Embed Size (px)
Citation preview

Cell Membrane Fluid–Mosaic Structure and Cancer Metastasis TRADUCIDO
ResumenLas células cancerosas están rodeadas por una membrana de fluido de mosaicoque proporciona una barrera estructural altamente dinámica con elmicroambiente, filtro de la comunicación y el transporte, el receptory la enzima plataforma. Esta estructura de formas debido a lapropiedades físicas de sus componentes, que pueden moverse lateralmentey de forma selectiva dentro del plano de la membrana y asociarse concomponentes similares o diferentes, formando específica y funcionaldominios. A través de los años, los datos se han acumulado en elcantidades, las estructuras y las movilidades de los constituyentes de la membranadespués de la transformación y durante la progresión y metástasis.Información más reciente ha demostrado la importancia de la especializadadominios de la membrana, tales como balsas de lípidos, proteína-lípidocomplejos, complejos de receptores, invadopodios, y otra celularestructuras en el proceso maligno. En la descripción de la macroestructuray la dinámica de las membranas plasmáticas, membraneassociatedestructuras del citoesqueleto y la matriz extracelular sontambién importante, lo que limita el movimiento de componentes de la membranay actuando como puntos de tracción para la motilidad celular. Estas

asociaciones pueden ser alteradas en las células malignas, y probablementetambién en torno a las células normales, la promoción de la invasión ycolonización metastásica. Además, los componentes pueden serliberado de las células como moléculas secretoras, enzimas, receptores,grandes complejos macromoleculares, vesículas de membrana, y exosomasque puede modificar el microambiente, proporcionar específicadiafonía, y facilitar la invasión, la supervivencia y el crecimiento delas células malignas. Cancer Res; 75 (7); 1169-1176. ? 2015 AACR.
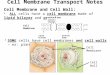
Figura 1.Una célula cancerosa hipotético proceso de cambio a un fenotipo invasivo y comenzar el proceso de invasión. A, una representación de la membrana celularque contiene estructuras de dominios de membrana y citoesqueleto asociada a la membrana y las estructuras extracelulares. La membrana celular se ha peladovuelta a la derecha para revelar la superficie de membrana y asociadas a la membrana elementos del citoesqueleto de fondo que forman barreras (corrales) que limitan lamovimientos laterales de algunas de las proteínas integrales de membrana. Además, las estructuras del citoesqueleto asociadas a la membrana están interactuando con indirectamenteproteínas integrales de membrana en la superficie de la membrana interna, junto con componentes de la matriz de ECM o en la superficie exterior. Aunque este diagrama

presenta posibles mecanismos de membrana integral restricción de movilidad de proteína, no representa con precisión los tamaños y estructuras de la integralproteínas de membrana, dominios de lípidos, o las estructuras del citoesqueleto asociadas a la membrana. B, exosomas o membrana intracelular vesículas se liberan devesículas más grandes que se fusionan con la membrana celular. MF, microfilamentos; MT, microtúbulos. Tomado de BBA Biomembranas, 1838 (6), GL Nicolson,El Fluid-Mosaico Modelo de Estructura de membrana: Todavía relevante para la comprensión de la estructura, función y dinámica de las membranas biológicas después de másde 40 años, desde 1451 hasta 1456, Derechos de Autor 2013, con permiso de Elsevier.
IntroducciónLas membranas celulares representan importantes barreras celulares yestructuras de primer contacto de las células normales y cancerosas. Extracelularseñales de iones, hormonas, citoquinas, enzimas, el crecimiento yfactores de motilidad, receptores, la matriz extracelular (ECM), otroelementos del estroma, y vesículas de membrana subcelular debe primerointeractuar con la membrana celular para iniciar los procesos de señalización.Por lo tanto, las membranas celulares o plasma son filtros celulares que puedentransmitir selectivamente señales y sustancias desde fuera de las células ya partir de células adyacentes en el interior de una célula. A la inversa, también puedeseñales de liberación y moléculas a otras células y la micro y macro-ambiente en un proceso complejo que se ha denominado"Biología celular sociales" (1). Además, las células son compartimentadaen orgánulos por diversas estructuras de la membrana intracelularesque son responsables de la biosíntesis, la producción de energía, la replicación,el transporte, el reciclaje, la destrucción, la secreción, y otraactividades celulares.Las membranas celulares están íntimamente involucrados en la bioquímicaeventos que definen tipos de cáncer, y en particular, son intensamenteinvolucrada en la metástasis del cáncer (2). Además, el establecimientode las metástasis también requiere una compleja interacción entre malignocélulas, las células normales, estroma y ECM en sus nuevos microambientes,y estas interacciones están mediadas principalmentea través de las membranas de células (3).

