Embed Size (px)
Citation preview

ii
Centro de Investigación en Alimentación y
Desarrollo, A. C.
PORCENTAJE DE OCUPACIÓN Y MODELACIÓN DEL
NICHO ECOLÓGICO DE LA ZORRA GRIS (UROCYON
CINEREOARGENTEUS) EN EL CAÑÓN DE LAS BARAJITAS,
SONORA.
Por:
Lucila del Carmen Armenta Méndez
TESIS APROBADA POR LA COORDINACIÓN DE ASEGURAMIENTO
DE CALIDAD Y APROVECHAMIENTO SUSTENTABLE DE
RECURSOS NATURALES
Como requisito parcial para obtener el grado de
MAESTRO EN CIENCIAS
Guaymas, Sonora Enero, 2015



ii
AGRADECIMIENTOS
A CONACYT.
Al CIAD Unidad Regional Guaymas, gracias al personal administrativo y docente por todas
los servicios que brindan para poder llevar a cabo un trabajo de investigación.
Al CIADE compañeros que están y estuvieron, gracias por: los raites, el café, las coyotas, las
semitas y las pláticas amenas.
A mi director de tesis el Dr. Juan Pablo Gallo, gracias por la confianza, el apoyo para este
proyecto, sobre todo por presentarme el Cañón de Las Barajitas (con mayúscula), muchas
gracias y ha seguir trabajando en pro de la conservación de los recursos naturales.
A Chito, Ana, Victoria, Dr. Gallo, Leo, Chelino, Néstooooooooor, Susana, Bárbara, Flaco y
Yas, gracias por ir, ayudar, opinar y soportar, ¡gracias compañeros! Un agradecimiento
especial a Chelino.
A Don José Luis y a los pescadores que nos llevaron y trajeron con bien (bien mojados).
Al Herbario USON, a Jesús por sus asesorías y el apoyo para la identificación de plantas, un
placer trabajar contigo, lo mismo que Ben Wilder.
A los profesores investigadores de la UES, Leopoldo Villarruel y Alberto Macías.
Al Dr. Alfonso Gardea por su revisión y oportunas correcciones.
A Mateo por permitirme el acceso a Las Barajitas; Jaime, Güero y Capitán por su hospitalidad.
A mis hermanos y sus ewoks.
A mis padres, gracias por cuidar a mi Ambar y por la educación que me han regalado, los
amo.

iii
DEDICATORIA
A mi hija Ambar, tus dibujos son la mejor bibliografía.
A mis padres, por su amor y apoyo sin fin…La trinidad del Desierto de Sonora.
A los estudiantes, luchadores intelectuales por una mejor sociedad.

iv
CONTENIDO
Página
LISTA DE FIGURAS vii
LISTA DE TABLAS viii
RESUMEN ix
ABSTRACT x
I. INTRODUCCION 1
II MARCO TEÓRICO 4
II.I Nicho ecológico 4
II.II Conceptos claves del nicho ecológico 5
II.III Modelación de nicho 5
II.IV Ocupación de las especies en su hábitat 8
II.V Zorra gris 9
II.VI Foto trampeo 11
II.VII Sierra El Aguaje y Cañón de Las Barajitas 12
II.VIII Clima 13
II.IX Bioclima 13
III. JUSTIFICACION 15
IV. HIPÓTESIS 16
V.OBJETIVO GENERAL 17
VI.OBJETIVOSESPECÍFICOS 17
VII. MATERIALES Y MÉTODOS 18
VII.I Área de estudio 18
VII.II Estimación de parámetros poblacionales 24
VII.II.I Análisis Fototrampeo 24
VII.III Trampeo de ratones 25
VII.III.I Análisis trampeo de ratones 25
VII.IV Descripción de la vegetación 26
VII.IV.I Análisis de los cuadrantes 26
VII.IV.II Análisis de los releves 27
VII.IV.III Acumulación de especies 28
VII.V Modelación del nicho ecológico 31
VII.VI Probabilidad de captura y ocupación 34
VIII Resultados y Discusión 35
VIII.I Fototrampeo 35
VIII.II Patrones de actividad 45
VIII.III Especies presa 47
VIII.IV Vegetación 49

v
CONTENIDO (Continuación)
VIII.V Porcentaje de ocupación y probabilidad de detección 51
VIII.VI Modelación del nicho ecológico 53
IX. CONCLUSIONES 63
X. RECOMENDACIONES 65
XI. BIBLIOGRAFÍA 67
XII. APÉNDICES 75
Apéndice 1 Listado florístico muestreado 76
Apéndice 2 Cambios estacionales en la vegetación al Noroeste del
Cañón 78

vi
LISTA DE FIGURAS
Figura Página
1 Zorra gris en zona costera del Cañón de Las Barajitas 10
2 Área de estudio 18
3 Uso de suelo y vegetación 19
4 Geología 20
5 Topografía e Hidrología 21
6 Fisiografía 22
7 Edafología 23
8 Pantalla interfaz gráfica de MaxEnt 33
9 Estaciones de fototrampeo verano 37
10 Estaciones de fototrampeo otoño 38
11 Estaciones fototrampeo invierno 39
12 Estaciones fototrampeo primavera 40
13 Frecuencia de aparición 41
14 Zorra gris y cría 44
15 Dos individuos de zorra gris 45
16 Patrones de actividad 47
17 Función de acumulación de especies 50
18 Mapa de la distribución general 55
19 Mapa de la distribución en Sonora 56
20 Mapa de la distribución en el Desierto Sonorense 57
21 AUC de la distribución general 58
22 AUC de la distribución en Sonora 59
23 AUC de la distribución en el Desierto Sonorense 60
24 Mapa de la validación en el Desierto Sonorense 61
25 AUC de la validación en el Desierto Sonorense 62

vii
LISTADO DE TABLAS
Tabla Página
1 Ejemplos de estudios con MaxEnt 7
2 Grados de presencia 27
3 Grados de fidelidad 27
4 Matriz de acumulación de especies vegetales 28
5 Variables bioclimáticas 32
6 Especies foto trampeadas 36
7 Relevé de vegetación zona costera 42
8 Relevé de vegetación aguaje 42
9 Relevé de vegetación aledaña al arroyo 43
10 Resultados Trampeo de ratones 49

viii
RESUMEN
Los modelos de ocupación y de nicho ecológico, son útiles para establecer las características
ecológicas de un lugar; con estas características se pueden analizar las condiciones que
favorecen la conservación de la biodiversidad de un área determinada. Se utilizó como modelo
a la zorra gris (Urocyon cinereoargenteus) para evaluar las características ecológicas del
Cañón de Las Barajitas, ubicado a 40 km al noroeste del puerto de Guaymas, en Sonora,
México.
El hábitat potencial de la zorra gris se modeló con el programa MaxEnt. La probabilidad de su
detección (P) y el porcentaje de ocupación (ψ) fueron estimados con el programa MARK. Los
patrones de actividad de la zorra gris y de las especies simpátricas fueron comparados usando
la frecuencia de aparición y la hora de detección con foto-trampeo. La evaluación de la
vegetación se determinó por el método de cuadrantes y el de Braun-Blanquet, mediante la
función de acumulación de especies. Se capturaron y liberaron roedores para registrar a las
especies presa que conforman la base de la cadena trófica.
El esfuerzo anual de muestreo fue de 124 días/noche trampa, obteniendo 575 fotografías de
zorra gris. El análisis de los patrones de actividad coloca a la zorra gris, al coyote, al gato
montés y al cacomixtle, como especies crepusculares - nocturnas. La evaluación de la
vegetación muestreada presenta una dominancia de la asociación Bursera-Jatropha-Ruellia.
Ruellia californica fue la planta más abundante (43.48%). La función de la acumulación de
especies vegetales mostró una relación lineal positiva (r²= 0.97).
En las capturas se identificaron a los roedores de las familias Heteromydae en sustrato
pedregoso y Muridae en sustrato arenoso y pedregoso.
De acuerdo a los criterios de información (AICc), se encontraron correlaciones positivas con
un intervalo de confianza del 95% entre especies simpátricas y la estacionalidad,
principalmente con el cacomixtle (1.66) y el coyote (1.61); así, bajo los modelos de MARK
los resultados fueron P= 2.85% y ψ= 14.37%, y P= 11.93% y ψ= 2.15%; estos modelos se
compararon con una prueba de Mann-Whitney y se encontró que las diferencias no son
significativas (P= 0.667).
Los resultados muestran que el Cañón de Las Barajitas es un ambiente adecuado para la zorra
gris, de acuerdo a la dinámica de comunidades de flora y relaciones de fauna observadas en

ix
este estudio, y que el modelo de nicho de MaxEnt coincide con las preferencias del hábitat de
la especie con un valor de AUC= 0.873.
Palabras clave: Cañón de Las Barajitas, costa central del Desierto Sonorense, foto- trampeo,
modelación ecológica, zorra gris.

x
ABSTRACT
Occupancy and ecological niche models are used to stablish the ecological characteristics of a
given area; based on such characteristics, analyses can be conducted on the conditions that
might favor biodiversity conservation of that area. Gray fox (Urocyon cinereoargenteus) was
used as a model to evaluate ecological characteristics of the Cañón de Las Barajitas, located
40 km NW of Guaymas Port in Sonora, Mexico.
Gray fox potential habitat was modeled by using the MaxEnt program. Detection probability
(P) and occupancy (ψ) were estimated with the MARK program. Gray foxes activity patterns
were compared with other sympatric species by using frequency and hour of detection on
camera traps. Vegetation evaluation was determined by the quadrant and the Braun-Blanquet
methods, by using the species accumulative function. Rodent capture and release was used to
record prey species conforming the trophic chain basis.
Sampling annual effort was 124 day/night camera traps, obtaining 575 gray fox photographs.
Activity patterns analyses place gray fox, coyote, bobcat and ringtail as crepuscular –
nocturnal species. Evaluation of sampled vegetation is dominated by the association Bursera-
Jatropha-Ruellia. Ruellia californica being the most abundant plant (43.48%). The
accumulated vegetation species function showed a positive lineal relationship (r²= 0.97).
Rodents captured represented the families Heteromydae over rocky substrate, and Muridae
over sandy and rocky substrates.
In accordance with information criteria (AICc), positive correlations were found with a 95%
confidence interval between sympatric species and seasonality; mainly with ringtail (1.66) and
coyote (1.61). Results from the MARK model were P= 2.85% and ψ= 14.37% for sympatric
species, and P= 11.93% and ψ= 2.15% for non-sympatric species; these models were
compared with the Mann-Whitney test and no significant differences (P= 0.667) were found.
Results show that Cañón de Las Barajitas is an optimal environment for gray fox, due to the
flora and fauna communities dynamics observed in this study. Also the ecological niche

xi
modeling with MaxEnt coincides with the species habitat preference reaching an AUC value
of 0.873.
Key words: Cañón de Las Barajitas, Sonoran Desert Central Coast, camera traps, ecological
modeling, gray fox.

1
I. INTRODUCCIÓN
Los ecosistemas de Sonora están fuertemente amenazados por perturbaciones
antropogénicas (agricultura, ganadería, extracción forestal, minería y desarrollo urbano)
el objetivo de los estudios ambientales es, por lo tanto, la conservación de áreas
naturales que alberguen diversidad de hábitats, por ser sistemas que ofrecen servicios
ambientales y son importantes refugios de biodiversidad, es decir son fuente de acervo
de recursos genéticos, provisión para polinizadores, prevención de la erosión y en
general, son ambientes que favorecen la conservación de las especies (Búrquez y
Martínez- Yrízar, 2007). Una de las consideraciones con respecto a las áreas protegidas
está en función de los acontecimientos externos; a medida que las sitios alrededor de ella
están siendo impactados, las reservas van quedando como islas, al reducirse el límite real
que dispone para la biota, con mayor afectación sobre las especies con desplazamientos
amplios, como los mamíferos depredadores, se supone que a corto plazo esto afecta a
todo el conjunto florístico-faunístico (Halffter, 1995).
Villaseñor (2004) menciona que mientras más diversidad total encontremos en una
región, debemos suponer que allí existe una mayor complejidad ecológica, ya que una
mayor diversidad solo podrá coexistir y mantenerse si allí se encuentran mecanismos
más complejos, que han evolucionado junto con las especies para regular las
interacciones entre sus miembros.
Bajo esta premisa, podemos utilizar contextos teóricos orientados para resolver
problemas de conservación en sistemas ecológicos específicos, aunque su aplicación no
sea universal, sino más bien regional o local; la teoría tanto a nivel regional como local
nos ayudará a escoger los tratamientos de conservación más apropiados, a manera de
hipótesis, para luego aplicarlos bajo un esquema de manejo experimental adaptativo a
cada lugar de estudio, es decir buscar in situ elementos para explicar de alguna forma
cómo ocurren los procesos naturales dentro de los ecosistemas no alterados o, con un
mínimo grado de perturbación, como es el caso de la zona de estudio de este trabajo, en
el Cañón de Las Barajitas, y de cómo la preservación de la biodiversidad de
determinados lugares es fundamental para mejorar la calidad de vida de la comunidad
humana a regional.

2
Algunos de esos contextos teóricos apuntan hacia el concepto de nicho ecológico y la
ocupación del hábitat por las especies, Hutchinson en 1957 define el nicho ecológico
como el rango total de las variables (p.e. temperatura, alimento, pH, etc.) a las que una
especie está adaptada y en las cuales vive y se sustituye a si misma; el nicho entonces es
una respuesta de los organismos al ambiente; por lo que la utilización de los recursos
que ofrece determinado lugar en determinado tiempo define el nicho ecológico de las
especies, por su parte, la estimación de la ocupación de las especies en su hábitat es un
conocimiento relevante, este parámetro se ha utilizado como una variable alternativa a la
abundancia, y para muchos tiene propósitos inferenciales como la selección del hábitat
y permite formular hipótesis acerca de la dinámica poblacional y la distribución de las
especies (Sollman et al. 2012).
El cañón de Las Barajitas forma parte de la serie de cañones que componen La Sierra El
Aguaje, ubicada en la subdivisión del Desierto Sonorense Costa Central del Golfo, esta
subdivisión está presente en ambos lados del Golfo de California, es la de menor área en
el estado y cubre una extensión aproximada de un millón de hectáreas, es conocida como
región Bursera-Jatropha (Shreve, 1951). Esta unidad se distribuye como una estrecha
banda a lo largo de la mayor parte de la costa de Sonora. Se le considera la tierra
tradicional de los comcáac (seris) (Felger y Moser, 1985). También se distribuye en las
casi novecientas islas e islotes del Golfo de California; Felger (en preparación) describe
un listado de vegetación del Cañón de Las Barajitas con un total de 283 especies
distribuidas en 69 familias que incluyen hierbas, arbustos, trepadoras, pastos, árboles,
plantas anuales y perennes. En cuanto a la diversidad faunística, la Comisión de Áreas
Naturales Protegidas ha reportado 799 especies de invertebrados, de los cuales 9 son
endémicos, 67 especies de mamíferos, así como 20 especies de reptiles, 10 de anfibios y
250 especies de aves. La Sierra El Aguaje está dentro del Coto de Caza Cajón del diablo
(RTP-18 CONABIO, 2000).
Para conocer y describir el nicho ecológico de la zorra gris (Urocyon cinereoargenteus)
en el cañón de Las Barajitas, se hizo la toma de datos in situ, para después proyectarlos

3
mediante modelos de ocupación y de nicho ecológico (por ejemplo, MARK y MaxEnt),
lo cual ayudará en la toma de decisiones acerca del manejo de determinados sitios donde
habita esta especie a nivel regional.
La zorra gris es un cánido pequeño, su longitud total es de 800 a 1,125 mm, los adultos
pesan de 3 a 7 kg, los machos son ligeramente más grandes que las hembras; se
distribuye desde el sur de Canadá hasta Colombia y Venezuela y puede encontrarse en
casi cualquier ecosistema; es una especie generalista dentro de los carnívoros (Fritzell y
Haroldson, 1981); De acuerdo con varios autores, podemos suponer que la zorra gris
contribuye a mantener la riqueza de especies vegetales en el Cañón de Las Barajitas ya
que:
1. Promovería la dispersión y germinación de semillas de plantas de importancia
estructural (Villalobos et al., 2014).
2. Controlaría la densidad poblacional de pequeños mamíferos, lo que implica
promoción de diversidad (Arnaud y Acevedo, 1990).
3. Podría ser una especie presa de puma, coyote, águilas y gato montés (Cypher,
2003).
4. Su tamaño corporal le permitiría tener poblaciones extensas ya que su ámbito
hogareño es variable y depende del tipo de ecosistema (0.8 km a 27.6 km, según
Yearsley y Samuel, 1982; Nicholson, 1982).
El modelado de nicho ecológico es un instrumento que nos permite analizar los factores
ecológicos asociados a diferentes poblaciones de una misma especie y que influyen en
diferentes grados y modos; información que analizada por diferentes algoritmos nos
posibilita proyectar a nivel geográfico el área potencial que ocupa la especie (Martínez
Meyer, 2010), la mayoría de los mecanismos que favorecen una relación positiva entre
abundancia-ocupación estará relacionada con los procesos ecológicos (Gastón, 1997);
así, con la modelación del nicho ecológico y la estimación de la ocupación de la zorra
gris en el Cañón de las Barajitas tendremos una línea base de estudio de las condiciones
y requerimientos asociados con la calidad del hábitat en el cual ocurre esta especie.

