.
Nº arco branquial
Anterior
Posterior
0100200300400500600700800900
1 2 3 4
Abu
ndancia L. salad
ensis
Nº arco branquial
Anterior
Posterior
0
50
100
150
200
250
1 2 3 4
Abu
ndancia D. ann
ulus n sp.
Nº arco branquial
Anterior
Posterior

Paula S. Marcotegui
95Capítulo 3. Ecología de Monogeneos
Gráfico 3.3.4.2. Distribución de la abundancia de Absonifibula bychowskyi (a), de Macrovalvitrema argentinensis n. comb. (b), de Ligophorus saladensis (c) y de Demidospermus annulus n sp. (d) en los arcos branquiales izquierdos y derechos.
a)
b)
c)
d)
05
1015202530354045
1 2 3 4
Abu
ndancia A. bycho
wskyi
Nº arco branquial
Derecha
Izquierda
0
5
10
15
20
25
1 2 3 4
Abu
ndancia M. argentin
enesisn.
comb.
Nº arco branquial
Derecha
Izquierda
0
100
200
300
400
500
600
700
1 2 3 4
Abu
ndancia L. salad
ensis
Nº arco branquial
Derecha
Izquierda
0
50
100
150
200
250
1 2 3 4
Abu
ndancia D. ann
ulus n sp
Nº arco branquial
Derecha
Izquierda

Paula S. Marcotegui
96Capítulo 3. Ecología de Monogeneos
Los ejemplares de A. bychowskyi exhibieron preferencia por los primeros y
segundos arcos branquiales (X2=77,858, p=0,0001). El 50% de los ejemplares se
halló en el primer arco y el 32% en el segundo arco (Gráfico 3.3.4.3). Los resultados
obtenidos del procedimiento de Marascuilo se exponen en la Tabla 3.3.4.3.
Tabla 3.3.4.3. Valores del Procedimiento de Marascuilo para evaluar las diferencias en la abundancia de Absonifibula bychowskyi entre los distintos arcos branquiales de Micropogonias furnieri.
Pares de arcos Valor Valor crítico
|1 ‐ 2| 0,180* 0,173
|1 ‐ 3| 0,385* 0,150
|1 ‐ 4| 0,434* 0,141
|2 ‐ 3| 0,205* 0,143
|2 ‐ 4| 0,254* 0,134
|3 ‐ 4| 0,049 0,102
*valores significativos
Gráfico 3.3.4.3. Distribución de las abundancias de Absonifibula bychowskyi en los diferentes arcos branquiales de Micropogonias furnieri.
La distribución de la abundancia de M. argentinensis n comb. en los diferentes arcos
branquiales presentó diferencias significativas (X2= 19,59, p= 0,0001). El 46% de los
individuos se ubicó en el segundo arco branquial, mientras que el primer y el cuarto
arco resultaron de menor preferencia (Gráfico 3.3.4.2). La Tabla 3.3.4.4 exhibe los
valores del procedimiento de Marascuilo.
50%
32%
11%
7% Nº arco
1
2
3
4

Paula S. Marcotegui
97Capítulo 3. Ecología de Monogeneos
Tabla 3.3.4.4. Valores del Procedimiento de Marascuilo para evaluar las diferencias en la abundancia de Macrovalvitrema argentinensis n. comb. entre los distintos arcos branquiales de Micropogonias furnieri.
Pares de arcos Valor Valor crítico
|1 ‐ 2| 0,305* 0,224
|1 ‐ 3| 0,085 0,203
|1 ‐ 4| 0,000 0,185
|2 ‐ 3| 0,220 0,238
|2 ‐ 4| 0,305* 0,224
|3 ‐ 4| 0,085 0,203
*valores significativos
Gráfico 3.3.4.4. Distribución de las abundancias de Macrovalvitrema argentinensis n. comb. en los diferentes arcos branquiales de Micropogonias furnieri.
La diferencias entre las intensidades medias de cada arco branquial cuando A.
bychowskyi y M. argentinensis n. comb. se presentaron en infecciones
monoespecíficas y cuando estos monogeneos co-ocurrieron no fueron significativas
(X2= 1,249, p=0,364). EL gráfico 3.3.4.5. muestra la distribución de las intensidades
medias en cada arco branquial en los diferentes casos.
15%
46%
24%
15% Nº Arco
1
2
3
4

Paula S. Marcotegui
98Capítulo 3. Ecología de Monogeneos
Gráfico 3.3.4.5. Intensidad media en cada arco branquial de Micropogonias furnieri en infestaciones monoespecíficas de Absonifibula bychowskyi y de Macrovalvitrema argentinensis n. comb. y cuando co-ocurren.
-Mugil liza
La mayor abundancia de los ejemplares de L. saladensis ocurre en los primeros
arcos branquiales, el 31% se halló en el primer arco branquial y el 29% en el segundo
arco branquial (Gráfico 3.3.4.6). El test de comparación de proporciones arrojó
diferencias significativas (X2=711,96; p=0,0001). La Tabla 3.3.4.5 exhibe los valores
del procedimiento de Marascuilo.
Tabla 3.3.4.5. Valores del Procedimiento de Marascuilo para evaluar las diferencias en abundancias de Ligophorus saladensis entre los distintos arcos branquiales de Mugil liza.
Pares de arcos Valor Valor crítico
|1 ‐ 2| 0,058* 0,027
|1 ‐ 3| 0,141* 0,026
|1 ‐ 4| 0,252* 0,024
|2 ‐ 3| 0,082* 0,026
|2 ‐ 4| 0,194* 0,023
|3 ‐ 4| 0,111* 0,021
*valores significativos
0
0,2
0,4
0,6
0,8
1
1
2
3
4
Intensidad
med
ia

Paula S. Marcotegui
99Capítulo 3. Ecología de Monogeneos
Gráfico 3.3.4.6. Distribución de las abundancias de Ligophorus saladensis en los diferentes arcos branquiales de Mugil liza.
-Parapimelodus valenciennis
La abundancia de D. annulus n sp. en los diferentes arcos branquiales arrojó
diferencias significativas (X2=84,85; p=0,0001). El 42 % de los especímenes se halló
en el primer arco branquial y el 25% en el tercero. El procedimiento de Marascuilo
arrojó diferencias significativas entre los arcos 2, 3 y 4 (ver tabla 3.3.4.6).
Tabla 3.3.4.6. Procedimiento de Marascuilo para evaluar las diferencias en abundancia de Demidospermus annulus n sp. entre los distintos arcos branquiales de Parapimelodus valenciennis.
Pares de arcos Valor Valor crítico
|1 ‐ 2| 0,026 0,052
|1 ‐ 3| 0,087* 0,050
|1 ‐ 4| 0,148* 0,048
|2 ‐ 3| 0,061* 0,049
|2 ‐ 4| 0,122* 0,047
|3 ‐ 4| 0,061* 0,045
*valores significativos
Gráfico 3.3.4.7. Distribución de las abundancias de Demidospermus annulus n sp. en los diferentes arcos branquiales de Parapimelodus valenciennis.
31%
29%
23%
17% Nº arco
1
2
3
4
42%
18%
25%
15% Nº arco
1
2
3
4

Paula S. Marcotegui
100Capítulo 3. Ecología de Monogeneos
3.3.5. Amplitud del nicho
La Tabla 3.3.5.1 exhibe los valores de amplitud de nicho a partir del índice de
Levin (B) estandarizado obtenidos para cada especie. Los valores bajos presentados
por todas las especies analizadas indican restricción en el nicho.
Tabla 3.3.5.1. Valores del índice de amplitud de nicho de Levin para cada especie analizada.
Promedio Desvío
A. bychowskyi 0,080 0,124
M. argentinensis n. comb. 0,104 0,107
L. saladensis 0,245 0,279
D. annulus n sp. 0,145 0,329
El 62% de las infrapoblaciones de A. bychowskyi presentó valores de
agregación intraespecífica (J) menores a 0, mientras que el 85% de las
infrapoblaciones de M. argentinensis n comb. presentaron valores de agregación
intraespecífica menores a 0. La agregación interespecífica entre A. bychowskyi y M.
argentinensis n comb. presentó en el 90% de las infracomunidades valores de cero,
indicando la ausencia de este tipo de agregación. La reducción de competencia
causada por la agregación interespecífica (A12) en el 46% de las infracomunidades
tomó valores mayores a 1, siendo el promedio de 1,26. Esto indica que la agregación
intraespecifica de A. bychowskyi y M. argentinensis n comb. es más fuerte que la
agregación interespecífica.
El 73,4% de las infrapoblaciones de L. saladensis presentó valores de
agregación intraespecífica mayores a 0. En el caso de D. annulus n sp., el 86% de las
infrapoblaciones presentó valores de agregación intraespecífica mayores a 0. De esta
forma las infrapoblaciones de estas dos especies presentan un patrón de agregación
intraespecífica mayor que la esperada por azar.
3.3.6. Relación abundancia-longitud y peso
A continuación se detallan los resultados obtenidos para cada especie
hospedadora.

