Embed Size (px)
Citation preview
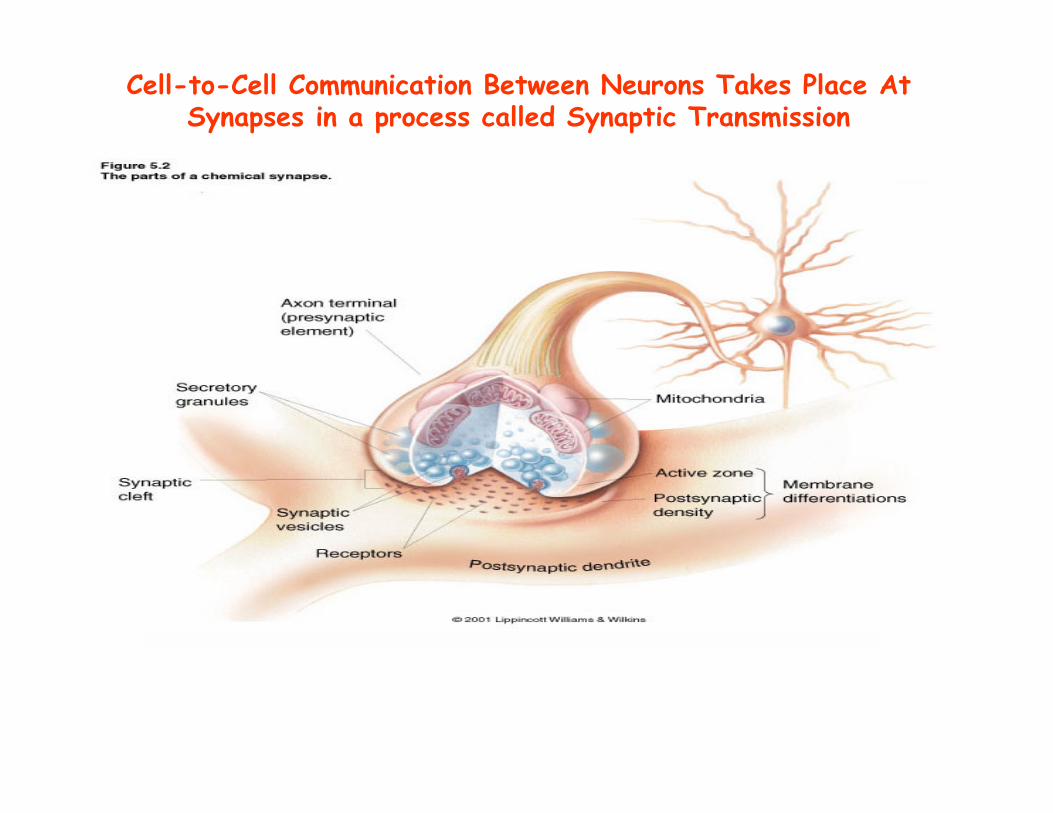
Cell-to-Cell Communication Between Neurons Takes Place At Synapses in a process called Synaptic Transmission
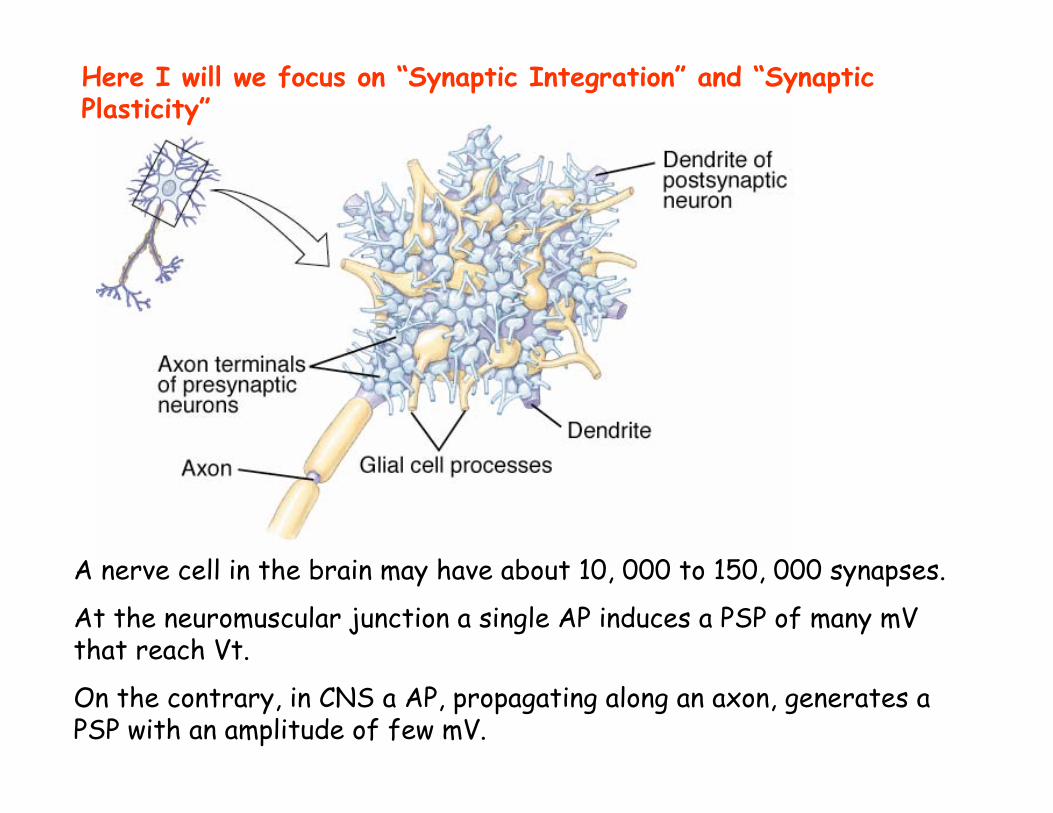
A nerve cell in the brain may have about 10, 000 to 150, 000 synapses.
At the neuromuscular junction a single AP induces a PSP of many mV that reach Vt.
On the contrary, in CNS a AP, propagating along an axon, generates a PSP with an amplitude of few mV.
Here I will we focus on “Synaptic Integration” and “Synaptic Plasticity”
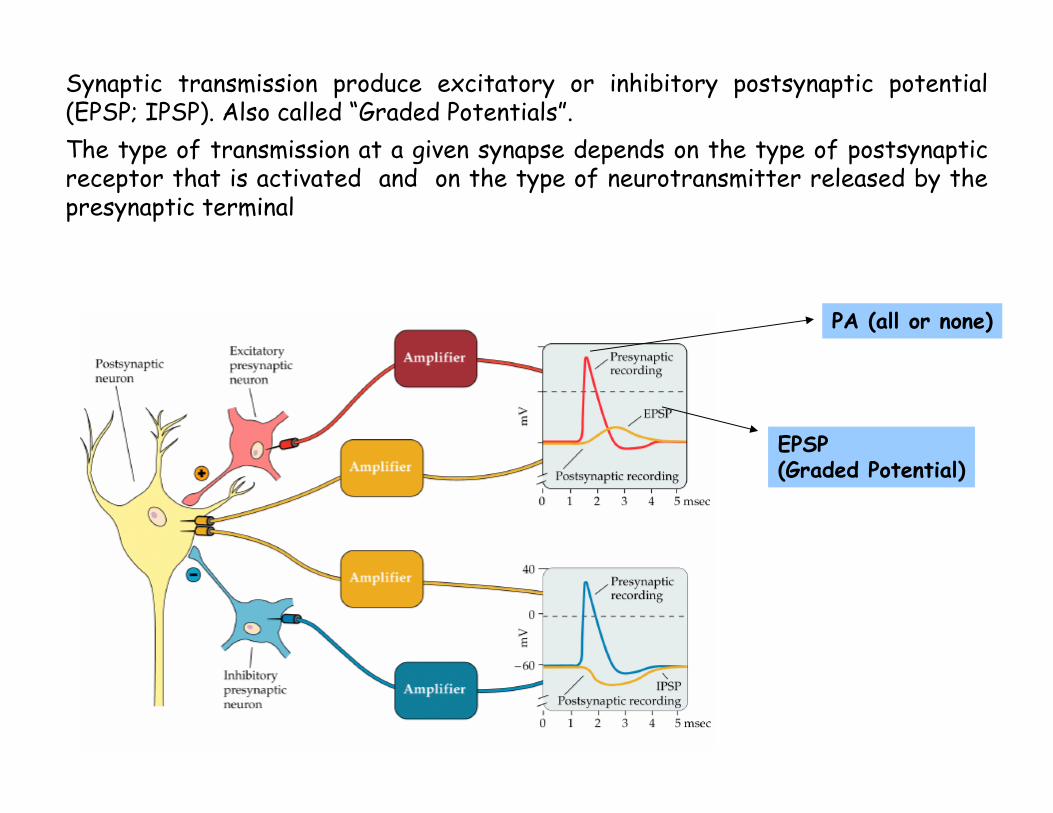
Synaptic transmission produce excitatory or inhibitory postsynaptic potential(EPSP; IPSP). Also called “Graded Potentials”.The type of transmission at a given synapse depends on the type of postsynaptic receptor that is activated and on the type of neurotransmitter released by the presynaptic terminal
PA (all or none)
EPSP (Graded Potential)
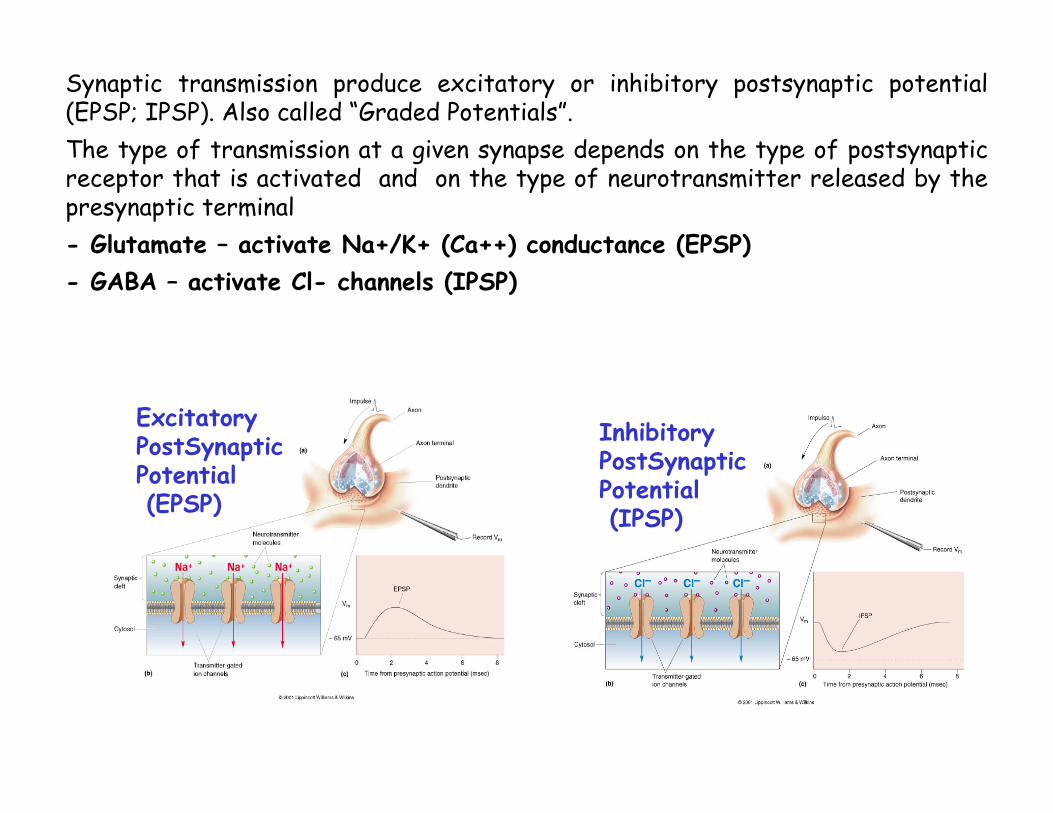
Excitatory PostSynapticPotential(EPSP)
Inhibitory PostSynapticPotential(IPSP)
Synaptic transmission produce excitatory or inhibitory postsynaptic potential(EPSP; IPSP). Also called “Graded Potentials”.The type of transmission at a given synapse depends on the type of postsynaptic receptor that is activated and on the type of neurotransmitter released by the presynaptic terminal- Glutamate – activate Na+/K+ (Ca++) conductance (EPSP)- GABA – activate Cl- channels (IPSP)
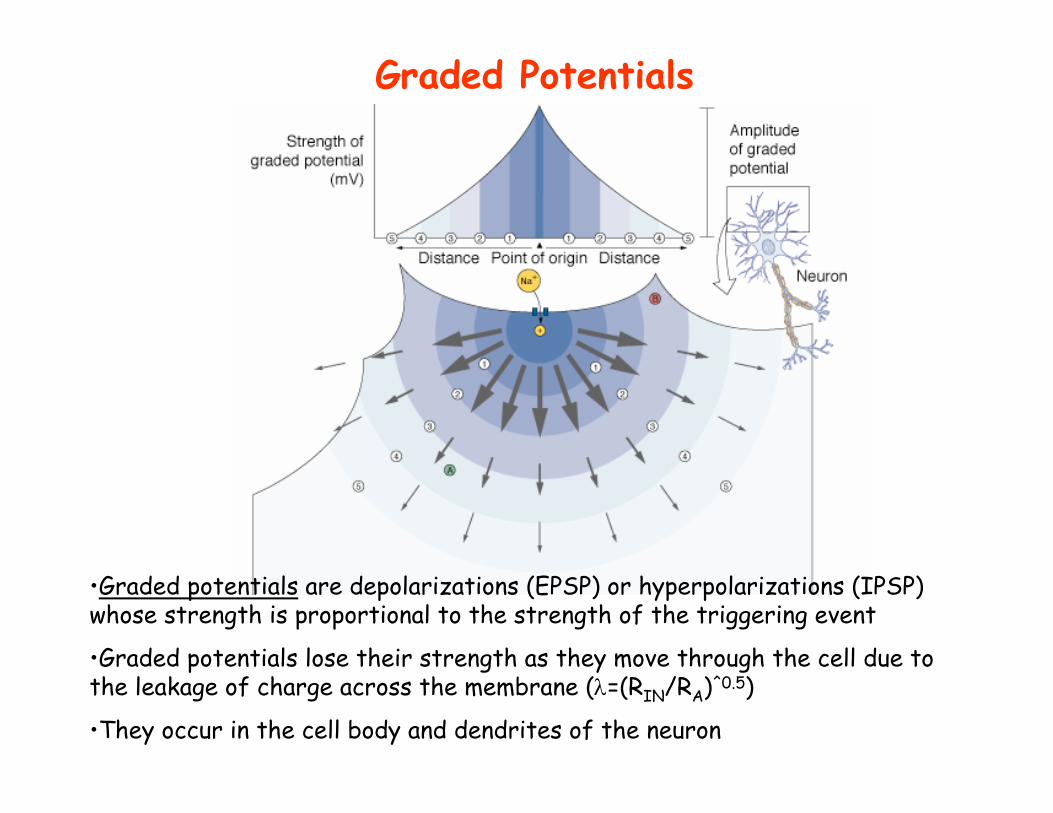
Graded Potentials
•Graded potentials are depolarizations (EPSP) or hyperpolarizations (IPSP) whose strength is proportional to the strength of the triggering event
•Graded potentials lose their strength as they move through the cell due to the leakage of charge across the membrane (λ=(RIN/RA)^0.5)
•They occur in the cell body and dendrites of the neuron

• Graded potentials travel through the neuron until they reach the trigger zone. If they depolarize the membrane above threshold voltage(about -55 mV in mammals), an action potential is triggered and it travels down the axon.
