Embed Size (px)
Citation preview

Catecholamine Inputs to Expiratory LaryngealMotoneurons in Rats
Wen-Jing Zhao,1 Qi-Jian Sun,2 Rui-Chen Guo,1 and Paul M. Pilowsky3*1Institute of Clinical Pharmacology, Qilu Hospital of Shandong University, Jinan, Shandong 250012, China2Australian School of Advanced Medicine, Macquarie University, Sydney, NSW 2109, Australia3Heart Research Institute and University of Sydney, Sydney, NSW 2042, Australia
ABSTRACTMany respiration-related interneurons and motoneurons
receive a catecholaminergic input, but the extent and
distribution of this input to recurrent laryngeal moto-
neurons that innervate intrinsic muscles of the larynx
are not clear. In the present study, we examined the
catecholaminergic input to expiratory laryngeal moto-
neurons in the caudal nucleus ambiguus by combining
intracellular labeling of single identified motoneurons,
with immunohistochemistry to reveal tyrosine hydroxy-
lase immunoreactive (catecholaminergic) terminal vari-
cosities. Close appositions were found between the two
structures, with 18 6 5 close appositions per moto-
neuron (n 5 7). Close appositions were more frequently
observed on distal rather than proximal dendrites. Axo-
somatic appositions were not seen. In order to deter-
mine the source of this input, microinjections of
cholera toxin B subunit (1%, 20 nl) were made into the
caudal nucleus ambiguus. Retrogradely labeled neurons,
located in the ipsilateral nucleus tractus solitarius and
the area postrema, were tyrosine hydroxylase-positive.
Our results not only demonstrate details of the extent
and distribution of potential catecholamine inputs to
the expiratory laryngeal motoneuron, but further indi-
cate that the inputs, at least in part, originate from the
dorsomedial medulla, providing a potential anatomical
basis for previously reported catecholaminergic effects
on the laryngeal adductor reflex. J. Comp. Neurol.
000:000–000, 2014.
VC 2014 Wiley Periodicals, Inc.
INDEXING TERMS: laryngeal motoneurons; tyrosine hydroxylase; nucleus tractus solitarius; intracellular recording;
respiration; cholera toxin B subunit; RRIDs AB: 10013220 and 477560
Catecholaminergic neurons comprise three subgroups
based on their complement of the catecholamine-
synthesizing enzymes, tyrosine hydroxylase (TH),
dopamine-beta-hydroxylase ("A1–7" cell groups) and
phenylethanolamine-N-methyltransferase ("C1–3" cell
groups) (Kalia et al., 1985). Catecholamine neurons
(TH-immunoreactive, TH-ir) are found in both the ventro-
lateral (A1/C1 cell groups) and the dorsomedial (A2/
C2 cell groups) medulla oblongata (H€okfelt et al., 1984;
Kalia et al., 1985; Pilowsky et al., 1990; Ellenberger
et al., 1990). In the ventrolateral medulla, TH-ir terminal
varicosities form close appositions with respiratory neu-
rons in the ventral respiratory column and motoneurons
in the rostral nucleus ambiguus (NA) (Sun et al., 1994).
Noradrenergic projections (TH-ir and dopamine-b-
hydroxylase-ir, but not phenylethanolamine-N-methyl-
transferase-ir) also arise from A5–7 cell groups and pro-
ject to the hypoglossal, facial, trigeminal motor nucleus
as well as motoneurons in the spinal cord (Card et al.,
1986; Grzanna et al., 1987; Lyons and Grzanna, 1988;
Aldes et al., 1990, 1992). Despite these extensive stud-
ies, it is still not clear if, and to what extent, recurrent
laryngeal motoneurons receive any TH inputs.
The caudal NA is mainly occupied by motoneurons
that project to, and regulate, intrinsic laryngeal muscles
via the recurrent laryngeal nerve (Bieger and Hopkins,
1987). Inspiratory laryngeal motoneurons innervate the
posteriod cricoarytenoid muscle, acting to open the
glottis during inspiration (Berkowitz et al., 1999). Expira-
tory laryngeal motoneurons (ELMs) innervate the thyro-
arytenoid, and other laryngeal constrictor muscles,
Grant sponsor: National Health and Medical Research Council ofAustralia; Grant number: 1024489; Grant sponsor: Australian ResearchCouncil; Grant numbers: DP110102110; G 11S 5957; Grant sponsor:National Nature Science of China; Grant number: 81202575.
*CORRESPONDENCE TO: Professor Paul M. Pilowsky, Heart ResearchInstitute, 7 Eliza St, Newtown 2042, Sydney, NSW, Australia.E-mail: [email protected]
Received October 21, 2013; Revised September 12, 2014;Accepted September 12, 2014.DOI 10.1002/cne.23677Published online Month 00, 2014 in Wiley Online Library(wileyonlinelibrary.com)VC 2014 Wiley Periodicals, Inc.
The Journal of Comparative Neurology | Research in Systems Neuroscience 00:00–00 (2014) 1
RESEARCH ARTICLE

acting to close the glottis and protect the airway from
aspiration during swallowing (Barillot et al., 1990; Yosh-
ida et al., 1998; Berkowitz et al., 2005; Ludlow, 2005).
The ELMs are partly controlled by monosynaptic con-
nections from respiratory neurons in the B€otzinger com-
plex and other ventral respiratory groups (Jiang and
Lipski, 1990; Ono et al., 2006; Sun et al., 2008). On
the other hand, during stimulation of the superior laryn-
geal nerve, ELMs are activated with a nonrespiratory burst
activity by neurons in the nucleus tractus solitarius (NTS)
around the obex level, which plays a critical role in coordi-
nating apnea, swallowing, and other airway protective
reflexes (Gestreau et al., 2000; Abu-Shaweesh et al.,
2001; Jean, 2001; Saito et al., 2002; Sun et al., 2011). In
addition, intravenous application of dopamine D1 receptor
antagonists is known to facilitate the laryngeal adductor
response in the rat (Feng et al., 2009). However, further
anatomical evidence of a dopamine input to the ELMs
remains unknown.
Given that TH-ir terminals and ELMs are both located
in the caudal NA (Kalia et al., 1985; Ludlow, 2005; Sun
et al., 2008), we hypothesized that there may be direct
contact between the two structures. This was examined
in the present study after intracellular labeling of ELMs
combined with immunohistochemistry to reveal TH-ir ter-
minals. In addition, in a separate tract-tracing study, our
findings not only confirm the existence of close apposi-
tions between the TH-ir varicosities and ELMs, but also
indicate that the TH inputs originate, at least in part,
from TH-ir neurons located in the dorsomedial medulla.
