Embed Size (px)
Citation preview

system’s trivalent fullerene anions (C603−) was
found in the ‘paramagnetic’ phases in both the b.c.c. and f.c.c. forms. Such a low-spin state favours quantum spin fluctuation and suppresses magnetic ordering, and could be understood in terms of a geometrical distor-tion (either static or dynamic) of the C60
3− anion known as the Jahn–Teller effect, which has so far not been detected in trivalent fuller-enes. Further studies of the interplay between superconductivity, magnetic ordering and structural arrangement promise to deepen our understanding of the physics that underpins superconducting materials. ■
Yoshihiro Iwasa is at the Quantum-Phase Electronics Center, School of Engineering, University of Tokyo, Tokyo 113–8656, and at CERG, RIKEN, Wako 351-0198, Japan.e-mail: [email protected]
1. Jérome, D. et al. J. Phys. Lett. 41, 95–98 (1980).2. Ganin, A. Y. et al. Nature Mater. 7, 367–371 (2008).3. Ganin, A. Y. et al. Nature 466, 221–225 (2010).4. Hebard, A. F. Phys. Today 45, 26–32 (1992).5. Jérome, D. Science 252, 1509–1514 (1991).6. Kanoda, K. Hyperfine Interact. 104, 235–249 (1997).7. Iwasa, Y. & Takenobu, T. J. Phys. Condens. Matter 15,
R495–R519 (2003). 8. Takabayashi, Y. et al. Science 323, 1585–1590 (2009).9. Jeglič, P. et al. Phys. Rev. B 80, 195424 (2009).
cell BIology
Sensing tension Andrew D. Doyle and Kenneth M. Yamada
Measuring the tension forces at specific sites in living cells is technically challenging. Now, a fluorescent biosensor protein can be used to characterize dynamic local changes in tension in migrating cells.
Cells constantly use molecular force1–3. Co ordinating protrusion and retraction helps cells to migrate or invade, and by using contractile tension they can stably adhere to each other or to the extracellular matrix4. Cells must also orchestrate multiple intracellular forces in order to divide. It is, however, tech-nically difficult to measure and map these complex tension forces in a single cell. An ideal tool would be a miniaturized biological equivalent of a mechanical-force gauge that can be inserted into sites of local tension or movement. On page 263 of this issue, Grashoff et al.5 describe such a powerful tool to map forces in individual cells. Specifically, they have developed a ‘second-generation’ biosensor to measure tension accurately in dynamic cell-adhesion structures, and they show how local forces change substantially as cells migrate.
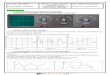
Grashoff and co-workers began by designing a tension-sensor module that contains a spring-like protein segment, based on a sequence from a spider silk protein, that stretches in response to tension. They then positioned fluor escent proteins at the ends of this spring-like pro-tein segment (Fig. 1a). Application of tension causes the sensor module to stretch, separ ating the fluorescent proteins and so leading to a readily detectable decrease in fluorescence (as measured by fluorescence resonance energy transfer; Fig. 1b). Grashoff et al. can thus make sensitive measurements of local tension (in the piconewton range) by using a single chimaeric molecule.
The authors inserted the fluorescent ten-sion-sensor module into vinculin, a protein that has force-related functions in cell ad hesion and migration. Vinculin is an excellent choice because its head domain binds to talin
— a protein that binds to cell-surface adhesion receptors, which interact with the extracellular matrix — and its tail domain binds to the actin cytoskeleton. Vinculin is therefore well posi-tioned to sense changes in tension between the interior and exterior of the cell. When vinculin is stretched between the actin cytoskeleton and cell-surface adhesion receptors, the tension-sensor module locates and measures changes in tension in living, migrating cells.
The biosensor has allowed Grashoff and colleagues to provide the first direct measure-ments of tension in focal adhesions — adhesive structures that are formed by cells migrating in tissue culture. The authors5 find that vinculin is under high tension in newly formed ad hesions at the front end of migrating cells, and that tension is decreased at the back of the cells as the ad hesions disassemble. This observation provides an elegant and direct quantification of the asymmetry of tension that was previ-ously proposed to occur in migrating cells. And, as expected, the tension on vinculin is lost when the contraction of actin filaments within cells, which is driven by the molecular motor myosin, is inhibited.
Another insight gained from using the bio sensor is that the recruitment of vinculin to adhesion sites and its activity are independent of its ability to transmit cellular forces. More-over, in the absence of vinculin, high forces result in the disassembly of cell adhesions, suggesting that vinculin helps to maintain the stability of ad hesions.
