Embed Size (px)
Citation preview

CARRERA DE ESPECIALIZACION EN BIOTECNOLOGIA INDUSTRIAL
FCEyN-INTI
Materia de Articulación CEBI_A4
Química Biológica
Docente a cargo:Marta Blanca Mazzetti
CEBI_A4_5b : Trasporte de electrones y fosforilación oxidativa
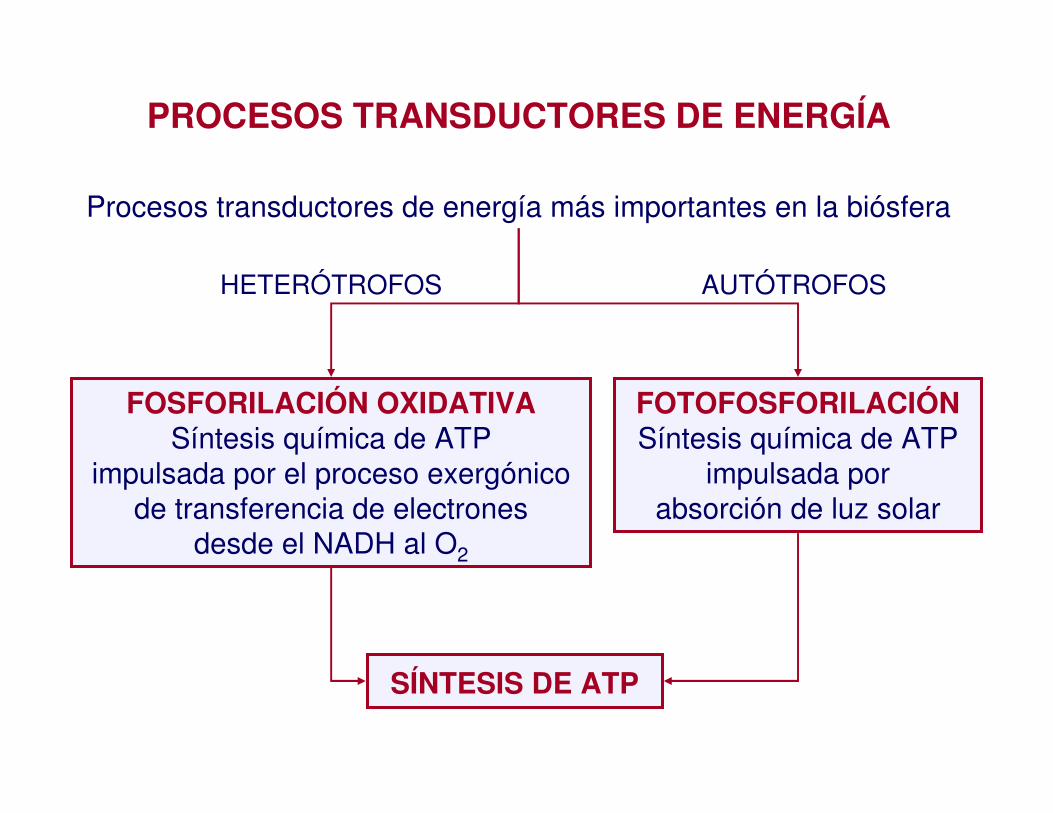
PROCESOS TRANSDUCTORES DE ENERGÍA
FOSFORILACIÓN OXIDATIVASíntesis química de ATP
impulsada por el proceso exergónicode transferencia de electrones
desde el NADH al O2
FOTOFOSFORILACIÓNSíntesis química de ATP
impulsada por absorción de luz solar
Procesos transductores de energía más importantes en la biósfera
HETERÓTROFOS AUTÓTROFOS
SÍNTESIS DE ATP

Respiración Celular
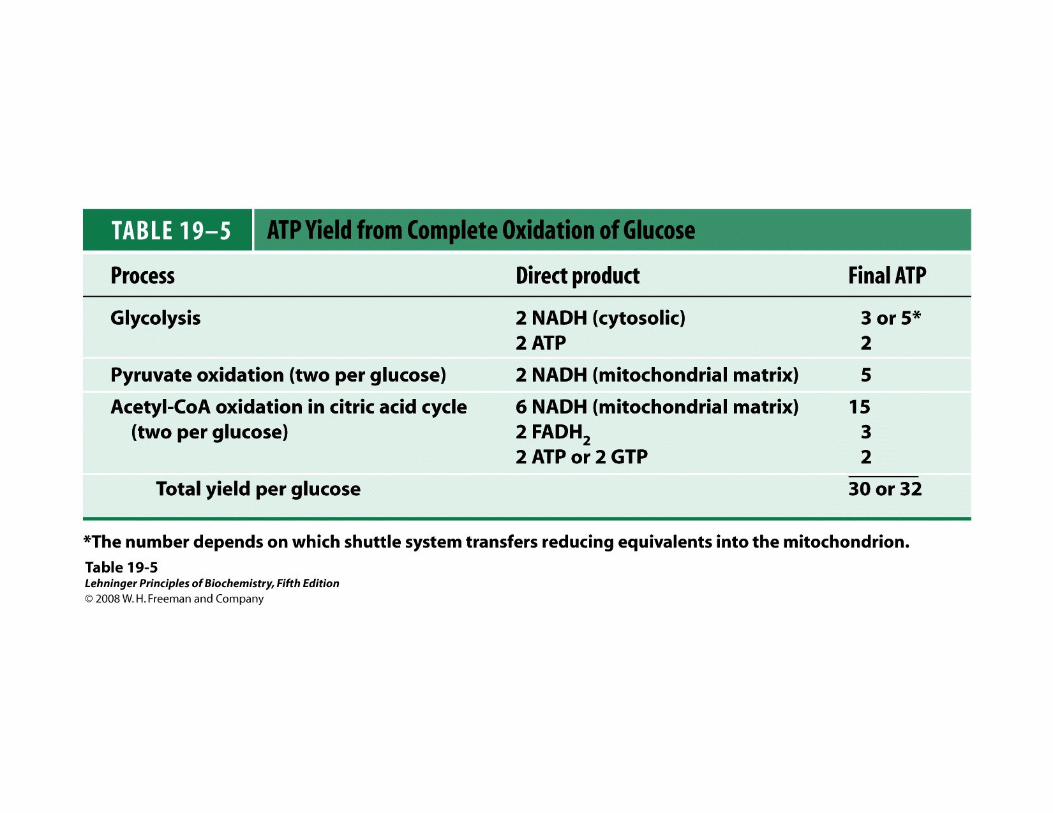
Es el conjunto de reacciones en las cuales el ác. pirúvico producido por la glucólisis se desdobla a CO2 y H2O y se producen 30 ATP.
En las células eucariontes la respiración se realiza en la mitocondria.
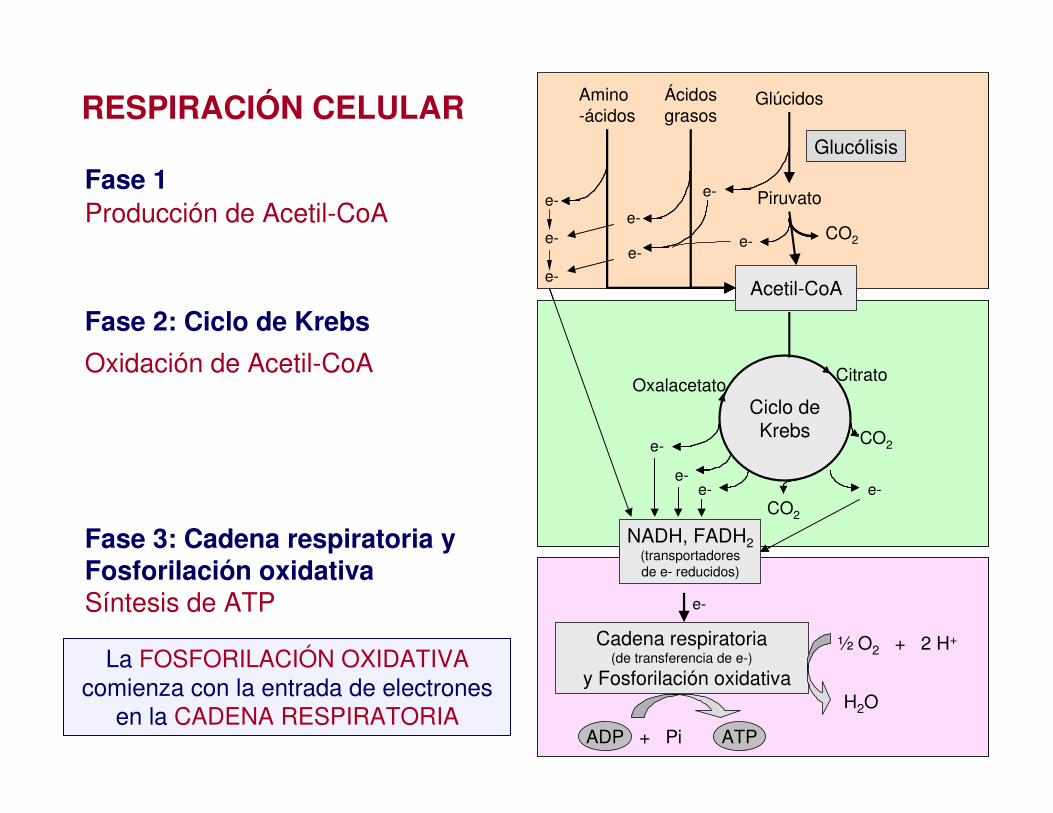
RESPIRACIÓN CELULAR
Fase 1
Fase 2: Ciclo de Krebs
Fase 3: Cadena respiratoria yFosforilación oxidativa
Producción de Acetil-CoA
Oxidación de Acetil-CoA
Síntesis de ATP
Amino-ácidos
Ácidosgrasos
Glúcidos
Glucólisis
Piruvato
CO2
Acetil-CoA
Cadena respiratoria(de transferencia de e-)
y Fosforilación oxidativa
Ciclo deKrebs
CitratoOxalacetato
CO2
CO2
NADH, FADH2(transportadoresde e- reducidos)
ADP + Pi ATP
½ O2 + 2 H+
H2O
e-
e-e-
e-
e-
e-e- e-
e-
e-e-
e-
Amino-ácidos
Ácidosgrasos
Glúcidos
Glucólisis
Piruvato
CO2
Acetil-CoA
Cadena respiratoria(de transferencia de e-)
y Fosforilación oxidativa
Ciclo deKrebs
CitratoOxalacetato
CO2
CO2
NADH, FADH2(transportadoresde e- reducidos)
ADP + Pi ATP
½ O2 + 2 H+
H2O
e-
e-e-
e-
e-
e-e- e-
e-
e-e-
e-
La FOSFORILACIÓN OXIDATIVAcomienza con la entrada de electrones
en la CADENA RESPIRATORIA

