Embed Size (px)
Citation preview

Cdc42 Stimulates Neurite Outgrowth and Formationof Growth Cone Filopodia and Lamellipodia
Michael D. Brown,1 Brandon J. Cornejo,2 Thomas B. Kuhn,2,* and James R. Bamburg2
1 Department of Anatomy and Neurobiology, Colorado State University, Fort Collins,Colorado 80523-16702 Department of Biochemistry and Molecular Biology, Colorado State University, Fort Collins,Colorado 80523-1870
Received 20 January 2000; accepted 21 February 2000
ABSTRACT: To assess the role of cdc42 duringneurite development, cmyc-tagged constitutively active(CA) and dominant negative (DN) cdc42 were expressedin dissociated primary chick spinal cord neurons usingadenoviral-mediated gene transfer. Three days after in-fection, >85% of the neurons in infected cultures ex-pressed cdc42 proteins, as detected by indirect immuno-fluorescence against cmyc. Growth cones of infectedneurons displayed 1.83- (CAcdc42) and 1.93-fold(DNcdc42) higher cmyc immunofluorescence per squaremicrometer than uninfected controls. CAcdc42 expres-sion stimulated growth cones, almost doubling growthcone size and number of filopodia, and increased neuritegrowth rates by 65–89%. In neurons plated onto fi-bronectin, the percent of growth cones with both filop-odia and lamellipodia increased from 71 to 92%. TotalTexas Red-phalloidin staining in these growth conesdoubled, and the percent of growth cones with F-actinlocalized to peripheral regions increased from 52% in
controls to 78% after CAcdc42 expression. Expressionof DNcdc42 did not significantly alter growth cone mor-phology or neurite growth rates. Addition of solublelaminin to spinal cord neurons resulted in the identicalphenotype as CAcdc42 expression, including changes ingrowth cone morphology, F-actin localization, and neu-rite growth rates. Significantly, expression of DNcdc42blocked the effects of laminin on growth cones. Theseresults show that cdc42 promotes neurite outgrowth andfilopodial and lamellipodial formation in growth conesand suggests that cdc42 and laminin share a commonsignaling pathway during neurite development. Addi-tion of laminin to CAcdc42-expressing neurons is inhib-itory to growth cones, indicating that laminin also mayactivate some other pathways. © 2000 John Wiley & Sons, Inc.
J Neurobiol 43: 352–364, 2000
Keywords:laminin; actin; neural development; rho GT-Pases; adenoviral mediated gene transfer
After proliferation and migration, newly formed neu-rons differentiate by extending axons and dendrites.The tip of a growing neurite (the growth cone) guidesthe rate and direction of neurite outgrowth by inter-
preting environmental cues, allowing neurons to makeprecise connections with target cells after they extendneurites over long distances. The specific connectionsthat are made determine the structure and, ultimately,the function of the neuron. After the nervous systemhas established initial neuronal connections, plasticityis limited, as demonstrated by the inability of centralnervous system axons to regenerate after injury. Twoimportant goals of neurobiology are to understand theprocess of neurite elongation during development andto assist neurites of adult neurons to regrow afterinjury.
Much research has focused on characterizing the
* Present address: Institute for Arctic Biology, P.O. Box757000, University of Alaska, Fairbanks, AK 99775-7000.
Correspondence to:J. R. Bamburg ([email protected]).
Contract grant sponsor: National Institutes of Health; contractgrant number: GM54004 and GM35126 (JRB).
Contract grant sponsor: Paralyzed Veterans of America SpinalCord Research Foundation; contract grant number: 1643 (TBK).© 2000 John Wiley & Sons, Inc.
352

environment surrounding developing neurons and un-derstanding how extracellular cues guide neurite out-growth. Many growth-promoting and growth-inhibit-ing molecules have been identified and characterized(reviewed by Goodman, 1996; Mueller, 1999). Dur-ing development, growth cones encounter solublemolecules, such as netrins; extracellular matrix mol-ecules, such as laminin and fibronectin; and cell mem-brane–bound molecules, such as cell adhesion mole-cules. Each of these extracellular molecules interactswith specific transmembrane proteins that activateintracellular signaling pathways to interpret and inte-grate these extracellular signals, ultimately leading tochanges in growth cone behavior.
Growth cone dynamics are largely dependent onthe assembly and reorganization of actin filaments. Inparticular, filopodia and lamellipodia, both actin-based structures, are important for neurite outgrowthand guidance (reviewed by Bentley and O’Conner,1994; Tanaka and Sabry, 1995). The small GTPases,cdc42, rac1, and rhoA, regulate filopodial, lamellipo-dial, and stress fiber formation in response to extra-cellular cues in nonneuronal cells (Nobes and Hall,1995; reviewed by Hall, 1998). In recent years, muchwork has focused on determining the role of theseproteins during neuronal development. Rac1 is in-volved in neurite extension (Luo et al., 1994; Kuhn etal., 1998), dendrite development (Luo et al., 1996a;Threadgill et al., 1997), and target innervation (Kauf-mann et al., 1998). Both rac1 and rho are involved ingrowth cone signaling in response to collapsing fac-tors (Jin and Strittmatter, 1997; Kuhn et al., 1999).
The importance of cdc42 during neuronal develop-ment has also been demonstrated. Expression ofCAcdc42 in the nervous system of developingDro-sophila embryos results in defects in neuronal posi-tion and a dramatic decrease in axonal and dendriticgrowth (Luo et al., 1994). However, whether this wasa result of abnormal neurite initiation, extension, orpathfinding was not determined. Cdc42 induces filo-podia in neuroblastoma N1E-115 cell growth cones(Kozma et al. 1997), is required for proper dendritedifferentiation in cortical neurons (Threadgill et al.,1997), and is involved in growth cone collapse in-duced by collapsin-1 (Kuhn et al., 1999).
Here, we characterize the role of cdc42 duringneurite outgrowth in dissociated primary chick spinalneurons. Specifically, we show that cdc42 activity isstimulatory to growth cones, resulting in largergrowth cones displaying more filopodia and fasterneurite outgrowth rates. We compare the effects tothose of soluble laminin and examine the pathwaysthese factors have in common.
MATERIALS AND METHODS
Reagents
Unless otherwise stated, all reagents were purchased fromSigma (St. Louis, MO).
Cell Culture
Spinal cords were dissected from 6- to 7-day-old chickembryos. Isolated spinal cords were trypsinized (4% tryp-sin, 0.2 mg/mL EDTA, 10 min, 37°C) and triturated. Dis-sociated cells were preplated on plastic for 1 h in Dulbec-co’s modified Eagle’s medium (DMEM) and 10% fetalbovine serum (FBS; Hyclone, Logan, UT). Nonadherentcells were resuspended in DMEM, 10% FBS, 12 nM flu-orodeoxyuridine, and 1% N3 supplement [10mg/mL bovineserum albumin (BSA), 100mg/mL transferrin, 10mg/mLinsulin, 32 ng/mL putrescine, 20 ng/mL triiodothyronine, 10ng/mL sodium selenite, 12.6 ng/mL progesterone, 200ng/mL corticosterone] at a density of 25,000 to 100,000cells per coverslip (223 22 mm; Carolina Biological Sup-ply, Burlington, NC). Coverslips were treated with poly-D-lysine (10mg/mL, borate buffer, pH 8.4). In some cases,coverslips were then coated with 2mg/cm2 fibronectin(Boehringer Mannheim, Indianapolis, IN) for 1 h at37°C.
Recombinant Adenoviruses
Recombinant adenoviruses capable of expressing cmyc-tagged constitutively active (CA) cdc42 (V12-G25K mu-tant, CAcdc42), dominant negative (DN) cdc42 (N17-G25Kmutant, DNcdc42), Enhanced Green Fluorescent Protein(EGFP), or LacZ were generated and purified as previouslydescribed (Kuhn et al., 1999). Expression of each gene wasunder the control of the adenovirus E1A promoter, which isrepressed in the 293 cells used to generate the replication-deficient virus. Virus titers were routinely between 13 108
and 13 109 plaque-forming units (pfu)/mL. Spinal neuronswere infected 16–18 h after plating with 200 to 300 pfu/neuron [multiplicity of infection (MOI)] in 400mL ofculture medium.