MATERIALS AND METHODS
Data were obtained from nine male Sprague–Dawley
rats (350–500 g, supplied by the Animal Resource
Centre, WA, Australia). All procedures were approved
by the Animal Care and Ethics Committee of Macquarie
University (http://www.nhmrc.gov.au/publications/
synopses/ea16syn.htm).
Animal preparationFive rats were anesthetized with sodium pentobarbi-
tone (72 mg/kg, ip, Virbac Animal Health, NSW, Aus-
tralia). The trachea, right femoral artery, and vein were
cannulated for artificial ventilation, blood pressure record-
ing and drug administration, respectively. Additional doses
of pentobarbitone sodium (3 mg/kg, iv) were adminis-
tered to maintain adequate anesthesia, as evidenced by a
stable blood pressure and phrenic nerve discharge, with
only minor autonomic responses (<10 mmHg increase in
blood pressure) in response to firm paw pinch. The left
phrenic and recurrent laryngeal nerves (RLN) were dis-
sected for recording or stimulation with standard bipolar
electrodes (Pilowsky et al., 1990). The rat was mounted
in a stereotaxic frame, paralyzed with pancuronium bro-
mide (1 mg/kg, with additional doses of 0.5 mg/kg, iv as
needed, AstraZeneca, Australia), and artificially ventilated
with oxygen-enriched air. Rectal temperature was main-
tained at 35–37�C with a homeothermic heating blanket
(Harvard Apparatus, Dover, MA). Part of the occipital and
parietal bones were removed to expose the dorsal sur-
face of medulla, and a laminectomy from C3 to C6 was
made to reduce brainstem movement.
Intracellular recording and labelingThe caudal NA was located by antidromic stimulation
of the RLN using low-impedance electrodes (5–10 Mx,
3M NaCl, Harvard Apparatus, SDR, Australia) to detect
field potentials. In three out of the five rats, when the
RLN recordings showed a weak respiratory activity the
left vagus was dissected for stimulation to locate the NA
and to identify ELMs (Gestreau et al., 2000). Intracellular
recordings were made with glass microelectrodes
(<1 lm tip diameter) containing 1.5% neurobiotin
(Vector Laboratories, Burlingame, CA) in 1M KCl (resist-
ance: 40–60 Mx). Only neurons with a stable membrane
potential more negative than 240 mV were labeled with
neurobiotin by using depolarizing current pulses (4–5 nA;
70% duty cycle for 3–5 minutes). After labeling, the elec-
trodes were rapidly removed from the cell and the
extracellular potential noted. One hour after labeling,
rats were perfused transcardially with 400 ml of saline,
followed by 400 ml of fixative containing 4% paraformal-
dehyde in 0.1M phosphate buffer (pH 7.4).
Microinjection of retrograde tracerA further four rats were anesthetized with urethane
(1.0–1.5 g/kg, ip) and maintained with additional doses
of pentobarbitone sodium (see above; 3 mg/kg, iv). The
right femoral vein was cannulated for drug administra-
tion. The left vagus nerve was dissected for stimulation
with standard bipolar electrodes. The rat was then
placed in a stereotaxic frame and a partial occipital cra-
niotomy performed to expose the dorsal surface of the
medulla oblongata. After the caudal NA was located by
antidromic stimulation of the vagus (10–15 mV, 0.2 ms),
cholera toxin B subunit (CTB; List Biological Laboratories,
Campbell, CA) was microinjected (1%, 20 nl) into the NA
at 0.5 mm caudal to the obex. After the CTB injection
(20–24 hours), the rat was perfused transcardially with
400 ml of saline, followed by 400 ml 4% paraformalde-
hyde in 0.1M phosphate buffer (pH 7.4).
ImmunohistochemistryThe medulla oblongata was removed from the rat
and post-fixed in the same fixative overnight. Vibrating
W.-J. Zhao et al.
2 The Journal of Comparative Neurology |Research in Systems Neuroscience

microtome sections (50 lm; Leica VT1200S) were
placed in 50% ethanol for 30 minutes. After three
washes with Tris phosphate-buffered saline (PBS) (Sun
et al., 1994), the sections from the intracellular labeling
experiments were placed in 10% normal horse serum
(Jackson ImmunoResearch, West Grove, PA) in Tris PBS
for 2 hours before incubation for 3 days in 10% normal
horse serum in Tris PBS (with 0.05% merthiolate) con-
taining ExtrAvidin-horseradish peroxidase (1:1,000 dilu-
tion, Sigma, St. Louis, MO, E2886) and the mouse
monoclonal antibody to TH (1:5,000; Sigma-Aldrich,
Cat. no. T1299, RRID: AB_477560) (Table 1). The mor-
phology of neurobiotin-filled neurons was revealed using
an imidazole-intensified diaminobenzidine reaction
(Pilowsky et al., 1991). Sections were then incubated
for 48 hours in a biotinylated donkey antimouse anti-
body (Jackson ImmunoResearch Laboratories, Lot no.
60651, 1:500 in 5% normal horse serum in Tris PBS)
and then incubated for 24 hours in ExtrAvidin-
horseradish peroxidase (1:1,500 dilution). TH-ir varicos-
ities were visualized with a nickel-intensified diamino-
benzidine reaction. Sections were mounted onto
gelatin-treated slides, dehydrated, and slides were cov-
erslipped with Ultramount No. 4 (Fronine, Australia).
Sections from the CTB microinjection experiments
were incubated for 48 hours in 10% normal horse
serum in Tris PBS (with 0.05% merthiolate) containing
goat anti-CTB (1:10,000 dilution; List Biological Labora-
tories, Campbell, CA, Cat. no. 703. RRID: AB
10013220. (Table 1), and mouse monoclonal antibody
to TH (1:2,000; Sigma-Aldrich, Cat. no. T1299, RRID:
AB_477560) (Table 1). After three washes in Tris PBS,
sections were incubated for 24 hours in 5% normal
horse serum in Tris PBS containing FITC-donkey anti-
sheep IgG (713-095-147, Jackson, 1:500 dilution) and
Cy3-donkey antimouse IgG (715-066-150, Jackson,
1:500 dilution) for visualization of CTB and TH-ir,
respectively. All incubations were performed at 4�C
with continuous gentle agitation and followed by 33
30-minute washes in Tris PBS. Sections were mounted
on microscope slides in Vectorshield mounting medium
(Vector Laboratories, Burlingame, CA).
