Embed Size (px)
Citation preview

Cell Death and Survival
TDP1 and PARP1 Deficiency Are Cytotoxic toRhabdomyosarcoma Cells
Hok Khim Fam1,2�, Cheryl Walton1,2�, Sheetal A. Mitra3
�, Miraj Chowdhury1,2
�, Nichola Osborne1,2, Kunho
Choi1,2, Guobin Sun1,4, Patrick C.W.Wong1,5, Maureen J. O'Sullivan5,6,7, Gulisa Turashvili8, Samuel Aparicio8,Timothy J. Triche3, Mason Bond1,4, Catherine J. Pallen1,4, and Cornelius F. Boerkoel1,2
AbstractRhabdomyosarcoma is the most common soft tissue sarcoma in children. Metastatic rhabdomyosarcoma in
children has a 5-year event-free survival rate of <30%, and a recent clinical trial with irinotecan, a topoisomeraseI inhibitor, failed to improve outcome. Therefore, it was surmised that failure of irinotecan may be the result of over-expression of the DNA repair enzyme tyrosyl-DNA phosphodiesterase (TDP1), which processes topoisomeraseI-DNA complexes resulting from topoisomerase I inhibitor treatment. Using human tissue microarrays and geneexpression arrays, a marked overexpression of TDP1 protein and mRNA in RMS tumors was observed. Critically,knockdown of TDP1 or inhibition of poly(ADP-ribose) polymerase-1 (PARP-1), an enzyme in the same complex asTDP1, sensitized rhabdomyosarcoma cell lines to analogues of irinotecan. Interestingly, BRCA1/2mutations or alteredexpression was not detectable in rhabdomyosarcoma cells; however, TDP1 knockdown and PARP-1 inhibition alonewere cytotoxic to a subset of rhabdomyosarcoma cells, suggesting that they harbor genetic lesions in DNA repaircomponents that have synthetic lethal interactions with loss of TDP1 or PARP1 function. Furthermore, culturingembryonal rhabdomyosarcoma cells in serum/nutrient–restrictedmedium increased cellular cytotoxicity uponPARP-1inhibition and was intrinsically cytotoxic to alveolar, though not embryonal rhabdomyosarcoma cells. The results ofthese studies suggest a compensatory role for TDP1 in rhabdomyosarcoma after topoisomerase-I based therapy andfurther demonstrate that TDP1 knockdown, PARP-1 inhibition, and dietary restriction have therapeutic validity.
Implications: Selective targeting of TDP1 and/or PARP-1 in rhabdomyosarcoma induces cytotoxicity andsensitizes to DNA damaging agents. Mol Cancer Res; 11(10); 1179–92. �2013 AACR.
IntroductionConventional chemotherapeutic agents have targeted gen-
eral cellular processes such as nucleotide synthesis, DNAreplication, and cell division. Because each of these processes
is required for homeostasis of normal tissues, these treat-ments cause significant morbidity and limit the doses ofdrugs that can be given for a cure. Despite these short-comings, combinations of cytotoxic agents have improvedcure rates for many malignancies such as pediatric acutelymphoblastic leukemia (1).Pediatric sarcomas and other cancers have not responded
as well to these conventional therapies (2). Improvement incancer-free survival therefore requires a better understandingof cancer biology and the identification of cancer-specifictargets (3). During the evolution of human cancers, cancercells accumulate genetic alterations such that each tumorcontains multiple genetic and epigenetic abnormalities (4).As illustrated by imatinib inhibition of BCR-ABL in chronicmyelogenous leukemia (5), the inhibition of a single onco-gene is sufficient to block cancer cell growth in some cancers.In other cancers, DNA maintenance pathways mutate dur-ing the evolution of the cancer (6) and inhibition of remain-ing DNA repair pathways causes cancer-cell death. This isillustrated by the synthetic lethal interaction of PARP-1inhibitors with deficits of homologous recombination suchas those arising from mutations of BRCA1 and BRCA2 (7).Sparing normal tissues, these 2 approaches show the poten-tial for preferential targeting of malignant cells.
Authors' Affiliations: 1Child and Family Research Institute; and 2Depart-ment of Medical Genetics, University of British Columbia, Vancouver,Canada; 3Department of Pathology, Children's Hospital Los Angeles, LosAngeles, CA; 4Department of Pediatrics, University of British Columbia,Vancouver, Canada; 5Department of Pathology and Laboratory Medicine,University of British Columbia, Vancouver, Canada; †6National Children'sResearchCentre andHistology Laboratory, Our Lady'sChildren’sHospital,Crumlin, Dublin; †7Trinity College Dublin, Ireland; 8Molecular Oncology andBreast Cancer Program, BC Cancer Research Centre, Vancouver, BC,Canada
Note: Supplementary data for this article are available at Molecular CancerResearch Online (http://mcr.aacrjournals.org/).
H.K. Fam, C. Walton, S.A. Mitra, and M. Chowdhury contributed equally tothis work.
Corresponding Author: Cornelius F. Boerkoel, University of British Colum-bia, 4500 Oak St., Room. C234, Vancouver, BC V6H 3N1, Canada. Phone:604-875-2157; Fax: 604-875-2376; E-mail: [email protected]/[email protected]
doi: 10.1158/1541-7786.MCR-12-0575
�2013 American Association for Cancer Research.
MolecularCancer
Research
www.aacrjournals.org 1179
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 2, 2013; DOI: 10.1158/1541-7786.MCR-12-0575

The question then arises as to whether such targets mightbe identifiable for the high-risk pediatric sarcomas (2).Rhabdomyosarcoma is one of the most common soft tissuesarcomas of childhood. Each year approximately 300 chil-dren in the United States are diagnosed with this disease andtwo-thirds are younger than 10 years old (8). The 2 majorsubtypes are embryonal and the more aggresive alveolarrhabdomyosarcoma (8). Although the overall cure increasedfrom 25% to 70% between the 1970s and the 1990s (8, 9),the 5-year failure-free survival rate is only 65% for childrenwith nonmetastatic alveolar rhabdomyosarcoma or embry-onal rhabdomyosarcoma at unfavorable sites (9). Survival iseven less for the approximately 30% of children presentingwith metastatic rhabdomyosarcoma (10).The genetic alterations associated with rhabdomyosarco-
ma include chromosomal translocations and deletions andthese correlate with outcome.Many embryonal rhabdomyo-sarcoma tumors have loss of heterozygosity at the 11p15.5locus (11) and 70% to 80% of alveolar rhabdomyosarcomahave chromosomal translocations t(2;13)(q35;q14) or t(1;13)(p36;q14) resulting in expression of PAX3/7-FOXO1chimeric proteins that function as transcription factors (12,13). The PAX3-FOXO1 transcription factor increasesexpression of the insulin growth factor 1 receptor (IGF-IR; 14) and inhibition of IGF-IR signaling shows promise inimpeding rhabdomyosarcoma tumor growth (15).Unlike recent studies implicating a role for the EWS–FLI
fusion protein in maintaining the PARP-1–mediated DNAdamage response in Ewing sarcoma, defects of DNA repairhave not been reported in rhabdomyosarcoma despite thegenomic rearrangements identified in these tumors (16, 17).Of particular interest is whether the recent failure of theirinotecan trials for metastatic or recurrent rhabdomyosarco-ma is attributable to compensatory increased expression of theDNA repair enzyme tyrosyl-DNA phosphodiesterase(Tdp1); resistance of some cancers to topoisomerase-I inhi-bitors has been attributed to overexpression ofTdp1 (18–21).Such overexpression might also suggest that rhabdomyosar-coma tumors have defective DNA double-strand break repairas expression of the Tdp1-PARP-1 complex is increased inother cancers with such deficits (22–24). To assess whetherTdp1 and PARP-1 might be targets for treatment of rhab-domyosarcoma,we profiled the expression ofTdp1 in humanpediatric tumors and analyzed the effect of topoisomerase-Iinhibition, TDP1 knockdown, PARP-1 inhibition, andnutrient restriction on rhabdomyosarcoma cell lines.
Materials and MethodsHuman subjectsHuman tissue andmaterial use for this study was approved
by the Institutional ReviewBoard of theUniversity of BritishColumbia (Vancouver, BC; H09-03301).
TMA constructionWe assembled pediatric tumor samples from cases referred
to BC Children's Hospital (Vancouver, BC) into 4 tissuemicroarray (TMA) blocks. The tumors that were readable(vs. total number of tumor samples) included alveolar
rhabdomyosarcoma (n ¼ 18/21), embryonal rhabdomyo-sarcoma (n ¼ 24/25), ganglioneuroma (n ¼ 12/14), neu-roblastoma (n ¼ 23/30), Ewing sarcoma (n ¼ 10/22),medulloblastoma (n ¼ 9/14), and Wilms tumor (n ¼24/24). Of the 46 rhabdomyosarcoma tumors, 14 weremetastatic. Molecular diagnosis conducted on 31 of 46 rhab-domyosarcoma tumors identified 18 fusion-positive and 13fusion-negative tumors. Hematoxylin–eosin images of allcores are provided online at http://www.cfri.ca/pallenlab/Additional TMAs were obtained from the Children's
Oncology Group (COG). These included 34 embryonalrhabdomyosarcoma and 39 alveolar rhabdomyosarcomatumors derived from individuals enrolled in protocols9602, 9150, D9902, D9802, and D9803.
ImmunohistochemistryTheTMAswere screened for Tdp1 expression according to
standard immunohistochemistry protocols. Briefly, 4 mmthick sections were cut from each block and immunostainedon aVentanaDiscoveryXT staining system (VentanaMedicalSystems). Sections were deparaffinized in xylene, dehydratedthrough 3 alcohol changes and transferred to Ventana WashSolution. Endogenous peroxidase activity was blocked in3% hydrogen peroxide. Antigen retrieval was conducted inVentana CC1 buffer. Slides were incubated with anti-Tdp1rabbit antibody (dilution1:200) for 32minutes anddevelopedin DAB for 10 minutes. Tissue sections were then counter-stained with hematoxylin and mounted. Preimmune serumwas used as a negative control in place of immune serum.Three independent observers scored Tdp1 expression.