4
II. MARCO TEÓRICO
II.I Nicho ecológico
La geobotánica en los inicios del siglo XX sistematizó el conocimiento de la distribución
de las especies vegetales y su relación con el clima, al complementar información
obtenida in situ con el nuevo conocimiento cartográfico del globo terrestre. Estos
avances permitieron construir una relación causal entre la distribución geográfica de las
especies vegetales y la distribución espacial del clima; posteriormente, desde la ecología,
surge el concepto de nicho ecológico, ampliando fuera del ámbito de la botánica la
relación entre los seres vivos y los factores abióticos. Este concepto tiene su origen en
los trabajos de los ecólogos Johnson, Grinell y Elton, donde se buscaba establecer el rol
de las especies e identificar la subdivisión del ambiente (hábitat) que era ocupado por
una especie. (Pliscoff y Fuentes-Castillo, 2011). Grinell (1924) describe al nicho como
la unidad de distribución final en la que cada especie está condicionada por sus
limitaciones instintivas y estructurales; para Elton (1927) el nicho es el estatus de un
animal en su comunidad, indicando qué hace y no solamente cómo se ve. La concepción
de nicho más utilizada es la desarrollada por Hutchinson (1957) que lo definió como el
rango total de las variables ambientales (p. e. alimento, temperatura, pH, etc.) a las
cuales una especie está adaptada y en las cuales vive y se substituye a si misma, esta
definición es clave en la conceptualización de los modelos de distribución de especies
(Pliscoff y Fuentes-Castillo, 2011) y permitió darle un enfoque cuantitativo, cuya base
está en dos descriptores: amplitud y sobre posición (Smith, 1982). La amplitud es la
suma total de la variedad de recursos utilizados por una especie, en cambio la
sobreposición, resulta de las demandas simultáneas sobre algunos recursos por parte de
dos o más especies (Giller, 1984).

5
II.II Conceptos clave del nicho ecológico.
Nicho fundamental o potencial: es donde la especie puede vivir, incluye todos los
aspectos o variables del espacio o bien, es el hypervolumen en ausencia de otras especies
(Hutchinson, 1957).
Nicho efectivo o realizado: es un subconjunto del nicho fundamental en el cual las
especies están restringidas debido a sus interacciones interespecíficas, es el espacio
ecológico y geográfico donde la especie vive; para Soberón y Nakamura (2009) “el
nicho realizado es la parte del nicho fundamental que las especies realmente usarían,
después de tomar en cuenta los efectos de competidores y depredadores”.
Se puede decir entonces que el concepto de nicho ecológico explica la manera en la cual
una especie hace uso de los recursos y sus relaciones con las especies (alimentos,
competencia y depredación) que el hábitat le ofrece en determinado tiempo y espacio,
considerándose que la energía que obtiene es la suficiente para tener una adecuación
exitosa, garantizando la transferencia de genes e ir perpetuando su especie bajo las
condiciones dadas.
II.III Modelación de nicho.
El modelado del nicho ecológico es un instrumento que nos permite analizar los factores
ecológicos en los cuales las especies pueden sobrevivir asociados a distintas poblaciones
de determinada especie y que la influyen en distintos grados y modos, información que
analizada por distintos tipos de algoritmos nos posibilita proyectar a nivel geográfico el
área potencial que ocupa la especie (Martínez, 2010).
La pregunta que se busca responder con la modelación es: ¿Cuáles son los factores que
determinan la distribución de los seres vivos? (McGill, 2010).
El propósito del modelado del nicho ecológico o de los modelados de distribución de
especies y del modelado de hábitat es el mismo: identificar los sitios adecuados para la
supervivencia de las poblaciones de una especie por medio de la identificación de sus
requerimientos ambientales (Soberón y Nakamura, 2009; Martínez, 2010).
Bajo esa perspectiva lo que estamos modelando es el nicho efectivo o realizado (nicho
realizado Grinelleano, sensu J. Soberon) y el resultado del análisis nos indica con cierto

6
valor de probabilidad (y su error estadístico asociado), el espacio geográfico que es
propicio para una especie, por lo que las especies podrían no estar en el espacio
predicho, ya sea por efecto de:
a) Interacciones bióticas con otros organismos (competencia, depredación, escasez de
alimento).
b) No se ha podido dispersar a esos lugares (por tiempo o barreras geográficas y
ecológicas).
c) Simplemente ha sido removida o se ha extendido (Martínez, 2010).
Se pueden distinguir cuatro grandes familias de técnicas de modelación de la
distribución geográfica de las especies/ecosistemas: las relacionadas a modelos
estadísticos de regresiones (Modelos Lineales Generalizados (GLM), Modelos Aditivos
Generalizados (GAM), los métodos de clasificación (Random Forest (RF), Boosted
regression trees (BRT)), los métodos de “sobre” (BIOCLIM, ENFA) y aquéllos basados
en algoritmos específicos (GARP, MAXENT) (Pliscoff y Fuentes-Castillo, 2011). Para
este trabajo se utilizará el programa MaxEnt, éste modela la distribución geográfica de
las especies, utiliza como insumos datos bioclimáticos (capas) y datos de presencia
(georreferenciación) de la especie de interés, la información bioclimática se obtiene de
http://www.worldclim.org/bioclim y ésta debe ser asociada a los registros de la especie,
los datos de presencia pueden obtenerse a través de encuestas, avistamientos, foto
trampeo, registro de rastros o registros de avistamientos que se se pueden obtener de la
red (gbif.org, madrean.org, manisnet.org, ornisnet.org, trópicos.org,
conabio.inaturalista.org/taxa), así como de colecciones de museos; MaxEnt utiliza solo
los sitios de presencia y las variables bioclimáticas asociadas a cada uno de esos puntos
de presencia, para modelar las distribuciones se basa en el principio de Máxima entropía,
es decir se extraen las correlaciones que presenten mayor orden en el ambiente y que
podrían influir en la presencia de la especie con una distribución más uniforme.
Para Baldwin (2009), el modelado con MaxEnt aún siendo un método nuevo que
requiere del desarrollo de técnicas enfocadas en la disminución del sesgo estadístico
producido por la obtención de los datos, tiene varias ventajas sobre otros programas: 1)

7
podemos obtener resultados no solo de distribución de especies sino de uso de hábitat de
diferentes especies en diferentes localidades (Tabla 1), 2) evita complicaciones
asociadas a las relacionadas con los métodos que analizan datos de presencia-ausencia
de las especies, 3) supera la eficacia de otros modelos -que al igual que MaxEnt- solo
utilizan datos de presencia de especies, 4) presenta poca sensibilidad a los errores
asociados con las geolocalizaciones y puede arrojar resultados con solo 5 registros de
presencia.
Tabla 1. Ejemplos de estudios de vida silvestre utilizando el programa de MaxEnt para generar
mapas de distribución de especies y estudios de uso del hábitat. Modificado de: Baldwin R., 2009,
Use of Maximum Entropy Modeling in Wildlife Research.
Referencia Especies Localidad
Pearson, R.G. et al, 2009 Geckos (Uroplatus spp.) Madagascar
Baldwin, R. B. y Bender L. C., 2008 Oso negro americano (Ursus
americanus)
Colorado, EUA
DeMtteo, K. E. y Loiselle, B. A.,
2008 en Baldwin R., 2009
Perro de monte (Speothos venaticus) Centroamérica y
Sudamérica
Suárez-Seoane, S. et al, 2008 Sisón común (Tetrax tetrax) España
Yost, A. C. et al, 2008 Gallo de salvia (Centrocercus
urophasianus)
Oregon, EUA
Boubli, J. P. y de Lima, M. G., 2009 Sakí barbudo de espalda roja
(Chiropotes israelita)
Uacarí de cabeza negra (Cacajao spp.)
Brasil
Roder, D. y Weinsheimer, F., 2009 Cuban treefrog (Osteopilus
sepentrionalis)
Caribe y Golfo de
México
Thorn, J. S. et al, 2009 Loris perezosos (Nycticebus spp.) Sureste de Asia
Hoenes, B. D. y Bender, L. C., 2010 Venado bura (Odocoileus hemionus)
Gacela órice (Oryx gazella)
Nuevo México, EUA
La pregunta si el nicho de una especie se mantiene en el tiempo y el espacio involucra
aspectos evolutivos (conservación del nicho) y ecológicos (estabilidad del nicho)
(Wiense y Graham, 2005; Pearman et al., 2008).
La modelación de nicho de especies que presentan poca sensibilidad a los cambios en el
ambiente y que presentan una distribución amplia puede considerarse un conocimiento
básico para mantener en equilibrio funcional los ecosistemas y ser importante para la

8
promoción de la biodiversidad en áreas específicas y así considerar estos lugares como
sitios prioritarios para la conservación los cuales deberán permanecer sin perturbación.
II.IV Ocupación de las especies.
Cambios aparentes en la abundancia de las especies animales se infiere a partir de los
cambios en los componentes vegetativos de sus hábitats, ésta es una de las principales
razones para realizar el monitoreo a nivel de especie. Dado que las especies son los
agentes fundamentales de la transferencia de materia y energía en los ecosistemas, la
dinámica de estos ecosistemas a menudo se impulsa por un pequeño número de especies
que tienen efectos desiguales en los procesos del mismo ecosistema. Debido a que los
resultados obtenidos mediante el monitoreo de las especies son un indicador de la
viabilidad de la población, el conocimiento de la dinámica espacio-temporal de la
abundancia de la especie se convierte en conocimiento fundamental para la evaluación
de los riesgos de las especies raras (Holmes, 1993; Skellam 1951 en Hui et al, 2012) y
especies que por su papel funcional y distribución geográfica son fundamentales para la
estabilidad de los ecosistemas, por lo tanto, la ocupación, definida como la proporción
de unidades de muestreo en las que una especie es detectada, es una variable de estado
que debe ser monitoreada, ya que se puede basar en observaciones en tiempo real
(observaciones directas y foto trampeo) de una especie en un sitio de muestreo o en
algún momento del pasado reciente, puesto que los patrones temporales y espaciales de
los datos de ocupación están relacionados con cambios en la abundancia de los animales
y, proporciona ideas sobre la probabilidad de persistencia de una especie (Noon, et al.
2012). Si bien se consideran enfoques indirectos, son una necesidad pragmática para
evaluar la diversidad global de plantas y animales (Wiens et al, 2008; Noon et al. 2009;
Caro 2010). Estimar la ocupación de una especie a nivel de paisaje es un método
factible, que relacionado con su papel funcional en el ecosistema puede ser de utilidad
para proponer objetivos de manejo específico para su conservación; el modelado de los
cambios en la ocupación del nicho por la especie será una forma de monitorear
respuestas a través del tiempo a escala de paisaje.
Los datos de detección y el modelo de probabilidad se combinan para formar una
función de verosimilitud obteniéndose estimaciones de ocupación y probabilidad de

9
detección a través del programa MARK (White y Burnham, 1999). Los modelos de
ocupación utilizan datos de detección/no detección (presencia/ausencia) a partir de
muestreos repetidos para estimar la probabilidad de detección, al mismo tiempo permite
incorporar covariables categóricas y cualitativas asociadas a las detecciones,
desarrollando así historias de captura de la especie en un tiempo y espacio determinado.
II.V. Zorra Gris.
Algunas especies dentro de un ecosistema pueden verse “favorecidas” por ambientes
impactados por sus hábitos alimenticios y capacidades adaptativas, por ser generalistas y
tener un amplio rango de distribución, mismas que pueden considerarse fundamentales
para la estabilidad del ecosistema (Brown, 2003). En este tipo de especies, que por ser
generalistas tienden a ser abundantes (Gastón et al., 2000) se encuentra la zorra gris,
Urocyon cinereoargenteus, que es un cánido pequeño, de hocico agudo, orejas erectas y
grandes, con una cola larga y densa, Fig. 1. El color general es gris en el dorso y blanco
en el pecho y vientre; entre el gris y el blanco normalmente hay una parte de color
pardo, de diferentes tonalidades. Los animales adultos pesan entre 2.5 y 7 kg. Su
distribución comprende desde el sur de Canadá hasta Colombia y Venezuela. Es el
carnívoro más ampliamente distribuido en México, ya que se encuentra en todo el país.
Pueden encontrarse casi en cualquier ambiente, es un animal solitario, activo tanto de día
como de noche y tanto en tierra como en los árboles. Es el único cánido que
regularmente sube a los árboles gracias a sus garras semi-retráctiles (Aranda, 2000). Es
por esta razón que es considerado uno de los miembros más primitivos de la familia de
los cánidos (Fritzell y Haroldson, 1982). Sus hábitos alimenticios comprenden una
amplia variedad de presas y vegetales; incluyen roedores (Microtus spp., Neotoma spp.,
Peromyscus spp., Reithrodontomys spp. y Sigmodon hispidus entre otros); invertebrados,
particularmente los ortópteros, y vegetales; en una frecuencia menor puede consumir
aves, además de especies domésticas y carroña de venados del género Odocoileus spp.
(Glover 1949, Hatfield 1939, Korschgen 1957, McGregor 1942, Nelson 1933, Salas
1988, Scott 1955). Debido a la relativamente alta abundancia y bajo valor económico de
las zorras grises, se cuenta con poca información sobre esta especie, principalmente en
Sonora. Se considera necesario el obtener información ecológica y demográfica básica

10
para cada uno de los hábitats más importantes ocupados por las zorras grises. Además,
se necesitan más datos sobre la respuesta a los paisajes alterados por el hombre; muy
poco se sabe sobre su estado y ecología fuera de EE.UU. y Canadá a pesar de que la
influencia de las zorras grises sobre las poblaciones de vertebrados más pequeños
pueden ser importantes (List, 2008).
Hay información acerca de la zorra gris que nos dice que está muy extendida en
bosques, matorrales, zonas de arbustos y hábitats rocosos en las regiones templadas y
tropicales de América del Norte y en las regiones montañosas septentrionales de
América del Sur. En cambio hay poco sobre su distribución en zonas desérticas. No hay
suficiente evidencia de que su población haya variado en cualquier parte de su área de
distribución y no se considera que la especie esté amenazada en la actualidad (Cypher et
al., 2008).
Figura 1. Zorra gris en zona costera del Cañón de Las Barajitas.

