Embed Size (px)
Citation preview

DEVELOPMENTAL GENETICS 12:423430 (1991)
Catalase and Superoxide Dismutase Gene Expression and Distribution During Stem Development in Maize ALBERT0 ACEVEDO AND JOHN G. SCANDALIOS Department of Genetics, North Carolina State University, Raleigh
ABSTRACT The temporal and spatial pat- terns of expression of the Cut and Sod genes en- coding the multiple catalases and superoxide dis- mutases in maize have been examined throughout stem development. Three stages of stem develop- ment have been defined based upon catalase ac- tivity profiles and stem internode elongation. At stage 1, catalase activity is law and internodes re- main short; at stage 2, catalase activity dramati- cally increases and internodes rapidly elongate; and at stage 3, catalase activity decreases to levels intermediate to stage 1 and 2, and internode elon- gation ceases. Zymogram analysis and immu- noassays show that only the CAT-3 catalase isozyme is present in the stem, even though both Cut I and Cut3 mRNAs accumulate throughout stem development. Cat2 mRNA is not detectable in the developing stem. In full-grown stems catalase is localized primarily in the sclerenchyma beneath the epidermis and around the vascular bundles and may possibly play a role in lignification. Unlike catalase, all the superoxide dismutase (SOD) isozymes and transcripts are present in the devel- oping stem. Thus, these two maior antioxidant gene-enzyme systems show differential patterns of expression during stem development in maize. 0 1992 Wiley-Liss, Inc.
Key words: Zeu mays, superoxide dismutase, differential gene expression, stem lignification
INTRODUCTION The stem is commonly an aerial tissue of the plant
body that bears the leaves and reproductive structures. The stem contributes to define the particular architec- ture of the plant due to its insertion between roots and reproductive structures. In maize (Zea mays L.), the stem is a segmented shaft that gradually tapers toward its top and develops as a succession of nodes which alternate with internodes [Kiesselbach, 19491. The maize stem passes through three distinct stages in its development from germination to anthesis. At stage 1, stem length is 0 to 15 cm, the internodes remain short,
and a t the time of tassel formation the stem is no more than 10 cm tall, although the plant may be 35 cm in height [Poethig, 19821. At stage 2, stem length is 16 to 85 cm, and the above-ground internodes begin to elon- gate rapidly through new cell formation by an inter- calary meristem that is located at the base of each in- ternode except for the first [Kiesselbach, 19491. Subsequently, cell elongation, which may occur in sev- eral internodes at the same time, causes the rapid growth in height of the maize plant [Kiesselbach, 19491. Finally, a t stage 3, stem length is over 86 cm, active internode elongation diminishes, and nodes and internodes make up the final height of the stem. Dur- ing the development of the stem a variable number of below-ground internodes remain short and form the stem crown [Poethig, 19821. The crown internodes give rise to the adventitious crown-root system, whereas the above-ground nodes determine the points of leaf at- tachment. The fact that the stem temporarily stores photosynthesis products [Gallais et al., 19801, coupled with the distribution of the reproductive structures along it, assigns to the stem an underlying role during the translocation of assimilates to the growing ear.
In maize, catalase (E.C. 1.11.1.6; H,02:H,02 oxi- doreductase; CAT) and superoxide dismutase (E.C. 1.15.1.1. -O;:.Og oxidoreductase; SOD) have been ex- tensively studied during the youngest developmental stages of the sporophytic phase and at the tricellular mature pollen stage of the gametophytic phase [Baum and Scandalios, 1979,1982; Tsaftaris et al., 1983; Skad- sen and Scandalios, 1987; Wadsworth and Scandalios, 1989; Acevedo and Scandalios, 1990; Redinbaugh et al., 1990; White et al., 19901. Similarly, the vascular course, developmental morphology, and programming of the maize stem have been described in detail [Kies- selbach, 1949; Kumazawa, 1961; Poethig, 1982; Irish
Received for publication October 29, 1991; accepted December 17, 1991.
Address reprint requests to John G. Scandalios, Department of Ge- netics, North Carolina State University, Box 7614, Raleigh, NC 27695-7614.
0 1992 WILEY-LISS, INC.

424 ACEVEDO AND SCANDALIOS
and Nelson, 1988; McDaniel and Poethig, 1988; Sussex, 19891. However, few studies on maize stem develop- ment have been done from a genetic and biochemical viewpoint [Jorgenson, 1931; Gallais et al., 19801.
The three maize structural genes (Catl , Cat2, and Cat3) encode three biochemically distinct isozymic forms of catalase (CAT-1, CAT-2, and CAT-3) [Scan- dalios, 1979; Roupakias et al., 19801. These genes ex- hibit a high degree of temporal and spatial regulation and respond to imposed environmental stresses [Scan- dalios, 19871. The multiple maize nuclear genes (Sodl , Sod2, So&, sod4, Sod4A, and sod5) encode the corre- sponding differentially compartmentalized superoxide dismutase isozymes: SOD-1 (chloroplastic), SOD-3 (mi- tochondrial), and SOD-2, SOD-4, SOD4A, and SOD-5 (cytosolic) [Baum and Scandalios, 1979, 1982; Cannon and Scandalios, 19891. The possibilities that a second isozyme for SOD-3 exists in the W64A maize line [Ace- vedo and Scandalios, 19901 and that Sod5 and either sod4 or Sod4A may be allelic [Baum and Scandalios, 1982; Cannon and Scandalios, 19891 cannot be ruled out. Unlike catalase, superoxide dismutase isozymes do not appear to be tissue specific in their expression be- cause they have been detected in all maize tissues ex- amined to date [Baum and Scandalios, 1982; Acevedo and Scandalios, 1990; White et al., 19901.
These two major antioxidant gene-enzyme systems are believed to play an underlying role in the defense of the maize plant by protecting cells from toxic oxygen species [Halliwell, 19741. Consequently, they provide two promising and differentially regulated systems with which to study gene expression throughout stem development.
Herein, we characterize the expression of catalase and superoxide dismutase in different stages of the de- veloping stem and discuss the possible physiological role of catalase during stem lignification.