Any discrepancies between the 3 scores were resolved byC.F. Boerkoel. Images of each tumor were acquired using aZeiss Axiovert 200microscope, a Zeiss AxiocamHR camera,and the Zeiss Axiovision imaging system.
Gene expression arraysTranscriptome-profiling data on primary rhabdomyosar-
coma tumors and 7 normal skeletal muscle specimensgenerated using Human Genome U133A Arrays (Affyme-trix) were obtained from the NCI caArray (trich-00099) andGEO (GSE873, GSE1462) databases. From the available185 samples in the caArray database, expression data on 147primary rhabdomyosarcoma were selected for analysis afterconfirmation of their diagnosis and PAX-FOXO1 translo-cation status. In addition, expression data on 5 randomlyselected samples each of non-rhabdomyosarcoma soft tissuesarcoma (NRSTS), osteosarcoma (OS), and Ewing sarcoma(ES) were obtained from GEO (GSE16088, GSE37372)and NCI caArray (trich-00099) databases. Tdp1 expressionwas compared among specimens using Partek GenomicsSuite Software (Partek) after quantile normalization byrobust multichip averaging and median summarization ofraw data using one-way ANOVA.
Cell cultureRH30 (alveolar rhabdomyosarcoma, PAX3-FOXO1) and
CW9019 (alveolar rhabdomyosarcoma, PAX7-FOXO1)cells were cultured in Dulbecco's modified Eagle medium
Fam et al.
Mol Cancer Res; 11(10) October 2013 Molecular Cancer Research1180
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 2, 2013; DOI: 10.1158/1541-7786.MCR-12-0575

(DMEM; Gibco BRL Life Technologies) supplementedwith 10% heat-inactivated FBS (Hyclone) and 1% anti-biotic–antimycotic (Gibco BRL Life Technologies). RD(embryonal rhabdomyosarcoma) cells were grown inDMEMsupplemented with 10% heat-inactivated FBS, 1% antibi-otic–antimycotic, 4 mL/L L-glutamine (Gibco BRL LifeTechnologies), 4.5 g/L glucose, and 1.5 g/L NaHCO3.Human skeletal myoblast cells were cultured using skBM-2 (Lonza) supplemented with 15% FBS, 1% antibiotic–antimycotic and 9 g/L D-glucose (final concentration 10 g/Lglucose). A204 (embryonal rhabdomyosarcoma) and Birch(embryonal rhabdomyosarcoma) cells were grown in RPMI-1640 media (Gibco BRL Life Technologies) supplementedwith 10% FBS and 1% antibiotic–antimycotic. Rhabdo-myosarcoma cell lines were procured from the LeibnizInstitute DSMZ-German Collection of Microorganismsand Cell Cultures or the American Type Culture Collection.The human skeletal muscle cell-line was obtained fromLonza (XM13A1). All cells were grown at 37�C with 5%CO2 in a humidified environment. Human skeletal musclecells under nutrient-restricted conditions were cultured inskBM-2 supplemented with 1% heat-inactivated FBS andcontaining 1 g/L D-glucose. RH30 and CW9019 cells undernutrient-restricted conditions were cultured in glucose-freeDMEM supplemented with 1% heat-inactivated FBS and1.0 g/L glucose. A204 and Birch cells under nutrient-restricted conditions were cultured in glucose-free RPMIsupplemented with 1% heat-inactivated FBS and 1.0g/Lglucose.
Anti-Tdp1 serum productionAn anti-human Tdp1 serum was generated in rabbits
against amino acids 1–152 of the human Tdp1 protein aspreviously described (25). The specificity of the antiserumwas confirmed by immunofluorescent comparison withTdp1þ/þ and Tdp1�/� mouse skeletal muscle (Supplemen-tary Fig. S1).
ImmunofluorescenceImmunofluorescence was conducted as described by Hir-
ano and colleagues (25). Rabbit anti-Tdp1 and mouse anti-a-tubulin (DM1A, Sigma) were diluted 1:100 and 1:400,respectively. Images were acquired using a Zeiss Axiovert200 microscope, a Zeiss AxiocamMR camera, and the ZeissAxiovision imaging system.
Immunoblot analysisImmunoblotting was conducted as previously described
(25). Antibodies used were anti-human TDP1 (1:1,000),mouse anti-GAPDH monoclonal antibody (1:5,000; Che-micon), mouse anti-a-tubulin (1:1,000; Abcam), rabbitanti-histone H2B (1:1,000; Upstate), rabbit anti-PARP-1(1:400, Abcam), mouse anti-Poly-ADP Ribosyl Polymer(PAR; 1:1000; Abcam) and rabbit anti-caspase-3 (1:1,000,Cell signaling).
qRT-PCRRNA was isolated from each cell line using the Qiagen
RNeasyMini Kit (Qiagen) and 3 mg were reverse transcribed
with qScript cDNA supermix (Quanta Biosciences). Quan-titative PCR was then done using PerfeCTa SybergreenMix(Quanta Biosciences) and the 7500 Applied BiosystemsqPCR machine. Data was analyzed using the 7500 software(v2.0.1). The sequences of the primers used for qRT-PCRare listed in Supplementary Table S1.
MTT assayCell proliferation and viability were measured by MTT
assay as previously described (26). Briefly, 5� 103 cells wereplated in each well of a 96-well plate and cultured in phenolred-free media. MTT solution was prepared by dissolving 5mg of MTT in 1 mL of 1� PBS. Following the describedtreatments, the culture media was supplemented with 10%MTT and incubated for 3 hours. This was followed by a 30-minute incubation in 100% dimethyl sulfoxide (DMSO).Spectrophotometry was done at 565 nm using the WallacVICTOR2 Multilabel Plate Reader (Beckman–Coulter).
TUNEL assayBriefly, 5� 103 cells were seeded per well in 6-well plates
and following the described treatments, apoptotic cells werevisualized by colorimetric labeling of free 30OH DNAtermini using the Apoptag Plus Peroxidase In Situ ApoptosisDetection Kit according to the manufacturer's protocol(EMD Millipore). The number of stained nuclei per 100total cells was determined for 3 independent replicates toquantify apoptosis.
Homogeneous caspase-3/7 assayThe activity of caspases 3 and 7, which are key effectors
of mammalian cell apoptosis, was measured using theApo-ONE Homogeneous Caspase-3/7 Kit according to themanufacturer's protocol (Promega). Briefly, 5 � 103 cellswere plated in each well of a 96-well plate and cultured for24 hours and following the described treatments, fluor-escence of the cleaved caspase-3, and -7 substrate wasmeasured at 520 nmusing theWallac VICTOR2MultilabelPlate Reader (Beckman–Coulter).
Comet assayDNA damage was assessed by the alkaline comet assay as
previously described (25). This assay is unable to distinguishbetween single-stranded versus double-stranded breaksbecause the alkaline conditions used also detect single-stranded break sites and convert alkali–labile apurinic andapyrimidinic sites to DNA breaks. As Tdp1 also participatesin the repair of apyrimidinic sites (27), the alkaline conver-sion of these sites to single-stranded breaks allows a greaterappreciation of the function of Tdp1. Briefly, rhabdomyo-sarcoma cell lines were seeded at a density of 3� 105 cells in a6-well (9.6 cm2) plate. After 24 hours, the cells were treatedfor 2 hours (37�C, 5%CO2) with 4 mmol/L, 8 mmol/L, and16 mmol/L camptothecin (CPT; Sigma), harvested by tryp-sinization, resuspended in 80 mL of 0.5% low melting pointagarose and pipetted onto microscope slides precoated with1% agarose. After addition of a second layer of 1% normalmelting point agarose, the cells were immersed in lysis
TDP1 and PARP-1 in Rhabdomyosarcoma
www.aacrjournals.org Mol Cancer Res; 11(10) October 2013 1181
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 2, 2013; DOI: 10.1158/1541-7786.MCR-12-0575

solution (2.5 M NaCl, 100 mmol/L EDTA, 10 mmol/LTrizma base, 1% Triton X-100, 10% DMSO) for 2 hours.The DNA was then denatured by incubation at 4�C inalkaline buffer (50 mmol/L NaOH, 1 mmol/L EDTA, 1%DMSO, pH > 13) for 25 minutes. Following electrophoresisat 25V and 4�C for 25minutes, the slides were neutralized in0.4MTris-HCl and stained with SYBR-Green (Invitrogen).Images were acquired using a Zeiss Axiovert 200microscope,a Zeiss AxiocamMRcamera and theZeiss Axiovision imagingsystem. A total of 100 comets were scored for each sampleusing theComet Assay IV Software (Perceptive Instruments).
TDP1 knockdownTDP1 was knocked down transiently by transfecting 8�
104 cells with 100 nmol/L of pooled siRNAs (Dharmacon)in a 24-well plate using Lipofectamine 2000 (Invitrogen).Knockdown was confirmed by immunoblotting and qRT-PCR. The sequences of the siRNAs are listed in Supple-mental Table S1.For stable knockdown of TDP1, 1 to 2 � 104 cells were
plated in each well of a 96-well plate and cultured overnight(37�C, 5%CO2). The culture mediumwas replaced for 4 to18 hours with serum-free culture medium, 3 mg/mL poly-brene and lentivirus carrying the SMARTvector 2.0 shRNA(Dharmacon). The infection was carried out at a multiplicityof infection (MOI) of 0.3, 1, 2, or 5. Stably infected cellswere selected with 0.5 mg/mL puromycin and ultimately theMOI that gave the best knockdown was used for each cellline. Knockdown of TDP1 was verified by immunoblottingand qRT-PCR. The sequences of the shRNAs are listed inSupplemental Table S2.
