Embed Size (px)
Citation preview

1
河川基金助成事業
河川敷の植物間相互作用から外来植物の 繁茂メカニズムを解明する
助成番号 28-5311-016 秋田県立大学 生物資源科学部 生物環境科学科
坂田 ゆず
平成 28 年度

様式6・2 1.研究者・研究機関 [概要版報告書]
助成番号 助成事業名 所属・助成事業者氏名
28-5311-016 河川敷の植物間相互作用から外来植物の
繁茂メカニズムを解明する (秋田県立大学・坂田ゆず)
助
成
事
業
の
要
旨
〔目 的〕 河川生態系には、氾濫や水辺の環境に適応した特徴的な植生が見られるが、近年の人為撹乱に
より、外来植物が優占する群落が増加している。またこれらの外来植物を利用する外来昆虫が、
河川敷沿いに分布を拡大し、在来植物への食害も見られる。また、外来植物の分布は、土壌などの
物理的な環境にも依存する。本研究の目的は、都市河川に分布する外来植物と在来植物の被度と食
害に与える影響を解明することである。本研究では特に、キク科を食害する外来昆虫のアワダチ
ソウグンバイに着目し、グンバイの侵入の有無によって、河川敷沿いに広く優占するキク科の外来
植物と在来植物の優占度及び食害の程度がどのように異なるかについて調べた。河川生態系におい
て外来植物が優占するメカニズムを解明し、日本の河川敷特有の在来植物を保全する上で有効な
指針となる。 〔内 容〕 河川敷には、在来・外来植物ともに多種のキク科多年生草本が見られる。本研究では、キク科を
広く食害する外来の植食性昆虫であるアワダチソウグンバイ(以下グンバイ)の侵入が河川敷を優
占するキク科の在来植物と外来植物の食害の程度の違いに及ぼす影響を野外調査と実験によって
調べた。グンバイの侵入していない秋田県雄物川(4地点)と、侵入して 10 年以上が経過してい
る京都府桂川(4地点)において、外来植物と在来植物の被度(2m×2m×10)を記録した。次に、
キク科とマメ科(外来の植食性昆虫の分布に違いが見られない対象種として)の外来植物と在来植
物それぞれ 5種において、植食性昆虫の個体数および食害率を測定した。また、各調査地点におい
て、土壌を採集しリン濃度と pHを測定した。また、グンバイの食害が見られたキク科植物 8種(在
来植物:ノコンギク、アキノキリンソウ、シマカンギク、アズマギク;外来植物:セイタカアワダ
チソウ、ヒメムカシヨモギ、オオブタクサオオハンゴンソウ)について、飼育環境下でグンバイの
成長を比較した。 〔結 果〕 在来昆虫は、秋田では在来のキク科植物を多く食害していたが、京都では外来のキク科植物を多
く食害していた。一方で外来昆虫は、外来植物を多く食害していた。グンバイを含めた外来昆虫の
個体数は、在来昆虫の個体数に負の影響を与えていた。秋田では在来植物と外来植物とで食害の程
度に違いは見られなかったが、京都では外来植物が在来植物よりも食害率が高く、特にキク科で顕
著にその差が見られた。外来植物の被度に土壌条件による影響は見られなかった。グンバイの食害
が見られたキク科植物 8 種について、飼育環境下でグンバイの成長を比較したところ、そのうち卵
が孵化し、成虫まで成長したのは 5 種であった。グンバイの成長について、在来植物・外来植物に
よる違いは見られなかったが、種によって次世代の個体数と生存率にも差が見られた。 以上により、グンバイの侵入によって、在来昆虫による食害が低下し、グンバイが外来植物を選
択的に食害することで、外来植物が在来植物に比べて多くの食害を受けていることが明らかになっ
た。グンバイの侵入は外来植物と在来植物の食害のされ方を変化させたが、優占度への影響は見ら
れなかった。一方で、飼育環境下では、在来植物と外来植物で大きな違いは見られなかったものの、
種によって違いが見られた。グンバイの侵入が河川敷のキク科在来植物に与える影響は、その地域
の植物群集の種組成によって大きく異なる可能性が考えられる。外来昆虫が生物学的防除として、
在来植物の保全に活用できるかどうかは、導入場所の植物の種組成などを考慮して慎重に考える必
要がある。 調査対象水系・河川 データベースに登
録するキーワード 部門 大分類 中分類 小分類
調査部門 環境 生態系 植生 ※データベースに登録するキーワードは、本冊子 P.43の表から代表的なものを一つ記入して下さい。