La fosforilación oxidativa, ocurre en lamitocondria, y se refiere a la síntesis
química de ATP (reacción endergónica)
impulsada por el proceso exergónico de transferencia de electrones desde el
NADH al O2 (cadena respiratoria).
FOSFORILACIÓN OXIDATIVA
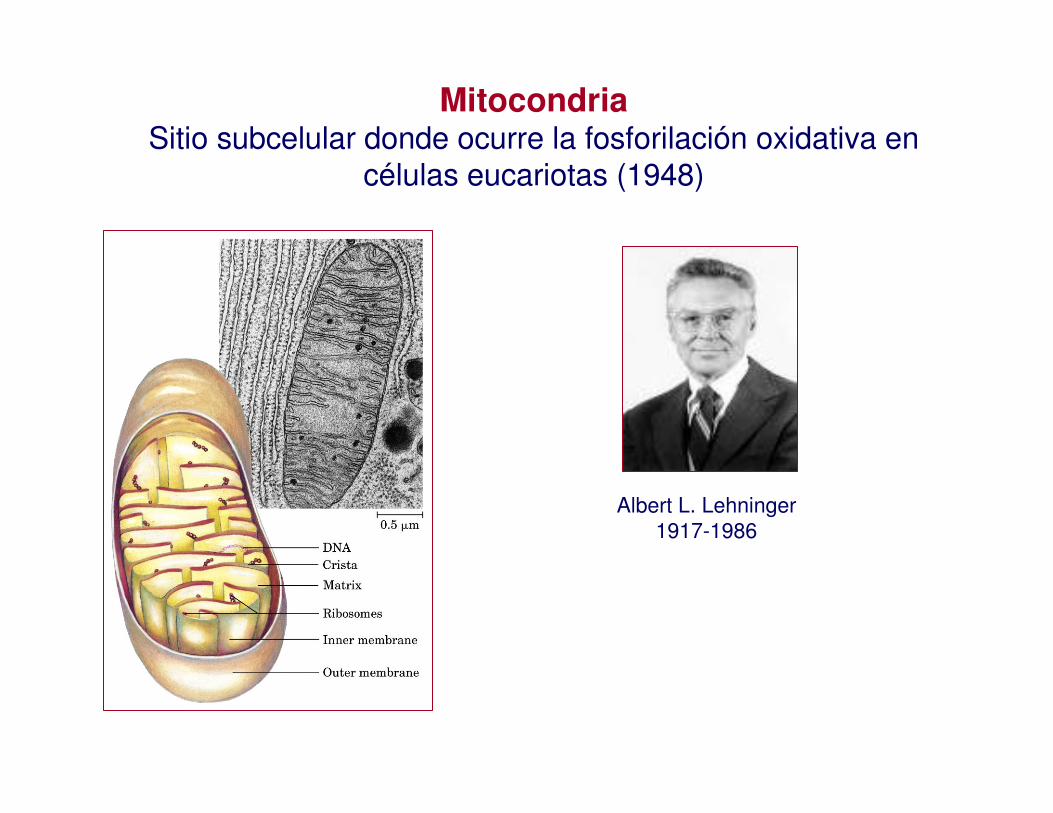
MitocondriaSitio subcelular donde ocurre la fosforilación oxidativa en
células eucariotas (1948)
Albert L. Lehninger1917-1986
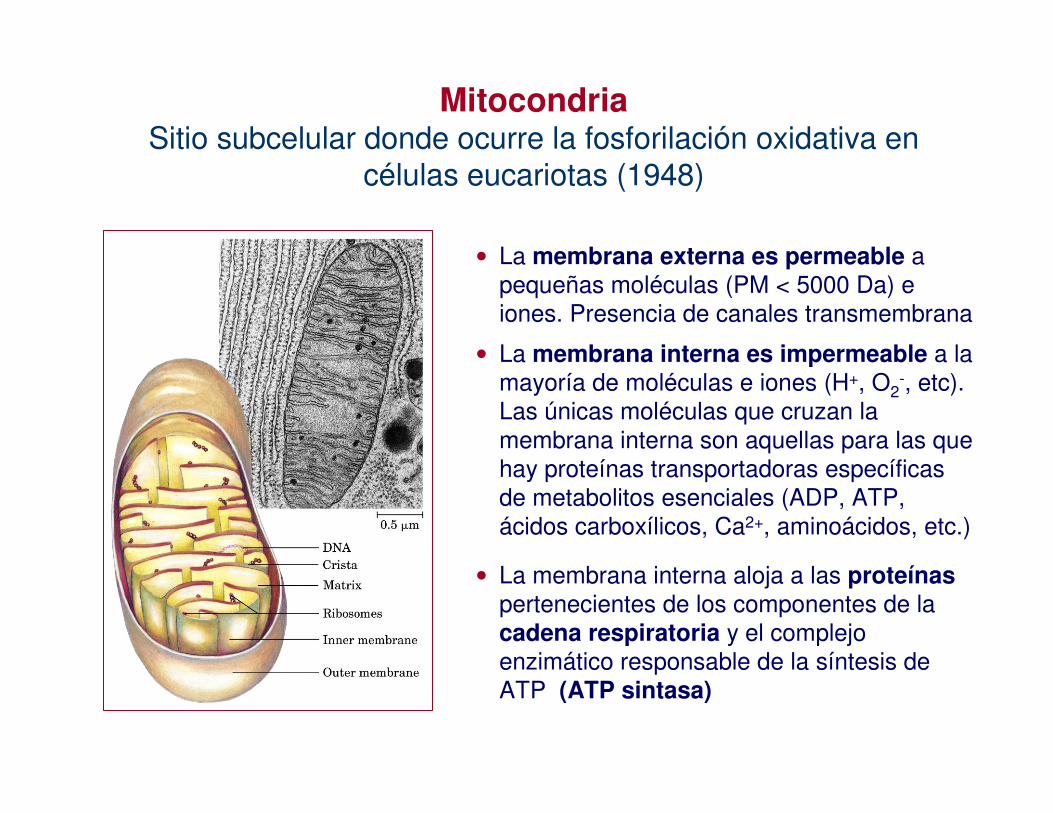
• La membrana externa es permeable a pequeñas moléculas (PM < 5000 Da) e iones. Presencia de canales transmembrana
• La membrana interna es impermeable a la mayoría de moléculas e iones (H+, O2
-, etc). Las únicas moléculas que cruzan la membrana interna son aquellas para las que hay proteínas transportadoras específicas de metabolitos esenciales (ADP, ATP, ácidos carboxílicos, Ca2+, aminoácidos, etc.)
• La membrana interna aloja a las proteínas pertenecientes de los componentes de la cadena respiratoria y el complejo enzimático responsable de la síntesis de ATP (ATP sintasa)
MitocondriaSitio subcelular donde ocurre la fosforilación oxidativa en
células eucariotas (1948)

La fosforilación oxidativa, ocurre en la mitocondria, y se refiere a la síntesis
química de ATP (reacción endergónica)
impulsada por el proceso exergónico de transferencia de electrones desde el
NADH al O2 (cadena respiratoria).
FOSFORILACIÓN OXIDATIVA

• Consta de una serie de transportadores de electrones, la mayoría proteínas integrales de la membrana interna, con grupos prostéticos capaces de aceptar y/o ceder 1 ó 2 electrones
• Cada componente de la cadena, acepta electrones del transportador precedente y se los transfiere al siguiente, en una secuencia específica
• Los transportadores electrónicos mitocondriales funcionan dentro de complejos proteicos ordenados en serie
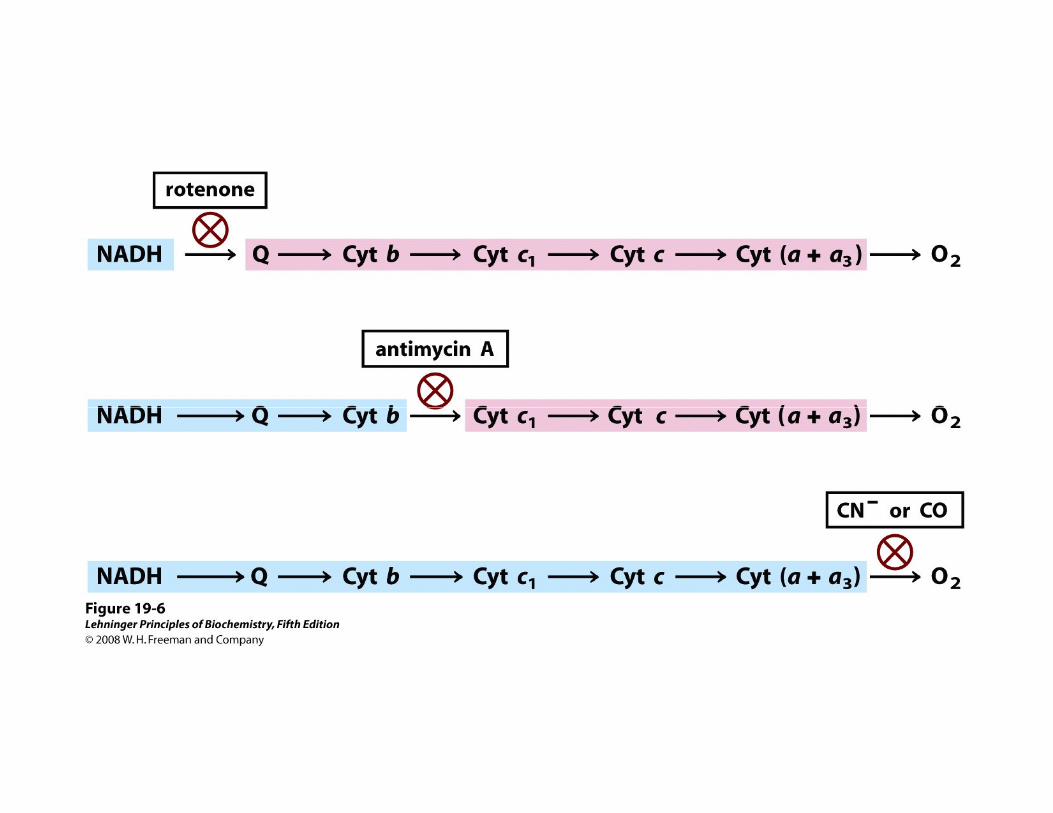
CADENA DE TRANSPORTE DE ELECTRONES MITOCONDRIALO CADENA RESPIRATORIA MITOCONDRIAL
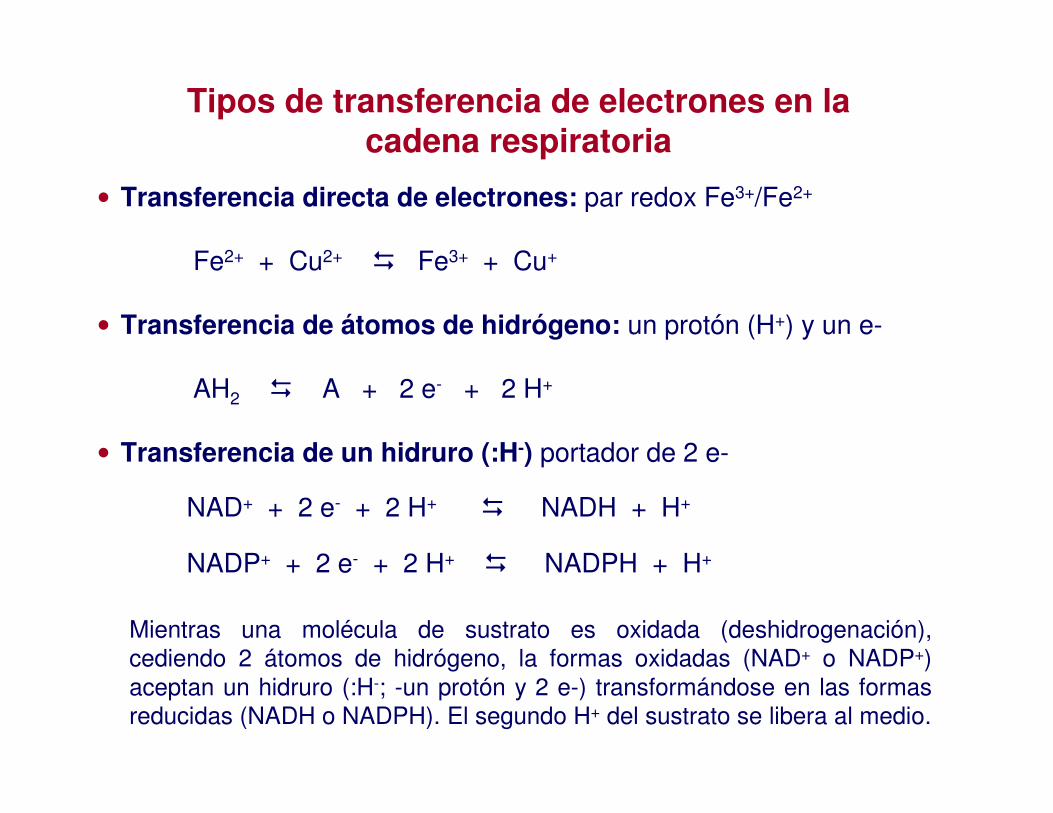
• Transferencia directa de electrones: par redox Fe3+/Fe2+
Fe2+ + Cu2+ � Fe3+ + Cu+
• Transferencia de átomos de hidrógeno: un protón (H+) y un e-
AH2 � A + 2 e- + 2 H+
• Transferencia de un hidruro (:H-) portador de 2 e-
Tipos de transferencia de electrones en la cadena respiratoria
Mientras una molécula de sustrato es oxidada (deshidrogenación), cediendo 2 átomos de hidrógeno, la formas oxidadas (NAD+ o NADP+) aceptan un hidruro (:H-; -un protón y 2 e-) transformándose en las formas reducidas (NADH o NADPH). El segundo H+ del sustrato se libera al medio.
NADP+ + 2 e- + 2 H+ � NADPH + H+
NAD+ + 2 e- + 2 H+ � NADH + H+