MATERIALS AND METHODS Materials
The maize (Zea mays L.) inbred line W64A, which is maintained by our laboratory, was the source of all the tissues analyzed in this report. Plants were grown in the field. Half parts of whole stems, nodes, and inter- nodes (dissected from plants harvested at different stages of stem development) were immediately pro- cessed for zymogram analysis. The other halves were frozen in liquid nitrogen and stored at - 70°C until pro- cessed for RNA extraction. Catalase isozyme markers were prepared from kernels and mature pollen that was collected and processed as described [Acevedo and Scandalios, 19901.
Kernels that were surface-sterilized for 10 min in 1% sodium hypochlorite and soaked in distilled water for 24 h were then placed on moistened germination paper in sterilized germination boxes. Germination and sub- sequent growth proceeded a t 25°C in a dark growth
chamber. Scutella and epicotyls, dissected from dark- grown seedlings at 4 and 8 days postimbibition, and pollen were processed for zymogram and immunoelec- trophoresis analyses to serve as catalase isozyme markers. Tissue for RNA isolation was frozen in liquid nitrogen and stored at -70°C.
Enzyme Assay Catalase was assayed spectrophotometrically as pre-
viously described [Beers and Sizer, 19521. An extrac- tion concentration of 1 g fresh weight tissue for 5 ml of 25 mM glycylglycine NaOH buffer, pH 7.4 (1 g.f.w./5 ml) was used. Total catalase activity is expressed as the change in absorbancelminlgram fresh weight (AM min1g.f.w.). Protein concentrations were determined as described [Lowry et al., 19511 using bovine serum al- bumin as standard. Each activity measurement repre- sents the average of two independent experiments with repetitions.
Zymogram Analysis For catalase, starch gel (12%) electrophoresis was
performed using a tris-citrate buffer system [Scandal- ios, 19681. For superoxide dismutase, electrophoresis was conducted using a lithium hydroxide-boric acid: tris-citrate buffer system [Baum and Scandalios, 19791. Histochemical staining of the zymograms was as pre- viously described [Scandalios, 1968; Baum and Scan- dalios, 19791.
Immunoelectrophoresis Analysis Rocket immunoelectrophoresis [Laurell, 19661 using
catalase monospecific antibodies was performed as modified by Tsaftaris and Scandalios [ 19811.
RNA Isolation and Analysis Total RNA was isolated as previously described
[Chirgwin et al., 19791. Total cellular RNA, which will henceforth be referred to as RNA, was separated on denaturing agarose gels (1.2%), transferred to nitrocel- lulose paper, and prehybridized at 42°C for 24 h. Hy- bridization with gene-specific sequences for Cat2 [Bethards et al., 19871, Cat1 and Cat3 [Redinbaugh et al., 19881, Sod2 [Cannon et al., 19871, sod4 and Sod4A [Cannon and Scandalios, 19891, and a full-length se- quence for Sod3 [White and Scandalios, 19881 pro- ceeded a t 42°C for 24 h with lo6 cpm/ml of probes radiolabeled with (a-32P)dCTP (800 Cilmmol). After hybridization, filters were washed, and autoradio- grams were produced as previously described [Acevedo and Scandalios, 19901.
Sample Preparation for Optical Microscopy Upper and basal nodes and internodes dissected from
full-grown stems were sectioned with a microtome. To determine the presence and distribution of lignin, cross-sections of nodes and internodes were placed on microscope slides and embedded with phloroglucinol-
1 4
fn ; 10 c 0 E .- 4- C
6 7
3
MAIZE STEM CATALASES AND SUPEROXIDE DISMUTASES 425
S t a g e s of Stem Development 1 2 3
0
0 4 0 8 0 1 2 0
Days After Plant ing
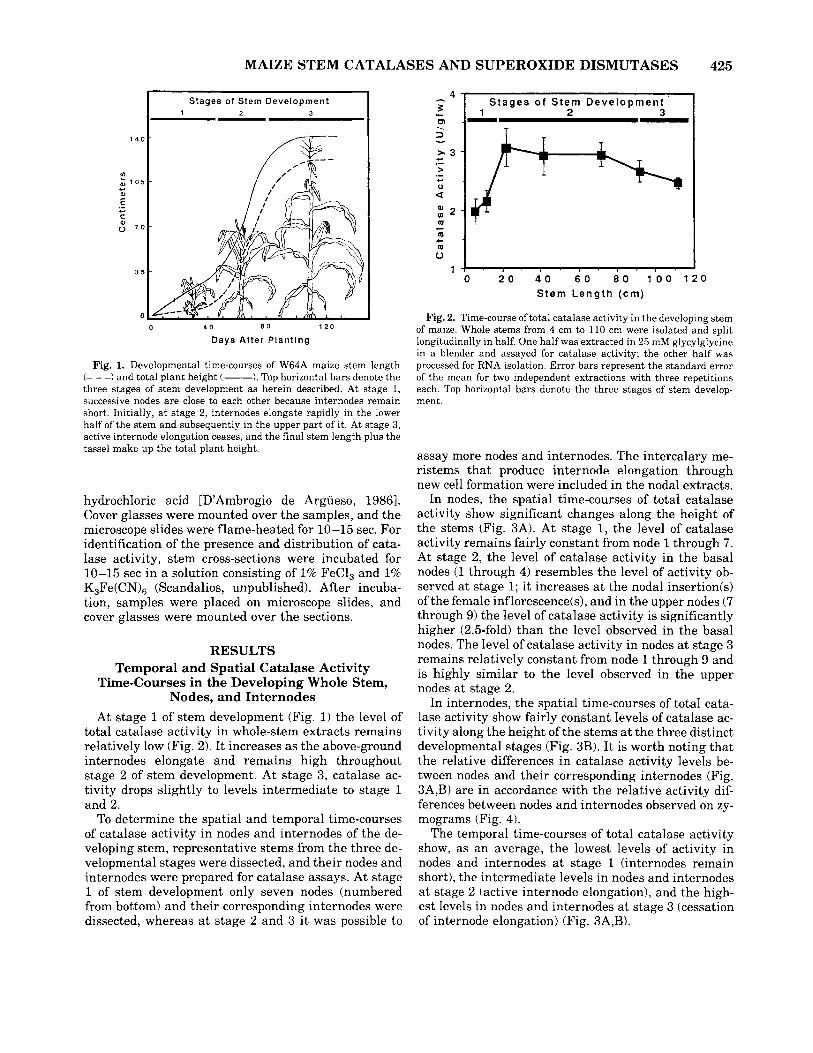
Fig. 1. Developmental time-courses of W64A maize stem length (- - -1 and total plant height (-). Top horizontal bars denote the three stages of stem development as herein described. At stage 1, successive nodes are close to each other because internodes remain short. Initially, a t stage 2, internodes elongate rapidly in the lower half of the stem and subsequently in the upper part of it. At stage 3, active internode elongation ceases, and the final stem length plus the tassel make up the total plant height.