様式6・3 1.研究者・研究機関 [自己評価シート]
助成番号 助成事業名 所属・助成事業者氏名
28-5311-016 河川敷の植物間相互作用から外来植物の
繁茂メカニズムを解明する (秋田県立大学・坂田ゆず)
助
成
事
業
実
施
成
果
の
自
己
評
価
〔計画の妥当性〕 本研究では、これまで注目されてこなかった、外来昆虫の侵入によって、外来植物が在来植物に
与える影響を解明することで、生物学的防除の外来種管理法や在来種の保全策として、研究成
果を応用できると考えた。外来昆虫のアワダチソウグンバイの侵入の有無に着目し、2つの都市河
川敷での野外調査と実験を行った。河川敷沿いの集団は選定済みであり、実験の場所も確保済みで
あったため効率的に研究を行うことができた。河川の土壌分析においても、助成事業者が在籍する
秋田県立大学生物環境科学科で協力して行うことができた。昆虫の飼育実験においても、一部の植
物を既に圃場で生育しており、研究開始前に円滑に研究を推進する準備ができていた。これらを総
合すると、本研究は妥当な計画であった。
〔当初目標の達成度〕 本研究では、予定していた野外調査と飼育実験を終えることができた。当初予定していた圃場実験
については、植物の生育が間に合わず行うことができなかったが、材料は十分に準備済みであり、
今後行う予定である。 〔事業の効果〕 本研究から、外来昆虫の侵入が必ずしも外来植物だけではなく、在来植物をも食害することで、分
布をさらに拡大している可能性が示された。また、外来植物が繁茂するメカニズムの1つとして、
植食性昆虫の食害が考えられることが示された。得られた成果は、今後河川植生の整備において、
外来植物を管理するにあたり、生物学的防除を導入する場合には、その地域の植物群集の種組成な
どを考慮して慎重に行う必要があることが明らかになった。本年度の成果は、日本生態学会におけ
る学会発表で公表したほか、H28 年度秋田県立大学卒業論文発表会においても発表を行った。今
後、論文発表を行う準備をすすめていく予定である。 【学会発表】坂田ゆず , 村上昌宏 「河川敷を優占するキク科の在来植物と外来植物における植食
性昆虫および食害の比較」日本生態学会第 64 回大会,東京,2017 年 3 月 〔河川管理者等との連携状況〕 本研究では、河川管理者との連携は特にありません。

2
目次 1.はじめに ……………………………………………………………………………………………………….3 1.1.河川における植物の撹乱………………………………………………………………………………...3 1.2. 外来植物の侵入と在来昆虫による食害………………………………………………………………..3 1.3. 外来昆虫の侵入…………………………………………………………………………………………3 1.4.外来昆虫アワダチソウグンバイの侵入…………………………………………………………………4 1.5.本研究の目的……………………………………………………………………………………………..4 2.調査地………………………………………………………………………………………………………….4 3.方法…………………………………………………………………………………………………………….4 3.1.野外調査…………………………………………………………………………………………………..4 3.1.1在来・外来植物の被度………………………………………………………………………………4 3.1.2食害率……………………………………………………………………………………………......5 3.1.3植食性昆虫の個体数…………………………………………………………………………….…..5 3.1.4土壌特性……………………………………………………………………………………...………5 3.2.飼育実験:キク科植物におけるグンバイの成長の比較………………………………………………..5 4.結果…………………………………………………………………………………………………………….6 4.1.在来・外来植物の被度……………………………………………………………………………………6 4.2.土壌特性が在来・外来植物の相対優占度に与える影響………………………………………………6 4.3.食害率……………………………………………………………………………………………………..6 4.4.植食性昆虫の個体数……………………………………………………………………………………...6 4.5.キク科植物におけるグンバイの成長の比較……………………………………………………………7 5.考察…………………………………………………………………………………………………………….7 5.1. グンバイの侵入が在来植物と外来植物の食害の程度の違いと優占度に及ぼす影響………………7 5.2. 外来植物と在来植物におけるグンバイの食害の程度の違い ………………………………………..8 5.3.まとめ ………………………………………………………………………………………………………8 6.引用文献 ………………………………………………………………………………………………………8 7.図表………………………………………………………………………………………………………...…11

