Embed Size (px)
Citation preview

1 58 Neuros¢'ience Research, 8 t 1990) 15~ 175 Elsevier Scientific Publishers Ireland i t d
NEURES 00343
Cerebellar corticonuclear and nucleocortical projections in the vermis of posterior lobe of the rat as studied with
anterograde and retrograde transport of WGA-HRP
Juha P~iiillysaho, Shoei Sugita and Hiroharu Noda Department of Visual Sciences, School of Optometry, Indiana University, Bloomington, IN (U.S.A.)
(Received 10 November 1989; Revised version received 18 January 1990; Accepted 13 February 1990)
Key words: Cerebellar corticonuclear projection; Cerebellar nucleocortical projection; Posterior cerebellar vermis; Horseradish peroxidase; Rats
SUMMARY
Corticonuclear (CN) and nucleocortical (NC) projections of the cerebellum were studied in rats by using antero- and retrograde transport of wheatgerm agglutinin-conjugated horseradish peroxidase (WGA-HRP). Following minute injections of WGA-HRP in different folia for vermal lobules VI-VIII. a rostrocaudal topography was observed in the CN projection; lobules Via and VIb. c projected to distinctly separate areas in the ipsilateral fastigial nucleus (FN): Via projected to the ventral part of the medial subdivision of the FN (FNm), whereas VIb, c projected to the dorsocaudal part of the caudomedial subdivision of the FN (FNcm) and to the caudal FNm. The terminals from lobules VIb, c and VII occupied partially overlapping areas m the caudal part of the ipsilateral FNcm and FNm. A few terminal fibers were also seen in the dorsolateral protuberance of the FN (dip). The fibers from vermal lobule VIII terminated in the ipsilateral rostral FNem and in the FNm. The latter terminal area was divided into two parts: a smaller dorsal and a larger ventral part that in turn overlapped almost completely with the terminal area of fibers from lobute Via. Retrogradely labeled fastigial neurons were found in the areas of the FN where Purkinje-cell (P-cell) axons from the respective injection sites terminated. In addition, retrogradely labeled fastigial neurons were scattered in areas outside the P-cell terminal fields on the injection side and in the contralateral FN.
INTRODUCTION
The afferent and efferent connections of the cerebellum have been the subject of a number of anatomical and physiological investigations. Although the topographical organization of the cerebellar connections has been studied extensively in the cat (see Ito 23 for review), our knowledge of similar cortical topography in other mammals is less extensive.
Despite a considerable large volume of work on the rat corticonuclear (CN) projec- tions 1,4,5,12.18,41-43, there are only a few reports t,t8 which have dealt exclusively with the rostrocaudal topography of CN projections involving the vermis of the posterior lobe. These studies have indicated that there is a great similarity between the cat and the rat in
Correspondence: Dr. Hiroharu Noda, Department of Visual Sciences, School of Optometry, Indiana University, 800 E. Atwater, Bloomington, IN 47405, U.S.A.
0168-0102/90/$03.50 © 1990 Elsevier Scientific Publishers Ireland Ltd.

159
the longitudinal zonal structure of the cerebellar cortex. However, they have not provided a comprehensive survey of how the CN projection from different lobules is organized within one cortical zone.
Because minute amounts of horseradish peroxidase (HRP) were used as an axonal tracer in the present study, the climbing- and mossy-fiber connections and the P-cell projections could be simultaneously investigated following a single injection. This enabled us to investigate an entire 'module' of afferent and efferent connections of each lobule of the cerebellar cortex.
In a preceding paper 33 we presented the topographical organization of the olivocere- bellar fibers to vermal lobules VI-VIII . In this paper we describe the rostrocaudal topographical organization of CN, nucleocortical (NC) and corticovestibular fibers of lobules Via, VIb, c, VII and VIII.
MATERIALS AND METHODS
Thirty female Sprague-Dawley strain albino rats weighing between 150 and 300 g were used. In order to study the morphology of the deep cerebellar nuclear complex of the rat, the brain of one rat was sectioned in the coronal plane and that of another rat in the parasagittal plane, and sections were stained with cresyl violet.