PARP inhibitionPARP inhibitors AG-014699 (Rucaparib), AZD-2281
(Olaparib), and ABT-888 (Veliparib) were obtained fromSelleck Chemicals. The LD50 for each inhibitor was deter-mined by seeding each wild-type rhabdomyosarcoma cellline at 5 � 103 cells per well in 96-well microtiter plates.After 24 hours, the PARP inhibitors were added to theculture medium to a final concentration of 0.1 mmol/L, 0.5mmol/L, 1 mmol/L, or 10 mmol/L. Cell viability and pro-liferation was measured in triplicate every 24 hours for 96hours by the MTT assay.
Tdp1 cleavage assayCells were lysed and their nuclear contents extracted as
previously described (28). Briefly, cells were pelleted andlysed in 1 mL of lysis buffer. The cells were placed on ice for10 minutes and vortexed every 2 minutes to allow completelysis. The nuclei were pelleted for 4 minutes at 2,000 rpm.Following removal of the supernatant, 3 times the volume ofthe pellet of nuclear extraction buffer was added. The pelletwas resuspended and incubated at 4�C for one hour to isolatenuclear proteins. The lysate was clarified for 10 minutes at11,000 rpm to obtain nuclear extracts. The supernatantcontaining nuclear proteins were diluted 1:10 in reactionbuffer (10 mmol/L Tris-HCl, 50 mmol/L KCl, 1 mmol/LEDTA, 1mmol/L DTT, 0.01% Brij-35, and pH 7.5).
Tdp1 enzymatic activity was determined by cleavage ofan artificial substrate (Dean and colleagues; manuscript inpreparation). Briefly, the assay was run at room temperature,in 96-well format with a final volume of 100 mL per well. Afinal concentration of 50 nmol/L DNA substrate was used(50-/56-TAMN/AGGATCTAAAAGACTT/3BHQ_1/-30).Kinetic analysis was conducted using a Varioskan PlateReader (ThermoScientific) at Ex557/Em582 for theTAMRAfluorophore. To establish Tdp1 cleavage activity, 20 readswere recorded at a kinetic interval of 45 seconds per read. A2.5 nmol/L of purified Tdp1 was used in the control samples.Purified Tdp1 was isolated as previously described.
Statistical analysisMicrosoft Excel was used to compute the group means
and SDs for all treatment and control groups from cellviability data derived from theMTT, caspase, and apoptosisassays. The GraphPad Prism software was used for two-wayANOVA to compare the differences in growth and viabilityunder varying treatment conditions over time. The F-ratiowas used to test the validity of interactions between treat-ment groups over the course of the experiment. Statisticalsignificance between individual groups was established bythe Student t test and a P value of less than 0.05 was judgedsignificant. The Bonferroni correction was applied to allcomparisons of replicate means for the duration of theexperiments.
Sequencing of TDP1, BRCA1 and BRCA2Genomic DNA was isolated from rhabdomyosarcoma cell
lines using the Gentra Puregene Blood Kit (Qiagen). RNAwas extracted from rhabdomyosarcoma cells using theQiagenRNeasy Kit (Qiagen) and cDNA synthesized using theqScript cDNA SuperMix. PCR amplification of the TDP1coding exons was conducted as previously described (29).PCR amplification of the BRCA1 cDNA and of the BRCA2exons was conducted using the primers listed in Supplemen-tary Table S4 and HotStar Taq Plus (Qiagen) with thefollowing conditions: Denaturation stage: 95�C for 5 min-utes, cycling stage (40 cycles): 95�C for 30 seconds, 57�C for30 seconds, 72�C for 1minute. End stage: 72�C, 10minutes,hold at 4�C. All amplicons were sequenced at Macrogen andthe sequences were aligned and analyzed using Sequencher4.3 (Gene Codes).
ResultsTdp1 is highly expressed in rhabdomyosarcoma tumorsTo determine whether Tdp1 is expressed in rhabdomyo-
sarcoma and other pediatric solid tumors, we immunohis-tochemically screened a pediatric tumor microarray consist-ing of 24 embryonal rhabdomyosarcoma, 18 alveolar rhab-domyosarcoma, 12 ganglioneuroma, 23 neuroblastoma, 10Ewing sarcoma, 9 medulloblastoma, and 24 Wilms tumorreadable tissue cores. This showed that Tdp1 was expressedwith the highest frequency in rhabdomyosarcoma tumors(Supplementary Fig. S2). To confirm this expression inrhabdomyosarcoma tumors, we obtained 34 embryonalrhabdomyosarcoma and 39 alveolar rhabdomyosarcoma
Fam et al.
Mol Cancer Res; 11(10) October 2013 Molecular Cancer Research1182
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 2, 2013; DOI: 10.1158/1541-7786.MCR-12-0575

tumors from the COG and observed Tdp1 expressionrespectively in 97% and 100% of the tumors. In comparisonwith adjacent noncancerous tissue, Tdp1 also seemed tohave increased expression in the cancer cells (Fig. 1A–C).To confirm TDP1 mRNA expression in rhabdomyosar-
coma tumors, we screened an independent soft tissuesarcoma cohort that included 20 rhabdomyosarcoma (spe-cifically, 10 PAX-FKHR–positive aRMS, 5 PAX-FKHR–negative aRMS, and 5 eRMS), 5 NRSTS, 5 osteosarcomas,and 5 Ewing sarcomas by Affymetrix expression arrays. Thisanalysis revealed that TDP1 mRNA was expressed in allsoft-tissue sarcomas (Fig. 1D). The mean (� SD)-normal-ized microarray expression levels for these soft-tissue sar-
comas were 189.6 (�31.9), 161.5 (�32.2), 164.9 (�38.1),and 148.7 (�16.0), respectively. TDP1 expression levels inrhabdomyosarcoma were significantly higher comparedwith the other sarcomas in the group (ANOVA P¼ 0.041).To investigate further the specific pattern ofTDP1 expres-
sion in rhabdomyosarcoma, transcriptome-profiling data on147 primary tumors were compared with that of normalskeletal muscle. This analysis indicated that Tdp1 wassignificantly overexpressed in all primary rhabdomyosarco-ma samples compared with normal skeletal muscle (Supple-mentary Table S2; Fig. 1E). In addition, themean expressionlevel in fusion-positive rhabdomyosarcoma tumors was sig-nificantly more than in fusion-negative tumors (233.1 vs.
Figure 1. Tdp1 expression inrhabdomyosarcoma (RMS)tumors. Photomicrographsshowing immunohistochemicaldetection of Tdp1 in alveolar RMS(aRMS; A), embryonal RMS (eRMS;B), and unaffected human skeletalmuscle tissue (C). Scale bar ¼ 10mm.D, graph of the TDP1 transcriptintensity in soft-tissue sarcomas asmeasured by Affymetrix array.NRSTS,Non-RhabdomyosarcomaSoft Tissue Sarcoma; OS,osteosarcoma; and ES, Ewingsarcoma. E, Tukey box plotsrepresenting Tdp1 expressionintensities across 7 normal skeletalmuscle samples, 44 PAX3-FKHRRMS, 17 PAX7-FKHR RMS, 9fusion-negative RMS, 69 eRMS,and 8 spindle RMS. The boxesrepresent median with interquartilerange; whiskers extend 1.5 timesthe interquartile distance or to thehighest or lowest point, whicheveris shorter. Dots represent outliers.
TDP1 and PARP-1 in Rhabdomyosarcoma
www.aacrjournals.org Mol Cancer Res; 11(10) October 2013 1183
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 2, 2013; DOI: 10.1158/1541-7786.MCR-12-0575
192.2; Mann–Whitney P < 0.001, data not shown). Thus,although TDP1 is highly expressed in rhabdomyosarcomain general (P < 0.001), it is most significantly overexpres-sed in fusion-positive rhabdomyosarcoma (P < 0.001) com-pared to normal skeletal muscle. This suggests a cor-relation between rhabdomyosarcoma subtypes and TDP1expression.
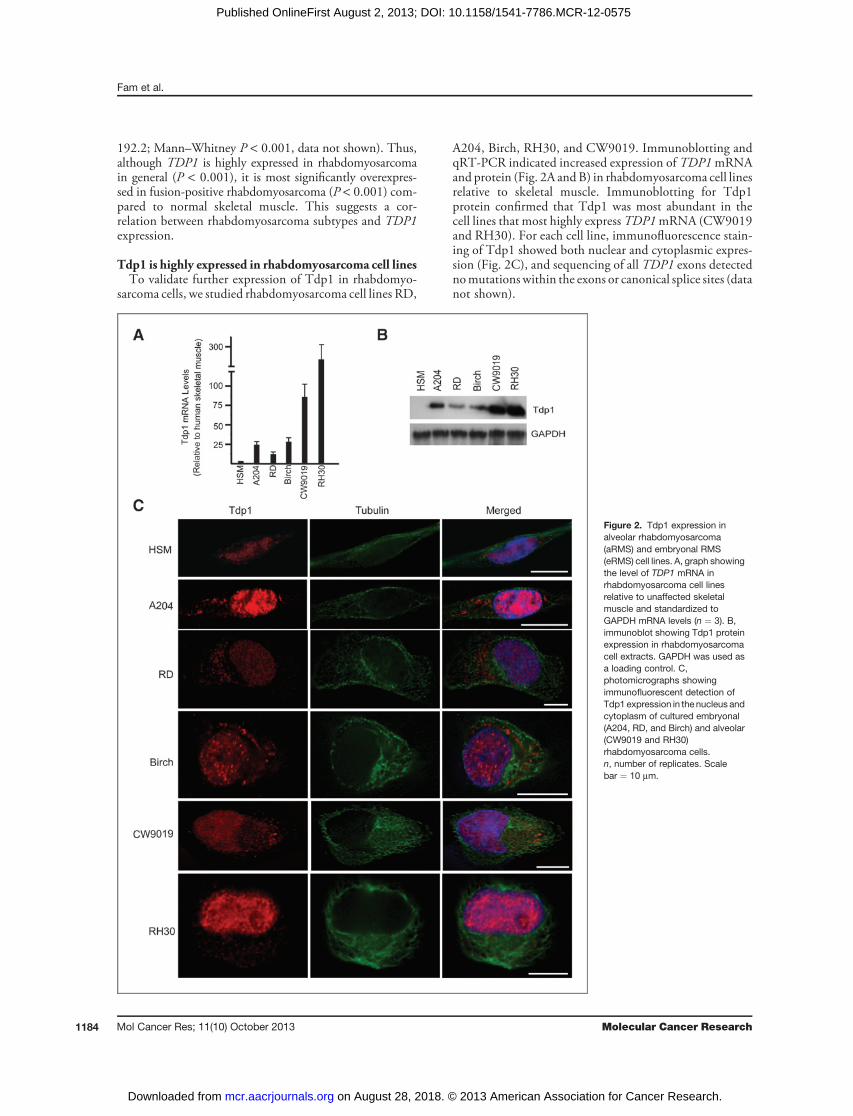
Tdp1 is highly expressed in rhabdomyosarcoma cell linesTo validate further expression of Tdp1 in rhabdomyo-
sarcoma cells, we studied rhabdomyosarcoma cell lines RD,
A204, Birch, RH30, and CW9019. Immunoblotting andqRT-PCR indicated increased expression of TDP1mRNAand protein (Fig. 2A and B) in rhabdomyosarcoma cell linesrelative to skeletal muscle. Immunoblotting for Tdp1protein confirmed that Tdp1 was most abundant in thecell lines that most highly express TDP1mRNA (CW9019and RH30). For each cell line, immunofluorescence stain-ing of Tdp1 showed both nuclear and cytoplasmic expres-sion (Fig. 2C), and sequencing of all TDP1 exons detectednomutations within the exons or canonical splice sites (datanot shown).