Polyacrylamide Gel Electrophoresis andWestern Blotting
Neuronal cell cultures were extracted with 2% sodium do-decyl sulfate (SDS), 10 mM Tris-HCl, pH 7.5, 10 mM NaF,5 mM dithiothreitol, and 2 mM EGTA. Soluble protein wasplaced in a boiling water bath for 5 min, precipitated withchloroform/methanol (Wessel and Flu¨gge, 1984), and resus-pended in SDS sample preparation buffer (Laemmli, 1970).Protein concentrations were determined using a dye-bindingfilter paper assay (Minamide and Bamburg, 1990). Separa-tion of proteins using SDS-PAGE (Laemmli, 1970) andelectroblotting (Towbin et al., 1979) onto polyvinylidenedifluoride (PVDF) membrane (Gelman Sciences, Ann Ar-
Cdc42 Stimulates Neurite Development 353

bor, MI) were performed as previously described (Bamburget al., 1991). Membranes were blocked with 5% nonfat drymilk in TBS (10 mM Tris-HCl, pH 8.0, 150 mM NaCl),washed with TBS/0.05% Tween 20, and incubated withprimary antibody (rabbit anti-cdc42, 1mg/mL; Santa CruzBiotechnology, Santa Cruz, CA) for 1 h. Membranes werethen washed with TBS/0.05% Tween 20 and incubated withsecondary antibody (alkaline phosphatase–conjugated goatanti-rabbit IgG, 1:30,000 dilution) for 1 h. CDP-Star(Tropix, Bedford, MA) was used for chemiluminescent de-tection. Chemiluminescent blots were digitized directly us-ing a Photometrics cooled CCD camera (12 bit, 4000-foldlinear range) and analyzed with Phoretix image analysissoftware. CDP-Star was removed by washing, and blotswere stained with nitroblue tetrazolium chloride and 5 bro-mo-4-chloro-3-indolylphosphatep-toluidine salt (LifeTechnologies, Gaithersburg, MD).
Immunocytochemistry
Cultured spinal neurons were fixed in 4% paraformalde-hyde, 0.1% gluteraldehyde, 100 mM sodium phosphate, pH7.4, 120 mM sucrose for 30 min at room temperature, andwashed three times with phosphate-buffered saline (PBS).For detection of cmyc-tagged GTPases, cells were perme-abilized with ice-cold methanol for 3 min, washed threetimes in TBS, and blocked with TBS supplemented with 2%goat serum and 1% BSA for 30 min. Coverslips wereincubated with a mouse monoclonal antibody against cmyc(9E10) (1 mg/mL; Santa Cruz Biotechnology) for 1 h,followed by a fluorescein-labeled goat-anti-mouse IgG (1:500; Molecular Probes, Eugene, OR) for 1 h. All antibodieswere diluted in TBS and 1% BSA. For detection of F-actin,cells were fixed as above, permeabilized with 0.1% TritonX-100 in PBS for 30 min, washed in PBS, and blocked withTBS containing 2% goat serum and 1% BSA. Cells wereincubated with Texas Red-phalloidin (1:50 in TBS, 1%BSA; Molecular Probes) for 30 min, then washed threetimes (5 min each) in TBS. In all cases, coverslips weremounted on slides using the Prolong Antifade Kit (Molec-ular Probes).
Image Analysis and Statistics
Images of neuronal growth cones were acquired using aNikon inverted microscope (Diaphot 200; Nikon Inc., Gar-den City, NY) equipped with a computer controlled CCDcamera (PXL Photometrics, Tucson, AZ) using a 603 1.40numerical aperture oil objective (Nikon). Images of neuriteswere obtained using a Nikon inverted microscope (Diaphot300) with a 203 objective (Nikon) and a Dage MTI tubevideo camera (VE-1000). Images were acquired and ana-lyzed using Metamorph imaging software (Universal Imag-ing Corporation, West Chester, PA).
One-way analysis of variance was used in comparingmultiple samples. Ap value, .05 was considered signifi-cant. Experimental data from CAcdc42- and DNcdc42-expressing neurons was considered significantly different
from controls only if it was statistically different (p , .05)than both uninfected and virus-infected controls.
RESULTS
Expression of Cdc42 Mutants in PrimarySpinal Neurons
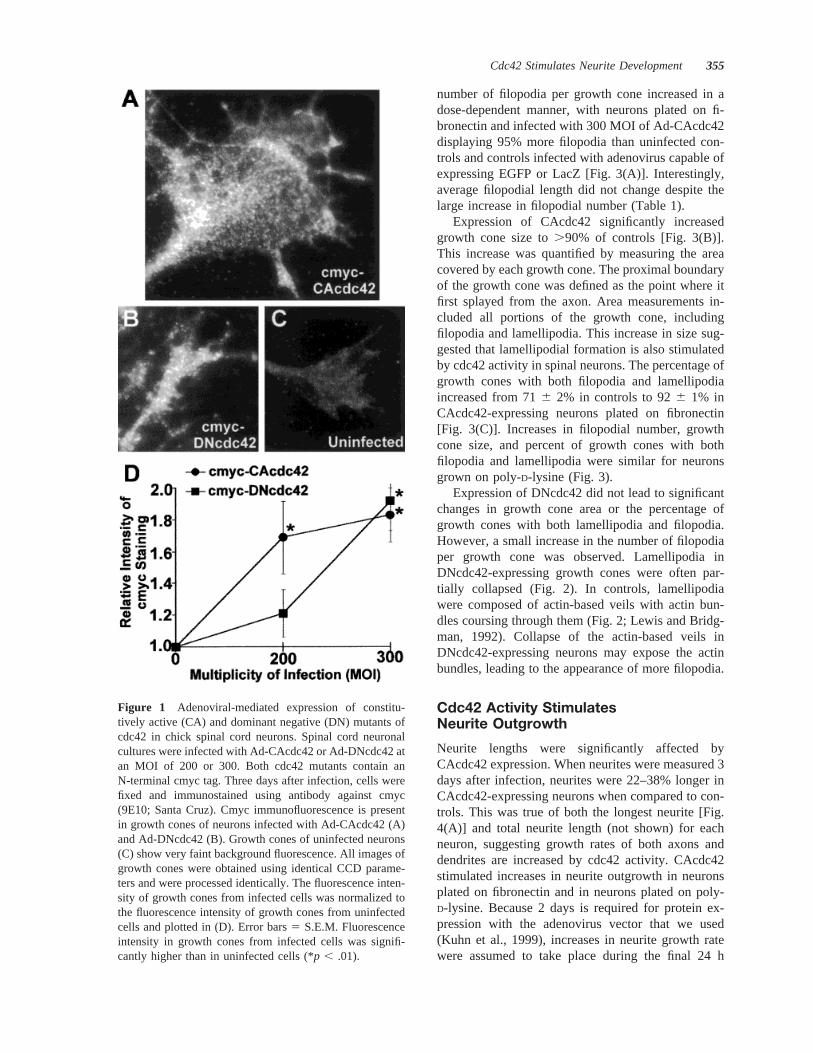
To assess the role of cdc42 during neurite develop-ment, CA and DN mutants of cdc42 were introducedinto dissociated primary chick spinal neurons usingrecombinant, replication-deficient adenovirus. Threedays after infection with adenovirus-carrying, cmyc-tagged CAcdc42 (Ad-CAcdc42) or DNcdc42 (Ad-DNcdc42), cdc42 proteins were detected in spinalneuron growth cones using indirect immunofluores-cence against cmyc. Growth cones of neurons in-fected with 300 MOI of Ad-CAcdc42 or Ad-DNcdc42displayed 1.83- and 1.93-fold higher levels of totalfluorescence per square micrometer when comparedto uninfected controls (Fig. 1). Immunofluorescencewas detected at similar levels in cell bodies of infectedneurons. More than 85% of the neurons in infectedcultures were cmyc-positive. In growth cones, cmycstaining was punctate in appearance and was notconsistently localized to any particular growth coneregion. Staining was found in lamellipodia, filopodia,the central region, and in the neurite shaft. Levels ofmutant cdc42 expression were dose-dependent, withlower MOIs of adenovirus leading to lower levels ofcdc42 expression.
To determine if adenoviral infection and CAcdc42or DNcdc42 expression affect cell survival, the num-ber of neurite-bearing neurons in virus-infected (200and 300 MOI) and control dishes were counted in 35random fields 3 days after infection. The density ofneurite-bearing neurons was not significantly differentin uninfected cultures (2.54 neurons/field) and cul-tures infected with either Ad-CAcdc42 (2.66 neurons/field) or Ad-DNcdc42 (2.66 neurons/field). Thus, in-fection and expression of cdc42 proteins did not affectneuronal survival when cultures were incubated for 3days after infection with 200 or 300 MOI. However,neuronal survival decreased when cultures were incu-bated for longer periods of time after infection andwhen MOIs higher than 300 were used, suggestinghigh levels of CAcdc42 or DNcdc42 are toxic to cells.