Tipos de transferencia de electrones en la cadena respiratoria
Equivalente de reducciónTérmino utilizado para designar a un equivalente
electrónico simple (un electrón transferido) que participa en una reacción de oxidación-reducción, sin importar si este equivalente está en forma de electrón per sé, átomo de hidrógeno ó ión hidruro

• Transportadores universales de electrones- Nucleótidos de nicotinamida (NAD+ y NADP+) - Nucleótidos de flavina (FMN y FAD)
• Otros grupos transportadores de electrones- Ubiquinona ó Coenzima Q (benzoquinona)- Citocromos- Hierro-sulfo
proteínas
Transportadores de electrones en la cadena respiratoria

• Transportadores universales de electrones- Nucleótidos de nicotinamida (NAD+ y NADP+) - Nucleótidos de flavina (FMN y FAD)
• Otros grupos transportadores de electrones- Ubiquinona ó Coenzima Q (benzoquinona)- Citocromos- Hierro-sulfo
proteínas
Transportadores de electrones en la cadena respiratoria
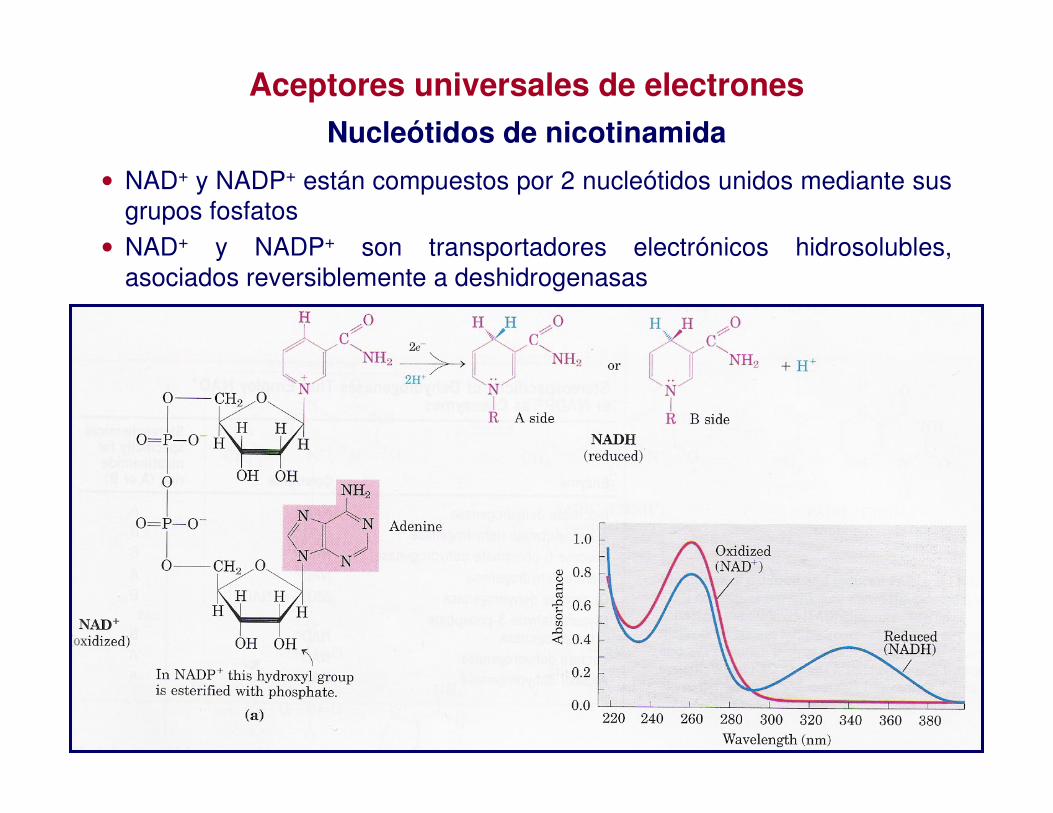
Aceptores universales de electrones
Nucleótidos de nicotinamida
• NAD+ y NADP+ están compuestos por 2 nucleótidos unidos mediante sus grupos fosfatos
• NAD+ y NADP+ son transportadores electrónicos hidrosolubles, asociados reversiblemente a deshidrogenasas
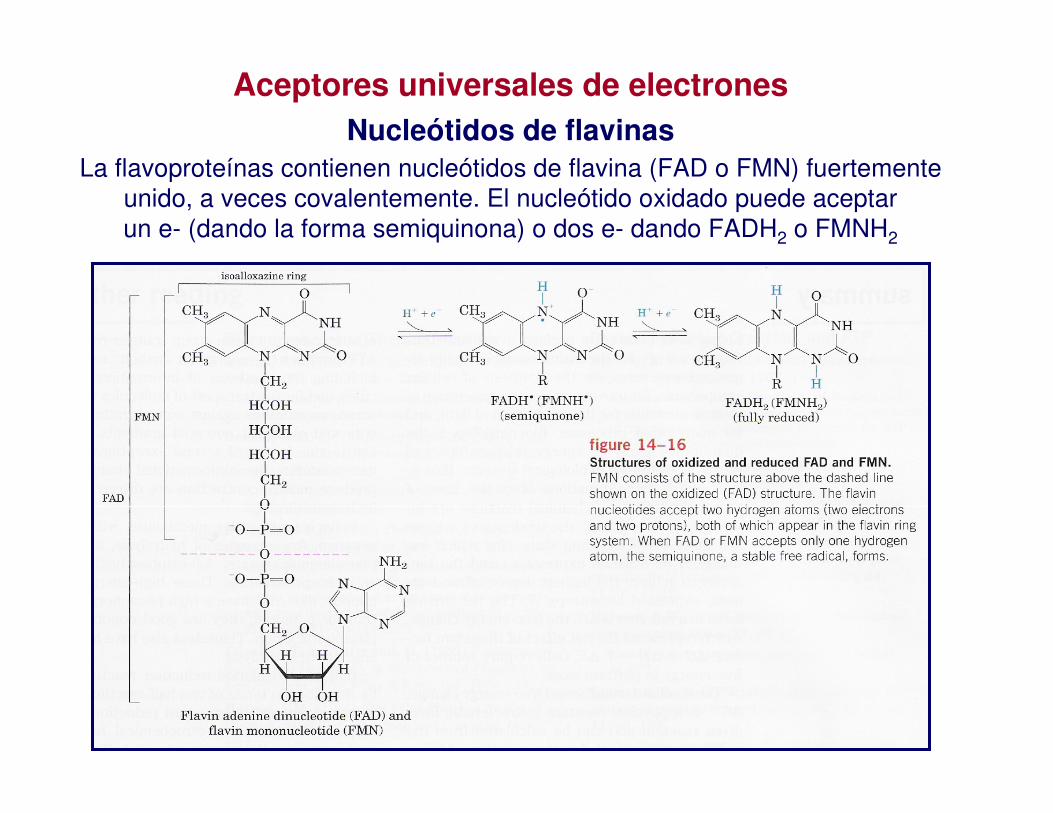
La flavoproteínas contienen nucleótidos de flavina (FAD o FMN) fuertemente unido, a veces covalentemente. El nucleótido oxidado puede aceptar un e- (dando la forma semiquinona) o dos e- dando FADH2 o FMNH2
Aceptores universales de electronesNucleótidos de flavinas