Propiedades físicas de las membranas celularesUn concepto importante que mantiene la estructura de la membrana celulares que los componentes de la membrana anfipáticas auto-asociados aexcluir a las interacciones del agua en sus superficies hidrofóbicas, mientras quelas porciones hidrófilas de sus estructuras interactúan con elambiente acuoso (4). Así, glycerolphospholipids de membranaauto-ensamblan para formar bicapas de lípidos (5) debido a la energíaproporcionado por el efecto hidrófobo y van der Waals (6).Membrana proteínas globulares integrales interactúan con la membranalípidos a través de sus estructuras acilo debido a fuerzas hidrofóbicasy mucho menos a las interacciones hidrofílicas entre la cabeza de lípidosgrupos y aminoácidos hidrófilos proteína (4, 6, 7).Las proteínas de membrana son operacionalmente de tres tipos: integral,periférico, y asociada a la membrana (7). Proteínas integrales songlobular y fuertemente unido a las membranas por principalmente hidrófobofuerzas y intercalan en la bicapa lipídica de la membrana,mientras que las proteínas de membrana periféricas están obligados a membranaspor fuerzas electrostáticas u otros. Membrana periféricalas proteínas pueden ser removidos de las membranas sin destruirmicroestructura membrana básica y son importantes en la prestación desitios de unión de membrana, andamios, tethering o membrana de soporteestructuras, membrana curvatura de promocióncomponentes y puntos de fijación para las enzimas solubles ymoléculas de señalización (7, 8).Cuando la membrana se distorsiona, deformado, comprimido, oexpandido, diferentes fuerzas y componentes reaccionan a la físicaperturbaciones (9, 10). Por ejemplo, cierta membrana periféricaproteínas pueden unirse y causar la deformación mediante la formación de crescentshapeda-helicoidales paquetes que se unen a las membranas a través de electrostáticay algunas interacciones hidrofóbicas (9), causando una curvatura pormembranas de doblado para adaptarse a la estructura de proteínas periférica (9, 10).La tercera clase de proteínas de membrana a menudo se aisló con la célulamembranas, pero en realidad están sin membrana de proteínas (7).
Se trata de estructuras de señalización del citoesqueleto y asociados en elsuperficie de la membrana interna de la célula, y en la superficie exterior que incluyenciertos componentes de ECM. Estos componentes asociados a la membranason partes de estructuras dinámicas implicadas en la estabilización delas membranas (y por tanto células) y componentes de la membrana de inmovilización.Ellos son especialmente importantes en las actividades celulares, talescomo la adhesión celular y la motilidad, el crecimiento, la endocitosis, exocitosis,transducción de señales, y otras actividades importantes (11, 12).El Fluid-Mosaico Modelo de las membranas celularesEn primer lugar se propone en 1972, el modelo de membrana de líquido mosaico(8) ha demostrado su utilidad en la descripción de nanoescala básicoestructura de la membrana celular por más de 40 años (11). Aunque estemodelo ha sido notablemente consistente con los datos recogidos enmembranas biológicas desde 1925, era inevitable que el

modelo original no podía explicar aspectos de la dinámica de la membrana.Por ejemplo, los conceptos que la membrana celular y dominiosestructuras asociadas a la membrana son importantes en el control delas movilidades laterales y la distribución de proteínas de la membrana celularaún no se habían descubierto (11-14).Modelos de estructura de la membrana celular producen unos pocos años despuésel modelo original (8) eran mucho menos homogénea (7). Ellosinformación adicional contenida en proteínas y lípidos agregacionesy su segregación en dominios de membrana, citoesqueleto,y las interacciones de ECM, entre otras características (7, 11). Sin embargo,en posteriores revisiones del modelo de mosaico fluido, todos los básicoslos elementos a nivel de nanoescala se mantuvieron (11-14). Sin embargo,los arreglos de lípidos y proteínas en más compactoestructuras y dominios que maximizan su naturaleza mosaico juntocon la adición de organización jerárquica membrana dinámicoproducido una descripción mucho más detallada de su organización(Fig. 1A; refs 11-14.).Al clasificar los diferentes lípidos de membrana y membrana integralproteínas en los dominios de membrana especializadas en función de supropiedades físicas, Mouritsen y Bloom (15) propusieron quetal clasificación se basó principalmente en las interacciones hidrofóbicasy algunas interacciones hidrófilas. Esto evita hidrófobodesajustes entre lípidos y proteínas, evitando así la membranadistorsiones (15).El modelo de membrana de fluido de mosaico también representó celularasimetría de la membrana (8). Las membranas celulares son asimétricas en elvisualización de sus componentes (11, 16). El hallazgo de la asimetríadistribuciones de diferentes