Cdc42 Activity is Stimulatory toNeuronal Growth Cones
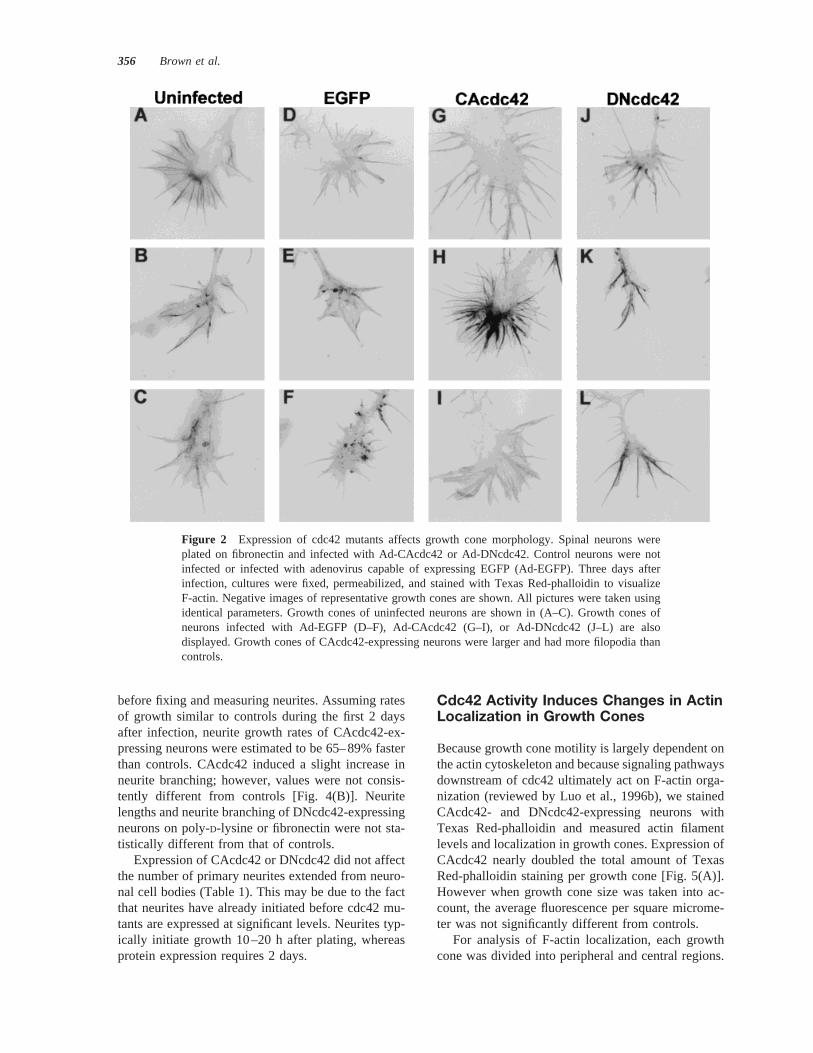
Expression of CAcdc42 induced significant changesin neuronal growth cone morphology (Figs. 2, 3). The
354 Brown et al.
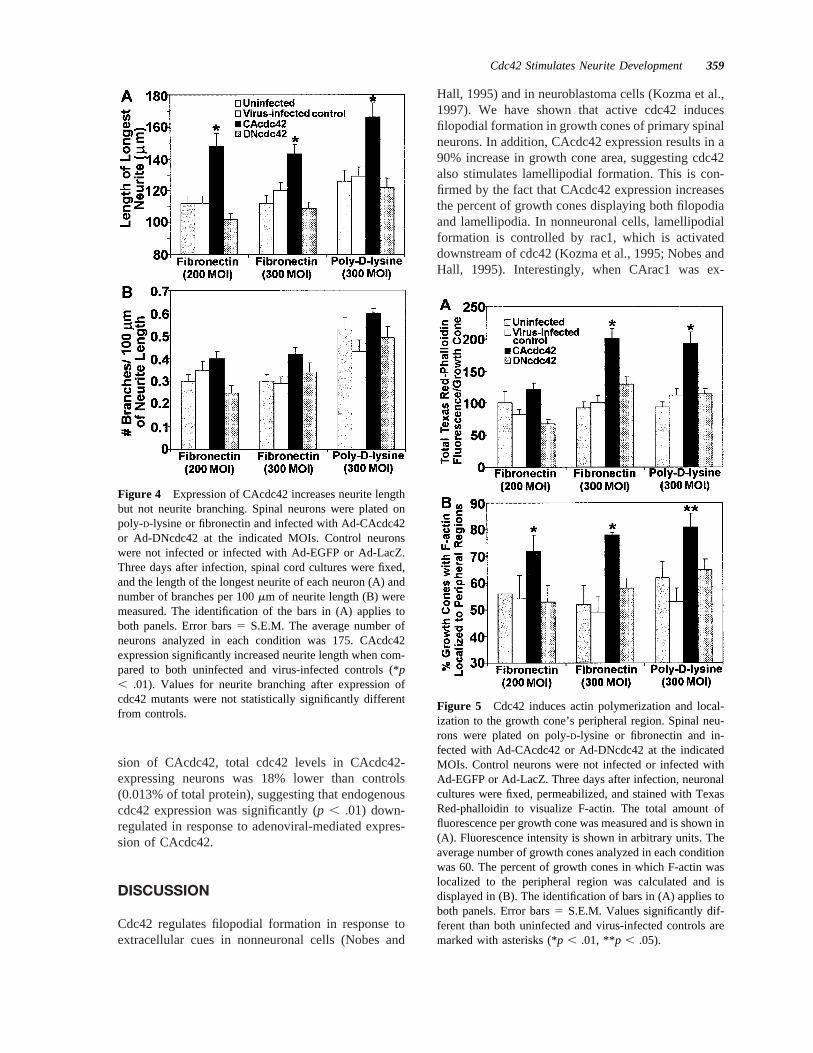
number of filopodia per growth cone increased in adose-dependent manner, with neurons plated on fi-bronectin and infected with 300 MOI of Ad-CAcdc42displaying 95% more filopodia than uninfected con-trols and controls infected with adenovirus capable ofexpressing EGFP or LacZ [Fig. 3(A)]. Interestingly,average filopodial length did not change despite thelarge increase in filopodial number (Table 1).
Expression of CAcdc42 significantly increasedgrowth cone size to.90% of controls [Fig. 3(B)].This increase was quantified by measuring the areacovered by each growth cone. The proximal boundaryof the growth cone was defined as the point where itfirst splayed from the axon. Area measurements in-cluded all portions of the growth cone, includingfilopodia and lamellipodia. This increase in size sug-gested that lamellipodial formation is also stimulatedby cdc42 activity in spinal neurons. The percentage ofgrowth cones with both filopodia and lamellipodiaincreased from 716 2% in controls to 926 1% inCAcdc42-expressing neurons plated on fibronectin[Fig. 3(C)]. Increases in filopodial number, growthcone size, and percent of growth cones with bothfilopodia and lamellipodia were similar for neuronsgrown on poly-D-lysine (Fig. 3).
Expression of DNcdc42 did not lead to significantchanges in growth cone area or the percentage ofgrowth cones with both lamellipodia and filopodia.However, a small increase in the number of filopodiaper growth cone was observed. Lamellipodia inDNcdc42-expressing growth cones were often par-tially collapsed (Fig. 2). In controls, lamellipodiawere composed of actin-based veils with actin bun-dles coursing through them (Fig. 2; Lewis and Bridg-man, 1992). Collapse of the actin-based veils inDNcdc42-expressing neurons may expose the actinbundles, leading to the appearance of more filopodia.
Cdc42 Activity StimulatesNeurite Outgrowth
Neurite lengths were significantly affected byCAcdc42 expression. When neurites were measured 3days after infection, neurites were 22–38% longer inCAcdc42-expressing neurons when compared to con-trols. This was true of both the longest neurite [Fig.4(A)] and total neurite length (not shown) for eachneuron, suggesting growth rates of both axons anddendrites are increased by cdc42 activity. CAcdc42stimulated increases in neurite outgrowth in neuronsplated on fibronectin and in neurons plated on poly-D-lysine. Because 2 days is required for protein ex-pression with the adenovirus vector that we used(Kuhn et al., 1999), increases in neurite growth ratewere assumed to take place during the final 24 h
Figure 1 Adenoviral-mediated expression of constitu-tively active (CA) and dominant negative (DN) mutants ofcdc42 in chick spinal cord neurons. Spinal cord neuronalcultures were infected with Ad-CAcdc42 or Ad-DNcdc42 atan MOI of 200 or 300. Both cdc42 mutants contain anN-terminal cmyc tag. Three days after infection, cells werefixed and immunostained using antibody against cmyc(9E10; Santa Cruz). Cmyc immunofluorescence is presentin growth cones of neurons infected with Ad-CAcdc42 (A)and Ad-DNcdc42 (B). Growth cones of uninfected neurons(C) show very faint background fluorescence. All images ofgrowth cones were obtained using identical CCD parame-ters and were processed identically. The fluorescence inten-sity of growth cones from infected cells was normalized tothe fluorescence intensity of growth cones from uninfectedcells and plotted in (D). Error bars5 S.E.M. Fluorescenceintensity in growth cones from infected cells was signifi-cantly higher than in uninfected cells (*p , .01).
Cdc42 Stimulates Neurite Development 355
before fixing and measuring neurites. Assuming ratesof growth similar to controls during the first 2 daysafter infection, neurite growth rates of CAcdc42-ex-pressing neurons were estimated to be 65–89% fasterthan controls. CAcdc42 induced a slight increase inneurite branching; however, values were not consis-tently different from controls [Fig. 4(B)]. Neuritelengths and neurite branching of DNcdc42-expressingneurons on poly-D-lysine or fibronectin were not sta-tistically different from that of controls.
Expression of CAcdc42 or DNcdc42 did not affectthe number of primary neurites extended from neuro-nal cell bodies (Table 1). This may be due to the factthat neurites have already initiated before cdc42 mu-tants are expressed at significant levels. Neurites typ-ically initiate growth 10–20 h after plating, whereasprotein expression requires 2 days.