• Transportadores universales de electrones- Nucleótidos de nicotinamida (NAD+ y NADP+) - Nucleótidos de flavina (FMN y FAD)
• Otros grupos transportadores de electrones- Ubiquinona ó Coenzima Q (benzoquinona)- Citocromos- Hierro-sulfo
proteínas
Transportadores de electrones en la cadena respiratoria
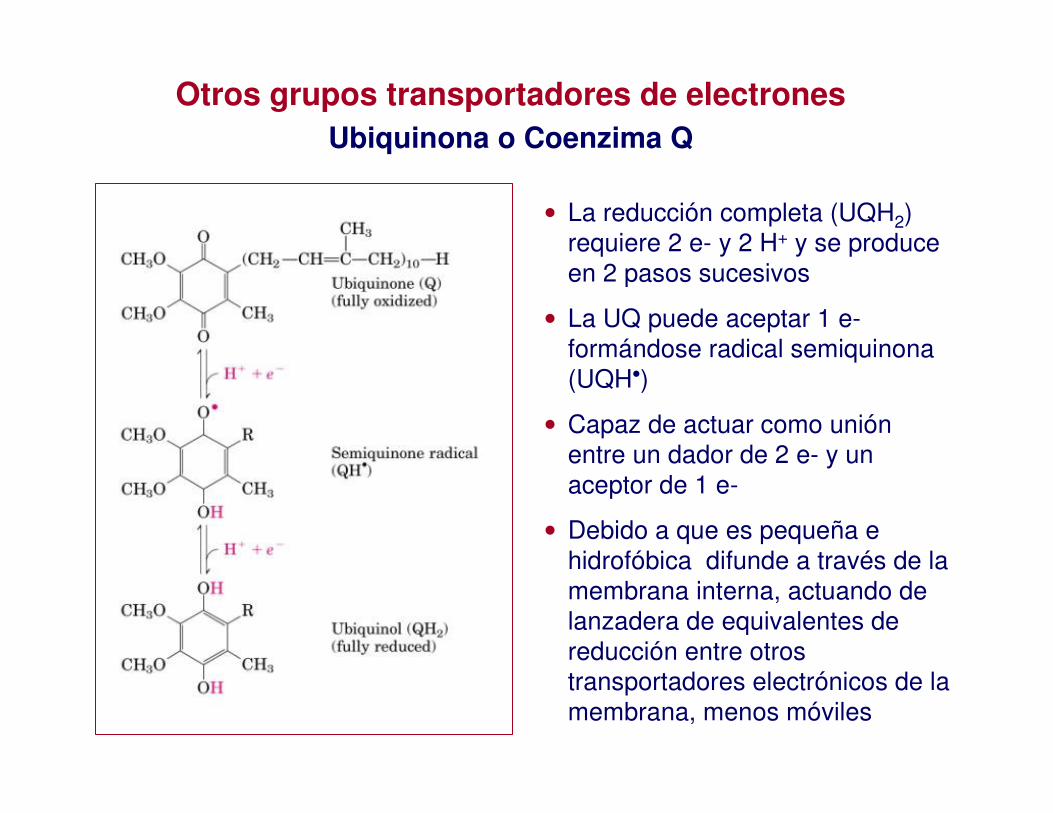
Otros grupos transportadores de electronesUbiquinona o Coenzima Q
• La reducción completa (UQH2) requiere 2 e- y 2 H+ y se produce en 2 pasos sucesivos
• La UQ puede aceptar 1 e-formándose radical semiquinona(UQH•)
• Capaz de actuar como unión entre un dador de 2 e- y un aceptor de 1 e-
• Debido a que es pequeña e hidrofóbica difunde a través de la membrana interna, actuando de lanzadera de equivalentes de reducción entre otros transportadores electrónicos de la membrana, menos móviles
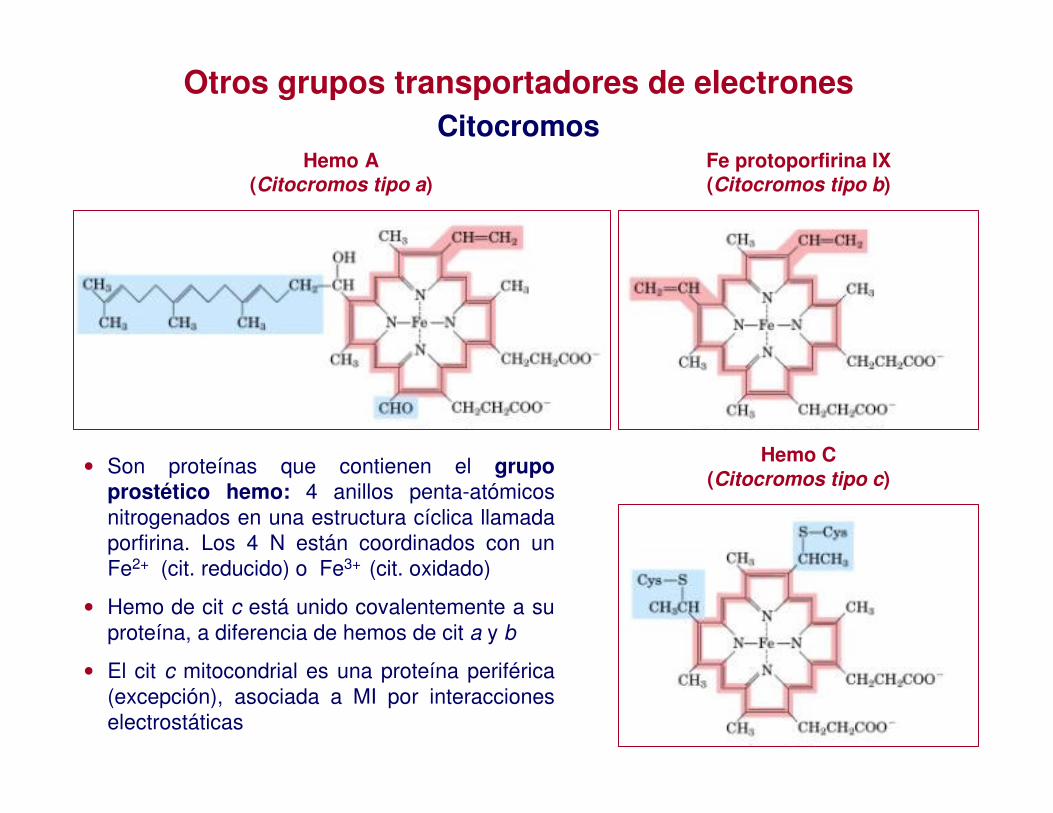
Otros grupos transportadores de electronesCitocromos
• Son proteínas que contienen el grupo prostético hemo: 4 anillos penta-atómicos nitrogenados en una estructura cíclica llamada porfirina. Los 4 N están coordinados con un Fe2+ (cit. reducido) o Fe3+ (cit. oxidado)
• Hemo de cit c está unido covalentemente a su proteína, a diferencia de hemos de cit a y b
• El cit c mitocondrial es una proteína periférica (excepción), asociada a MI por interacciones electrostáticas
Hemo A(Citocromos tipo a)
Hemo C(Citocromos tipo c)
Fe protoporfirina IX(Citocromos tipo b)
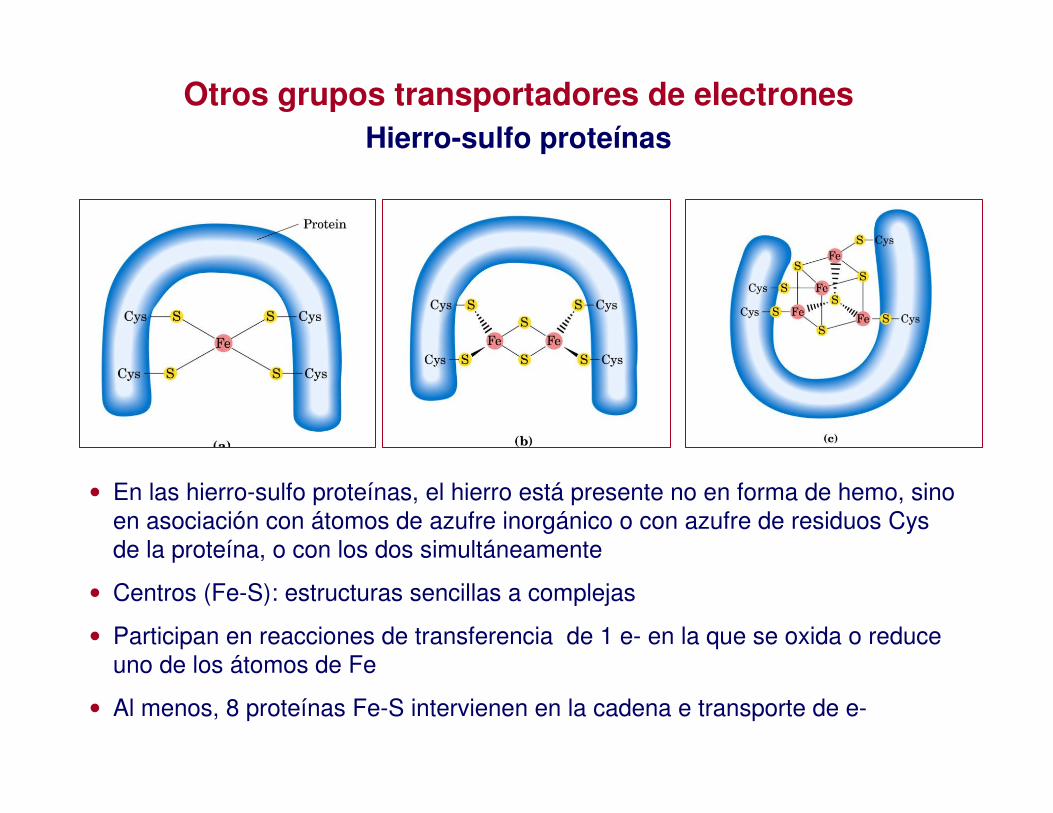
Otros grupos transportadores de electronesHierro-sulfo proteínas
• En las hierro-sulfo proteínas, el hierro está presente no en forma de hemo, sino en asociación con átomos de azufre inorgánico o con azufre de residuos Cysde la proteína, o con los dos simultáneamente
• Centros (Fe-S): estructuras sencillas a complejas
• Participan en reacciones de transferencia de 1 e- en la que se oxida o reduce uno de los átomos de Fe
• Al menos, 8 proteínas Fe-S intervienen en la cadena e transporte de e-

Potenciales de reducción estándar de los transportadores de la cadena respiratoria mitocondrial
Hemi-reacción Eo’ (mV)
NAD+ + 2H+ + 2e- → NADH + H+ - 320
FMN + 2H+ + 2e- → FMNH2 - 290
Fe3+-S-proteína (Complejo I) + 1e- → Fe2+-S-proteína - 270
Fumarato + 2H+ + 2e- → Succinato - 31
FAD + 2H+ + 2e- → FADH2 - 10
Fe3+-S-proteína (Complejo II) + 1e- → Fe2+-S-proteína 20
Ubiquinona + 2H+ + 2e- → Ubiquinol (UQH2) 50
Citocromo bk (Fe3+) + 1e- → Citocromo bk (Fe2+) 77
Citocromo bT (Fe3+) + 1e- → Citocromo bT (Fe2+) 190
Citocromo c1 (Fe3+) + 1e- → Citocromo c1 (Fe2+) 220
Citocromo c (Fe3+) + 1e- → Citocromo c (Fe2+) 254
Citocromo a (Fe3+) + 1e- → Citocromo a (Fe2+) 290
Citocromo a3 (Fe3+) + 1e- → Citocromo a3 (Fe2+) 550
½ O2 + 2H+ + 2e- → H2O 816
Los electrones fluyen espontáneamente desde los transportadores de Eo’ más bajo hacia los transportadores con Eo’ más elevado. En la tabla, los de más arriba reducen a los de mas abajo.
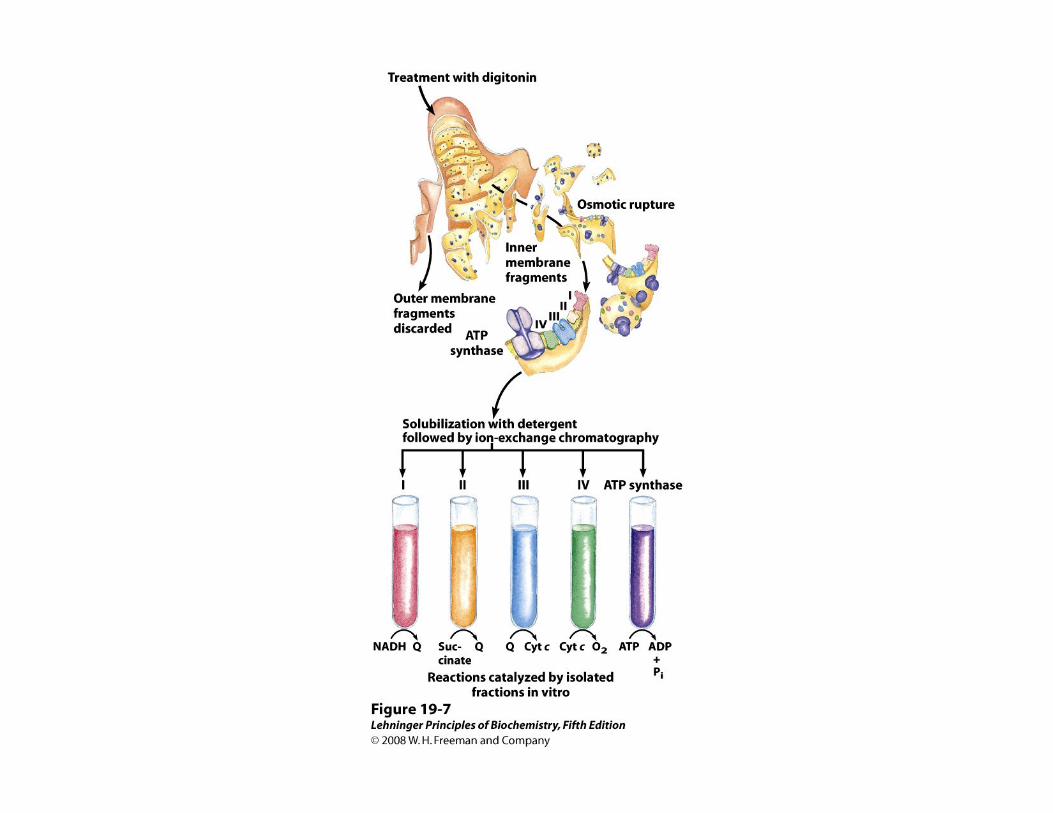
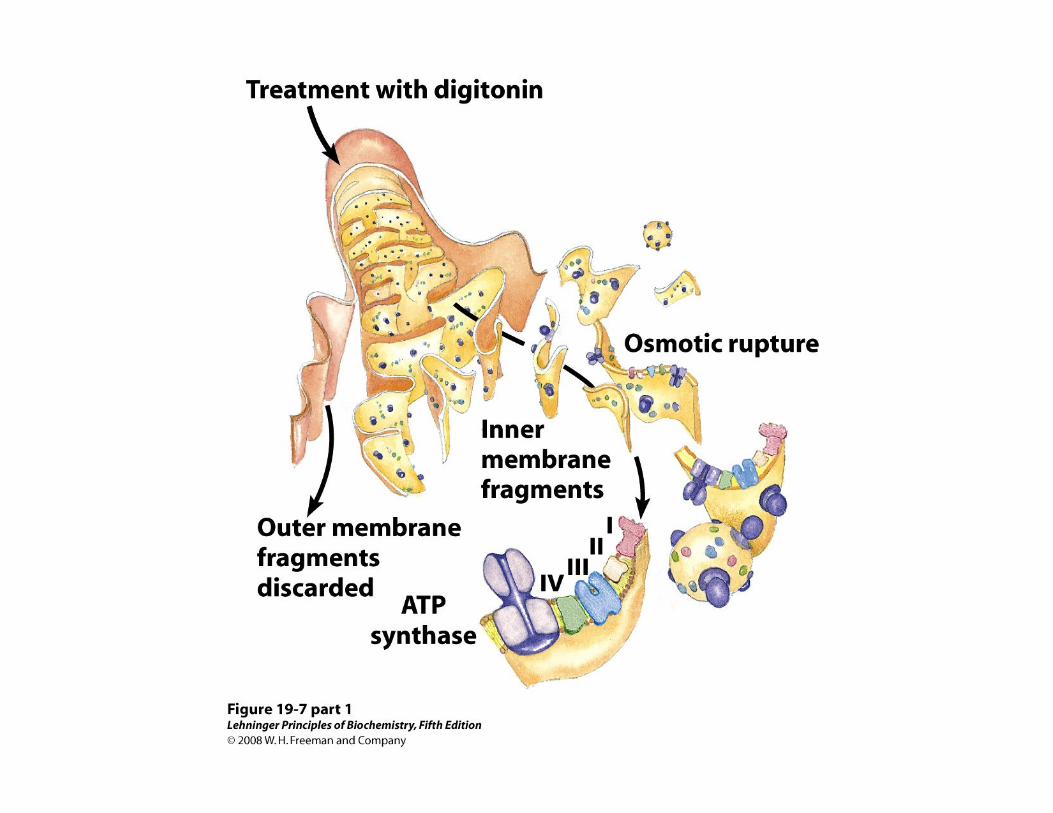