lípidos, proteínas y glicoproteínasentre las valvas interior y exterior de las membranas celulares es probableuniversal (11, 16, 17). Por otra parte, la interrupción de la membranaasimetría en las membranas celulares está asociada con la activación celular,adhesión, agregación, la apoptosis, el reconocimiento por las células fagocíticas,entre otros eventos. De nota, también se asocia con patológicoprocesos (17, 18).Cytoskeletal- y ECM-Membrana CelularInteraccionesLo que el modelo de membrana de líquido mosaico original, faltaba erala integración de esta estructura con otros elementos celulares(7, 11). Citoesqueleto y ECM interacciones son conocidos para alterarmembrana celular macroestructura por las restricciones en la libertad demovimiento (movilidad lateral) de proteínas de membrana y tambiéncausando movimientos globales de estos y otros componentes porla inmovilización a estructuras celulares o extracelulares (7, 11, 12).Este proceso puede resultar en la endocitosis de algunos macromolecularcomplejos en la superficie celular. La agrupación de receptores, la formación de dominio,ensamblaje de placa submembrane, internalización, la acidificaciónde los endosomas resultantes, la degradación, y la membranareciclaje son parte del reciclaje de membrana normal (19, 20). Los

movilidad de los componentes integrales de membrana también se puede controlarpor célula-célula y las interacciones célula-ECM (21).Complejos de adhesión de la célula y del receptor que están inmovilizadospor ECM o diversas interacciones son capaces de comunicarseseñales que se transmiten a través de un ensamblado dinámicamentecitoesqueleto o la generación de fuerzas mecánicas que se puede movercélulas o resistir esfuerzos mecánicos exteriores (22). Esta seriemontaje de componentes especializados (ECM, integral de membranaproteínas, proteínas de membrana periféricas, proteínas adaptadoras, citoesqueletoelementos, entre otros) pueden haber evolucionado para convertirseñales bioquímicas en las fuerzas mecánicas que son importantes enel comportamiento celular. Aunque muchos de los periférica de membranaproteínas han sido identificados como componentes involucrados en citoesqueletointeracciones con membranas (23), lípidos de membranatambién son importantes en estas interacciones como lípidos especializadosdominios o "balsas lipídicas" (20, 24).Dominios de membrana son estructuras dinámicas que se pueden generarpor ligando o ion de unión, interacciones hidrofóbicas (u otroeventos) y pueden ensamblarse en superestructuras transmembrana complejos.Estos complejos reclutan proteínas periféricas adicionalesen la superficie de la membrana interna de la célula para formar transmembranaplacas que son competentes para iniciar la señalización celular a través deprocesos enzimáticos o someterse a más apego al citoesqueletoelementos (3, 22, 25, 26).Las membranas celulares deben considerarse completamente integradamechanostructures dentro de los tejidos. Ellos interactúan continuamentey enlazar con diversas estructuras intracelulares a los componentesfuera de la célula, mientras que la recepción de las señales y los contactos de lamicroambiente y pasando estas señales en para provocar apropiadorespuestas celulares. También envían mensajes, mantienenla polaridad celular, y las propiedades mecánicas mientras se someterotación constante de sus componentes constituyentes. Por lo tanto, laestructura básica de las membranas celulares se ha desarrollado desde el originalconcepto homogéneo a uno que contiene "dominios" específicosde diferentes tamaños que forman membrana específicos de regulación yestructuras mecánicas que están vinculados a otro intra y extracelularestructuras (11, 12) que están implicados en muchas celularpropiedades características de las células normales y cancerosas (3, 11).Membrana Celular lípidos y proteínasInteraccionesAdemás de sus distribuciones asimétricas través de la membrana,lípidos de membrana también se distribuyen de manera desigual en elplano de membrana (5, 27, 28). El colesterol es particularmente importanteen la organización de la membrana celular y se encuentra a menudo en específicadominios de membrana (27-29). Esto se cree que es debido, en parte, aLa afinidad de colesterol tanto para las fases fluidas y sólidas de membranas(29). Particiones de colesterol en líquido ordenada / -disorderedfases a más o menos en la misma medida y cambia las propiedades