Cdc42 Activity Induces Changes in ActinLocalization in Growth Cones
Because growth cone motility is largely dependent onthe actin cytoskeleton and because signaling pathwaysdownstream of cdc42 ultimately act on F-actin orga-nization (reviewed by Luo et al., 1996b), we stainedCAcdc42- and DNcdc42-expressing neurons withTexas Red-phalloidin and measured actin filamentlevels and localization in growth cones. Expression ofCAcdc42 nearly doubled the total amount of TexasRed-phalloidin staining per growth cone [Fig. 5(A)].However when growth cone size was taken into ac-count, the average fluorescence per square microme-ter was not significantly different from controls.
For analysis of F-actin localization, each growthcone was divided into peripheral and central regions.
Figure 2 Expression of cdc42 mutants affects growth cone morphology. Spinal neurons wereplated on fibronectin and infected with Ad-CAcdc42 or Ad-DNcdc42. Control neurons were notinfected or infected with adenovirus capable of expressing EGFP (Ad-EGFP). Three days afterinfection, cultures were fixed, permeabilized, and stained with Texas Red-phalloidin to visualizeF-actin. Negative images of representative growth cones are shown. All pictures were taken usingidentical parameters. Growth cones of uninfected neurons are shown in (A–C). Growth cones ofneurons infected with Ad-EGFP (D–F), Ad-CAcdc42 (G–I), or Ad-DNcdc42 (J–L) are alsodisplayed. Growth cones of CAcdc42-expressing neurons were larger and had more filopodia thancontrols.
356 Brown et al.
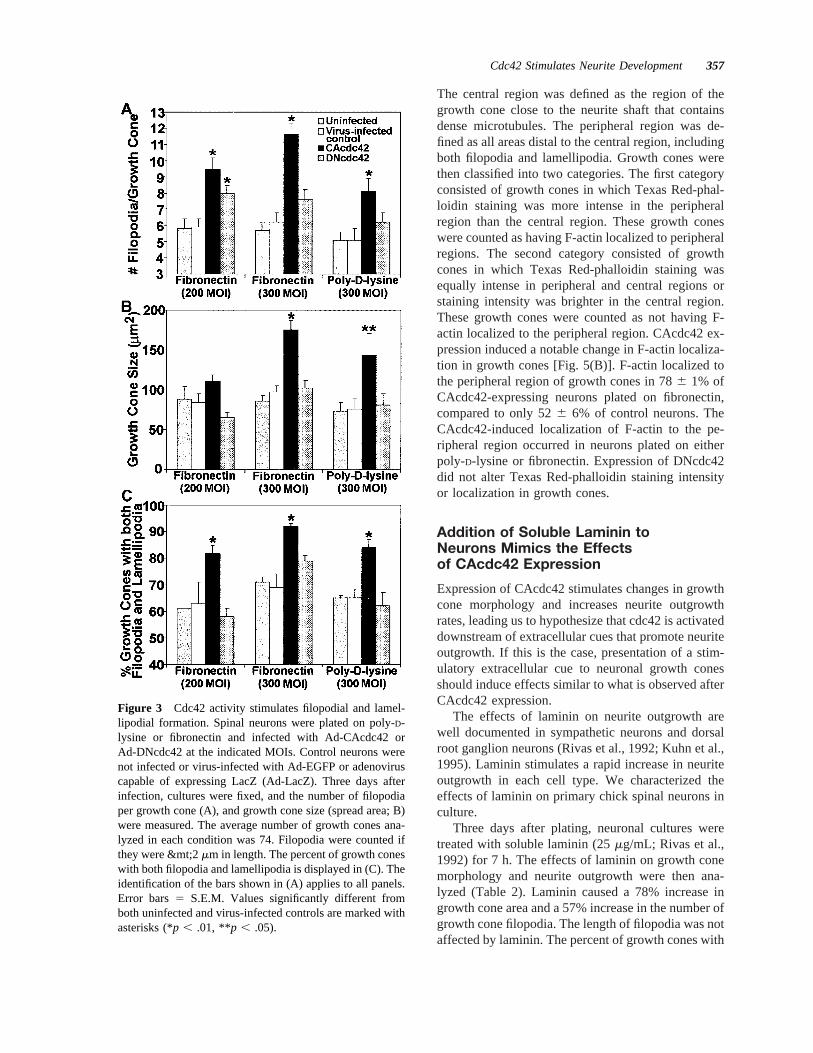
The central region was defined as the region of thegrowth cone close to the neurite shaft that containsdense microtubules. The peripheral region was de-fined as all areas distal to the central region, includingboth filopodia and lamellipodia. Growth cones werethen classified into two categories. The first categoryconsisted of growth cones in which Texas Red-phal-loidin staining was more intense in the peripheralregion than the central region. These growth coneswere counted as having F-actin localized to peripheralregions. The second category consisted of growthcones in which Texas Red-phalloidin staining wasequally intense in peripheral and central regions orstaining intensity was brighter in the central region.These growth cones were counted as not having F-actin localized to the peripheral region. CAcdc42 ex-pression induced a notable change in F-actin localiza-tion in growth cones [Fig. 5(B)]. F-actin localized tothe peripheral region of growth cones in 786 1% ofCAcdc42-expressing neurons plated on fibronectin,compared to only 526 6% of control neurons. TheCAcdc42-induced localization of F-actin to the pe-ripheral region occurred in neurons plated on eitherpoly-D-lysine or fibronectin. Expression of DNcdc42did not alter Texas Red-phalloidin staining intensityor localization in growth cones.
Addition of Soluble Laminin toNeurons Mimics the Effectsof CAcdc42 Expression
Expression of CAcdc42 stimulates changes in growthcone morphology and increases neurite outgrowthrates, leading us to hypothesize that cdc42 is activateddownstream of extracellular cues that promote neuriteoutgrowth. If this is the case, presentation of a stim-ulatory extracellular cue to neuronal growth conesshould induce effects similar to what is observed afterCAcdc42 expression.
The effects of laminin on neurite outgrowth arewell documented in sympathetic neurons and dorsalroot ganglion neurons (Rivas et al., 1992; Kuhn et al.,1995). Laminin stimulates a rapid increase in neuriteoutgrowth in each cell type. We characterized theeffects of laminin on primary chick spinal neurons inculture.
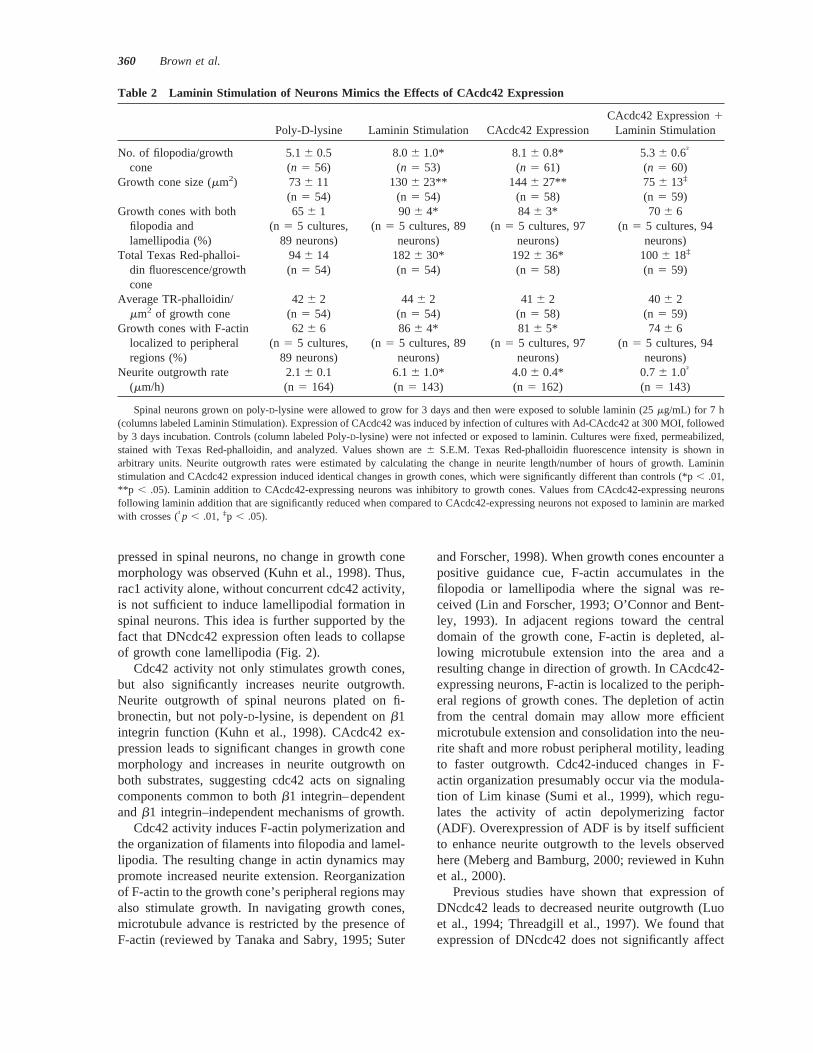
Three days after plating, neuronal cultures weretreated with soluble laminin (25mg/mL; Rivas et al.,1992) for 7 h. The effects of laminin on growth conemorphology and neurite outgrowth were then ana-lyzed (Table 2). Laminin caused a 78% increase ingrowth cone area and a 57% increase in the number ofgrowth cone filopodia. The length of filopodia was notaffected by laminin. The percent of growth cones with
Figure 3 Cdc42 activity stimulates filopodial and lamel-lipodial formation. Spinal neurons were plated on poly-D-lysine or fibronectin and infected with Ad-CAcdc42 orAd-DNcdc42 at the indicated MOIs. Control neurons werenot infected or virus-infected with Ad-EGFP or adenoviruscapable of expressing LacZ (Ad-LacZ). Three days afterinfection, cultures were fixed, and the number of filopodiaper growth cone (A), and growth cone size (spread area; B)were measured. The average number of growth cones ana-lyzed in each condition was 74. Filopodia were counted ifthey were &mt;2mm in length. The percent of growth coneswith both filopodia and lamellipodia is displayed in (C). Theidentification of the bars shown in (A) applies to all panels.Error bars5 S.E.M. Values significantly different fromboth uninfected and virus-infected controls are marked withasterisks (*p , .01, **p , .05).