11
II.VI Foto Trampeo.
A pesar de que las trampas cámara han estado disponibles desde principio del siglo XX
(Cuttler y Swann, 1999) su utilización en inventarios de fauna silvestre y en inves-
tigación ecológica es relativamente reciente, Royama (1959, 1970), en uno de los
trabajos pioneros con trampas cámara logró identificar los componentes de la dieta de un
ave paseriforme llevados a los nidos por medio de una cámara activada con un
disparador automático. A partir de estos estudios, las trampas cámara han sido utilizadas
para documentar y estudiar la depredación en nidos de aves, la dieta, los patrones de
actividad de diversas especies, así como para registrar la presencia de ciertas especies en
un área determinada (Cuttler y Swann, 1999). Una de las formas más sencillas de
registrar la presencia de especies en un área determinada es por medio de observaciones
directas a lo largo de transectos establecidos, sin embargo, la observación directa de
muchas especies es un evento raro y ocasional, en estos casos los rastros, huellas y
excrementos son muy valiosos, ya que en la mayoría de los casos permite identificar al
mamífero hasta el nivel de especie, mediante esto podemos determinar que especies
están presentes en un área determinada, finalmente, las trampas cámara pueden
considerarse una herramienta no invasiva para detectar presencia de especies de una
manera más eficaz (Chávez et al., 2013). Muchas de las publicaciones con estudios de
trampas cámaras han tenido el objetivo de estimar la abundancia y el estudio es
diseñado para determinar el tamaño de una pequeña poblacional en un tiempo y espacio
dentro de un sistema (Karanth 1995; Karanth y Nichols 2000; Silver et al. 2004), dado
que es prácticamente imposible contar a todos los individuos de una población dentro de
un área específica, es necesario registrar las características de una proporción de la
población para hacer inferencias sobre la población real (Chávez et al., 2013).

12
II. VII La Sierra El Aguaje y el Cañón de Las Barajitas.
Forma parte de una larga región volcánica, la cual se origina por enormes erupciones
resultantes de un colapso de decenas de calderas a lo largo de kilómetros. Se localiza en
el límite sur del Desierto Sonorense, cerca de Guaymas y el límite oriental en el centro
de Sonora, presenta una amplia transición al matorral espinoso (Felger, 1999). En su
conjunto la Sierra El Aguaje tiene un aspecto agreste, elevándose abruptamente a más de
300 msnm, las máximas altitudes que alcanza esta sierra son de 740 msnm en el Cerro
Los Algodones o Cerro Agua Caliente y de 860 msnm en el Cerro El Parapeto o El
Tambor (Gallo-Reynoso y González-Martínez, 2003; Ventura, 2003; INEGI, carta
H12C89). La Sierra El Aguaje se caracteriza por su gran número de cañones de los
cuales once son los principales: San Pedro, Las Barajitas, Los Anegados, El Orejano, El
Alacrán, San Vicente, La Balandrona, Las Pirinolas, La Cantera, El Nacapule y el
Guálamo, en estos cañones se encuentran grandes bancos de obsidiana, cuarzo, geodas,
así como un sin número de cuevas producto de la erosión eólica y por derrumbamientos;
también existen sitios arqueológicos y pinturas rupestres. En ellos es posible encontrar
tinajas con aguas de escurrimiento de tipo volcánico, algunas son perennes, mientras que
otras son intermitentes y dependen de las lluvias de invierno y de verano para almacenar
agua (Gallo-Reynoso y González-Martínez, 2003).
El Cañón de las Barajitas se localiza 40 km al noroeste de Guaymas, el acceso es por el
mar, los segmentos norte y sur del Cañón de Las Barajitas se cortan en un colapso
riolítico que corresponden a bloques de roca que se deslizan uno sobre la otra, en la boca
del cañón, el drenaje pasa de roca volcánica a aluvión cuaternario consistiendo de arenas
no consolidadas y grava derivada de las montañas (Felger, 1999). Esta región se localiza
en una zona geotérmica potencial en la que se encuentran manantiales de aguas térmicas
como el Aguaje de Robinson y el Aguaje de Santa Úrsula; en uno de estos aguajes las
aguas alcanzan temperaturas de 40°C (Gallo-Reynoso y González-Martínez, 2003).
Esta característica de tener aguajes y lloraderos de agua ha dado nombre a la sierra en su
conjunto y ha permitido un considerable aumento en la biodiversidad al incluir una rica
flora y vegetación subtropical y fauna que está ausente en otras partes del Desierto
Sonorense y que presentan endemismos (Gallo-Reynoso y González-Martínez, 2003).

13
II. VIII Clima
El área de estudio, cuenta con un tipo de clima descrito como muy seco de tipo desértico
o BW, el cual es el más seco de los secos (INEGI, 1985; 2000), este clima se describe
con regímenes de lluvia en verano, porcentaje de lluvia invernal entre 5 y 10.2, con
temperatura media anual > de 22°C, la temperatura media del mes más frío es < 18°C.
Felger (1999), describió el tipo de clima como árido, con régimen de lluvias de dos
estaciones y con una alta variabilidad. Las lluvias de invierno-primavera derivan de las
tormentas frontales del Pacífico, que ofrecen lluvia o llovizna durante muchas horas; las
lluvias de verano resultan de los vientos monzónicos de origen tropical y se caracterizan
por violentas tormentas con rayos dramáticos y lluvias fuertes pero de corto tiempo. Los
desiertos que presentan dos temporadas de lluvia o régimen de lluvias bimodal muestran
una flora con afinidad Neotropical (Peinado et al., 2008, 2009).
II. IV Bioclima y Vegetación.
Dentro del cinturón subtropical anticiclónico es un área en la cual las masas de aire
descienden asociadas con zonas de alta presión atmosférica, y prevalecen los cielos
claros. Es donde ocurren los grandes desiertos de Norteamérica, la franja latitudinal
entre este cinturón y la zona de convergencia intertropical es dominada por vientos del
Este. La transición de condiciones de tropical árido a tropical semiárido pueden
observarse en conexiones con los tipos de hábitat representados por tipos de vegetación,
de forma que hacia el norte de Guaymas y el Río Sonora los vientos del Este empiezan a
tener su efecto, hay presencia de vegetación tropical semiárida, confinada a depresiones
limítrofes. En el Sur de Guaymas las plantas típicas desérticas dejan de observarse y los
matorrales espinosos y zonas arbóreas empiezan a dominar. Pequeñas áreas de
vegetación desértica se observan en los sitios más áridos del imperante bioclima
Tropical Semiárido (Peinado et al., 2010). Dicho bioclima está dominado por
comunidades vegetales xeromorficas, deciduas, espinosas, hojas pinnadas, árboles o
arbustos con varios troncos de 2-10 m de altura, fuerte composición de suculentas, los
arbustos tienden a crecer como árboles, los árboles con hojas pequeñas son dominantes,
principalmente leguminosas. El cardón (Pachycereus pecten aborigen) y el mauto

14
(Lysiloma divaricatum) son los mayores bioindicadores de zonas tropicales semiáridas
(Peinado et al., 2010).
La diversidad de plantas de Sonora refleja la mezcla de grandes provincias
biogeográficas. Los trópicos del Nuevo Mundo alcanzan su límite septentrional en
Norteamérica en el Este de Sonora, hay especies tropicales que alcanzan su límite Norte
en la transición del matorral espinoso costero al Desierto sonorense cerca de Guaymas
(ca. 28° N), o en poblaciones aisladas en cañones o cumbres de sierras en el desierto
(Van Devender et al., 2009).
Shreve (1951) designa la región del Cañón de Las Barajitas como un segmento de la
subdivisión Costa Central del Golfo, del Desierto Sonorense. Las altas elevaciones de la
Sierra El Aguaje, especialmente la parte Norte y muchos de los hábitats riparios,
sostienen vegetación que puede ser clasificada como matorral espinoso, en ocasiones se
asemeja a bosque tropical deciduo (Felger, 1966; Felger y Lowe, 1976), ésta ocurre en
ambos lados del Golfo de California con muchas especies en común (Shreve, 1964). Las
plantas típicas de Baja California en la costa sonorense son relictas de la Edad de Hielo.
Los interglaciares más calientes y más secos como el presente Holoceno dieron como
resultado contracciones repetidas de los rangos de distribución de estas plantas en
poblaciones más pequeñas y aisladas, sitios ideales para la especiación (Van Devender,
et al., 2009).

15
III. JUSTIFICACIÓN
Las especies de amplia distribución tienden a ser abundantes y muestran flexibilidad
biológica; la pérdida del hábitat y del nicho ecológico, invariablemente se traducirá en
pérdida de especies, conllevando a la disminución de la biodiversidad; de tal manera que
el conocimiento del nicho ecológico y la ocupación de la zorra gris (Urocyon
cinereoargenteus), dentro del área de estudio darán la línea base y otras pautas para su
conservación, y al mismo tiempo de su entorno a nivel ecosistema; por lo que la
conservación deberá estar enfocada hacia la prevención de la fragmentación y pérdida de
sitios que posean una riqueza específica para esta especie. Estas zonas por lo general
albergan especies de amplia distribución que forman parte de su alimentación, ya que
por sus hábitos tienden a ser fundamentales para mantener el equilibrio de los
ecosistemas y ser promotores de la biodiversidad.
El modelado de la distribución y la ocupación de la zorra gris en el Cañón de Las
Barajitas como zona de baja perturbación, proveerá de herramientas para la toma de
decisiones sobre la protección de la zorra gris y su entorno, a partir del conocimiento de
su nicho ecológico en este cañón, y con este modelo aplicarlo a otras zonas en donde se
ha observado a esta especie para detectar aquellas áreas en donde sea probable su
distribución y de esta manera conocer su estado de conservación. El modelado de
ocupación del hábitat implica el estatus de una población, y puede ser una herramienta
fiable capaz de medir su variación poblacional en el espacio y el tiempo y permite
describir factores asociados a la presencia y nicho ecológico de la zorra gris en un
ambiente con poca perturbación.

16
IV. HIPÓTESIS
La probabilidad de detección y la ocupación de la zorra gris estarán relacionadas con la
diversidad florística del Cañón de Las Barajitas, lo que permitirá modelar su distribución
potencial donde los sitios con mayor diversidad florística presentarán mayor
probabilidad de ocurrencia de esta especie.

17
V. OBJETIVO GENERAL
Estimar la ocupación y modelar el nicho ecológico de la zorra gris en el Cañón de Las
Barajitas.
VI. OBJETIVOS ESPECÍFICOS
Estimar la probabilidad de detección de la zorra gris en el Cañón de Las
Barajitas.
Análisis de los patrones de actividad de la zorra gris y especies simpátricas del
nicho trófico.
Generar registros de las especies presa base de la cadena trófica.
Evaluar la diversidad florística en el Cañón de Las Barajitas.

18
VII. MATERIALES Y MÉTODOS.
VII. I Área de Estudio: Cañón de Las Barajitas.
El área de estudio (figura 2) corresponde en su totalidad a la provincia llanura sonorense
y a la subprovincia aparatos volcánicos aislados y a la región sierra escarpada compleja
Sierra Libre (INEGI, 1981), con las siguientes clasificaciones: uso de suelo: Matorral
Sarcocaule (figura 3); suelos leptosoles y regosoles, rocas ígneas intrusivas en la sierra y
planicies de origen aluvial (figura 4); (Carta geológica 1:250 000 Sierra Libre H12-11)
(figuras 5-7).
El área total muestreada fue de 5.48 km².
Figura 2.- Fotografía satelital de Google Earth del Cañón de Las Barajitas donde se
indica el polígono del área de estudio (sombreado verde) con vista hacia el N. Fecha 22
de junio del 2010, satélite DigitalGlobe.