de las fases de lípidos (30). Los esfingolípidos son también importantesen la formación de dominios de lípidos de membrana ordenados (31).Esfingomielinas y fosfatidilcolinas constituyen másde un medio de fosfolípidos de la membrana de plasma y forman laprincipales socios para el colesterol (32). De hecho, esfingomielinas ycolesterol son de vital importancia en la formación de pequeños dominios, ordenados de lípidos ("balsas lipídicas") que generalmente están rodeadas porlípidos en fase líquida (32, 33).La formación de balsas de lípidos es una dinámica y reversibleproceso que confiere propiedades funcionales de señalización a célulalas membranas. Como se mencionó, las balsas de lípidos se caracterizan porenriquecimientos de colesterol y esfingolípidos, que se celebranunidas por enlaces de hidrógeno, el emparejamiento cargo y hidrofóbicay las fuerzas de van der Waals (32-34). Sus componentes puedenintercambiar rápidamente con los lípidos de membrana mayor, así como conlípidos en otras balsas (35). Las balsas de lípidos son generalmente de tamaño nanométrico(<300 nm de diámetro, la mayoría de 10-200 nm?) dominios que puedencontener proteínas integrales de membrana y periféricos. Las proteínassecuestrada en balsas de membrana pueden convertir estos dominiosen las estructuras de transducción de señales funcionales importantes enendocitosis, la regulación de la muerte celular, y otros eventos relevantespara la terapia del cáncer (34, 36).Proteínas integrales de membrana pueden interactuar dentro diferentedominios de la membrana, sino que también deben interactuar con la membranalípidos para producir una membrana plasmática intacta (8, 11, 15, 37).Específicamente, las porciones de sus estructuras deben emparejar directamente conlas cadenas acilo de fosfolípidos de la membrana o el hidrófoboporciones de otros lípidos de membrana. Esto se logracoincidente hidrofóbica (15, 29, 37). El concepto de hidrófobojuego entre el núcleo hidrófobo del lípidobicapa e hidrófobos tramos de aminoácidos en integralproteínas de membrana es esencial para la comprensión de cómo las célulasmembranas forman una estructura estable (11, 29, 38). Si el hidrófoboporciones de esta estructura son coincidentes, un elásticodistorsión de la matriz lipídica alrededor de la membrana integralproteína se produce (15, 29, 37). Esto puede producir proteína conformacionalcambios, la función de proteínas potencialmente efectuar yinteracciones proteína-proteína, tales como la agregación de proteínas enmembrana super-estructuras (37, 39). Además, hay otrofuerzas físicas, tales como fuerzas de presión laterales, fase lateralcambios, la curvatura de la membrana, interacciones iónicas, entre otrafuerzas, que deben ser considerados para producir un sin tensión globalestructura de la membrana (11, 39).Restricciones de membrana de la célula en la movilidad yOrganización jerárquicaLas restricciones a los movimientos laterales de membrana integralproteínas se han atribuido a restricciones extracelulares, tales comoECM, la formación de dominios de membrana especializadas (lípido

balsas de lípidos y dominios más grandes), grandes complejos de proteínas, ybarreras de membrana periférica en la superficie de la membrana interna(Fig 1A;.. Refs 11, 12). Jacobson y sus colegas (13) han resumidolos movimientos laterales de las proteínas de membrana en distintacategorías: (i) la reclusión transitoria por la proteína obstáculogrupos (postes de cerca también llamada proteína o piquetes); (Ii) transitoriaconfinamiento en dominios definidos o corrales por un citoesqueletomalla; (Iii) dirigida movimiento debido al apego al citoesqueleto;y (iv) la difusión aleatoria en la membrana de fluido.Por lo tanto, la descripción original de proteínas integrales de membranadifundir libremente en el plano de la membrana se refiere a uno solo deestas categorías (8).Ahora se cree que una porción sustancial de membrana integralproteínas no es capaz de difusión lateral libre en elmembrana celular; se limitan, al menos transitoriamente, a los pequeñosdominios de la membrana por barreras en la superficie de la membrana interior(11-14). Sin embargo, las proteínas integrales pueden escapar de una deestos dominios a un dominio adyacente. Incluso pueden escapar de ladominios por completo, a menos que se someten a la agregación y sutamaño impide movimientos extradomain. En consecuencia, las capacidadesde proteínas de membrana para moverse entre dominios adyacentespuede estar relacionada con sus tamaños, los tamaños de las barreras citoplasmáticos,y las complejas interacciones de estas barreras con el citoesqueleto(14) y ECM (11).Las áreas aproximadas de los dominios de los receptores de la membrana celularse han estimado para variar desde 0,04 hasta 0,24 mm2, y ellos tiempos de tránsito aproximados de receptores de membrana en estos membranadominios pueden variar de 3 a 30 segundos (14, 40). Por lo tanto, céluladominios de la membrana pueden variar en diámetro desde 2 hasta 300 nm. Porejemplo, los dominios del citoesqueleto-vallada que contiene actina haber sidoencontrado en diámetros aproximados de 40 a 300 nm, balsa lipídicadominios en el rango de 2 a 20 nm, y dinámica integralcomplejos de proteínas de membrana en los dominios de 3 a 10 nm ende diámetro (14, 40). La presencia de diferentes tipos de membrana de la céluladominios