Los transportadores mitocondriales de electrones funcionan dentro de complejos proteicos
H2O
Malato
NAD FMN - (Fe-S)6
Glutamato
UQ
Succinato
(Fe-S)3
cit bK . cit bT (Fe-S) . cit c1 cit c cit aa3
O2
FAD
I
II
IIIIV
• Cada componente de la cadena, acepta electrones del transportador precedente y se los transfiere al siguiente, en una secuencia específica
• Los transportadores electrónicos mitocondriales funcionan dentro de complejos proteicos ordenados en serie
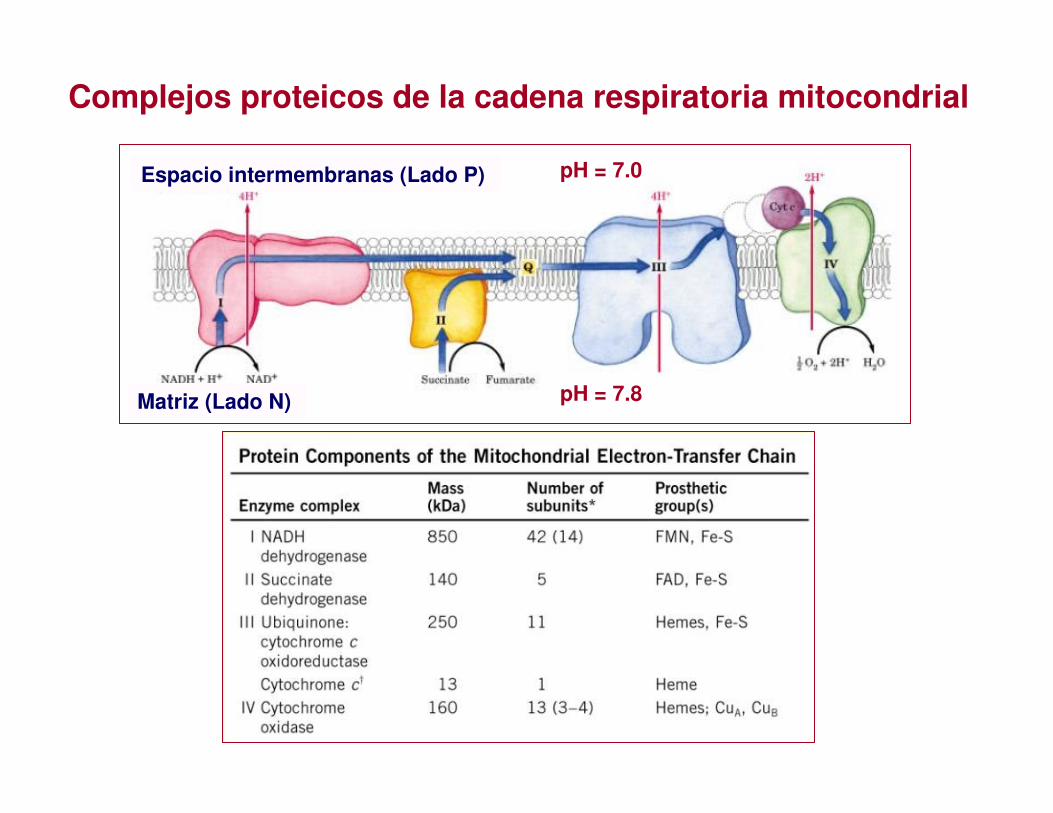
Complejos proteicos de la cadena respiratoria mitocondrial
pH = 7.0 Espacio intermembranas (Lado P)
pH = 7.8 Matriz (Lado N)
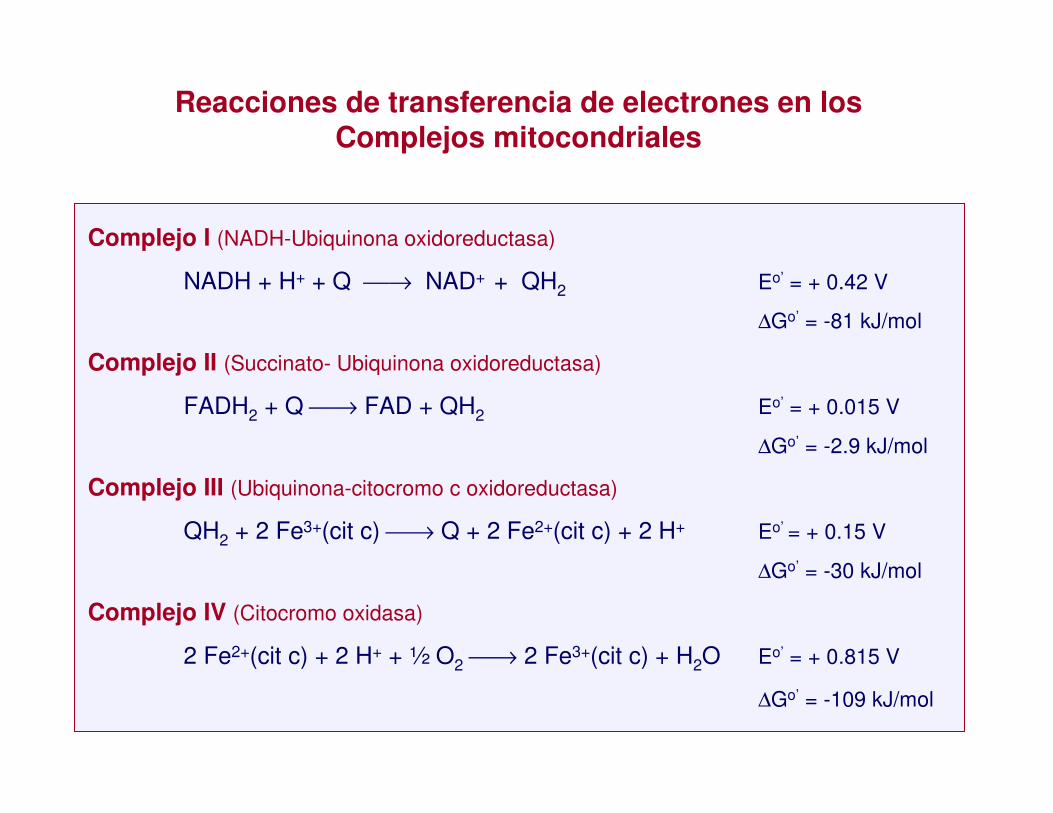
Reacciones de transferencia de electrones en los Complejos mitocondriales
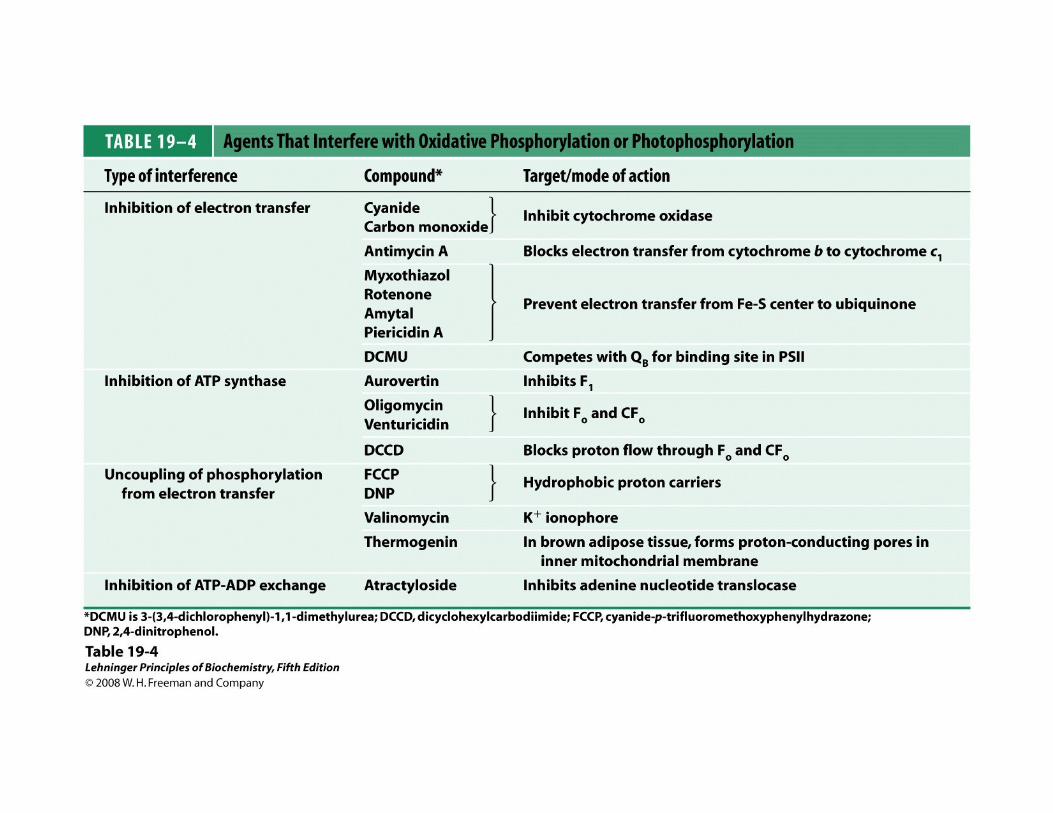
Complejo I (NADH-Ubiquinona oxidoreductasa)
NADH + H+ + Q → NAD+ + QH2 Eo’ = + 0.42 V
∆Go’ = -81 kJ/mol
Complejo II (Succinato- Ubiquinona oxidoreductasa)
FADH2 + Q → FAD + QH2 Eo’ = + 0.015 V
∆Go’ = -2.9 kJ/mol
Complejo III (Ubiquinona-citocromo c oxidoreductasa)
QH2 + 2 Fe3+(cit c) → Q + 2 Fe2+(cit c) + 2 H+ Eo’ = + 0.15 V
∆Go’ = -30 kJ/mol
Complejo IV (Citocromo oxidasa)
2 Fe2+(cit c) + 2 H+ + ½ O2 → 2 Fe3+(cit c) + H2O Eo’ = + 0.815 V
∆Go’ = -109 kJ/mol

