Embed Size (px)
Citation preview

J PlantPhysiol. Vol. 141. pp. 502-504(1993) Short Communication
Cauliflower Mosaic Virus Particles Alter the Sensitivity of Arabidopsis thaliana Seedlings to 2,4-0
ANDREW BANNISTER, ANDREW J. MAULE, and SIMON N. COVEY
Department of Virus Research, John Innes Institute, John Innes Centre, Colney Lane, Norwich NR4 7UH, UK
Received October 5,1992 . Accepted November 12,1992
Summary
Seed obtained from A rabidopsis thaliana plants can be germinated on agar plates and seedlings remain viable in the presence of low concentrations of the growth substance 2,4-D. However, when seed obtained from plants infected with cauliflower mosaic virus (CaMV) was similarly germinated in 2,4-D, seedlings developed cotyledons but then rapidly bleached and senesced. Bleached plants were also observed when purified particles of CaMV were mixed with seed obtained from healthy plants and germinated in 0.23-2.3 J.1M 2,4-D. The growth substance alone was found to cause bleaching only when increased above 2.3 J.1M. The effect of CaMV and 2,4-D on Arabidopsis was still observed after UV-inactivation of virus particles suggesting that the effect was caused by a change in the sensitivity of Arabidopsis to 2,4-D rather than virus replication.
Key words: A rabidopsis thaliana, cauliflower mosaic virus, 2,4.0 sensitivity.
Abbreviations: 2,4-D = 2,4-dichlorophenoxyacetic acid; CaMV = cauliflower mosaic virus.
Introduction
Arabidopsis thaliana is widely used as a model plant for physiological, pathological and molecular genetic studies (Meyerowitz and Pruitt, 1985). We have been exploiting cauliflower mosaic virus (CaMV) infection of Arabidopsis to understand the mechanisms of plant/virus interaction which lead to development of symptoms. Systemic infection following mechanical inoculation of susceptible species such as A rabidopsis with CaMV leads to vein clearing and chlorotic leaf symptoms, and plant stunting (Melcher, 1985; Stratford and Covey, 1989). As part of a programme to obtain large numbers of CaMV-infected Arabidopsis plants to screen genetic variants for altered response to the virus, we have experimented with various methods of inoculating seed. Although CaMV is not generally seed-transmitted, virus particles have been detected in the coats of seed obtained from CaMV-infected Arabidopsis (Melcher, 1985). In experiments designed to lower the susceptibility of seedlings to
© 1993 by Gustav Fischer Verlag, Stuttgart
endogenous CaMV particles, we observed an unexpected effect of 2,4-D upon seedling viability.
Materials and Methods
Seeds of Arabidopsis thaliana ecotype Columbia (Col-O), were surface sterilized by immersion for 20 min in 10 % bleach, rinsed 5 times in sterile water and sown on agar containing Murashige & Skoog salt mixture (Flow Laboratories) in Petri dishes (50-100 seeds per 5 cm dish or 200 - 500 seeds per 9 cm dish). Petri dishes were supplemented with 2,4-dichlorophenoxyacetic acid (2,4-D) and incubated at 21°C under 16 h illumination. For flowering, individual plants were removed from agar and pricked out into compost and grown in a 16h photoperiod.
Cauliflower mosaic virus isolate Cabb B-JI (Stratford and Covey, 1989) was propagated in turnip plants (Brassica rapa rapifera cv. «Just Right») and in Arabidopsis following mechanical inoculation. CaMV virions were purified as described by Hull et al. (1976). CaMV virions were UV-inactivated by incubation on a UV transil-