Paula S. Marcotegui
101Capítulo 3. Ecología de Monogeneos
-Micropogonias furnieri
El 83% de los individuos de A. bychowskyi se halló en hospedadores de tallas
entre los 5,1 y 10 cm de longitud total. La talla mínima de infestación fue de 3,37 cm
y la talla máxima fue 27,2 cm, la cual corresponde a los individuos de la talla máxima
examinados.
El 79,7% de los ejemplares de M. argentinensis n comb. se halló en peces de
tallas entre los 5,1 y 12,5 cm de longitud total. La talla mínima de infestación en las
corvinas fue 5,22, mientras que la talla máxima correspondió a los 16,69 cm.
Se observó correlación positiva entre la longitud del hospedador y la
abundancia de M. argentinensis n comb. (rs=0,211, p=0,002). El peso del hospedador
estuvo correlacionado positivamente con la abundancia de M. argentinensis n. comb.
(rs=0,159, p=0,023) y negativamente con A. bychowskyi (rs=-0,139, p=0,046). No se
observó correlación entre la longitud total del hospedador y la abundancia de A.
bychowskyi (rs= -0,0733; p=0,295). Los gráficos de 3.3.6.1 a 3.3.6.4 exhiben la
distribución de la abundancia de A. bychowskyi y M. argentinensis n comb. en función
de la longitud y el peso del hospedador.
Gráfico 3.3.6.1. Distribución de la abundancia de Absonifibula bychowskyi en función de la longitud total del hospedador.
y = 0,0119x + 0,4684
0
1
2
3
4
5
6
7
0 5 10 15 20 25 30
Abu
ndan
cia A. bycho
wskyi
Longitud Total del hospedador (cm)

Paula S. Marcotegui
102Capítulo 3. Ecología de Monogeneos
Gráfico 3.3.6.2. Distribución de la abundancia de Absonifibula bychowskyi en función del peso total del hospedador.
Gráfico 3.3.6.3. Distribución de la abundancia de Macrovalvitrema argentinensis n comb. en función de la longitud total del hospedador.
Gráfico 3.3.6.4. Distribución de la abundancia de Macrovalvitrema argentinensis n comb. en función del peso total del hospedador.
y = 0,0025x + 0,5476
0
1
2
3
4
5
6
7
0 20 40 60 80 100 120
Abu
ndan
cia A. bycho
wskyi
Peso total del hospedador (gr)
y = 0,0119x + 0,4684
0
1
2
3
4
5
6
7
0 5 10 15 20 25 30Abu
ndan
cia M. argen
tine
nsisn comb.
Longitud total del hospedador (cm)
y = 0,0108x + 0,2038
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
0 20 40 60 80 100 120
Anu
ndan
cia M. argen
tine
nsis n com
b.
Peso total del hospedador (gr)

Paula S. Marcotegui
103Capítulo 3. Ecología de Monogeneos
-Mugil liza
El 68,72% de los individuos de L. saladensis se halló entre los 15,1 y 22 cm de
longitud total del hospedador. La talla mínima de infestación fue 3,78 cm, mientras
que la talla máxima fue 32 cm, la cual corresponde a los individuos de talla máxima
examinados.
Se observó correlación positiva entre la abundancia de L. saladensis y la
longitud total (rs=0,250; p=0,002) y el peso total (rs=0,272; p=0,001) del pez. Los
gráficos 3.3.6.5 y 3.3.6.6 exhiben la distribución de la abundancia de L. saladensis en
función de la longitud y del peso del hospedador.
Gráfico 3.3.6.5. Distribución de la abundancia de Ligophorus saladensis en función de la longitud total del hospedador.
Gráfico 3.3.6.6. Distribución de la abundancia de Ligophorus saladensis en función del peso total del hospedador.
y = 1,6448x ‐ 3,2442
0
50
100
150
200
250
300
350
400
0 5 10 15 20 25 30 35
Abu
ndancia L. salad
encis
Longitud total del hospedador (cm)
y = 0,1473x + 12,805
0
50
100
150
200
250
300
350
400
0 50 100 150 200 250 300 350
Abu
ndancia L. salad
ensis
Peso total del hospedador (gr)

Paula S. Marcotegui
104Capítulo 3. Ecología de Monogeneos
-Parapimelodus valenciennis
Entre los 12 y los 17,5 cm de longitud total del hospedador se halló el 47,2%
de los individuos de D. annulus n sp. La talla mínima de infestación de los
hospedadores fue 5,9 cm, mientras que la talla máxima fue de 22,3 cm, la cual
corresponde a los individuos de talla máxima examinados.
No se observó correlación entre la abundancia de D. annulus n sp. y la longitud
(rs=-0,044; p=0,637) y el peso (rs=-0,052; p=0,576) del pez. Los gráficos 3.3.6.7 y
3.3.6.8 exhiben la distribución de la abundancia de D. annulus n sp. en función de la
longitud y del peso del hospedador.
Gráfico 3.3.6.7. Distribución de la abundancia de Demidospermus annulus n sp. en función de la longitud total del hospedador.
Gráfico 3.3.6.8. Distribución de la abundancia de Demidospermus annulus n sp. en función del peso total del hospedador.
y = ‐0,3291x + 14,26
0
10
20
30
40
50
60
70
80
90
0 5 10 15 20 25Abu
ndan
cia Dem
udospe
rmus ann
ulus n
sp.
Longitud Total del hospedador (cm)
y = ‐0,1009x + 13,473
0
10
20
30
40
50
60
70
80
90
0 20 40 60 80 100 120
Abu
ndan
cia Dem
idospe
rmus ann
ulus n sp.
Peso total del hospedador(gr)

Paula S. Marcotegui
105Capítulo 3. Ecología de Monogeneos
-Odontesthes argentinensis
Los hospedadores de 3 a 5 cm de longitud total presentaron el 48,48% de los
ejemplares de G. mauri n sp., mientras que aquellos con tallas entre 12 y 15 cm
presentaron el 28% de los ejemplares hallados. La talla mínima de infestación fue de
3,35 cm, mientras que la talla máxima fue de 18,24, la cual corresponde a los
ejemplares de la máxima longitud examinados de este hospedador.
No se observó correlación entre la abundancia individual de Gyrodactylus mauri
n sp. y la longitud (rs=-0,060; p=0,547) y el peso total del hospedador (rs=-0,060;
p=0,547). Los gráficos 3.3.6.9 y 3.3.6.10 exhiben la distribución de la abundancia de
Gyrodactylus mauri n sp. en función de la longitud y del peso del hospedador.
Gráfico 3.3.6.9. Distribución de la abundancia de Gyrodactylus mauri n sp. en función de la longitud total del hospedador.
Gráfico 3.3.6.10. Distribución de la abundancia de Gyrodactylus mauri n sp. en función del peso total del hospedador.
y = ‐0,0176x + 2,4272
0
2
4
6
8
10
12
14
16
0 5 10 15 20 25Abu
ndan
cia Gyrod
actylus mau
rin sp.
Longitud total (cm)
y = ‐0,0084x + 2,3447
0246810121416
0 10 20 30 40 50 60 70 80Abu
ndan
cia Gyrod
actylus mau
rin sp.
Peso total del hospedador