La fosforilación oxidativa, ocurre en la mitocondria, y se refiere a la síntesis
química de ATP (reacción endergónica)
impulsada por el proceso exergónico de transferencia de electrones desde el
NADH al O2 (cadena respiratoria).
FOSFORILACIÓN OXIDATIVA
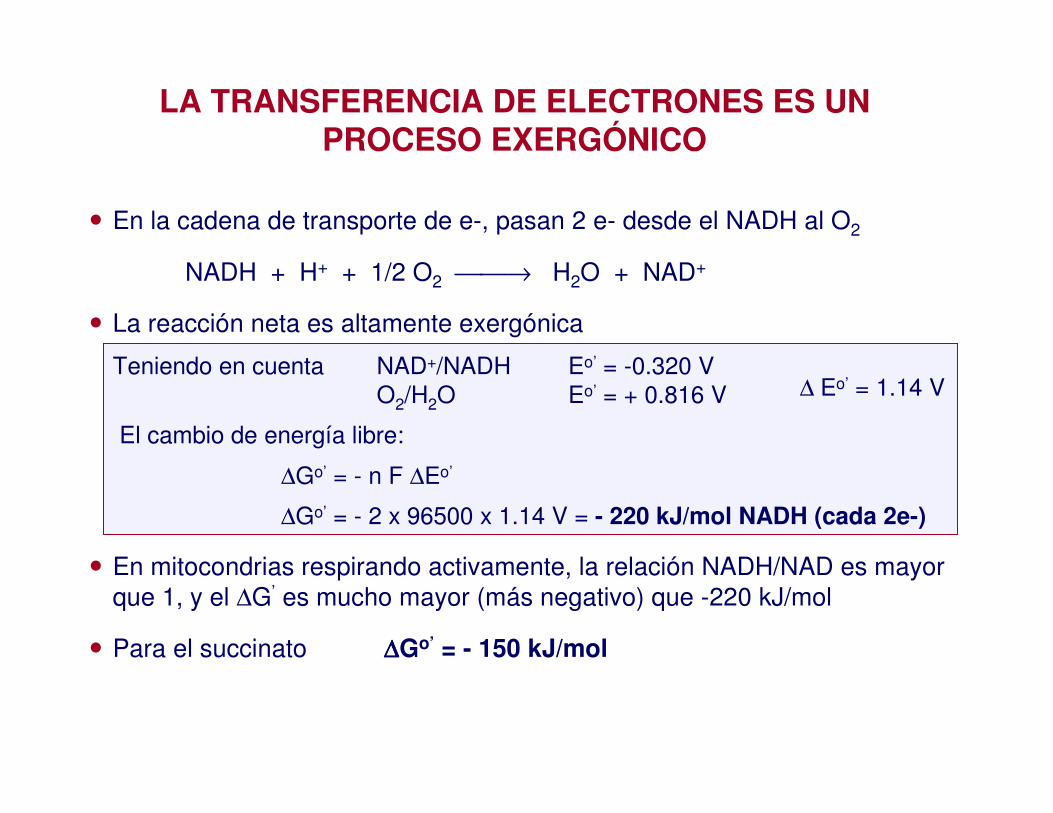
LA TRANSFERENCIA DE ELECTRONES ES UN PROCESO EXERGÓNICO
• En la cadena de transporte de e-, pasan 2 e- desde el NADH al O2
NADH + H+ + 1/2 O2 → H2O + NAD+
• La reacción neta es altamente exergónica
Teniendo en cuenta NAD+/NADH Eo’ = -0.320 V O2/H2O Eo’ = + 0.816 V
El cambio de energía libre:
∆Go’ = - n F ∆Eo’
∆Go’ = - 2 x 96500 x 1.14 V = - 220 kJ/mol NADH (cada 2e-)
• En mitocondrias respirando activamente, la relación NADH/NAD es mayor que 1, y el ∆G’ es mucho mayor (más negativo) que -220 kJ/mol
• Para el succinato ∆∆∆∆Go’ = - 150 kJ/mol
∆ Eo’ = 1.14 V

• La síntesis de ATP, a partir de ADP y Pi, es un proceso endergónico:
ADP + Pi → ATP + H2O ∆∆∆∆Go’ = 30.5 kJ/mol
• En condiciones fisiológicas, en eritrocitos[ATP] = 2.25 mM[ADP] = 0.25 mM y [Pi] = 1.65 mMA 25 oC, pH = 7.0 ∆∆∆∆G’ = 51.8 kJ/mol
En las condiciones celulares, la oxidación mitocondrial de NADH o de succinato libera una energía superior a la
necesaria para la síntesis de ATP.
La fosforilación oxidativa mitocondrial no plantea un problema termodinámico
LA SÍNTESIS DE ATP ES UN PROCESO ENDERGÓNICO
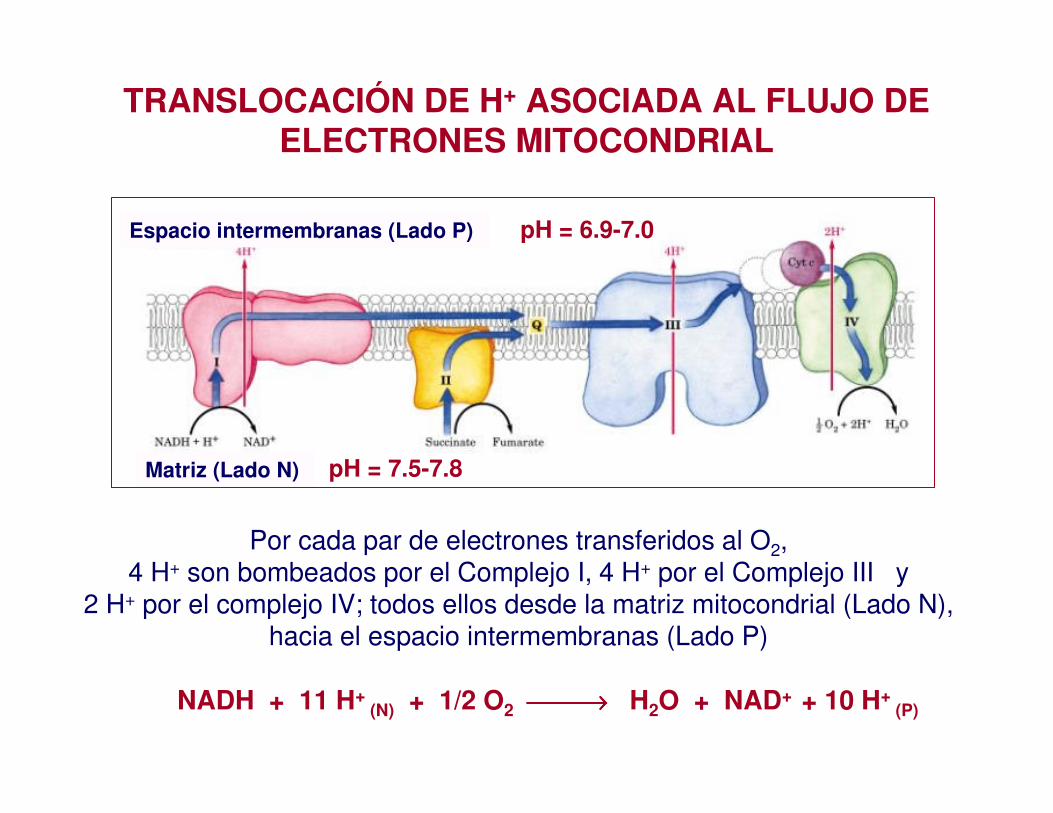
TRANSLOCACIÓN DE H+ ASOCIADA AL FLUJO DE ELECTRONES MITOCONDRIAL
pH = 6.9-7.0 Espacio intermembranas (Lado P)
pH = 7.5-7.8 Matriz (Lado N)
Por cada par de electrones transferidos al O2,4 H+ son bombeados por el Complejo I, 4 H+ por el Complejo III y
2 H+ por el complejo IV; todos ellos desde la matriz mitocondrial (Lado N), hacia el espacio intermembranas (Lado P)
NADH + 11 H+ (N) + 1/2 O2 →→→→ H2O + NAD+ + 10 H+
(P)
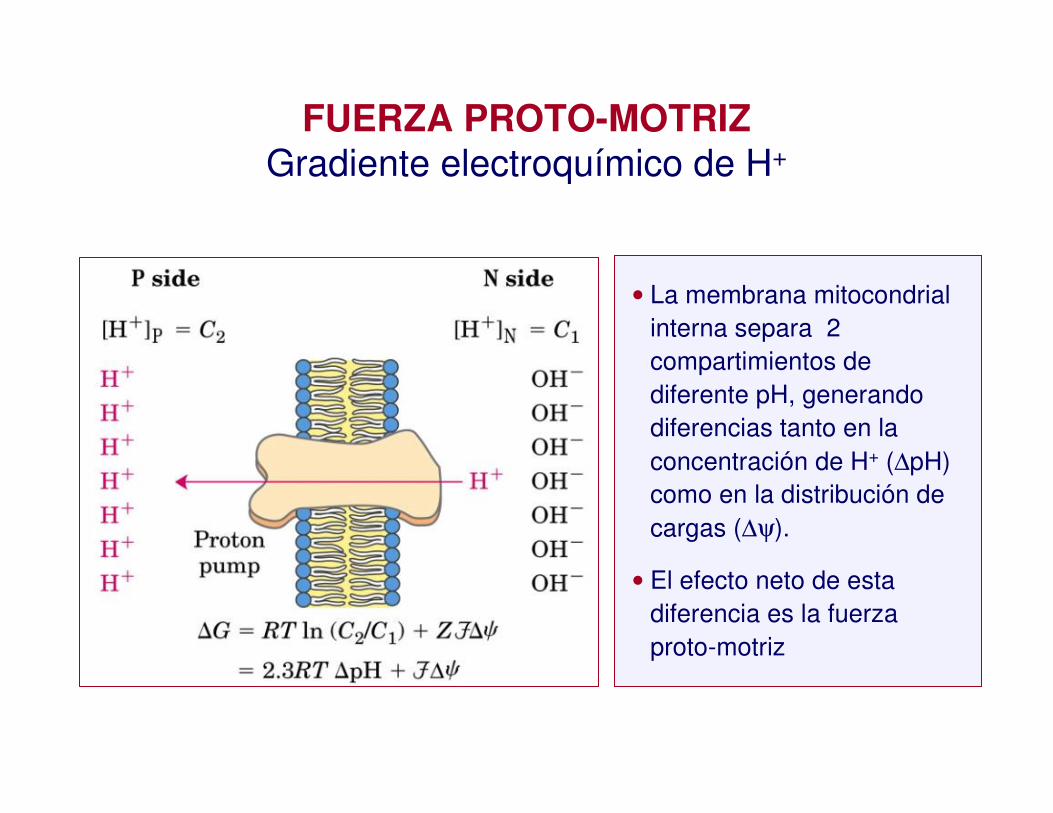
• La membrana mitocondrial interna separa 2 compartimientos de diferente pH, generando diferencias tanto en la concentración de H+ (∆pH) como en la distribución de cargas (∆ψ).
• El efecto neto de esta diferencia es la fuerza proto-motriz
FUERZA PROTO-MOTRIZGradiente electroquímico de H+

Potencial electroquímico para el H+
Para 10 H+ que translocan a través de la membrana
• Aproximadamente, 200 kJ/mol de los 220 kJ liberados por la oxidación de 1 mol de NADH son conservados en el gradiente electroquímico de H+ (fuerza proto-motriz)
• Cuando los H+ vuelvan espontáneamente a través de su gradiente electroquímico, hacia la matriz mitocondrial, la energía estará disponible para realizar trabajo

Potencial electroquímico para el H+
• La fuerza proto-motriz (∆∆∆∆p), expresa en mV (220-230 mV) el potencial electroquímico del H+ que es utilizado en la síntesis endergónica de ATP a partir de ADP y Pi

La fosforilación oxidativa, ocurre en la mitocondria, y se refiere a la síntesis
química de ATP (reacción endergónica)
impulsada por el proceso exergónico de transferencia de electrones desde el
NADH al O2 (cadena respiratoria).
FOSFORILACIÓN OXIDATIVA