hydrochloric acid [D’Ambrogio de Argiieso, 19861. Cover glasses were mounted over the samples, and the microscope slides were flame-heated for 10-15 sec. For identification of the presence and distribution of cata- lase activity, stem cross-sections were incubated for 10-15 sec in a solution consisting of 1% FeC1, and 1% K,Fe(CN), (Scandalios, unpublished). After incuba- tion, samples were placed on microscope slides, and cover glasses were mounted over the sections.
RESULTS Temporal and Spatial Catalase Activity
Time-Courses in the Developing Whole Stem, Nodes, and Internodes
At stage 1 of stem development (Fig. 1) the level of total catalase activity in whole-stem extracts remains relatively low (Fig. 2). It increases as the above-ground internodes elongate and remains high throughout stage 2 of stem development. At stage 3, catalase ac- tivity drops slightly to levels intermediate to stage 1 and 2.
To determine the spatial and temporal time-courses of catalase activity in nodes and internodes of the de- veloping stem, representative stems from the three de- velopmental stages were dissected, and their nodes and internodes were prepared for catalase assays. At stage 1 of stem development only seven nodes (numbered from bottom) and their corresponding internodes were dissected, whereas a t stage 2 and 3 it was possible to
4 I S t a g e s o f S t e m D e v e l o p m e n t
3 s CD .- - 1 2 - .
0 2 0 4 0 6 0 8 0 100 120 Stem L e n g t h ( c m )
Fig. 2. Time-course of total catalase activity in the developing stem of maize. Whole stems from 4 cm to 110 cm were isolated and split longitudinally in half. One half was extracted in 25 mM glycylglycine in a blender and assayed for catalase activity; the other half was processed for RNA isolation. Error bars represent the standard error of the mean for two independent extractions with three repetitions each. Top horizontal bars denote the three stages of stem develop- ment.
assay more nodes and internodes. The intercalary me- ristems that produce internode elongation through new cell formation were included in the nodal extracts.
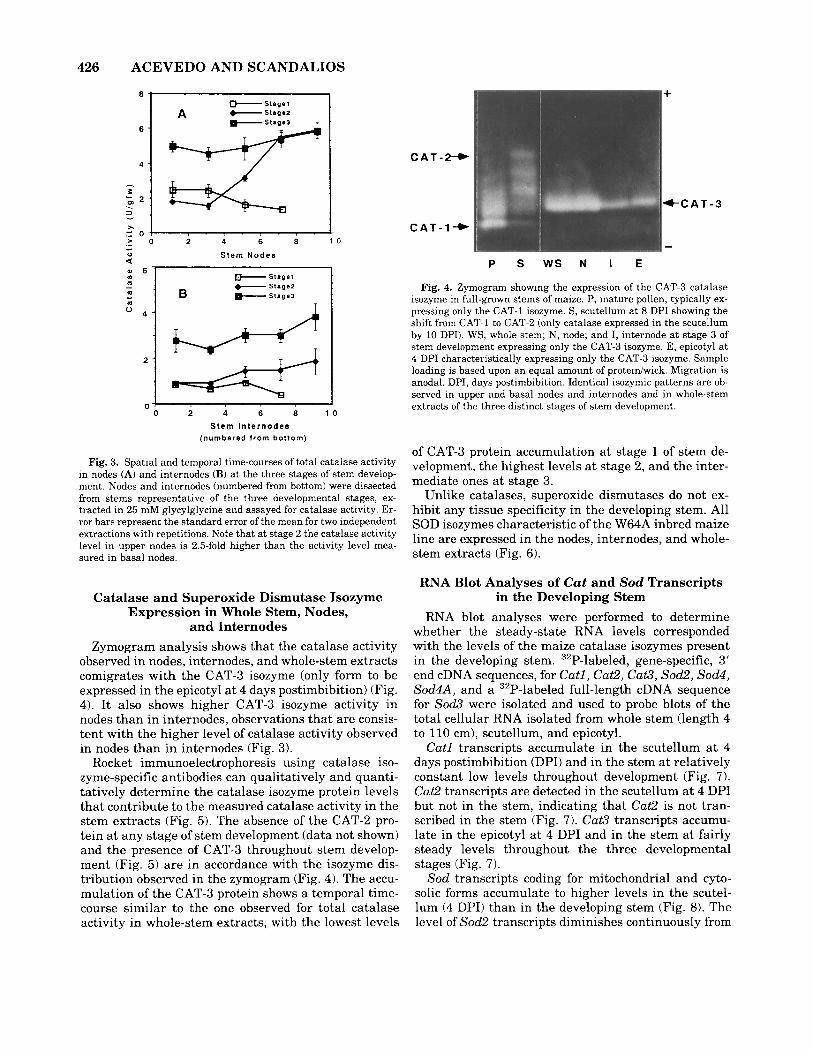
In nodes, the spatial time-courses of total catalase activity show significant changes along the height of the stems (Fig. 3A). At stage 1, the level of catalase activity remains fairly constant from node 1 through 7. At stage 2, the level of catalase activity in the basal nodes (1 through 4) resembles the level of activity ob- served a t stage 1; i t increases at the nodal insertion(s) of the female inflorescence(s), and in the upper nodes (7 through 9) the level of catalase activity is significantly higher (2.5-fold) than the level observed in the basal nodes. The level of catalase activity in nodes a t stage 3 remains relatively constant from node 1 through 9 and is highly similar to the level observed in the upper nodes at stage 2.