Paula S. Marcotegui
106Capítulo 3. Ecología de Monogeneos
La intensidad media de los ejemplares de D. annulus n. sp. en los diferentes
arcos branquiales en función de la longitud del hospedador no presentó diferencias
significativas (Q= 7,72; p=0,0052). El gráfico 3.3.6.11 exhibe la distribución de la
intensidad media de esta especie en cada arco branquial en función de la longitud del
hospedador.
Al aumentar el tamaño del hospedador aumentó la intensidad media y esto
provocó una mayor concentración de los ejemplares de L. saladensis en los dos
primeros arcos branquiales de M. liza (Q=21,057; p=0,000) (gráfico 3.3.6.12 a).
Al aumentar la longitud de M. furnieri la intensidad de los ejemplares de A.
bychowskyi tendió a aumentar en los segundos y terceros arcos (Q=11,94; p=0,008),
mientras que no existieron diferencias significativas en la distribución de M.
argentinensis n comb. en función de la longitud del hospedador (Q=3,78; p=0,28).
Los gráficos 3.3.6.12 b y c exhiben la distribución de la intensidad media en cada arco
branquial en función de la longitud del hospedador.
Gráfico 3.3.6.11. Distribución de la intensidad media de Demidospermus annulus n sp. en los diferentes arcos branquiales en función de la longitud del hospedador.
5,1‐7,57,6‐10
10,1‐12,512,6‐15
15,1‐17,517,6‐20
20,1‐22,5
0
5
10
15
20
1
2
3
4
Clases de longitud (cm)
Intensidad media

Paula S. Marcotegui
107Capítulo 3. Ecología de Monogeneos
Gráfico 3.3.6.12. Distribución de la intensidad media de los monogeneos en los diferentes arcos branquiales en función de la longitud del hospedador. a) L. saladensis, b) A. bychowskyi, c) M. argentinensis n comb.
a)
b)
c)
2,5‐55,1‐7,5
7,6‐1010,1‐12,5
12,6‐1517,6‐20
20,1‐22,522,6‐25
25,1‐27,527,6‐30
30,1‐32,5
0
5
10
15
20
1
2
3
4
Clases de longitud (cm)
Intensidad media
2,5‐55,1‐7,5
7,6‐1010,1‐12,5
12,6‐1525,1‐27,5
0
0,5
1
1,5
2
1
2
3
4
Clases de longitud (cm)
Intensidad media
2,5‐55,1‐7,5
7,6‐1010,1‐12,5
15,1‐17,5
00,20,40,6
0,8
1
1,2
1
2
3
4
Clases de longitud (cm)
Intensidad media

Paula S. Marcotegui
108Capitulo 3. Ecología de los Monogeneos
3.3.7. Estacionalidad de los distintos índices ecológicos
No se observaron diferencias significativas en la prevalencia e intensidad media
entre las diferentes estaciones del año. Las tablas 3.3.7.1 a 3.3.7.5 exhiben los
valores del test de Kruskal-Wallis realizado para cada especie de monogeneo. En los
gráficos 3.3.7.1 a 3.3.7.15 se observan los patrones de variación estacional de los
índices estudiados, representando la media y los desvíos.
-Micropogonias furnieri
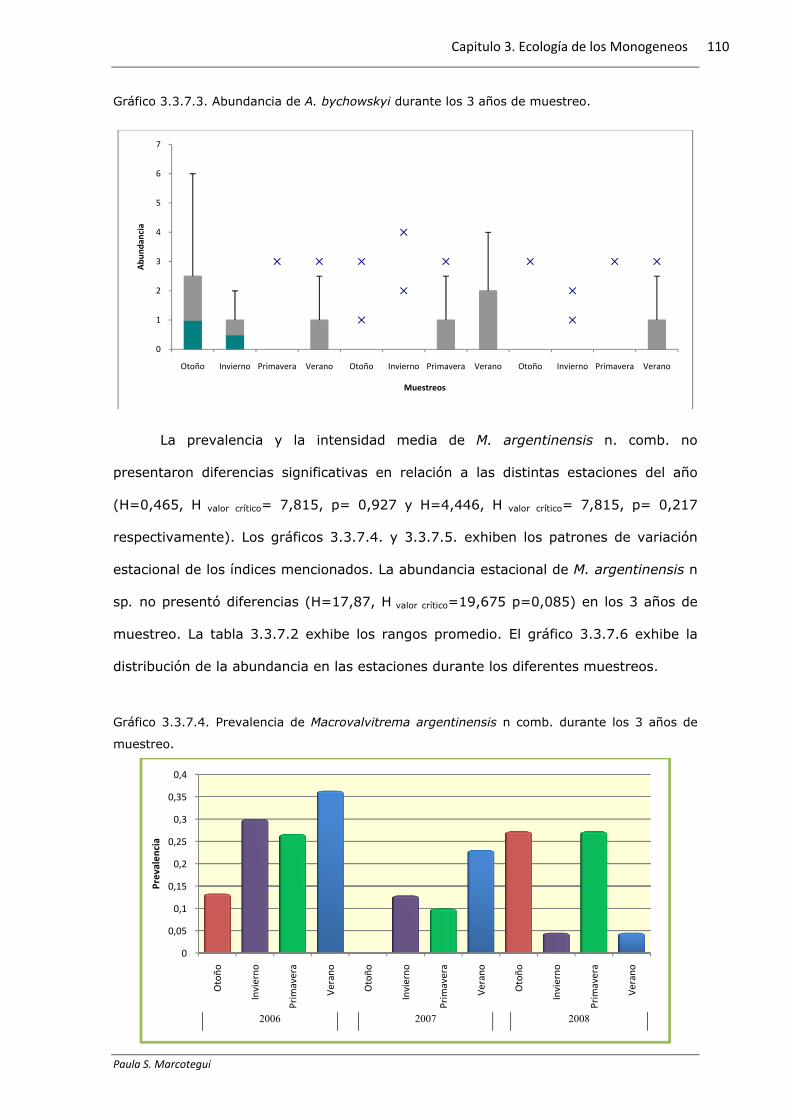
La prevalencia y la intensidad media de A. bychowskyi no presentaron
diferencias significativas en relación a las distintas estaciones del año (H=2,226, H valor
crítico= 7,815, p= 0,527 y H=3,147, H valor crítico= 7,815, p= 0,37 respectivamente). Los
gráficos 3.3.7.1. y 3.3.7.2. exhiben los patrones de variación estacional de los índices
mencionados. La abundancia estacional de A. bychowskyi presentó diferencias
(H=26,707, H valor crítico=19,675 p=0,05). Sin embargo no puede establecerse un
patrón estacional de la abundancia, debido a que se hallaron diferencias significativas
entre las mismas estaciones de los diferentes años de muestreo. La tabla 3.3.7.1
exhibe los rangos promedio y las diferencias significativas halladas.
Gráfico 3.3.7.1. Prevalencia estacional de Absonifibula bychowskyi durante los 3 años de
muestreo.
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Prevalen
cia
2006 2007 2008