TEORÍA QUIMIOSMÓTICA MITOCONDRIAL(P. Mitchell, 1961)
Peter Mitchell1920-1992
The Nobel Prize in Chemistry 1978
"for his contribution to the understanding
of biological energy transfer through the
formulation of the chemiosmotic theory"
El gradiente electroquímico de protones a través de la membrana interna mitocondrial (fuerza proto-motriz)
es crucial para los procesos de transducción de energía y síntesis de ATP
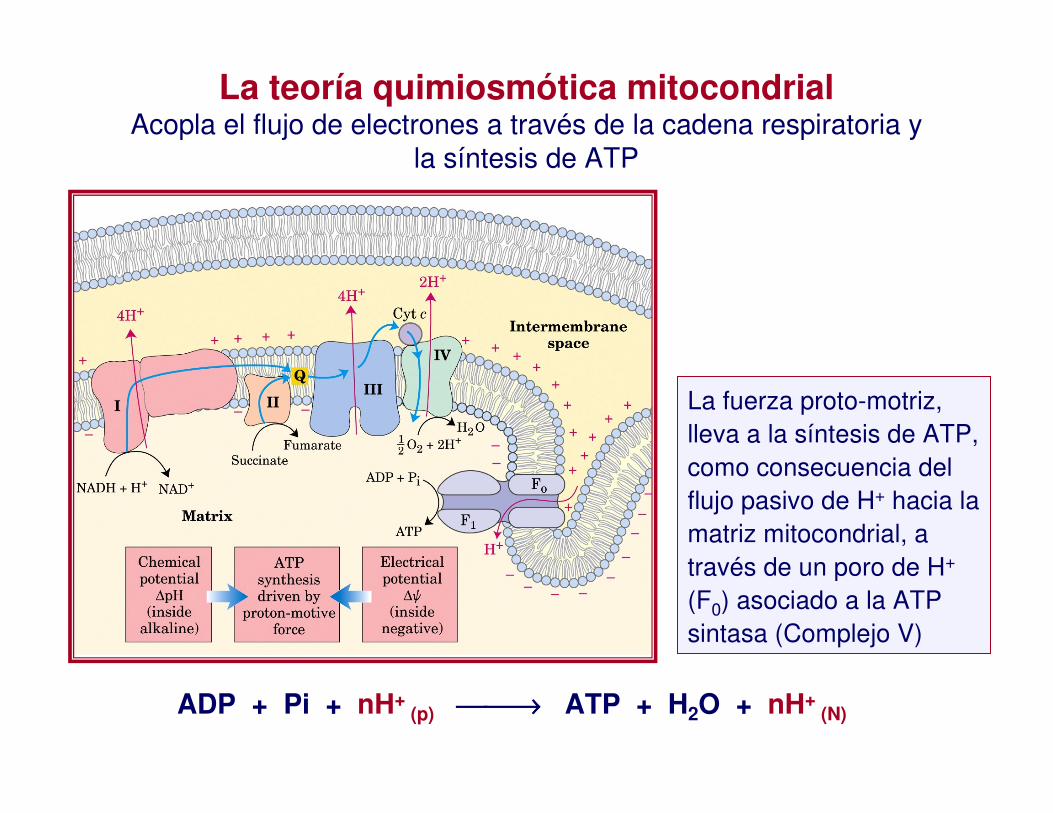
La teoría quimiosmótica mitocondrialAcopla el flujo de electrones a través de la cadena respiratoria y
la síntesis de ATP
La fuerza proto-motriz, lleva a la síntesis de ATP, como consecuencia del flujo pasivo de H+ hacia la matriz mitocondrial, a través de un poro de H+
(F0) asociado a la ATP sintasa (Complejo V)
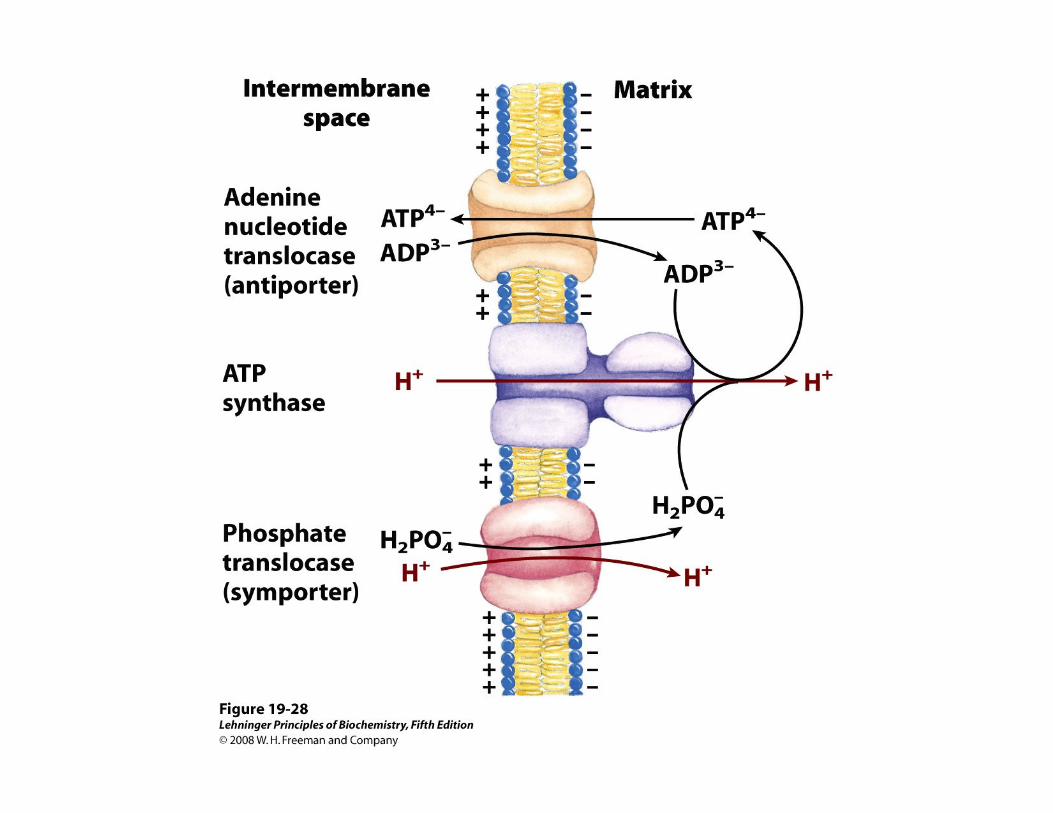
ADP + Pi + nH+ (p) →→→→ ATP + H2O + nH+
(N)

Postulados de la teoría quimiosmótica mitocondrial(P. Mitchell, 1961)
• La membrana interna es impermeable a los H+
• El transporte de e- a través de la cadena respiratoria está asociado al transporte de H+ desde la matriz hacia el espacio intermembranas
• Se conserva la energía de oxidación de los procesos metabólicos en forma de potencial electroquímico, ya que se genera un gradiente electroquímico de H+
• La cadena respiratoria está acoplada a la síntesis de ATP
• Las [H+] en las 2 fases acuosas (EIM y M) separadas por la membrana interna, constituyen la fuerza responsable (fuerza proto-motriz) de la formación de ATP: el flujo de H+ a favor de su gradiente electroquímico proporciona la energía libre para la síntesis de ATP a partir de ADP y de Pi, por acción de la F1-ATPasa de la membrana mitocondrial.
• Se sintetizan 3 ATP por cada par de e- pasados al O2 si el dador es NADH, y 2 ATP por cada par de e- pasados al O2 si el dador es succinato.