Graded Potentials Above Threshold Voltage Trigger Action Potentials
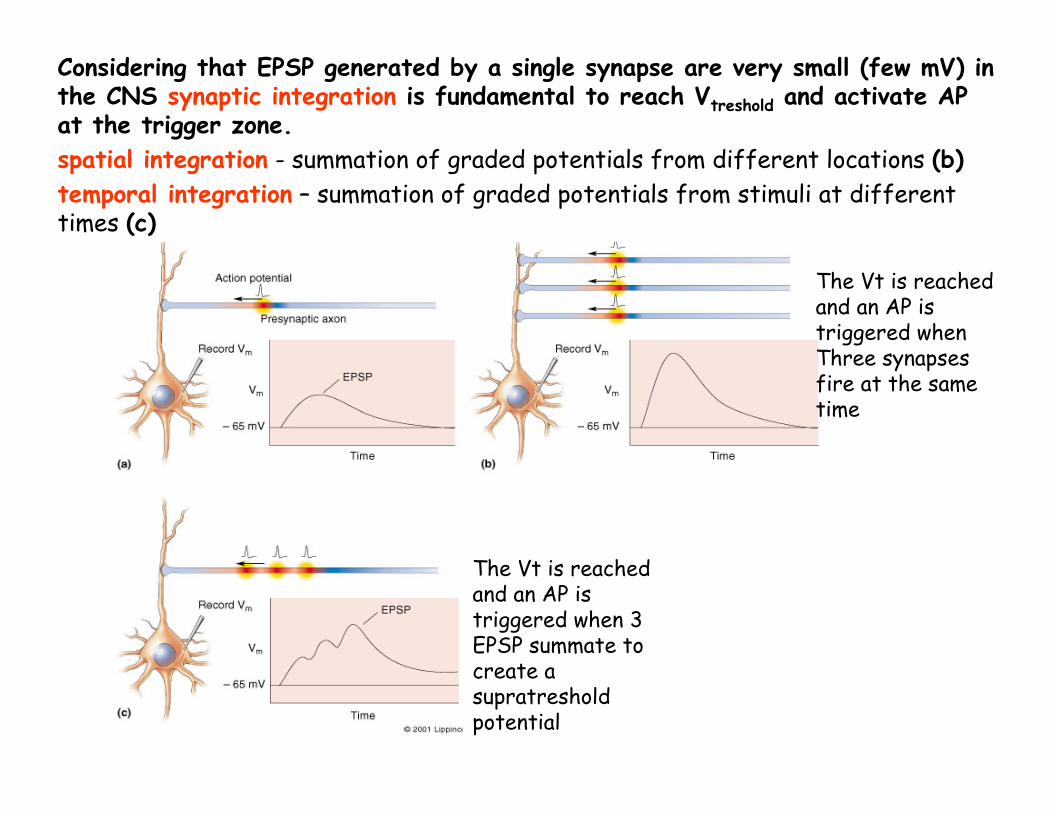
Considering that EPSP generated by a single synapse are very small (few mV) in the CNS synaptic integration is fundamental to reach Vtreshold and activate AP at the trigger zone.spatial integration - summation of graded potentials from different locations (b)temporal integration – summation of graded potentials from stimuli at different times (c)
The Vt is reachedand an AP is triggered when 3 EPSP summate to create a supratreshold potential
The Vt is reachedand an AP is triggered when Three synapses fire at the sametime
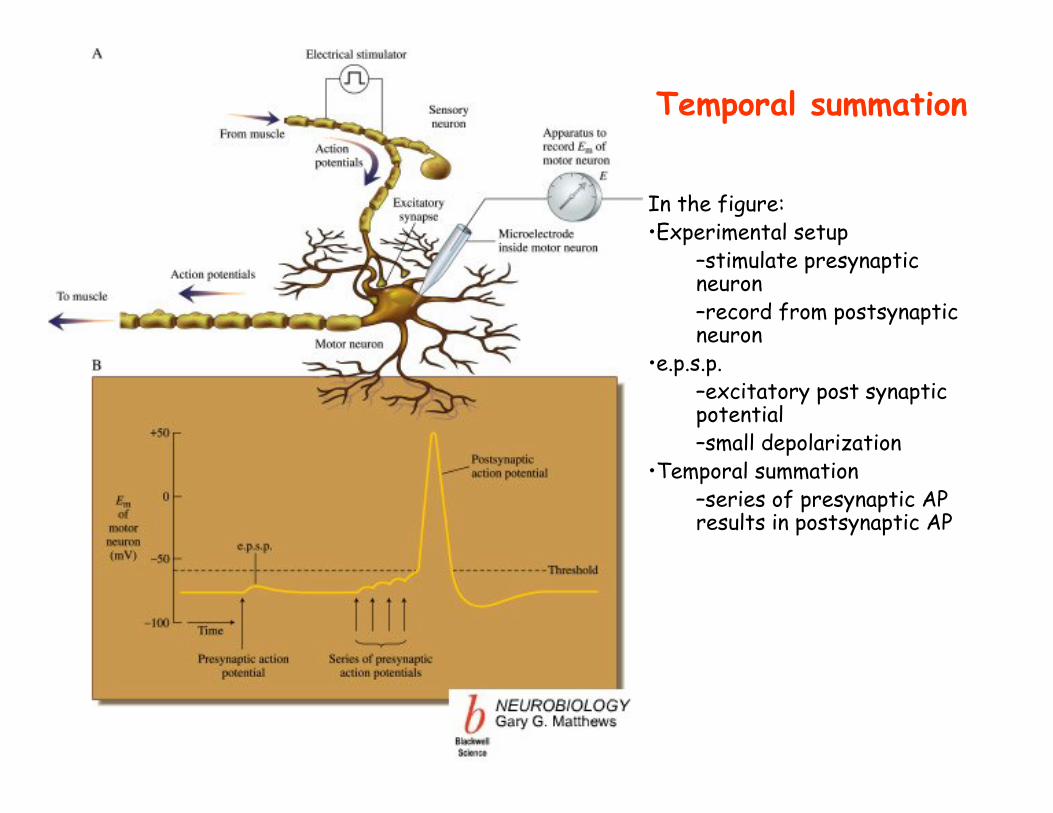
In the figure:•Experimental setup
–stimulate presynapticneuron–record from postsynaptic neuron
•e.p.s.p.–excitatory post synaptic potential–small depolarization
•Temporal summation–series of presynaptic AP results in postsynaptic AP
Temporal summation
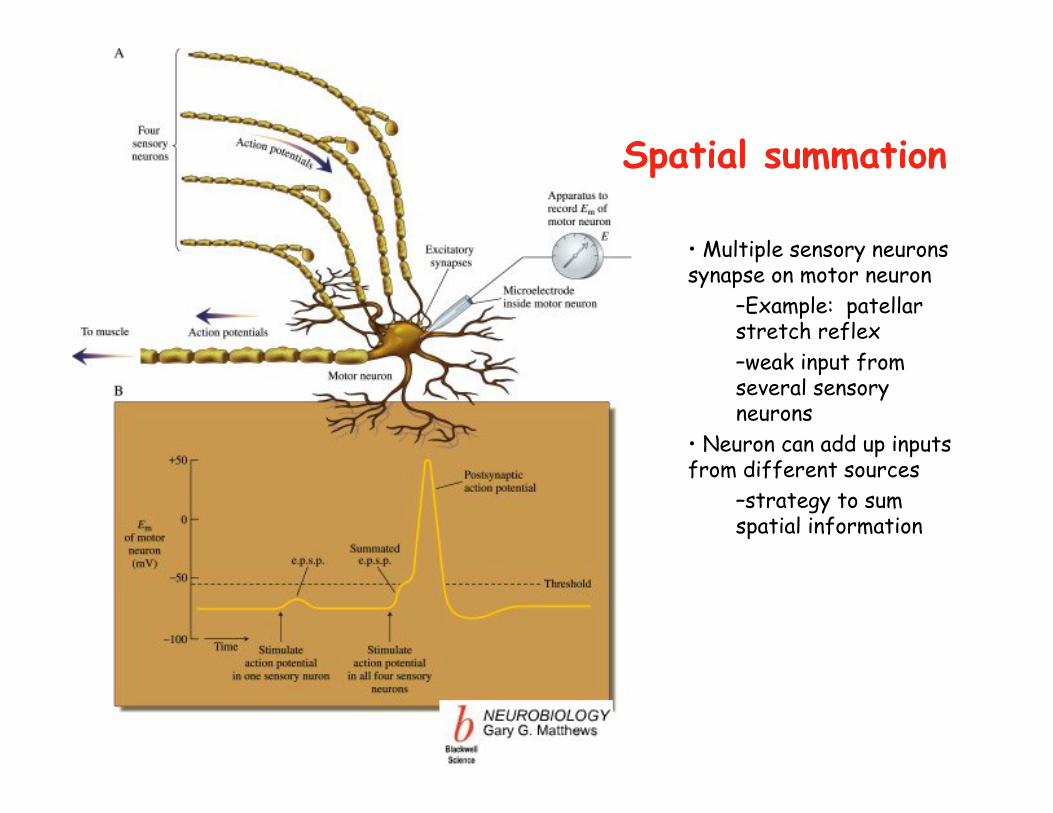
• Multiple sensory neurons synapse on motor neuron
–Example: patellar stretch reflex–weak input from several sensory neurons
• Neuron can add up inputs from different sources
–strategy to sum spatial information
Spatial summation
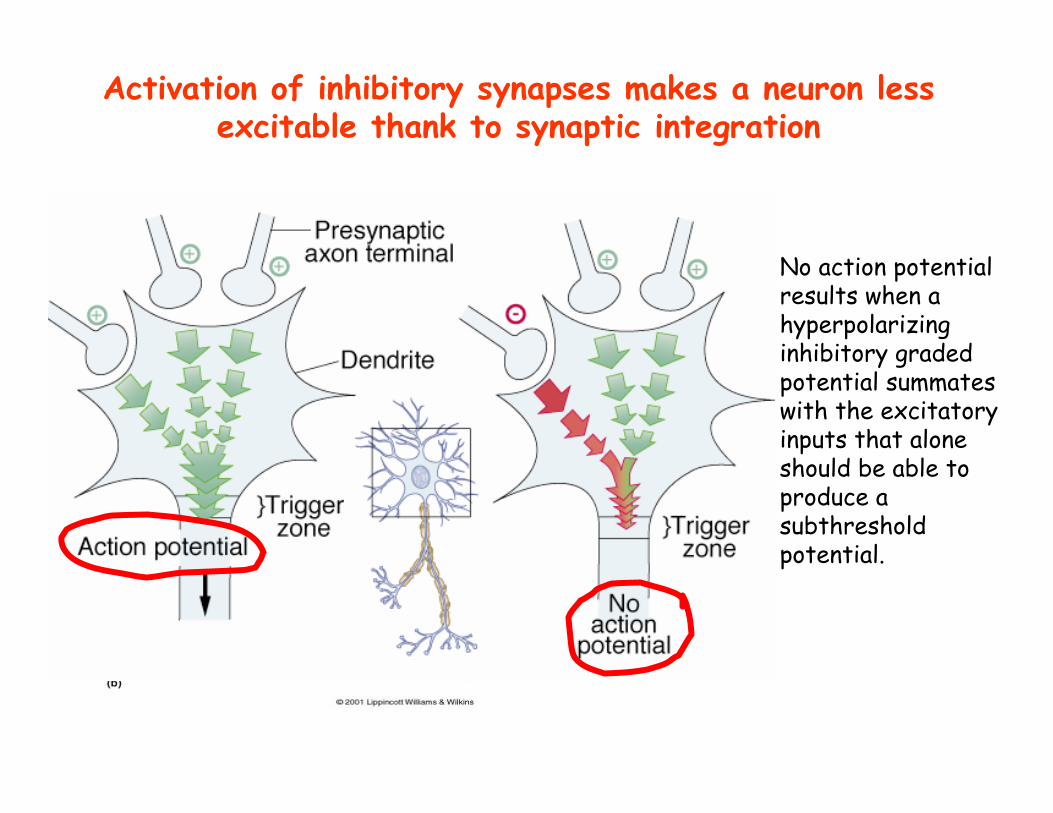
No action potential results when a hyperpolarizing inhibitory graded potential summates with the excitatory inputs that alone should be able to produce a subthresholdpotential.
Vt
Activation of inhibitory synapses makes a neuron less excitable thank to synaptic integration
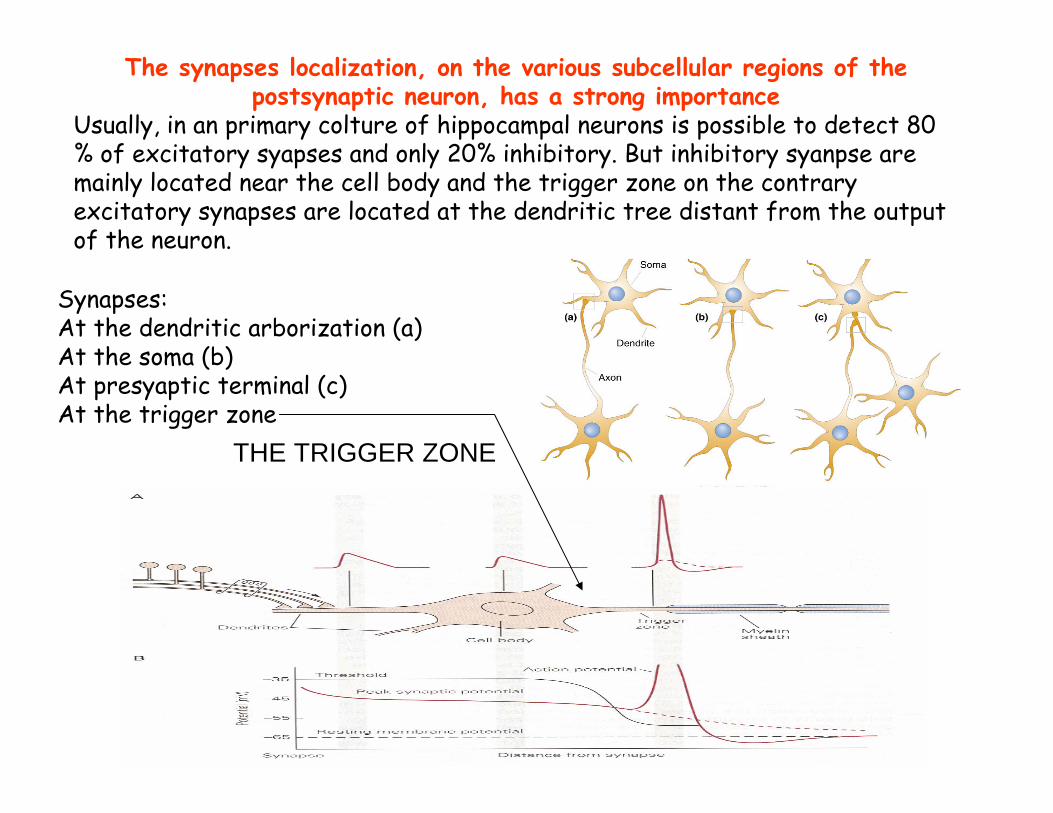
Synapses:At the dendritic arborization (a)At the soma (b)At presyaptic terminal (c)At the trigger zone
THE TRIGGER ZONE
The synapses localization, on the various subcellular regions of the postsynaptic neuron, has a strong importance
Usually, in an primary colture of hippocampal neurons is possible to detect 80 % of excitatory syapses and only 20% inhibitory. But inhibitory syanpse are mainly located near the cell body and the trigger zone on the contrary excitatory synapses are located at the dendritic tree distant from the output of the neuron.