Paula S. Marcotegui
109Capitulo 3. Ecología de los Monogeneos
Gráfico 3.3.7.2. Intensidad media de Absonifibula bychowskyi durante los 3 años de muestreo.
Tabla 3.3.7.1. Resultados del test a posteriori. Rangos promedio y diferencias significativas halladas entre las abundancias de Absonifibula bychowskyi en las diferentes estaciones durante los tres años de muestreo.
Muestreo n Rango Promedio Diferencias significativas
1 15 138,33 (3)(4)(5)(9)(10)(11) 2 10 120,05 (1)(8) 3 15 79,83 1 4 27 99,7 (1)(8) 5 23 84,54 6 11 93,68 7 18 113,81 (3)(5)(11) 8 31 123,56 1 9 10 84 1 10 13 93,54 (1)(8) 11 11 82,86 12 22 106,11
0
0,5
1
1,5
2
2,5
3
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Intensidad
med
ia
2006 2007 2008
Paula S. Marcotegui
110Capitulo 3. Ecología de los Monogeneos
Gráfico 3.3.7.3. Abundancia de A. bychowskyi durante los 3 años de muestreo.
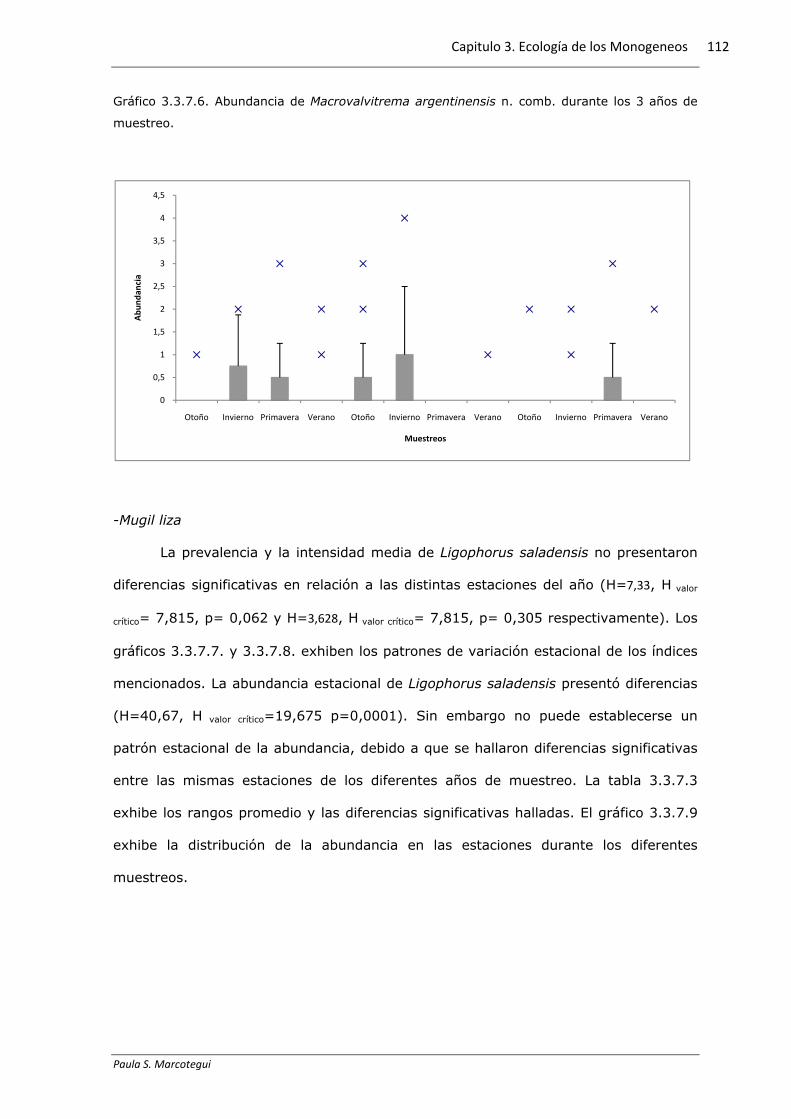
La prevalencia y la intensidad media de M. argentinensis n. comb. no
presentaron diferencias significativas en relación a las distintas estaciones del año
(H=0,465, H valor crítico= 7,815, p= 0,927 y H=4,446, H valor crítico= 7,815, p= 0,217
respectivamente). Los gráficos 3.3.7.4. y 3.3.7.5. exhiben los patrones de variación
estacional de los índices mencionados. La abundancia estacional de M. argentinensis n
sp. no presentó diferencias (H=17,87, H valor crítico=19,675 p=0,085) en los 3 años de
muestreo. La tabla 3.3.7.2 exhibe los rangos promedio. El gráfico 3.3.7.6 exhibe la
distribución de la abundancia en las estaciones durante los diferentes muestreos.
Gráfico 3.3.7.4. Prevalencia de Macrovalvitrema argentinensis n comb. durante los 3 años de
muestreo.
0
1
2
3
4
5
6
7
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
Abu
ndan
cia
Muestreos
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Prevalen
cia
2006 2007 2008

Paula S. Marcotegui
111Capitulo 3. Ecología de los Monogeneos
Gráfico 3.3.7.5. Intensidad media de Macrovalvitrema argentinensis n comb. durante los 3 años
de muestreo.
Tabla 3.3.7.2. Valores obtenidos a partir del Test de Kruskal-Wallis de Macrovalvitrema
argentinensis n comb.
Muestreo n Rango promedio
1 15 99,6 2 10 116,55 3 15 116,2 4 27 94,44 5 23 115,3 6 11 124,68 7 18 87 8 31 99,19 9 10 97,65 10 13 109,73 11 11 116,41 12 22 91,84
0
0,5
1
1,5
2
2,5Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Intensidad
med
ia
2006 2007 2008
Paula S. Marcotegui
112Capitulo 3. Ecología de los Monogeneos
Gráfico 3.3.7.6. Abundancia de Macrovalvitrema argentinensis n. comb. durante los 3 años de
muestreo.
-Mugil liza
La prevalencia y la intensidad media de Ligophorus saladensis no presentaron
diferencias significativas en relación a las distintas estaciones del año (H=7,33, H valor
crítico= 7,815, p= 0,062 y H=3,628, H valor crítico= 7,815, p= 0,305 respectivamente). Los
gráficos 3.3.7.7. y 3.3.7.8. exhiben los patrones de variación estacional de los índices
mencionados. La abundancia estacional de Ligophorus saladensis presentó diferencias
(H=40,67, H valor crítico=19,675 p=0,0001). Sin embargo no puede establecerse un
patrón estacional de la abundancia, debido a que se hallaron diferencias significativas
entre las mismas estaciones de los diferentes años de muestreo. La tabla 3.3.7.3
exhibe los rangos promedio y las diferencias significativas halladas. El gráfico 3.3.7.9
exhibe la distribución de la abundancia en las estaciones durante los diferentes
muestreos.
0
0,5
1
1,5
2
2,5
3
3,5
4
4,5
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
Abu
ndan
cia
Muestreos

Paula S. Marcotegui
113Capitulo 3. Ecología de los Monogeneos
Gráfico 3.3.7.7. Prevalencia de Ligophorus saladensis durante los 3 años de muestreo.
Gráfico 3.3.7.8. Intensidad media de Ligophorus saladensis durante los 3 años de muestreo.
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
1
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Prevalen
cia
0
10
20
30
40
50
60
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Intensidad
med
ia
2006 2007 2008
2006 2007 2008

Paula S. Marcotegui
114Capitulo 3. Ecología de los Monogeneos
Tabla 3.3.7.3. Valores obtenidos a partir del Test de Kruskal-Wallis de Ligophorus saladensis.
Muestreo n Rango promedio Diferencias significativas
1 11 65,45 (5)(9) 2 10 85,95 (4)(5)(6) 3 9 52,11 (5)(9) 4 7 30,14 (2)(5)(7)(9)(10)(11) 5 15 117,33 (1)(2)(3)(4)(6)(7)(8)(10)(11)(12) 6 20 54,58 (2)(5)(9)(10) 7 16 71,37 (4)(5)(9) 8 11 61,91 (5)(9) 9 11 110,55 (1)(3)(4)(6)(7)(8)(11)(12) 10 12 84,96 (4)(5)(6) 11 16 74,41 (4)(5)(9) 12 10 66,6 (5)(9)
Gráfico 3.3.7.9. Abundancia de Ligophorus saladensis durante los 3 años de muestreo.
-Parapimelodus valenciennis
La prevalencia y la intensidad media de M. argentinensis n. comb. no
presentaron diferencias significativas en relación a las distintas estaciones del año
(H=2,35, H valor crítico=7,815, p=0,502 y H=5,635 H valor crítico=7,815, p=0,131
respectivamente). Los gráficos 3.3.7.10. y 3.3.7.11. exhiben los patrones de variación
estacional de los índices mencionados. La abundancia estacional de Demidospermus
annulus n sp. no presentó diferencias (H=14,99, H valor crítico=19,675 p=0,183) en los 3
0
20
40
60
80
100
120
140
160
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
Abu
ndan
cia
Muestreos