Síntesis de ATP por ATP sintasa
John Walker
The Nobel Prize in Chemistry 1997
"for their elucidation of the enzymatic
mechanism underlying the synthesis of
adenosine triphosphate (ATP)"
Paul Boyer
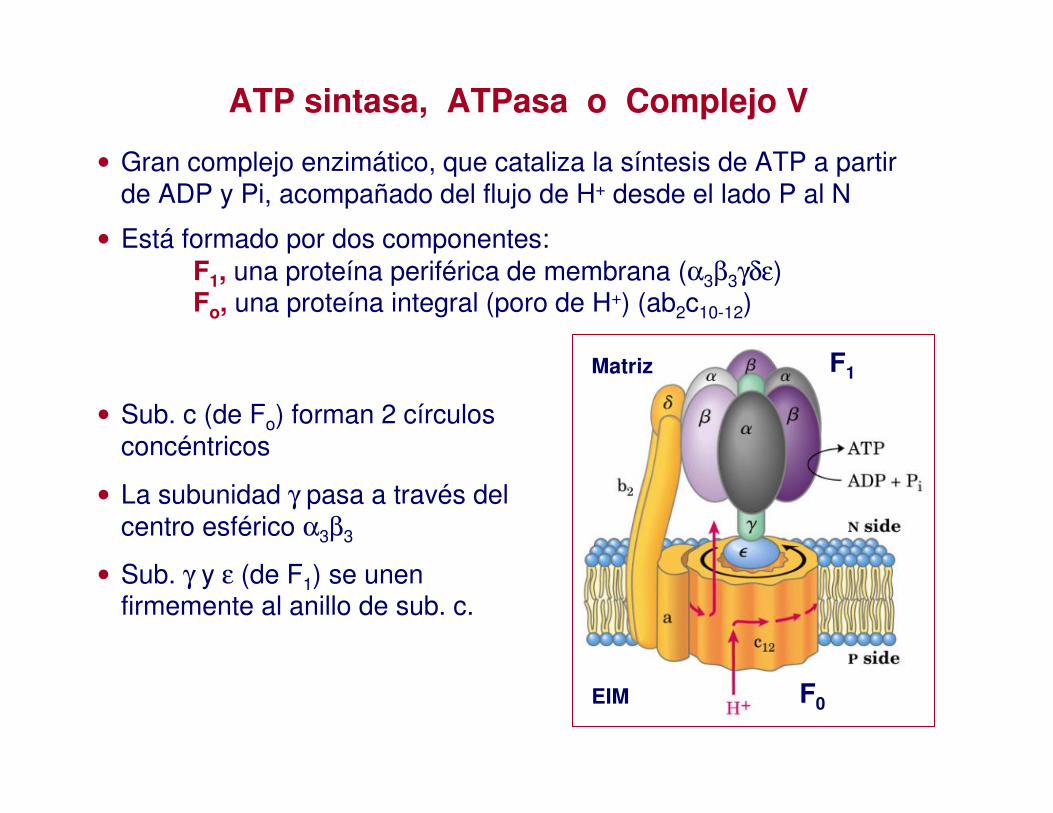
ATP sintasa, ATPasa o Complejo V
• Gran complejo enzimático, que cataliza la síntesis de ATP a partir de ADP y Pi, acompañado del flujo de H+ desde el lado P al N
• Está formado por dos componentes:F1, una proteína periférica de membrana (α3β3γδε)Fo, una proteína integral (poro de H+) (ab2c10-12)
• Sub. c (de Fo) forman 2 círculos concéntricos
• La subunidad γ pasa a través del centro esférico α3β3
• Sub. γ y ε (de F1) se unen firmemente al anillo de sub. c.
F1
F0
Matriz
EIM
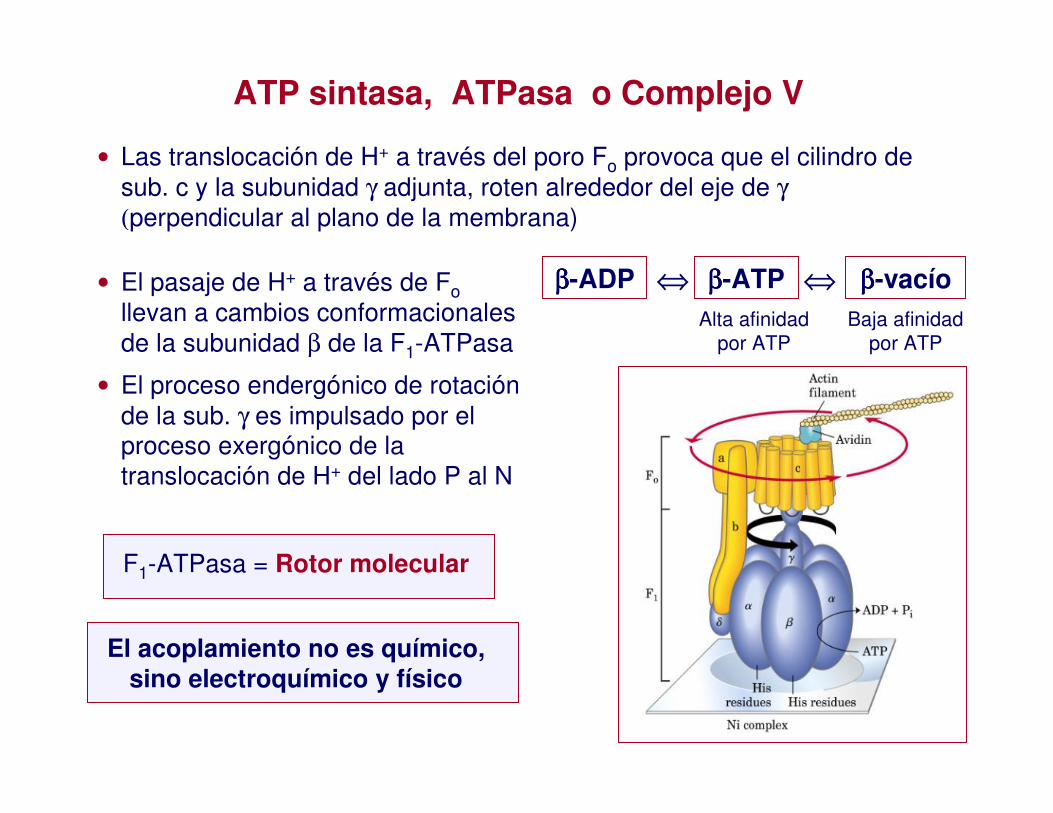
• Las translocación de H+ a través del poro Fo provoca que el cilindro de sub. c y la subunidad γ adjunta, roten alrededor del eje de γ
(perpendicular al plano de la membrana)
ATP sintasa, ATPasa o Complejo V
ββββ-ADP ββββ-ATP ββββ-vacío⇔ ⇔Alta afinidad
por ATPBaja afinidad
por ATP
• El pasaje de H+ a través de Follevan a cambios conformacionalesde la subunidad β de la F1-ATPasa
• El proceso endergónico de rotación de la sub. γ es impulsado por el proceso exergónico de la translocación de H+ del lado P al N
F1-ATPasa = Rotor molecular
El acoplamiento no es químico, sino electroquímico y físico
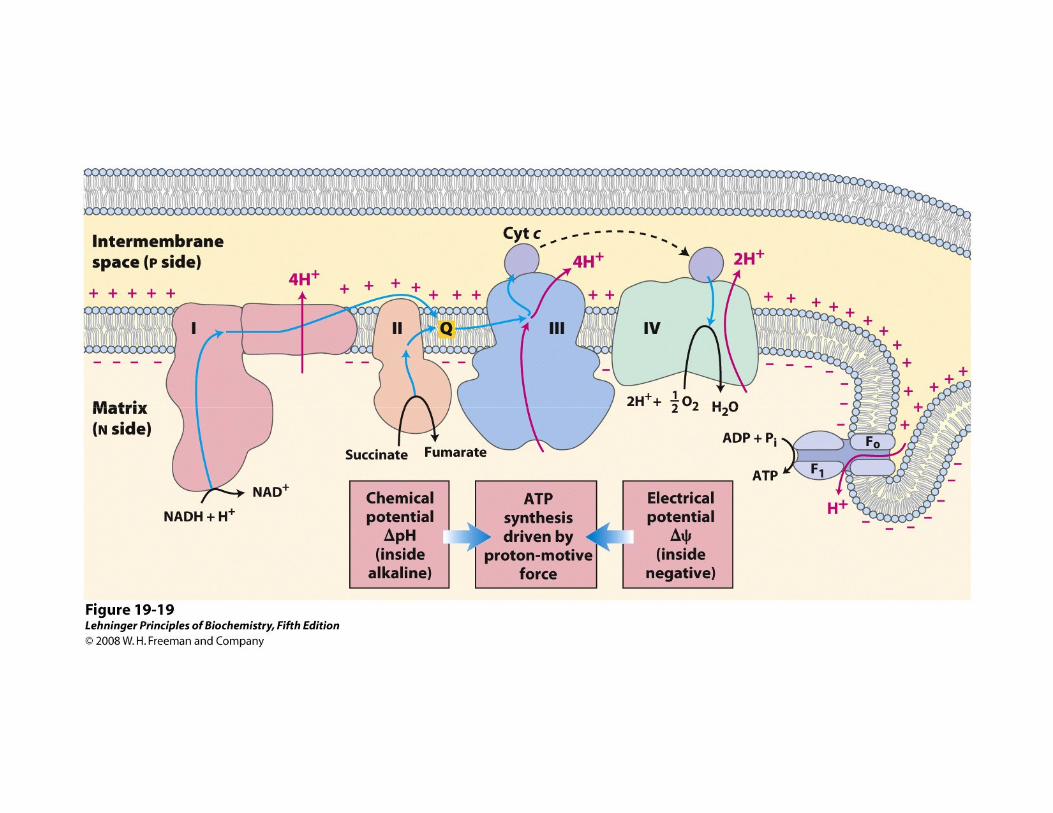

La FOSFORILACIÓN OXIDATIVA se refiere a la síntesis química de ATP impulsada por el proceso exergónico de transferencia de
electrones desde el NADH al O2
RESUMEN
• Ocurre en la mitocondria
• La fosforilación oxidativa comienza con la entrada de e- en la cadena respiratoria
• Los e- pasan a través de una serie de transportadores incluídos en la membrana interna mitocondrial. Los e- fluyen espontáneamente desde los transportadores de Eo’ más bajo hacia los de Eo’ más elevado
• Los transportadores electrónicos mitocondriales funcionan dentro de complejos proteicos ordenados en serie
• La transferencia mitocondrial de e- es un proceso exergónico, que libera energía suficiente para la síntesis de ATP
• El transporte de e- está asociado al transporte de H+ desde la matriz hacia el EIM (fuerza proto-motriz)
• El flujo de H+ a favor de su gradiente electroquímico proporciona la energía libre para la síntesis de ATP, por acción de la ATP sintasa (rotor molecular)
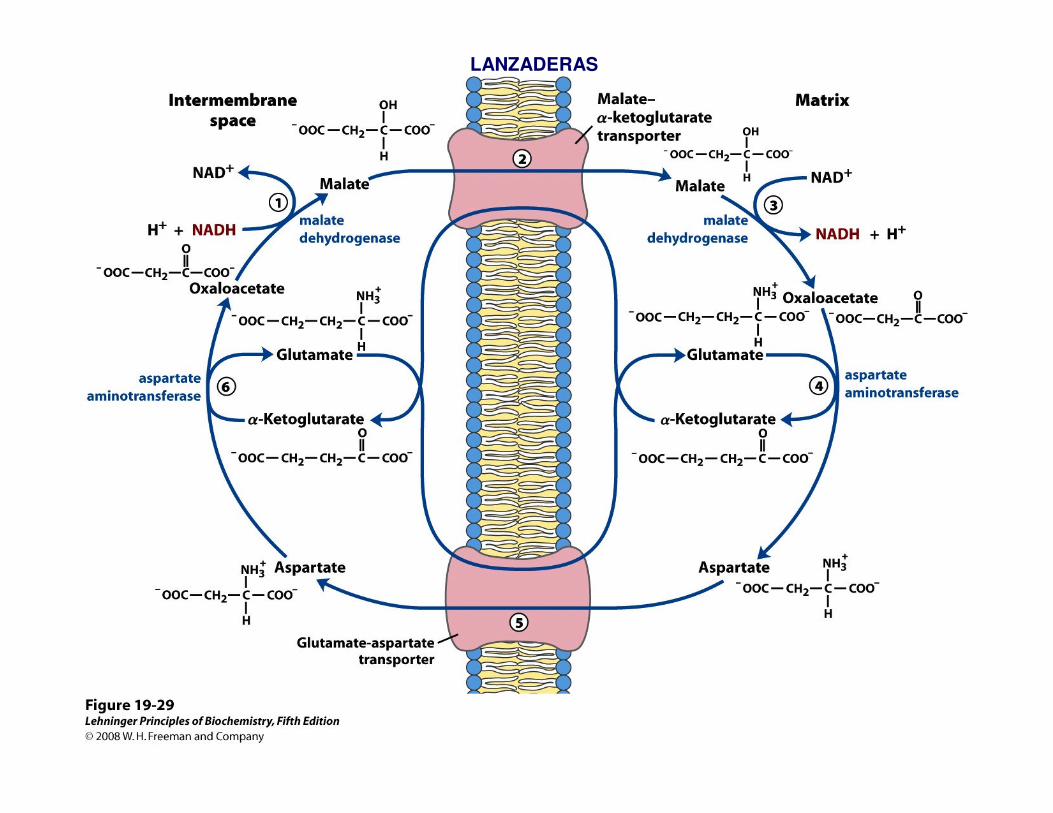
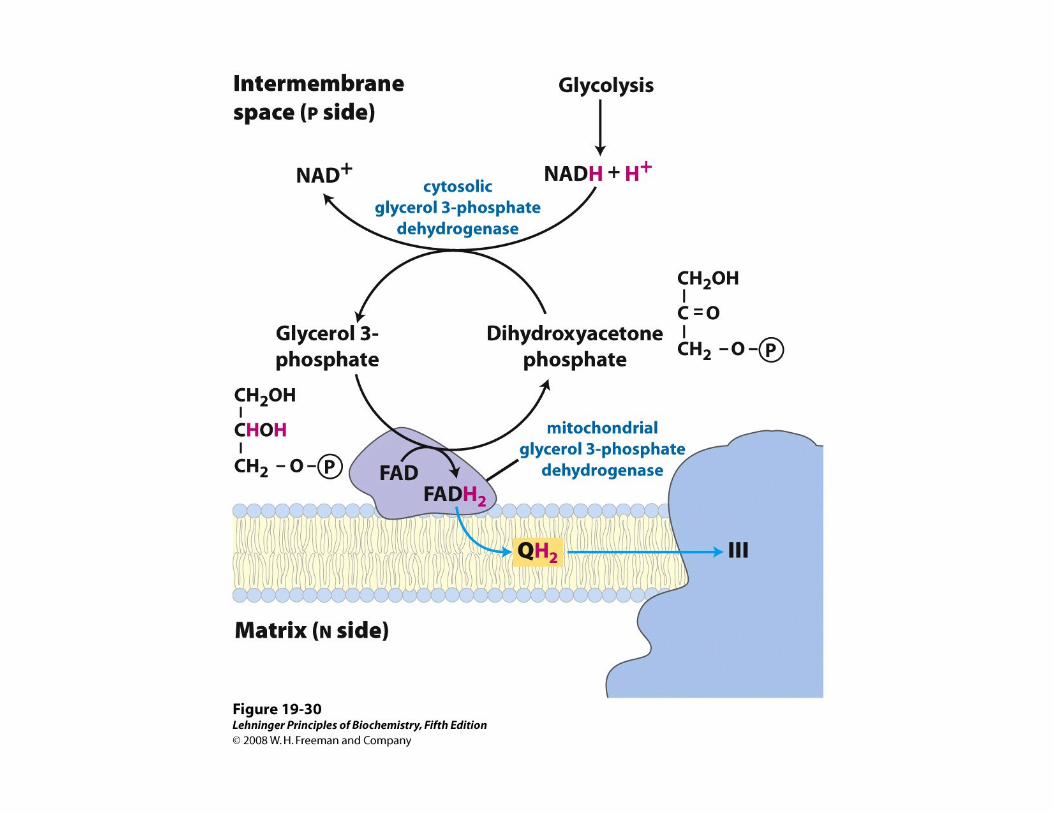
LANZADERAS