y la presencia de proteínas de membrana selectivaen estos dominios sugerir otro nivel de la composición de la membranacomplejidad más allá de la membrana de fluido de mosaico originalesestructura (11-14).Las membranas celulares deben responder rápidamente a intracelular yseñales extracelulares y otros eventos microambientales. Hacereste rápidamente, puede ser más eficiente tener receptores preposicionadaen la membrana celular dentro de los dominios de señalización de modo quepueden someterse a agregación rápida en la señalización supramolecularestructuras (14). La partición de las membranas plasmáticas para limitarla dinámica de sus componentes de proteínas de membrana integrales (enmenos parte del tiempo) para corrales citoesqueleto-vallado, o tetheringellos directa o indirectamente a citoesqueleto asociada a la membrana

elementos, pueden crear dominios de membrana relativamente estables dedensidades de receptores de aumento (40).La transducción de señales, la activación celular, la identificación, la diferenciación,y otros eventos de membrana iniciada complejos puederequerir la presencia de densidad de los receptores mejoradas dentro específicadominios de la membrana para promover selectivamente la señalización celular.Kusumi y sus colegas (14, 40) han propuesto que las célulasmembranas poseen arquitecturas jerárquicas que constan de unamembrana básica de fluido de mosaico plus Varios microand membranadominios de tamaño nanométrico o compartimentos definidos por citoesqueletoesgrima y proteínas postes de cerca, balsas de lípidos y otros lípidosdominios, dominios de lipoproteínas de membrana integral oligoméricos,y otras estructuras. Esta estructura compleja se representa de manera simplistaen la Fig. 1A. Dentro de la estructura jerárquica (incluyendodominios de la membrana) componentes proteicos están limitados en suvelocidades de difusión a los que son de cinco a 50 veces más lento que cuandolos mismos componentes son libres para difundir lateralmente en la membranasin restricción (41).Membrana Celular Fusion y VesículaTransporteLas células paquete recién materiales sintetizados para su transporte adiversos orgánulos y al exterior de las células mediante la incorporación de ellosen pequeñas vesículas de membrana que se entregan al objetivo específicodominios de la membrana (42). Este proceso también se usa para rápidamentereparar el daño a la membrana plasmática y otras membranas celulares(43). Las células tumorales invasoras utilizan exocitosis y otra dirigidavías para liberar enzimas degradadoras, biorreguladores, yotras moléculas en el microambiente para facilitar la invasiónde ECM y estroma de tejido y evadir las defensas del huésped (Figura 1;.. refs 44-46). También es un mecanismo que se utiliza para mostrar reciénreceptores sintetizados, enzimas y otras moléculas sobre lasuperficie exterior de la membrana celular y facilitar su volumen de negocios.Un evento crítico en la entrega de materiales a través de pequeñas vesículas esfusión de la membrana (47). Eventos de fusión de membrana también son evidentesa nivel celular, por ejemplo, cuando las células cancerosas adyacentessometerse a fusión con otras células cancerosas o células normales para producircélulas aneuploides, complementación, y otras característicasencontrado en las células neoplásicas progresivos (38, 48). La fusión de membranasobviamente no es un evento asociado al cáncer; se produce durante muchosprocesos normales celulares, tales como la fertilización, la formación de mioblastos,y la homeostasis ósea (38).El transporte de vesículas Dirigida y la fusión dentro de las células dependeen cierta medida de la composición de lípidos, distribución, y acilación.Por ejemplo, ciertos lípidos, tales como los esfingolípidos yesteroles encontrados en balsas de lípidos, se concentran en las vesículas destinadospara fusionarse con la membrana plasmática (42, 49, 50). Polifosfoinosítidos específicascon sus proteínas atados también pueden ayudarvesículas directos a sitios particulares de membrana (51, 52). Membrana

fusión depende de fusión especializada unión a la membranamaquinaria compuesta de proteínas específicas (SNARE, SNAP, ySM proteínas, entre otros) que tiran de las membranas adyacentesjuntos para promover la fusión bicapa lipídica (53).El ensamblaje de las proteínas fusogénicas en la membrana celularconstituye una microdominio membrana dinámica especializada llamadaun porosome (54). En algunas células normales, aparecen porosomesultraestructuralmente como "piscinas" de medición de 0,5 a 2 mm de diámetroque contiene depresiones de 100 a 180 nm (55). Porosomes sonresponsable de dirigir la exocitosis a sitios particulares en el celularsuperficie. Esto es importante para la migración celular y la invasión dirigidade ECM y el estroma, así como para la función normal de las célulassecretar necesarias