3
1.はじめに
1.1.河川における植物の撹乱
河川敷は森林などほかの地域に比べて自然撹乱が起こりやすい。河川の氾濫や増水などにより、河川
植生が撹乱を受ける(宮脇と鷲谷 2004)。日本において河川敷は、撹乱耐性のあるカワラノギクやカワ
ラハハコなどの固有の植物が見られる貴重な生息地である。外来植物は撹乱依存的な特性を持っており、
在来植物よりも栄養生長期間は短く、種子の時間的・空間的散布能力が大きい種類が多い(鷲谷と矢原 1996)。そのため河川敷では撹乱後の植生の回復が早く、外来植物の侵入・定着を引き起こしやすい。
そして、日本の帰化植物のほとんどが、撹乱依存種としての性格を持っている(鷲谷と矢原 1996)。日
本では穀物や農産物の輸入に伴い、雑草種子が頻繁に日本に持ち込まれている。また、外来牧草や外来
マメ科樹木などが「緑化材料」として大量に意図的導入され、それらは緑化場所から逸出して広がり,
河原など,自然の生息・生育場所でも優占種となっている(鷲谷 2007)。緑化用のイネ科の外来牧草類
や,穀物の輸入に伴う北米原産のキク科外来植物の侵入が推測されるとともに, 河原固有の在来植物の衰
退が著しい(浦口ら 2003)。植物の生育・繁殖には植食者の食害などによる生物的要因と土壌条件など
の非生物的な要因が影響を与えていると考えられる。 外来植物の優占に影響を与える可能性がある要因として、天敵の植食性昆虫による食害などの生物的
な要因と、土壌特性などの非生物的要因がある。日本では、降水量が多いことから酸性土壌が広く分布
しており、日本の在来植物は、酸性土壌に対する耐性を持った植物が多い一方で、外来植物は、比較的
pH が高く(pH5.7 以上)、さらに有機体リン酸が 20mg/100g 以上で外来植物が多く生育する傾向にあ
るという報告例がある(平舘ら 2008)。本来日本の土壌は外来種にとって生育しにくいものであるが、
農地にまかれるリンなどによって外来植物の蔓延しやすい環境を作り出してしまっている可能性も指摘
されている(平舘 2012)。河川においても、地域や流域によって土壌特性や撹乱のしやすさが異なるた
め、外来植物の生育のしやすさに違いがある可能性がある。 1.2. 外来植物の侵入と在来昆虫による食害
植物の天敵となる植食者は植物の生育・繁殖に大きな影響をもたらす。一般に外来植物は侵入した土
地で生育や繁殖に負の影響を与える植食性昆虫が少ないことが、侵入地での拡大を招いていると提唱さ
れてきた(ERH仮説:Crawley 1987)。さらに、外来植物は侵入地で植食性昆虫への防御にかかるコスト
が低下した分、資源を繁殖や成長に投資できるため、在来植物に比べて成長しやすいことが報告されて
いる(EICA仮説:Blossey and Notzold 1995)。しかし一方で、外来植物は在来昆虫に対する抵抗性を持
っていないため、在来植物よりも昆虫による食害を受けやすいという報告例がある(Keeler and Chew 2008)。さらに近年、昆虫の中でも、スペシャリストとジェネラリストによって、外来植物への食害の
及ぼし方が異なることにも注目されている(Müller-Schärer et al. 2004)。 このように在来昆虫が外来植物に与える食害については多様なパターンがあり、環境条件や、昆虫の
生態学的特徴によって異なると考えられる。 1.3. 外来昆虫の侵入
これまで外来植物の生物学的防除として植食性昆虫が原産地から意図的に導入されてきた(Klinken and Edwards 2002)。また、近年では外来植物を追って外来昆虫が非意図的に侵入している現象が世界
中で起きている(Yamanaka et al. 2015)。日本においても、239種ほどの外来昆虫の侵入が確認されて
いる(Morimoto and Kiritani 1995)。しかし、外来昆虫は、適応していないために抵抗性を持たない在

4
来植物と、原産地において適応している外来植物のどちらを食害しやすいかについて理解が進んでおら
ず、さらに外来昆虫が在来昆虫の食害に及ぼす影響も注目されてこなかった。 1.4.外来昆虫アワダチソウグンバイの侵入
キク科植物を食害する外来昆虫であるアワダチソウグンバイ(以下グンバイ)は、北米原産で日本で
は 2000年に初めて侵入が報告された。グンバイは関西では侵入して10年以上たっており、侵入してい
る宮城・山形以南の地域ではキク科植物の他サツマイモやナスなどが被害を受けている(Kato and Ohbayashi 2009, 宮城県病害虫防除所 2013, 埼玉県病害虫防除所 2007, 福島県病害虫防除所 2008, 長野県病害虫防除所 2008, 熊本県病害虫防除所 2007, 香川県病害虫防除所 2005)が、北東北にはま
だ侵入していない。そこで、本研究では、グンバイとキク科植物を材料に用いて、外来昆虫の分布の有
無によって在来植物と外来植物の食害のされ方、及び植物の被度がどのように変化するのか調べた。グ
ンバイの侵入によってキク科外来植物が食害されることで在来植物が増加し、外来植物の被度に影響を
及ぼすのではないかと考えた。 1.5.本研究の目的
本研究では、①グンバイの侵入が河川敷における在来植物と外来植物の食害の程度の違いと優占度に
及ぼす影響、②グンバイにおける外来植物・在来植物の食害の程度の違いが遺伝的または可塑的に決ま
っているのかを明らかにすることを目的とした。 グンバイの侵入している地域と侵入していない地域の河川敷で、各流域ごとに植物の被度、土壌特性
の違いを考慮したうえで、在来/外来植物の食害の程度の違いを調べた。次に、インキュベータを用いた
温度、湿度、照度の同一条件下で、グンバイにキク科在来/外来植物各 4種のうち 1種を与え、グンバイ
の成長を比較した。 2.調査地
グンバイの侵入していない秋田と侵入して 10 年以上経っている京都の河川において上流から下流ま
でそれぞれ 4地点を調査地とした(下記地図)。
3.方法
3.1.野外調査
3.1.1在来・外来植物の被度
昆虫による植物の食害の蓄積が見られる 2016 年 9 月にグンバイの侵入していない秋田と、侵入して
1.京都調査地 2.秋田調査地