In internodes, the spatial time-courses of total cata- lase activity show fairly constant levels of catalase ac- tivity along the height of the stems a t the three distinct developmental stages (Fig. 3B). It is worth noting that the relative differences in catalase activity levels be- tween nodes and their corresponding internodes (Fig. 3A,B) are in accordance with the relative activity dif- ferences between nodes and internodes observed on zy- mograms (Fig. 4).
The temporal time-courses of total catalase activity show, as an average, the lowest levels of activity in nodes and internodes at stage 1 (internodes remain short), the intermediate levels in nodes and internodes a t stage 2 (active internode elongation), and the high- est levels in nodes and internodes a t stage 3 (cessation of internode elongation) (Fig. 3A,B).
426 ACEVEDO AND SCANDALIOS
121 . 2
U ' 0 2 4 6 8 1 0
Stem Internodes (numbered from bottom)
Fig. 3. Spatial and temporal time-courses of total catalase activity in nodes (A) and internodes (B) at the three stages of stem develop- ment. Nodes and internodes (numbered from bottom) were dissected from stems representative of the three developmental stages, ex- tracted in 25 mM glycylglycine and assayed for catalase activity. Er- ror bars represent the standard error of the mean for two independent extractions with repetitions. Note that at stage 2 the catalase activity level in upper nodes is 2.5-fold higher than the activity level mea- sured in basal nodes.
+
CAT-2-
Catalase and Superoxide Dismutase Isozyme Expression in Whole Stem, Nodes,
and Internodes Zymogram analysis shows that the catalase activity
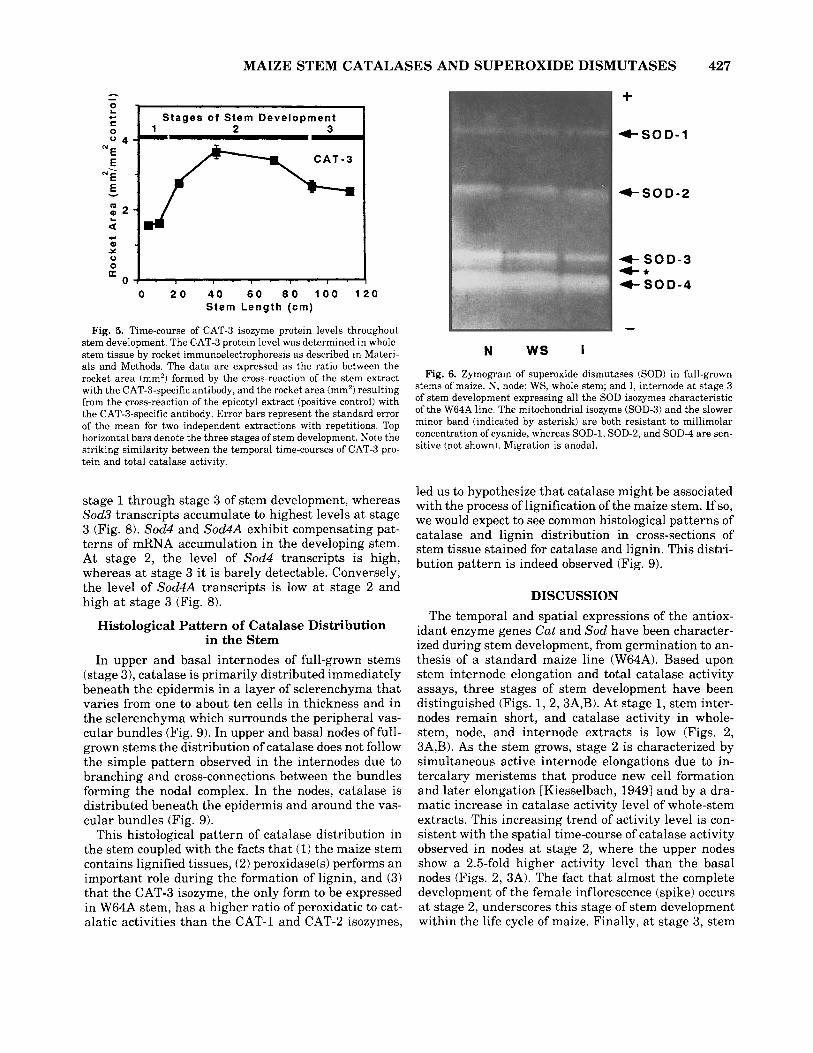
observed in nodes, internodes, and whole-stem extracts comigrates with the CAT-3 isozyme (only form to be expressed in the epicotyl a t 4 days postimbibition) (Fig. 4). It also shows higher CAT-3 isozyme activity in nodes than in internodes, observations that are consis- tent with the higher level of catalase activity observed in nodes than in internodes (Fig. 3).
Rocket immunoelectrophoresis using catalase iso- zyme-specific antibodies can qualitatively and quanti- tatively determine the catalase isozyme protein levels that contribute to the measured catalase activity in the stem extracts (Fig. 5). The absence of the CAT-2 pro- tein a t any stage of stem development (data not shown) and the presence of CAT-3 throughout stem develop- ment (Fig. 5) are in accordance with the isozyme dis- tribution observed in the zymogram (Fig. 4). The accu- mulation of the CAT-3 protein shows a temporal time- course similar to the one observed for total catalase activity in whole-stem extracts, with the lowest levels
+CAT-3
C A T - l - ) -
P S W S N I E
Fig. 4. Zymogram showing the expression of the CAT-3 catalase isozyme in full-grown stems of maize. P, mature pollen, typically ex- pressing only the CAT-1 isozyme. S, scutellum at 8 DPI showing the shift from CAT-1 to CAT-2 (only catalase expressed in the scutellum by 10 DPI). WS, whole stem; N, node; and I, internode at stage 3 of stem development expressing only the CAT-3 isozyme. E, epicotyl a t 4 DPI characteristically expressing only the CAT-3 isozyme. Sample loading is based upon an equal amount of proteiniwick. Migration is anodal. DPI, days postimbibition. Identical isozymic patterns are ob- served in upper and basal nodes and internodes and in whole-stem extracts of the three distinct stages of stem development.
of CAT-3 protein accumulation a t stage 1 of stem de- velopment, the highest levels at stage 2, and the inter- mediate ones a t stage 3.