Antibody characterizationThe suppliers specifications of the primary antibodies
used in this study are summarized in Table 1. Primary
antibodies were titrated on 4% paraformaldehyde-fixed
rat brainstem sections. The optimal concentrations of
the primary antibodies were determined as the dilution
at which the maximum number of immunoreactive
structures was observed with a minimal level of back-
ground staining.
The monoclonal anti-TH was derived from a hybrid-
oma produced by fusion of mouse myeloma cells and
splenocytes from an immunized mouse. Rat TH was
used as the immunogen. The antibody recognizes a sin-
gle band of �60 kDa on western blot from rat brain
(manufacturer’s technical information), and stained cat-
echolaminergic cells present in the brain and spinal
cord, and in sympathetic (Shi et al., 2008), chromaffin,
and entero-chromaffin systems. We confirmed that cell
morphology and regional distribution patterns of TH-
labeled neurons were identical to the results obtained
in previous studies (Pilowsky et al., 1990; Kawano
et al., 2006; Shi et al., 2008). A goat antiserum raised
against the CTB subunit (choleragenoid) was used. This
antibody recognizes a single band of CTB by western
blot (Xu et al., 2009). Goat antiserum to CTB did not
bind to any endogenous epitopes in the rat brain sec-
tions (Ge et al., 2010).
Biotinamide (neurobiotin) is an amino derivative of
biotin often used as an intracellular label for neurons.
Compared to biocytin and other neuronal labels, it is
more soluble, iontophoreses better, remains in cells lon-
ger, and can be fixed with formalin or glutaraldehyde.
Detection of neurobiotin using an avidin-peroxidase
technique revealed that only the one or two neuronal
cell somata that had been filled intracellularly were
unambiguously stained in any one brain.
Data analysisThe intracellularly labeled neurons were recon-
structed by light microscopy (340 objective) with a
camera lucida drawing tube. The criterion for a close
TABLE 1.
Primary Antibodies Used
Antigen Description of Immunogen
Source, Host Species, Cat.#,
Clone or Lot#, RRID Concentration/ Dilution
Tyrosine hydroxylase Rat tyrosine hydroxylase Sigma-Aldrich, mouse monoclonal,Cat#T1299, RRID: AB 477560
1 mg/ml, 1:5,000 (light),2.5 mg/ml, 1:2,000 (fluorescence)
Cholera Toxin Bsubunit (CTB)
B subunit (choleragenoid) List Biological Laboratories, goatantiserum, Cat#703, RRID: AB10013220
1:10,000
Catecholamine inputs to laryngeal motoneurons
The Journal of Comparative Neurology | Research in Systems Neuroscience 3
apposition was the presence of a side-by-side contact
formed between two structures with no discernible
intervening gap under an oil immersion objective lens
(3100). For estimation of the number and phenotype
of neurons in tract-tracing study, a one-in-four series of
50-lm coronal sections that contained the retrogradely
labeled neurons were chosen. All sections were exam-
ined and photographed using a Zeiss Axioimager Z2
microscope (Carl Zeiss Microimaging) with a Zeiss Axio-
cam HRm monochrome digital camera and Zeiss Imag-
ing software (AxioVision Rel. 4.8). Data are presented
as mean 6 SD.
RESULTS
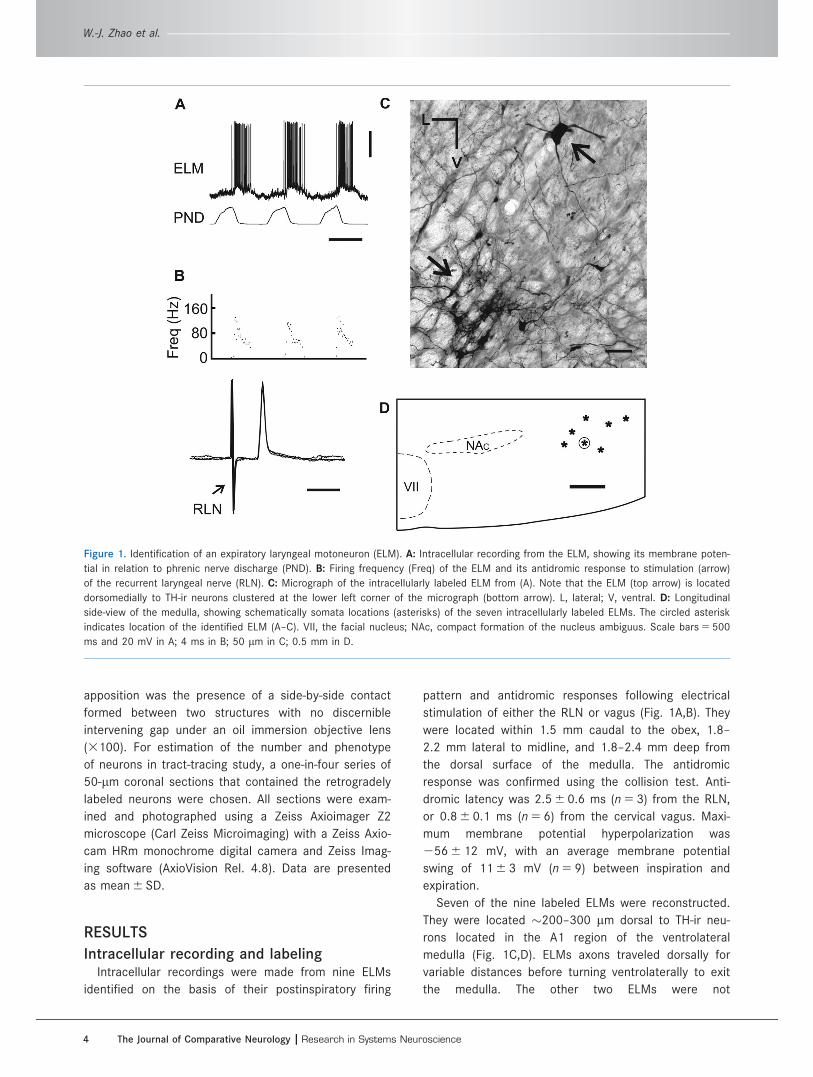
Intracellular recording and labelingIntracellular recordings were made from nine ELMs
identified on the basis of their postinspiratory firing
pattern and antidromic responses following electrical
stimulation of either the RLN or vagus (Fig. 1A,B). They
were located within 1.5 mm caudal to the obex, 1.8–
2.2 mm lateral to midline, and 1.8–2.4 mm deep from
the dorsal surface of the medulla. The antidromic
response was confirmed using the collision test. Anti-
dromic latency was 2.5 6 0.6 ms (n 5 3) from the RLN,
or 0.8 6 0.1 ms (n 5 6) from the cervical vagus. Maxi-
mum membrane potential hyperpolarization was
256 6 12 mV, with an average membrane potential
swing of 11 6 3 mV (n 5 9) between inspiration and
expiration.