Paula S. Marcotegui
115Capitulo 3. Ecología de los Monogeneos
años de muestreo. La tabla 3.3.7.4 exhibe los rangos promedio y las diferencias
significativas halladas. El gráfico 3.3.7.12 exhibe la distribución de la abundancia en
las estaciones durante los diferentes muestreos.
Gráfico 3.3.7.10. Prevalencia de Demidospermus annulus n sp. durante los 3 años de muestreo.
Gráfico 3.3.7.11. Intensidad media de Demidospermus annulus n sp. durante los 3 años de
muestreo.
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Prevalen
cia
0
5
10
15
20
25
30
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Intensidad
med
ia
2006 2007 2008
2006 2007 2008

Paula S. Marcotegui
116Capitulo 3. Ecología de los Monogeneos
Tabla 3.3.7.4. Valores obtenidos a partir del Test de Kruskal-Wallis de Demidospermus annulus
n sp.
Muestreo n Rango promedio1 10 43,95 2 8 60,44 3 16 39,47 4 10 49,5 5 10 71 6 10 59,6 7 10 60,65 8 9 71,39 9 10 71 10 6 61,33 11 9 71,72 12 10 69,3
Gráfico 3.3.7.12. Abundancia de Demidospermus annulus n sp. durante los 3 años de muestreo.
-Odontesthes argentinensis
La prevalencia y la intensidad media de Gyrodactylus mauri n sp. no presentaron
diferencias significativas en relación a las distintas estaciones del año (H=3,4, H valor
crítico=7,815, p=0,333 y H=1,865 H valor crítico=7,815, p=0,601 respectivamente). Los
gráficos 3.3.7.13. y 3.3.7.14. exhiben los patrones de variación estacional de los
índices mencionados. La abundancia estacional de Demidospermus annulus n sp. no
presentó diferencias (H=4,08, H valor crítico=19,675 p=0,967) en los 3 años de muestreo.
La tabla 3.3.7.5 exhibe los rangos promedio y las diferencias significativas halladas. El
gráfico 3.3.7.15 exhibe la distribución de la abundancia en las estaciones durante los
diferentes muestreos.
0
10
20
30
40
50
60
70
80
90
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
Abu
ndan
cia
Muestreos

Paula S. Marcotegui
117Capitulo 3. Ecología de los Monogeneos
Gráfico 3.3.7.13. Prevalencia de Gyrodactylus mauri n sp. durante los 3 años de muestreo.
Gráfico 3.3.7.14. Intensidad media de Gyrodactylus mauri n sp. durante los 3 años de
muestreo.
Tabla 3.3.7.5. Valores obtenidos a partir del Test de Kruskal-Wallis de Gyrodactylus mauri n sp.
Muestreo n Rango promedio 1 7 61,71 2 7 46,14 3 9 55,17 4 18 50 5 16 45,59 6 7 44,14 7 5 51,6 8 5 59,9 9 9 55,11
10 5 45,7 11 5 50,2 12 9 58,89
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Prevalen
cia
0
1
2
3
4
5
6
7
8
9
10
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Otoño
Invierno
Prim
avera
Verano
Intensidad
med
ia
2006 2007 2008
2006 2007 2008

Paula S. Marcotegui
118Capitulo 3. Ecología de los Monogeneos
Gráfico 3.3.7.15. Abundancia de Gyrodactylus mauri n sp. durante los 3 años de muestreo.
3.3.8. Relación de los parámetros poblacionales con las variables ambientales
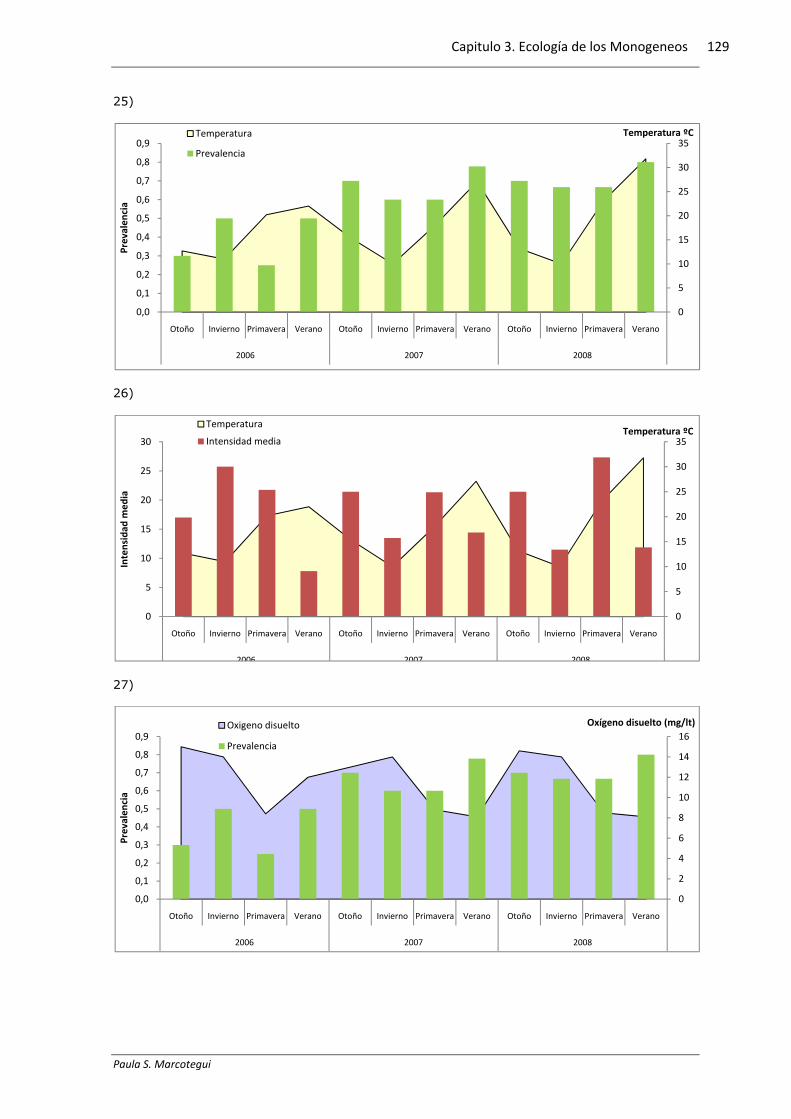
La temperatura (H=10; p=0,018) el oxígeno disuelto (H=7,69; p=0,034) y la
conductividad (H=7,48; p=0,049) presentaron diferencias significativas en las
diferentes estaciones del año, mientras que el pH se mantuvo estable y no presentó
diferencias significativas (H=3,88; p=0,27). La tabla 3.3.8.1 muestra los valores de
temperatura, oxígeno disuelto, conductividad y pH registrados en cada muestreo. El
gráfico 3.3.8.1 exhibe las medianas y los desvíos estándar obtenidos de cada
parámetro analizado en las diferentes estaciones del año.
Tabla 3.3.8.1 Valores de temperatura, oxígeno disuelto, conductividad y pH obtenidos durante
los 3 años de muestreo.
Muestreos Temperatura (ºC) Oxígeno disuelto (mg/lt) Conductividad (mS) pH
2006
Otoño 12,7 15 0,012 8,06
Invierno 11 14 0,104 8,02
Primavera 20,2 8,4 0,067 8,47
Verano 22 12 0,0422 8,01
2007
Otoño 15,4 13 0,0113 8,9
Invierno 10 14 0,106 8,02
Primavera 18 8,8 0,055 8,47
Verano 27,1 8,1 0,105 8,49
2008
Otoño 13,2 14,6 0,0118 8,07
Invierno 10 14 0,106 8,02
Primavera 23,1 8,5 0,057 8,15
Verano 31,8 8,1 0,353 8,96
0
2
4
6
8
10
12
14
16
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
Abu
ndan
cia
Muestreos

Paula S. Marcotegui
119Capitulo 3. Ecología de los Monogeneos
Gráfico 3.3.8.1 Distribución de los valores de cada parámetro analizado en las diferentes estaciones del año.
7,75
12,75
17,75
22,75
27,75
32,75
37,75
Otoño Invierno Primavera Verano
Tempe
ratura (ºC)
Otoño ‐ Verano
7,3
9,3
11,3
13,3
15,3
17,3
Otoño Invierno Primavera VeranoOxígeno
disue
lto (m
g/lt)
Oxígeno
0
0,1
0,2
0,3
0,4
Otoño Invierno Primavera VeranoCond
uctivida
d (m
S/cm
)
Conductividad
8,005
8,205
8,405
8,605
8,805
9,005
9,205
Otoño Invierno Primavera Verano
pH
pH