Cdc42 Stimulates Neurite Development 357
both filopodia and lamellipodia increased from 626 6% in controls to 866 4% in laminin-treatedcultures. Neurites grew nearly three times faster afterthe addition of laminin. Laminin treatment did notaffect the number of primary neurites per neuron orthe degree of neurite branching. The total amount ofTexas Red-phalloidin staining per growth cone nearlydoubled; however, the amount of Texas Red-phalloi-din staining per square micrometer of growth conewas not different from controls. Laminin induced anincrease from 626 6% to 866 4% in the percent ofgrowth cones with F-actin localized to peripheral re-gions. The effects of laminin very closely resembledthe effects of CAcdc42 expression in every measuredaspect (Table 2).
DNcdc42 Blocks the Effects of SolubleLaminin on Growth Cones
The fact that laminin and CAcdc42 induce strikinglysimilar changes in growth cone morphology and neu-rite outgrowth suggests that cdc42 and laminin sharea common signaling pathway. If this is the case,expression of DNcdc42 should block the effects oflaminin on neurons. To test this hypothesis, we treatedDNcdc42-expressing neurons (3 days after infectionwith Ad-DNcdc42) with soluble laminin. Expressionof DNcdc42 completely blocked all effects of solublelaminin on growth cones, including the effects ongrowth cone size and filopodial number, percent ofgrowth cones with lamellipodia and filopodia, total
F-actin levels, F-actin localization in growth cones,and rate of neurite outgrowth (Fig. 6).
Laminin is Inhibitory to CAcdc42-Expressing Neurons
We also investigated the response of CAcdc42-ex-pressing neurons to laminin stimulation. Surprisingly,addition of laminin to CAcdc42-expressing neuronshad a negative effect on growth cones. Before laminintreatment, growth cones of CAcdc42-expressing neu-rons were larger and had more filopodia than controls(Fig. 3). These effects were attenuated by the additionof laminin (Table 2), which caused a significant re-duction in growth cone size and number of filopodia.In addition, neurite outgrowth slowed considerably.Thus, laminin is inhibitory to growth cones ofCAcdc42-expressing neurons.
CAcdc42 Downregulates EndogenousCdc42 Expression
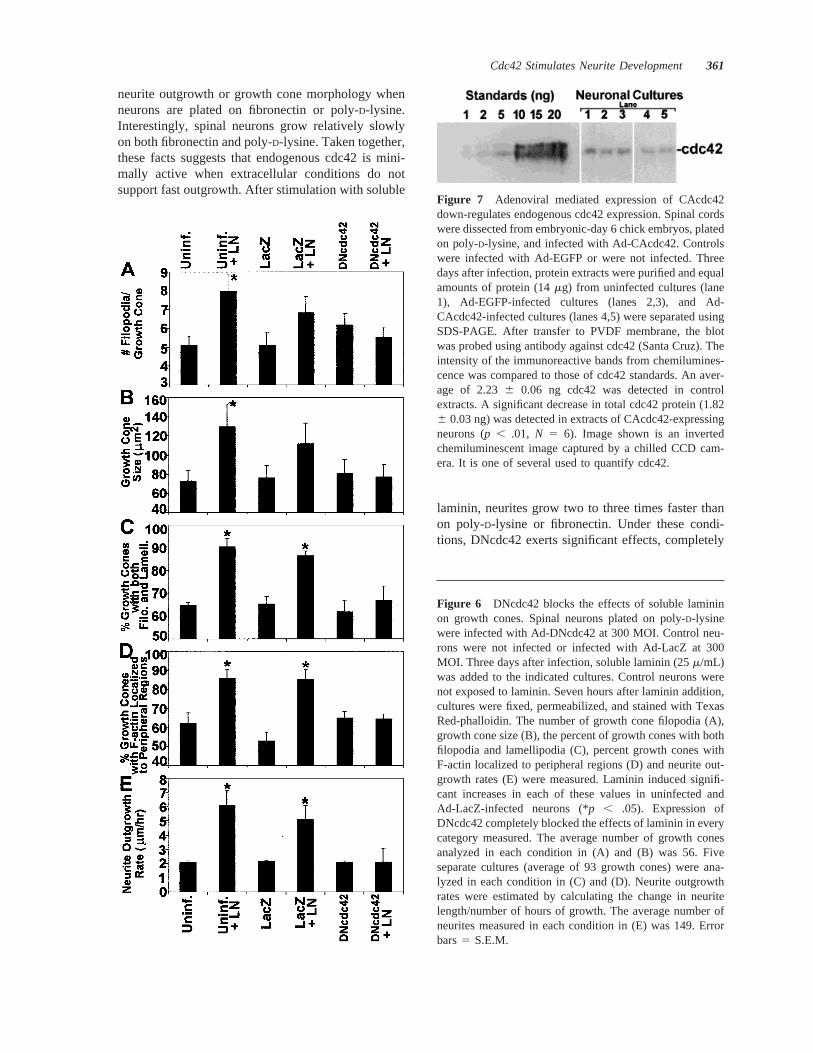
We examined the effects of CAcdc42 expression onendogenous cdc42 levels by extracting protein frominfected and uninfected primary chick spinal neuronalcultures. Western blots indicate that cdc42 is presentin uninfected and Ad-EGFP–infected spinal neuronsas 0.016% of total protein (Fig. 7), consistent withother studies that have demonstrated the presence ofcdc42 in primary neurons (Olenik et al., 1997;Threadgill et al., 1997). Interestingly, despite expres-
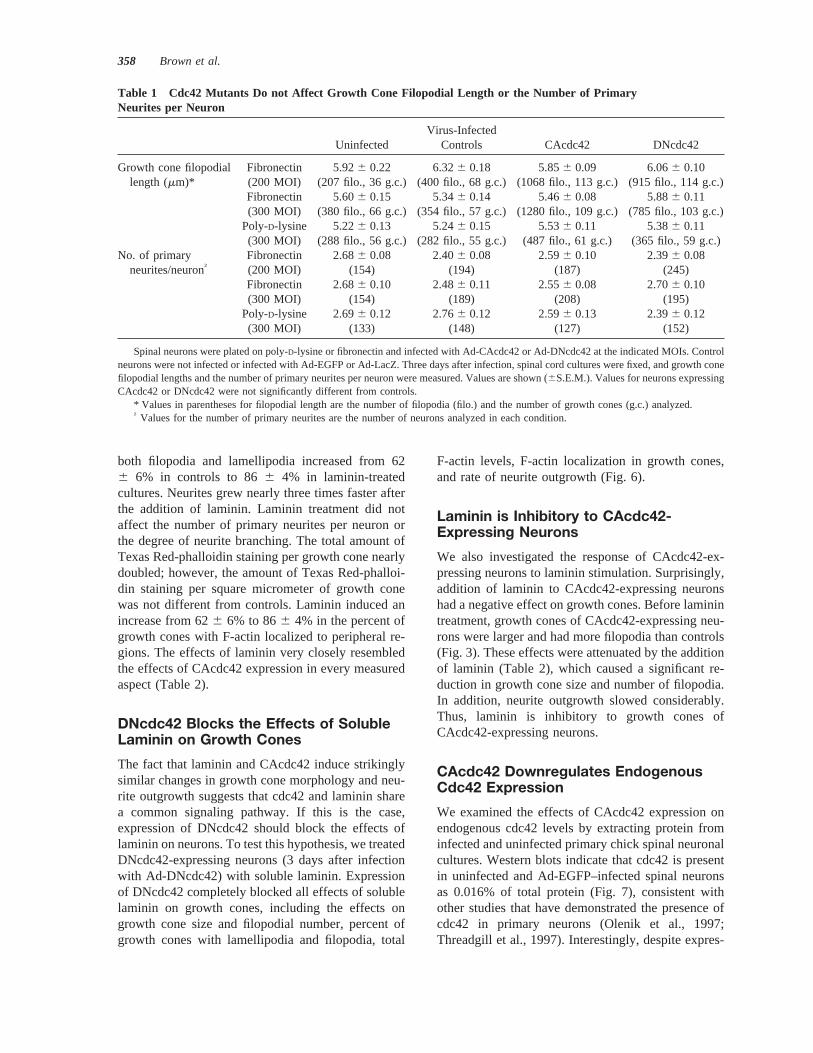
Table 1 Cdc42 Mutants Do not Affect Growth Cone Filopodial Length or the Number of PrimaryNeurites per Neuron
UninfectedVirus-Infected
Controls CAcdc42 DNcdc42
Growth cone filopodial Fibronectin 5.926 0.22 6.326 0.18 5.856 0.09 6.066 0.10length (mm)* (200 MOI) (207 filo., 36 g.c.) (400 filo., 68 g.c.) (1068 filo., 113 g.c.) (915 filo., 114 g.c.)