Seven of the nine labeled ELMs were reconstructed.
They were located �200–300 lm dorsal to TH-ir neu-
rons located in the A1 region of the ventrolateral
medulla (Fig. 1C,D). ELMs axons traveled dorsally for
variable distances before turning ventrolaterally to exit
the medulla. The other two ELMs were not
Figure 1. Identification of an expiratory laryngeal motoneuron (ELM). A: Intracellular recording from the ELM, showing its membrane poten-
tial in relation to phrenic nerve discharge (PND). B: Firing frequency (Freq) of the ELM and its antidromic response to stimulation (arrow)
of the recurrent laryngeal nerve (RLN). C: Micrograph of the intracellularly labeled ELM from (A). Note that the ELM (top arrow) is located
dorsomedially to TH-ir neurons clustered at the lower left corner of the micrograph (bottom arrow). L, lateral; V, ventral. D: Longitudinal
side-view of the medulla, showing schematically somata locations (asterisks) of the seven intracellularly labeled ELMs. The circled asterisk
indicates location of the identified ELM (A–C). VII, the facial nucleus; NAc, compact formation of the nucleus ambiguus. Scale bars 5 500
ms and 20 mV in A; 4 ms in B; 50 lm in C; 0.5 mm in D.
W.-J. Zhao et al.
4 The Journal of Comparative Neurology |Research in Systems Neuroscience
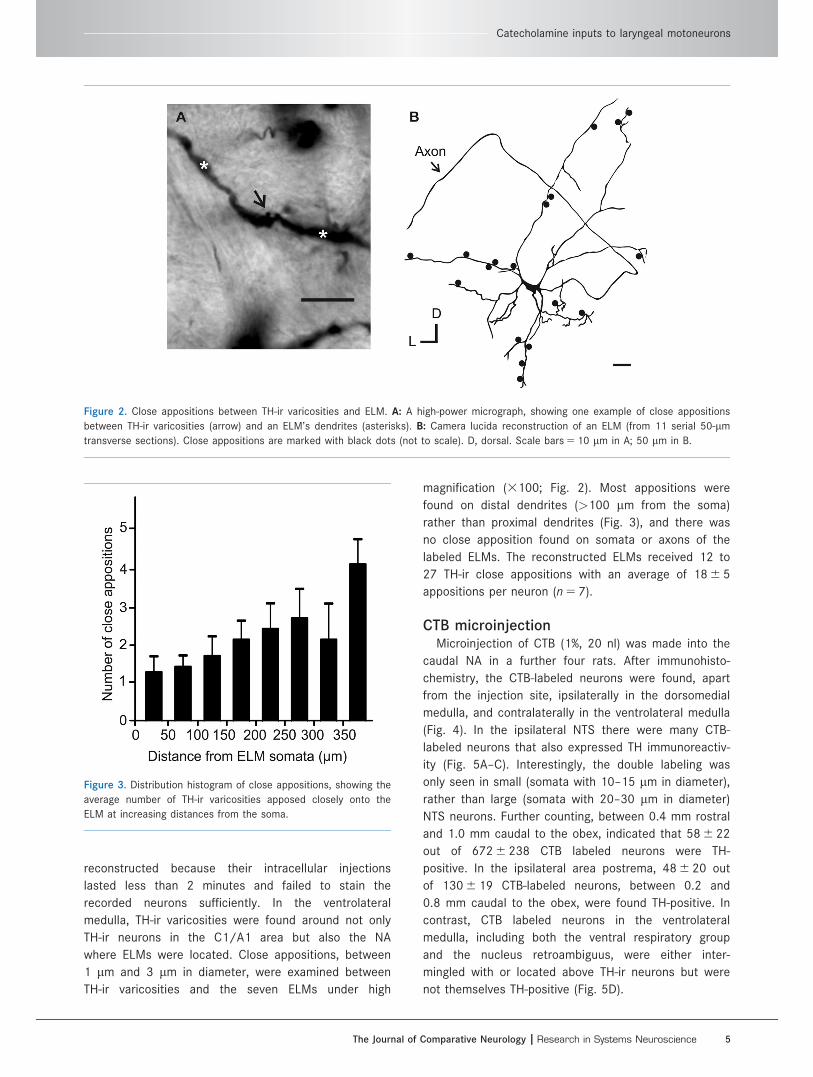
reconstructed because their intracellular injections
lasted less than 2 minutes and failed to stain the
recorded neurons sufficiently. In the ventrolateral
medulla, TH-ir varicosities were found around not only
TH-ir neurons in the C1/A1 area but also the NA
where ELMs were located. Close appositions, between
1 lm and 3 lm in diameter, were examined between
TH-ir varicosities and the seven ELMs under high
magnification (3100; Fig. 2). Most appositions were
found on distal dendrites (>100 lm from the soma)
rather than proximal dendrites (Fig. 3), and there was
no close apposition found on somata or axons of the
labeled ELMs. The reconstructed ELMs received 12 to
27 TH-ir close appositions with an average of 18 6 5
appositions per neuron (n 5 7).