Figure 2. Tdp1 expression inalveolar rhabdomyosarcoma(aRMS) and embryonal RMS(eRMS) cell lines. A, graph showingthe level of TDP1 mRNA inrhabdomyosarcoma cell linesrelative to unaffected skeletalmuscle and standardized toGAPDH mRNA levels (n ¼ 3). B,immunoblot showing Tdp1 proteinexpression in rhabdomyosarcomacell extracts. GAPDH was used asa loading control. C,photomicrographs showingimmunofluorescent detection ofTdp1 expression in the nucleus andcytoplasm of cultured embryonal(A204, RD, and Birch) and alveolar(CW9019 and RH30)rhabdomyosarcoma cells.n, number of replicates. Scalebar ¼ 10 mm.
Fam et al.
Mol Cancer Res; 11(10) October 2013 Molecular Cancer Research1184
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 2, 2013; DOI: 10.1158/1541-7786.MCR-12-0575

TDP1 knockdown preferentially increases CPTsensitivity of rhabdomyosarcoma cell lines comparedwith control skeletal myoblastsAs Tdp1 expression is elevated in rhabdomyosarcoma
cells relative to skeletal muscle, this might explain theunresponsiveness of rhabdomyosarcoma to treatment withthe irinotecan, an analogue of camptothecin (CPT; ref. 18).
Consistent with this, siRNA knockdown of TDP1 by 70%to 85% decreased the viability of all CPT-treated rhabdo-myosarcoma cell lines except RD (Supplementary Fig. S3Aand Fig. 3A). In contrast, siRNA TDP1 knockdown ofnearly 90% (Supplementary Fig. S3A) did not significantlydecrease the viability of CPT-treated control skeletal myo-blasts. As complete loss of functional Tdp1 dramatically
0 48 960
200
400
600
800
Hours
Control
CPT (1 µmol/L)
Tdp1 KD + CPT (1 µmol/L)
0 48 960
200
400
600
800
0 48 960
200
400
600
800
0 48 960
200
400
600
800
0 48 960
200
400
600
800
Hours
0 48 960
200
400
600
800
A204
Birch RH30
RD
CW9019
HSM
A204
Birch RH30
RD
CW9019
HSM
A
B
******
******
****** ***
***
******
***
***
**
Contro
l
TDP1 KD
CPT
TDP1 KD +
CPT
Tai
l mom
ent
0
2
4
6
Tai
l mom
ent
0
2
4
6
Tai
l mom
ent
0
2
4
6
0
2
4
6
0
2
4
6
0
2
4
6
Contro
l
TDP1 KD
CPT
TDP1 KD +
CPT
Rel
ativ
e ce
ll vi
abili
ty (
%)
Rel
ativ
e ce
ll vi
abili
ty (
%)
Rel
ativ
e ce
ll vi
abili
ty (
%)
Figure 3. TDP1 knockdownincreases the sensitivity ofrhabdomyosarcoma cell lines toCPT treatment. A, graphs showingthe percent proliferation ofrhabdomyosarcoma cells andcontrol myoblasts over a 96-hourperiod during which the cells (i)received no CPT, (ii) were treatedwith 1 mmol/L CPT, or (iii) weretreated with 1 mmol/L CPT aftertransient TDP1 knockdown (n¼ 4).B, graphs showing the results of analkaline comet assay, whichanalyzes the number of DNAbreaks (tail moment). The assaywas performed after 2 hours ofculture under the conditions statedabove. Each dot represents asingle cell/comet. A total of 100cells were scored for eachtreatment. Error bars, mean � SD;��, P < 0.01; ���, P < 0.001.
TDP1 and PARP-1 in Rhabdomyosarcoma
www.aacrjournals.org Mol Cancer Res; 11(10) October 2013 1185
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 2, 2013; DOI: 10.1158/1541-7786.MCR-12-0575

increased the sensitivity of mouse embryonic fibroblaststo CPT (Supplementary Fig. S3B), we hypothesized thatresidual Tdp1 activity provided this resistance of skeletalmyoblasts toCPT and indeed observed 50% residual activityamong myoblasts with 85% knockdown of TDP1 mRNA(Supplementary Fig. S4).To determine whether the increased sensitivity of rhab-
domyosarcoma cells to CPT arose from accumulation ofDNA breaks, we used alkaline comet assays to compare 1mmol/L CPT treatment alone and 1 mmol/L CPT treatmentplus knockdown of TDP1. The combined treatmentinduced 5-fold more DNA strand breaks in A204, Birch,and CW9019 cells and 15-foldmore in RH30 cells over cellstreated with CPT alone (Fig. 3B). In contrast, combinedtreatment did not significantly increase the amount of DNAbreaks over CPT treatment alone in control myoblasts.To determine whether this differential sensitivity of rhab-
domyosarcoma cells versus control myoblasts was attribut-able to different proliferation rates that gave rise to differ-ences in the likelihood of collision between the replicationapparatus and DNA-TopoI complexes, we analyzed theconsequences of TDP1 knockdown on the breast cancercell lineMDA-231. The proliferation rate ofMDA-231 cellswas 16 hours (Supplementary Fig. S5A); a rate that iscomparable with that of CW9019 cells (Fig. 3A) andapproximately 3-fold faster than normal myoblasts (48hours; Fig. 3A). TDP1 expression in MDA-231 cells wascomparable with that of the myoblasts (Supplementary Fig.S5B). A 1 mmol/L CPT treatment of MDA-231 cells after75% TDP1 knockdown did not significantly increase theamount of DNA breaks compared with CPT treatmentalone (Supplementary Fig. S5C and S5D).
PARP-1 expression is increased in rhabdomyosarcomacell lines and its inhibition preferentially increases theCPT sensitivity of rhabdomyosarcoma cell linescompared with control skeletal myoblastsAs PARP-1 and Tdp1 are in the same DNA-repair
complex, we hypothesized that PARP-1 inhibition alsosensitizes rhabdomyosarcoma cells to CPT. As observedfor Tdp1, rhabdomyosarcoma cells expressed more PARP-1 mRNA and protein than control skeletal myoblasts(Supplementary Fig. S6A and S6B). The administrationof the PARP-1–specific inhibitor AG-014699 (Rucaparib)reduced the amount of PARP-1 (poly-ADP ribosyl poly-mers, PAR) products in all cell lines (Fig. 4, insets). Asshown by comet assay (Supplementary Fig. S7), treatmentwith 10 mmol/L rucaparib increased the number of DNAbreaks in all rhabdomyosarcoma cell lines exposed to 1mmol/L CPT by 4 to 5-fold compared with treatmentwith 1 mmol/L CPT alone. In contrast, the same treat-ment on control myoblasts only increased the sensitivityto 1 mmol/L CPT by 2-fold.To ascertain whether inhibition of PARP-2 in conjunc-
tion with inhibition of PARP-1would bemore effective thaninhibition of PARP-1 alone, we tested inhibitors targetingboth PARP-1 and PARP-2. In contrast with Tdp1 andPARP-1, PARP-2 mRNA and protein expression are unal-
tered or decreased in most rhabdomyosarcoma cells linescompared with control myoblasts (Supplementary Fig. S6Aand S6C). As measured by comet assay, AZD-2281 (Ola-parib) and ABT-888 (Veliparib), which inhibit both PARP-1 and PARP-2, sensitized rhabdomyosarcoma cells less toCPT than did Rucaparib (Supplementary Fig. S7). AsRucaparib (Ki ¼ 1.4 nmol/L) is a more potent inhibitor ofPARP-1 than are Olaparib (Ki ¼ 5 nmol/L), and Veliparib(Ki ¼ 5.2 nmol/L), we hypothesize that the difference ineffectiveness of these 3 compounds is the differential inhi-bition of PARP-1.