Paula S. Marcotegui
120Capitulo 3. Ecología de los Monogeneos
La prevalencia de Demidospermus annulus n sp. estuvo correlacionada con la
concentración de oxígeno disuelto. El resto de los parámetros químicos analizados no
presentaron correlación con ninguno de los índices poblacionales para las diferentes
especies de monogeneos estudiados. La Tabla 3.3.8.2 exhibe los valores obtenidos de
correlación de Spearman. El gráfico 3.3.8.2 muestra la distribución de la prevalencia e
intensidad media en relación con los parámetros químicos estudiados.
Tabla 3.3.8.2. Valores de correlación de Spearman entre los distintos parámetros químicos
analizados, la prevalencia de cada especie.
Temperatura Oxígeno Conductividad pH
Valor observado p Valor observado p Valor observado p Valor observado p
A. bychowskyi P 0,06 0,85 0,124 0,702 0,158 0,624 ‐0,117 0,718
M. argentinensis n. comb. P 0,12 0,72 0,08 0,81 ‐0,20 0,54 ‐0,52 0,09
L. saladensis P ‐0,51 0,09 0,44 0,74 ‐0,25 0,44 0,07 0,83
D. annulus n sp. P 0,38 0,23 0,34 0,0,02* 0,20 0,53 0,56 0,06
Gyrodactylus mauri n sp. P 0,19 0,55 0,04 0,89 ‐0,29 0,37 0,26 0,42
*valores significativos. P=prevalencia

Paula S. Marcotegui
121Capitulo 3. Ecología de los Monogeneos
Gráfico 3.3.8.2. Relación de la abundancia, la prevalencia y la intensidad media para cada especie de monogeneo y los diferentes parámetros ambientales: 1-8: Absonifibula bychowskyi, 9-18-p: Macrovalvitrema argentinensis n comb., 19-24 Ligophorus saladensis, 25-32: Demidospermus annulus n sp., 33-40: Gyrodactylus mauri n sp.
1)
2)
3)
0
5
10
15
20
25
30
35
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Temepratura ºC
Prevalen
cia
Temperatura
Prevalencia
0
5
10
15
20
25
30
35
0
0,5
1
1,5
2
2,5
3
3,5
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Temepratura º C
Intensidad
med
ia
Temperatura
Intensidad media
0
2
4
6
8
10
12
14
16
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Oxígeno disuelto (mg/lt)
Prevalen
cia
Oxigeno disuelto
Prevalencia

Paula S. Marcotegui
122Capitulo 3. Ecología de los Monogeneos
4)
5)
6)
0
2
4
6
8
10
12
14
16
0
0,5
1
1,5
2
2,5
3
3,5
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Oxígeno disuelto (mg/lt)
Intensidad
med
ia
Oxigeno disuelto
Intensidad media
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Conductivida mS/cm)
Prevalen
cia
Conductividad
Prevalencia
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0
0,5
1
1,5
2
2,5
3
3,5
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Conductividad (mS/cm)
Intensidad
med
ia
Conductividad
Intensidad media

Paula S. Marcotegui
123Capitulo 3. Ecología de los Monogeneos
7)
8)
9)
7,4
7,6
7,8
8
8,2
8,4
8,6
8,8
9
9,2
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
pH
Prevalen
cia
pH
Prevalencia
7,4
7,6
7,8
8
8,2
8,4
8,6
8,8
9
9,2
0
0,5
1
1,5
2
2,5
3
3,5
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
pH
Intensidad
med
ia
pH
Intensidad media
0
5
10
15
20
25
30
35
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
TemperaturaºC
Prevalen
cia
Temperatura
Prevalencia

Paula S. Marcotegui
124Capitulo 3. Ecología de los Monogeneos
10)
11)
12)
0
5
10
15
20
25
30
35
0
0,5
1
1,5
2
2,5
3
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
TemeperaturaºCIntensidad
med
iaTemperatura
Intensidad media
0
2
4
6
8
10
12
14
16
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Oxígeno disuelto mg/lt)
Prevalen
cia
Oxigeno disuelto Prevalencia
0
2
4
6
8
10
12
14
16
0
0,5
1
1,5
2
2,5
3
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Oxígeno disuelto (mg/lt)
Intensidad
med
ia
Oxigeno disuelto
Intensidad media

Paula S. Marcotegui
125Capitulo 3. Ecología de los Monogeneos
13)
14)
15)
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Conductividad (mS/cm)Prevalen
cia
Conductividad
Prevalencia
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0
0,5
1
1,5
2
2,5
3
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Conductividad (mS/lt)
Intensidad
med
ia
Conductividad
Intensidad media
7,4
7,6
7,8
8
8,2
8,4
8,6
8,8
9
9,2
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
pH
Prevalen
cia
pH
Prevalencia

Paula S. Marcotegui
126Capitulo 3. Ecología de los Monogeneos
16)
17)
18)
7,4
7,6
7,8
8
8,2
8,4
8,6
8,8
9
9,2
0
0,5
1
1,5
2
2,5
3
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
pHIntensidad
med
iapH
Intensidad media
0
5
10
15
20
25
30
35
0
0,2
0,4
0,6
0,8
1
1,2
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Temperatura ºC
Prevalen
cia
Temperatura
Prevalencia
0
5
10
15
20
25
30
35
0
10
20
30
40
50
60
70
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Temperatura ºC
Intensidad
med
ia
Temperatura
Intensidad

Paula S. Marcotegui
127Capitulo 3. Ecología de los Monogeneos
19)
20)
21)
0
2
4
6
8
10
12
14
16
0
0,2
0,4
0,6
0,8
1
1,2
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Oxígeno disuelto (mg/lt)
Prevalen
cia
Oxigeno disuelto
Prevalencia
0
2
4
6
8
10
12
14
16
0
10
20
30
40
50
60
70
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Oxígeno disuelto (mg/lt)
Intensidad
med
ia
Oxigeno disuelto
Intensidad
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0
0,2
0,4
0,6
0,8
1
1,2
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Conductividad (mS/cm)
Prevalen
cia
Conductividad
Prevalencia

Paula S. Marcotegui
128Capitulo 3. Ecología de los Monogeneos
22)
23)
24)
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0
10
20
30
40
50
60
70
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Conductividad (mS/cm)Intensidad
med
iaConductividad Intensidad
7,4
7,6
7,8
8
8,2
8,4
8,6
8,8
9
9,2
0
0,2
0,4
0,6
0,8
1
1,2
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
pH
Prevalen
cia
pH
Prevalencia
7,4
7,6
7,8
8
8,2
8,4
8,6
8,8
9
9,2
0
10
20
30
40
50
60
70
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
pH
Intensidad
med
ia
pH
Intensidad
Paula S. Marcotegui
129Capitulo 3. Ecología de los Monogeneos
25)
26)
27)
0
5
10
15
20
25
30
35
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Temperatura ºCPrevalen
cia
Temperatura
Prevalencia
0
5
10
15
20
25
30
35
0
5
10
15
20
25
30
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Temperatura ºC
Intensidad
med
ia
Temperatura
Intensidad media
0
2
4
6
8
10
12
14
16
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Oxígeno disuelto (mg/lt)
Prevalen
cia
Oxigeno disuelto
Prevalencia

Paula S. Marcotegui
130Capitulo 3. Ecología de los Monogeneos
28)
29)
30)
0
2
4
6
8
10
12
14
16
0
5
10
15
20
25
30
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Oxígeno disuelto (mg/lt)Intensidad
med
iaOxigeno disuelto
Intensidad media
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Conductividad mS/cm)
Prevalen
cia
Conductividad
Prevalencia
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0
5
10
15
20
25
30
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Conductividad (mS/cm)
Intensidad
med
ia
Conductividad
Intensidad media