The remaining 28 rats were anesthetized by intraperitoneal injection of sodium amobarbital (60 mg/kg) and placed in a stereotaxic head holder. A trephine hole was made in the skull and the dura matter was exposed. The midline was determined as the mid-distance between the lateral boundaries of the posterior vermis.
Each animal received a single HRP injection into the cerebellar cortex either under visual guidance or by using the stereotaxic coordinates derived from previous experi- ments. All the rats were injected with 10% wheatgerm agglutinin-conjugated HRP (WGA-HRP) and 10% HRP (Sigma type VI) in 0.005-0.02 #1 Ringer solution in the cerebellar cortex through a glass micropipette (20 #g tip diameter) attached to a 1 #1 Hamilton microsyringe. The micropipette was placed in each lobule 1.0-1.5 mm under the cortical surface. After injection, the pipette was left in situ for 15 min to prevent the enzyme from leaking through the track of the retracted cannula.
After 24-28 h, the animals were re-anesthetized and perfused with 500 ml warm (36-37 ° C) Ringer solution, with 1% paraformaldehyde and 1.25% glutaraldehyde in 0.1 M phosphate buffer (pH 7.4), followed by 10% sucrose in the same buffer. The brains were removed and placed in phosphate buffer with 30% sucrose and stored at 4 ° C.
Frozen sections (40/~m thick) of the brains were cut either in the parasagittal or in the coronal plane and treated for HRP histochemical reaction with tetramethylbenzidine (TMB), following the method described by Mesulam 3o. The sections were then counter- stained with neutral red.
In j ec t i on s i tes
The injection sites were determined by reconstructing the parasagittal or the coronal sections of the brains. The extent of injection site was evaluated in TMB-reacted sections. HRP-labeled P-cells were mapped on camera-lucida drawings. All P-cells in the close vicinity of the injection were labeled, although the density of labeling decreased with the distance from the center of injection. When the white matter near the center of injection was examined, labeled axons were seen en m a s s e (see Fig. 4). However, there was no labeling in the white matter beneath the lightly labeled P-cells at a distance from the center of injection. Hence, we only reconstructed the dense central core impregnated with

160
the enzyme close to the tip of the micropipette. The injection sites are shown in Figure I and in Table 1, where the effective injection site is slightly overestimated to include both antero- and retrograde labeling.
Anterograde labeling In each experiment, anterogradely labeled P-cell axon terminals were carefully mapped
onto large-size camera-lucida drawings of the deep cerebellar nuclei and the Deiters' nucleus. The results were schematically shown in a drawing of 6 parasagittal sections from the medial fastigial nucleus (FN) to the medial portion of the interposed nuclei (IN) (see, e.g., Fig. 3). We incorporated the description of Korneliussen 25 for the anatomical subdivisions of the cerebellar nuclei in the rat.
A quantitative study was attempted for each experiment, in order to evaluate the extent of HRP spread and the efferent projections from posterior vermal lobules. The number of anterogradely labeled terminals of P-cells was estimated in each deep cerebel- lar nuclei and Deiters' nucleus. A unit area of 70/ t in 2 was determined by using a scale in the light microscope and the number of unit areas in each nucleus was counted. The density of terminals in each unit area was weighted by a factor of 3, 2 and 1, corresponding to numerous, many and a few labeled terminals, respectively. The number of these weighted terminal areas was then counted for each nucleus and expressed as relative percentages of the total number of terminals (see Table I).
Retrograde labeling Retrogradely labeled cells were located in the FN and the posterior IN, and their
location was reported in the same schematic drawings as the anterogradely labeled P-cell axons (see, e.g., Fig. 3). Only cell bodies with a complete nucleus were mapped and counted. The results of the quantitative study of labeled cells are listed in Table II.