Grashoff and colleagues’ probe is not the first tension biosensor but rather the latest advance in biological force sensing. A conceptually related approach has been used6 to measure tension-mediated stretching of the
extracellular protein fibronectin. Other ten-sion biosensors have been designed to report differences in the forces on the actin-binding protein α-actinin at the front and back of migrating cells7 and to determine the levels of actin–myosin association in living cells8. Yet another tension biosensor has been expressed successfully in the nematode Caenorhabditis elegans7, suggesting that these approaches can be applied in vivo.
Some proteins have also been shown to have an intrinsic ability to sense and respond to force. For example, stretch activates certain ion channels in membranes, and force activates talin to bind to vinculin9 and the signalling protein p130Cas to promote downstream phos-phorylation and signalling10. What is special about the vinculin biosensor5, however, is that it provides the high sensitivity and accurate
High tensionLow fluorescence
Low tensionHigh fluorescence
b
a
TailHead
Vinculin
+
Vinculin-tension sensor
Extracellularmatrix
Cell membrane
Adhesionreceptor
CytoplasmTalin
Tension-sensor module
SSP
Increasedtension
Actin cytoskeleton
Figure 1 | Tension biosensor. a, To measure localized cellular forces, Grashoff et al.5 designed a tension-sensor module by using an elastic protein segment — 40 amino acids long and derived from the sequence of flagelliform, a spider silk protein (SSP) — flanked by two fluorescent proteins (blue and green). They inserted this sensor module between the head and tail domains of vinculin, a protein component of cell-adhesion structures. b, Increased cellular tension across the vinculin-tension sensor stretches the SSP region, resulting in decreased fluorescence as the two fluorescent proteins are pulled apart (as measured by fluorescence resonance energy transfer). In this way, tension is quantified by measuring changes in fluorescence. (Figure not to scale.)
192
NATURE|Vol 466|8 July 2010NEWS & VIEWS
© 20 Macmillan Publishers Limited. All rights reserved10

quantification needed for mapping local forces, as well as allowing in-depth studies of force-dependent biological pro cesses.
Molecular tension gauges such as the vinculin biosensor have the potential to define many cellular forces, including the forces that move and guide chromosomes as cells undergo mitotic division, the complex local forces on membranes as cells migrate and as the axonal projections of neurons extend, and the mechanical features of bone remodelling and tumour-cell invasion. But how practical will it be to use these tension-sensor modules as part of other proteins, at other cellular sites, or for measuring other types of intracellular force?
The answer might depend on whether protein candidates that have certain key features in common with vinculin can be found. The protein into which the tension-sensor module is inserted must be present at a cellular site where it can be stretched by the force being measured. The protein needs to retain its function after the sensor module has been inserted, and, conversely, the func-tion of the protein must not affect that of the sensor. If such an ideal protein cannot be found to carry the tension-sensor module, it might be possible to generate a different type of chimaeric force biosensor by positioning the module between two different proteins or between the binding sites of these proteins. Another advance would be to increase the sensitivity with which force can be detected; the current approach5 involves analysing hundreds of molecules at a particular site
rather than single-molecule force detection.An intriguing possibility is that tension
biosensors might be applied systematically to measure forces within intact tissues and ani-mals7: for example, within the extracellular matrix in vivo, between cells, between a cell and its surrounding connective tissue, in the establishment of the body plan of embryos, and during the shaping of the internal structures of developing organs. The ability to visualize and quantify local forces will help to bridge the gap between the extensive knowledge about tissue-specific expression of genes or proteins and the forces that mould a functional, living organism. ■
Andrew D. Doyle and Kenneth M. Yamada are in the Laboratory of Cell and Developmental Biology, National Institute of Dental and Craniofacial Research, National Institutes of Health, Bethesda, Maryland 20892, USA. e-mails: [email protected]; [email protected]
1. Mofrad, M. R. K. & Kamm, R. D. Cellular Mechanotransduction (Cambridge Univ. Press, 2009).
2. Vogel, V. & Sheetz, M. Nature Rev. Mol. Cell Biol. 7, 265–275 (2006).
3. Orr, A. W., Helmke, B. P., Blackman, B. R. & Schwartz, M. A. Dev. Cell 10, 11–20 (2006).
4. Ridley, A. J. et al. Science 302, 1704–1709 (2003).5. Grashoff, C. et al. Nature 466, 263–266 (2010).6. Klotzsch, E. et al. Proc. Natl Acad. Sci. USA 106,
18267–18272 (2009).7. Meng, F., Suchyna, T. M. & Sachs, F. FEBS J. 275,
3072–3087 (2008).8. Iwai, S. & Uyeda, T. Q. P. Proc. Natl Acad. Sci. USA 105,
16882–16887 (2008).9. del Rio, A. et al. Science 323, 638–641 (2009).10. Sawada, Y. et al. Cell 127, 1015–1026 (2006).