TDP1 knockdown and PARP-1 inhibition alone andtogether are more cytotoxic to rhabdomyosarcoma cellsthan to control skeletal myoblastsWe observed that either TDP1 knockdown or PARP-1
inhibition impaired proliferation of the rhabdomyosarcomacell lines (Fig. 4A), as measured using theMTT assay. About70% to 90% siRNA knockdown of TDP1 reduced prolif-eration of A204, Birch, and RH30 by 30%, 50%, and 30%respectively, but had no effect on controlmyoblasts (Fig. 4A).Of the PARP-1 inhibitors tested, Rucaparib was most det-rimental to rhabdomyosarcoma cell proliferation (Supple-mentary Table S3). Treatment with 10 mmol/L Rucaparibfor 96 hours led to a 50% or greater decrease in proliferationof 4 of 5 rhabdomyosarcoma cell lines compared withuntreated cells, but only a 10% decrease in the proliferationof control myoblasts (Supplementary Table S3 and Fig.S4A). Excepting Birch, this reduction in proliferation is 2-fold more than that observed with TDP1 knockdown alone(Fig. 4A), suggesting that either PARP-1 inhibition is moreeffective than TDP1 knockdown or that PARP-1 inhibitionis affecting pathways essential for rhabdomyosarcoma cellsurvival other than those involving Tdp1.Investigating whether TDP1 knockdown and PARP-1
inhibition are additive, we found that the combination of70% to 90% knockdown of TDP1 and 10 mmol/L Ruca-parib decreased A204, Birch, RH30, and CW9019 cellproliferation more than either treatment alone; this treat-ment had no effect on control myoblasts (Fig. 4A). Over thecourse of 96 hours, A204, Birch, and RH30 cell numbersdeclined by 70% to 80%, and as measured by caspase-3/7expression and terminal deoxynucleotidyl transferase–medi-ated dUTP nick end labeling (TUNEL)-positive staining,this toxicity correlated with induction of apoptosis (Fig. 4Band C), implying that cytotoxicity was responsible for thedecline in proliferation. In addition, suggestive of TDP1knockdown and PARP-1 inhibition targeting the samepathway, constitutive TDP1 knockdown (SupplementaryFig. S3C and S3D) resulted in the selection of cells that wereresistant to 10 mmol/L rucaparib treatment, whereas cellsconstitutively expressing a nontargeting shRNA remainedsensitive to PARP-1 inhibition (Supplementary Fig. S8A).
Rhabdomyosarcoma cell lines do not harbor pathogenicBRCA1 or BRCA2 mutationsOn the basis of the effect of PARP-1 inhibitors on cancer
cells with BRCA1 or BRCA2 mutations or other defects of
Fam et al.
Mol Cancer Res; 11(10) October 2013 Molecular Cancer Research1186
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 2, 2013; DOI: 10.1158/1541-7786.MCR-12-0575

DNA double-strand break repair (7, 30), we postulated thatthe cytotoxicity of TDP1 knockdown and PARP-1 inhibi-tion on rhabdomyosarcoma cells could arise from loss offunctional BRCA1 or BRCA2. However, qRT-PCR did notdetect a significant loss of BRCA1 or BRCA2 mRNA andsequencing of theBRCA1 cDNA andBRCA2 exons detectedno pathogenic mutations (Supplementary Tables S4 and S5,Supplementary Fig. S8B). Therefore, BRCA1 and BRCA2perturbations did not explain the sensitivity of rhabdomyo-sarcoma cells to PARP-1 inhibition.In the absence of BRCA1 and BRCA2 mutations, rever-
sion of such mutations does not explain the PARP-1inhibitor resistance of rhabdomyosarcoma cells with con-stitutive TDP1 knockdown. Also, as expression of BRCA1mRNA in rhabdomyosarcoma cells with constitutive Tdp1knockdown declined or stayed the same relative to rhab-
domyosarcoma cells expressing a nontargeting shRNA,increased BRCA1 expression did not account for thePARP-1 inhibitor resistance (Supplementary Fig. S8C).However, as resistance to PARP-1 inhibitors is also cor-related with a reduction in TP53BP1 expression (31), wetested for this and found that A204, Birch, RD, and RH30cells surviving selection for constitutive TDP1 knockdownhad a 50% reduction in TP53BP1 expression (Supple-mentary Fig. S8C).
Serum and glucose restriction is cytotoxic to Birch andCW9019 cell linesDespite the absence of deleterious BRCA1 and BRCA2
mutations, the sensitivity of rhabdomyosarcoma cells toTdp1 knockdown and PARP-1 inhibition suggests theyharbor a defect of DNA repair. Therefore, as glucose and
0 48 960
200
400
600
800
1,000
1,200
0 48 960
200
400
600
800
1,000
1,200
0 48 960
200
400
600
800
1,000
1,200
Hours0 48 96
0
200
400
600
800
1,000
1,200
Hours
0 48 960
200
400
600
800
1,000
1,200
0 48 960
200
400
600
800
1,000
1,200
Control
Tdp1 KD
Both
0
20
40
60
0
10
20
30
A204
Birch
RD
CW9019
RH30
HSM
Caspase activity TUNEL
Rucaparib
Rucaparib
Both
Control
Tdp1 KD
A
B C
Arb
itrar
y flu
ores
cenc
e un
its (
×105
)
PAR
GAPDH
PAR
GAPDH
PAR
GAPDH
PAR
GAPDH
PAR
GAPDH
PAR
GAPDH
******
***
******
*********
*****
***
***
*** ******
******
******
*****
***
******
***
A204
Birch
RD
CW90
19Rh3
0HSM A20
4Birc
h RD
CW90
19Rh3
0HSM
Cel
l via
bilit
y (%
) C
ell v
iabi
lity
(%)
Cel
l via
bilit
y (%
)
Apo
ptot
ic c
ells
(pe
r 15
0 ce
lls)
Figure 4. Combined TDP1knockdown and PARP-1 inhibitionare more cytotoxic torhabdomyosarcoma than eitheralone. A, graph of the number ofrhabdomyosarcoma cells andcontrol myoblasts over a 96-hourperiod during which the cells weresubjected to (i) nontargeting siRNAtreatment, (ii) TDP1 siRNAtreatment (iii) 10 mmol/L PARP-1inhibitor (rucaparib) treatment, or(iv) combined rucaparib and TDP1siRNA treatment (n ¼ 4). Insetwithin graphs are photographs ofimmunoblots showing PARpolymer expression after 24-hourrucaparib treatment. GAPDH isused as a loading control. B, graphof a caspase-3/7 activation assayillustrating caspase-3/7 cleavagefor all cell lines under conditions 1through4 above (n¼3). C, graphofTUNEL assay showing the numberof terminally apoptotic cells asmeasured by DNA strand breakassay for all cell lines underconditions 1 through 4 above(n ¼ 3). Error bars, mean � SDamong treatments 2, 3, and 4.��, P < 0.01; ���, P < 0.001.
TDP1 and PARP-1 in Rhabdomyosarcoma
www.aacrjournals.org Mol Cancer Res; 11(10) October 2013 1187
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 2, 2013; DOI: 10.1158/1541-7786.MCR-12-0575
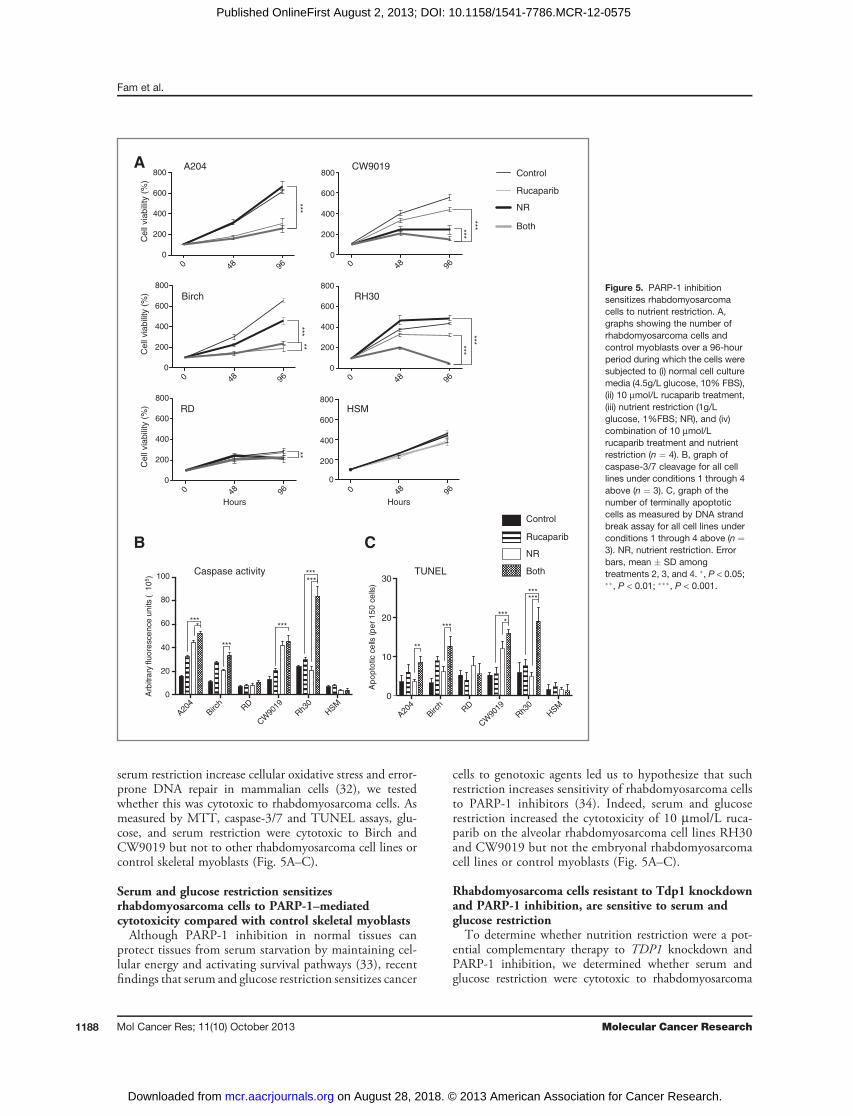
serum restriction increase cellular oxidative stress and error-prone DNA repair in mammalian cells (32), we testedwhether this was cytotoxic to rhabdomyosarcoma cells. Asmeasured by MTT, caspase-3/7 and TUNEL assays, glu-cose, and serum restriction were cytotoxic to Birch andCW9019 but not to other rhabdomyosarcoma cell lines orcontrol skeletal myoblasts (Fig. 5A–C).