proteínas, glicoproteínas, enzimas, biorreguladores,y otras moléculas importantes.Las membranas celulares y invadopodiosPara facilitar la invasión celular, la invasión de las células tumorales pueden extenderprotuberancias de la membrana de actina-ricos especializadas llamadas "invadopodios"que penetran en los alrededores de ECM, estroma, y el sótano(Fig. 1.; refs 56, 57) membranas. Estos célula especializadaestructuras exhiben y están asociados con degradación extracelularenzimas y contienen polímeros de actina intracelular y sureguladores, como cortactin, cofilina, N-WASP, Arp2 / 3, y facin(57-59). Los homólogos de células normales comparables de invadopodiosson llamados podosomes, y estas estructuras son evidentes enmuchas células normales en condiciones de ECM y el sótanoinvasión de la membrana durante la embriogénesis, la cicatrización de heridas,respuestas inflamatorias, y la regeneración de órganos (58, 59). Diferenciasencontrado entre invadopodios y podosomes son queque a menudo difieren en tamaño, forma, densidad y estabilidad en particular,invadopodios son normalmente estables durante períodos de tiempo mucho más largosen comparación con podosomes (59).Invadopodios son estimulados para formar por diversos factores. Estasincluir factores de crecimiento, transformación oncogénica, epithelial-transición mesenquimal (EMT), la hipoxia, receptores de adhesión,quimiocinas, y actividad de la enzima degradativa. Por lo generalrequerir apego inicial a ECM y la vinculación con el citoesqueletocomponentes para iniciar la formación (57-59). Beaty y Condeelis(60) han propuesto que hay cuatro etapas (presumiblemente después deApego ECM) que participan en la formación invadopodios: (i) el montajede una estructura de núcleo de actina (y proteína accesoria), (ii) la activaciónde quinasas asociadas, montaje de polímeros de actina, la contrataciónde los componentes de adhesión, y su vinculación transmembrana,(Iii) polimerización de la actina y la elongación y invadopodialestabilización, y (iv) de filamentos y microtúbulos intermediola contratación, el alargamiento de la estructura madura junto con ECMdegradación. Aunque existe cierta incertidumbre en cuanto a la exactaetapa en la que los receptores de adhesión son importantes en invadopodiosformación (inicial o en la maduración, etapas de elongación;

ref. 60), no hay duda de que la adhesión es un importante tempranoelemento del proceso invadopodios (56, 59, 61).Invadopodios puede requerir lípidos especializada membrana celulardominios para iniciar su organización. De hecho, los lípidos especializadobalsas se han encontrado en los sitios de formación invadopodios, ybalsas similares son conocidos por estar involucrados en vesículas de membranael tráfico, la exocitosis, y la polimerización de actina en la membrana celularsuperficie interior (62). Por lo tanto, la formación de microdominios de membranaes un aspecto importante de la formación de invadopodios ypodosomes y probablemente otros procesos que requieren dirigidosdistorsiones de membrana, adhesión, la fusión de vesículas, degradación de la matriz,y otros procesos.Para que se produzca la invasión de células, las células invasoras deben tener la capacidadpara degradar las barreras de la matriz y migrar a lo largo de las vías de invasióngenerada por la reestructuración y la destrucción de ECM. Por lo tanto,La degradación de ECM es un paso importante en el proceso metastásico(44, 61-64). Varias enzimas degradadoras de ECM se asocian conaparecen enzimas invadopodios aislado (56, 59), y degradativaspara ser lanzado por exocitosis cerca de sitios de invadopodios (61, 65).Además, las membranas celulares en los sitios de invadopodios parecenunirse y orientar mecánicamente componentes de la matriz aflojadosparalelo a las superficies celulares para ayudar en la generación de fuerza mecánicaen los túneles de ECM (61, 65). Por lo tanto, las membranas celulares, degradativaenzimas, y invadopodios puede mecánica y enzimáticamentereestructurar ECM para facilitar el movimiento celular y la invasiónmediante la remodelación de ECM en estructuras tubulares en el que las células cancerosaspuede invadir. Eventualmente movimientos celulares colectivas pueden formar en unaestructura multicelda masiva invasivo que penetra a lo largo de latubos generados por invadopodios y células entonces individuales (65).Las membranas celulares, vesículas extracelulares, yLos exosomasLas células tumorales liberan naturalmente pequeñas (0,1 a 2 mm de diámetro) extracelularvesículas de membrana (EV) derivados del plasma en ciernesmembranas y por separado aún más pequeño (<100 nm de diámetro)microvesículas llamadas exosomas de vesículas intracelulares exocitado(Fig. 1; refs 66, 67.). Las vesículas de membrana liberada puedecontener diversas moléculas, tales como pequeños fragmentos de ADN,microRNAs, proteínas (enzimas, biomoduladores y receptores),y los hidratos de carbono (67-69). Junto con su plasma enriquecidocomponentes de la membrana y diversos receptores de las células, que puedenmediar en una forma de comunicación por transferencia entre tumorlas células, dando como resultado el intercambio de materiales celulares. También puedemediar en la comunicación entre tumorales y normales células en elmicroambiente (67-69). Esto no es una propiedad única detumorales células vesículas liberadas de las células normales se encuentran enprácticamente todas líquido extracelular donde parecen desempeñar un papelen la comunicación normal de las células y la regulación de la inflamación,coagulación, el desarrollo y otros procesos fisiológicos normales