Unlike catalases, superoxide dismutases do not ex- hibit any tissue specificity in the developing stem. All SOD isozymes characteristic of the W64A inbred maize line are expressed in the nodes, internodes, and whole- stem extracts (Fig. 6).
RNA Blot Analyses of Cut and Sod Transcripts in the Developing Stem
RNA blot analyses were performed to determine whether the steady-state RNA levels corresponded with the levels of the maize catalase isozymes present in the developing stem. 32P-labeled, gene-specific, 3' end cDNA sequences, for Catl, Cat2, Cat3, Sod2, sod4, Sod4A, and a 32P-labeled full-length cDNA sequence for Sod3 were isolated and used to probe blots of the total cellular RNA isolated from whole stem (length 4 to 110 cm), scutellum, and epicotyl.
Catl transcripts accumulate in the scutellum a t 4 days postimbibition (DPI) and in the stem a t relatively constant low levels throughout development (Fig. 7). Cat2 transcripts are detected in the scutellum a t 4 DPI but not in the stem, indicating that Cat2 is not tran- scribed in the stem (Fig. 7). Cat3 transcripts accumu- late in the epicotyl at 4 DPI and in the stem at fairly steady levels throughout the three developmental stages (Fig. 7).
Sod transcripts coding for mitochondria1 and cyto- solic forms accumulate to higher levels in the scutel- lum (4 DPI) than in the developing stem (Fig. 8). The level of Sod2 transcripts diminishes continuously from
MAIZE STEM CATALASES AND SUPEROXIDE DISMUTASES 427
$ I t a g e s of Stem Development 0 2 V A
I : 1 . , . , . , . , , , , I 0
a 0 2 0 4 0 6 0 8 0 1 0 0 1 2 0
Stem Length (em)
Fig. 5. Time-course of CAT-3 isozyme protein levels throughout stem development. The CAT-3 protein level was determined in whole- stem tissue by rocket immunoelectrophoresis as described in Materi- als and Methods. The data are expressed as the ratio between the rocket area (mm2) formed by the cross-reaction of the stem extract with the CAT-3-specific antibody, and the rocket area (mrn') resulting from the cross-reaction of the epicotyl extract (positive control) with the CAT-3-specific antibody. Error bars represent the standard error of the mean for two independent extractions with repetitions. Top horizontal bars denote the three stages of stem development. Note the striking similarity between the temporal time-courses of CAT-3 pro- tein and total catalase activity.
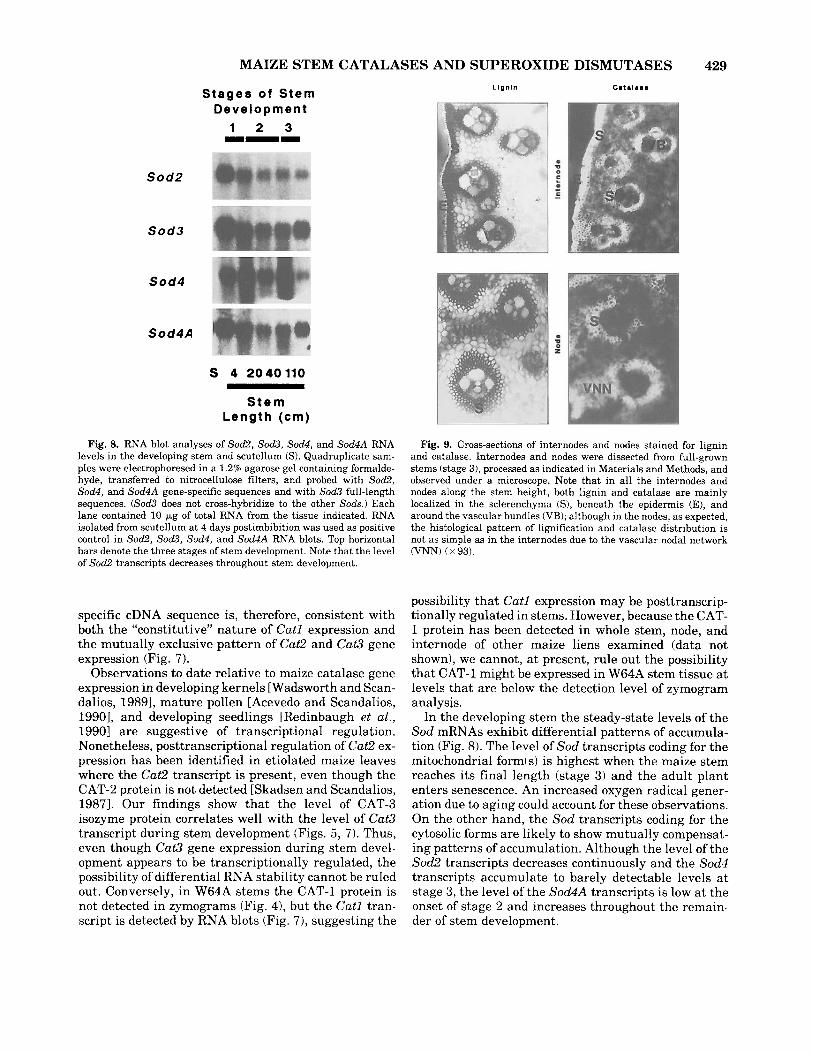
stage 1 through stage 3 of stem development, whereas Sod3 transcripts accumulate to highest levels a t stage 3 (Fig. 8). sod4 and Sod4A exhibit compensating pat- terns of mRNA accumulation in the developing stem. At stage 2, the level of sod4 transcripts is high, whereas at stage 3 i t is barely detectable. Conversely, the level of Sod4A transcripts is low a t stage 2 and high a t stage 3 (Fig. 8).
Histological Pattern of Catalase Distribution in the Stem
In upper and basal internodes of full-grown stems (stage 3), catalase is primarily distributed immediately beneath the epidermis in a layer of sclerenchyma that varies from one to about ten cells in thickness and in the sclerenchyma which surrounds the peripheral vas- cular bundles (Fig. 9). In upper and basal nodes of full- grown stems the distribution of catalase does not follow the simple pattern observed in the internodes due to branching and cross-connections between the bundles forming the nodal complex. In the nodes, catalase is distributed beneath the epidermis and around the vas- cular bundles (Fig. 9).