Synaptic transmission is plastic
The ability to learn and form memories, is based on the ability of neurons to change the way in which they communicate with each other- that is, through synaptic plasticity
Plastic means that synapses function and structure can be moulded and changed in anuse-dependent way.
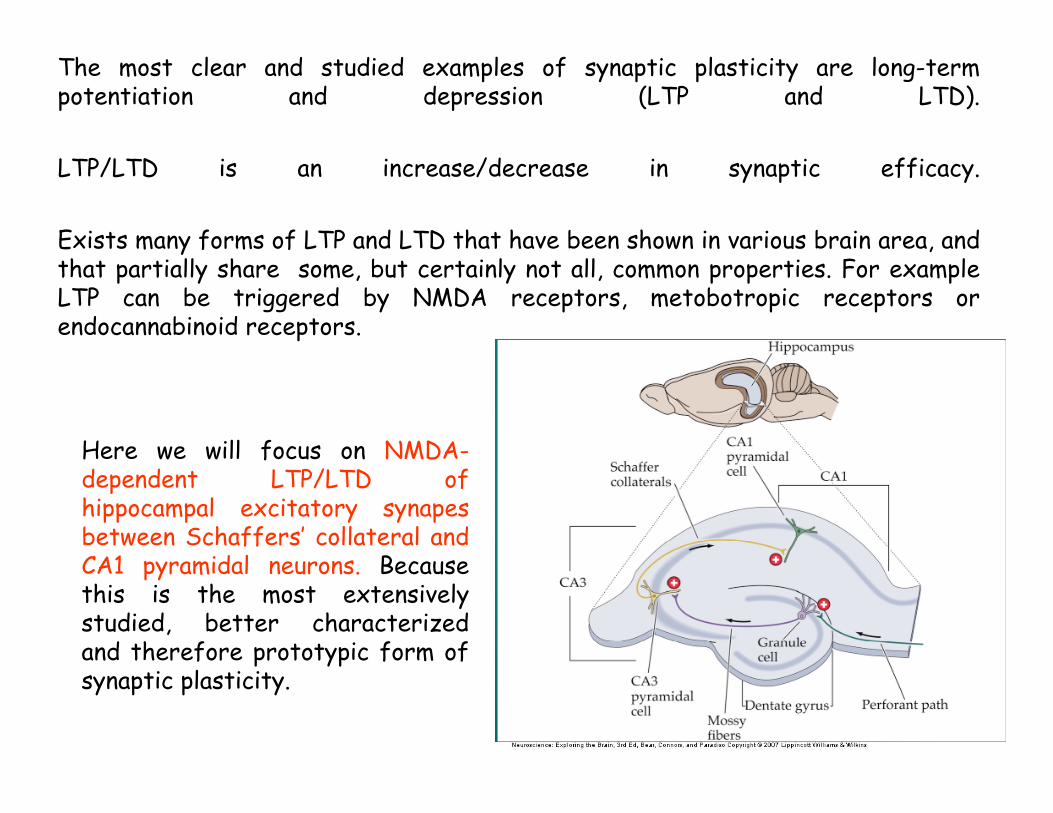
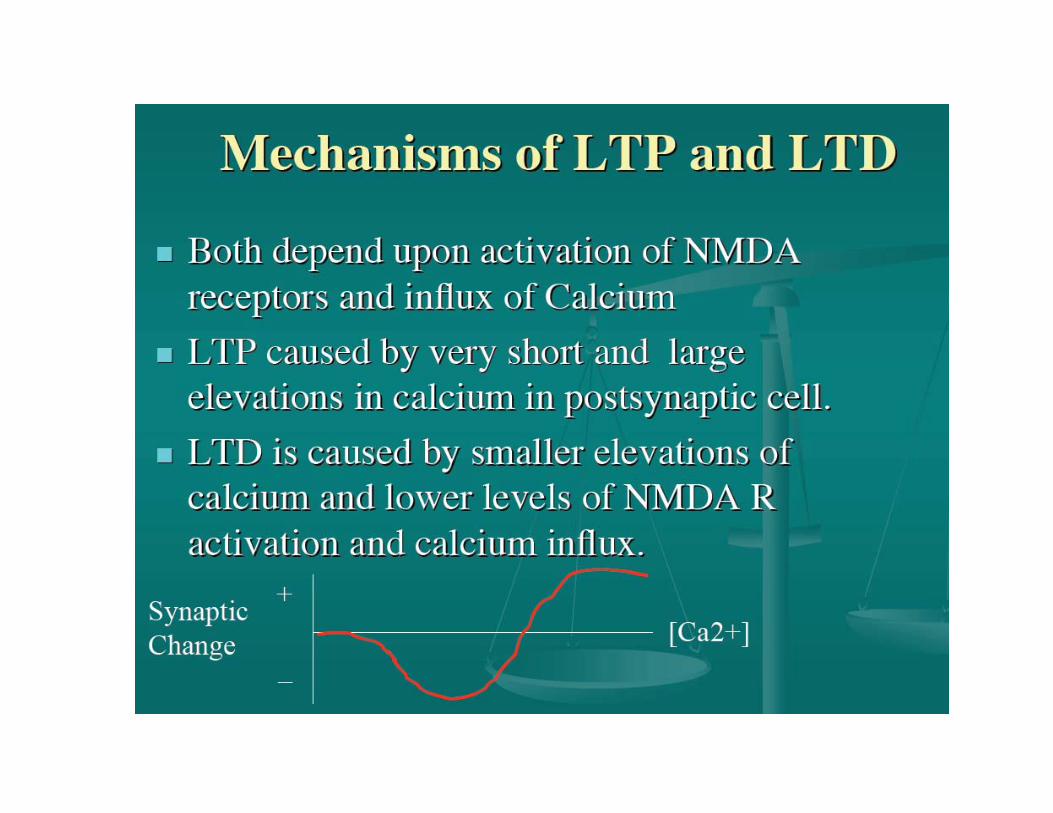
The most clear and studied examples of synaptic plasticity are long-term potentiation and depression (LTP and LTD).
LTP/LTD is an increase/decrease in synaptic efficacy.
Exists many forms of LTP and LTD that have been shown in various brain area, and that partially share some, but certainly not all, common properties. For example LTP can be triggered by NMDA receptors, metobotropic receptors or endocannabinoid receptors.
Perforant path
Mossy fibers
Schaffer collateral
Here we will focus on NMDA-dependent LTP/LTD of hippocampal excitatory synapesbetween Schaffers’ collateral and CA1 pyramidal neurons. Because this is the most extensively studied, better characterized and therefore prototypic form of synaptic plasticity.

• epilepsy surgery in patient H.M. • studied by Brenda Milner and colleagues for over 40 years.• declarative memory impaired• procedural memory fine
Why LTP/LTD are better characterized in the hippocampus?Because the hippocampus was the first brain area that was proved necessary for some forms of learning and memory
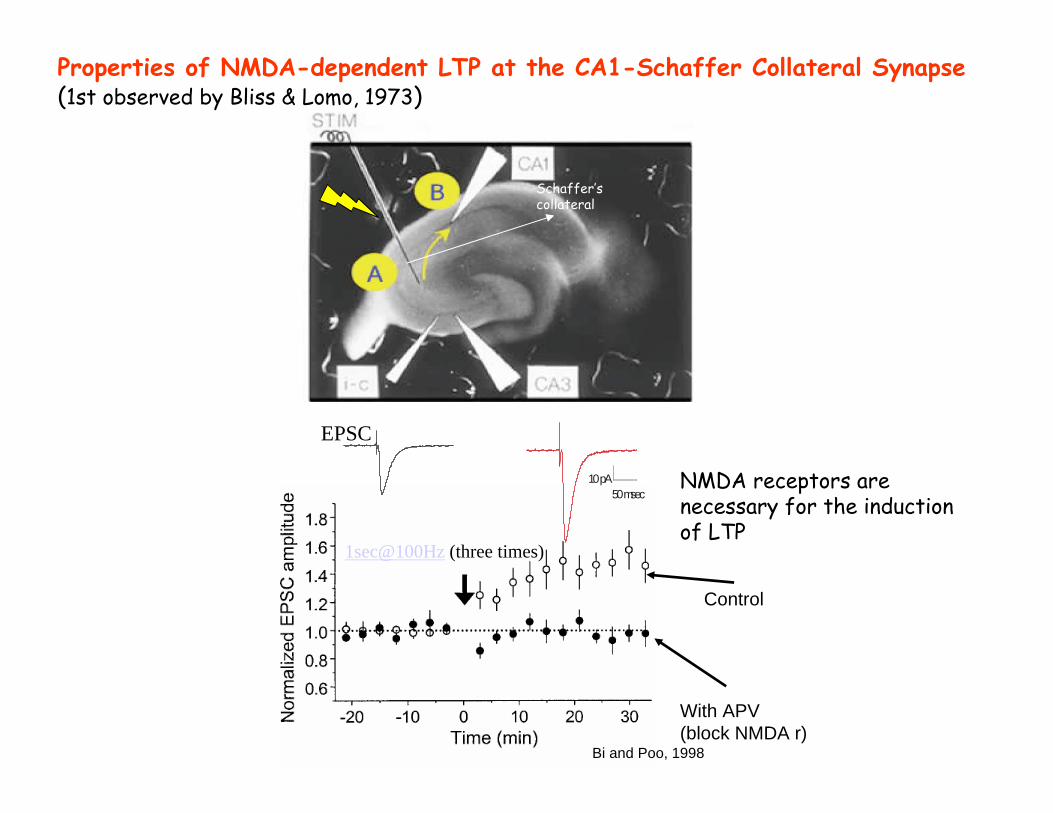
Properties of NMDA-dependent LTP at the CA1-Schaffer Collateral Synapse(1st observed by Bliss & Lomo, 1973)
NMDA receptors are necessary for the induction of LTP
Bi and Poo, 1998
Control
With APV(block NMDA r)
10pA50msec
EPSC
1sec@100Hz (three times)
Schaffer’s collateral
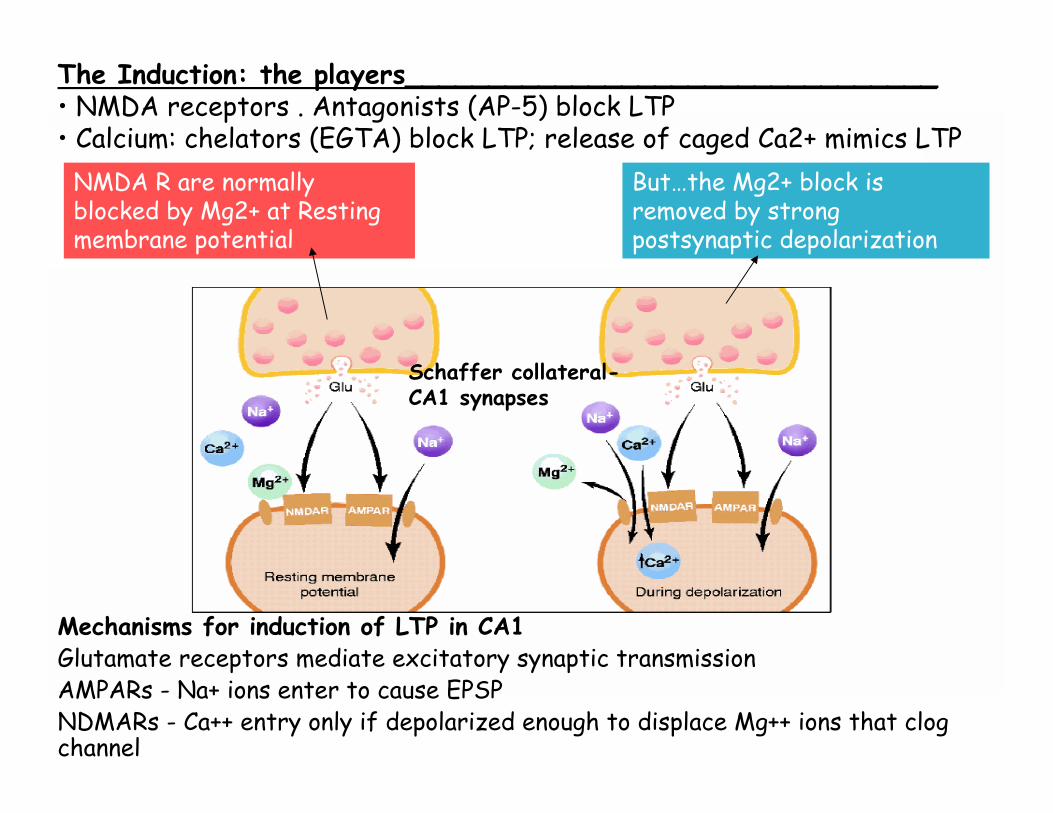
NMDA R are normally blocked by Mg2+ at Resting membrane potential
But…the Mg2+ block is removed by strong postsynaptic depolarization