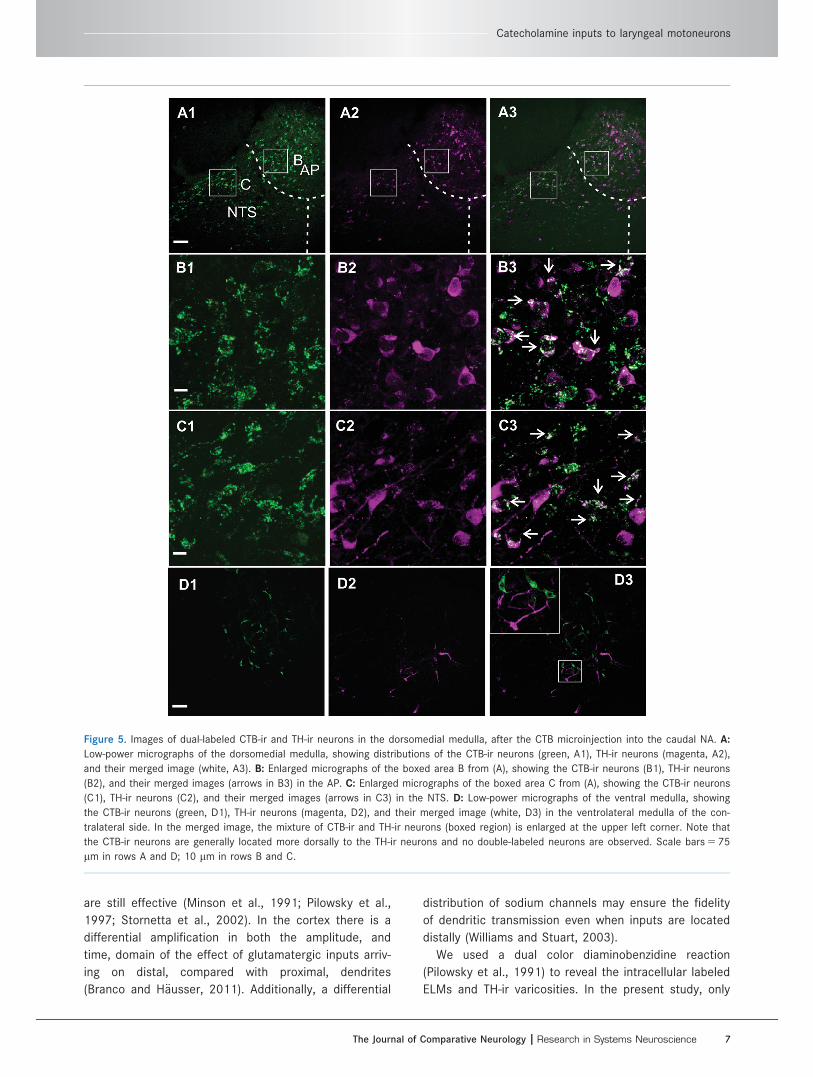
CTB microinjectionMicroinjection of CTB (1%, 20 nl) was made into the
caudal NA in a further four rats. After immunohisto-
chemistry, the CTB-labeled neurons were found, apart
from the injection site, ipsilaterally in the dorsomedial
medulla, and contralaterally in the ventrolateral medulla
(Fig. 4). In the ipsilateral NTS there were many CTB-
labeled neurons that also expressed TH immunoreactiv-
ity (Fig. 5A–C). Interestingly, the double labeling was
only seen in small (somata with 10–15 lm in diameter),
rather than large (somata with 20–30 lm in diameter)
NTS neurons. Further counting, between 0.4 mm rostral
and 1.0 mm caudal to the obex, indicated that 58 6 22
out of 672 6 238 CTB labeled neurons were TH-
positive. In the ipsilateral area postrema, 48 6 20 out
of 130 6 19 CTB-labeled neurons, between 0.2 and
0.8 mm caudal to the obex, were found TH-positive. In
contrast, CTB labeled neurons in the ventrolateral
medulla, including both the ventral respiratory group
and the nucleus retroambiguus, were either inter-
mingled with or located above TH-ir neurons but were
not themselves TH-positive (Fig. 5D).
Figure 2. Close appositions between TH-ir varicosities and ELM. A: A high-power micrograph, showing one example of close appositions
between TH-ir varicosities (arrow) and an ELM’s dendrites (asterisks). B: Camera lucida reconstruction of an ELM (from 11 serial 50-lm
transverse sections). Close appositions are marked with black dots (not to scale). D, dorsal. Scale bars 5 10 lm in A; 50 lm in B.
Figure 3. Distribution histogram of close appositions, showing the
average number of TH-ir varicosities apposed closely onto the
ELM at increasing distances from the soma.
Catecholamine inputs to laryngeal motoneurons
The Journal of Comparative Neurology | Research in Systems Neuroscience 5

DISCUSSION
In this study, we demonstrate for the first time close
appositions between TH-ir varicosities and ELMs in the
rat. A subsequent tract-tracing study after CTB microin-
jection into the caudal NA revealed that retrogradely
labeled neurons were mainly present in the ipsilateral
NTS and area postrema, with a proportion of them colo-
calized with TH-immunoreactivity. These results suggest
that catecholamine projections may provide a direct
input to the ELMs in control of the laryngeal constrictor
muscles, laying a potential anatomical basis for previ-
ously reported dopamine effects on laryngeal adductor
reflex (Feng et al., 2009).
Catecholamine inputs to the ELMsIn the present study, we demonstrate that ELMs
receive close appositions from TH-ir varicosities. The
close appositions were found more commonly on the
distal rather than the proximal dendrites. No TH-ir vari-
cosity was observed to make close appositions with
somata or axons of the labeled ELMs. In comparison
with previous studies, TH-ir close appositions had a
similar distribution and density onto the respiratory
neurons in the B€otzinger complex and other ventral
respiratory neurons (Sun et al., 1994). In contrast,
fewer TH-ir varicosities were apposed closely to respira-
tory motoneurons in the rostral NA (Sun et al., 1994).
In addition, a similar pattern of TH inputs to motoneur-
ons in the hypoglossal nucleus has been reported, in
which TH-ir terminals were shown to form synaptic con-
nections with the hypoglossal motoneurons more fre-
quently on the dendrites rather than the somata (Aldes
et al., 1990). Based on our results that TH inputs to
ELMs are directed to their distal dendrites with a mod-
est number, we believe that the catecholamine inputs
may be more important in modulating responses to
other inputs, rather than in dominating, the firing of the
ELMs. Given the location on the distal dendrites, such
inputs may affect the electrotonic size of the neuron
differentially, so that distally located glutamatergic TH
inputs (all brainstem TH neurons are also glutamatergic)
Figure 4. Distribution of the retrogradely labeled CTB neurons in the medulla. A: Microinjection site of CTB (20 nl) into the caudal NA. CC,
central canal. B: Enlarged micrograph from boxed region B from (A), showing distribution of the CTB-labeled neurons in the dorsomedial
medulla. AP, area postrema; NTSi, ipsilateral nucleus tractus solitaries (NTS); NTSc, contralateral NTS. Please note that the CTB labeled
neurons are located in the ipsilateral, but not contralateral, NTS and AP. C: Enlarged micrograph from boxed region C from (A), showing
distribution of the CTB-labeled neurons in the ventrolateral medulla (circled by dashed line), with ventral surface indicated by the arrow.