19
Figura 3.- Uso de Suelo y vegetación (Carta geológica 1:250 000 Sierra Libre H12-11
INEGI 1981).
20
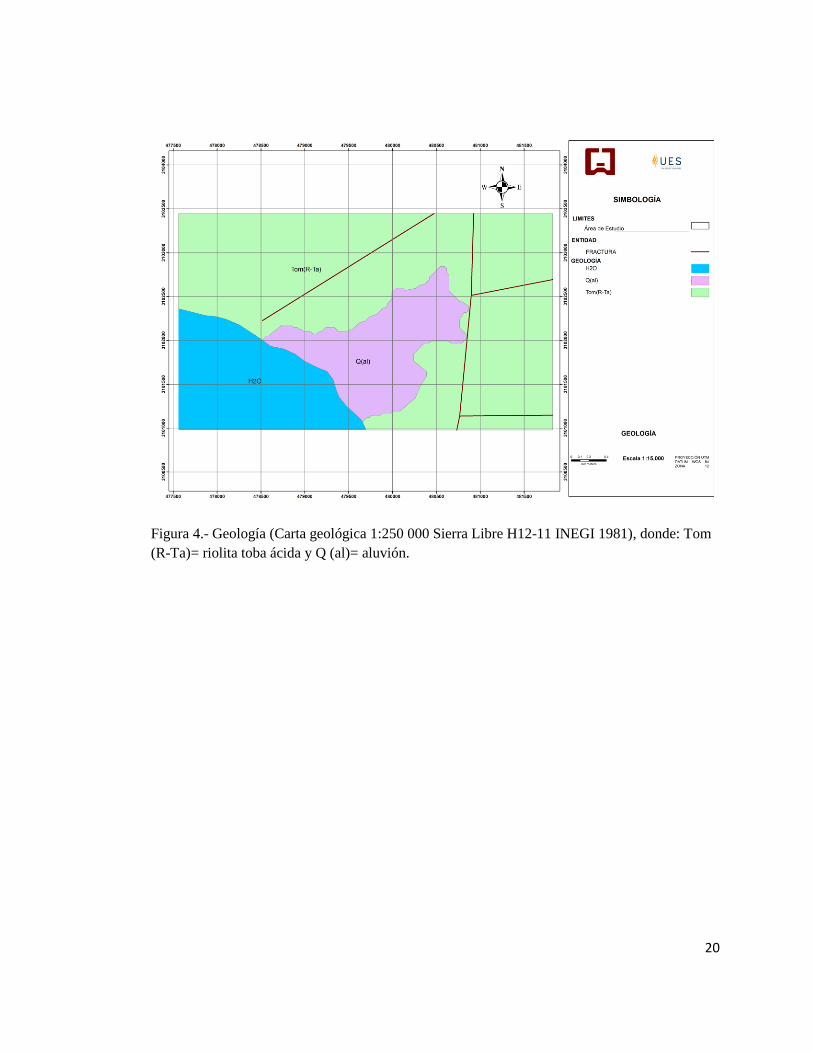
Figura 4.- Geología (Carta geológica 1:250 000 Sierra Libre H12-11 INEGI 1981), donde: Tom
(R-Ta)= riolita toba ácida y Q (al)= aluvión.
21
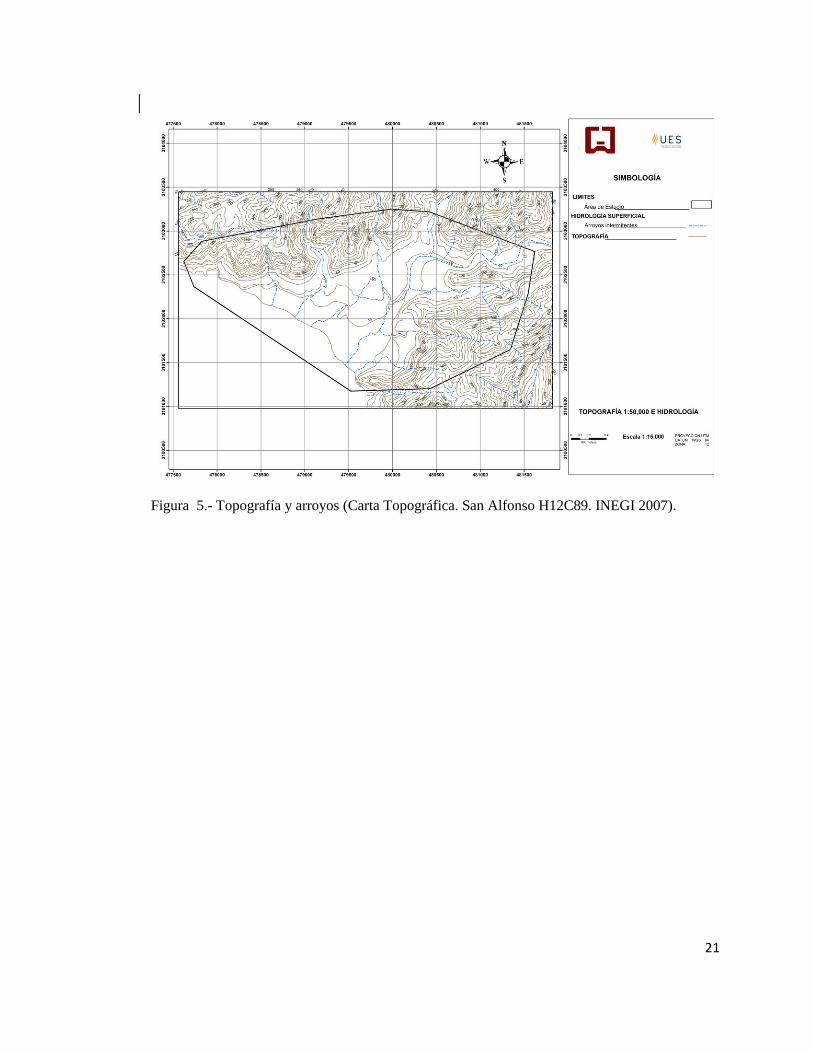
Figura 5.- Topografía y arroyos (Carta Topográfica. San Alfonso H12C89. INEGI 2007).

22
Figura 6.- Fisiografía (Carta Fisiográfica. Tijuana. Escala 1:1’000,000. INEGI 2006).
23
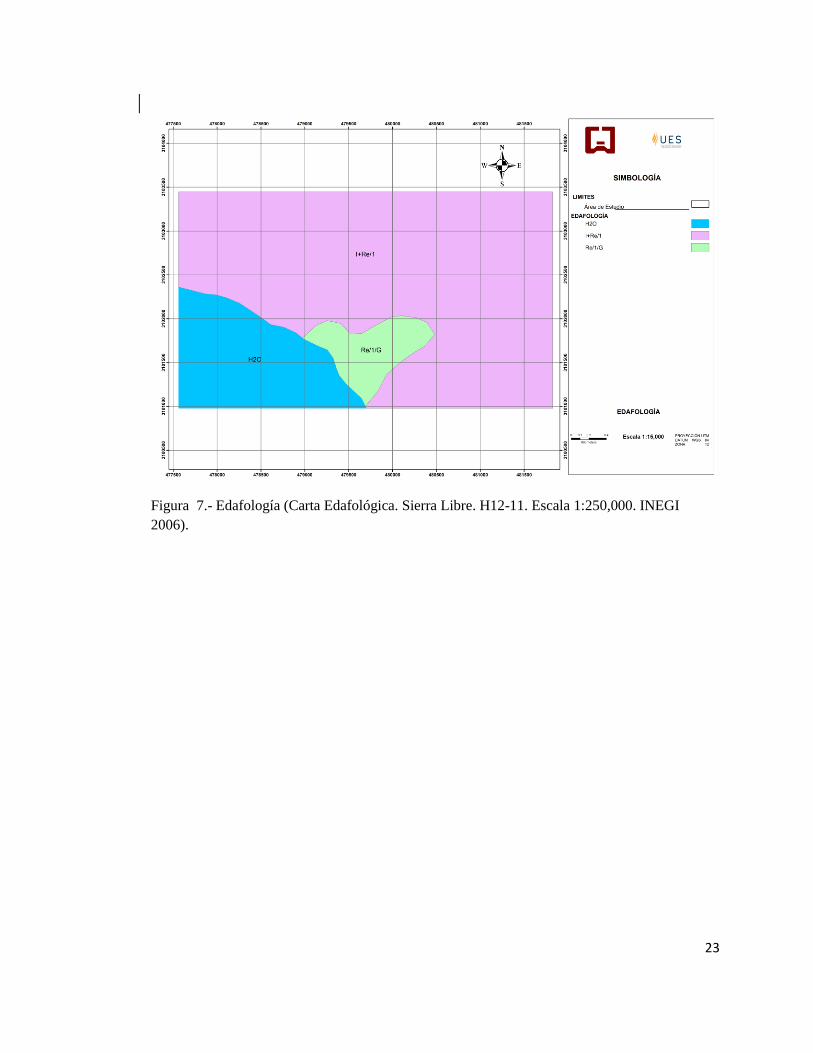
Figura 7.- Edafología (Carta Edafológica. Sierra Libre. H12-11. Escala 1:250,000. INEGI
2006).

24
VII.II Estimación de la Presencia de Zorra Gris y sus Parámetros Poblacionales
Mediante el Uso de Cámaras Trampa.
1. Se realizó una salida de reconocimiento del sitio en la cual se buscaron rastros de
la especie de interés, así como caminos de fauna, letrinas y huellas.
2. Localizados los rastros se establecieron de 12-14 estaciones de foto trampeo a lo
largo de un año, con cámaras trampa marca Wildview Xtreme 4, Bushnell
Trophy Cam, y Bushnell Trophy Cam HD, éstas se colocaron en troncos de
árboles o lugares que brindaron soporte al equipo, a 50 cm aproximadamente de
altura, la orientación de la cámara debió ser hacia el norte, evitando la puesta y la
salida de sol.
3. Cada estación de foto trampeo tuvo al menos 500 m de separación de las otras.
4. Se colocó un atrayente que consiste en una mezcla de sardina enlatada en salsa
de tomate, pasas y manzanas.
5. La cámara trampa se programó para que tomase tres fotos segundo una vez que
se detecta movimiento.
6. Cada cámara trampa estuvo en funcionamiento entre 20 a 39 días.
Cada lugar con cámara trampa se define como una estación de foto trampeo, estas
estaciones varían en cada muestreo con el fin de cubrir la mayor área posible a lo largo
de un año siguiendo las estaciones anuales; las cámaras permanecieron en campo por al
menos 25 días (Chávez et al., 2013).
VII.II.I Análisis.
Abundancia relativa= número de registros independientes / esfuerzo de muestreo.
Esfuerzo de muestreo= días trampa o noches trampa expresado como el producto de
multiplicar el número de trampas activas por el número de días o noches en los que las
cámaras trampa estuvieron activas.

25
VII.III Trampeo de Ratones.
Para conocer la disponibilidad de especies presa de pequeños roedores para la zorra gris,
se realizó trampeo de ratones utilizando trampas tipo Sherman (White et al, 1998); 20
trampas fueron colocadas cada 15 m, en lugares donde se observaron madrigueras o
presencia de roedores, las trampas se colocaron por la noche y se revisaron al amanecer,
dejando atrayente fuera y dentro de la trampa, éste consistió en una mezcla de 1 kg de
avena, 200 g de alimento para pájaros y varias gotas de vainilla. Estas trampas son para
captura en vivo, es decir, no mata o causa daño a los animales, los individuos capturados
fueron medidos, pesados, sexuados, fotografiados para su posterior identificación y
liberados.
VII.III.I Análisis de las capturas de ratones.
Frecuencia absoluta= total de especies en cada trampeo.
Frecuencia relativa= número de individuos por especie/ frecuencia absoluta.

26
VII. IV Descripción de la Vegetación.
Para estimar la acumulación de especies vegetales en los lugares donde se detectó la
presencia de zorra gris, se muestrearon 14 sitios cercanos a las estaciones de foto
trampeo, 11 por el método de cuadrantes con un tamaño de 25 x 50 m, y tres por releves
de vegetación. En cada cuadrante se registraron las especies de plantas perennes
presentes y la cantidad de cada una de ellas para obtener la diversidad y la densidad de
especies, así como la altura de 5 individuos al azar de cada especie para determinar la
estructura vertical de la comunidad vegetal (Wilder, comunicación personal, 2014), se
estimó un valor de importancia para cada cuadrante que resultó de la suma de la
densidad relativa, frecuencia relativa y dominancia de la estructura vertical relativa.
El muestreo por el método de releves de vegetación (Braun-Blanquet et al., 1932)
consistió en determinar un área mínima en la cual a cada especie presente se le asigna un
valor según sea su abundancia basándose en el cálculo de la cobertura visual como
sigue: 3) >75%, 2) 50-75%, 1) 25-50%, P= presente cuando la cobertura sea <5% y R=
rara cuando se encuentre un solo individuo presente, se anotan características del
sustrato y altura del sitio muestreado.
VII.IV.I Análisis de los Cuadrantes.
Para construir una tabla para cada cuadrante se aplicaron las siguientes fórmulas
(Martínez et al., 2013; Gea, 2005).
Densidad= Número de individuos por unidad de superficie.
Dominancia en estructura vertical= Altura promedio de cada especie.
Altura relativa= Altura de cada especie/ suma total de alturas promedio x 100.
Frecuencia= Cantidad de cada especie por cuadrante.
Frecuencia relativa= Frecuencia de una especie x100/ valores de frecuencia de todas las
spp.
Valor de importancia= densidad +altura relativa +frecuencia relativa.
El valor de importancia se utilizó como una covariable relacionada a la probabilidad de
captura y ocupación de la zorra gris mediante el programa MARK (White, 2001).