Fibronectin 5.606 0.15 5.346 0.14 5.466 0.08 5.886 0.11(300 MOI) (380 filo., 66 g.c.) (354 filo., 57 g.c.) (1280 filo., 109 g.c.) (785 filo., 103 g.c.)
Poly-D-lysine 5.226 0.13 5.246 0.15 5.536 0.11 5.386 0.11(300 MOI) (288 filo., 56 g.c.) (282 filo., 55 g.c.) (487 filo., 61 g.c.) (365 filo., 59 g.c.)
No. of primary Fibronectin 2.686 0.08 2.406 0.08 2.596 0.10 2.396 0.08neurites/neuron† (200 MOI) (154) (194) (187) (245)
Fibronectin 2.686 0.10 2.486 0.11 2.556 0.08 2.706 0.10(300 MOI) (154) (189) (208) (195)
Poly-D-lysine 2.696 0.12 2.766 0.12 2.596 0.13 2.396 0.12(300 MOI) (133) (148) (127) (152)
Spinal neurons were plated on poly-D-lysine or fibronectin and infected with Ad-CAcdc42 or Ad-DNcdc42 at the indicated MOIs. Controlneurons were not infected or infected with Ad-EGFP or Ad-LacZ. Three days after infection, spinal cord cultures were fixed, and growth conefilopodial lengths and the number of primary neurites per neuron were measured. Values are shown (6S.E.M.). Values for neurons expressingCAcdc42 or DNcdc42 were not significantly different from controls.
* Values in parentheses for filopodial length are the number of filopodia (filo.) and the number of growth cones (g.c.) analyzed.† Values for the number of primary neurites are the number of neurons analyzed in each condition.
358 Brown et al.
sion of CAcdc42, total cdc42 levels in CAcdc42-expressing neurons was 18% lower than controls(0.013% of total protein), suggesting that endogenouscdc42 expression was significantly (p , .01) down-regulated in response to adenoviral-mediated expres-sion of CAcdc42.
DISCUSSION
Cdc42 regulates filopodial formation in response toextracellular cues in nonneuronal cells (Nobes and
Hall, 1995) and in neuroblastoma cells (Kozma et al.,1997). We have shown that active cdc42 inducesfilopodial formation in growth cones of primary spinalneurons. In addition, CAcdc42 expression results in a90% increase in growth cone area, suggesting cdc42also stimulates lamellipodial formation. This is con-firmed by the fact that CAcdc42 expression increasesthe percent of growth cones displaying both filopodiaand lamellipodia. In nonneuronal cells, lamellipodialformation is controlled by rac1, which is activateddownstream of cdc42 (Kozma et al., 1995; Nobes andHall, 1995). Interestingly, when CArac1 was ex-
Figure 5 Cdc42 induces actin polymerization and local-ization to the growth cone’s peripheral region. Spinal neu-rons were plated on poly-D-lysine or fibronectin and in-fected with Ad-CAcdc42 or Ad-DNcdc42 at the indicatedMOIs. Control neurons were not infected or infected withAd-EGFP or Ad-LacZ. Three days after infection, neuronalcultures were fixed, permeabilized, and stained with TexasRed-phalloidin to visualize F-actin. The total amount offluorescence per growth cone was measured and is shown in(A). Fluorescence intensity is shown in arbitrary units. Theaverage number of growth cones analyzed in each conditionwas 60. The percent of growth cones in which F-actin waslocalized to the peripheral region was calculated and isdisplayed in (B). The identification of bars in (A) applies toboth panels. Error bars5 S.E.M. Values significantly dif-ferent than both uninfected and virus-infected controls aremarked with asterisks (*p , .01, **p , .05).
Figure 4 Expression of CAcdc42 increases neurite lengthbut not neurite branching. Spinal neurons were plated onpoly-D-lysine or fibronectin and infected with Ad-CAcdc42or Ad-DNcdc42 at the indicated MOIs. Control neuronswere not infected or infected with Ad-EGFP or Ad-LacZ.Three days after infection, spinal cord cultures were fixed,and the length of the longest neurite of each neuron (A) andnumber of branches per 100mm of neurite length (B) weremeasured. The identification of the bars in (A) applies toboth panels. Error bars5 S.E.M. The average number ofneurons analyzed in each condition was 175. CAcdc42expression significantly increased neurite length when com-pared to both uninfected and virus-infected controls (*p, .01). Values for neurite branching after expression ofcdc42 mutants were not statistically significantly differentfrom controls.
Cdc42 Stimulates Neurite Development 359
pressed in spinal neurons, no change in growth conemorphology was observed (Kuhn et al., 1998). Thus,rac1 activity alone, without concurrent cdc42 activity,is not sufficient to induce lamellipodial formation inspinal neurons. This idea is further supported by thefact that DNcdc42 expression often leads to collapseof growth cone lamellipodia (Fig. 2).
Cdc42 activity not only stimulates growth cones,but also significantly increases neurite outgrowth.Neurite outgrowth of spinal neurons plated on fi-bronectin, but not poly-D-lysine, is dependent onb1integrin function (Kuhn et al., 1998). CAcdc42 ex-pression leads to significant changes in growth conemorphology and increases in neurite outgrowth onboth substrates, suggesting cdc42 acts on signalingcomponents common to bothb1 integrin–dependentandb1 integrin–independent mechanisms of growth.
Cdc42 activity induces F-actin polymerization andthe organization of filaments into filopodia and lamel-lipodia. The resulting change in actin dynamics maypromote increased neurite extension. Reorganizationof F-actin to the growth cone’s peripheral regions mayalso stimulate growth. In navigating growth cones,microtubule advance is restricted by the presence ofF-actin (reviewed by Tanaka and Sabry, 1995; Suter
and Forscher, 1998). When growth cones encounter apositive guidance cue, F-actin accumulates in thefilopodia or lamellipodia where the signal was re-ceived (Lin and Forscher, 1993; O’Connor and Bent-ley, 1993). In adjacent regions toward the centraldomain of the growth cone, F-actin is depleted, al-lowing microtubule extension into the area and aresulting change in direction of growth. In CAcdc42-expressing neurons, F-actin is localized to the periph-eral regions of growth cones. The depletion of actinfrom the central domain may allow more efficientmicrotubule extension and consolidation into the neu-rite shaft and more robust peripheral motility, leadingto faster outgrowth. Cdc42-induced changes in F-actin organization presumably occur via the modula-tion of Lim kinase (Sumi et al., 1999), which regu-lates the activity of actin depolymerizing factor(ADF). Overexpression of ADF is by itself sufficientto enhance neurite outgrowth to the levels observedhere (Meberg and Bamburg, 2000; reviewed in Kuhnet al., 2000).