Serum and glucose restriction sensitizesrhabdomyosarcoma cells to PARP-1–mediatedcytotoxicity compared with control skeletal myoblastsAlthough PARP-1 inhibition in normal tissues can
protect tissues from serum starvation by maintaining cel-lular energy and activating survival pathways (33), recentfindings that serum and glucose restriction sensitizes cancer
cells to genotoxic agents led us to hypothesize that suchrestriction increases sensitivity of rhabdomyosarcoma cellsto PARP-1 inhibitors (34). Indeed, serum and glucoserestriction increased the cytotoxicity of 10 mmol/L ruca-parib on the alveolar rhabdomyosarcoma cell lines RH30and CW9019 but not the embryonal rhabdomyosarcomacell lines or control myoblasts (Fig. 5A–C).
Rhabdomyosarcoma cells resistant to Tdp1 knockdownand PARP-1 inhibition, are sensitive to serum andglucose restrictionTo determine whether nutrition restriction were a pot-
ential complementary therapy to TDP1 knockdown andPARP-1 inhibition, we determined whether serum andglucose restriction were cytotoxic to rhabdomyosarcoma
0 48 960
200
400
600
800
Hours
0 48 960
200
400
600
800
0 48 960
200
400
600
800
0 48 960
200
400
600
800
0 48 960
200
400
600
800
0 48 960
200
400
600
800
Hours
A204
Birch
RD
CW90
19Rh3
0HSM
Control
Rucaparib
Caspase activity TUNEL
A204
Birch
RD
CW90
19Rh3
0HSM
Control
Rucaparib
Both
A
B C
NR
Both
A204
Birch
RD
CW9019
RH30
HSM
Arb
itrar
y flu
ores
cenc
e un
its (×
105 )
0
20
40
60
80
100
0
10
20
30
Ap
op
totic
ce
lls (p
er 1
50
ce
lls)
NR***
*****
**
************
****
***
***
******
**
******
*
******
Cel
l via
bilit
y (%
)C
ell v
iabi
lity
(%)
Cel
l via
bilit
y (%
)
Figure 5. PARP-1 inhibitionsensitizes rhabdomyosarcomacells to nutrient restriction. A,graphs showing the number ofrhabdomyosarcoma cells andcontrol myoblasts over a 96-hourperiod during which the cells weresubjected to (i) normal cell culturemedia (4.5g/L glucose, 10% FBS),(ii) 10 mmol/L rucaparib treatment,(iii) nutrient restriction (1g/Lglucose, 1%FBS; NR), and (iv)combination of 10 mmol/Lrucaparib treatment and nutrientrestriction (n ¼ 4). B, graph ofcaspase-3/7 cleavage for all celllines under conditions 1 through 4above (n ¼ 3). C, graph of thenumber of terminally apoptoticcells as measured by DNA strandbreak assay for all cell lines underconditions 1 through 4 above (n ¼3). NR, nutrient restriction. Errorbars, mean � SD amongtreatments 2, 3, and 4. �, P < 0.05;��, P < 0.01; ���, P < 0.001.
Fam et al.
Mol Cancer Res; 11(10) October 2013 Molecular Cancer Research1188
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 2, 2013; DOI: 10.1158/1541-7786.MCR-12-0575

cells resistant to 10 mmol/L PARP-1 inhibition and consti-tutive TDP1 knockdown.We found that serum and glucoserestriction did cause apoptosis of A204, Birch, RH30, andCW9019 cells resistant to PARP-1 inhibitors, whereascontrol myoblasts were minimally affected (Fig. 6A–C).
DiscussionWe have shown that Tdp1 and PARP-1 are highly
expressed in rhabdomyosarcoma tumors and that as predictedfrom the functions of Tdp1 and PARP-1 in DNA repair,knockdown of TDP1 and inhibition of PARP-1 sensitizedrhabdomyosarcoma cells to CPT treatment. Suggestive of anintrinsic defect of DNA repair in rhabdomyosarcoma celllines, TDP1 knockdown and PARP-1 inhibition alone or in
combination also caused apoptosis of rhabdomyosarcomacells. In addition, stressing the DNA repair and replicationsystem through glucose and serum restriction reduced via-bility of some rhabdomyosarcoma cells and enhanced sensi-tivity to PARP-1 inhibitors. Remarkably, each of thesemanipulations preferentially affected tumor cells comparedwith control skeletal myoblasts.Tdp1 and PARP-1 are members of a DNA single-strand
break repair complex that also includes XRCC1, Ligase III,and PNKP (35). Within this complex, Tdp1 processestopoisomerase-I–DNA complexes and blocked 30-termini(27). Tdp1 mutant cell lines and knockout mice haveincreased sensitivity to several anticancer agents (25, 36).These observations have consequently led to the suggestionthat Tdp1 overexpression in tumor cells confers resistance to
A
0 48 960
200
400
600
800 Control KD
Tdp1 KD
Control + NR
Tdp1 KD + NR
0 48 960
200
400
600
800
0 48 960
200
400
600
800
0 48 960
200
400
600
800
0 48 960
200
400
600
800
0 48 96
HoursHours
A204
Birch
RD
CW9019
RH30
HSM
0
20
40
60
80
100
Arb
itrar
y flu
ores
cenc
e un
its (×
105 )
Control KD
Tdp1 KD
Control KD + NR
Tdp1 KD + NR
Caspase activity TUNEL
B C
***
***
****
******
***
***
***
***
******
***
***
******
0
200
400
600
800
A204
Birch
RD
CW90
19Rh3
0HSM
A204
Birch
RD
CW90
19Rh3
0HSM
0
10
20
30
Apo
ptot
ic c
ells
(pe
r 15
0 ce
lls)
Cel
l via
bilit
y (%
)C
ell v
iabi
lity
(%)
Cel
l via
bilit
y (%
)
Figure 6. Nutrient restriction is aneffective alternative for treatingrhabdomyosarcoma cells resistantto TDP1 knockdown and PARP-1inhibition. A, graphs showing thenumber of rhabdomyosarcomacells and control myoblasts over a96-hour period for (i) cells treatedwith nontargeting shRNA, (ii) cellstreated with TDP1 shRNA, (iii)nontargeting shRNA cells culturedunder nutrient restriction, or (iv)TDP1 shRNA cells cultured undernutrient restriction (n¼ 4). B, graphof caspase-3/7 cleavage for all celllines under conditions 1 through 4above (n ¼ 3). C, graph of thenumber of terminally apoptoticcells as measured by DNA strandbreak assay for all cell lines underconditions 1 through 4 above (n ¼3). Error bars, mean � SD amongtreatments 2, 3, and 4. �, P < 0.05;���, P < 0.001.
TDP1 and PARP-1 in Rhabdomyosarcoma
www.aacrjournals.org Mol Cancer Res; 11(10) October 2013 1189
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 2, 2013; DOI: 10.1158/1541-7786.MCR-12-0575

anticancer agents, particularly the CPT analogues (19). Theincreased expression of Tdp1 in rhabdomyosarcoma com-pared with unaffected skeletal muscle and the increasedsensitivity of rhabdomyosarcoma cells to CPT after TDP1knockdown or PARP-1 inhibition suggest a possible answerfor the failure of irinotecan to show efficacy against rhabdo-myosarcoma (18).For the rhabdomyosarcoma cell lines RD, A204, Birch,
RH30, and CW9019, this markedly elevated expression ofTdp1 is not explicable as a compensatory reaction to aTDP1hypomorphic allele or to aberrant Tdp1 subcellular locali-zation. As many cancers compensate for defects of DNArepair, which are permissive for rapid tumor evolution, byoverexpression of other DNA repair pathways (37, 38), wehad wondered whether rhabdomyosarcoma cells might alsohave a defect of DNA repair and whether the overexpressionof Tdp1 was a compensatory survival mechanism. Findingthat TDP1 knockdown and PARP-1 inhibition preferen-tially decreased rhabdomyosarcoma cell viability over that ofcontrol myoblasts, we can now hypothesize that rhabdo-myosarcoma tumors have a damaged DNA repair processthat is at least partially compensated for by processes inwhich Tdp1 and PARP-1 are involved.Tdp1 and PARP-1 are involved in repairing both single-
and double-strand DNA breaks (36, 39, 40). However,PARP-1 has in addition been shown to participate in theprocesses of DNA replication and transcription (41, 42). Therole of PARP-1 in these latter functions might partiallyexplain why PARP-1 inhibition had a more deleterious effecton rhabdomyosarcoma cells than did TDP1 knockdown.Alternatively, PARP-1 inhibition might have reduced DNA-repair activity more than TDP1 knockdown as the knock-down alone left more than 10% expression of Tdp1. Sup-porting the last explanation, 85% knockdown of TDP1diminished Tdp1 cleavage activity by 50% in myoblasts andwas insufficient to induce CPT hypersensitivity, a property oftissues and organisms fully deficient for Tdp1 (25, 35, 36).Even though TDP1 knockdown was insufficient to cause
hypersensitivity of control myoblasts to CPT, it was suffi-cient to decrease the viability of rhabdomyosarcoma cells andto increase their sensitivity to CPT. These observations raisethe possibility of selectively targeting Tdp1 and PARP-1 tosensitize cancer but not noncancerous cells. In addition,Tdp1 knockdown or PARP-1 inhibition alone were cyto-toxic to a subset of cells in all rhabdomyosarcoma cell linesanalyzed.However, because some cells were resistant to thesetreatments, rhabdomyosarcoma cells have or rapidly developresistance to TDP1 knockdown and PARP-1 inhibition.In other cancers, resistance to PARP-1 inhibition occursthrough partial correction of defects in homologous recom-bination-mediated DNA double-strand break repair and caninclude reversion ofmutations inBRCA1 or loss ofTP53BP1expression (31). Two recent studies in mice showed that lossof TP53BP1 in BRCA1 mutant cells leads to PARP-1inhibitor resistance and that TP53BP1 loss did not alleviatesensitivity to PARP-1 inhibition without the accompanyingBRCA1 loss-of-functionmutation (43, 44). Therefore, as wedid not find mutations in BRCA1, we hypothesize that if the