RESULTS
HRP injections into vermes of 28 rats resulted in labeling of neurons as well as terminals in the deep cerebellar and vestibular nuclei in 16 cases. The failure of axonal transport of HRP in 12 cases was possibly caused by disturbance of the local circulation associated with injection, or by the leakage of the enzyme into the subarachnoidal space.
In all successful cases, anterogradely labeled P-cell axons could be traced in the folial white matter along the entire course from the injection site to the ipsilateral FN, where the fibers in most cases formed a dense terminal plexus (see, e.g., Fig. 4). A plexus of thin and varicose fibers found within localized parts of the FN was considered as a terminal area. In some cases anterogradely stained axons vanished as preterminal fibers. Thicker fibers which branched inside or just outside the nuclei were considered as preterminal fibers. When HRP encroached upon the lateral border of the posterior vermis, plexi were observed also in the posterior IN and vestibular complex. Retrogradely labeled NC neurons were present in the same areas and were usually located among anterogradely labeled terminal fibers.
In the presentation given below the animals are divided into 4 groups: those with injection in (1) vermal lobule Via, (2) verrnal folia VIb, c, (3) vermal tobule VII, and (4) vermal lobule VIII. Because of the anatomical differences between the anterior and posterior halves of lobule VI 33, the lobule VI cases were separated into two different groups.
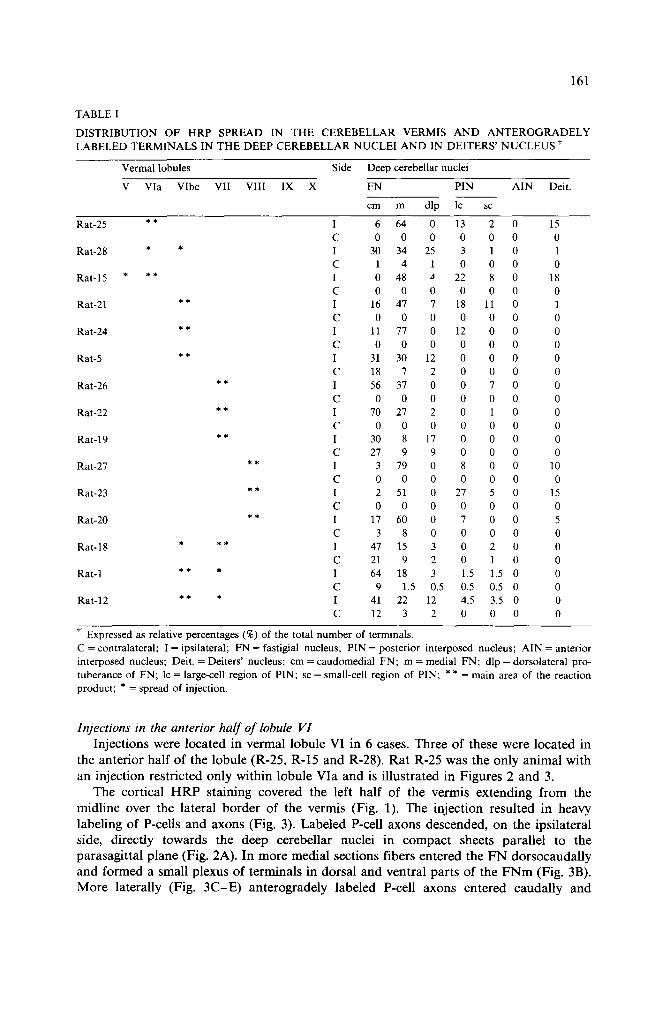
161
TABLE I
DISTRIBUTION OF HRP SPREAD IN THE CEREBELLAR VERMIS A N D ANTEROGRADELY LABELED TERMINALS IN THE DEEP CEREBELLAR NUCLEI AND IN DEITERS' NUCLEUS f
Vermal lobules Side Deep cerebellar nuclei
V Via Vlbc VII VIII IX X FN PIN AIN Deit.