Schaffer collateral-CA1 synapses
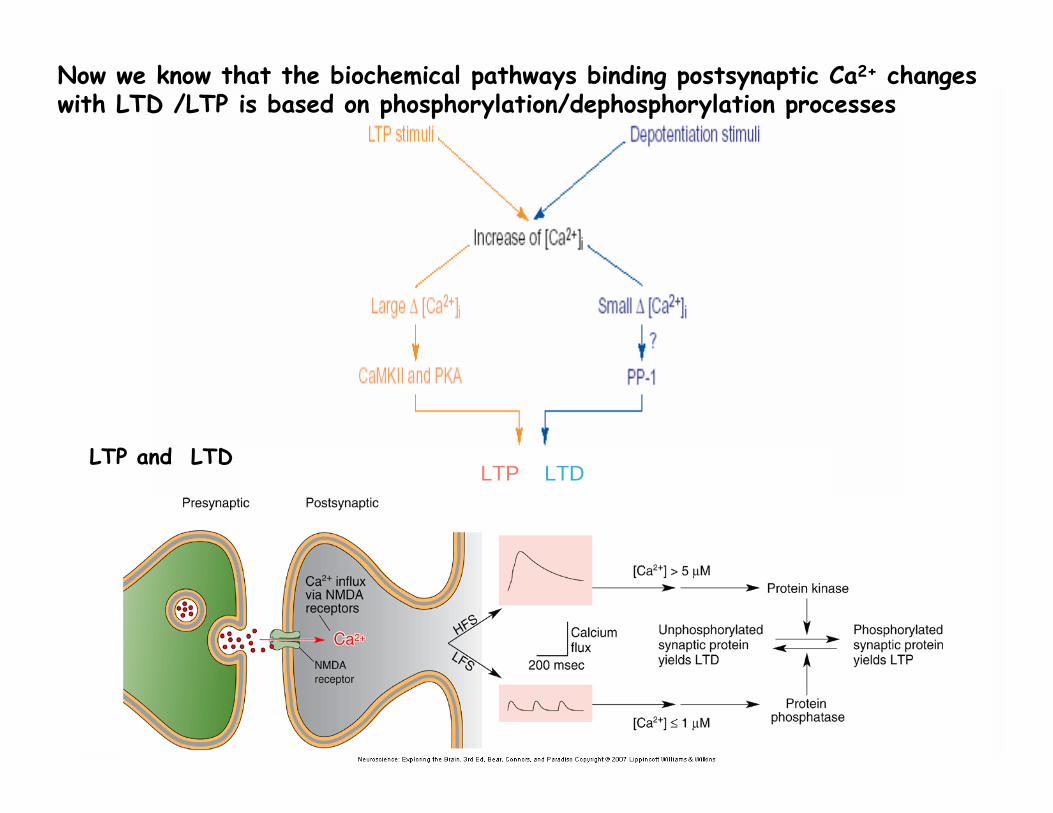
The Induction: the players________________________________ • NMDA receptors . Antagonists (AP-5) block LTP• Calcium: chelators (EGTA) block LTP; release of caged Ca2+ mimics LTP
Mechanisms for induction of LTP in CA1Glutamate receptors mediate excitatory synaptic transmissionAMPARs - Na+ ions enter to cause EPSPNDMARs - Ca++ entry only if depolarized enough to displace Mg++ ions that clog channel
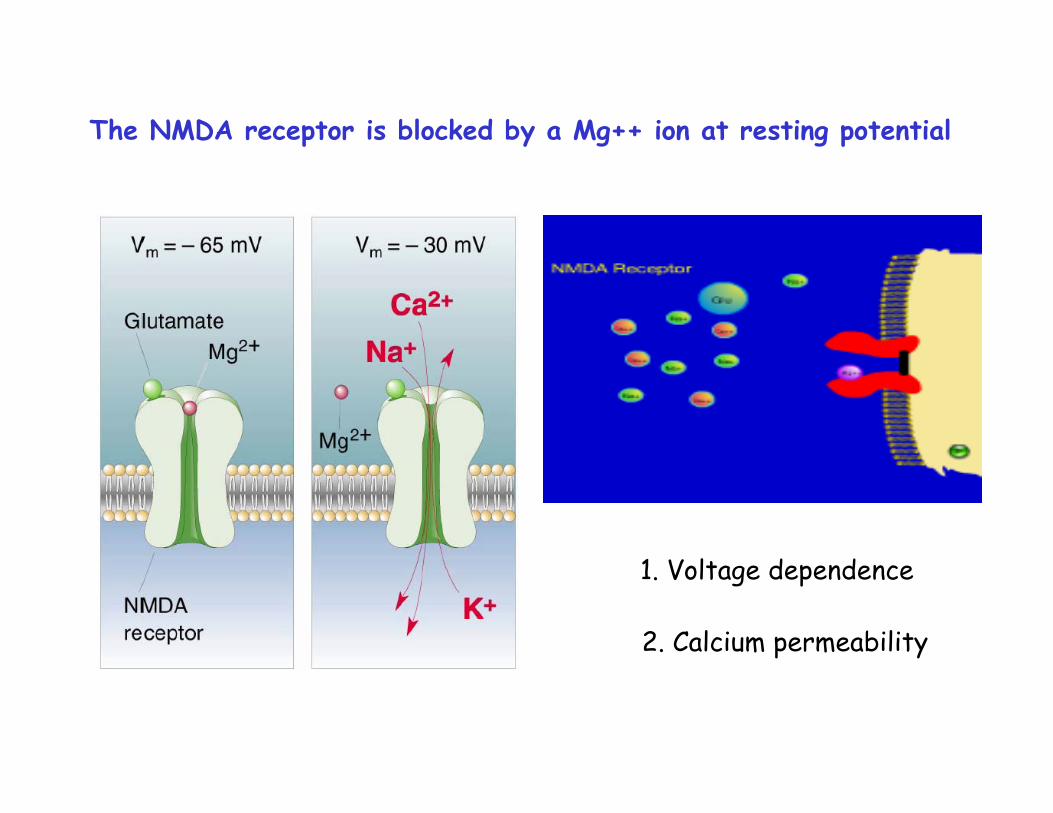
The NMDA receptor is blocked by a Mg++ ion at resting potential
1. Voltage dependence
2. Calcium permeability
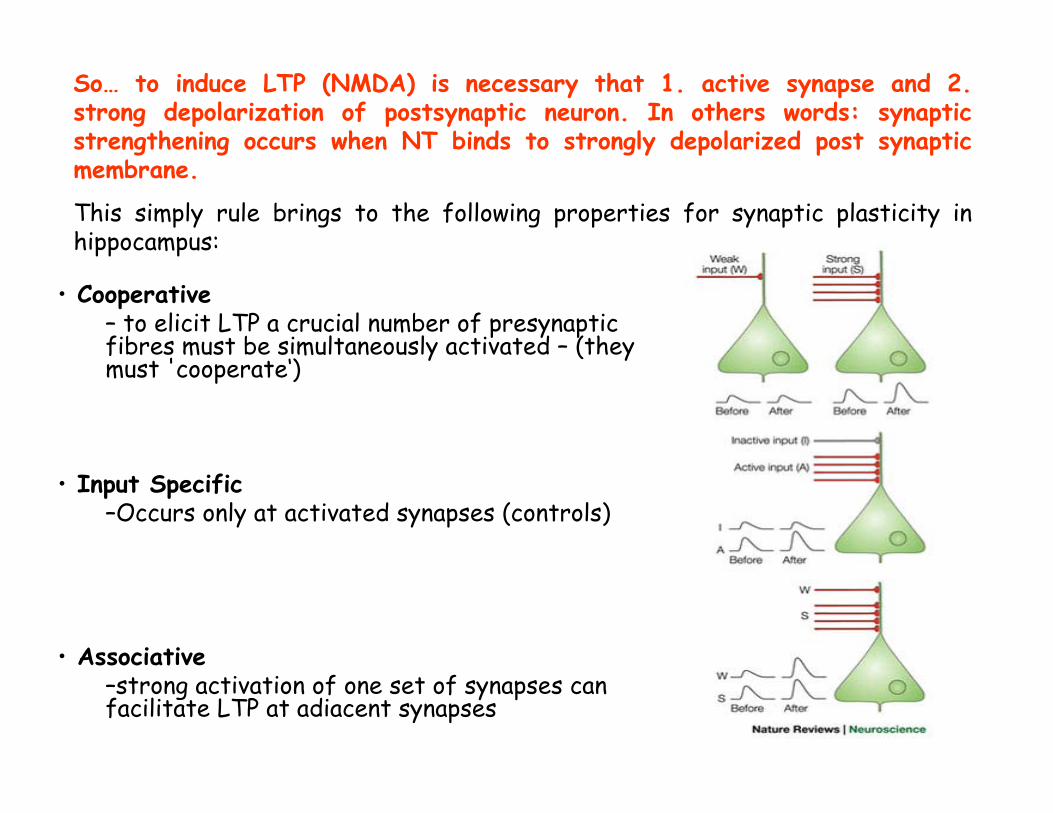
• Cooperative– to elicit LTP a crucial number of presynaptic fibres must be simultaneously activated – (they must 'cooperate‘)
• Input Specific–Occurs only at activated synapses (controls)
• Associative–strong activation of one set of synapses can facilitate LTP at adiacent synapses
So… to induce LTP (NMDA) is necessary that 1. active synapse and 2.strong depolarization of postsynaptic neuron. In others words: synaptic strengthening occurs when NT binds to strongly depolarized post synaptic membrane.
This simply rule brings to the following properties for synaptic plasticity in hippocampus:
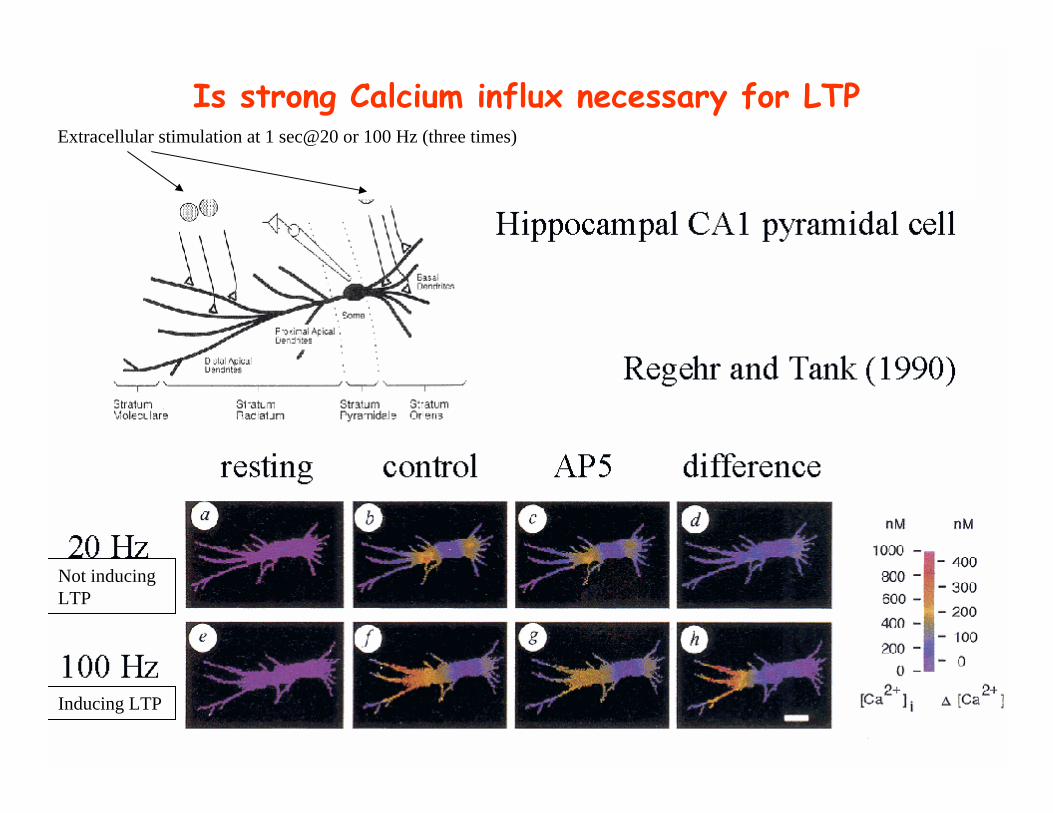
Is strong Calcium influx necessary for LTP
Not inducingLTP
Inducing LTP
Extracellular stimulation at 1 sec@20 or 100 Hz (three times)
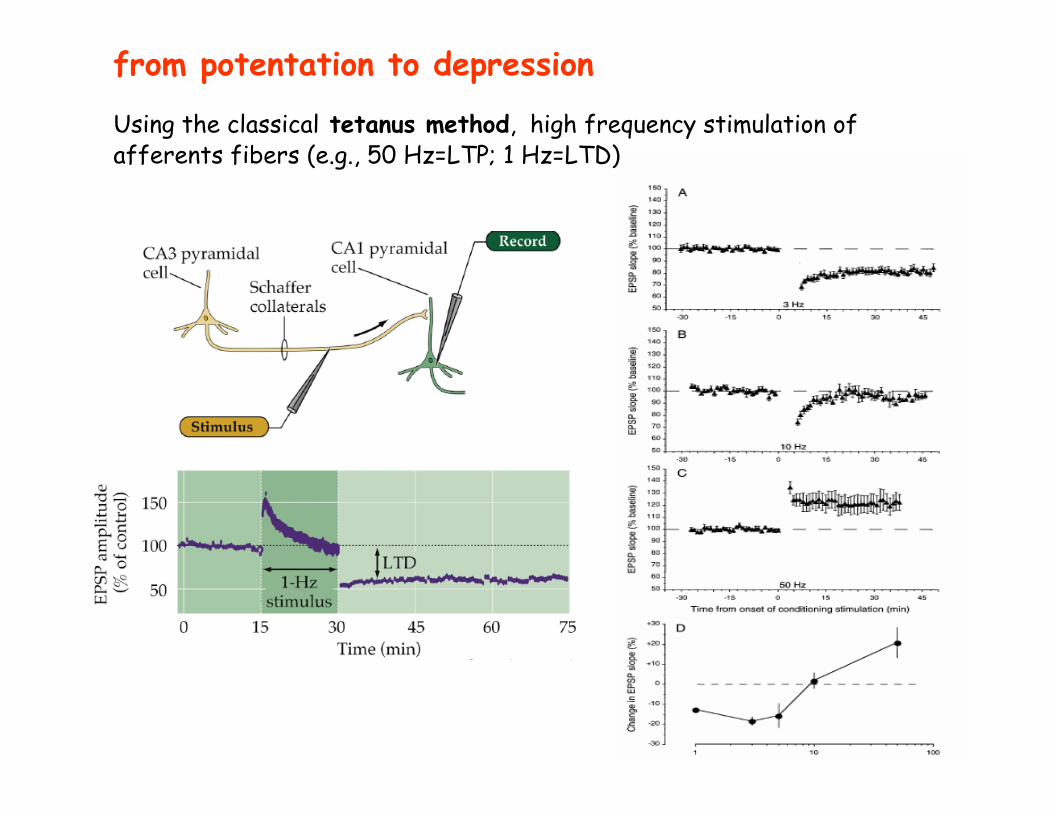
from potentation to depression
Using the classical tetanus method, high frequency stimulation of afferents fibers (e.g., 50 Hz=LTP; 1 Hz=LTD)
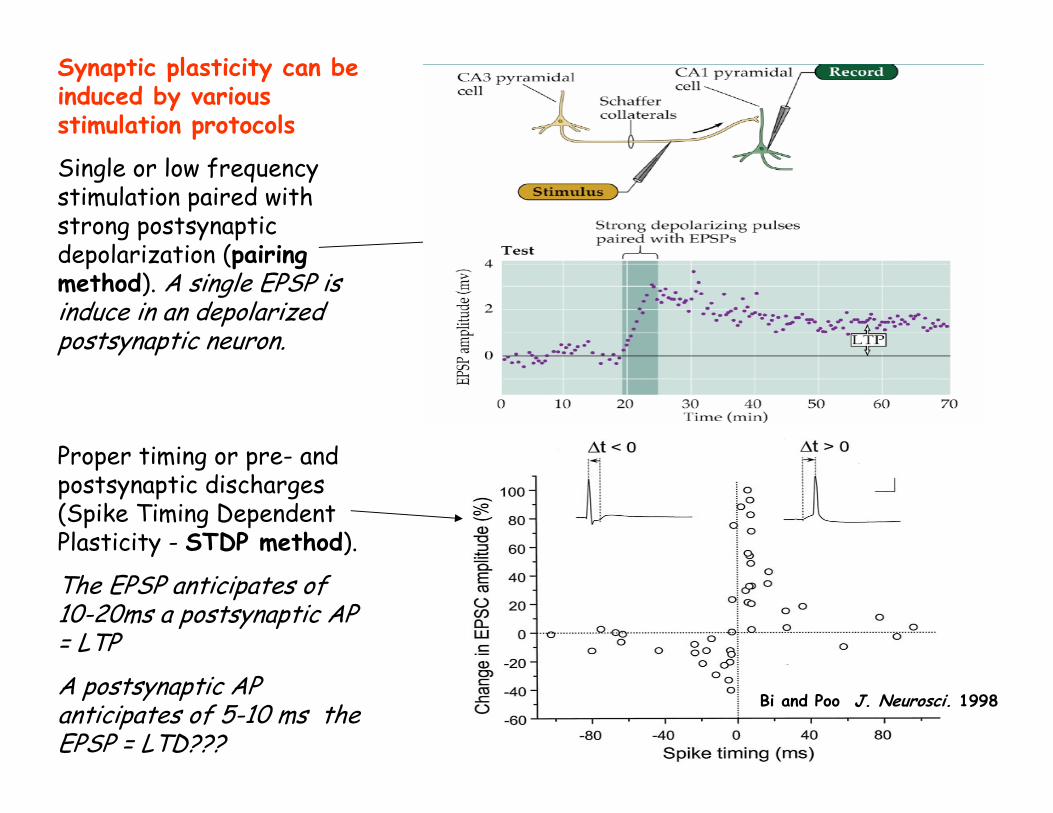
Synaptic plasticity can be induced by various stimulation protocols
Single or low frequency stimulation paired with strong postsynaptic depolarization (pairing method). A single EPSP is induce in an depolarized postsynaptic neuron.