5
10年以上が経過している京都のそれぞれ4地点の調査地ごとに、15個ずつ2m×2mプロットをつくり、
植物種ごとに被度を記録した。 解析手法:植物の被度に及ぼす影響をポアソン分布を仮定した一般化線形混合モデル(以下GLMM)
を構築して解析を行った。1 プロット中の植物種当たりの被度(キク科のみ、マメ科のみについても)
を応答変数とし、地域(秋田・京都)、河川流域(上流・中流上・中流下・下流)、植物種の在来・外来、地
域×植物種の在来・外来の交互作用を説明変数とし、植物の科(キク科とマメ科のみについては、種を)
ランダム効果とした。 3.1.2.食害率
植生調査で優占していた上位 2 つの科であり、外来昆虫の分布に地域差のあるキク科と地域差のない
マメ科において、8つの調査地で共通して生育が確認された種(表 1)を対象に、食害の割合と植食性昆虫
の個体数の測定を行った。各植物について食害の割合を4段階(0, 1~30, 31~60, 61~100%)に分類
して測定した。 解析手法:キク科、マメ科、キク科+マメ科における食害の大きさに及ぼす影響について、GLMMを
用いて解析した。葉 1 枚当たりの食害の積算値を応答変数とし、地域(秋田・京都)、河川流域(上流・中
流上・中流下・下流)、葉の枚数、植物種の在来・外来、植物の科(キク科+マメ科のみ)、地域×植物種
の在来・外来と地域×植物種の在来・外来×植物の科の交互作用を説明変数とし、植物の種をランダム
効果とした。 3.1.3.植食性昆虫の個体数
食害割合を見た植物個体において、植食昆虫の種、個体数、外来種・在来種の別を記録した。 解析手法:在来昆虫、グンバイ以外の外来昆虫、グンバイ、全体の昆虫の個体数に及ぼす影響につい
て、それぞれGLMMを用いて解析した。昆虫の数を応答変数とし、地域(秋田・京都)、河川流域(上流・
中流上・中流下・下流)、植物種の在来・外来、地域×植物種の在来・外来の交互作用を説明変数とし、
植物の種をランダム効果とした。 3.1.4.土壌特性
各流域において、5 か所から土壌を採取し、混合したものを風乾してトルオーグ法とブレイ法で有機
態リン酸を分析し、ガラス電極法(H₂O, KCl)でpHを測定した。 解析手法:土壌条件が植物の被度に及ぼす影響をポアソン分布を仮定した一般化線形モデル(以下
GLM)を用いて解析し、結果を表 3に示した。キク科の外来植物の被度またはマメ科の外来植物の被度
を応答変数とし、地域(秋田・京都)、河川流域(上流・中流上・中流下・下流)、pH(H₂O または KCl)、可給態リン酸(トルオーグ法またはブレイ 2法)を説明変数とした。 3.2.飼育実験:キク科植物におけるグンバイの成長の比較
キク科の在来種と外来種各 4種において 10サンプル用意し、1サンプルにつき葉とグンバイ雄雌各 2個体ずつを入れた。温度25℃、照度 3L(10000lux)湿度 70%の同一条件下で飼育を行った。気温は秋
田と京都の 7月と 9月の気温を平均した値として、25℃に設定した。グンバイの食害の有無、産卵の有
無、孵化に要した時間、孵化した幼虫数、成虫数を測定し、生存率を算出した。 解析手法:グンバイの生育が在来植物と外来植物で異なるかについて、GLMM を用いて解析した。