27
VII.IV.II Análisis de los Releves de Vegetación.
En primer lugar para cada especie se calcula el índice de presencia (tabla 2), después se
determina la fidelidad que es la mayor o menor restricción de cada especie a la
comunidad objeto de estudio (tabla 3), una vez obtenidas las tablas mencionadas se
caracterizaron y describieron a detalle.
Tabla 2. Grados de presencia (constancia).
Índice Presencia (%)
R 0-5%
P 5%
1 25-50%
2 50-75%
3 >75%
Tabla 3. Grados de fidelidad. Los grados de presencia y los grados de fidelidad son
utilizados para evaluar la vegetación muestreada por el método de Braun-Blanquet,
modificada de: El método fitosociológico, Dr. Francisco Alcaraz, Universidad de
Murcia, España, 2013.
Valor Carácter
3 Especie característica
2 Especie selectiva
1 Especie preferente
P Especie compañera
R Especie accidental
28
VII.IV.III Acumulación de Especies.
La función de la acumulación de especies vegetales, obedece al supuesto de que se
predice la riqueza total de un sitio, como el valor del número de especies en la cual una
curva de acumulación de especies alcanza la asíntota, para modelos de regresión lineal
se calcula como la relación a/b (Soberón y Llorente, 1993). Conforme la lista de
especies aumenta, la probabilidad de añadir una especie nueva a la lista disminuye de
forma exponencial, hasta llegar a cero. Estas funciones de acumulación de especies
basadas en un modelo adecuado permiten la predicción de la riqueza específica
(Moreno, 2001).
Para la función de la acumulación de especies, se creó una matriz en la cual se anotaron
los cuadrantes y relevés muestreados y cómo se fueron incrementando el número de
especies conforme el muestreo aumentaba (Tabla 4), esta matriz se analizó con el
programa SigmaPlot 11.0
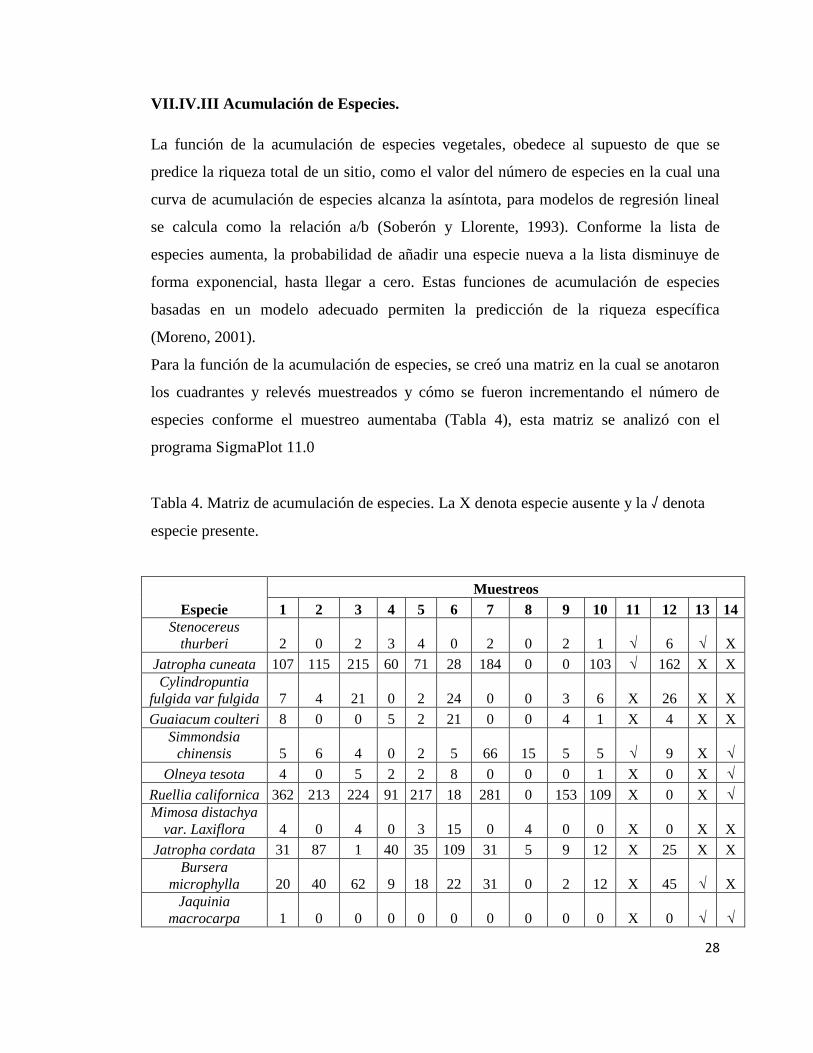
Tabla 4. Matriz de acumulación de especies. La X denota especie ausente y la √ denota
especie presente.
Especie
Muestreos
1 2 3 4 5 6 7 8 9 10 11 12 13 14
Stenocereus
thurberi 2 0 2 3 4 0 2 0 2 1 √ 6 √ X
Jatropha cuneata 107 115 215 60 71 28 184 0 0 103 √ 162 X X
Cylindropuntia
fulgida var fulgida 7 4 21 0 2 24 0 0 3 6 X 26 X X
Guaiacum coulteri 8 0 0 5 2 21 0 0 4 1 X 4 X X
Simmondsia
chinensis 5 6 4 0 2 5 66 15 5 5 √ 9 X √
Olneya tesota 4 0 5 2 2 8 0 0 0 1 X 0 X √
Ruellia californica 362 213 224 91 217 18 281 0 153 109 X 0 X √
Mimosa distachya
var. Laxiflora 4 0 4 0 3 15 0 4 0 0 X 0 X X
Jatropha cordata 31 87 1 40 35 109 31 5 9 12 X 25 X X
Bursera
microphylla 20 40 62 9 18 22 31 0 2 12 X 45 √ X
Jaquinia
macrocarpa 1 0 0 0 0 0 0 0 0 0 X 0 √ √
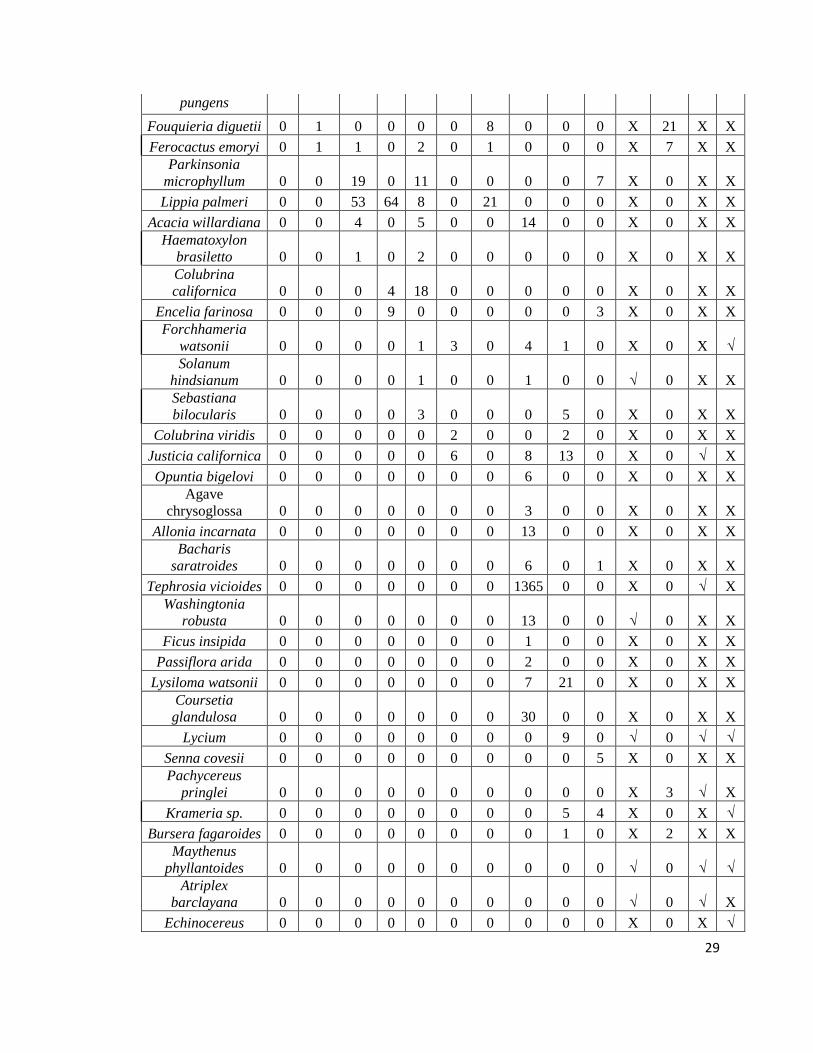
29
pungens
Fouquieria diguetii 0 1 0 0 0 0 8 0 0 0 X 21 X X
Ferocactus emoryi 0 1 1 0 2 0 1 0 0 0 X 7 X X
Parkinsonia
microphyllum 0 0 19 0 11 0 0 0 0 7 X 0 X X
Lippia palmeri 0 0 53 64 8 0 21 0 0 0 X 0 X X
Acacia willardiana 0 0 4 0 5 0 0 14 0 0 X 0 X X
Haematoxylon
brasiletto 0 0 1 0 2 0 0 0 0 0 X 0 X X
Colubrina
californica 0 0 0 4 18 0 0 0 0 0 X 0 X X
Encelia farinosa 0 0 0 9 0 0 0 0 0 3 X 0 X X
Forchhameria
watsonii 0 0 0 0 1 3 0 4 1 0 X 0 X √
Solanum
hindsianum 0 0 0 0 1 0 0 1 0 0 √ 0 X X
Sebastiana
bilocularis 0 0 0 0 3 0 0 0 5 0 X 0 X X
Colubrina viridis 0 0 0 0 0 2 0 0 2 0 X 0 X X
Justicia californica 0 0 0 0 0 6 0 8 13 0 X 0 √ X
Opuntia bigelovi 0 0 0 0 0 0 0 6 0 0 X 0 X X
Agave
chrysoglossa 0 0 0 0 0 0 0 3 0 0 X 0 X X
Allonia incarnata 0 0 0 0 0 0 0 13 0 0 X 0 X X
Bacharis
saratroides 0 0 0 0 0 0 0 6 0 1 X 0 X X
Tephrosia vicioides 0 0 0 0 0 0 0 1365 0 0 X 0 √ X
Washingtonia
robusta 0 0 0 0 0 0 0 13 0 0 √ 0 X X
Ficus insipida 0 0 0 0 0 0 0 1 0 0 X 0 X X
Passiflora arida 0 0 0 0 0 0 0 2 0 0 X 0 X X
Lysiloma watsonii 0 0 0 0 0 0 0 7 21 0 X 0 X X
Coursetia
glandulosa 0 0 0 0 0 0 0 30 0 0 X 0 X X
Lycium 0 0 0 0 0 0 0 0 9 0 √ 0 √ √
Senna covesii 0 0 0 0 0 0 0 0 0 5 X 0 X X
Pachycereus
pringlei 0 0 0 0 0 0 0 0 0 0 X 3 √ X
Krameria sp. 0 0 0 0 0 0 0 0 5 4 X 0 X √
Bursera fagaroides 0 0 0 0 0 0 0 0 1 0 X 2 X X
Maythenus
phyllantoides 0 0 0 0 0 0 0 0 0 0 √ 0 √ √
Atriplex
barclayana 0 0 0 0 0 0 0 0 0 0 √ 0 √ X
Echinocereus 0 0 0 0 0 0 0 0 0 0 X 0 X √

30
scopolorum
Carnegia
giganteae 0 0 0 0 0 0 0 0 0 0 X 0 √ X
Macroptilium
atrupurpureum 0 0 0 0 0 0 0 0 0 0 X 0 √ X
Gossypium
davidsoni 0 0 0 0 0 0 0 0 0 0 X 0 √ X
Hibiscus denudatus 0 0 0 0 0 0 0 0 0 0 X 0 √ X
Hofmeisteria
crassifolia 0 0 0 0 0 0 0 0 0 0 X 0 √ X
Bebbia juncea 0 0 0 0 0 0 0 0 0 0 X 0 √ X
Stegnosperma
halimofolium 0 0 0 0 0 0 0 0 0 0 X 0 √ X
Randia sonorensis 0 0 0 0 0 0 0 0 0 0 X 0 X √
Sideroxylon
occidentale 0 0 0 0 0 0 0 0 0 0 X 0 X √
Marsdenia edulis 0 0 0 0 0 0 0 0 0 0 X 0 X √
Prospois
glandulosa var.
Torreyana 0 0 0 0 0 0 0 0 0 0 X 0 X √
Ziziphus amole 0 0 0 0 0 0 0 0 0 0 X 0 X √
Sapindus
saponaria 0 0 0 0 0 0 0 0 0 0 X 0 X √
Phaulothamnus
spinescens 0 0 0 0 0 0 0 0 0 0 X 0 X √
Ziziphus obtusifolia 0 0 0 0 0 0 0 0 0 0 X 0 X √
Especies
acumuladas 11 13 17 19 22 24 24 34 37 38 41 42 49 57

31
VII.V Modelación de Nicho Ecológico.
Se utilizó el programa MaxEnt, este es un programa de acceso libre y lo que hace es
modelar la distribución potencial de las especies, utilizando como datos solamente los
sitios de presencia y variables ambientales asociadas a cada uno de esos puntos de
presencia.
La información sobre las colectas y poblaciones de distintas especies que interesa
modelar tienen que contar con coordenadas geográficas, que permite su ubicación
espacial y permite su posterior relación con los datos medioambientales, ecológicos,
geológicos, etc.; recabados para el sitio o la región en donde se colectó u observó el
organismo (Martínez, 2010).
Se utilizaron puntos de presencia de la zorra gris obtenidos con foto trampeo en el
Cañón de las Barajitas (para la validación del modelo); registros obtenidos del proyecto
“Cuatro Gatos” de la organización no gubernamental Sky Island Alliance y de la página
www.gbif.org. Las variables bioclimáticas se obtuvieron de la página
http://www.worldclim.org/bioclim, mismas que se indican en la Tabla 5.

32
Tabla 5. Código y descripción de las variables bioclimáticas obtenidas de WorldClim
utilizadas en el análisis (http://www.worldclim.org/bioclim).
Clave
BIO1
BIO2
BIO3
BIO4
BIO5
BIO6
BIO7
BIO8
BIO9
BIO10
BIO11
BIO12
BIO13
BIO14
BIO15
BIO16
BIO17
BIO18
Descripción
Annual Mean Temperature
Mean Diurnal Range (Mean of monthly (max temp - min temp))
Isothermality (P2/P7) (* 100)
Temperature Seasonality (standard deviation *100)
Max Temperature of Warmest Month
Min Temperature of Coldest Month
Temperature Annual Range (P5-P6)
Mean Temperature of Wettest Quarter
Mean Temperature of Driest Quarter
Mean Temperature of Warmest Quarter
Mean Temperature of Coldest Quarter
Annual Precipitation
Precipitation of Wettest Month
Precipitation of Driest Month
Precipitation Seasonality (Coefficient of Variation)
Precipitation of Wettest Quarter
Precipitation of Driest Quarter
Precipitation of Warmest Quarter
Una vez obtenidas las georeferencias de los sitios de presencia y las capas bioclimáticas,
se ingresan los datos en hojas de Excel, posteriormente estos datos se transformaron o
guardaron en formatos que puedan ser leídos por los programas de modelación de
nicho. Esto se hace por medio de Sistemas de Información Geográfica (Martínez,
2010). El programa se cargó con variables o capas bioclimáticas en formato ASCII, con
datos de presencia, con nombre de la especie y coordenadas en grados decimales
guardados en archivos de formato CSV (disponible en Excel). El resultado son mapas de
probabilidad de distribución en ASCII, una hoja de resultados en Html con imágenes de
los mismos mapas en (*png) y una serie de estadísticos de validación. Los datos se
cargan fácilmente por medio de una interfaz gráfica (Martínez, 2010), como la que se
muestra en la Fig. 8.

33
Figura 8.- Pantalla de la interfaz gráfica del programa MaxEnt.

34
VII.VI Estimación de Probabilidad de Captura y Ocupación del Hábitat de la
Zorra Gris.
Utilizando las capturas de zorra gris se hace una historia de captura para cada cámara en
la que se indica la presencia con el número uno y la ausencia de la especie con el cero,
se analizan e incluyen las covariables asociadas a cada estación de foto trampeo que
pueden ser cualitativas y categóricas, se crea un archivo .txt que es leído por el programa
MARK, el cual estima la probabilidad de la detección y el porcentaje de ocupación
basándose en la teoría de la probabilidad; todas las estimaciones son de máxima
verosimilitud, es decir que el valor de las variables estudiadas hacen máxima la
probabilidad de obtener la muestra observada (Edwards, 1992).
Se seleccionan los mejores modelos mediante el criterio de selección Akaike (1977) para
muestras pequeñas (AICc), éste enfatiza la bondad del modelo, seleccionando el mejor
modelo entre los modelos candidatos, mediante la maximización de la log-verosimilitud
esperada de un modelo determinado, mediante el método de máxima verosimilitud
(Félix, 2011). Se otorga el mayor peso al que represente un mayor soporte para
asemejar una realidad (Akaike, 1987).
Para el diseño de varios modelos se seleccionaron variables categóricas y variables
cualitativas asociadas a las capturas de zorra gris, éstas fueron: tipo de hábitat (cueva,
arroyos, sitios perturbados, caminos, aguajes y pie de montaña) y estación del año
(primavera, verano, otoño e invierno). Como variables cualitativas se consideraron:
Valor de importancia de la vegetación, presencia de especies simpátricas y altitud de la
estación de fototrampeo. Los modelos generados a partir de hipótesis se corrieron en el
programa MARK y se seleccionaron los dos mejores modelos (los de mayor peso), para
determinar mediante los resultados observados qué factores pueden ser determinantes
para la detección de la zorra gris y qué porcentaje de ocupación presenta en un espacio y
tiempo dado, bajo los enfoques de los modelos seleccionados.

35
VIII. RESULTADOS Y DISCUSIÓN
VIII. I Fototrampeo.
El esfuerzo de muestreo anual (días/noches de foto trampeo) fue de 164 días/noches y se
obtuvieron un total de 18,672 fotos, de las cuales 7,459 incluyeron fotos de animales,
575 corresponden a zorra gris, el resto fueron de especies simpátricas en su nicho trófico
(coyote, Canis latrans, cacomixtle, Bassariscus astutus y gato montés, Lynx rufus). Así
como de diversa fauna compuesta por: venado cola blanca (Odoicoileus virginianus),
coatí (Nassua narica), mapache (Procyon lotor), pécari (Tayassu tajacu), lepóridos
(Silvilagus audobonii y Lepus alleni) y el murciélago (Macrotus californicus). Especies
del Orden Rodentia (Neotoma albigula, Tamias dorsalis, Spemorphilus variegatus).
Aves rapaces (Cathartes aura, Buteo Jamaicensis ), gallináceas (Calliplepa gambelii y
Geococcyx californianus) y paseriformes (Corvus corax, Catherpes mexicanus, Icterus
cucullatus, Carpodacus mexicanus, Cardinalis cardinalis, y Pipilo chlorurus). Tabla 6.