Proper timing or pre- and postsynaptic discharges (Spike Timing Dependent Plasticity - STDP method).
The EPSP anticipates of 10-20ms a postsynaptic AP = LTPA postsynaptic AP anticipates of 5-10 ms the EPSP = LTD???
Pairing induced plasticity
Bi and Poo J. Neurosci. 1998
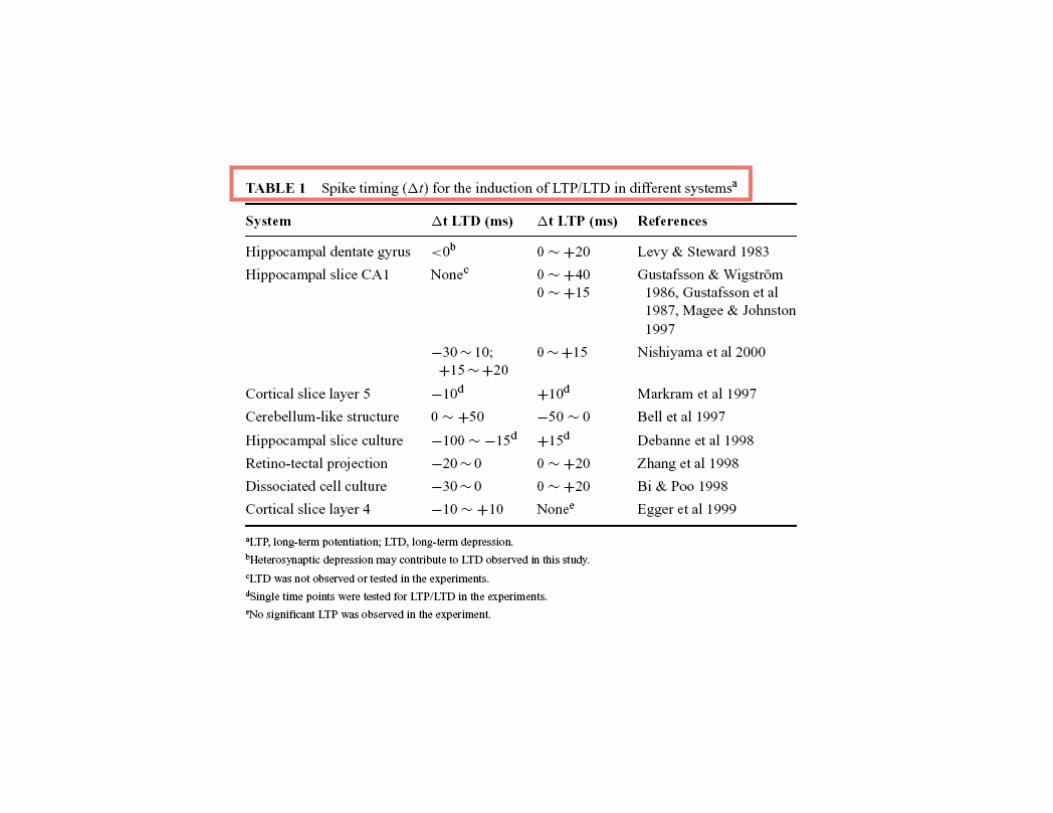
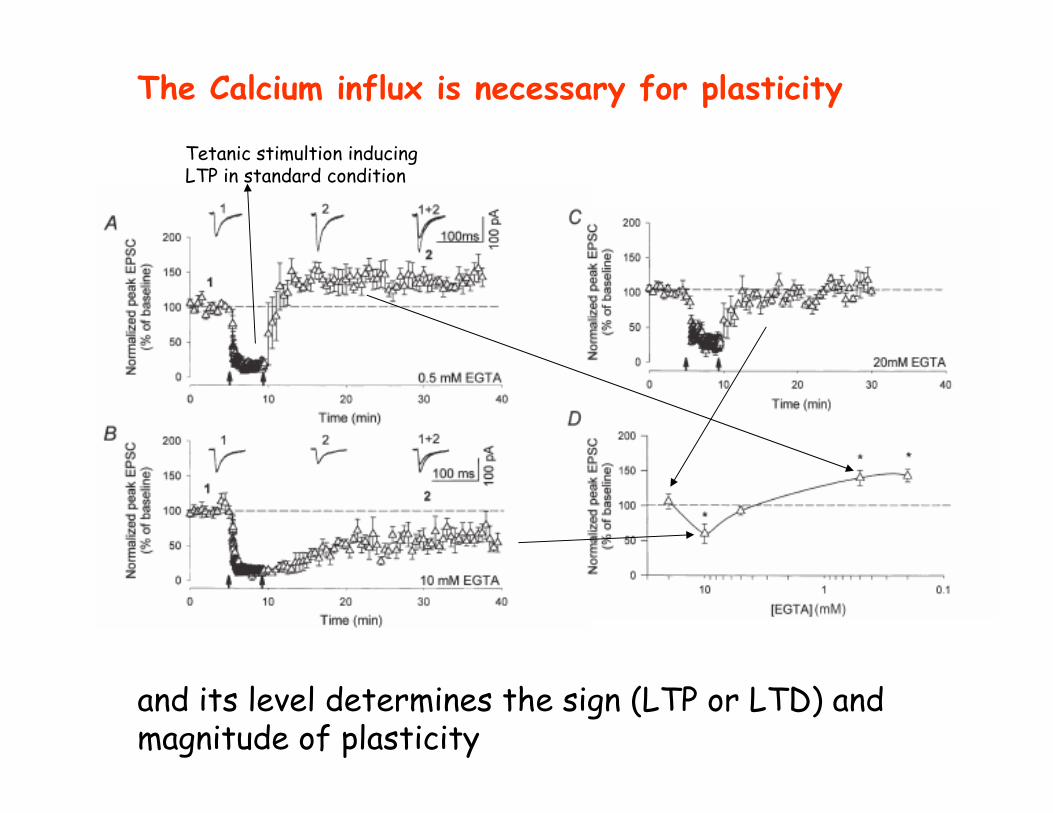
The Calcium influx is necessary for plasticity
and its level determines the sign (LTP or LTD) and magnitude of plasticity
Tetanic stimultion inducingLTP in standard condition
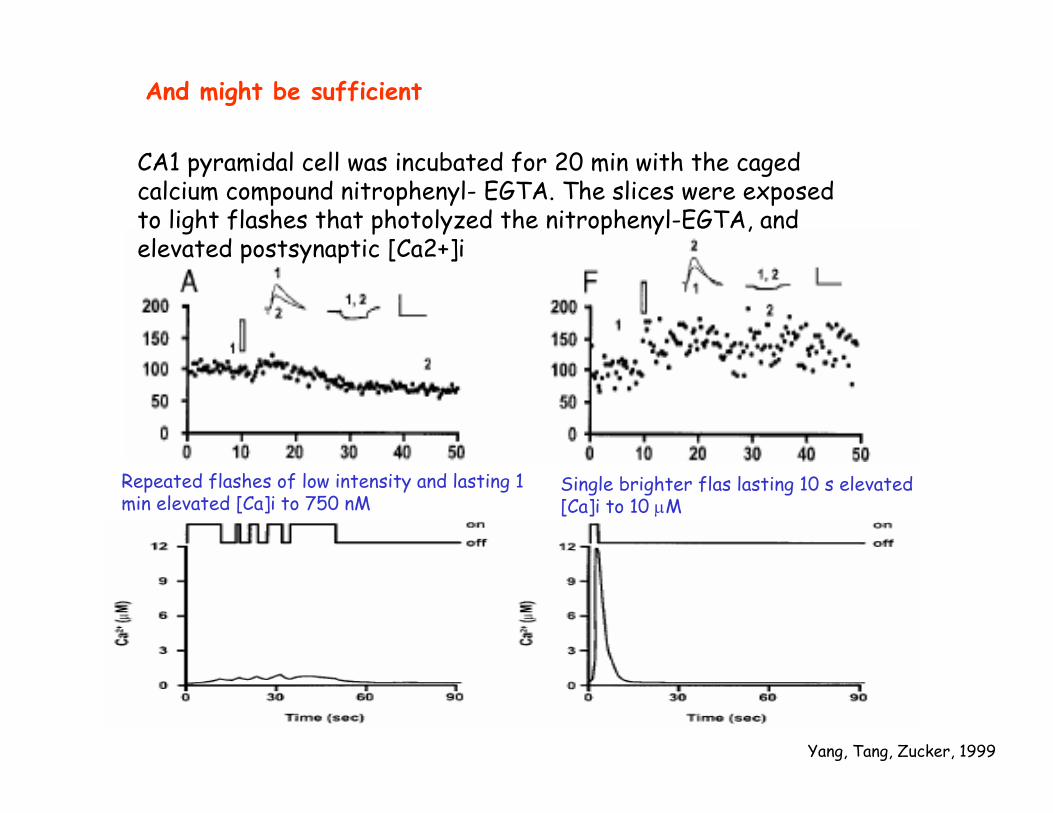
And might be sufficient
Yang, Tang, Zucker, 1999
CA1 pyramidal cell was incubated for 20 min with the caged calcium compound nitrophenyl- EGTA. The slices were exposed to light flashes that photolyzed the nitrophenyl-EGTA, and elevated postsynaptic [Ca2+]i
Repeated flashes of low intensity and lasting 1 min elevated [Ca]i to 750 nM
Single brighter flas lasting 10 s elevated[Ca]i to 10 µM
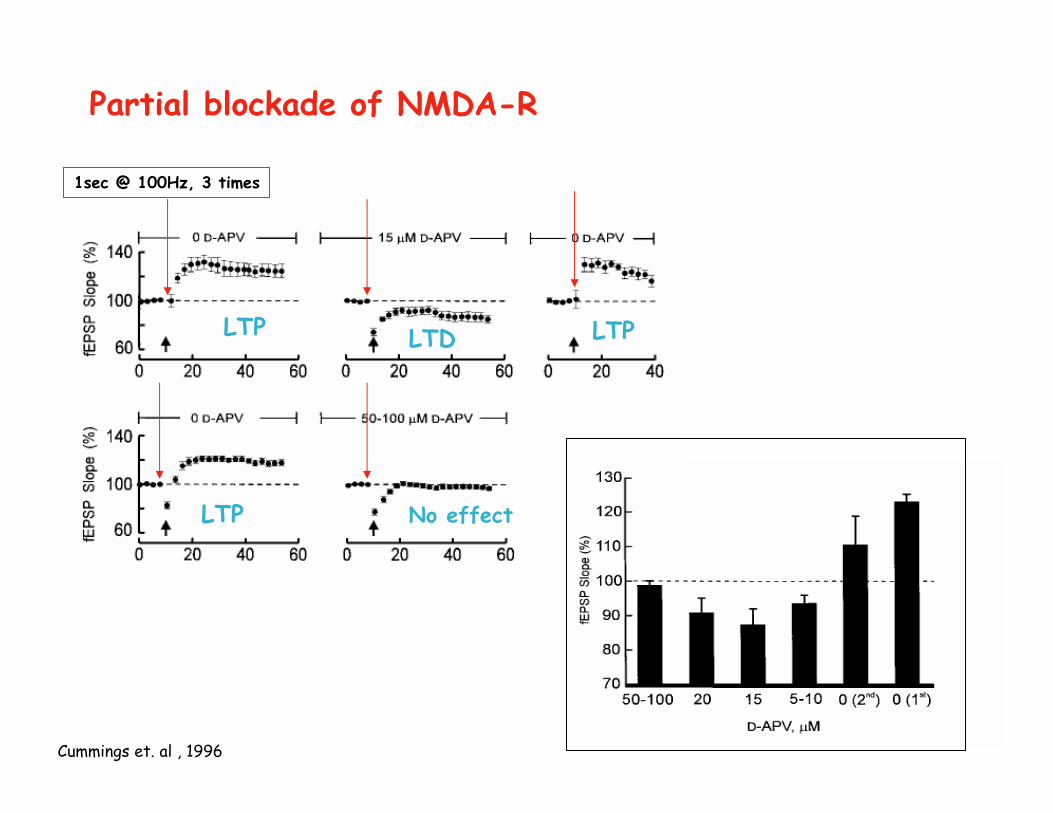
Partial blockade of NMDA-R
Cummings et. al , 1996
1sec @ 100Hz, 3 times
LTP LTD LTP
LTP No effect
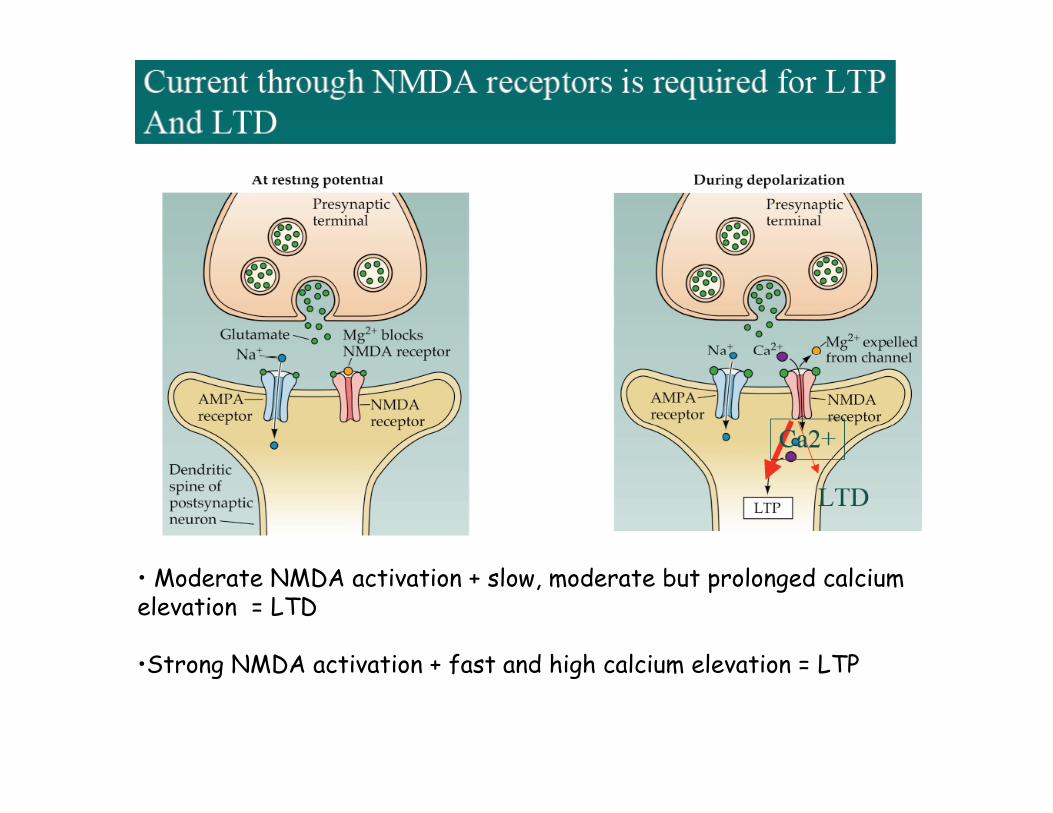
• Moderate NMDA activation + slow, moderate but prolonged calciumelevation = LTD
•Strong NMDA activation + fast and high calcium elevation = LTP
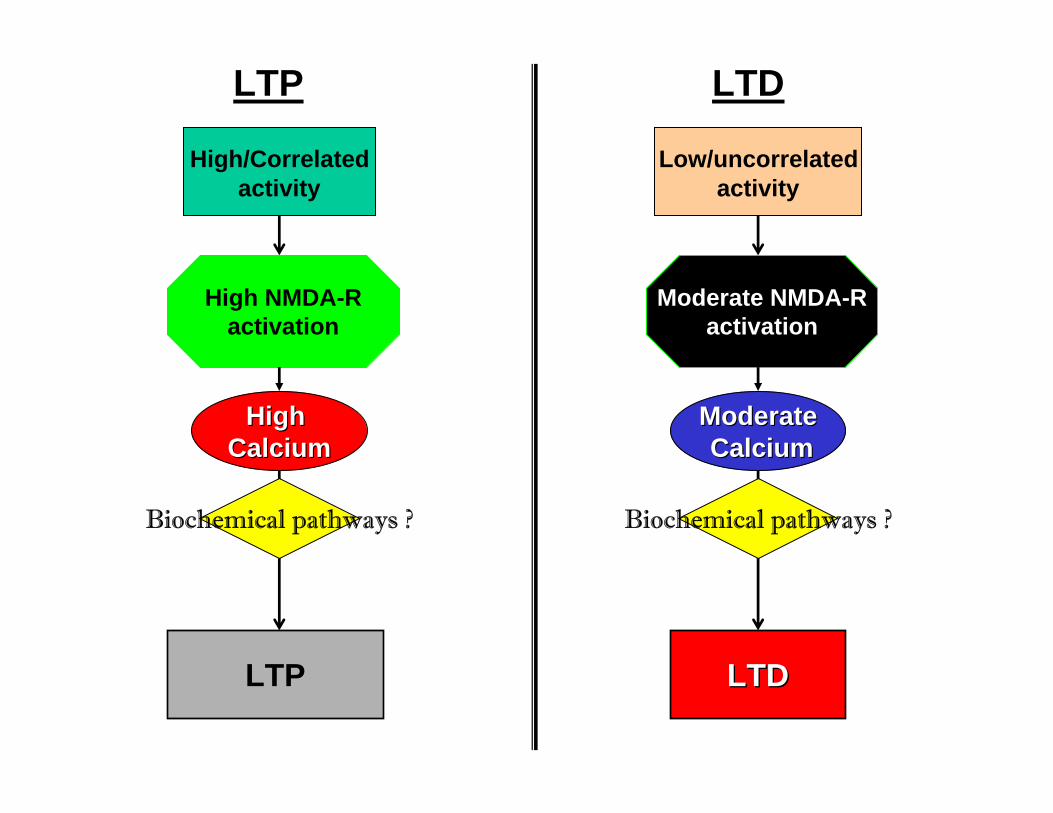
High/Correlated activity
High High CalciumCalcium
LTP
LTP
Low/uncorrelated activity
ModerateModerateCalciumCalcium
LTD
LTDLTD
Biochemical pathways ? Biochemical pathways ?