6
産卵の有無、孵化までの日数、孵化までの日数、成虫数、生存率を応答変数とし、植物種の在来・外来
を説明変数とし、植物の種をランダム効果とした。 上記全てのGLMM(GLM)において、説明変数の効果の有用性はカイの 2乗分布(F 分布)を仮定した尤
度比検定によって確認した。また、解析には、lme4パッケージ(Bates et al. 2016)と carパッケージ(Fox and weisborg 2011)を用いて、R.3.3.2を使用した。 4.結果
4.1.在来・外来植物の被度
秋田と京都両地域の河川においてキク科、マメ科、イネ科の順に被度が大きかった。解析結果から、
植物の被度には、流域、地域、植物の在来・外来が影響を与えていた(表 2-1)。秋田の方が京都よりも
被度が大きく、外来植物よりも在来植物の被度の方が有意に大きかった(表 2-1)。また、在来・外来と
地域の交互作用項が影響を与えていた。京都では外来植物の方が在来植物より被度が小さかったが、秋
田では両者の差は京都ほど顕著でなかった(図 3)。また、京都の外来植物の被度は秋田に比べて小さか
った。
キク科においては流域の影響を受けており、京都より秋田で被度が大きく、外来植物より在来植物の
被度が大きかった。マメ科は秋田よりも京都で被度が大きかった(表 2-2,図 4)。在来植物と外来植物の
被度の大きさに違いはなかった。マメ科は植物の種組成が秋田と京都で大きく異なっていた(表 2-3,図
5)。
4.2.土壌特性が在来・外来植物の相対優占度に与える影響
キク科とマメ科の外来植物の被度は、どちらも秋田より京都で大きく、流域が影響を与えていた。今
回の結果では pH、リン酸含有量は、外来植物の被度に影響を与えておらず、土壌特性に関わらず在来植
物と外来植物の被度が同程度または在来植物の被度が大きい傾向を示した(表 3,図 6)。
4.3.食害率
解析結果では食害率が、キク科、マメ科、キク科+マメ科の全てで、秋田よりも京都の方が有意に高か
った(表 4)。また、すべての項目に河川が影響しており、葉数が正の影響を与えていた。植物の在来・
外来による影響は見られなかった(表 4)。地域×在来・外来の交互作用項が影響を与えていたことから、
在来植物と外来植物での食害の差が秋田と京都で違っており、京都では外来植物が在来植物よりも食害
率が高かった(表 4)。さらに、地域×在・外×科の交互作用項が影響を与えており、マメ科よりもキク
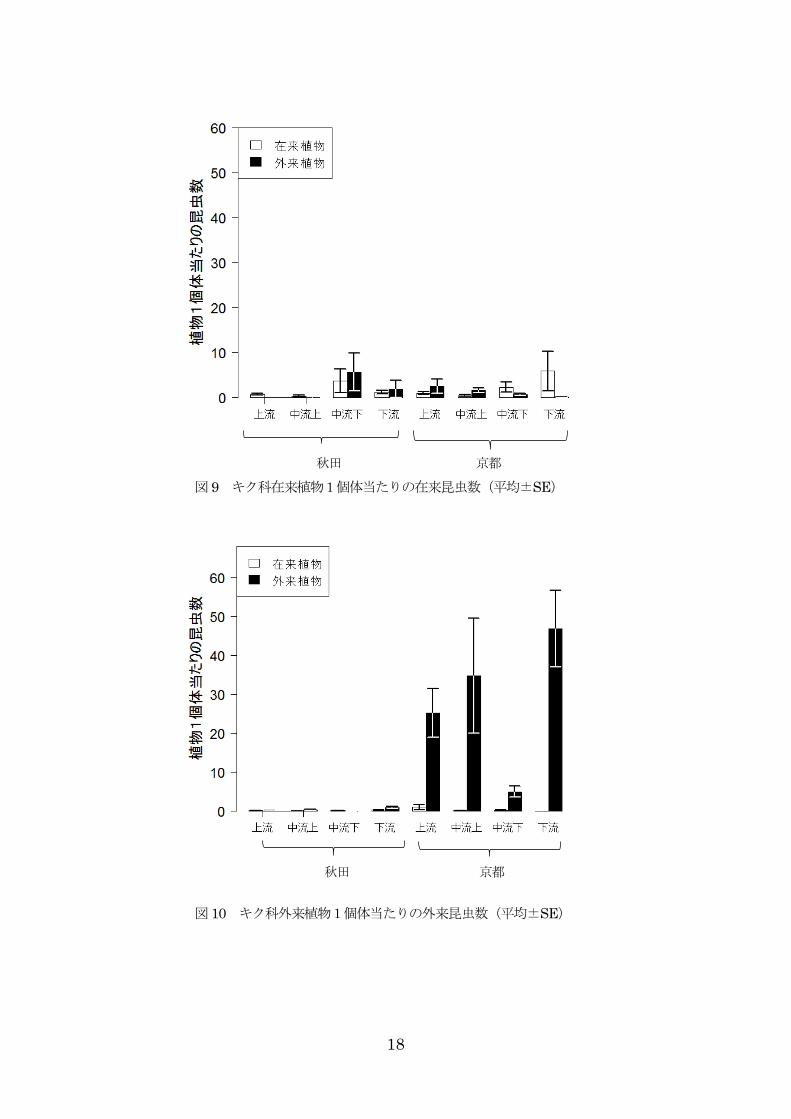
科で顕著にその差が見られた(図 7,8)。
4.4.植食性昆虫の個体数
昆虫に注目してみると在来昆虫、外来昆虫(グンバイ以外)、全体の個体数は秋田より京都の方が多く
見られた(表 5)。また、すべてにおいて流域が影響しており、葉の数は在来昆虫の数には影響していな
かったが、それ以外の昆虫の個体数には正の影響が見られた(表 5)。地域×在来・外来の交互作用も在
来昆虫、外来昆虫、全体の個体数に影響を与えていた。
昆虫全体の個体数には、植物の科が影響を与えていた(表 5)。キク科はマメ科に比べて昆虫全体の個
体数が多かった(表 5)。また、秋田では個体数に在来植物と外来植物で大きな差は見られなかったが、

7
京都では在来植物に比べて外来植物で個体数が多かった(図 14)。
在来昆虫の個体数では、植物の在来/外来と科による影響は見られなかった(表 5)。外来昆虫が在来
昆虫に対して負の影響を与えていた。マメ科では、どちらの地域でも在来植物を食害しているが、キク
科では秋田で在来植物よりも外来植物を食害し、京都で外来植物よりも在来植物を食害していた(図
9,11)。
外来昆虫は、マメ科では両地域で在来植物を多く食害していたが、キク科では京都で外来植物をより
食害していた(表 5,図 10,12)。また、在来昆虫が外来昆虫に負の影響を与えており、京都で観察された
外来昆虫の個体数うち、ほとんどがグンバイであった。
グンバイにおいては、在来植物よりも外来植物を食害しており(図 13)、在来昆虫からは負の影響を
受けていたが、外来昆虫からは正の影響を受けていた(表 5)。
植物 1個体あたりの昆虫数をギルド別にみると、秋田ではキク科の在来植物で、吸汁性の昆虫とゴー
ル性の昆虫が多かった。京都では、キク科の外来植物を吸汁する昆虫が大部分を占めていた(図 15)。こ
のほとんどの個体数をグンバイが占めていた。また、在来昆虫のタマバエ科のスペシャリストの昆虫が
ヨモギにおいて見られたことが影響していると考えられる。マメ科では、在来植物と外来植物で顕著な
差は見られなかったが、両地域において葉食性の昆虫が多く、京都では在来植物において吸汁性昆虫が
多い結果となった(図 16)。これらはマルカメムシなどの在来のカメムシや、ジェネラリストの葉植生の
昆虫が見られたことが影響していると考えられる。
4.5.キク科植物におけるグンバイの成長の比較
今回調査した 8種のうちシマカンギク以外の 7種でグンバイの卵が確認できた(図 17.1)。その後孵化
し、成虫まで成長したものはそのうちセイタカアワダチソウ、ヒメムカシヨモギ、アキノキリンソウ、
ノコンギク、アズマギクの 5種であった(図 17.5)。ヒメムカシヨモギは 10サンプルのうち 2サンプル
のみで卵が確認できた(表 6)。オオブタクサとオオハンゴンソウの 2 種は産卵が見られたが、孵化した
ものは見られなかった。孵化が見られた植物種の中で、孵化までの日数は全体で大きな差は見られず、
約 20 日前後であった。成虫数は、幼虫数と同様のパターンが見られたが、約 70~80%の生存率であっ
た。ヒメムカシヨモギだけは生存率が 27.8%と低い値になっていた。以上のことから、グンバイの 1世
代の生活史における成長において、植物種によってばらつきが見られた。しかし、解析結果よりすべて
の測定項目において在来植物・外来植物による違いは見られなかった(表 7)。
5.考察
5.1. グンバイの侵入が在来植物と外来植物の食害の程度の違いと優占度に及ぼす影響
在来昆虫の個体数は、植物の在来・外来の影響を受けていなかったが、外来昆虫は外来植物よりも在
来植物で多く見られた。しかし、グンバイは外来植物の方を食害していたことから、外来昆虫の中でも
種によって、在来植物と外来植物に与える食害の影響は異なると考えられる。また、京都ではグンバイ
が外来植物を多く食害したため、京都で秋田に比べて、特にキク科において外来植物が在来植物に比べ
て植食昆虫の個体数、食害率が大きかったと考えられる。
また、在来昆虫と外来昆虫の個体数は、負の関係が見られたことから、秋田では、在来昆虫は外来植
物上で多く見られたが、京都ではグンバイを避けて在来植物上で多く見られた要因である可能性がある。
植物の被度、食害率の結果から、グンバイの侵入している京都でキク科の外来植物が秋田よりも食害を