36
Tabla 6.- Número de registros de cada especie foto trampeada.
Especie
No. fotografías obtenidas para
cada especie en las cámaras
trampa
Procyon lotor 2,652
Canis latrans 1,241
Tamias dorsalis 947
Tayassu tajacu 667
Urocyon cinereoargenteus 575
Bassariscus astutus 470
Odocoileus virginianus 186
Macrotus californicus 62
Neotoma albigula 36
Lynx rufus 15
Spemorphilus variegatus 12
Nassua narica 8
Lepóridos 52
Aves 536
En cada estación del año se registraron capturas en diferentes sitios, permaneciendo
constante el sitio del aguaje para tomarlo como referencia para la modelación en
MARK. La estación climática con mayores sitios de detección fue la primavera y los
lugares con mayor frecuencia de aparición fueron aquéllos en los que la cobertura
vegetal fue >75% (Figuras 9-12).
Bajo el supuesto de que todas las trampas tienen la misma probabilidad de captura, la
zorra gris pudo encontrarse en cualquier sitio dentro del hábitat que representa el Cañón
de Las Barajitas, así que las estaciones de foto trampeo en las que hubo más frecuencia
de captura (Figura 13) presentaron la siguiente similitud: ser sitios que pudieran brindar
seguridad para escapar de posibles depredadores por el tipo de dosel de la vegetación,
valor considerado mediante el método de releves de vegetación asociado a la densidad
de las especies (Tablas 7, 8 y 9).

37
Figura 9.- Estaciones de foto trampeo en verano. Los círculos verdes significan
presencia de zorra gris, los círculos rojos denotan ausencia de zorra gris.

38
Figura 10.-Estaciones de fototrampeo en otoño. Los círculos verdes significan presencia
de zorra gris, los círculos rojos denotan ausencia de zorra gris.

39
Figura 11.- Estaciones de fototrampeo en invierno. Los círculos verdes significan
presencia de zorra gris, los círculos rojos denotan ausencia de zorra gris.

40
Figura 12.- Estaciones de fototrampeo en primavera. Los círculos verdes significan
presencia de zorra gris, los círculos rojos denotan ausencia de zorra gris.

41
Figura 13.- Frecuencia de aparición (%) en los diferentes sitios de detección (presencia)
de la zorra gris.

42
Tabla 7. Evaluación de la comunidad vegetal costera por el método Braun-Blanquet. R=
especie rara y P= especie presente.
Especies Familia Índice Carácter
Maythenus phillantoides Celastraceae 3 sp característica
Simmondsia chinensis Simmondsiaceae 1 sp preferente
Jatropha cuneata Euphorbiaceae 2 sp selectiva
Lycium Solanaceae 1 sp preferente
Washingtonia robusta Palmae P sp compañera
Atriplex barclayana Chenopodiaceae P sp compañera
Forchhammeria watsonii Capparaceae R sp accidental
Stenocereus thurberi Cactaceae R sp accidental
Fabaceae Fabaceae 2 sp selectiva
Echnocereus scopulurum Cactaceae R sp accidental
Tabla 8. Evaluación de la comunidad vegetal del aguaje costero por el método de Braun-
Blanquet. R= especie rara y P= especie presente.
Especies Familia Índice Carácter
Washingtonia robusta Poaceae 3 sp característica
Jaquinia macrocarpa Theophrastaceae 1 sp preferente
Carnegia giganteae Cactaceae 2 sp selectiva
Pachycereus pringlei Cactaceae 1 sp preferente
Stenocereus thurberi Cactaceae 1 sp preferente
Bursera microphyllum Burseraceae 2 sp selectiva
Lycium Solanaceae P sp compañera
Maythenus pillantoides Celastraceae R sp accidental
justicia californica Acanthaceae P sp compañera
Macroptilium atroporpureum Fabaceae P sp compañera
Gossypium davidsonii Malvaceae P sp compañera
Hibiscus denudatus Malvaceae R sp accidental
Hofmeisteria crassifolia Asteraceae P sp compañera
Bebia juncea Asteraceae P sp compañera
Tephrosia vicioides Fabaceae 1 sp preferente
Stegnosperma halimifolium Phytolaccaceae P sp compañera

43
Tabla 9. Evaluación de la comunidad vegetal aledaña al arroyo principal por el método
de Braun-Blanquet.
Especies Familia Índice Carácter
Randia thurberi Rubiaceae 1 sp preferente
Simmondisa chinensis Euphorbiaceae 2 sp selectiva
Maythenus pillantoides Celastraceae 1 sp preferente
Forchammeria watsoni Capparaceae 1 sp preferente
Jaquinia macrocarpa Theophrastaceae 1 sp preferente
Lycium Solanaceae 1 sp preferente
Olneya tesota Fabaceae 1 sp preferente
Krameria sonorae Krameriaceae 1 sp preferente
Ruellia californica Acanthaceae 2 sp selectiva
Sideroxylon occidentale Sapotaceae 3 sp característica
Sapindus saponaria Sapindaceae 3 sp característica
Marsdenia edulis Asclepiadaceae 1 sp preferente
Prosopis glandulosa var torreyana Fabaceae 3 sp característica
Ziziphus amole Rhamnaceae 3 sp característica
Phaulothamnus spinescens Phytolacaceae 3 sp característica
Ziziphus obtusifolia Rhamnaceae 2 sp selectiva
Se consideraron 95 fotografías como registros individuales de zorra gris, 2 registros con
2 individuos (hembra y cría; y hembra y macho), ver figuras 14 y 15. Se consideró a una
foto independiente si pasaron 30 minutos de la siguiente foto de la misma especie
(Srbek-Araujo y Chiarello, 2005); así, según la fórmula de Chávez et al. (2013) para
estimar la abundancia relativa por medio del fototrampeo la relación es 95/129 por lo
que el IAR=0.73, para estandarizar el valor de la abundancia relativa y compararlo con
otros estudios la fórmula es IAR=Número de registros independientes/100 días trampa
(Monroy-Vilchis et al., 2009) resultando así un valor de 0.95, la abundancia relativa
reportada para una selva baja caducifolia es 1.53 (Monroy-Vilchis et al., 2010)
encontrando así que esta especie es más abundante en ambientes menos áridos, o con
vegetación más abierta, aunque se ha concluido que prefieren hábitats boscosos
(Yearsley y Samuel, 1980). Su presencia puede ser significativa en ambientes en los
cuales convergen especies vegetales áridas y subtropicales, si la densidad de la zorra gris

44
es de 1.2-2.1 individuos por km², dependiendo de la localidad, época y método de
estimación (Fritzell y Haroldson, 1982) aplicando la fórmula de Chávez, et al. (2013)
para el área de estudio la densidad será de 0.173 individuos por km².
Las foto-capturas en las cuales se observan la hembra con su cría en agosto (Fig. 14) y
los 2 individuos en febrero (Fig. 15) coinciden con lo citado por Fritzell y Haroldson
(1982) acerca de los meses en que la zorra gris se reproduce e indican que nacen entre
los meses de marzo y abril, y que las crías a los 4 meses son capaces de ser
independientes, por lo que la cría fotografiada en el mes de agosto, posiblemente tenga
esa edad. De los 2 individuos foto-capturados en el mes de febrero, pudieran tratarse de
hembra y macho en etapa reproductiva. En toda el área no hubo más foto-capturas de 2
individuos en los meses correspondientes en los que esta especie se reproduce, se puede
inferir que para el Cañón de Las Barajitas, la abundancia poblacional es al menos de 2
individuos en edad reproductiva.
Figura 14.- Las flechas indican a la hembra de zorra gris con su cachorro en el mes de agosto
2013, Aguaje Cañón de Las Barajitas.

45
Figura 15.- Dos individuos adultos de zorra gris en el Cañón de Las Barajitas en febrero de
2014.
VII.II Patrones de Actividad de Zorra Gris y Especies Simpátricas.
Para determinar el nicho ocupado en relación a su patrón de actividad, se analizaron los
registros de presencia de la zorra gris y especies simpátricas en intervalos de 2 horas a lo
largo del día, se determinó así la frecuencia de aparición (FA) correspondiente a cada
intervalo. El análisis de los patrones de actividad de la zorra gris y de las especies
simpátricas que ocurren en el Cañón de Las Barajitas se describe en la figura 4, donde se
puede observar que todas estas especies muestran una FA en horarios nocturnos y
crepusculares principalmente, el coyote es la especie que se puede considerar más activa
durante todo el día, pero tanto esta especie como las demás especies simpátricas
muestran preferencias por los horarios con menor luz.

46
Los coyotes (8-20 kg), los gatos monteses (5-15 kg) y las zorras grises (3-5kg) (Fedriani
et al., 2000) son especies simpátricas en la mayor parte de Norteamérica (Hill et al.,
1987) este mismo comportamiento se observa en el Cañón de las Barajitas, donde
además a las especies mencionadas se añade el cacomixtle (Fig. 16). Sargent (1982) y
Wooding (1985) sugieren que la relación simpátrica entre la zorra gris y el coyote ocurre
debido a las capacidades semi-arborícolas que posee la primera, lo que le proporciona
mecanismos efectivos para escapar de sus depredadores. Esta característica puede ser
considerada como una respuesta a la presión por depredación por el gato montés y el
coyote; al observarse que tiene los mismos patrones de actividad, dentro del horario
crepuscular y nocturno, se detecta la presencia de estas especies en intervalos de horas
similares. Al igual que el coyote es un depredador potencial de esta especie, así se puede
considerar que la zorra gris podría formar parte de la dieta de ambos carnívoros en caso
de: 1) escaséz de recursos alimenticios, 2) por depredación directa en las madrigueras de
esta especie, 3) la competencia es más probable que ocurra cuando un recurso
compartido es limitado (Begon et al., 1996). Adicionalmente las especies que son objeto
de depredación y presentan interacciones con un competidor más dominante suelen
desarrollar conductas de aislamiento que influyen en su abundancia y distribución
(Fedriani et al., 2000; Lindström et al., 1995).
El índice de la amplitud del nicho de alimentación anual en una región tropical para la
zorra gris y el coyote es de 15.32 y 14.95, respectivamente (Guerrero et al., 2002), este
resultado pudiera verse reflejado de igual forma en el Cañón de las Barajitas por la
presencia de frutos suculentos de la vegetación característica de ambientes subtropicales
que convergiendo en una zona desértica promueve la variabilidad de fuentes de
alimentación para especies carnívoras generalistas, ya que en los análisis cualitativos de
las heces fecales (n= 22) de la zorra gris en el Cañón de Las Barajitas se observaron
semillas de especies arbóreas como la uvalama (Vitex mollis), el San Juanico (Jacquinia
macrocarpa), la palma (Washingtonia robusta), el jito (Forchammeria watsonii) y el
limoncillo (Ziziphus amole).

47
Figura 16.- Patrones de actividad de la zorra gris a lo largo de un año (mayo 2013-abril 2014) el
eje de las “Y” refleja la FA.
VIII.III Especies Presa.
Se capturaron un total de 17 individuos (frecuencia absoluta) en 160 trampas Sherman
colocadas en transectos con diferente tipo de suelo a lo largo de un año con 2
repeticiones por temporada, los individuos capturados fueron de 4 géneros diferentes
correspondientes a dos familias: Heteromydae de los géneros: Dipodomys merriami
(n=2), Neotoma albigula (n=2) y Chaetodipus sp. (n=4) y Muridae con el género
Peromyscus sp. (n= 9), los primeros son especies granívoras nocturnas, prefieren
hábitats de cobertura escasa y alta pedregosidad; por otro lado las especies de la familia
Muridae son omnívoros (según la disponibilidad de alimento y temporada), con hábitos
trepadores (Michel et al., 2007), por los resultados observados también son especies
00:00 2:00 4:00 6:00 8:00 10:00 12:00 14:00 16:00 18:00 20:00 22:000
2
4
6
8
10
Cannis latrans
00:00 2:00 4:00 6:00 8:00 10:00 12:00 14:00 16:00 18:00 20:00 22:000
2
4
6
8
10
Urocyon cinereoargenteus
00:00 2:00 4:00 6:00 8:00 10:00 12:00 14:00 16:00 18:00 20:00 22:000
2
4
6
8
10
Bassariscus astutus
Hora
00:00 2:00 4:00 6:00 8:00 10:00 12:00 14:00 16:00 18:00 20:00 22:00
0
2
4
6
8
10
Primavera
Verano
Otono
Invierno
Felis rufus
Hora

48
nocturnas. Esto es un reflejo de la diversidad de alimento que presenta el Cañón de las
Barajitas, ya que Morris (1987) y Tews et al. (2004) mencionan que en los hábitats
terrestres, son los atributos de la comunidad vegetal los que determinan la estructura
física del ambiente y los que tienen influencia sobre la distribución e interacción de las
especies animales (roedores); por otra parte para Michel et al. (2007), los atributos del
hábitat, tanto a nivel del paisaje como a nivel local, están asociados a la composición y
dinámica de la comunidad de roedores. Aunque para Sonora, Hafner, (2012) dice que
no es particularmente rico en proporción de roedores, las 53 especies documentadas por
éste autor representan el 22% del total de las 241 especies mexicanas de roedores; en
cambio Sonora es relativamente rico en la familia Heteromydae (37.5% del total de la
familia Heteromydae en México), la alta representación de esta familia que
generalmente está adaptada a regiones áridas, refleja que los desiertos son la regiones
dominantes en el estado (Hafner, 2012). El alimento específico para la zorra gris varía
según la temporada y la localidad, Villalobos et al., 2014 menciona que esta especie se
comporta como insectívoro-granívoro durante todo el año con una marcada preferencia
hacia la frugivoría durante la temporada seca asociando este resultado con la fenología
de la fructificación; pero los mamíferos, particularmente del orden Rodentia constituyen
la base de su dieta (Fritzell y Haroldson, 1982; Arnaud y Acevedo, 1990) por lo que la
diversidad de estas especies constituye un beneficio para la obtención de proteínas,
como también favorecerá la diversidad florística específica del Cañón de Las Barajitas
lo que implicará determinado valor de ocupación del hábitat y nicho ecológico. En la
tabla 10 se describen los resultados de las capturas de los trampeos de roedores de mayo
2013-abril 2014, que representan el esfuerzo de captura de 20 trampas Sherman en 16
transectos.

49
Tabla 10. Especies de roedores capturados en en Cañón de las Barajitas con trampas
Sherman en 4 estaciones del año (FR, frecuencia relativa).
Especie Temporada Tipo de Suelo No. Individuos FR
Peromyscus sp. Primavera Gravoso 5 0.29
Chaetodipus sp. Primavera Gravoso 1 0.06
Dipodomys merriami. Primavera/verano Gravoso 2 0.12
Chaetodipus sp. Verano Gravoso/arenoso 1 0.06
Neotoma albigula Verano Gravoso 2 0.12
Peromyscus sp. Otoño Arenoso/gravoso 4 0.24
Chaetodypus sp. Invierno Arenoso 2 0.12
VIII.IV Vegetación.
La regularidad de la floración y la fructificación de las especies es de considerable
importancia en el sostenimiento de la fauna de determinada localidad (De la Luz et al.,
1996); en los diferentes cuadrantes muestreados se contabilizaron un total de 3,233
individuos pertenecientes a 26 especies diferentes, distribuidas en 13 familias, la familia
más representativa en diversidad fue la Fabaceae, pero las de mayor frecuencia y
dominancia fueron la Euphorbiaceae, Burseraceae y Acanthaceae, estando presente en
todos los cuadrantes muestreados. Hace falta hacer un estudio de la fenología floral de
esta región.
La comunidad vegetal que compone al Cañón de Las Barajitas pertenece a la
denominada Costa Central del Desierto Sonorense, conocida también como asociación
Bursera-Jatropha (Shreve, 1951). En esta evaluación de la flora en el Cañón de Las
Barajitas se observa que la asociación dominante de plantas es Bursera-Jatropha-
Ruellia, esta última de la familia Acanthaceae, es un arbusto desértico que presentó la
mayor abundancia en todos los cuadrantes (43.48%)
Respecto a la función de correlación lineal de la acumulación de especies, los valores de
a= 3.30 y b= 5.82 con una r²= 0.97, significan que a mayor muestreos el número de
especies diferentes aumenta y que el número de muestreos realizados no fueron los
suficientes para que la función llegara a la asíntota de su distribución, pero sí es
suficiente para representar una función lineal positiva como se observa en la figura 17.