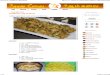
Fig. 1: Effects of 2,4-D and CaMV particles on growth of Arabi· do psis seedlings. Seeds were obtained from (a) healthy plants and germinated without 2,4-D (b) CaMV-infected plants without 2,4-D (c) healthy plants with 2.31!M 2,4-D (d) CaMV-infected plants with 2.3 I!M 2,4-D grown in a 16 h photoperiod for 21 days.
luminator (Chromatovue TM40) under the short wavelength setting.
Results
In an attempt to influence the susceptibility of Arabidopsis to CaMY with 2,4-D, we first established that sterilised Arabidopsis seed germinates normally in the presence of 2.3 ~M 2,4-D (Fig. 1 c). Although 100 % germination was observed, plants became stunted after three weeks propagation, compared with those grown without 2,4-D (Fig. 1 a), but remained viable indefinitely. Tissue in contact with the agar medium containing 2,4-D proliferated callus (Fig. 1 c). Seed obtained from Arabidopsis plants systemically-infected with CaMY germinated (70% germination rate) normally both in the presence and absence of 2,4-D. Seed from CaMY-infected plants continued to develop normally in the absence of the growth substance (Fig. 1 b) whereas that cultured in the presence of 2.3 ~M 2,4-D produced seedlings which became bleached (Fig. 1 d), failed to develop true leaves, and then rapidly progressed to senescence.
To determine whether the 2,4-D-induced senescence of Arabidopsis seedlings obtained from CaMY-infected plants was due to the presence of virus, seed obtained from healthy plants was mixed with a preparation of purified CaMY to a level of 0.5 ~g/ mL in the agar and cultured with and without 2.3 ~M 2,4-D. Seedlings germinated in the presence of only virus (0.5 ~g/mL) or only 2,4-D (2.3 ~M) developed normally whereas those germinated with a mixture of virus and
100
III 80 Cl
-= =c Q)
60 Q) III "C Q) .c 40 u III Q)
:c "if. 20
0
Arabidopsis sensitivity to 2,4-D and CaMV 503
0.23 0.45 1.36 2.3
2,4-D (~M)
-CaMV
4.5 13.6
Fig.2: Bleaching of Arabidopsis seedlings by 2,4-D in the presence and absence of CaMV particles. Seed was germinated in agar containing different concentrations of 2,4-D (shown as a logarithmic scale) either in the absence (-CaMV) or presence (+CaMV) of virus particles at O.5I!g/mL. The percentage of plants showing bleaching was determined after 21 days growth in a 16h photoperiod.
2,4-D, at these respective concentrations, became bleached and senesced as before.
Since the presence of CaMY virions was required to elicit senescence, albeit only in the presence of 2,4-D, we were interested to determine whether this was due to the growth substance lowering the tolerance of Arabidopsis to CaMY infection. However, we detected no viral products in the 2,4-Daffected plants indicative of viral replication (data not shown). To confirm our suspicion that the effect was not caused by virus infection per se, CaMY virions were treated with increasing doses of UY light to inactivate the particles. Exposure of CaMY virions to UY light for periods greater than 15 min was necessary to abolish their infectivity for turnip. However, none of the UY treatments affected the ability of CaMY virions (0.5 ~g/mL) to induce bleaching of Arabidopsis in the presence of 2.3 ~M 2,4-D.
To determine the minimum concentration of 2,4-D capable of causing bleaching, the growth substance was titrated against a constant amount (0.5 ~g/mL) of purified virus present in the agar on which Arabidopsis seed from healthy plants was germinated. Bleaching was observed with levels of 2,4-D above 0.23 ~M and increased to 100 % bleaching at a concentration of 1.36 ~M and above (Fig. 2). To test whether 2,4-D alone could induce bleaching, seedlings were germinated in 2,4-D in the absence of virus. Bleaching was first observed at a significantly higher concentration of 2,4-D (4.5 ~M) than in the presence of virus (Fig. 2). Complete bleaching was seen without virus at 13.6 ~M 2,4-D, a concentration one order of magnitude higher than the same effect observed in the presence of CaMY (Fig. 2).
To show if the concentration of virus particles was also important in altering the response of Arabidopsis seedlings to 2,4-D, the amount of virus was varied in agar containing 2.3 ~M 2,4-D. Complete bleaching was observed at virus concentrations of 0.4 ~g/mL and above. At 0.3 ~g/mL virus, approximately 70 % of the seedlings showed bleaching whilst no bleaching was apparent at a virus concentration of 0.1 ~g/

504 ANDREW BANNISTER, ANDREW J. MAULE, and SIMON N. COVEY
mL. Also, we were not able to obtain any bleaching of seedlings at greatly increased levels virus (50Ilg/mL) in the absence of 2,4-D.
Discussion
We have observed an unexpected effect of CaMV particles in enhancing senescence of A rabidopsis seedlings in the presence of 2,4-D. This effect appeared to be caused not as a result of CaMV replication but rather by lowering the threshold of sensitivity of plants to 2,4-D. The possibility that toxic factors co-purify with virus particles during their preparation can be dismissed by our initial observation that seed obtained from CaMV-infected Arabidopsis in the absence of exogenously added virus also increased sensitivity to 2,4-D. Although CaMV is not seed-transmitted, CaMV particles have been detected in coats of seed recovered from infected Arabidopsis plants (Melcher, 1985) and we presume that their presence at this location increases sensitivity to 2,4-D.
The finding that non-infectious CaMV particles increase the sensitivity of Arabidopsis to 2,4-D suggests that the effect is caused by some property of the coat protein possibly by
affecting uptake of 2,4-D into the plants. Somewhat intriguingly, we have been unable to induce the effect with purified particles of a mild strain of CaMV called Bari 1 (Stratford and Covey, 1989). Infection by the mild CaMV strain causes attenuated symptoms in our indicator host plant (turnip) but we do not observe any symptoms upon inoculation of A rabidopsis with Bari 1 (our unpublished observations). This property might also be related to symptom development in plants since the mild CaMV strain failed to induce the 2,4-D effect.
Acknowledgements
This work was supported in part by a grant from the AFRC under its Arabidopsis Programme.
References
HULL, R., R. J. SHEPHERD, and J. D. HARVEY: Journal of General Virology 31,93-100 (1976).
MELCHER, U.: Botanic Gazette 150, 139-147 (1985). MEYEROWITZ, E. M. and R. E. PRUITT: Science 229, 1214-1218
(1985). STRATFORD, R. and S. N. COVEY: Virology 172, 451-459 (1989).