8
多く受けているが、外来のキク科植物の被度は低下していなかった。このことから、個体数や食害率で
は測定できない、在来昆虫による植物の適応度に与える影響が、在来植物に比べて外来植物上で大きか
ったことが、食害率が外来植物上で大きいにもかかわらず被度に与える影響は見られなかったことの理
由の一つかもしれない。
一方で、外来昆虫であるグンバイの個体数は、他の外来昆虫の個体数からは正の影響を受けていた。
そのため、今後、他の外来昆虫がグンバイの侵入を促進させ、グンバイの侵入は外来植物上の在来昆虫
の個体数を減少させる可能性が考えられる。
京都では、グンバイが侵入してから 15年以上経過しているが、今後の時間経過に伴い、植物被度に影
響が与えられるかについて、より長期的な観測を続けていく必要があるだろう。また、土壌 pHと可給態
リン酸含有量といった土壌特性は、植物の被度に影響を及ぼしていなかった。その理由として、先行研
究に比べて、調査地点間の土壌特性の変異が小さかったため、影響が見られなかったことが考えられる。
5.2. 外来植物と在来植物におけるグンバイの食害の程度の違い
飼育実験よりグンバイの孵化までに日数、生長期間は植物種によらず一定であることが分かった。し
かし、次世代の個体数は植物種ごとに大きなばらつきがあった。そして、孵化した幼虫は植物を食害し、
成虫になることができたため、親が食害できる植物は幼虫も食害できると考えられる。
次世代の個体数・生存率は外来植物と在来植物では違いが見られなかった。しかし、在来植物の中で
もアズマギクは次世代の個体数が多く、アキノキリンソウは少ない、また外来昆虫の中でもセイタカア
ワダチソウでは個体数が多く、ヒメムカシヨモギでは少ないなどのように、グンバイの食害の程度の違
いは植物種によって遺伝的に異なっていると考えられる。しかし、野外では外来植物が在来植物に比べ
て大きく食害されていた。そのため、グンバイは多くの種を食害する能力を持っているが、植物種によ
って嗜好性が異なるため、野外ではその環境にある植物の中から選択的に、外来植物を食害していたと
考えられる。しかし、在来植物のみの環境にグンバイが侵入した場合、その植物を食べる能力があれば
グンバイは繁殖できる可能性がある。植物種ごとにグンバイの食害を受けるものと受けないものを調査
し、河川環境の植物群集の種組成に応じて、食害への影響を把握する必要があると考えられる。
5.3.まとめ
以上の結果より、グンバイの侵入は、在来昆虫の外来植物上の食害を低下させ、在来植物と外来植物
両方の食害を増加させ、特に外来植物の食害を増加させたが、外来植物の優占度には影響を与えていな
かった。また、野外集団においては、京都で外来植物が在来植物に比べて多くの食害を受けていたが、
飼育実験では、種によって遺伝的に異なってはいるものの在来・外来に大きな違いは見られなかったこ
とから、グンバイの侵入が食害に与える影響は植物種によって異なると考えられる。そして、グンバイ
は外来植物の被度だけでなく在来植物も食害する可能性があるため、生物学的防除として活用できるか
どうかは導入できるかどうかは導入場所の植物の種組成などを慎重に考える必要がある。
6.引用文献
Kato A. and Ohbayashi N.(2009)Habitat expansion of an exotic lace bug, Corythucha marmorata (Uhler) (Hemiptera: Tingidae), on the Kii Peninsula and Shikoku Island in western Japan Island in western Japan. Entomological Science 12, 130–134
Blossey B. and Notzold R. (1995) Evolution of increased competitive ability in invasive