Paula S. Marcotegui
131Capitulo 3. Ecología de los Monogeneos
31)
32)
33)
7,4
7,6
7,8
8
8,2
8,4
8,6
8,8
9
9,2
0,0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
0,9
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
pH
Prevalen
cia
pH
Prevalencia
7,4
7,6
7,8
8
8,2
8,4
8,6
8,8
9
9,2
0
5
10
15
20
25
30
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
pH
Prevalen
cia
pH
Intensidad media
0
5
10
15
20
25
30
35
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Temperatura ºC
Prevalen
cia
Temperatura
Prevalencia

Paula S. Marcotegui
132Capitulo 3. Ecología de los Monogeneos
34)
35)
36)
0
5
10
15
20
25
30
35
0
2
4
6
8
10
12
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Temperatura ºC
Intensidad
med
ia
Temperatura
Intensidad media
0
2
4
6
8
10
12
14
16
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Oxígeno disuelto (mg/lt)
Prevalen
cia
Oxigeno disuelto
Prevalencia
0
2
4
6
8
10
12
14
16
0
2
4
6
8
10
12
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Oxígeno disuelto (mg/lt)
Intensidad
med
ia
Oxigeno disuelto
Intensidad media

Paula S. Marcotegui
133Capitulo 3. Ecología de los Monogeneos
37)
38)
39)
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Conductividad mS/cm)
Prevalen
cia
Conductividad
Prevalencia
0
0,05
0,1
0,15
0,2
0,25
0,3
0,35
0,4
0
2
4
6
8
10
12
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
Conductividad (mS/cm)
Intensidad
med
ia
Conductividad
Intensidad media
7,4
7,6
7,8
8
8,2
8,4
8,6
8,8
9
9,2
0
0,1
0,2
0,3
0,4
0,5
0,6
0,7
0,8
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
pH
Prevalen
cia
pH
Prevalencia

Paula S. Marcotegui
134Capitulo 3. Ecología de los Monogeneos
40)
Los resultados de los MLG exhiben que ninguno de los parámetros ambientales
analizados tiene un efecto significativo sobre la intensidad media de las diferentes
especies de monogeneos estudiadas. Las tablas 3.3.8.3 a 3.3.8.7 exhiben los
resultados de los análisis.
Tabla 3.3.8.3. Resultados del MLG entre la intensidad media de Absonifibula bychowskyi y los parámetros ambientales.
SS MS F p Intensidad media 0,47 0,47 0,50 0,50 Temperatura 0,20 0,20 0,21 0,66 Oxígeno 0,35 0,35 0,36 0,56
pH 0,08 0,08 0,08 0,78 Conductividad 0,20 0,20 0,21 0,66
Tabla 3.3.8.4. Resultados del MLG entre la intensidad media de Macrovalvitrema argentinensis n. comb. y los parámetros ambientales.
SS MS F p Intensidad media 2,89 2,89 7,30 0,03 Temperatura 0,00 0,00 0,001 0,97 Oxígeno 0,87 0,87 2,19 0,18
pH 2,05 2,05 5,19 0,06 Conductividad 0,17 0,17 0,42 0,54
7,4
7,6
7,8
8
8,2
8,4
8,6
8,8
9
9,2
0
2
4
6
8
10
12
Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano Otoño Invierno Primavera Verano
2006 2007 2008
pH
Intensidad
med
iapH
Intensidad media

Paula S. Marcotegui
135Capitulo 3. Ecología de los Monogeneos
Tabla 3.3.8.5. Resultados del MLG entre la intensidad media de Ligophorus saladensis y los parámetros ambientales.
SS MS F p Intensidad media 111,03 111,028 0,34 0,58 Temperatura 481,73 481,73 1,49 0,26 Oxígeno 164,76 164,76 0,51 0,50
pH 453,96 453,96 1,40 0,27 Conductividad 72,43 72,43 0,22 0,65
Tabla 3.3.8.6. Resultados del MLG entre la intensidad media de Demidospermus annulus n sp. y los parámetros ambientales.
SS MS F p Intensidad media 4,16 4,16 0,11 0,75 Temperatura 55,49 55,49 1,42 0,27 Oxígeno 65,48 65,48 1,67 0,24
pH 10,90 10,90 0,28 0,61 Conductividad 31,49 31,49 0,80 0,40
Tabla 3.3.8.7. Resultados del MLG entre la intensidad media de Gyrodactylus mauri n sp. y los parámetros ambientales.
SS MS F p
Intensidad media 6,95 6,95 0,87 0,38 Temperatura 6,85 6,85 0,86 0,38 Oxígeno 13,50 13,50 1,70 0,23
pH 0,95 0,95 0,12 0,74 Conductividad 3,88 3,88 0,42 0,51
3.4. Discusión
Los patrones de abundancia, prevalencia e intensidad media podrían estar
influenciados por las características propias de las especies y poblaciones parásitas,
las características del hospedador y/o las características del ambiente.
Según Brown (1984), las especies capaces de explotar un amplio rango de
recursos (generalistas) exhiben una gran abundancia a escala local y una amplia
distribución regional. En el caso de los parásitos, Morand & Guégan (2000) no
confirman esta hipótesis y no hallan un efecto de la especificidad por el hospedador en
la abundancia y la prevalencia parasitarias, mientras que Knipes & Janovy (2009),
estudiando 3 especies de monogeneos del género Dactylogyrus, parásitos de
ciprínidos en Nebraska, encuentran evidencias para sustentar la hipótesis de Brown,

Paula S. Marcotegui
136Capitulo 3. Ecología de los Monogeneos
estableciendo que los ectoparásitos generalistas poseen mayores prevalencias y
abundancias que los ectoparásitos especialistas.
En general, las especies de monogeneos de mayor tamaño requieren más
recursos per capita que las especies pequeñas. Debido a que existe una cantidad finita
de espacio y energía disponible en un hábitat determinado, las especies de mayor
tamaño no pueden existir en una alta abundancia. Simková et al. (2000) estudiando la
coexistencia de 9 especies de Gyrodactylus sp. parásitas de Rutilus rutilus, hallaron
que el tamaño corporal de los parásitos estuvo negativamente correlacionado con la
agregación interespecífica, mientras que Poulin & Justine (2008) no registran una
relación entre el tamaño del monogeneo y la abundancia. Todas las especies halladas
durante el desarrollo del presente trabajo manifestaron una alta especificidad por el
hospedador, no encontrándose ninguna de ellas en más de una especie de pez. Esto
podría explicar los bajos valores de prevalencia y abundancia hallados sobre todo en
A. bychowskyi y en M. argentinensis n comb., parásitos de Micropogonias furnieri. En
cuanto al tamaño corporal, estos dos monogeneos poseen un gran tamaño en relación
con el área branquial. Los bajos valores de agregación interespecífica entre A.
bychowskyi y M. argentinensis n comb. concuerdan con los de Simková et al. (2000)
para Dacylogyrus del Hemisferio Norte.
La competencia interespecífica puede jugar un rol importante tanto en la
abundancia como en la determinación de la preferencia por hemibranquias o
especificidad por el sitio de fijación de los monogeneos en el hospedador. Rohde
(1978) estableció que numerosas especies de monogeneos presentan restricción en el
nicho, aun cuando no existe competencia interespecífica, por lo que deben existir
factores intrínsecos los cuales pueden jugar un rol en la determinación de nicho de los
monogeneos. Gutiérrez (1997) explica los patrones de agrupación en monogeneos
parásitos de Siluriformes del Rio de la Plata, identificando la formación de ¨grupos
reproductivos¨ en los filamentos branquiales. La formación de estos grupos
reproductivos genera un patrón de distribución agregada dentro del hospedador y de
esta forma, un espacio libre mayor que el ocupado. Gutiérrez & Martorelli (1999 b)