This histological pattern of catalase distribution in the stem coupled with the facts that (1) the maize stem contains lignified tissues, (2) peroxidase(s) performs an important role during the formation of lignin, and (3) that the CAT-3 isozyme, the only form to be expressed in W64A stem, has a higher ratio of peroxidatic to cat- alatic activities than the CAT-1 and CAT-2 isozymes,
+ +SOD-1
4 S O D - 2
4 S O D - 3 4-* 4- SOD-4
N ws I Fig. 6. Zymogram of superoxide dismutases (SOD) in full-grown
stems of maize. N, node; WS, whole stem; and I, internode a t stage 3 of stem development expressing all the SOD isozymes characteristic of the W64A line. The mitochondria1 isozyme (SOD-3) and the slower minor band (indicated by asterisk) are both resistant to millimolar concentration of cyanide, whereas SOD-1, SOD-2, and SOD-4 are sen- sitive (not shown). Migration is anodal.
led us to hypothesize that catalase might be associated with the process of lignification of the maize stem. If so, we would expect to see common histological patterns of catalase and lignin distribution in cross-sections of stem tissue stained for catalase and lignin. This distri- bution pattern is indeed observed (Fig. 9).
DISCUSSION The temporal and spatial expressions of the antiox-
idant enzyme genes Cat and Sod have been character- ized during stem development, from germination to an- thesis of a standard maize line (W64A). Based upon stem internode elongation and total catalase activity assays, three stages of stem development have been distinguished (Figs. 1, 2,3A,B). At stage 1, stem inter- nodes remain short, and catalase activity in whole- stem, node, and internode extracts is low (Figs. 2, 3A,B). As the stem grows, stage 2 is characterized by simultaneous active internode elongations due to in- tercalary meristems that produce new cell formation and later elongation [Kiesselbach, 19491 and by a dra- matic increase in catalase activity level of whole-stem extracts. This increasing trend of activity level is con- sistent with the spatial time-course of catalase activity observed in nodes at stage 2, where the upper nodes show a 2.5-fold higher activity level than the basal nodes (Figs. 2, 3A). The fact that almost the complete development of the female inflorescence (spike) occurs a t stage 2, underscores this stage of stem development within the life cycle of maize. Finally, a t stage 3, stem

428 ACEVEDO AND SCANDALIOS
S t a g e s o f S t e m D e v e l o p m e n t 1 2 3 I--
C a t l
S
C a t 2
S
C a t 3
E 4 20 40110
S t e m L e n g t h ( c m )
Fig. 7. RNA blot analyses of Catl, Cat2, and Cut3 RNA levels in the developing stem, scutellum (S), and epicotyl (El. Triplicate stem RNA samples were electrophoresed in a 1.29 agarose gel containing formaldehyde, transferred to nitrocellulose filters, and probed with Catl, Cat2, and Cat3 gene-specific sequences as indicated. Each lane contained 10 Fg of total RNA from the tissue indicated. RNA isolated from scutellum and epicotyl a t 4 days postimbibition was used as positive controls in Catl, Cut 2, and Cat3 RNA blots. Top horizontal bars denote the three stages of stem development. Note that although Cat1 and Cat3 transcripts accumulate at the three stages of stem development, no Cat2 transcripts are detected throughout stem de- velopment
nodes and internodes make up the final plant height, and catalase activity in whole stems gently decreases to levels intermediate to stage 1 and 2, whereas the level of catalase activity in the spatial time-courses of node and internode extracts remains relatively con- stant along the stem length (Figs. 2, 3A,B).
The histological pattern of catalase distribution and the differential vascularization in maize stem nodes and internodes may explain the differential levels of catalase activity observed throughout stem develop- ment. We have demonstrated in cross-sections of nodes and internodes of full-grown stems that catalase is lo- calized primarily in the sclerenchyma beneath the epi- dermis and around the vascular bundles (Fig. 9). In addition, the concentration of vascular bundles in nodes is higher than in internodes due to branching and cross-connections between bundles forming the nodal network [Kumazawa, 19611. Therefore, as we have demonstrated in zymogram and catalase activity assays, it is reasonable to observe in nodes higher lev-
els of activity than in internodes a t each of the three stages of stem development examined (Figs. 3A,B, 4).
Zymogram analysis of whole-stem, node, and inter- node extracts shows that CAT-3 is the only detectable catalase isozyme present throughout W64A stem de- velopment (Fig. 4). Furthermore, the CAT-3 protein is likely to be predominant in stem tissue. This is sup- ported by the immunological data that further verify the absence of CAT-2 and the presence of CAT-3 isozyme proteins in developing stems and by the strik- ing similarity between the temporal time-course of to- tal catalase activity and CAT-3 protein (Figs. 2, 5). Unlike catalase, all SOD isozymes are present in the stem, and their corresponding activities appear to be similar in whole stems, nodes, and internodes (Fig. 6). These observations indicate that the maize SODs do not exhibit any obvious tissue specificity and are in accordance with our previous reports [Baum and Scan- dalios, 1982; Acevedo and Scandalios, 19901. To date, nulls for CAT-2 [Tsaftaris and Scandalios, 19811 and CAT-3 [Wadsworth and Scandalios, 19901 have been found and characterized. However, like the CAT-1 isozyme, the apparent absence of any SOD null pheno- type among the many lines we have examined, in ad- dition to its apparent “constitutive” expression, sug- gests that the expression of the SODs may be crucial for the viability of the maize plant.
It is known that the developing stem of maize under- goes lignification and that during this process the CoA esters of 4-coumaric, ferulic, and sinapic acids are re- duced to their corresponding alcohols, which are then polymerized to form lignin through the action of a per- oxidase [Hahlbrock and Grisebach, 19791. It is also known that the CAT-3 isozyme, the only form detected in the stem, has enhanced peroxidatic activity [Scan- dalios, unpublished observations; Havir and McHale, 19891. Taken together, these data and the overlapping histological patterns of lignification and catalase dis- tribution (Fig. 9) suggest that catalase might play a role during the lignification process of the maize stem.