Previous studies have shown that expression ofDNcdc42 leads to decreased neurite outgrowth (Luoet al., 1994; Threadgill et al., 1997). We found thatexpression of DNcdc42 does not significantly affect
Table 2 Laminin Stimulation of Neurons Mimics the Effects of CAcdc42 Expression
Poly-D-lysine Laminin Stimulation CAcdc42 ExpressionCAcdc42 Expression1
Laminin Stimulation
No. of filopodia/growth 5.16 0.5 8.06 1.0* 8.16 0.8* 5.36 0.6†
cone (n 5 56) (n 5 53) (n 5 61) (n 5 60)Growth cone size (mm2) 736 11 1306 23** 144 6 27** 75 6 13‡
(n 5 54) (n 5 54) (n 5 58) (n 5 59)Growth cones with both 656 1 906 4* 84 6 3* 70 6 6
filopodia andlamellipodia (%)
(n 5 5 cultures,89 neurons)
(n 5 5 cultures, 89neurons)
(n 5 5 cultures, 97neurons)
(n 5 5 cultures, 94neurons)
Total Texas Red-phalloi- 946 14 1826 30* 1926 36* 1006 18‡
din fluorescence/growthcone
(n 5 54) (n 5 54) (n 5 58) (n 5 59)
Average TR-phalloidin/ 426 2 446 2 416 2 406 2mm2 of growth cone (n 5 54) (n 5 54) (n 5 58) (n 5 59)
Growth cones with F-actin 626 6 866 4* 81 6 5* 74 6 6localized to peripheralregions (%)
(n 5 5 cultures,89 neurons)
(n 5 5 cultures, 89neurons)
(n 5 5 cultures, 97neurons)
(n 5 5 cultures, 94neurons)
Neurite outgrowth rate 2.16 0.1 6.16 1.0* 4.06 0.4* 0.76 1.0†
(mm/h) (n 5 164) (n5 143) (n5 162) (n5 143)
Spinal neurons grown on poly-D-lysine were allowed to grow for 3 days and then were exposed to soluble laminin (25mg/mL) for 7 h(columns labeled Laminin Stimulation). Expression of CAcdc42 was induced by infection of cultures with Ad-CAcdc42 at 300 MOI, followedby 3 days incubation. Controls (column labeled Poly-D-lysine) were not infected or exposed to laminin. Cultures were fixed, permeabilized,stained with Texas Red-phalloidin, and analyzed. Values shown are6 S.E.M. Texas Red-phalloidin fluorescence intensity is shown inarbitrary units. Neurite outgrowth rates were estimated by calculating the change in neurite length/number of hours of growth. Lamininstimulation and CAcdc42 expression induced identical changes in growth cones, which were significantly different than controls (*p, .01,**p , .05). Laminin addition to CAcdc42-expressing neurons was inhibitory to growth cones. Values from CAcdc42-expressing neuronsfollowing laminin addition that are significantly reduced when compared to CAcdc42-expressing neurons not exposed to laminin are markedwith crosses (†p , .01, ‡p , .05).
360 Brown et al.
neurite outgrowth or growth cone morphology whenneurons are plated on fibronectin or poly-D-lysine.Interestingly, spinal neurons grow relatively slowlyon both fibronectin and poly-D-lysine. Taken together,these facts suggests that endogenous cdc42 is mini-mally active when extracellular conditions do notsupport fast outgrowth. After stimulation with soluble
laminin, neurites grow two to three times faster thanon poly-D-lysine or fibronectin. Under these condi-tions, DNcdc42 exerts significant effects, completely
Figure 7 Adenoviral mediated expression of CAcdc42down-regulates endogenous cdc42 expression. Spinal cordswere dissected from embryonic-day 6 chick embryos, platedon poly-D-lysine, and infected with Ad-CAcdc42. Controlswere infected with Ad-EGFP or were not infected. Threedays after infection, protein extracts were purified and equalamounts of protein (14mg) from uninfected cultures (lane1), Ad-EGFP-infected cultures (lanes 2,3), and Ad-CAcdc42-infected cultures (lanes 4,5) were separated usingSDS-PAGE. After transfer to PVDF membrane, the blotwas probed using antibody against cdc42 (Santa Cruz). Theintensity of the immunoreactive bands from chemilumines-cence was compared to those of cdc42 standards. An aver-age of 2.236 0.06 ng cdc42 was detected in controlextracts. A significant decrease in total cdc42 protein (1.826 0.03 ng) was detected in extracts of CAcdc42-expressingneurons (p , .01, N 5 6). Image shown is an invertedchemiluminescent image captured by a chilled CCD cam-era. It is one of several used to quantify cdc42.
Figure 6 DNcdc42 blocks the effects of soluble lamininon growth cones. Spinal neurons plated on poly-D-lysinewere infected with Ad-DNcdc42 at 300 MOI. Control neu-rons were not infected or infected with Ad-LacZ at 300MOI. Three days after infection, soluble laminin (25m/mL)was added to the indicated cultures. Control neurons werenot exposed to laminin. Seven hours after laminin addition,cultures were fixed, permeabilized, and stained with TexasRed-phalloidin. The number of growth cone filopodia (A),growth cone size (B), the percent of growth cones with bothfilopodia and lamellipodia (C), percent growth cones withF-actin localized to peripheral regions (D) and neurite out-growth rates (E) were measured. Laminin induced signifi-cant increases in each of these values in uninfected andAd-LacZ-infected neurons (*p , .05). Expression ofDNcdc42 completely blocked the effects of laminin in everycategory measured. The average number of growth conesanalyzed in each condition in (A) and (B) was 56. Fiveseparate cultures (average of 93 growth cones) were ana-lyzed in each condition in (C) and (D). Neurite outgrowthrates were estimated by calculating the change in neuritelength/number of hours of growth. The average number ofneurites measured in each condition in (E) was 149. Errorbars5 S.E.M.
Cdc42 Stimulates Neurite Development 361

blocking laminin-induced effects on growth conesand neurites, suggesting that endogenous cdc42 isactive when neurons are in a growth-promotingenvironment. It is possible that higher levels ofDNcdc42 expression may be needed to detect ef-fects in neurons plated on poly-D-lysine or fi-bronectin. However, the fact that DNcdc42 expres-sion completely blocks laminin-induced effectssuggests that levels are sufficiently high to blockendogenous cdc42 function. Moreover, incubationof neurons for longer than 3 days after infectionwith Ad-DNcdc42 or infection with higher MOIs ofAd-DNcdc42 leads to neuronal death. Therefore,higher levels of DNcdc42 cannot be obtained with-out compromising cell survival.
It is striking that CAcdc42 expression and lamininstimulation cause virtually identical phenotypes inevery measured aspect of growth cone morphologyand neurite outgrowth (Table 2). Significantly, ex-pression of DNcdc42 blocks all laminin-induced ef-fects on neurons. Taken together, these resultsstrongly suggest that cdc42 is a component of a lami-nin-activated signaling pathway. Given that laminin’seffects on growth cones and neurite outgrowth areblocked by DNcdc42, a most surprising finding wasthat addition of laminin to neurons already expressingCAcdc42 has a marked inhibitory effect on growthcones and neurite outgrowth. Although CAcdc42-ex-pressing neurons have larger growth cones with morefilopodia than controls, acute exposure of CAcdc42-expressing neurons to laminin induces a significantdecrease in growth cone size, number of filopodia,neurite growth rate, and percent of growth cones withboth filopodia and lamellipodia. Several alternativescould explain the negative effect of laminin inCAcdc42-expressing motor neurons. First, chronicexpression of CAcdc42 could induce changes in geneexpression. Indeed, the total amount of cdc42 proteinin CAcdc42-expressing neurons is 18% less thancdc42 levels in uninfected neurons when measured byWestern blot 3 days after infection (Fig. 7). Thus, oneneuronal response to adenoviral-mediated expressionof CAcdc42 is down-regulation of endogenous cdc42expression. With less endogenous cdc42 present inthe growth cone, exposure to laminin may activatealternative pathways that are inhibitory to growthcones. For example, the guanine nucleotide exchangefactors Dbl and Ost activate both cdc42 and rho andare predominantly found in neural tissue (Horii et al.,1994; Colucci-D’Amato et al., 1995; Lorenzi et al.,1999). If laminin normally activates cdc42 via Dbl orOst, a relative lack of endogenous cdc42 in growthcones of CAcdc42-expressing neurons would allowDbl and Ost to bind to and activate rho to a greater
degree than normal. In fact, rho activation leads togrowth cone collapse and neurite retraction (Jalink etal., 1994; Tigyi et al., 1996; Kozma et al., 1997).
An alternate explanation for the inhibitory effect oflaminin on outgrowth from CAcdc42-expressing neu-rons arises from recent studies on laminin’s effect oncyclic nucleotide levels. Poo and colleagues havedemonstrated that changes in the levels of cyclicnucleotide can alter growth cone responses upon in-teraction with attractive or repulsive guidance cues(Song and Poo, 1999; Song et al., 1998). Growthcones advancing on poly-D-lysine are attracted bynetrin and exhibit rather high levels of cAMP. How-ever, when advancing on laminin, growth cones arerepelled by netrin coinciding with low levels of cAMP(Hopker et al., 1999). Acute exposure of growth conesto laminin or chronic exposure to laminin as a sub-strate results in a drastic drop in cAMP levels ingrowth cones (Ho¨pker et al., 1999). In uninfectedspinal neurons, laminin stimulation, and subsequentactivation of cdc42, may promote growth cone ad-vance, with cAMP levels falling within a permissiverange for stimulatory growth. However, chronic ex-pression of CAcdc42 could negatively regulate levelsof cAMP, and the additional decline in cAMP inducedby acute laminin addition now becomes inhibitory. Adetailed study of the effects of cAMP (and cGMP;Caroni, 1998) levels on the outgrowth of neuronsexpressing CA and DNcdc42 will be needed to deter-mine if this model is correct. Interestingly, proteinkinase A phosphorylates rhoA, allowing Rho-GDI tobind to and inactivate rhoA. Thus, a very low level ofcAMP due to chronic expression of CAcdc42 andacute addition of laminin could release rhoA inhibi-tion, which could ultimately result in growth coneinhibition (Lang et al., 1996).