decreasedTP53BP1 expression is relevant to the resistance toPARP-1 inhibition, then there must be other mutations ofDNA repair in the rhabdomyosarcoma cell lines to comple-ment the reduced TP53BP1 expression. In addition, thepresence or development of cells resistant to TDP1 knock-down and PARP-1 inhibition suggest that targeting Tdp1and PARP-1 in rhabdomyosarcoma needs to be part of amultimodal therapy.Targeting the altered energymetabolism common among
solid tumors is a potential component of multimodaltherapy (45). Altered tumor metabolism arises becausealthough solid tumor cells require a high metabolic rate tosupport their rapid proliferation (46), they come to rely oninefficient anaerobic metabolism as they outgrow theirblood supply. In a recent study, glucose deprivation inglioblastoma cells triggered mitochondria-derived reactiveoxygen species (ROS) production that caused ROS-medi-ated cell death (47). As Tdp1 and PARP-1 contribute to thebase excision repair processes targeting oxidatively damagednuclear and mitochondrial DNA (48–50), the complemen-tation between inhibition of Tdp1 or PARP-1 and glucosedeprivation has a logical basis. Further studies are required;however, to test this hypothesis as was recently done forother cell lines (49, 51). Nonetheless, the observation doessuggest that nutritional restriction is a consideration fortreatment of rhabdomyosarcoma.A final therapeutic consideration arising from our study is
that none of the 5 rhabdomyosarcoma cell lines respondedidentically to any one therapy, let alone all therapies. Thissuggests that the genetic and epigenetic defects intrinsic toeach rhabdomyosarcoma tumor are likely different. In theunderstanding of these differences resides the ability totarget therapies to the tumor and minimally harm normaltissues. The 3 potential therapeutic approaches identifiedherein all preferentially affected rhabdomyosarcoma celllines compared with control myoblasts and thus providedirection for tumor stratification, analysis of genomic stud-ies of rhabdomyosarcoma tumors and a basis for futurepreclinical trials.In summary, we have shown that TDP1 knockdown and
PARP-1 inhibition increase the sensitivity of rhabdomyo-sarcoma cells to CPT analogues and have a direct toxic effecton some rhabdomyosarcoma cell types. We also find thatglucose and serum restriction can be cytotoxic to rhabdo-myosarcoma cells and sensitize them to PARP-1 inhibition.Although further study is needed to understand the molec-ular mechanisms underlying the effects of these therapieson rhabdomyosarcoma cells and to determine whetherthey have relevance in vivo, these or similar therapies havethe potential to improve outcomes for individuals withrhabdomyosarcoma.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
DisclaimerThe contents of this manuscript have not been published, in whole or in part, prior
to or simultaneous with our submission of the manuscript to Molecular CancerResearch. The necessary ethics committee approval was secured for the study reported.
Fam et al.
Mol Cancer Res; 11(10) October 2013 Molecular Cancer Research1190
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 2, 2013; DOI: 10.1158/1541-7786.MCR-12-0575

Authors' ContributionsConception and design: S.A. Mitra, M. Chowdhury, M. Bond, C.J. Pallen, C.F.BoerkoelDevelopment of methodology:H.K. Fam, S.A. Mitra, M. Chowdhury, K. Choi, C.F. BoerkoelAcquisition of data (provided animals, acquired and managed patients, providedfacilities, etc.): H.K. Fam, C. Walton, S.A. Mitra, G. Sun, P.C.W. Wong, G.Turashvili, S. Aparicio, T.J. Triche, C.J. PallenAnalysis and interpretation of data (e.g., statistical analysis, biostatistics, compu-tational analysis):H.K. Fam, S.A. Mitra, M. Chowdhury, N. Osborne, K. Choi, G.Sun, G. Turashvili, S. Aparicio, C.J. Pallen, C.F. BoerkoelWriting, review, and/or revision of the manuscript: H.K. Fam, C. Walton, S.A.Mitra, M. Chowdhury, M.J. O'Sullivan, G. Turashvili, C.J. Pallen, C.F. BoerkoelAdministrative, technical, or material support (i.e., reporting or organizing data,constructing databases): M. Chowdhury, K. Choi, M.J. O'Sullivan, T.J. TricheStudy supervision: C.J. Pallen, C.F. Boerkoel
AcknowledgmentsThe authors thankDrs. Kirk Schultz, GreggMorin, andDouglasHawkins for their
criticism, suggestions and review of this manuscript. The authors also thank YokoShimizu for assistance with the Tdp1 cleavage assay. Tissue samples from the COG
were provided by the Cooperative Human Tissue Network, which is funded by theNational Cancer Institute. Other investigators may have received specimens from thesame subjects.
Grant SupportThis work was supported in part by grants from theMichael Smith Foundation for
Health Research [CI-SCH-O1899(07-1) to C.F. Boerkoel], the Child & FamilyResearch Institute (to C.F. Boerkoel), the Child & Family Research Institute PediatricOncology Research Program (to M. Bond, C.F. Boerkoel, and C.J. Pallen), and theCanadian Institutes of Health Research (PPP-248178). G. Sun and M. Chowdhurywere supported by a Michael Cuccione Studentship for Oncology Research. C.J.Pallen holds an investigatorship from the Child & Family Research Institute. C.F.Boerkoel is a scholar of the Michael Smith Foundation for Health Research and aClinical Investigator of the Child & Family Research Institute.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be herebymarked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
Received October 4, 2012; revised June 11, 2013; accepted June 21, 2013;published OnlineFirst August 2, 2013.
References1. Stanulla M, Schrappe M. Treatment of childhood acute lymphoblastic
leukemia. Semin Hematol 2009;46:52–63.2. Hingorani P, Kolb EA. Past, present, and future of therapies in pediatric
sarcomas. Future Oncol 2010;6:605–18.3. Stegmeier F, Warmuth M, Sellers WR, Dorsch M. Targeted cancer
therapies in the twenty-first century: lessons from imatinib. Clin Phar-macol Ther 2010;87:543–52.
4. Cahill DP, Kinzler KW, Vogelstein B, Lengauer C. Genetic instabilityand darwinian selection in tumours. Trends Cell Biol 1999;9:M57–60.
5. Pinilla-Ibarz J, Quintas-Cardama A. New agents in the treatment ofchronic myelogenous leukemia. J Natl Compr Canc Netw 2009;7:1028–37.
6. Bartek J, Lukas J, Bartkova J. DNA damage response as an anti-cancer barrier: damage threshold and the concept of "conditionalhaploinsufficiency". Cell Cycle 2007;6:2344–7.
7. Fong PC, Boss DS, Yap TA, Tutt A, Wu P, Mergui-Roelvink M, et al.Mergui-Roelvink M, Inhibition of poly(ADP-ribose) polymerase intumors from BRCA mutation carriers. N Engl J Med 2009;361:123–34.
8. Dagher R, Helman L. Rhabdomyosarcoma: an overview. Oncologist1999;4:34–44.
9. Arndt CA, Stoner JA, Hawkins DS, Rodeberg DA, Hayes-Jordan AA,Paidas CN, et al. Vincristine, actinomycin, and cyclophosphamidecompared with vincristine, actinomycin, and cyclophosphamide alter-nating with vincristine, topotecan, and cyclophosphamide for inter-mediate-risk rhabdomyosarcoma: Children's Oncology Group StudyD9803. J Clin Oncol 2009;27:5182–8.
10. Ellison LF, Pogany L, Mery LS. Childhood and adolescent cancersurvival: a period analysis of data from the Canadian Cancer Registry.Eur J Cancer 2007;43:1967–75.
11. Zhan S, Shapiro DN, Helman LJ. Activation of an imprinted allele of theinsulin-like growth factor II gene implicated in rhabdomyosarcoma.J Clin Invest 1994;94:445–8.
12. Barr FG, Galili N, Holick J, Biegel JA, Rovera G, Emanuel BS. Rear-rangement of the PAX3 paired box gene in the paediatric solid tumouralveolar rhabdomyosarcoma. Nat Genet 1993;3:113–7.
13. Davis RJ, D'Cruz CM, Lovell MA, Biegel JA, Barr FG. Fusion of PAX7 toFKHR by the variant t(1;13)(p36;q14) translocation in alveolar rhabdo-myosarcoma. Cancer Res 1994;54:2869–72.
14. Ayalon D, Glaser T, Werner H. Transcriptional regulation of IGF-Ireceptor gene expression by the PAX3-FKHR oncoprotein. GrowthHorm IGF Res 2001;11:289–97.
15. Kolb EA, Gorlick R, Houghton PJ, Morton CL, Lock R, Carol H, et al.Initial testing (stage 1) of a monoclonal antibody (SCH 717454) againstthe IGF-1 receptor by the pediatric preclinical testing program. PediatrBlood Cancer 2008;50:1190–7.
16. Brenner JC, Feng FY, Han S, Patel S, Goyal SV, Bou-Maroun LM, et al.PARP-1 inhibition as a targeted strategy to treat Ewing's sarcoma.Cancer Res 2012;72:1608–13.
17. Garnett MJ, Edelman EJ, Heidorn SJ, Greenman CD, Dastur A, LauKW, et al. Systematic identification of genomic markers of drugsensitivity in cancer cells. Nature 2012;483:570–5.
18. Mascarenhas L, Lyden ER, Breitfeld PP, Walterhouse DO, DonaldsonSS, Paidas CN, et al. Randomized phase II window trial of twoschedules of irinotecan with vincristine in patients with first relapseor progression of rhabdomyosarcoma: a report from the Children'sOncology Group. J Clin Oncol 2010;28:4658–63.
19. Liu C, Zhou S, Begum S, Sidransky D, Westra WH, Brock M, et al.Increased expression and activity of repair genes TDP1 and XPF innon–small cell lung cancer. Lung Cancer 2007;55:303–11.