The Catl transcript has been found in all maize tis- sues screened so far [Wadsworth and Scandalios, 1989; Acevedo and Scandalios, 1990; Redinbaugh et al., 19901. On the other hand, it has been demonstrated that the Cat2 and Cat3 transcripts accumulate in a “mutually exclusive” manner in maize developing ker- nels [Wadsworth and Scandalios, 19891 and in the tis- sues of the developing maize seedling [Redinbaugh et al., 19901. In this context, although both Cat2 and Cat3 are expressed in green maize leaf, there is evidence that the products of the Cat2 and Cat3 genes are dif- ferentially distributed between the dimorphic photo- synthetic cell types in the leaf (i.e., CAT-2 is found primarily in the bundle-sheath cells, whereas CAT-3 is found primarily in the mesophyll cells [Tsaftaris et al., 19831). The fact that stem RNA of the three develop- mental stages hybridizes with the Catl and Cat3 gene- specific cDNA sequences but not with the Cat2 gene-
MAIZE STEM CATALASES AND SUPEROXIDE DISMUTASES 429
Sod2
Stages of Stem Development
1 2 3 ---
S 4 2040110
S t e m Length (cm)
Fig. 8. RNA blot analyses of So&, So&, sod4, and Sod4A RNA levels in the developing stem and scutellum (S). Quadruplicate sam- ples were electrophoresed in a 1.2% agarose gel containing formalde- hyde, transferred to nitrocellulose filters, and probed with So&, sod4, and Sod4A gene-specific sequences and with Sod3 full-length sequences. (Sod3 does not cross-hybridize to the other Sods.) Each lane contained 10 pg of total RNA from the tissue indicated. RNA isolated from scutellum a t 4 days postimbibition was used as positive control in So&, So&, sod4, and Sod4A RNA blots. Top horizontal bars denote the three stages of stem development. Note that the level of Sod2 transcripts decreases throughout stem development.
specific cDNA sequence is, therefore, consistent with both the “constitutive” nature of Catl expression and the mutually exclusive pattern of Cat2 and Cat3 gene expression (Fig. 7).
Observations to date relative to maize catalase gene expression in developing kernels [ Wadsworth and Scan- dalios, 19891, mature pollen [Acevedo and Scandalios, 19901, and developing seedlings [Redinbaugh et al., 19901 are suggestive of transcriptional regulation. Nonetheless, posttranscriptional regulation of Cat2 ex- pression has been identified in etiolated maize leaves where the Cat2 transcript is present, even though the CAT-2 protein is not detected [Skadsen and Scandalios, 19871. Our findings show that the level of CAT-3 isozyme protein correlates well with the level of Cat3 transcript during stem development (Figs. 5, 7). Thus, even though Cat3 gene expression during stem devel- opment appears to be transcriptionally regulated, the possibility of differential RNA stability cannot be ruled out. Conversely, in W64A stems the CAT-1 protein is not detected in zymograms (Fig. 4), but the Catl tran- script is detected by RNA blots (Fig. 7), suggesting the
Llgnln cataia..
V
z
Fig. 9. Cross-sections of internodes and nodes stained for lignin and catalase. Internodes and nodes were dissected from full-grown stems (stage 3) , processed as indicated in Materials and Methods, and observed under a microscope. Note that in all the internodes and nodes along the stem height, both lignin and catalase are mainly localized in the sclerenchyma (S), beneath the epidermis (E), and around the vascular bundles (VB); although in the nodes, as expected, the histological pattern of lignification and catalase distribution is not as simple as in the internodes due to the vascular nodal network (VNN) ( x 93).
possibility that Catl expression may be posttranscrip- tionally regulated in stems. However, because the CAT- 1 protein has been detected in whole stem, node, and internode of other maize liens examined (data not shown), we cannot, at present, rule out the possibility that CAT-1 might be expressed in W64A stem tissue at levels that are below the detection level of zymogram analysis.
In the developing stem the steady-state levels of the Sod mRNAs exhibit differential patterns of accumula- tion (Fig. 8). The level of Sod transcripts coding for the mitochondria1 form(s) is highest when the maize stem reaches its final length (stage 3) and the adult plant enters senescence. An increased oxygen radical gener- ation due to aging could account for these observations. On the other hand, the Sod transcripts coding for the cytosolic forms are likely to show mutually compensat- ing patterns of accumulation. Although the level of the Sod2 transcripts decreases continuously and the sod4 transcripts accumulate to barely detectable levels a t stage 3, the level of the Sod4A transcripts is low at the onset of stage 2 and increases throughout the remain- der of stem development.

430 ACEVEDO AND SCANDALIOS
Ongoing research on the regulatory region of the maize catalase and superoxide dismutase genes will greatly contribute to a better understanding of gene expression and regulation during the development of the maize plant.
ACKNOWLEDGMENTS We gratefully acknowledge Olga Casal of Centro de
Investigaciones de Celulosa y Papel, INTI, Migueletes, Argentina, for her assistance with optical microscopy. The critical reading and constructive comments on the manuscript by J. M. Chandlee and J. Varner are grate- fully acknowledged. We thank Suzanne Quick for ex- pert typing of the manuscript. This work was sup- ported, in part, by a research grant from the U.S. Environmental Protection Agency to J.G.S. and by an INTA-BID Fellowship to A.A.
REFERENCES Acevedo A, Scandalios JG (1990): Expression of the catalase and su-
peroxide dismutase genes in mature pollen in maize. Theor Appl Genet 80:705-711.
Baum JA, Scandalios JG (1979): Developmental expression and in- tracellular localization of superoxide dismutases in maize. Differ- entiation 13:133-140.
Baum JA, Scandalios JG (1982): Multiple genes controlling superox- ide dismutase expression in maize. J Hered 73:95-100.
Beers RF, Sizer IW (1952): A spectrophotometric method for measur- ing the breakdown of hydrogen peroxide by catalase. J Biol Chem 195:133-140.
Bethards LA, Skadsen RW, Scandalios J G (1987): Isolation and char- acterization of a cDNA clone for the Cat2 gene in maize and its homology with other catalases. Proc Natl Acad Sci USA 84:6830- 6834.