Scale bars 5 500 lm in A; 100 lm in B,C.
W.-J. Zhao et al.
6 The Journal of Comparative Neurology |Research in Systems Neuroscience
are still effective (Minson et al., 1991; Pilowsky et al.,
1997; Stornetta et al., 2002). In the cortex there is a
differential amplification in both the amplitude, and
time, domain of the effect of glutamatergic inputs arriv-
ing on distal, compared with proximal, dendrites
(Branco and H€ausser, 2011). Additionally, a differential
distribution of sodium channels may ensure the fidelity
of dendritic transmission even when inputs are located
distally (Williams and Stuart, 2003).
We used a dual color diaminobenzidine reaction
(Pilowsky et al., 1991) to reveal the intracellular labeled
ELMs and TH-ir varicosities. In the present study, only
Figure 5. Images of dual-labeled CTB-ir and TH-ir neurons in the dorsomedial medulla, after the CTB microinjection into the caudal NA. A:
Low-power micrographs of the dorsomedial medulla, showing distributions of the CTB-ir neurons (green, A1), TH-ir neurons (magenta, A2),
and their merged image (white, A3). B: Enlarged micrographs of the boxed area B from (A), showing the CTB-ir neurons (B1), TH-ir neurons
(B2), and their merged images (arrows in B3) in the AP. C: Enlarged micrographs of the boxed area C from (A), showing the CTB-ir neurons
(C1), TH-ir neurons (C2), and their merged images (arrows in C3) in the NTS. D: Low-power micrographs of the ventral medulla, showing
the CTB-ir neurons (green, D1), TH-ir neurons (magenta, D2), and their merged image (white, D3) in the ventrolateral medulla of the con-
tralateral side. In the merged image, the mixture of CTB-ir and TH-ir neurons (boxed region) is enlarged at the upper left corner. Note that
the CTB-ir neurons are generally located more dorsally to the TH-ir neurons and no double-labeled neurons are observed. Scale bars 5 75
lm in rows A and D; 10 lm in rows B and C.
Catecholamine inputs to laryngeal motoneurons
The Journal of Comparative Neurology | Research in Systems Neuroscience 7

side-by-side contacts between reconstructed ELMs and
TH-ir varicosities were counted as close appositions.
While this criterion reduces the chance of false-positive
counts, it also inevitably underestimates the actual
number of close appositions on ELMs. On the other
hand, a close apposition is not definitive evidence of
the existence of synapse. Every close apposition cannot
be guaranteed to form a synaptic connection between
the two structures. In a previous study, we demon-
strated that about half of the close appositions
between substance-P-containing varicosities and sympa-
thetic preganglionic neurons formed synapses or direct
contacts when examined by electron microscopy (Pilow-
sky et al., 1992). Nevertheless, examination of close
appositions provides a useful and effective way to
assess the distribution of neurochemical inputs over
the entire somatodendritic tree of neurons and to com-
pare neurochemical inputs among different neuronal
groups (Paul et al., 1992; Murphy et al., 1995; Sun
et al., 2003). Given that our criteria will underestimate
synaptic input, but on the other hand some true synap-
ses may be missed or falsely classified, we believe that
our estimate may be reasonably accurate.
Origin of the catecholamine inputsAfter microinjection of CTB, we found that the retro-
gradely labeled neurons were located ipsilaterally in the
dorsomedial medulla and contralaterally in the ventro-
lateral medulla, including the nucleus retroambiguus
(Holstege, 1989). These results are in good agreement
with previous studies (Hayakawa et al., 2000; Ono
et al., 2006; Van Daele and Cassell, 2009). A novel
finding of this study, however, is that a proportion of
the CTB-labeled neurons in the NTS and area postrema
were also TH-ir-containing. In contrast, none of the
CTB-labeled neurons in the ventrolateral medulla were
TH-positive. The much closer proximity of unlabeled TH
neurons in the ventrolateral medulla to the injection
site argues against the findings being due to nonspe-
cific uptake by axons of passage. Similarly, the finding
of labeling in the contralateral, but not ipsilateral, NTS
also argues against the notion that the findings
occurred as a result of nonspecific uptake. However,
we cannot exclude that the injection also involves the
neurons surrounding the caudal NA, resulting in a small
portion of the retrogradely labeled neurons that do not
originate from the motoneurons but neurons of other
systems. Our study also agrees with those previous
studies that demonstrate monosynaptic inputs from
contralateral neurons of the ventral respiratory group to
recurrent laryngeal motoneurons (Ono et al., 2006;
Shiba et al., 2007). Although respiratory neurons them-
selves are not TH-positive (Ellenberger et al., 1990;
Pilowsky et al., 1990), we cannot rule out the possibility
that TH-ir neurons from other brainstem regions may
provide inputs to the ELMs, since a tracing time of less
than 24 hours in the present study may limit transport
to sites more rostral than the brainstem. The extensive
surgical preparation necessary to identify the injection
sites precluded recovery from anesthesia in these
experiments, which were all acute and terminal. For
example, noradrenergic neurons are known to project
from A5–7 cell groups to hypoglossal, facial, trigeminal
motor nucleus as well as motoneurons in the spinal
cord (Card et al., 1986; Grzanna et al., 1987; Lyons
and Grzanna, 1988; Aldes et al., 1992).