cm m dip lc sc
Rat-25 * * I 6 64 0 13 2 0 15 C 0 0 0 0 0 0 0
Rat-28 * * I 30 34 25 3 1 0 1 C 1 4 1 0 0 0 0
Rat-I 5 * * * I 0 48 ,J 22 8 0 18 C 0 0 0 0 0 0 0
Rat-21 * * I 16 47 7 18 11 0 1 C 0 0 0 0 0 0 0
Rat-24 * * I 11 77 0 12 0 0 0 C 0 0 0 0 0 0 0
Rat-5 * * I 31 30 12 0 0 0 0 C 18 7 2 0 0 0 0
Rat-26 * * I 56 37 0 0 7 0 0 C 0 0 0 0 0 0 0
Rat-22 * * I 70 27 2 0 1 0 0 C 0 0 0 0 0 0 0
Rat-19 * * I 30 8 17 0 0 0 0 C 27 9 9 0 0 0 0
Rat-27 * * I 3 79 0 8 0 0 10 C 0 0 0 0 0 0 0
Rat-23 * * I 2 51 0 27 5 0 15 C 0 0 0 0 0 0 0
Rat-20 * * I 17 60 0 7 0 0 5 C 3 8 0 0 0 0 0
Rat-18 * ** I 47 15 3 0 2 0 0 C 21 9 2 0 1 0 0
Rat-1 * * * I 64 18 3 1.5 1.5 0 0 C 9 1.5 0.5 0.5 0.5 0 0
Rat-12 * * * I 41 22 12 4.5 3.5 0 0 C 12 3 2 0 0 0 0
Expressed as relative percentages (%) of the total number of terminals. C = contralateral; I = ipsilateral; FN = fastigial nucleus; PIN = posterior interposed nucleus; AIN = anterior interposed nucleus; Deit. = Deiters' nucleus; cm = caudomedial FN; m = medial FN; dip = dorsolateral pro- tuberance of FN; lc = large-cell region of PIN; sc = small-cell region of PIN; ** = main area of the reaction product; * = spread of injection.
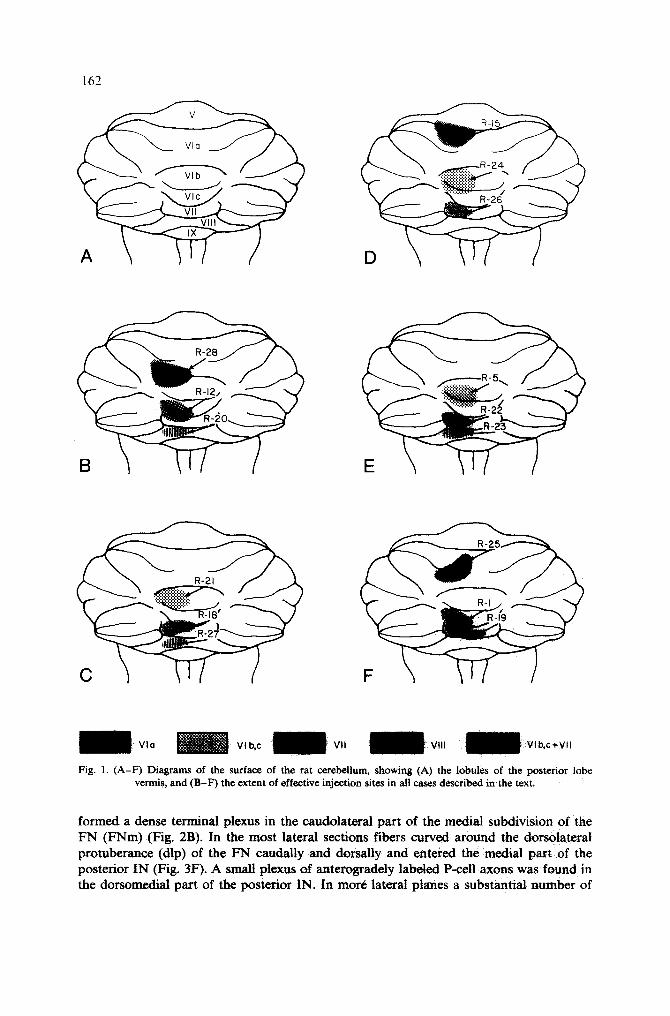
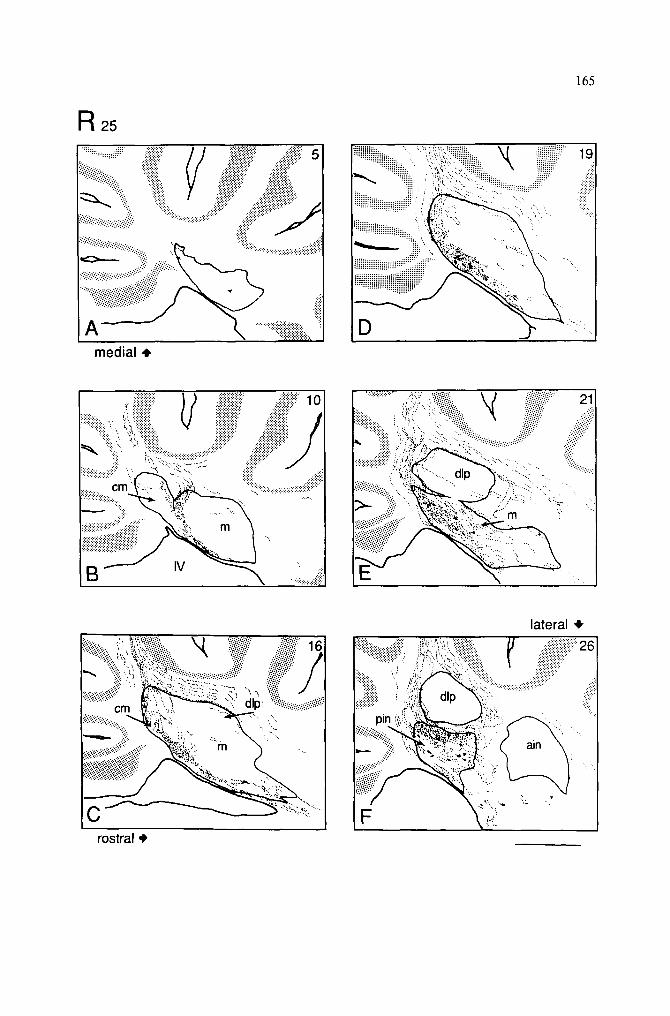