9
nonindigenous plants: a hypothesis. Ecology 83:887-889 平舘俊太郎(2012)セイタカアワダチソウを抑えて在来植生に戻す.農業環境技術研究所研究成果発表会 平舘俊太郎,楠本良延,森田沙綾香(2008)外来植物の侵入は土壌pHと有効態リン酸に関連している. 農
業環境技術研究所研究成果情報 主要研究情報 7 深野裕也(2012)外来雑草の進化生態学-天敵昆虫に対する防御の急激な進化. 関雑研会報 23: 34-42 Parker J., Burkepile D.E., Hay M.E.(2006)Opposing Effects of Native and Exotic Herbivores on
Plant Invasions. Science 311:1459-1461 Keeler M.S. and Chew F.S. (2008) Escaping an evolutionary trap: preference and performance of a
native insect on an exotic invasive host. Oecologia 156:559–568 宮脇成生,鷲谷いづみ (2004)生物多様性保全のための河川における侵略的外来植物の管理. 応用生態工
学 6:195−209 Morimoto N. and Kiritani K. (1995) Fauna of exotic insects in Japan. 農業環境技術研究所報告
12:87-120 Klinken D.R and Edwards OR.(2002)Is host-specificity of weed biological control agents likely to
evolve rapidly following establishment?. Ecology Letters, 5:590-596 Keane R.M. and Crawley M.J.(2002)Exotic plant invasions and the enemy release hypothesis.
Trends in Ecology & Evolution 17:164-170 Yamanaka T., N. Morimoto, G.M. Nishida, K. Kiritani, S. Moriya, A.M. Liebhold(2015)Comparison
of insect invasions in North America, Japan and their Islands. Biol Invasions 17:3049-3061 Bezemer T.M., Harvey J.A., Cronin J.T.(2013)Response of Native Insect Communities to Invasive
Plants. Annu. Rev. Entomol 59:119–41 浦口晋平,渡邉泉,久野勝治,星野義延,藤井義晴(2003)多摩川中流域の河川敷植生構成種の他感作用. 雑
草研究 48-117-129 鷲谷いづみ(2007)外来種の定着と侵略性の生態学的要因. Nippon Suisan Gakkaishi 73
1117−1120 鷲谷いづみ,矢原徹一(1996)保全生態学入門. 文一総合出版,:103-104 Morrison W., Hay M.(2011)Herbivore preference for native vs. exotic plants: generalist herbivores
from multiple continents prefer exotic plants that are evolutionarily naïve. PLoS ONE6:e17227
Sakata Y., Yamasaki M., Isagi Y., Ohgushi T.(2014)Ecology, 95:2569–2578 宮城県病害虫防除所(2013)特殊報第 1号 香川県病害虫防除所(2005)病害虫発生予察特殊報第1号 熊本県病害虫防除所(2007)病害虫発生予察特殊報第1号 埼玉県病害虫防除所(2007)特殊報第 3号 長野県病害虫防除所(2008)病害虫発生予察特殊報第4号 福島県病害虫防除所(2008)病害虫発生予察特殊報第3号 ・助成事業者紹介
坂田ゆず
現職:秋田県立大学 生物資源科学部 生物環境科学科 助教、博士(理学)

10
主な著書、原著論文:
Y. Sakata, M. Yamasaki, Y. Isagi, T. Ohgushi (2014) An exotic herbivorous insect drives the evolution of resistance in the exotic perennial herb Solidago altissima. Ecology 95: 2569-2578. Y. Sakata, J. Itami, Y. Isagi, T. Ohgushi (2015) Multiple and mass introductions from limited origins: genetic diversity and structure of Solidago altissima in t he native and invaded range. Journal of Plant Research 128: 909-921.
・研究協力者
村上昌宏
現職:岩手県職員、秋田県立大学 生物資源科学部卒

11
7.図表
表 1 食害率と植食性昆虫の調査を行った植物種
表に○が書かれているものがその地点で食害を調査した植物となっている。秋田と京都で共通
している種を中心に調査した。1つの流域のみに生育するものやその地域に生育していなかっ
たものは同じ科の近縁種を調査した。
図1 雄物川1プロットあたりの優占していた
植物の科ごとの被度 図 2 淀川・桂川 1 プロットあたりの優占していた
植物の科ごとの被度

12
表 2-1 在来植物と外来植物の被度に影響を及ぼす要因について GLMMで解析した結果
秋田 京都
図 3 流域ごとの植物の被度
1プロット当たりの平均被度±SE
要因 被度χ² Pr
地域 8.6 0.003
流域 104.4 <0.001在/外 6.9 0.03在/外×地域 5.0 0.03

13
表 2-2 キク科在来植物と外来植物の被度に影響を及ぼす要因について GLMMで解析した結果
要因 被度
χ² Pr
地域 19.2 <0.001流域 20.3 <0.002在/外 17.2 <0.003在/外×地域 0.89 0.3
秋田 京都
図 4 流域ごとのキク科植物の被度
1プロット当たりの平均被度±SE

14
表 2-3 マメ科在来植物と外来植物の被度に影響を及ぼす要因についてのGLMMで解析した結果
要因 被度
χ² Pr
地域 59.2 <0.001流域 55.9 <0.001在/外 2.2 0.3在/外×地域 1.2 0.3
秋田 京都
図 5 流域ごとのマメ科植物の被度
1プロット当たりの平均被度±SE