(69, 70). En varios tipos de cáncer, también pueden afectar tumorinteracciones con el microambiente y promover la progresión,angiogénesis, invasión y metástasis (69, 70-72).Hay varios factores que pueden influir en la liberación de EV y exosomas.Entre ellas se encuentran la energía celular, los niveles intracelulares dede calcio, los cambios en los fosfolípidos de membrana, y otros reguladoresde las interacciones del citoesqueleto de la membrana, la membrana de acciónenzimas, y otros efectores de la exocitosis, la hipoxia, yestrés oxidativo y de corte (67, 71). La liberación de EV y exosomastambién se ven afectados por invadopodios, que puede mejorarsu comportamiento invasivo de liberación y unidad del tumor (72). PublicadoEV y exosomas son bastante heterogéneos en su composicióny reflejar la célula a célula de las variaciones en los cargos, membranas, ycélulas de las que se derivan (70-73). De importancia esque la mayoría de las células normales y benignos no se liberan grandes cantidadesde EV y exosomas hasta que progresan a invasivo,fenotipos metastásico (73).EV y exosomas de células-lanzado tumor pueden tener profundosefectos sobre el microambiente del tumor (70-72). Además desu suministro de ácidos nucleicos, fosfolípidos bioactivos, degradativaenzimas, receptores, factores de crecimiento y de la motilidad, yotro tipo de carga que puede afectar a la invasión y colonización metastásica,estas vesículas también pueden estimular la angiogénesis, estromalreacciones, y la liberación de paracrina y otras moléculas bioactivasa partir de células circundantes normales que condicionan el tumormicroambiente (67, 70, 71, 73). Por ejemplo, la observaciónque la señalización microRNAs en microvesículas puede mejorarla migración de células endoteliales y promover la angiogénesis indicaque estas vesículas tienen importante la comunicación molecularPropiedades (74). Shed vesículas de membrana tumoral pueden romperabajo, liberando su contenido al medio ambiente extracelular,y esto tiene consecuencias importantes para la invasión y celularmotilidad. Un ejemplo es el de liberación EV de un derivado de tumorglicoproteína de membrana (EMMPRIN) que estimula los fibroblastosliberación de metaloproteinasas de la matriz que facilitan tumorla invasión y metástasis (75). Exosomas EV y también se puedeinvolucrados en las células linfoides de reclutamiento que estimulan tumorcrecimiento. Por ejemplo, el bloqueo de la secreción de exosome inhibiendoGTPasas pequeñas resultados en el crecimiento del tumor disminuyó y pulmónmetástasis en un modelo de cáncer de mama por la disminución de los neutrófilos estimulada-proliferación de células tumorales (76).EV y exosomas también pueden proteger neoplásica de células-lanzado Tumorlas células de los productos químicos nocivos como quimioterapéuticodrogas, fosfolípidos oxidados, la irradiación, las respuestas inmunes,y señales de muerte celular (67, 77). El tratamiento humano y animallíneas celulares de cáncer de pulmón con la irradiación y la hipoxia que no lo hacenestimular la apoptosis resultados en la mejora de la liberación de EV queactivar y estromal chemoattract y células endoteliales. Una vez

activado con EV derivado del tumor, las células del estroma luego sueltevarios factores pro-angiogénicos. Cuando esta célula estromal acondicionadoSe utilizaron medios de comunicación para estimular las células tumorales, el resultado fuepotencial mejorada metastásico in vivo (78).Recientemente, el uso de EV y exosomas para la modulación de lamicroambiente o la producción de nuevos agentes terapéuticos para la orientaciónde moléculas bioactivas específicas a sitios específicos, tales como el cerebro, tieneha propuesto (79). Ya sea que estos enfoques resultan útilespermanecen clínicamente por verse, pero este nuevo enfoque podría seruna interesante forma de dirigir fármacos a sitios secundarios.Las membranas de la célula y la InvasivaFenotipoLa transición de un fenotipo epitelial a mesenquimal celularo EMT en células de carcinoma se ha propuesto como uno de los llamativacambios que acompañan a la invasión y metástasis (80, 81). Diferente Alos múltiples cambios