High NMDA-Ractivation
Moderate NMDA-Ractivation
LTP LTD
Now we know that the biochemical pathways binding postsynaptic Ca2+ changes with LTD /LTP is based on phosphorylation/dephosphorylation processes
LTP and LTD
LTPHigh/Correlated
activity
High High CalciumCalcium
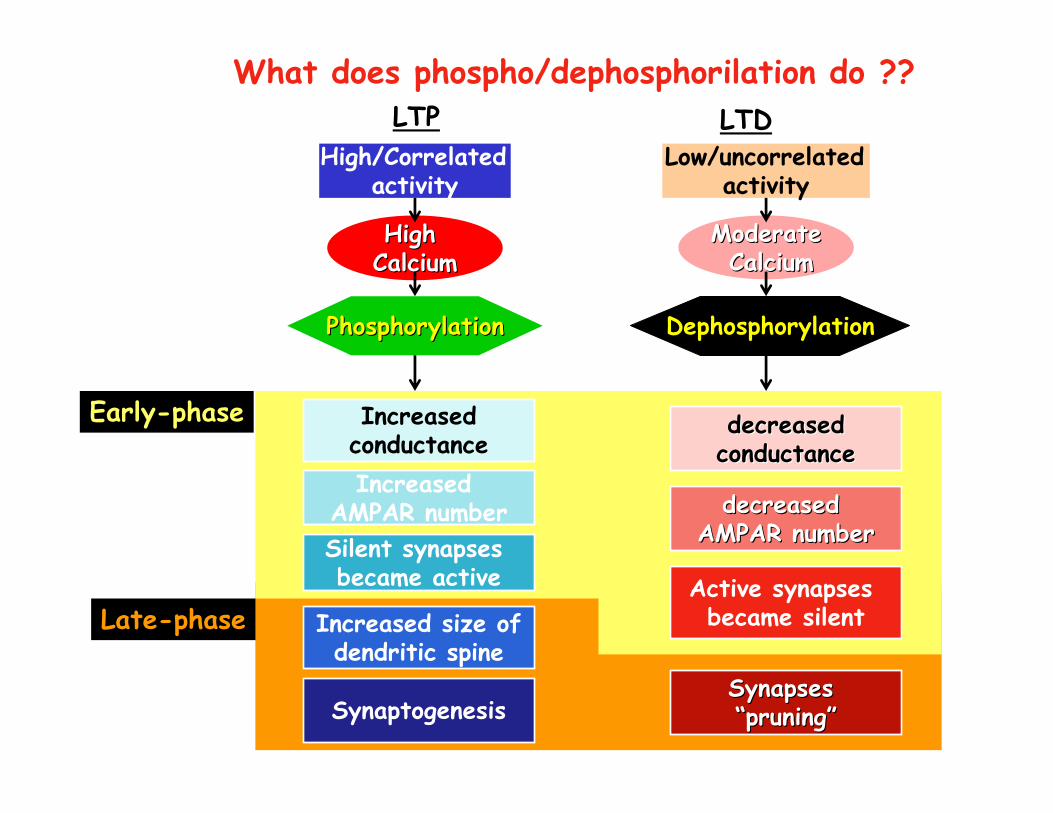
PhosphorylationPhosphorylation
IncreasedconductanceIncreased
AMPAR number
LTDLow/uncorrelated
activity
ModerateModerateCalciumCalcium
Dephosphorylation
decreaseddecreasedconductanceconductance
decreased decreased AMPAR numberAMPAR number
Increased size ofdendritic spine
Silent synapses became active
SynaptogenesisSynapses Synapses ““pruningpruning””
Active synapses became silent
What does phospho/dephosphorilation do ??
Early-phase
Late-phase

The two phases of synaptic plasticity
1. LTP/D (1-6 hrs) – blocked by NMDA antagonists, Ca chelators and protein Kinase/phosphatase inhibitors; not blocked by protein synthesis inhibitors
2. LTP/D (6-24 hrs) – blocked by translation inhibitors – not blocked by transcription inhibitors (require translation of mRNA already available at synaptic sites)
3. LTP/D (days) blocked by trascription inhibitor (require prodction of new mRNA at neclear level)
Early E-LTP/DPhosphorilation/Dephosphorilation
Late L-LTP/DGene expression & structural changes
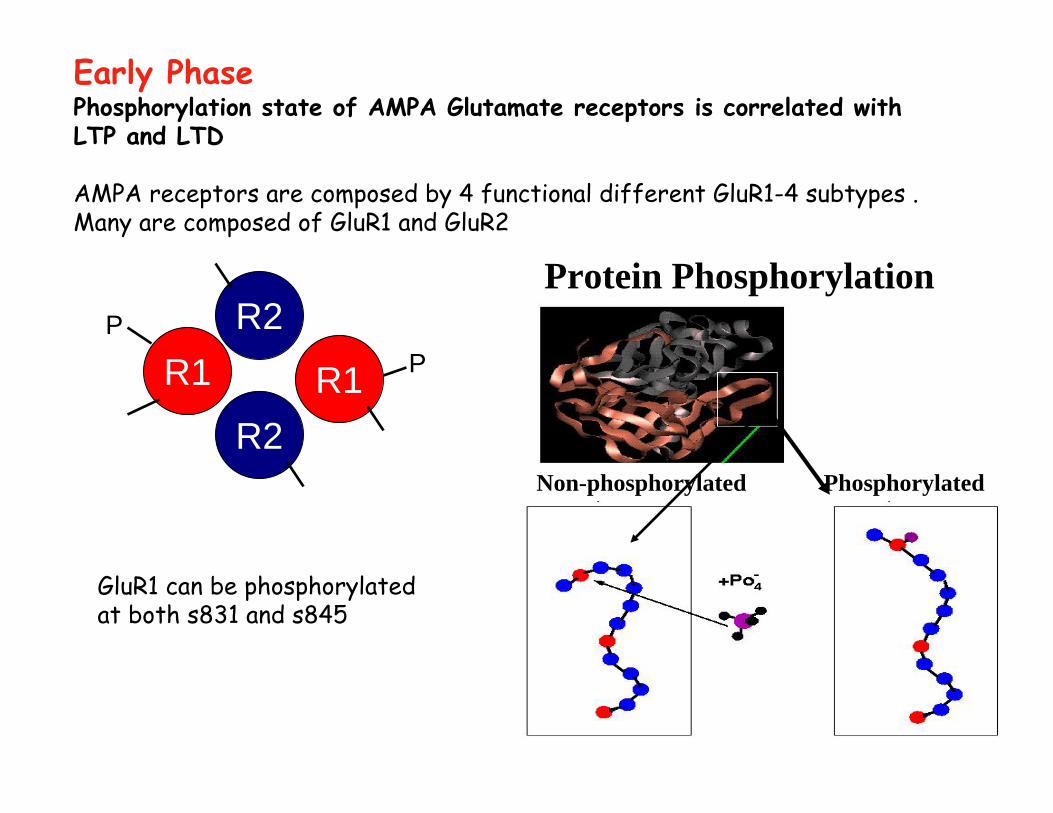
Phosphorylation state of AMPA Glutamate receptors is correlated with LTP and LTD
AMPA receptors are composed by 4 functional different GluR1-4 subtypes . Many are composed of GluR1 and GluR2
R1R2
R2R1 P
P
Early Phase
Protein Phosphorylation
Non-phosphorylated Phosphorylated
GluR1 can be phosphorylatedat both s831 and s845
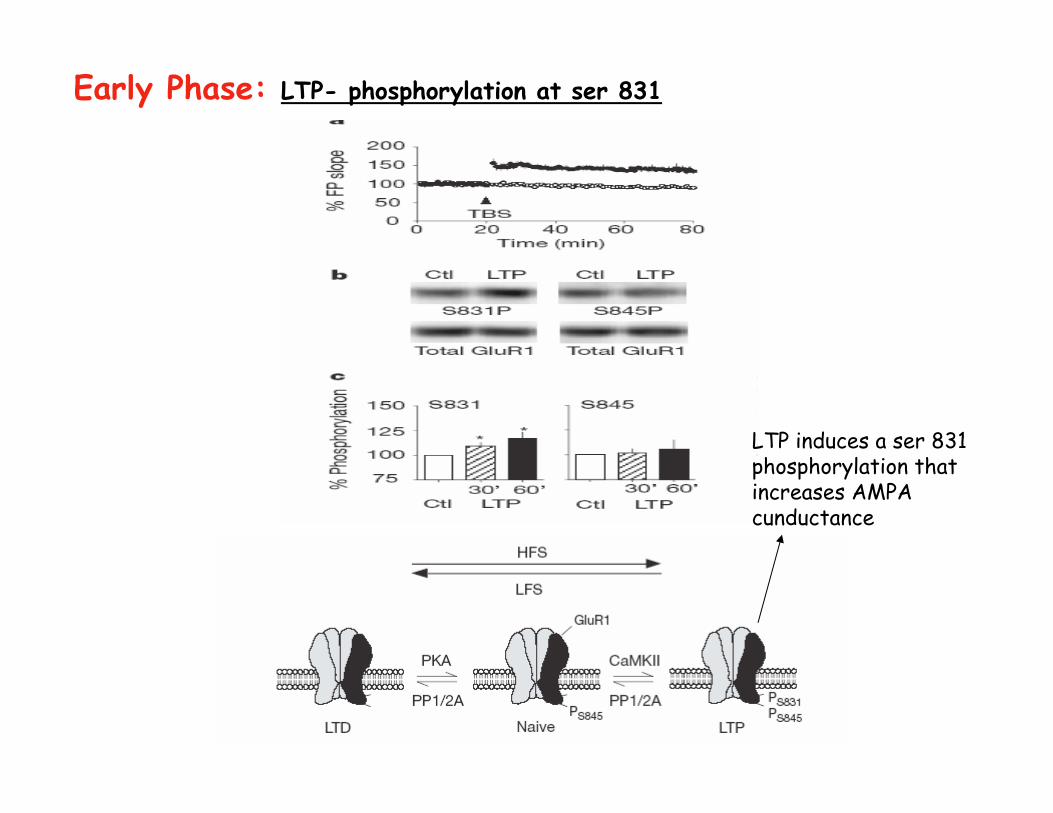
Early Phase: LTP- phosphorylation at ser 831
LTP induces a ser 831 phosphorylation that increases AMPA cunductance
Lee et al. 2000
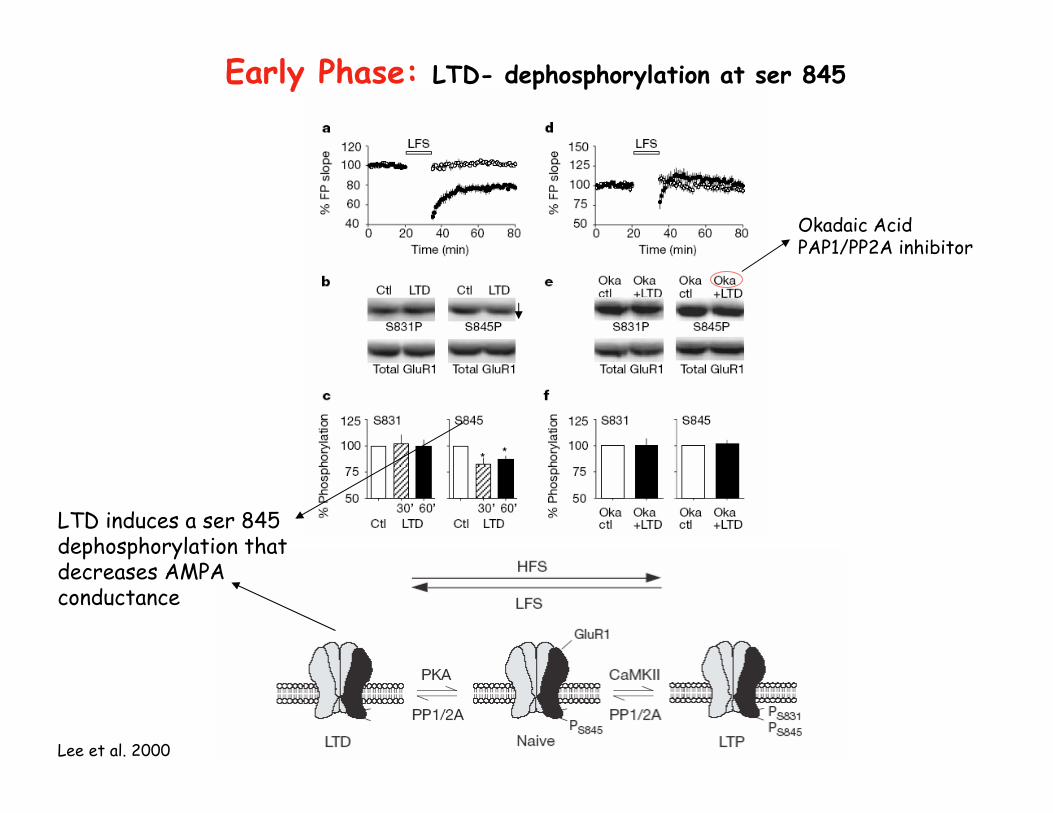
Early Phase: LTD- dephosphorylation at ser 845
LTD induces a ser 845 dephosphorylation that decreases AMPA conductance
Okadaic AcidPAP1/PP2A inhibitor
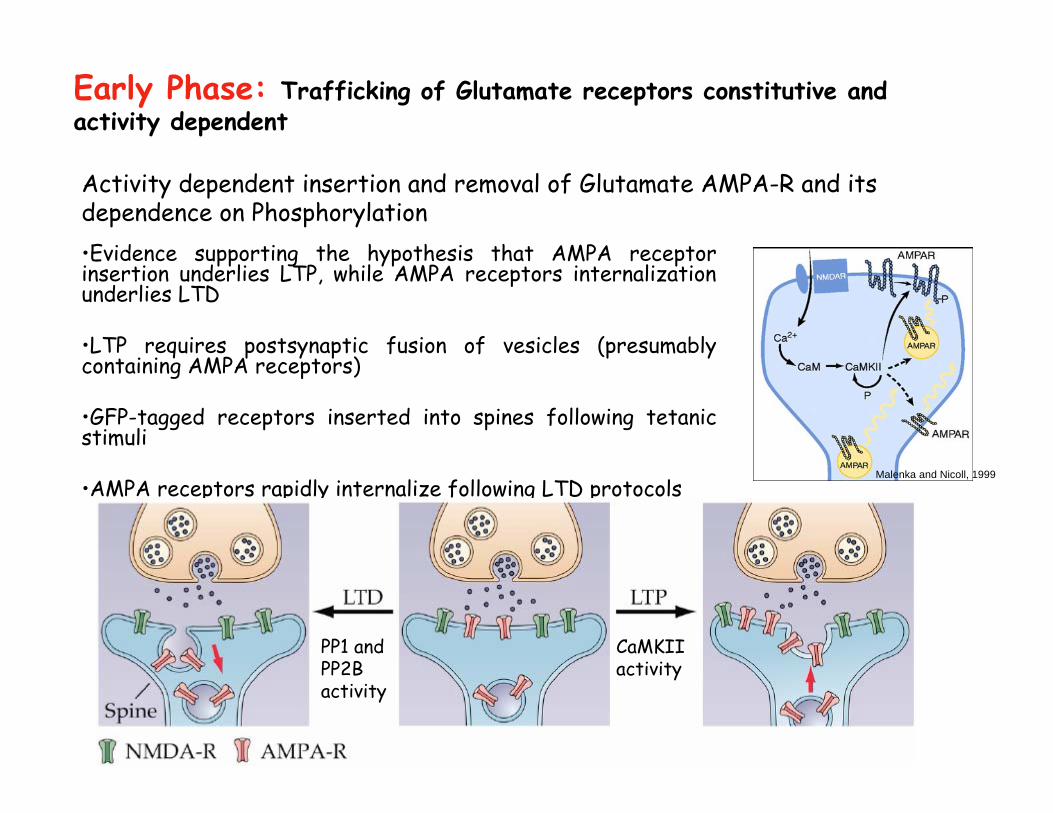
Activity dependent insertion and removal of Glutamate AMPA-R and its dependence on Phosphorylation•Evidence supporting the hypothesis that AMPA receptor insertion underlies LTP, while AMPA receptors internalization underlies LTD
•LTP requires postsynaptic fusion of vesicles (presumably containing AMPA receptors)