50
Esto representa para la estructura vegetal del hábitat de la zorra gris, un ecosistema en
condiciones estables dada la relación que existe entre éste y la diversidad para mantener
los procesos ecosistémicos.
Figura 17. Función de acumulación de especies vegetales, resultado de la evaluación del
muestreo de la vegetación del Cañón de Las Barajitas.

51
VIII.V Probabilidad de Detección y Ocupación.
Para Peterson et al. (2011) existen factores del medio ambiente que definirán los
patrones de ocurrencia de las especies, considerando entonces que la detección y la
ocupación de las especies está dada en función a su nicho efectivo o realizado, es decir,
su persistencia al sitio es por que hay fuente de alimento y menor riesgo de depredación
lo que beneficia su éxito reproductivo. Usando el programa MARK (White, 2001), se
seleccionó los modelos de regresión lineal múltiple con mayor peso para la
probabilidad de detección (P) y la ocupación (ψ) de la zorra gris, bajo el criterio de
información Akaike (1977):
Modelo A:
P= -1.78
P= e− 1.78
/1+e− 1.78
, por lo tanto la probabilidad de detección bajo ninguna
covariable asociada será de 14.37%.
Ψ= -3.81-0.04 vegetación-0.01 altura -1.89 otoño -0.08 invierno -2.79 primavera
Psi= e− 3.81
/1+e− 3.81
, por lo tanto el porcentaje de ocupación correlacionada a
las variables de vegetación, altura y estación del año es de 2.15%.
Modelo B:
P= -3.52-0.10 otoño-1.06 invierno-1.46 primavera-1.66 -0.04 cacomixte-0.04 gato
montés-1.61 coyote
P= e− 3.52
/1+e− 3.52
, por lo tanto la probabilidad de detección correlacionada
a las variables de estación del año y presencia de especies simpátricas es de 2.85%.
Ψ= -1.99-1.97 otoño-1.06 invierno-1.46 primavera-1.66 -0.04 cacomixte-0.04 gato
montés-1.61 coyote

52
Psi= e− 1.99
/1+e− 1.99
, por lo tanto el porcentaje de ocupación
correlacionado a las variables estación del año y especies simpátricas es de 11.93%
Para un área de 5.48 km² puede considerarse que los valores de ocupación son altos,
dadas las condiciones climáticas del Cañón de Las Barajitas, ya que según los modelos,
la presencia de la zorra gris en este sitio está en función de las variables climáticas dadas
por las estaciones del año, presentando mayor influencia negativa en el verano y su
abundancia no está relacionada directamente con la diversidad florística, pero sí podría
serlo por la fenología floral.
Para probar si las diferencias entre los modelos son estadísticamente significativas se
aplicó una prueba de Mann-Whitney y se encontró que la diferencia en los valores de la
mediana (-2.6 y -2.9) entre los 2 grupos no es suficiente para excluir la posibilidad de
que las diferencias entre las probabilidades de detección y los porcentajes de ocupación
se deban a la variabilidad del muestreo, por lo que no hay una diferencia
estadísticamente significativa con una P= 0.667.
Se puede deducir que las condiciones bióticas dadas por la estacionalidad climática
proporcionan a la zorra gris un hábitat que le permite establecerse, según el modelo de
ocupación (Gastón, 1997) como una población cerrada, si se determina que la
ocupación del hábitat y el nicho ecológico medido con un modelo de relación log-lineal
permite identificar los efectos significativos entre los recursos y condiciones
ambientales, y éstos supondrán un volumen abstracto que afectará el nicho ecológico
propuesto por Hutchinson (1957).

53
VIII. VI Modelación del Nicho Ecológico.
Las bases de los modelos de la distribución de las especies o de su nicho ecológico son
representaciones cartográficas de las relaciones matemáticas o estadísticas entre la
distribución real conocida y un conjunto de variables independientes de un espacio para
la presencia de una especie en función de las variables empleadas para generar dicha
distribución, este valor representa una probabilidad de presencia para la detección
(Mateo et al., 2011), se corrieron tres modelos de distribución: 1) General (fig. 18), 2)
En Sonora (fig. 19) y 3) En el Desierto Sonorense (fig. 20). De los cuales las variables
bioclimáticas que fueron determinantes para generar los mapas fueron las siguientes:
• Mapa de la distribución general: bios 7, 15 y 3 con un AUC= 0.820 (fig. 21).
• Mapa de la distribución en el estado de Sonora: bios 19, 2 y 9 con un
AUC=0.821(fig. 22).
• Mapa de la distribución en el Desierto Sonorense: bios 15, 6 y 18 con
AUC=0.873 (fig. 23).
Las gráficas AUC, son equivalentes de U de Mann-Whitney ya que en realidad
corresponden a la probabilidad de que tomado al azar un par de casos, uno de presencia
y otro de ausencia (o pseudoausencia), el modelo que adjudica a la presencia un valor
mayor de idoneidad o probabilidad y esto para todos los posibles pares en la muestra,
este valor está comprendido entre 0 y 1; un valor de 1 indica que todos los casos se han
clasificado y uno de 0.5 que el modelo no es diferente de clasificar y representa un mal
modelo (Mateo et al., 2011).
Los mapas generados con MaxEnt, indican que la probabilidad de la presencia de la
zorra gris para el área de estudio está en un rango de 0.46-0.62 que es un valor
aceptable, dado que es una especie generalista con preferencia por los hábitats boscosos
(Fritzell y Haroldson, 1982).
Se validó el modelo del Desierto Sonorense con las tres bios más importantes (fig. 24) y
los valores de probabilidad para la presencia de la especie en el área de estudio
presentaron el mismo rango de probabilidad con un valor de AUC= 0.821 (fig.25).

54
Para las especies con hábitos similares de alimentación, la división del hábitat o el
nicho, así como las diferencias en los patrones de actividad puede facilitar la
convivencia (Solman et al., 2011). El nicho ecológico de la zorra gris descrito por sus
hábitos alimenticios mediante el análisis de excretas encontradas en el Cañón de Las
Barajitas y comparado con la literatura referente a su dieta, los patrones de actividad
estimados para la zorra gris y las especies simpátricas así como los modelos de
ocupación correlacionados con variables categóricas y cualitativas del hábitat indican
que esta especie dentro de un hábitat con poca perturbación encuentra los recursos que le
permiten subsistir y perpetuarse en un espacio potencialmente viable para su adecuación
exitosa y en equilibrio con su entorno, y quizás sería conveniente considerar que la
presencia de las especies omnívoras con hábitos generalistas son fundamentales para la
autorregulación de los sistemas naturales y por medio de la modelación de su
distribución se pueden ubicar sitios que cuenten con características similares y ser de
importancia para establecer, en el caso de requerirse, manejo del área, en base a las
dinámicas poblacionales de estas especies, dado los datos reales relacionados a la
presencia de la zorra gris, lo que permite inferir zonas potencialmente idóneas en
función de sus características ambientales.
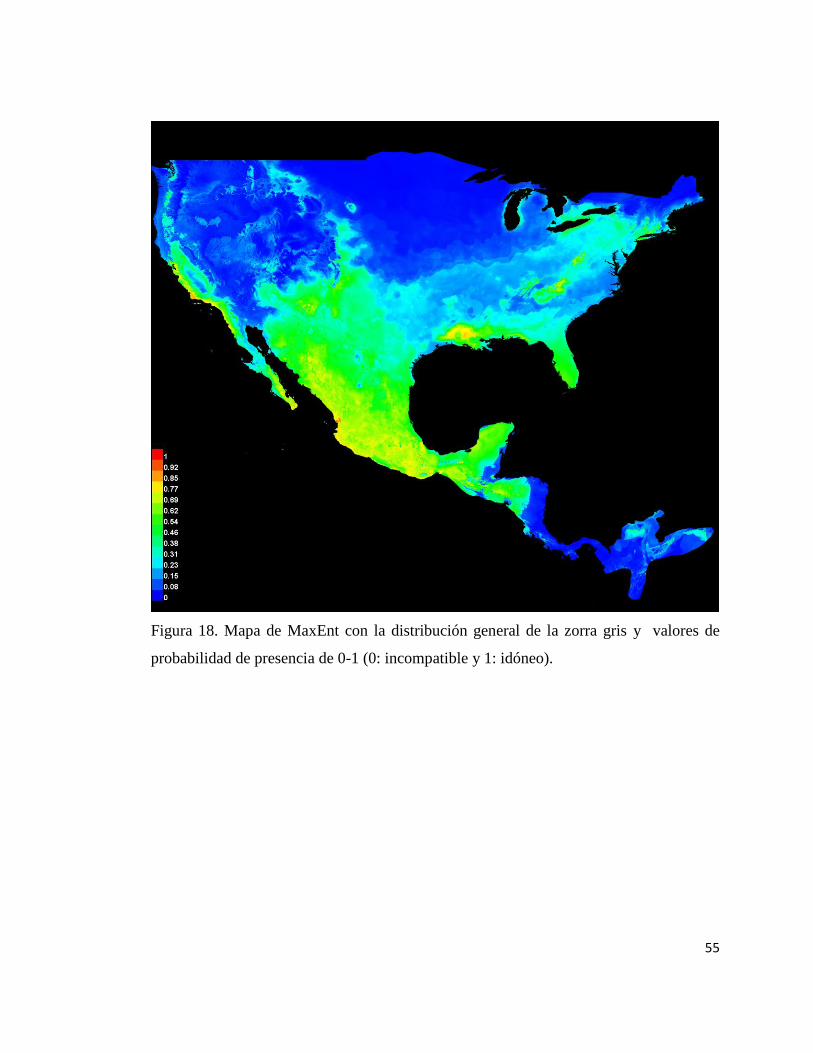
55
Figura 18. Mapa de MaxEnt con la distribución general de la zorra gris y valores de
probabilidad de presencia de 0-1 (0: incompatible y 1: idóneo).

56
Figura 19. Mapa de MaxEnt de la distribución potencial de la zorra gris en Sonora.

57
Figura 20. Mapa de MaxEnt de la distribución potencial de la zorra gris en el Desierto
Sonorense.

58
Figura 21. Gráfica del modelo general considerado aceptable, es decir la
distribución actual se explica moderadamente bien a partir de las variables
bioclimáticas (Felicísimo et al., 2012).

59
Figura 22. Gráfica del modelo del estado de Sonora, considerado modelo
aceptable, al igual que el de la distribución general (Felicísimo et al., 2012).

60
Figura 23. Gráfica del modelo del Desierto Sonorense, que indica que la
distribución actual se explica bien a partir de las variables bioclimáticas y se
considera un modelo bueno (Felicísimo et al., 2012).
.

61
Figura 24. Mapa de la validación del modelo para el Desierto Sonorense.

62
Figura 25. Gráfica de la validación del modelo del Desierto Sonorense que indica
que la distribución actual se explica bien a partir de las variables bioclimáticas y
se considera un modelo bueno (Felicísimo et al., 2012).

63
IX. CONCLUSIONES
Las tasas de captura están parcialmente asociadas a la abundancia, por lo que
pueden ser útiles para hacer estimaciones sobre ésta en el caso de las especies
que visitan con más frecuencia las estaciones de muestreo (foto-trampeo).
La permanencia de una especie omnívora que se encuentre en el centro de la
cadena trófica en un hábitat con poca perturbación, favorecerá las interacciones
positivas entre los componentes del ecosistema y lo mantendrán estable en
términos de la regulación de las especies herbívoras y -por lo mismo- permite
una mayor biodiversidad.
Dado que todas las cámaras trampa tienen la misma probabilidad de captura, se
puede suponer que los sitios con mayor frecuencia de aparición están ubicados
cerca de su centro de actividad, lo que puede indicar, tanto que la probabilidad de
detección será mayor en estos sitios, como la preferencia en el uso del hábitat.
Los modelos numéricos de los procesos naturales son importantes herramientas
para la conservación de las especies por que permiten generar líneas base para
comparar la sensibilidad de las especies a los cambios en los parámetros
biológicos que las mantienen con determinado estatus ecológico.
Estimar la ocupación de una especie a nivel de paisaje es un método factible de
monitoreo que, relacionado con el papel funcional de la especie, puede funcionar
como un indicador de cuál especie es más vulnerable y cual especie es más
persistente, en un tiempo y espacio determinado realizando estudios en el mismo
sitio a largo plazo ya que las reducciones poblacionales son ejemplos para
entender la respuesta adaptativa al cambio demográfico.

64
El modelo de ocupación de la zorra gris presupone poblaciones cerradas en los
sitios de muestreo donde no se presentan cambios demográficos o dispersión
hacia otras áreas.
Las variables estructurales de la composición florística del Cañón de Las
Barajitas varían significativamente a lo largo de un año, y se pueden considerar
dos estacionalidades fuertemente marcadas.
Ya que los resultados muestran que solo existe efecto de la estación del año para
la presencia de la zorra gris, se puede suponer que existe una complementariedad
entre el nicho ecológico de la zorra gris y de las especies simpátricas,
principalmente el coyote, al compartir el hábitat y presentar una frecuencia de
aparición similar a la de la zorra gris.

65
X. RECOMENDACIONES
La alta diversidad encontrada en el Cañón de las Barajitas sugiere que la toma de
decisiones acerca del uso de suelo en el Cañón de Las Barajitas debe enfocarse a
favor de la conservación.
Los modelos de acumulación de especies podrían ser útiles para fines de
conservación dado que se podrían formalizar inventarios y así se conocerá si
estos modelos pueden tener un valor predictivo al extrapolar y obtener los
valores asintóticos de las listas de especies acumuladas.
Para considerar a la zorra gris como un legítimo dispersor de semillas viables
para la germinación en el Cañón de Las Barajitas hace falta hacer un estudio
sobre el grado de germinación de las plantas que esta especie incluye en su dieta.
Es recomendable hacer estudios en hábitats fragmentados de la ocupación de la
zorra gris basados en los modelos de especialización ecológica (nicho).
Para conocer y cumplir los objetivos globales y regionales de la conservación de
la biodiversidad es necesario una mejor comprensión de los rangos de la
distribución de las especies, por lo que es necesario un estudio del ámbito
hogareño de la zorra gris en una región árida - tropical.
Para evaluar la demografía de una población y determinar si se trata de una
población fuente se sugiere que se detecte y monitoree a los juveniles,
particularmente machos de zorra gris ya que son los dispersores.