Functional implicationsIn the present study, we demonstrate that ELMs
receive an apparently modest number of close appositions
from TH-ir varicosities. The precise relationship of input
numbers to functional effects observed is unclear. Never-
theless, it is noteworthy that most of the neurotransmitter
products of TH neurons are metabotropic, so that at
times of extreme activity, when release from these sites
is likely to include catecholamine containing dense core
vesicles, the functional effects of even a small number of
inputs may be quite large. Effects of catecholamine inputs
to respiratory neurons are known to be different, depend-
ing on catecholamine subpopulations (Murakoshi et al.,
1985; Errchidi et al., 1990; Li et al., 2008). For example,
noradrenaline application to the brainstem has predomi-
nantly an inhibitory effect on respiratory neurons, whereas
dopamine application increased the respiratory frequency
(Murakoshi et al., 1985; Errchidi et al., 1990). During elec-
tromyographic recording from the thyroarytenoid muscle,
a blockade of dopamine D1 receptor was found to reduce
the response latency, but increased the response ampli-
tude of the laryngeal adductor response (Feng et al.,
2009). The thyroarytenoid is one of the main laryngeal
constrictor muscles that are controlled by the ELMs
(Ludlow, 2005). More functional studies are obviously
needed to clarify what exact role played by the TH inputs
to the ELMs.
CONFLICT OF INTEREST
We declare that there is no known or potential conflict
of interest including any financial, personal or other rela-
tionships with other people or organizations within 3 years
of beginning the submitted work that could inappropri-
ately influence, or be perceived to influence, our work.
ROLE OF AUTHORS
All authors had full access to all the data in the
study and take responsibility for the integrity of the
W.-J. Zhao et al.
8 The Journal of Comparative Neurology |Research in Systems Neuroscience

data and the accuracy of the data analysis. Study con-
cept and design: Paul M. Pilowsky, Qi-Jian Sun, Wen-
Jing Zhao, and Rui-Chen Guo. Acquisition of data: Wen-
Jing Zhao and Qi-Jian Sun. Analysis and interpretation of
data: Wen-Jing Zhao and Qi-Jian Sun. Drafting of the
article: Wen-Jing Zhao. Critical revision of the article for
important intellectual content: Paul M. Pilowsky. Statis-
tical analysis: Wen-Jing Zhao. Obtained funding: Paul M.
Pilowsky, Wen-Jing Zhao, and Qi-Jian Sun. Administra-
tive, technical, and material support: Qi-Jian Sun and
Paul M. Pilowsky. Study supervision: Paul M. Pilowsky.
LITERATURE CITEDAbu-Shaweesh JM, Dreshaj IA, Haxhiu MA, Martin RJ. 2001.
Central GABAergic mechanisms are involved in apneainduced by SLN stimulation in piglets. J Appl Physiol 90:1570–1576.
Aldes LD, Shaw B, Chronister RB, Haycock JW. 1990. Cate-cholamine-containing axon terminals in the hypoglossalnucleus of the rat: An immuno-electron microscopicstudy. Exp Brain Res 81:167–178.
Aldes LD, Chapman ME, Chronister RB, Haycock JW. 1992.Sources of noradrenergic afferents to the hypoglossalnucleus in the rat. Brain Res Bull 29:931–942.
Barillot JC, Grelot L, Reddad S, Bianchi AL. 1990. Dischargepatterns of laryngeal motoneurones in the cat: an intra-cellular study. Brain Res 509:99–106.
Bautista TG, Sun QJ, Zhao WJ, Pilowsky PM. 2010. Cholinergicinputs to laryngeal motoneurons functionally identified invivo in rat: a combined electrophysiological and micro-scopic study. J Comp Neurol 518:4903–4916.
Berkowitz RG, Sun QJ, Goodchild AK, Pilowsky PM. 2005.Serotonin inputs to laryngeal constrictor motoneurons inthe rat. Laryngoscope 115:105–109.
Berkowitz RG, Sun QJ, Chalmers J, Pilowsky PM. 1999. Identi-fication of posterior cricoarytenoid (PCA) motoneurons inthe rat. Ann Otol Rhinol Laryngol 108:1033–1041.
Bieger D, Hopkins DA. 1987. Viscerotopic representation of theupper alimentary tract in the medulla oblongata in the rat:the nucleus ambiguus. J Comp Neurol 262:546–562.
Branco T, H€ausser M. 2011. Synaptic integration radients insingle cortical pyramidal cell dendrites. Neuron 9:885–892.
Card JP, Riley JN, Moore RY. 1986. The motor trigeminalnucleus of the rat: analysis of neuronal structure and thesynaptic organization of noradrenergic afferents. J CompNeurol 250:469–484.
Ellenberger HH, Feldman JL, Zhan WZ. 1990. Subnuclear orga-nization of the lateral tegmental field of the rat. II: Cate-cholamine neurons and ventral respiratory group. J CompNeurol 294:212–222.
Errchidi S, Hilaire G, Monteau R. 1990. Permanent release ofnoradrenaline modulates respiratory frequency in thenewborn rat: an in vitro study. J Physiol (Lond) 429:497–510.
Feng X, Henriquez VM, Walters JR, Ludlow CL. 2009. Effectsof dopamine D1 and D2 receptor antagonists on laryn-geal neurophysiology in the rat. J Neurophysiol 102:1193–1205.
Ge SN, Ma YF, Hioki H, Wei YY, Kaneko T, Mizuno N, Gao GD,Li JL. 2010. Coexpression of VGLUT1 and VGLUT2 in tri-geminothalamic projection neurons in the principal sen-sory trigeminal nucleus of the rat. J Comp Neurol 518:3149–3168.
Gestreau C, Gr�elot L, Bianchi AL. 2000. Activity of respiratorylaryngeal motoneurons during fictive coughing and swal-lowing. Exp Brain Res 130:27–34.
Grzanna R, Chee WK, Akeyson EW. 1987. Noradrenergic pro-jections to brainstem nuclei: evidence for differential pro-jections from noradrenergic subgroups. J Comp Neurol263:76–91.
Hayakawa T, Takanaga A, Maeda S, Ito H, Seki M. 2000.Monosynaptic inputs from the nucleus tractus solitarii tothe laryngeal motoneurons in the nucleus ambiguus ofthe rat. Anat Embryol 202:411–420.
H€okfelt T, Everitt BJ, Theodorsson-Norheim E, Goldstein M.1984. Occurrence of neurotensin-like immunoreactivityin subpopulations of hypothalamic, mesencephalic, andmedullary catecholamine neurons. J Comp Neurol 222:543–559.
Holstege G. 1989. Anatomical study of the final commonpathway for vocalization in the cat. J Comp Neurol 284:242–252.
Jean A. 2001. Brain stem control of swallowing: Neuronal net-work and cellular mechanisms. Physiol Rev 81:929–969.