15
表 3 土壌条件がキク科とマメ科の被度に及ぼす要因についてGLMMで解析した結果
要因 キク科H₂O+truog KCl+truog H2O+bray KCl+brayF Pr F Pr F Pr F Pr
地域 235.1 0.002 237.5 0.002 128.1 0.02 112.7 0.03流域 332.3 0.003 253.1 0.02 228.6 0.02 234.3 0.02H2O 60.5 0.1 32.2 0.2KCl 33.9 0.2 11.4 0.5truog 3.1 0.7 7.8 0.6bray 4.3 0.7 14.7 0.4要因 マメ科
H₂O+truog KCl+truog H2O+bray KCl+bray
F Pr F Pr F Pr F Pr
地域 354.1 0.0002 252.0 0.002 187.3 0.006 104.2 0.05流域 411.7 0.0008 292.3 0.01 339.8 0.003 330.9 0.01H2O 165.4 0.009 89.6 0.05KCl 61.6 0.1 14.8 0.5truog 0.3 0.9 3.6 0.7bray 0.5 0.9 32.7 0.3
図 6 土壌 pHとリン酸が被度に与える影響

16
図 8 マメ科葉1枚当たりの食害率(平均±SE)
秋田 京都
秋田 京都
図 7 キク科とマメ科における葉 1枚当たりの食害率(平均±SE)の比較

17
表 4 キク科とマメ科の食害率に及ぼす要因について GLMMで解析した結果
表 5 昆虫の個体数に及ぼす要因について GLMMで解析した結果
要因 キク科 マメ科 全体 全体χ² Pr χ² Pr χ² Pr χ² Pr
地域 4852.2 <0.001 102.1 <0.001 4002.4 <0.001 4127.2 <0.001流域 248.4 <0.001 34.3 <0.001 297.6 <0.001 295.6 <0.001葉数 4344.8 <0.001 452.6 <0.001 2701.7 <0.001 3091.4 <0.001在/外 3.3 0.07 2.9 0.09 0.1 0.7 0.1 0.7科 25.4 <0.001 20.7 <0.001
地域×在/外 326.5 <0.001 80.7 <0.001 1158.8 <0.001 1201.5 <0.001地域×在/外×科 104.3 <0.001
要因 在来昆虫 外来昆虫 グンバイ 全体χ² Pr χ² Pr χ² Pr χ² Pr
地域 6.7 0.01 280.4 <0.001 194.0 <0.001流域 93.1 <0.001 1076.1 <0.001 1020.4 <0.001 481.8 <0.001葉数 3.2 0.07 40.2 <0.001 244.7 <0.001 44.7 0.23在/外 1.5 0.2 6.9 0.009 12.2 <0.001 3.4 0.07科 3.4 0.06 43.0 <0.001 72.3 <0.001在来昆虫 8.8 0.003 11.1 0.0009 1132.2 <0.001外来昆虫 6.8 0.009 402.2 <0.001 3903.5 <0.001地域×在/外 19.3 <0.001 631.6 <0.001 597.1 <0.001
18
図 9 キク科在来植物1個体当たりの在来昆虫数(平均±SE)
図 10 キク科外来植物1個体当たりの外来昆虫数(平均±SE)
秋田 京都
秋田 京都

19
図 12 マメ科外来植物1個体当たりの外来昆虫数(平均±SE)
図 11 マメ科在来植物1個体当たりの在来昆虫数(平均±SE)
秋田 京都

20
図 13 キク科外来植物1個体当たりのグンバイ数(平均±SE)
京都
図 14 植物全体の1個体当たりの昆虫数(平均±SE)
秋田 京都

21
図 15 キク科植物1個体当たりのギルド別昆虫数(平均±SE) 秋田 京都
図 16 マメ科植物1個体当たりのギルド別昆虫数(平均±SE) 秋田 京都

22
図 17.1 産卵までの日数 図 17.2 孵化までの日数
空欄は卵ができなかったもの、または成虫に
ならなかったものを示す。
図 17.3 孵化した幼虫数 図 17.4 成虫数
(成虫数/幼虫数×100)
図 17.5 生存率(%)

23
表 6 キク科植物によるグンバイの成長の比較
表 7 グンバイの生育に及ぼす要因について GLMMで解析した結果
要因 産卵までの日数 孵化日数 幼虫数 成虫数 生存率χ² Pr χ² Pr χ² Pr χ² Pr χ² Pr
在/外 0.35 0.6 1.28 0.3 1.12 0.3 1.34 0.2 0.07 0.8
平均 産卵した個数 孵化した個数 産卵までの孵化日数 幼虫数 成虫数 生存率(%)セイタカアワダチソウ 10 10 6.4 22.6 7.9 6.4 82.3ヒメムカシヨモギ 2 2 7.0 23.0 5.5 2.5 27.8オオブタクサ 9 0 10.2 0 0 0 0オオハンゴンソウ 8 0 6.5 0 0 0 0アキノキリンソウ 10 6 11.6 17.5 7.2 4.8 69.3ノコンギク 10 9 4.9 23.6 9.1 7.9 86.1アズマギク 10 10 4.2 19.5 12.6 10.4 80.9シマカンギク 0 0 0 0 0 0 0





