20. Walterhouse DO, Lyden ER, Breitfeld PP, Qualman SJ, Wharam MD,Meyer WH. Efficacy of topotecan and cyclophosphamide given in aphase II window trial in children with newly diagnosed metastaticrhabdomyosarcoma: a Children's Oncology Group study. J Clin Oncol2004;22:1398–403.
21. Pappo AS, Lyden E, Breitfeld P, Donaldson SS, Wiener E, Parham D,et al. Two consecutive phase II window trials of irinotecan alone or incombination with vincristine for the treatment of metastatic rhabdo-myosarcoma: the Children's Oncology Group. J Clin Oncol 2007;25:362–9.
22. Domagala P, Huzarski T, Lubinski J, Gugala K, Domagala W. Immu-nophenotypic predictive profiling of BRCA1-associated breast cancer.Virchows Arch 2011;458:55–64.
23. Goncalves A, Finetti P, Sabatier R, Gilabert M, Adelaide J, Borg JP,et al. Poly(ADP-ribose) polymerase-1 mRNA expression in humanbreast cancer: a meta-analysis. Breast Cancer Res Treat 2011;127:273–81.
24. NoshoK,YamamotoH,MikamiM,Taniguchi H, Takahashi T, Adachi Y,et al. Overexpression of poly(ADP-ribose) polymerase-1 (PARP-1) inthe early stage of colorectal carcinogenesis. Eur J Cancer 2006;42:2374–81.
25. Hirano R, Interthal H, Huang C, Nakamura T, Deguchi K, Choi K, et al.Spinocerebellar ataxia with axonal neuropathy: consequence of aTdp1 recessive neomorphic mutation? EMBO J 2007;26:4732–43.
26. Mosmann T. Rapid colorimetric assay for cellular growth and survival:application to proliferation and cytotoxicity assays. J Immunol Meth-ods 1983;65:55–63.
27. Murai J, HuangSY,DasBB,Dexheimer TS, TakedaS,PommierY, et al.Tyrosyl-DNA phosphodiesterase 1 (TDP1) repairs DNA damageinduced by topoisomerases I and II and base alkylation in vertebratecells. J Biol Chem 2012;287:12848–57.
28. Jensen PW, Falconi M, Kristoffersen EL, Simonsen AT, Cifuentes JB,Marcussen LB, et al. Real-time detection of TDP1 activity using a
TDP1 and PARP-1 in Rhabdomyosarcoma
www.aacrjournals.org Mol Cancer Res; 11(10) October 2013 1191
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 2, 2013; DOI: 10.1158/1541-7786.MCR-12-0575

fluorophore-quencher coupled DNA-biosensor. Biosens Bioelectron2013;48C:230–7.
29. Boerkoel CF, Takashima H, Garcia CA, Olney RK, Johnson J, Berry K,et al.Charcot-Marie-Tooth disease and relatedneuropathies:mutationdistribution andgenotype-phenotype correlation. AnnNeurol 2002;51:190–201.
30. Tutt A, Robson M, Garber JE, Domchek SM, Audeh MW, Weitzel JN,et al. Oral poly(ADP-ribose) polymerase inhibitor olaparib in patientswith BRCA1 or BRCA2 mutations and advanced breast cancer: aproof-of-concept trial. Lancet 2010;376:235–44.
31. Aly A, Ganesan S. BRCA1, PARP, and 53BP1: conditional syntheticlethality and synthetic viability. J Mol Cell Biol 2011;3:66–74.
32. Siggens L, Figg N, Bennett M, Foo R. Nutrient deprivation regulatesDNA damage repair in cardiomyocytes via loss of the base-excisionrepair enzyme OGG1. FASEB J 2012;26:2117–24.
33. Mathews MT, Berk BC. PARP-1 inhibition prevents oxidative andnitrosative stress-induced endothelial cell death via transactivation ofthe VEGF receptor 2. Arterioscler Thromb Vasc Biol 2008;28:711–7.
34. Lee C, Raffaghello L, Brandhorst S, Safdie FM, Bianchi G, Martin-Montalvo A, et al. Fasting cycles retard growth of tumors and sensitizea range of cancer cell types to chemotherapy. Sci Transl Med 2012;4:124ra27.
35. El-Khamisy SF, SaifiGM,WeinfeldM, Johansson F, Helleday T, LupskiJR, et al. Defective DNA single-strand break repair in spinocerebellarataxia with axonal neuropathy-1. Nature 2005;434:108–13.
36. Katyal S, el-Khamisy SF, Russell HR, Li Y, Ju L, Caldecott KW, et al.TDP1 facilitates chromosomal single-strand break repair in neuronsand is neuroprotective in vivo. EMBO J 2007;26:4720–31.
37. Messina A, Puccinelli E, Gervasi PG, Longo V. Expression andinducibility of CYP1A1, 1A2, 1B1 by beta-naphthoflavone andCYP2B22, CYP3Asby rifampicin in heart regions and coronary arteriesof pig. Res Vet Sci 2013;94:77–83.
38. Chiarugi A. A snapshot of chemoresistance to PARP inhibitors. TrendsPharmacol Sci 2012;33:42–8.
39. Plo I, Liao ZY, Barcel�o JM, Kohlhagen G, Caldecott KW, Weinfeld M,et al. Association of XRCC1 and tyrosyl DNA phosphodiesterase(Tdp1) for the repair of topoisomerase I-mediated DNA lesions. DNARepair (Amst) 2003;2:1087–100.
40. Schultz N, Lopez E, Saleh-Gohari N, Helleday T. Poly(ADP-ribose)polymerase (PARP-1) has a controlling role in homologous recombi-nation. Nucleic Acids Res 2003;31:4959–64.
41. Ray Chaudhuri A, Hashimoto Y, Herrador R, Neelsen KJ, Fachi-netti D, Bermejo R, et al. Topoisomerase I poisoning results inPARP-mediated replication fork reversal. Nat Struct Mol Biol2012;19:417–23.
42. Kim MY, Mauro S, G�evry N, Lis JT, Kraus WL. NADþ-dependentmodulation of chromatin structure and transcription by nucleosomebinding properties of PARP-1. Cell 2004;119:803–14.
43. Bouwman P, Aly A, Escandell JM, Pieterse M, Bartkova J, van derGulden H, et al. 53BP1 loss rescues BRCA1 deficiency and is asso-ciated with triple-negative and BRCA-mutated breast cancers. NatStruct Mol Biol 2010;17:688–95.
44. Bunting SF, Call�en E,WongN,ChenHT, Polato F, GunnA, et al. 53BP1inhibits homologous recombination in Brca1-deficient cells by block-ing resection of DNA breaks. Cell 2010;141:243–54.
45. Marin-Valencia I, Yang C, Mashimo T, Cho S, Baek H, Yang XL, et al.Analysis of tumormetabolism revealsmitochondrial glucose oxidationin genetically diverse human glioblastomas in the mouse brain in vivo.Cell Metab 2012;15:827–37.
46. Gatenby RA, Gillies RJ. Why do cancers have high aerobic glycolysis?Nat Rev Cancer 2004;4:891–9.
47. GrahamNA, TahmasianM, Kohli B, KomisopoulouE, ZhuM, Vivanco I,et al. Glucose deprivation activates a metabolic and signaling ampli-fication loop leading to cell death. Mol Syst Biol 2012;8:589.
48. Das BB, Dexheimer TS, Maddali K, Pommier Y. Role of tyrosyl-DNAphosphodiesterase (TDP1) in mitochondria. Proc Natl Acad Sci U S A2010;107:19790–5.
49. Luo X, Kraus WL. On PAR with PARP: cellular stress signaling throughpoly(ADP-ribose) and PARP-1. Genes Dev 2012;26:417–32.
50. FamHK, Chowdhury MK, Walton C, Choi K, Boerkoel CF, Hendson G.Expression profile and mitochondrial colocalization of Tdp1 in periph-eral human tissues. J Mol Histol 2013;44:481–94.
51. Rodriguez-Vargas JM, Ruiz-Maga~na MJ, Ruiz-Ruiz C, Majuelos-Mel-guizo J, Peralta-Leal A, Rodríguez MI, et al. ROS-induced DNA dam-age and PARP-1 are required for optimal induction of starvation-induced autophagy. Cell Res 2012;22:1181–98.
Fam et al.
Mol Cancer Res; 11(10) October 2013 Molecular Cancer Research1192
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 2, 2013; DOI: 10.1158/1541-7786.MCR-12-0575

2013;11:1179-1192. Published OnlineFirst August 2, 2013.Mol Cancer Res Hok Khim Fam, Cheryl Walton, Sheetal A. Mitra, et al. CellsTDP1 and PARP1 Deficiency Are Cytotoxic to Rhabdomyosarcoma
Updated version
10.1158/1541-7786.MCR-12-0575doi:
Access the most recent version of this article at:
Material
Supplementary
http://mcr.aacrjournals.org/content/suppl/2013/08/02/1541-7786.MCR-12-0575.DC1
Access the most recent supplemental material at:
Cited articles
http://mcr.aacrjournals.org/content/11/10/1179.full#ref-list-1
This article cites 51 articles, 16 of which you can access for free at:
Citing articles
http://mcr.aacrjournals.org/content/11/10/1179.full#related-urls
This article has been cited by 4 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mcr.aacrjournals.org/content/11/10/1179To request permission to re-use all or part of this article, use this link
on August 28, 2018. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst August 2, 2013; DOI: 10.1158/1541-7786.MCR-12-0575
![Survival Analysis - University of Washingtonfaculty.washington.edu/heagerty/Courses/VA-survival/...It’s life and death... Survival function: S(t) = P [T > t] The survival function](https://img.pdfslide.net/doc/110x75/611600f94ef3f41cc655565e/survival-analysis-university-of-itas-life-and-death-survival-function.jpg)




