Paula S. Marcotegui
137Capitulo 3. Ecología de los Monogeneos
concluyen que la distribución diferencial de los parásitos en los arcos branquiales es
resultado tanto del incremento del área branquial como de las diferencias en la
corriente ventilatoria. Gutiérrez (1997) determina como comportamiento común en las
especies de Demidospermus sp. la preferencia por los dos primeros pares de arcos
branquiales. Los resultados del presente trabajo difieren levemente, ya que los
ejemplares analizados presentaron preferencia por los primeros y terceros arcos
branquiales. La preferencia de A. bychowskyi por los dos primeros arcos branquiales
concuerda con los resultados de Martorelli et al. (2007) para esta misma especie en
ejemplares de M. furnieri del mismo sitio de muestreo. Del mismo modo, L. saladensis
presentó preferencia por los primeros arcos branquiales, al igual que lo reportado para
otras especies del mismo género (Fernández Bargiela, 1987).
Generalmente, las especies congéneres exhiben predilecciones diferentes
cuando co-ocurren en un mismo hospedador y ocupan sitios adyacentes, mientras que
especies no relacionadas son capaces de ocupar el mismo sitio (Rohde, 1991). Yan et
al. (2006) demostraron que la competencia intraespecífica también puede influenciar
la distribución de parásitos en los diferentes arcos branquiales. Los resultados del
presenten trabajo sugieren que la competencia no sería un factor que genera la
restricción de nicho, debido a que aun en ausencia de otros monogeneos, el patrón de
ocupación es agrupado y los valores de amplitud del nicho son bajos.
Las comunidades pueden clasificarse como aislacionistas, en las cuales las
interacciones entre especies no resulta importante y las comunidades interactivas,
donde ocurren interacciones entre sus especies componentes (Holmes & Price, 1986).
Para las comunidades aislacionistas se predice la ausencia de saturación y todas las
especies pueden coexistir en esta comunidad debido a que no existe restricción del
nicho. Shorrocks (1996) propone que la coexistencia se ve favorecida por la reducción
de la competencia en base a la utilización de diferentes recursos. Este autor postula
que si las especies están distribuidas de manera que la interacción interespecífica es
menor que la intraspecífica, la coexistencia se ve facilitada. Morand et al. (1999)

Paula S. Marcotegui
138Capitulo 3. Ecología de los Monogeneos
hallaron que la agregación intraespecífica se incrementa en relación con la agregación
interespecífica cuando la riqueza de ectoparásitos se incrementa.
Uno del los factores más reconocidos como influyentes de los patrones de
distribución y de abundancia de los parásitos es el tamaño del hospedador. Sasal &
Morand (1998) postulan que en el caso de los monogeneos parásitos de peces marinos
del Mediterráneo, el tamaño del hospedador es el principal factor que afecta la riqueza
de especies, hallando una relación inversa entre la abundancia y la longitud del
hospedador. Estos autores establecieron que ello podría demostrar que la influencia
generada por el hospedador sobre los parásitos es mayor que la influencia de los
parásitos sobre los hospedadores. Bakke at al. (2002), corroboraron esa hipótesis en
base a que, al aumentar el tamaño del hospedador aumenta la respuesta inmune ante
la infestación parasitaria, lo cual impediría el establecimiento de nuevos individuos. La
correlación negativa registrada en el presente trabajo entre la abundancia de A.
bychowskyi y el peso del hospedador corroboran esta hipótesis. Lawler & Overstreet
(1976) postulan que la alta prevalencia en ejemplares juveniles puede deberse a la
mayor densidad que presentan los peces más pequeños en los estuarios, favoreciendo
este comportamiento la transmisión. Angelescu & Prenski (1987) establecen que en
los peces adultos disminuye progresivamente la capacidad de filtración de los primeros
arcos branquiales, lo cual se traduciría en la disminución de la retención de los estados
infectivos de los parásitos. Martorelli et al. (2007) postularon que la ausencia de A.
bychowskyi en los hospedadores de tamaño mayores a 22 cm podría deberse al
incremento de salinidad. Recientemente, Portes Santos & Timi (2009) postulan que el
ciclo de vida de las especies de la subfamilia Absonifibulininae se desarrolla en
ambientes mixohalinos. La relación inversa entre tamaño del hospedador y la
abundancia de A. bychowskyi concuerda con los resultados de Sardella et al. (1995) y
corroboran la hipótesis de restricción del ciclo de A. bychowskyi a ambientes
mixohalinos postulada por Portes Santos & Timi (2009).
De las restantes especies analizadas en el presente trabajo, M. argentinensis n
comb. y L. saladensis exhibieron correlación positiva entre la abundancia y el tamaño

Paula S. Marcotegui
139Capitulo 3. Ecología de los Monogeneos
del hospedador. Estos resultado concuerdan con lo expuesto por Alves & Luque
(2001), quienes reportan la correlación entre la longitud de M. furnieri y el monogeneo
M. sinaloense en Brasil. Este tipo de relación ha sido explicada en base al aumento de
la superficie branquial, lo cual permite el establecimiento de nuevos ejemplares
(Morand et al., 2002 b; Tombi & Bilong Bilong, 2004). Gutiérrez & Martorelli (1999 b)
y Simková et al. (2006) establecen que al aumentar el tamaño del pez, se incrementa
la corriente ventilatoria. Gutiérrez & Martorelli (1999 b) y Gutiérrez (1997), no hallan
una relación entre la longitud y el peso de siluriformes del Río de La Plata y la
abundancia y la riqueza específica de monogeneos del género Demidospermus. Los
resultados del presente trabajo concuerdan con los hallazgos de estos autores, ya que
no ha sido hallada una correlación entre el tamaño del hospedador y la abundancia de
D. annulus n sp.
En relación a la distribución espacial en las diferentes longitudes de los
hospedadores, Yang et al. (2006) atribuyen las variaciones en la distribución de los
monogeneos Polyabris mamaevi Ogawa & Egusa, 1980 y Tetrancistrum nebulosi
Young, 1967 parásitos de Signatus fuscescens, principalmente al aumento de la
intensidad media de los monogeneos y estableciendo que no hay un efecto de la
longitud del hospedador sobre la distribución de estos parásitos. Los resultados de A.
bychowskyi en el presente trabajo concuerdan con esta hipótesis. Mientras que, en el
caso de L. saladensis, al aumentar la longitud de M. liza aumenta preferencialmente la
intensidad en los dos primeros arcos branquiales. Kadlec et al. (2003) atribuyen estas
variaciones a las variaciones en el tamaño del hospedador, teniendo en cuenta que al
aumentar el mismo, aumenta la superficie branquial, especialmente en los dos
primero arcos que en general son de mayor tamaño.
Los parámetros ambientales analizados se encuentran dentro de los rangos
previamente reportados por otros autores para el área de muestreo (Gabellone et al.,
2008; Schenone et al., 2008).
Existen numerosas hipótesis para explicar las variaciones de abundancia y
prevalencia en relación a las características particulares del ambiente. Raymond et al.

Paula S. Marcotegui
140Capitulo 3. Ecología de los Monogeneos
(2006) explican las variaciones en la abundancia de monogeneos en base a las
concentraciones de oxígeno disuelto. El oxígeno disuelto puede contribuir a la
selección de hábitat de Afrodyplozoon polycotyleus parásito de peces del genero
Barbus en Uganda. De acuerdo con este autor, las especies de monogeneos tolerantes
a la hipoxia, poseen ventajas competitivas en un hábitat con bajas concentraciones de
oxígeno disuelto, si los posibles parásitos branquiales de ese pez son sensibles a los
niveles de oxígeno disuelto. Esto también puede relacionarse con la corriente
ventilatoria, ya que cuanto menor es la concentración de oxígeno, la corriente
ventilatoria aumenta y este aumento favorece la tasa de transmisión de estos
monogeneos. En las especies analizadas durante el presente trabajo, la única
correlación positiva hallada fue entre Demidospermus annulus n sp. y la concentración
de oxígeno. Sin embargo, se necesitan nuevos estudios en áreas con mayor diferencia
en la concentración de oxígeno disuelto para corroborar estos resultados.
Los parámetros poblacionales no presentaron diferencias estacionales para
ninguna de las especies de monogeneos analizadas. Gutiérrez (1997) halló una fuerte
relación entre la estación y la abundancia, el número de sectores ocupados, la
diversidad, y la frecuencia de los grupos reproductivos por pez de las especies de
Demidospermus sp. parásitos de siluriformes del Río de la Plata. Los diferentes
resultados hallados obtenidos durante el presente trabajo, podrían deberse a las
características particulares de cada especie.





