Cannon RE, Scandalios JG (1989): Two cDNAs encode two nearly identical CuiZn superoxide dismutase proteins in maize. Mol Gen Genet 219:l-8.
Cannon RE, White JA, Scandalios JG (1987): Cloning of cDNA for maize superoxide dismutase 2 (SOD-2). Proc Natl Acad Sci USA 84:179-183.
Chirgwin JM, Przybyla AE, MacDonald RJ , Rutter MJ (1979): Isola- tion of biologically active RNA from sources enriched in ribonu- cleases. Biochemistry 18:5294-5299.
D’Ambrogio de Argiieso A (1986): “Manual de Tecnicas en Histologia Vegetal.” Buenos Aires: Hemisferio Sur, SA, p. 59.
Gallais A, Hugueet L, Berthet H, Bertin G, Braqua B, Mourguet A, Traineau R (1980): Preliminary evaluation of brown midrib maize hybrids for their feeding and agronomic value in France. In Pollmer WG, Phipps RH (eds): “Improvement of Quality Traits of Maize for Grain and Silage Use.” The Hague: Martinus Nijhoff Publishers, pp 319-339.
Hahlbrock K, Grisebach H (1979): Enzymic controls in the biosynthe- sis of lignin and flavonoids. Annu Rev Plant Physiol 30:105-130.
Halliwell B (1974): Supzroxide dismutase, catalase and glutathione peroxidase: Solutions to the problems of living with oxygen. New Phytol 73:1075-1086.
Havir EA, McHale NA (1989): Enhanced-peroxidatic activity in spe- cific catalase isozymes of tobacco, barley, and maize. Plant Physiol 91:812-815.
Irish EE, Nelson TM (1988): Development of maize plants from cul- tured shoot apices. Planta 175:9-12.
Jorgenson LR (1931): Brown midrib in maize and its linkage rela- tions. J Am SOC Agron 23549-557.
Kiesselbach TA (1949): The structure and reproduction of corn. Univ Nebraska, Agric Exp Sta Res Bull 161:25-30.
Kumazawa M (1961): Studies on the vascular course in maize plant. Phytomorphology 11:128-139.
Laurel1 C-B (1966): Quantitative estimation of proteins by electro- phoresis in agarose gel containing antibodies. Anal Biochem 15: 45-52.
Lowry OH, Rosebrough NJ, Farr AL, Randall R J (1951): Protein mea- surement with the folin phenol reagent. J Biol Chem 193:265-275.
McDaniel CN, Poethig RS (1988): Cell-lineage patterns in the shoot apical meristem of the germinating maize embryo. Planta 175:13- 22.
Poethig RS (1982): Maize-the plant and its parts. In Sheridan WF (edf: “Maize for Biological Research.” Charlottesville, VA: Plant Molecular Biology Association, pp 9-18.
Redinbaugh MG, Sabre M, Scandalios JG (1990): The distribution of catalase activity, isozyme protein, and transcript in the tissues of the developing maize seedling. Plant Physiol 92:375-380.
Redinbaugh MG, Wadsworth GJ, Scandalios JG (1988): Characteriza- tion of catalase transcripts and their differential expression in maize. Biochim Biophys Acta 951:104-116.
Roupakias DG, McMillin DE, Scandalios JG (1980): Chromosomal location of the catalase structural genes in Zea mays, using B-A translocations. Theor Appl Genet 58:211-218.
Scandalios JG (1968): Genetic control of multiple molecular forms of catalase in maize. Ann NY Acad Sci 151:274-293.
Scandalios JG (1979): Control of gene expression and enzyme differ- entiation. In Scandalios JG (ed): “Physiological Genetics.” New York: Academic Press, pp 63-1 07.
Scandalios JG (1987): The antioxidant enzyme genes Cat and Sod of maize: Regulation, functional significance, and molecular biology. In Rattazzi MC, Scandalios JG, Whitt GS (eds): “Isozymes: Current Topics in Biological and Medical Research, Volume 14, Molecular and Cellular Biology.” New York: Alan R. Liss, Inc., pp 19-44.
Skadsen RW, Scandalios JG (1987): Translational control of photo- induced expression of the Cat2 catalase gene during leaf develop- ment in maize. Proc Natl Acad Sci USA 84:2785-2789.
Sussex IM (1989): Developmental programming of the shoot me- ristem. Cell 56225-229.
Tsaftaris AS, Bosabalidis AM, Scandalios J G (1983): Cell-type-spe- cific gene expression and acatalasemic peroxisomes in a null Cat2 catalase mutant of maize. Proc Natl Acad Sci USA 80:4455- 4459.
Tsaftaris AS, Scandalios JG (1981): Genetic and biochemical charac- terization of a Cat2 catalase null mutant of Zea mays. Mol Gen Genet 181:158-163.
Wadsworth GJ, Scandalios J G (1989): The differential expression of the maize catalase genes during kernel development: The role of steady-state mRNA levels. Dev Genet 10:304-310.
Wadsworth GJ, Scandalios JG (1990): Molecular characterization of a catalase null allele at the Caf3 locus in maize. Genetics 125:867- 872.
White J A , Plant S, Cannon RE, Wadsworth GJ, Scandalios JG (1990): Developmental analysis of steady-state levels of CuiZn and Mn su- peroxide dismutase mRNAs in maize tissues. Plant Cell Physiol 31:1163-1167.
White JA, Scandalios JG (1988): Isolation and characterization of a cDNA for mitochondria1 manganese superoxide dismutase (SOD-3) of maize and its relation to other manganese superoxide dismu- tases. Biochim Biophys Acta 951%-70.












![Free Radic Biol Med. The induction of human superoxide dismutase … · 2018-02-01 · superoxide dismutase (SOD) and catalase, by virtue of their ... retinol [12], malathion [13],](https://img.pdfslide.net/doc/110x75/5f41fa3e32b0d31ee07cf958/free-radic-biol-med-the-induction-of-human-superoxide-dismutase-2018-02-01-superoxide.jpg)
