Because enhanced cdc42 activity leads to in-creased neurite outgrowth in embryonic neurons, itis tempting to speculate that cdc42 activity mayalso stimulate neurite outgrowth in the adult ner-vous system. High amounts of cdc42 mRNA andprotein are found in neurons of the adult rat hip-pocampus (Olenik et al., 1997), a brain regionknown to undergo synaptic remodeling. Cdc42mRNA and protein have also been detected in theadult cerebellum, thalamus, and neocortex (Oleniket al., 1997), where cdc42 may be involved insynaptic plasticity. After injury, adult central ner-vous system neurons have little capacity to regen-erate. Expression of CAcdc42 or activation of en-dogenous cdc42 in neurons after injury maypromote neurite regeneration.
362 Brown et al.

REFERENCES
Bamburg JR, Minamide LS, Morgan TE, Hayden SM, Giu-liano KA, Koffer A. 1991. Purification and characteriza-tion of low-molecular weight actin-depolymerizing pro-teins from brain and cultured cells. Methods Enzymol196:125–140.
Bentley D, O’Connor TP. 1994. Cytoskeletal events ingrowth cone steering. Curr Opin Neurobiol 4:43–48.
Caroni P. 1998. Driving the growth cone. Science 281:1465–1466
Colucci-D’Amato GL, Santelli G, D’Alessio A, ChiappettaG, Mineo A, Manzo G, Vecchio G, de Franciscis V.1995. Dbl expression driven by the neuron specific eno-lase promoter induces tumor formation in transgenic micewith a p53(1/2) genetic background. Biochem BiophysRes Commun 216:762–770.
Goodman CS. 1996. Mechanisms and molecules that con-trol growth cone guidance. Annu Rev Neurosci 19:341–377.
Hall A. 1998. Rho GTPases and the actin cytoskeleton.Science 279:509–514.
Hopker VH, Shewan D, Tessier-Lavigne M, Poo M-m, HoltC. 1999. Growth cone attraction to netrin-1 is convertedto repulsion by laminin-1. Nature 401:69–73.
Horii Y, Beeler JF, Sakaguchi K, Tachibana M, Miki T.1994. A novel oncogene, ost, encodes a guanine nucleo-tide exchange factor that potentially links rho and racsignaling pathways. EMBO J 13:4776–4786.
Jalink K, van Corvan EJ, Hengeveld T, Morii N, NarumiyaS, Moolenaar WH. 1994. Inhibition of lysophosphatidate-and thrombin-induced neurite retraction and neuronal cellrounding by ADF ribosylation of the small GTP-bindingprotein rho. J Cell Biol 126:801–810.
Jin Z, Strittmatter SM. 1997. Rac1 mediates collapsin-1-induced growth cone collapse. J Neurosci 17:6256–6263.
Kaufmann N, Wills ZP, Van Vactor D. 1998.DrosophilaRac1 controls motor axon guidance. Development 125:453–461.
Kozma R, Ahmed S, Best A, Lim L. 1995. The ras-relatedprotein cdc42Hs and bradykinin promote formation ofperipheral actin microspikes and filopodia in Swiss 3T3fibroblasts. Mol Cell Biol 15:1942–1952.
Kozma R, Sarner S, Ahmed S, Lim L. 1997. Rho familyGTPases and neuronal growth cone remodelling: rela-tionship between increased complexity induced bycdc42Hs, rac1, and acetylcholine and collapse induced byrhoA and lysophosphatidic acid. Mol Cell Biol 17:1201–1211.
Kuhn TB, Schmidt MF, Kater SB. 1995. Laminin andfibronectin guideposts signal sustained but opposite ef-fects to passing growth cones. Neuron 14:275–285.
Kuhn TB, Brown MD, Bamburg JR. 1998. Rac1-dependentactin filament organization in growth cones is necessaryfor b1 integrin-mediated advance but not for growth onpoly-D-lysine. J Neurobiol 37:524–540.
Kuhn TB, Brown MD, Wilcox CL, Raper JA, Bamburg JR.1999. Myelin and collapsin-1 induce motor neuron
growth cone collapse through different pathways: inhibi-tion of collapse by opposing mutants of rac1. J Neurosci19:1965–1975.
Kuhn TB, Meberg PJ, Brown MD, Bernstein BW, Mi-namide LS, Jensen JR, Okada K, Soda EA, Bamburg JR.2000. Regulating actin dynamics in neuronal growthcones by ADF/cofilin and rho family GTPases. J Neuro-biol, (In press).
Laemmli UK. 1970. Cleavage of structural proteins duringthe assembly of the head of bacteriophage T4. Nature227:680–684.
Lang P, Gesbert F, Delespine-Carmagnat M, Stancou R,Pouchelet M, Bertoglio J. 1996. Protein kinase A phos-phorylation of RhoA mediates the morphological andfunctional effects of cAMP in cytotoxic lymphocytes.EMBO J 15:510–519.
Lewis AK, Bridgman PC. 1992. Nerve growth cone lamel-lipodia contain two populations of actin filaments thatdiffer in organization and polarity. J Cell Biol 119:1219–1243.
Lin C-H, Forscher P. 1993. Cytoskeletal remodeling duringgrowth cone-target interactions. J Cell Biol 121:1369–1383.
Lorenzi MV, Castagnino P, Chen Q, Hori Y, Miki T. 1999.Distinct expression patterns and transforming propertiesof multiple isoforms of ost, an exchange factor for RhoAand cdc42. Oncogene 18:4742–4755.
Luo L, Lioa YJ, Jan LY, Jan YN. 1994. Distinct morpho-genetic functions of similar small GTPAses:DrosophilaDrac1 is involved in axonal outgrowth and myoblastfusion. Genes Dev 8:1787–1802.
Luo L, Hensch TK, Ackerman L, Barbel S, Jan LY, Jan YN.1996a. Differential effects of the rac1 GTPase on Pur-kinje cell axons and dendritic trunks and spines. Nature379:837–840.
Luo L, Jan L, Jan YN. 1996b. Small GTPases in axonoutgrowth. Perspect Dev Neurobiol 4:199–204.
Meberg PJ, Bamburg JR. 2000. Increases in neurite out-growth mediated by overexpression of actin depolymer-izing factor. J Neurosci 20:2459–2469.
Minamide LS, Bamburg JR. 1990. A filter paper dye-bind-ing assay for quantitative determination of protein with-out interference from reducing agents or detergents. AnalBiochem 190:66–70.
Mueller BK. 1999. Growth cone guidance: first steps to-wards a deeper understanding. Annu Rev Neurosci 22:351–388.
Nobes CD, Hall A. 1995. Rho, rac, and cdc42 GTPasesregulate the assembly of multi-molecular focal com-plexes associated with actin stress fibers, lamellipodia,and filopodia. Cell 81:53–62.
O’Connor TP, Bentley D. 1993. Accumulation of actin insubsets of pioneer growth cone filopodia in response toneural and epithelial guidance cues in situ. J Cell Biol123:935–948.
Olenik C, Barth H, Just I, Aktories K, Meyer DK. 1997.Gene expression of the small GTP-binding proteins
Cdc42 Stimulates Neurite Development 363

RhoA, RhoB, Rac1, and Cdc42 in adult rat brain. BrainRes Mol Brain Res 52:263–269.
Rivas RJ, Burmeister DW, Goldberg DJ. 1992. Rapid ef-fects of laminin on the growth cone. Neuron 8:107–115.
Song H-j, Poo M-m. 1999. Signal transduction underlyinggrowth cone guidance by diffusible factors. Curr OpinNeurobiol 9:355–363.
Song H-j, Ming G, He Z, Lehmann M, McKerracher L,Tessier-Lavigne M, Poo M-m. 1998. Conversion of neu-ronal growth cone responses from repulsion to attractionby cyclic nucleotides. Science 281:1515–1518.
Sumi T, Matsumoto K, Takai Y, Nakamura T. 1999. Cofilinphosphorylation and actin cytoskeletal dynamics regu-lated by rho- and Cdc42-activated LIM-kinase 2. J CellBiol 147:1519–1532.
Suter DM, Forscher P. 1998. An emerging link betweencytoskeletal dynamics and cell adhesion molecules ingrowth cone guidance. Curr Opin Neurobiol 8:106–116.
Tanaka E, Sabry J. 1995. Making the connection: cytoskel-etal rearrangements during growth cone guidance. Cell83:171–176.
Threadgill R, Bobb K, Ghosh A. 1997. Regulation of den-dritic growth and remodeling by rho, rac, and cdc42.Neuron 19:625–634.
Tigyi G, Fischer DJ, Sebok A, Yang C, Dyer DL, Miledi R.1996. Lysophosphatidic acid-induced neurite retractionin PC12 cells: control by phosphoinositide-Ca21 signal-ing and Rho. J Neurochem 66:537–548.
Towbin H, Staehelin T, Gordon J. 1979. Electrophoretictransfer of proteins from polyacrylamide gels to nitrocel-lulose sheets: procedure and some applications. Proc NatlAcad Sci USA 76:4350–4354.
Wessel D, Flugge UI. 1984. A method for the quantita-tive recovery of protein in dilute solution in the pres-ence of detergents and lipids. Anal Biochem 138:141–143.
364 Brown et al.





