The complex organization of biological func-tions into precise regions of space and time is essential for the functioning of living systems. Yet our understanding of the way in which small molecules self-assemble to form struc-tures that contain different kinds of compart-ments, and our ability to achieve this in the lab, are still mostly lacking. Chemists are, however, starting to unravel the molecular design rules that underpin such self-assembly, as reported by Pal et al.1 in the Journal of the American Chemical Society. They have prepared mol-ecules that sort themselves, when mixed together in water, so that distinct types of mol-ecules self-assemble into different domains.
Compartmentalization is one of the pre-requisites for the emergence of life. The cell membrane, for example, keeps intracellular
components together and shields them from a hostile environment. Further compartmen-talization within the cell separates otherwise incompatible reactions and biological func-tions. Scientists have long been intrigued by nature’s ability to achieve multiple levels of compartmentalization — from nano metre to micrometre scales and even larger — starting from the self-assembly of molecular compo-nents, and this has motivated them to inves-tigate how compartmentalization can be achieved using artificial molecules.
Such investigations have revealed that sim-ple micelles — aggregates of molecules — form in water from small surfactant molecules, and that vesicles — membrane-enclosed bags — form from lipids. But such assemblies comprise at best only a single hydrophobic
SUPrAmolecUlAr chemISTry
more than the sum of its partsJan H. van Esch
Can small molecules in test tubes form assemblies containing different hydrophobic domains, like those found in cells? Yes, finds a study, suggesting new ways of isolating incompatible compounds in water.
compartment and a single aqueous compart-ment. Architectures containing several differ-ent compartments have mainly been obtained by the self-assembly of polymer mixtures, and especially of block copolymers, the molecules of which consist of two or more chemically distinct polymer chains connected together2. When polymer mixtures or block copolymers are driven to assemble, chemically similar regions of the molecules aggregate into macro- or microdomains. This phase separation is thermodynamically favoured because of the small entropy of mixing. By contrast, mixing together two different small surfactant mol-ecules generally leads to the formation of mixed assemblies containing both types of molecule, because of the much larger entropy of mixing of small molecular systems compared with polymeric systems3.
Pal et al.1 show that this classical view needs to be revised, an insight that opens the way to the fabrication of much more complicated, multi-compartment architectures from self-assembling small molecules. The authors have studied the self-assembly of small amphiphiles — molecules that have both hydrophobic and hydrophilic parts. These particular amphiphiles contain two additional hydrogen-bonding groups within their hydrophobic domain. Pal et al. find that, in water, the amphiphiles aggre-gate into rod-like micellar assemblies because of a combination of hydrogen bonding and hydrophobic interactions.
Most interestingly, the authors find that the distance between the hydrogen-bonding groups can be used to control the aggregation behaviour of mixtures of their amphiphiles. When they mixed together two or three dif-ferent analogues of the amphiphiles in which the distance between the hydrogen-bonding groups was the same, the analogues assembled with each other to form mixed micelles (Fig. 1, overleaf). But if the number of carbon atoms in the hydrocarbon chain between the hydrogen-bonding groups differed by as little as one atom, the different amphiphiles aggregated separately to form distinct hydrophobic domains. Similar behaviour has been observed in self-assembled monolayers of molecules on surfaces4, but Pal and colleagues’ work goes much further, by demonstrating that the same principle can create two or more kinds of hydrophobic compartments that coexist in water.
The formation of distinct hydrophobic domains in water is well known, for example in biological membranes, in which domains called lipid rafts can be seen5. Lipid rafts are important for many processes, including cel-lular signalling events, but the way in which their component molecules become organized is still unclear. Pal and colleagues’ system1 is a good example of molecular self-sorting6, in which differences in interactions between the constituent molecules of a mixture create a thermodynamic impetus for the formation of separate domains.
As pointed out by the authors1 and others6,
193
NATURE|Vol 466|8 July 2010 NEWS & VIEWS
© 20 Macmillan Publishers Limited. All rights reserved10