•GFP-tagged receptors inserted into spines following tetanicstimuli
•AMPA receptors rapidly internalize following LTD protocols
Early Phase: Trafficking of Glutamate receptors constitutive and activity dependent
Malenka and Nicoll, 1999
PP1 and PP2B activity
CaMKIIactivity
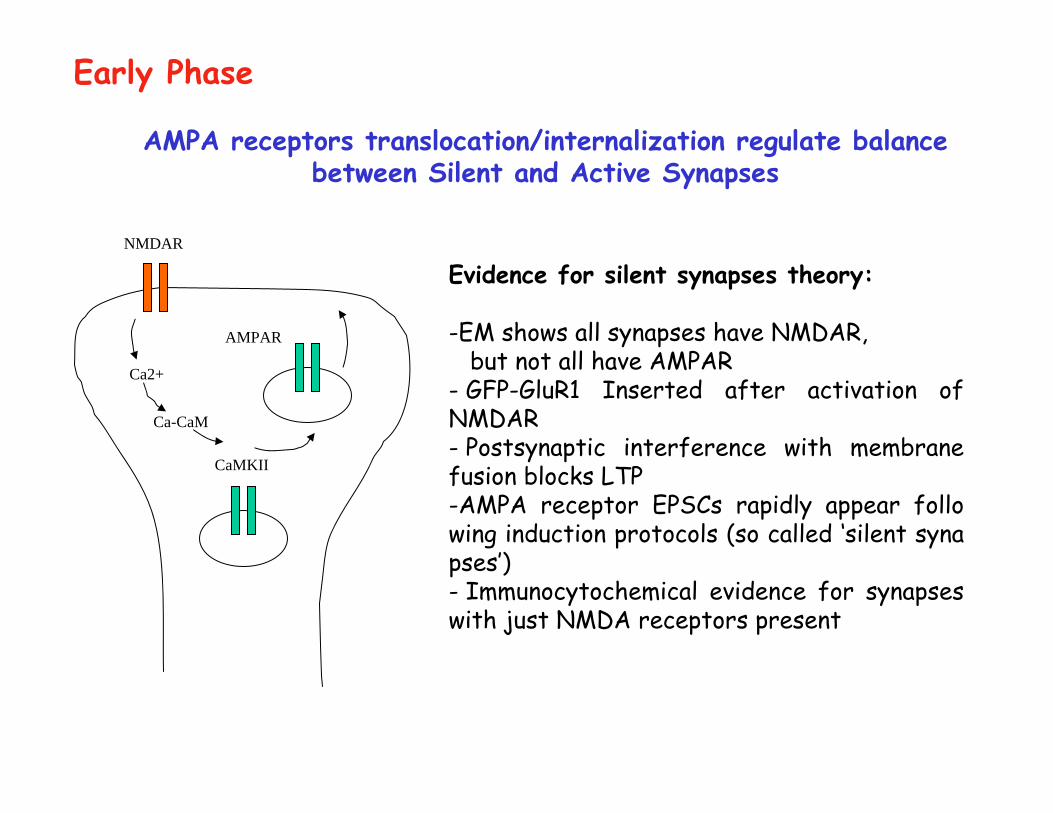
Ca2+
Ca-CaM
CaMKII
AMPA receptors translocation/internalization regulate balance between Silent and Active Synapses
Evidence for silent synapses theory:
-EM shows all synapses have NMDAR, but not all have AMPAR
- GFP-GluR1 Inserted after activation of NMDAR- Postsynaptic interference with membrane fusion blocks LTP-AMPA receptor EPSCs rapidly appear follo wing induction protocols (so called ‘silent synapses’)- Immunocytochemical evidence for synapses with just NMDA receptors present
NMDAR
AMPAR
Early Phase
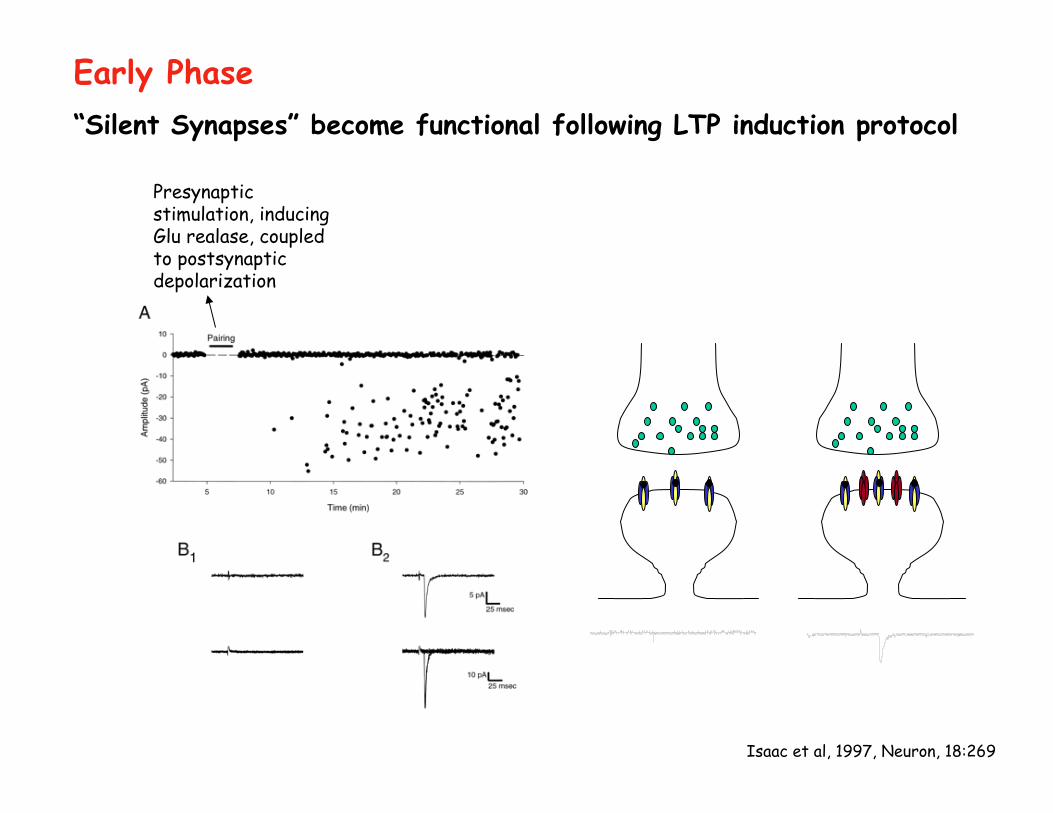
Isaac et al, 1997, Neuron, 18:269
“Silent Synapses” become functional following LTP induction protocol
Early Phase
Presynaptic stimulation, inducing Glu realase, coupled to postsynaptic depolarization

The Late Phase of LTP - It is probably responsible for long-term memory
- Synaptic plasticity due to the Early Phase is consolidated by the Late Phase
- Dependent on gene expression & structural changes of the synspes
- It is protein synthesis dependent, blocked by translation inhibitors (anysomicin)
- The Late Phase involves the activation of immediate early genes (positive and negative transcription factors: CREB, Zif268, REST, etc) which in turn switch on/off the expression on many genes involved in synaptic function and structure.
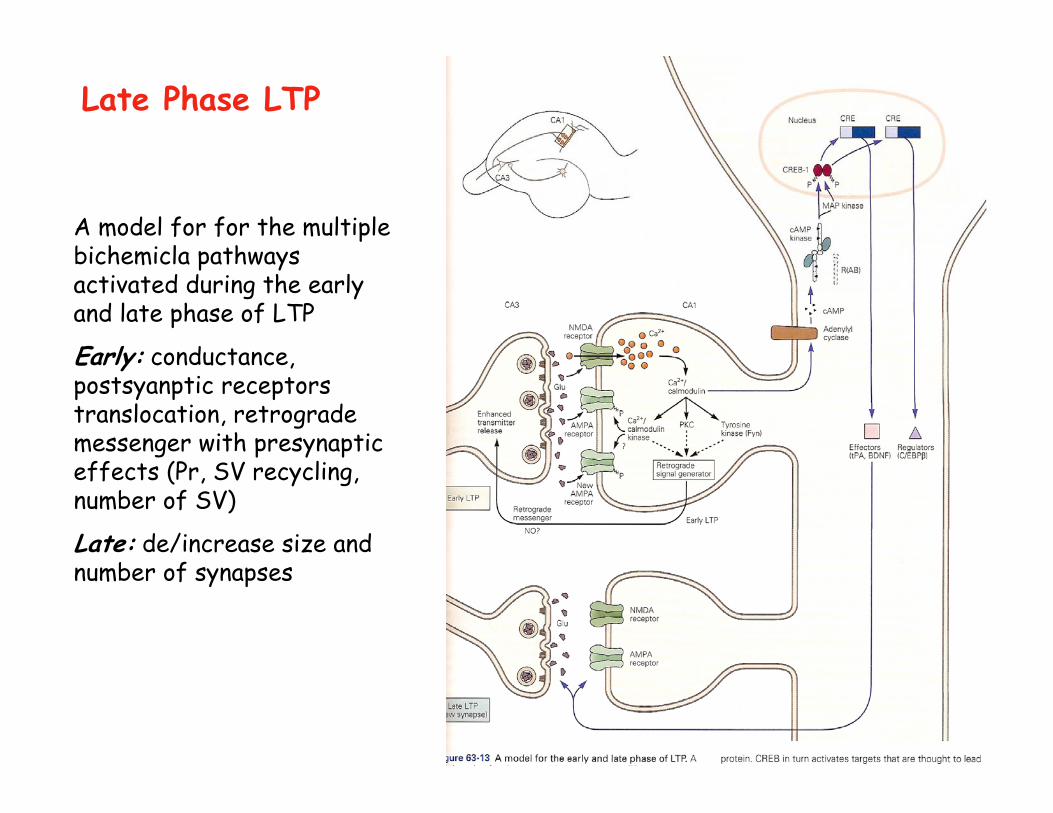
Late Phase LTP
A model for for the multiple bichemicla pathways activated during the earlyand late phase of LTP
Early: conductance, postsyanptic receptors translocation, retrograde messenger with presynaptic effects (Pr, SV recycling, number of SV)
Late: de/increase size and number of synapses
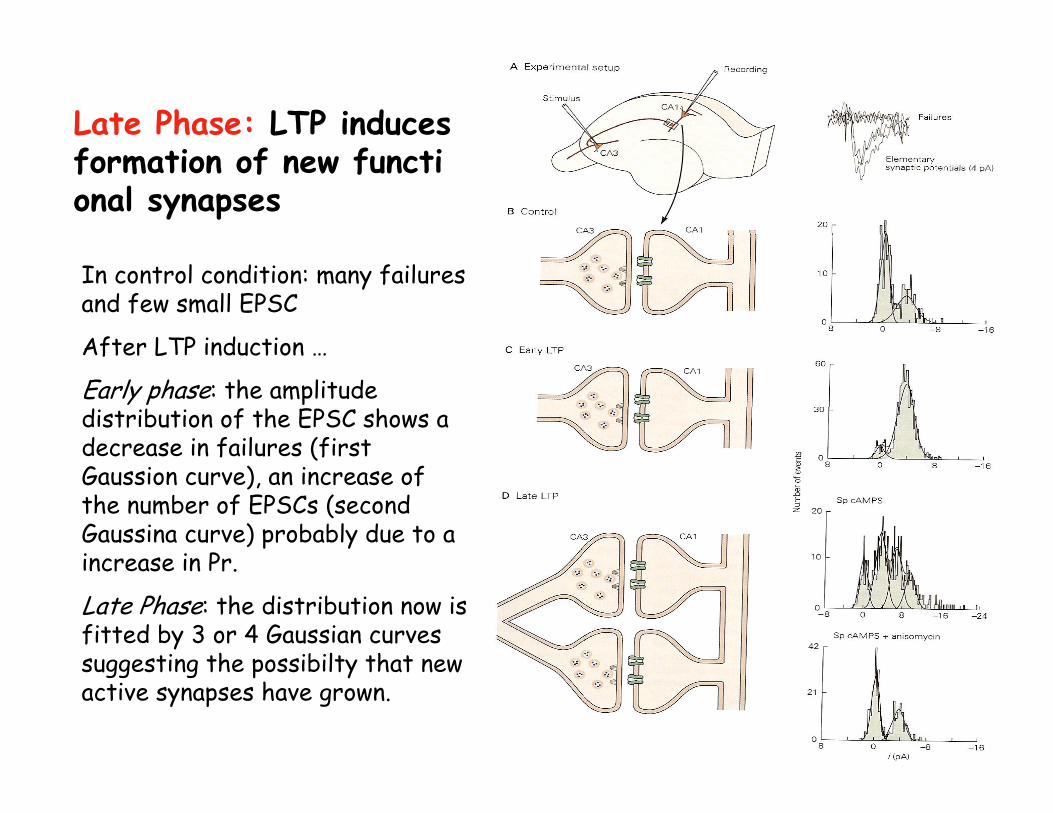
Late Phase: LTP induces formation of new functional synapses
In control condition: many failures and few small EPSC
After LTP induction …
Early phase: the amplitude distribution of the EPSC shows adecrease in failures (first Gaussion curve), an increase of the number of EPSCs (second Gaussina curve) probably due to a increase in Pr.