Injections in the anterior half of lobule 111 I n j e c t i o n s w e r e l o c a t e d in v e r m a l l o b u l e V I i n 6 c a s e s . T h r e e o f t h e s e w e r e l o c a t e d in
t h e a n t e r i o r h a l f o f t h e l o b u l e ( R - 2 5 , R - 1 5 a n d R - 2 8 ) . R a t R - 2 5 w a s t h e o n l y a n i m a l w i t h
a n i n j e c t i o n r e s t r i c t e d o n l y w i t h i n l o b u l e V i a a n d is i l l u s t r a t e d in F i g u r e s 2 a n d 3.
T h e c o r t i c a l H R P s t a i n i n g c o v e r e d t h e l e f t h a l f o f t h e v e r m i s e x t e n d i n g f r o m t h e
m i d l i n e o v e r t h e l a t e r a l b o r d e r o f t h e v e r m i s ( F i g . 1). T h e i n j e c t i o n r e s u l t e d i n h e a v y
l a b e l i n g o f P - c e l l s a n d a x o n s ( F i g . 3). L a b e l e d P - c e l l a x o n s d e s c e n d e d , o n t h e i p s i l a t e r a l
s i d e , d i r e c t l y t o w a r d s t h e d e e p c e r e b e l l a r n u c l e i i n c o m p a c t s h e e t s p a r a l l e l t o t h e
p a r a s a g i t t a l p l a n e ( F i g . 2 A ) . I n m o r e m e d i a l s e c t i o n s f i b e r s e n t e r e d t h e F N d o r s o c a u d a l l y
a n d f o r m e d a s m a l l p l e x u s o f t e r m i n a l s i n d o r s a l a n d v e n t r a l p a r t s o f t h e F N m ( F i g . 3B) .