genéticos que son típicos de la transformación in vivoy la tumorigénesis, EMT parece ser principalmente epigenéticay conducido en su mayor parte por las señales microambientales(82). Los cambios que se producen, como la pérdida de la adhesión celular yuniones celulares, modificaciones en la forma celular, la adquisición de la célulamotilidad, la liberación de factores de crecimiento y de la motilidad y degradanteenzimas, entre otros cambios, son característicos de invasiva,células malignas en el sitio primario (Fig 1;.. refs 80, 81, 83). Enmenos tres vías de regulación polaridad celular de control diferente,adherencia, uniones celulares, y otras propiedades relacionadas (82). Estasvías son impulsados por microambiente tumoral diafonía yeventos de señalización a partir de la superficie de la membrana celular ya continuación, a lo largo de los circuitos de señalización pertinentes dentro de las células (83). Por otra parte,una vez que las células de carcinoma maligno han hecho metástasis a secundariasitios, pueden parecer volverá a epitelial similarmorfologías junto con reexpresión de marcadores epiteliales,lo que indica que EMT tiene transitoria y potencialmente reversiblecaracterísticas (80, 81). Aunque los cambios de EMT se han asociadocon la invasión y metástasis en tumores animales, haysigue siendo un debate en curso sobre si este fenómeno es verdaderamenterepresentante de las vías malignos en los cánceres humanos (84).De hecho, Tarin (85) ha argumentado que los fenotipos de células invasorasocurrir después de los tejidos normales se dañan y durante la embriogénesisy que existe una falta de evidencia patológica convincente enlos seres humanos que EMT se produce cuando carcinomas metastatizan.Un nuevo enfoque para el desarrollo de terapias para altamente malignocélulas de carcinoma ha sido la de dirigir la metástasis relacionadacambios, tales como componentes de la superficie celular que participan en uniones celulares,de adhesión, la motilidad, el crecimiento, la señalización, y otras firmasde un fenotipo invasivo. Un lugar que esto podría seres importante en la identificación y tipificación de circulantelas células tumorales (CTC; Ref. 86). Enumeración de CTC y escribiendo

Biomarcadores de CTC se están desarrollando para evaluar el riesgo de metástasisla enfermedad y es de esperar predicen los efectos de la terapia para prevenirmetástasis (87). CTC se cree que están directamente relacionados con vástagocélulas, la fuente presuntivo de metástasis, así Zhang ycolegas (88) aislaron CTC de pacientes con cáncer de mamasin metástasis cerebrales, ellos crecieron en la cultura, y se someteellos a la metástasis ensayos en ratones inmunodeprimidos. Ellosencontrado que CTC de pacientes con cáncer de mama podría serseleccionado para una "firma cerebro de metástasis" único (EpCAM /HER2þ / EGFRþ / HPSEþ / Notch1þ) que puede explicar, en parte, lacapacidad del CTC para formar metástasis cerebrales. Utilizando el no seleccionadoCTC EpCAM de los padres y el cerebro de metástasis markerselectedCTC, se encontró que sólo estos últimos eran altamenteinvasivas y capaces de formar metástasis cerebrales cuando xenographeden ratones desnudos (88). Aunque este informe involucradosSólo unos pocos pacientes, los estudios futuros pueden ser capaces de desbloquear laFirmas CTC de metas para la metástasis de secundaria específicasitios. Tal información podría ser más útil quebiopsias en la predicción de la enfermedad metastásica, especialmente sitio específicometástasis, y, finalmente, la prevención de metástasis de formacióny el tratamiento de las metástasis que se han formado (86, 87).Comentarios FinalesSi bien se han logrado enormes progresos en los últimosdécada en la determinación de los papeles estructurales y funcionales de la célulamembranas en procesos malignos, queda mucho por descubrira nivel celular y molecular. En particular, se sabe muy poco acerca de cómo las señales microambientales y diafoníapuede cambiar rápidamente los fenotipos de cáncer y células normalesdentro del microambiente tumoral y cómo varios de señalizaciónvías, desde la membrana celular a diversos orgánulos celulares,y viceversa, controlar estas interacciones complejas. Descodificación delplasticidad de este proceso puede ser esencial en metastásico explicarcomportamiento (89).Estamos empezando a comprender la superficie celular ypropiedades circundantes de células malignas (y las células normales)que son importantes en la explicación de la metástasis a sitios secundariosasí como las propiedades de los órganos diana para la colonización metastásica.Esta información no será fácil próxima, peroserá esencial en el desarrollo final de la nueva terapéuticaenfoques para limitar o destruyen metástasis. También seráimportante en la reducción de los síntomas del cáncer y la eliminaciónlos efectos adversos de la terapia del cáncer (90).