66
Son necesarios estudios de radio telemetría para determinar el ámbito hogareño,
junto con un estudio de la fenología floral, dada la relación que existe entre los
componentes florísticos y la fauna. Éstos proveerán datos más sólidos acerca de
la importancia y la plasticidad que presenta la zorra gris, lo que le ha permitido
permanecer, a pesar del acelerado crecimiento de infraestructura urbana, como
una especie que no está en peligro de extinción

67
XI. BIBLIOGRAFÍA
Akaike, H. (1974). A new look at the statistical model identification. Automatic Control,
IEEE Transactions on, 19(6), 716-723.
Akaike, H. (1987). Factor analysis and AIC. Psychometrika, 52(3), 317-332.
Alcaraz-Ariza, F.J. (2013). El método fitosociológico, Geobotánica, tema 11.
Universidad de Murcia, España. Acceso en:
http://www.um.es/docencia/geobotanica/ficheros/tema11.pdf
Aranda, M. (2000). Huellas y otros rastros de los mamíferos grandes y medianos de
México. CONABIO e INECOL A.C., Xalapa, Veracruz. México. 212 pp.
Arroyo-Cabrales, J., Medellín, R. A. y Domínguez-Castellanos (2005). Lista actualizada
de los mamíferos de México. Revista Mexicana de Mastozoología 9, 2171 pp.
Arnaud, G., y Acevedo, M. (1990). Hábitos alimenticios de la zorra gris, Urocyon
cinereoargenteus (Carnívora: Canidae) en la región meridional de Baja California,
México. Revista de Biología Tropical, 32, 499-502.
Ávila, V. y Lamberton, M. (2013). Wildlife Survey and Monitoring in the Sky Island
Region with an Emphasis on Neotropical Felids. USDA Forest Service Proceedings
RMRS-P-67.
Barajas Gea C.I. (2005). Evaluación de la Diversidad de la flora en el Campus Juriquilla
de la UNAM, Centro de Geociencias, Bol-e, Vol 1, No.2.
Benayas, J. M. R. (2009). La rareza de las especies. Investigación y ciencia, 63.

68
Buckley, H. L., y Freckleton, R. P. (2010). Understanding the role of species dynamics
in abundance–occupancy relationships. Journal of Ecology, 98(3), 645-658.
Braun-Blanquet J., Fuller, G., y Conrad, H. (1932), Plant sociology: the study of plant
communities. Mc Graw Hill Book Co. NewYork; London. 472 pp
Brown, J. H. (2003). Macroecolgía, Primera edición. Fondo de cultura económica,
México D.F. 397 pp.
Callaway, R. M. (1995). Positive interactions among plants. The Botanical Review,
61(4), 306-349.
Castillo-Gámez, R.A., J.P. Gallo-Reynoso, J.Egido-Villarreal y W. Caire. (2010).
Mamíferos. En: F.E. Molina-Freaner y T. R. Van Devender, eds. Diversidad Biológica
de Sonora. UNAM, México, pp.42-436.
Cervantes F., y Ballesteros, C. (2012). Estudios sobre la biología de roedores silvestres
mexicanos. UNAM-UAM, México
Cuarón, A. D. (2008). Effects of Land-Cover Changes on Mammals in a Neotropical
Region: a Modeling Approach. Conservation Biology, 14(6), 1676-1692.
Chamberlain, M. J., y Leopold, B. D. (2000). Spatial use patterns, seasonal habitat
selection, and interactions among adult gray foxes in Mississippi. The Journal of wildlife
management, 742-751.
Chávez, C., A de la Torre, H. Bárcenas, R.A. Medellín, H. Zarza y G. Ceballos. (2013).
Manual de fototrampeo para estudio de fauna silvestre. El jaguar en México como
estudio de caso. Alianza WWF-Telcel, Universidad Nacional Autónoma de México,
México.

69
Dell’Arte, G. L., y Leonardi, G. (2005). Effects of habitat composition on the use of
resources by the red fox in a semi-arid environment of North Africa. Acta Oecologica,
28(2), 77-85.
Felicísimo, A. M., Muñoz, J., Mateo, R. G., y Villalba, C. J. (2012). Vulnerabilidad
de la flora y vegetación españolas ante el cambio climático. Revista Ecosistemas, 21(3),
1-6.
Felger S. Richard (1999). The Flora of Cañón de Nacapule: A Desert Bounded Tropical
Canyon near Guaymas, Sonora. México. Proceedings of the San Diego Society of
Natural History No. 35.
Felger, R. S., Wilder, B. T., y Reynoso, J. P. G. (2011). Floristic diversity and long-term
vegetation dynamics of San Pedro Nolasco Island, Gulf of California, Mexico. San
Diego Society of Natural History.
Fonturbel, F. y Jiménez, J., (2011). Conectando la ecología de una especie con la
conservación de ecosistemas: el caso del monito de monte (Dromiciops gliroides).
Revista Chagual 9:61-64, 2011
Fordham, D. A., Akcakaya, H. R., Araújo, M. B., Keith, D. A., y Brook, B. W. (2013).
Tools for integrating range change, extinction risk and climate change information into
conservation management. Ecography, 36(9), 956-964.
Fleming, T. H. (1977). Response of two species of tropical Heteromyid rodents to
reduced food and water availability. Journal of Mammalogy, 102-106.
Fritzell, E. K. y K. J. Haroldson. (1982). Urocyon cinereoargenteus. Mammalian
species 189:1-6

70
González, A. (2008) Distribución de la zorra del desierto Vulpes macrotis en relación a
sus presas y a su principal depredador, el coyote Canis latrans, Instituto de Ecología,
A.C
Guerrero, S., Badii, M. H., Zalapa, S. S., y Flores, A. E. (2002). Dieta y nicho de
alimentación del coyote, zorra gris, mapache y jaguarondi en un bosque tropical
caducifolio de la costa sur del estado de Jalisco, México. Acta Zoológica Mexicana, 86,
119-137.
Harrison, R. L. (1997). A comparison of gray fox ecology between residential and
undeveloped rural landscapes. The Journal of wildlife management, 112-122.
Hui, C., Boonzaaier, C., y Boyero, L. (2012). Estimating changes in species abundance
from occupancy and aggregation. Basic and Applied Ecology, 13(2), 169-177.
Klimova A, Munguia-Vega A., Hoffman J., y Culver M. (2014) Genetic diversity and
demography of two endangered captive pronghorn subspecies from the Sonoran Desert.
Journal of Mammalogy In-Press
Listado de regiones terrestres prioritarias de la Comisión Nacional para uso de la
Biodiversidad (2000) acceso en:
http://www.conabio.gob.mx/conocimiento/regionalizacion/doctos/rtp_018.pdf
León, J. L., Benet, L. R. C., y Estrada, M. C. (1996). Fenología floral de una comunidad
árido-tropical de Baja California Sur, México. Acta Botánica Mexicana, 35, 45-64.
López, B. A., y Tonatiuh, C. (2011). Estructura de comunidades de mamíferos
carnívoros bajo influencia de manejo forestal en Santa Catarina Ixtepeji, Oaxaca (Tesis
Doctoral).

71
Llorente B, J. (1993). The use of species accumulation functions for the prediction of
species richness. Conservation biology, 7(3), 480-488.
McAuliffe, J. R. (1986). Herbivore-Limited Establishment of a Sonoran Desert Tree,
Cercidium Microphyllum. Ecology, 276-280.
MacKenzie, D. I., y Nichols, J. D. (2004). Occupancy as a surrogate for abundance
estimation. Animal biodiversity and conservation, 27(1), 461-467.
Martinez-Yrízar, A.; Felger, R. S. y Búrquez, A. (2009). Los ecosistemas terrestres: un
diverso capital natural. En: Molina-Freaner, F. E. y Van Devender, T. R. (Eds.), La
diversidad biológica de Sonora. UNAM. México. Pp 129-156
Martínez, N. (2010). Apuntes sobre Modelación de Nichos Ecológicos. Laboratorio de
Evolución Molecular y Experimental, del Instituto de Ecología de la UNAM-México.
Monroy-Vilchis, O., Rodríguez-Soto, C., Zarco-González, M., y Urios, V. (2009).
Cougar and jaguar habitat use and activity patterns in central Mexico. Animal Biology,
59(2), 145-157.
Moreno, C. E. (2001). Manual de métodos para medir la biodiversidad. Universidad
Veracruzana.
Acceso en: http://entomología.rediris.es/sea/manytes/métodos.pdf
Munson, S. M., Webb, R. H., y Hubbard, J. A. (2011). A comparison of methods to
assess long-term changes in Sonoran Desert vegetation. Journal of Arid Environments,
75(11), 1228-1231.

72
Noon, B. R., Bailey, L. L., Sisk, T. D., y McKelvey, K. S. (2012). Efficient
Species‐Level Monitoring at the Landscape Scale. Conservation Biology, 26(3), 432-
441.
Odum, E. (2003), Ecología: el vínculo entre las ciencias naturales y las sociales,
Vigésima tercera edición. Compañía editorial continental. México, D. F. 295 pp.
Peinado, M., Macías, M. Á., Ocaña-Peinado, F. M., Aguirre, J. L., y Delgadillo, J.
(2011). Bioclimates and vegetation along the Pacific basin of Northwestern Mexico.
Plant ecology, 212(2), 263-281.
Peterson, A. T., Papes, M., y Soberón, J. (2008). Rethinking receiver operating
characteristic analysis applications in ecological niche modeling. Ecological Modelling,
213(1), 63-72.
Pliscoff, P y Fuentes-Castillo T. (2011). Modelación de la distribución de especies y
ecosistemas en el tiempo y en el espacio y una revisión de las nuevas herramientas y
enfoques disponibles. Revista de Geografía Norte Grande, 48:61-79.
Raxworthy, C. J., Ingram, C. M., Rabibisoa, N., y Pearson, R. G. (2007). Applications
of ecological niche modeling for species delimitation: a review and empirical evaluation
using day geckos (Phelsuma) from Madagascar. Systematic biology, 56(6), 907-923.
Rivera-Rivera, Emerenciano, Enríquez, Paula L., Flamenco-Sandoval, Alejandro, y
Rangel-Salazar, José Luis. (2012). Occupancy and abundance of nocturnal raptors
(Strigidae) in the Selva El Ocote Biosphere Reserve, Chiapas, Mexico. Revista
mexicana de biodiversidad, 83(3), 742-752
Robichaux, R. H. (1999). Ecology of Sonoran Desert plants and plant communities (pp.
36-67). Tucson, USA: The University of Arizona Press.

73
Rzedowski, J. (1991). "Diversidad y orígenes de la flora fanerogámica de México". Acta
Botánica Mexicana 14: 3-21.
Sollmann, R., Furtado, M. M., Hofer, H., Jácomo, A. T., Tôrres, N. M., y Silveira, L.
(2012). Using occupancy models to investigate space partitioning between two
sympatric large predators, the jaguar and puma in central Brazil.Mammalian Biology-
Zeitschrift für Säugetierkunde, 77(1), 41-46.
Sollmann, R., Mohamed, A., Samejima, H., y Wilting, A. (2013). Risky business or
simple solution–Relative abundance indices from camera-trapping. Biological
Conservation, 159, 405-412.
The IUCN Red List of Threatened Species, acceso en:
http://www.iucnredlist.org/details/22780/0
Tewksbury, J. J., y Lloyd, J. D. (2001). Positive interactions under nurse-plants: spatial
scale, stress gradients and benefactor size. Oecologia, 127(3), 425-434.
Treves, A. y U. Karanth. (2003). Human-carnivore conflict and perspectives on
carnivore management worldwide. Conservation Biology 17:1491-1499.
Valenzuela, D., y Ceballos, G. (2000). Habitat selection, home range, and activity of the
white-nosed coati (Nassua narica) in a Mexican tropical dry forest. Journal of
mammalogy, 81(3), 810-819.
Velázquez, A., Mas, J. F., Díaz Gallegos, J. R., Mayorga-Saucedo, R., Alcántara, P. C.,
Castro, R. y Palacio, J. L., (2002). Patrones y tasas de cambio de uso del suelo en
México. Gaceta ecológica, (62), 21-37.

74
Villaseñor, J. L. (2004). Los géneros de plantas vasculares de la flora de México. Boletín
de la Sociedad Botánica de México, 75, 105-135.
White, G. C., Burnham, K. P., y Anderson, D. R. (2001, June). Advanced features of
program MARK. In Wildlife, land, and people: priorities for the 21st century.
Proceedings of the second international wildlife management congress. The Wildlife
Society, Bethesda, Maryland, USA (pp. 368-377).

75
XII. APÉNDICES

76
Apéndice 1. Listado florístico de la vegetación muestreada.
Especie Familia
Stenocereus thurberi Cactaceae
Jatropha cuneata Euphorbiaceae
Cylindropuntia fulgida var fulgida Cactaceae
Guaiacum coulteri Zygophyllaceae
Simmondsia chinensis Simmondsiaceae
Olneya tesota Fabaceae
Ruellia californica Acanthaceae
Mimosa distachya var. laxiflora Fabaceae
Jatropha cordata Euphorbiaceae
Bursera microphylla Burseraceae
Jaquinia macrocarpa pungens Theophrastaceae
Fouquieria diguetii Fouqueriaceae
Ferocactus emoryi Cactaceae
Parkinsonia microphyllum Fabaceae
Lippia palmeri Verbenaceae
Acacia willardiana Fabaceae
Haematoxylon brasiletto Fabaceae
Colubrina californica Rhamnaceae
Encelia farinosa Asteraceae
Forchhameria watsonii Capparaceae
Solanum hindsianum Solanaceae
Sebastiana bilocularis Euphorbiaceae
Colubrina viridis Euphorbiaceae
Justicia californica Rhamnaceae
Opuntia bigelovi Cactaceae
Agave chrysoglossa Agavaceae
Allonia incarnata Nyctaginaceae
Bacharis saratroides Asteraceae
Tephrosia vicioides Asteraceae
Washingtonia robusta Palmae
Ficus insipida Moraceae
Passiflora arida Passifloraceae
Lysiloma watsonii Fabaceae
Coursetia glandulosa Fabaceae
Lycium Solanaceae
Senna covesii Fabaceae

77
Apéndice 1 (Continuación).
n=59
Especie Familia
Pachycereus pringlei Cactaceae
Krameria sp. Krameriaceae
Bursera fagaroides Burseraceae
Maythenus phyllantoides Celastraceae
Atriplex barclayana Chenopodiaceae
Echinocereus scopolorum Cactaceae
Carnegia giganteae Cactaceae
Macroptilium atrupurpureum Fabaceae
Gossypium davidsoni Malvaceae
Hibiscus denudatus Malavaceae
Hofmeisteria crassifolia Asteraceae
Bebbia juncea Asteraceae
Stegnosperma halimofolium Phytolaccaceae
Randia sonorensis Rubiaceae
Sideroxylon occidentale Sapotaceae
Marsdenia edulis Asclepiadaceae
Prospois glandulosa var. Torreyana Fabaceae
Ziziphus amole Rhamnaceae
Sapindus saponaria Sapindaceae
Phaulothamnus spinescens Phytolaccaceae
Ziziphus obtusifolia Rhamnaceae

78
Apéndice 2. Cambios estacionales en la vegetación al Noroeste del Cañón de Las
Barajitas
Primavera

79
Verano

80
Otoño

81
Invierno





