M o r e l a t e r a l l y ( F i g . 3 C - E ) a n t e r o g r a d e l y l a b e l e d P - c e l l a x o n s e n t e r e d c a u d a l l y a n d
162
R-I ,,
Vlo ~ Vlb,c c+Vlt
Fig. 1. (A-F) Diagrams of the surface of the rat cerebellum, showing (A) the lobules of the posterior lobe vermis, and (B-F) the extent of effective injection sites in all cases described in the text,
formed a dense terminal plexus in the caudotateral part of the medial subdivision of the FN (FNm) (Fig. 2B). In the most lateral sections fibers curved around the dorsolateral protuberance (dlp) of the FN caudally and dorsally and entered the medial part of the posterior IN (Fig. 3F). A small plexus of anterogradely labeled P-cell axons was found in the dorsomedial part of the posterior IN. In more lateral planes a substantial number of
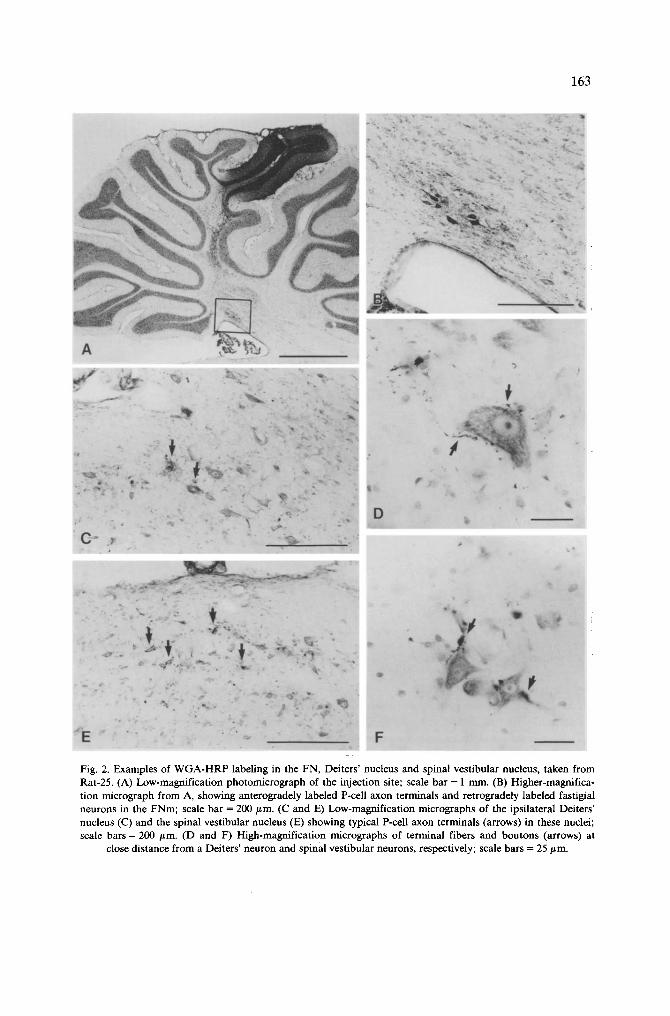
163
C ~ ;t~
f j" A1 . -
% F
I ~e
Fig. 2. Examples of WGA-HRP labeling in the FN, Deiters' nucleus and spinal vestibular nucleus, taken from Rat-25. (A) Low-magnification photomicrograph of the injection site; scale bar = 1 nun. (B) Higher-magnifica- tion micrograph from A, showing anterogradely labeled P-cell axon terminals and retrogradely labeled fastigial neurons in the FNm; scale bar = 200/~m. (C and E) Low-magnification micrographs of the ipsilateral Deiters' nucleus (C) and the spinal vestibular nucleus (E) showing typical P-cell axon terminals (arrows) in these nuclei; scale bars = 200 #m. (D and F) High-magnification micrographs of terminal fibers and boutons (arrows) at
close distance from a Deiters' neuron and spinal vestibular neurons, respectively; scale bars = 25 #m.