Late Phase: the distribution now is fitted by 3 or 4 Gaussian curves suggesting the possibilty that new active synapses have grown.
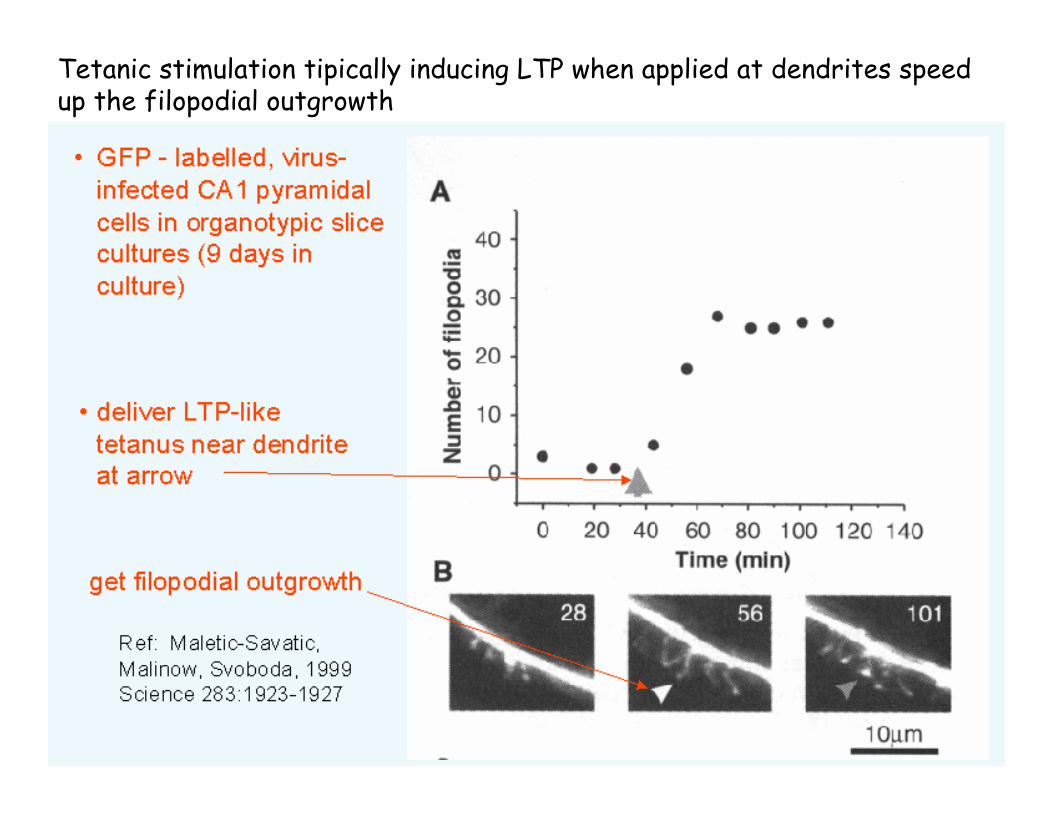
Late Phase LTPTetanic stimulation tipically inducing LTP when applied at dendrites speed up the filopodial outgrowth
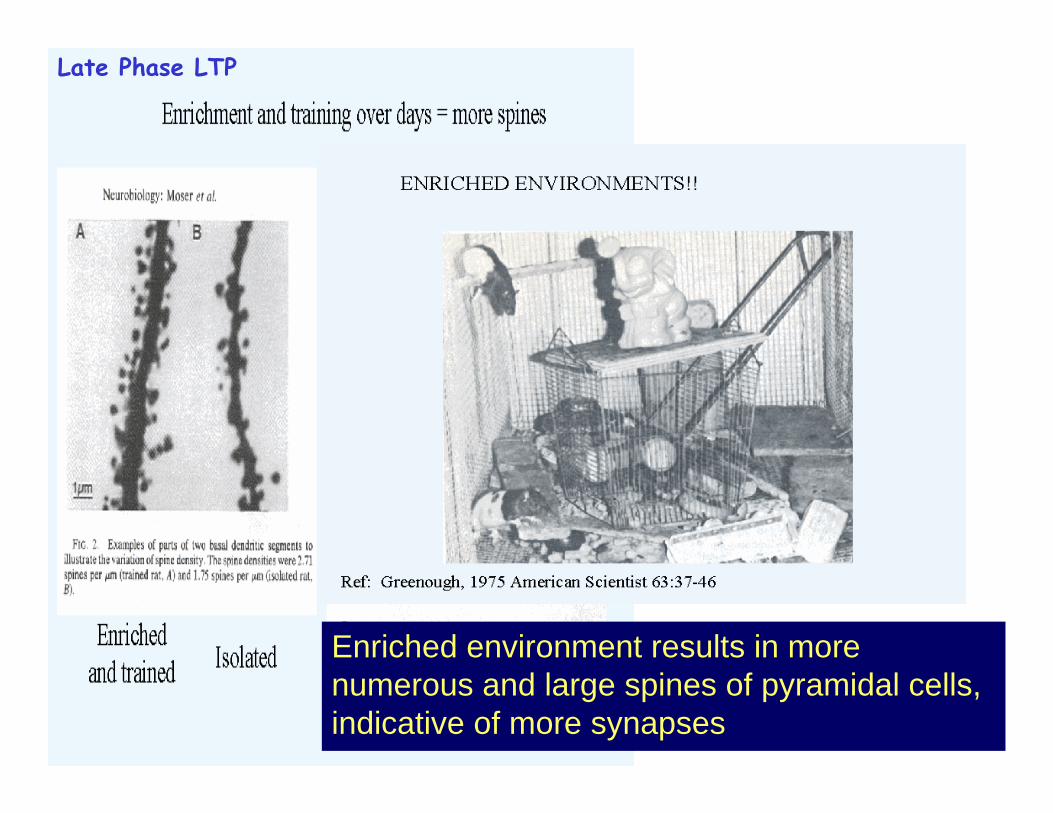
Enriched environment results in more numerous and large spines of pyramidal cells, indicative of more synapses
Late Phase LTP
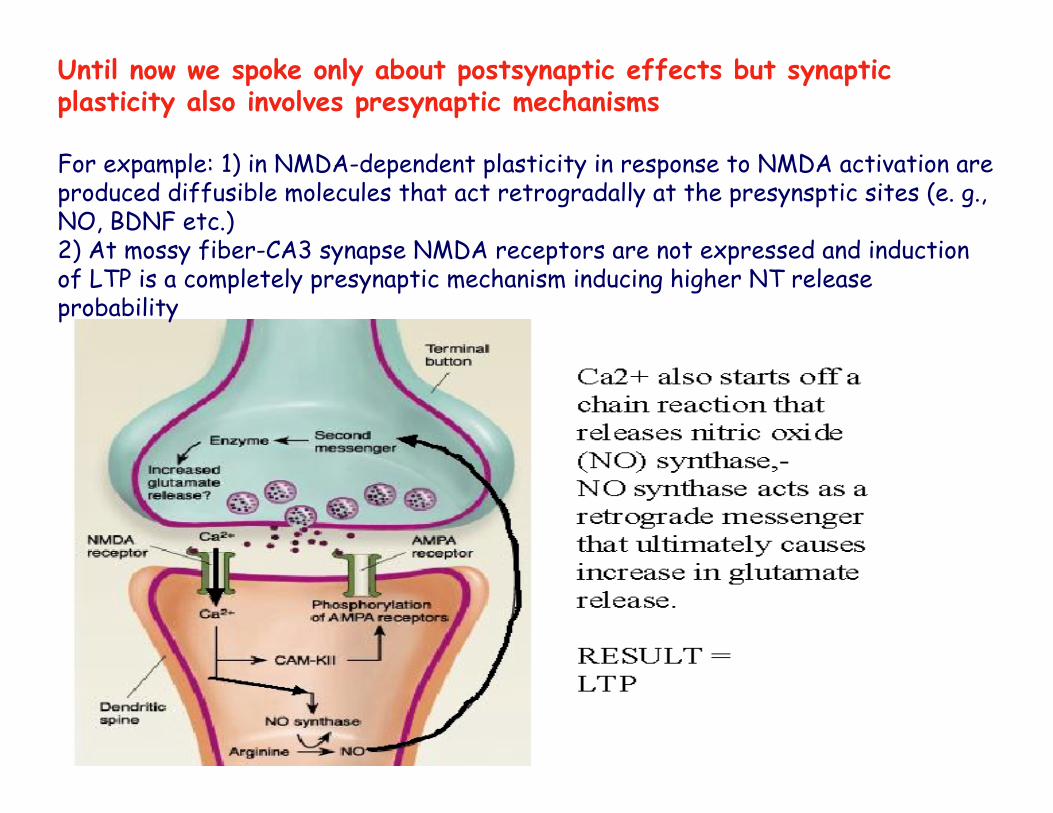
Until now we spoke only about postsynaptic effects but synaptic plasticity also involves presynaptic mechanisms
For expample: 1) in NMDA-dependent plasticity in response to NMDA activation are produced diffusible molecules that act retrogradally at the presynsptic sites (e. g., NO, BDNF etc.)2) At mossy fiber-CA3 synapse NMDA receptors are not expressed and induction of LTP is a completely presynaptic mechanism inducing higher NT release probability

To have an idea of the complexity of mechanisms involved in synaptic plasticity take a look at this table where are summarized molecules implicated in LTP and LTD as “Mediators” (required) or Modulators (non-essential, but involved)
Table updated at 2001

One of the greatest challenges in neuroscience is to determine how synaptic plasticity in neurons is linked with learning and memory in
alive animals
• Lesion studies of those brain areas where plasticity is particularly evident (for ex. “hippocampi lesions induced impairment of L&M”)• Pharmacological Studies (for ex. “NMDA antagonists induced impairment of L&M”)• Genetic manipulations (for ex. “knockout and other genetic alterations of various synaptic plasticity mediators impaired L&M”)
Three main strategies to find the relation between synaptic plasticity in neurons and L&M in alive animals
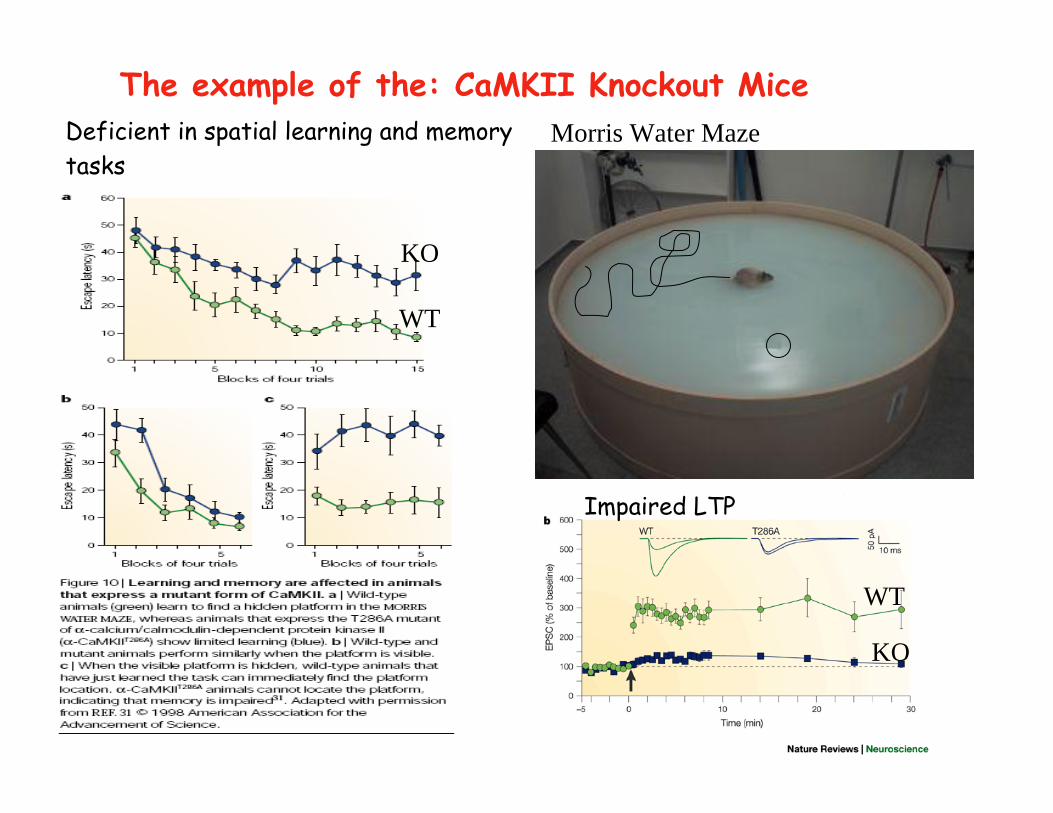
Deficient in spatial learning and memorytasks
The example of the: CaMKII Knockout Mice
WT
KO
WT
KO
Impaired LTP
Morris Water Maze

Concluding Remarks on SYNAPTIC PLASTICTYClear and definitive evidence showing that synaptic plasticity is themain mechanism underlying L&M are still lacking. There are multipletypes of memory and the exact role of synaptic plasticity in trace storage would depend very much on the neural network in which it was embedded.
Probably synaptic plasticity is only the starting point of more complex mechanisms that can be evaluated only evaluating how information is processed into the entire neural network to storememory traces.
To deepen these aspects we require to develop microelectrode arrays technologies for the simultaneous recording of hundreds/thousands of single cells and bioelectronic devices to detect in awake animals synaptic strength changes associated with L&M.
These new approaches combined with more classical pharmacologicaland genetic intervention is the way to advance our understanding on this fascinating topic.

IIT
Of course, the sophisticated nature of the field and such diversetechnological requirements dictate a multidisciplinary collaborative approach ofmany laboratories.