Jiang C, Lipski J. 1990. Extensive monosynaptic inhibition ofventral respiratory group eurons by augmenting neuronsin the Botzinger complex in the cat. Exp Brain Res 81:639–648.
Kalia M, Fuxe K, Goldstein M. 1985. Rat medulla oblongata.II. Dopaminergic, noradrenergic (A1 and A2) and adrener-gic neurons, nerve fibers, and presumptive terminal proc-esses. J Comp Neurol 233:308–332.
Kawano M, Kawasaki A, Sakata-Haga H, Fukui Y, Kawano H,Nogami H, Hisano S. 2006. Particular subpopulations ofmidbrain and hypothalamic dopamine neurons expressvesicular glutamate transporter 2 in the rat brain.J Comp Neurol 498:581–592.
Li A, Emond L, Nattie E. 2008. Brainstem catecholaminergicneurons modulate both respiratory and cardiovascularfunction. Adv Exp Med Biol 605:371–376.
Ludlow CL. 2005. Central nervous system control of the laryn-geal muscles in humans. Resp Physiol Neurobiol 147:205–222.
Lyons WE, Grzanna R. 1988. Noradrenergic neurons withdivergent projections to the motor trigeminal nucleusand the spinal cord: a double retrograde neuronal label-ing study. Neuroscience 26:681–693.
Minson J, Pilowsky P, Llewellyn-Smith I, Kaneko T, Kapoor V,Chalmers J. 1991. Glutamate in spinally projecting neuronsof the rostral ventral medulla. Brain Res 555:326–331.
Murakoshi T, Suzue T, Tamai S. 1985. A pharmacologicalstudy on respiratory rhythm in the isolated brainstem-spinal cord preparation of the newborn rat. Br J Pharma-col 86:95–104.
Murphy SM, Pilowsky PM, Sun QJ, Llewellyn-Smith IJ. 1995.Thyrotropin-releasing hormone-immunoreactive varicos-ities synapse on rat phrenic motoneurons. J Comp Neu-rol 359:310–322.
Ono K, Shiba K, Nakazawa K, Shimoyama I. 2006. Synapticorigin of the respiratory-modulated activity of laryngealmotoneurons. Neuroscience 140:1079–1088.
Pilowsky PM, Jiang C, Lipski J. 1990. An intracellular study ofrespiratory neurons in the rostral ventrolateral medullaof the rat and their relationship to catecholamine-containing neurons. J Comp Neurol 301:604–617.
Pilowsky PM, Lipski J, Prestidge R, Jiang C. 1991. Dual fluo-rescence combined with a two-color immunoperoxidasetechnique: a new way of visualizing diverse neuronal ele-ments. J Neurosci Methods 36:185–193.
Pilowsky P, Llewellyn-Smith IJ, Lipski J, Chalmers J. 1992. Sub-stance P immunoreactive boutons form synapses with
Catecholamine inputs to laryngeal motoneurons
The Journal of Comparative Neurology | Research in Systems Neuroscience 9

feline sympathetic preganglionic neurons. J Comp Neurol320:121–135.
Pilowsky PM, Sun QJ, Llewellyn-Smith I, Arnolda L, ChalmersJ, Minson J. 1997. Phosphate-activated glutaminaseimmunoreactivity in brainstem respiratory neurons.J Auton Nerv Syst 63:85–90.
Saito Y, Ezure K, Tanaka I. 2002. Swallowing-related activitiesof respiratory and non-respiratory neurons in the nucleusof solitary tract in the rat. J Physiol (Lond) 540:1047–1060.
Shi H, Cui H, Alam G, Gunning WT, Nestor A, Giovannucci D,Zhang M, Ding HF. 2008. Nestin expression defines bothglial and neuronal progenitors in postnatal sympatheticganglia. J Comp Neurol 508:867–878.
Shiba K, Nakazawa K, Ono K, Umezaki T. 2007. Multifunc-tional laryngeal premotor neurons: their activities duringbreathing, coughing, sneezing, and swallowing.J Neurosci 27:5156–5162.
Stornetta RL, Sevigny CP, Guyenet PG. 2001. Vesicular gluta-mate transporter NPI/VGLUT2 mRNA is present in C1and several other groups of brainstem catecholaminergicneurons. J Comp Neurol 444:191–206.
Sun QJ, Pilowsky P, Minson J, Arnolda L, Chalmers J,Llewellyn-Smith IJ. 1994. Close appositions betweentyrosine hydroxylase immunoreactive boutons and respi-ratory neurons in the rat ventrolateral medulla. J CompNeurol 340:1–10.
Sun QJ, Berkowitz RG, Goodchild AK, Pilowsky PM. 2003. Sub-stance P inputs to laryngeal motoneurons in the rat.Resp Physiol Neurobiol 137:11–18.
Sun QJ, Berkowitz RG, Pilowsky PM. 2008. GABAA mediatedinhibition and post-inspiratory pattern of laryngeal con-strictor motoneurons in rat. Resp Physiol Neurobiol 162:41–47.
Sun QJ, Bautista TG, Berkowitz RG, Zhao WJ, Pilowsky PM.2011. The temporal relationship between non-respiratoryburst activity of expiratory laryngeal motoneurons andphrenic apnoea during stimulation of the superior laryn-geal nerve in rat. J Physiol (Lond) 589:1819–1830.
Van Daele DJ, Cassell MD. 2009. Multiple forebrain systemsconverge on motor neurons innervating the thyroaryte-noid muscle. Neuroscience 162:501–524.
Williams SR1, Stuart GJ. 2003. Voltage- and site-dependentcontrol of the somatic impact of dendritic IPSPs.J Neurosci 23:7358–7367.
Xu L, Ryugo DK, Pongstaporn T, Johe K, Koliatsos VE. 2009.Human neural stem cell grafts in the spinal cord ofSOD1 transgenic rats: differentiation and structural inte-gration into the segmental motor circuitry. J Comp Neu-rol 514:297–309.
Yoshida Y, Yatake K, Tanaka Y, Imamura R, Fukunaga H,Nakashima T, Hirano M. 1998. Morphological observationof laryngeal motoneurons by means of cholera toxin Bsubunit tracing technique. Acta Otolaryngol Suppl 98–105.
W.-J. Zhao et al.
10 The Journal of Comparative Neurology | Research in Systems Neuroscience