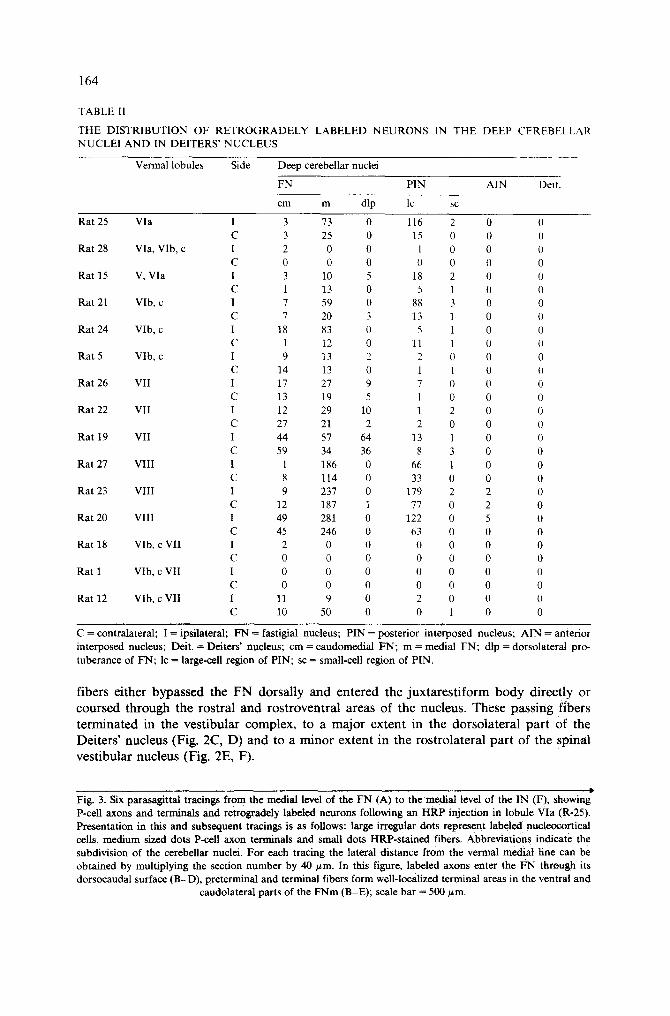
1 6 4
TABLE I1
THE DISTRIBUTION OF R E T R O G R A D E L Y LABELED N E U R O N S IN THE DEEP CEREBELLAR NUCLEI A N D IN DEITERS' N U C L E U S
Vermal lobules Side Deep cerebellar nuclei
FN PIN AIN Deit.
cm m dip lc sc
Rat 25 Via I 3 73 0 116 2 0 0 C 3 25 0 15 0 0 0
Rat 28 Via, VIb, c l 2 0 0 1 0 0 0 C 0 0 0 0 0 0 0
Rat 15 V, Via I 3 10 5 18 2 0 0 C 1 13 0 5 1 0 0
Rat 21 VIb, c 1 7 59 0 88 3 0 0 C 7 20 3 13 1 0 0
Rat 24 VIb, c I 18 83 0 5 1 0 0 C 1 12 0 11 1 0 0
Rat 5 Vlb, c I 9 13 2 2 0 0 0 C 14 13 0 1 1 0 0
Rat 26 VII I 17 27 9 7 0 0 0 C 13 19 5 1 0 0 0
Rat 22 VII I 12 29 10 1 2 0 0 C 27 21 2 2 0 0 0
Rat 19 VII I 44 57 64 13 1 0 0 C 59 34 36 8 3 0 0
Rat 27 VIII I 1 186 0 66 1 0 0 C 8 114 0 33 0 0 0
Rat 23 VIII I 9 237 0 179 2 2 0 C 12 187 1 77 0 2 0
Rat 20 VIII I 49 281 0 122 0 5 0 C 45 246 0 63 0 0 0
Rat 18 VIb, c VII I 2 0 0 0 0 0 0 C 0 0 0 0 0 0 0
Rat 1 VIb, c VII I 0 0 0 0 0 0 0 C 0 0 0 0 0 0 0
Rat 12 VIb, c VII I 11 9 0 2 0 0 0 C 10 50 0 0 1 0 0
C = contralateral; I = ipsilateral; FN = fastigial nucleus; PIN = posterior interposed nucleus; AIN = anterior interposed nucleus; Deit. -- Deiters' nucleus; cm = caudomedial FN; m = medial FN; dip = dorsolateral pro- tuberance of FN; lc = large-cell region of PIN; sc = small-cell region of PIN.
f i b e r s e i t h e r b y p a s s e d t h e F N d o r s a l l y a n d e n t e r e d t h e j u x t a r e s t i f o r m b o d y d i r e c t l y o r
c o u r s e d t h r o u g h t h e r o s t r a l a n d r o s t r o v e n t r a l a r e a s o f t h e n u c l e u s . T h e s e p a s s i n g f i b e r s
t e r m i n a t e d i n t h e v e s t i b u l a r c o m p l e x , t o a m a j o r e x t e n t i n t h e d o r s o l a t e r a l p a r t o f t h e
D e i t e r s ' n u c l e u s ( F i g . 2 C , D ) a n d t o a m i n o r e x t e n t i n t h e r o s t r o l a t e r a l p a r t o f t h e s p i n a l
v e s t i b u l a r n u c l e u s ( F i g . 2 E , F ) .
Fig. 3. Six parasagittal tracings from the medial level of the FN (A) to the medial level of the IN (F), showing P-cell axons and terminals and retrogradely labeled neurons following an HRP injection in lobule Via (R-25). Presentation in this and subsequent tracings is as follows: large irregular dots represent labeled nucleoeortical cells, medium sized dots P-cell axon terminals and small dots HRP-sta ined fibers. Abbreviat ions indicate the subdivision of the cerebellar nuclei. For each tracing the lateral distance from the vermal medial line can be obtained by multiplying the section number by 40 t~m. In this figure, labeled axons enter the I~N through its dorsocaudal surface (B-D) , preterminal and terminal fibers form welMocalized terminal areas in the ventral and
caudolateral parts of the F N m (B-E); scale bar = 500/~m.
R ~
m e d i a l 4,
9
165
".x::::::r:.:~':" ..",( ~,.' ~'~.'~ ..;{.~.'~k!~-~i-k~k~
:"37'.. L~:~:~,, ~'~:~" !.~,~. : . . . . p... - ~ - y . ~ ! ~ . ~ '
....... i~::~i~:: qh,.!'! ...:~.~. : :'~k-,. ... ~'~" ii::::i~ )"i.-~k!'~J""?" "~:!:~..,""~"."~""~''
. : ! i i ? : : : . ' '~ ."." . ' " ' i.'.'.'.'.'.:. :~ .:i!.?.~"~" "
' M ' . " ' I
~, ~?~;~ ...~.:.:...~ • .~, : . '~,~ C:~:~..'.'~.;~.
.. ~k~ ~" , ' "~.E4.'.,~ ~:('(:.:'.";"' .<:4.~'" "v.. : ~ 9 % ~ . . '
• "iiiii:
m
..(~:. ~,~!~::. :' .:~. :~ ...
r o s t r a l 4,
l a t e r a l ~,
%ii;j,~ ,~:~u.~,i~.i:~:~,~ii~iii~,. ~ iii~i~iiii;;i '~ 2c ~:~,~,,,,,,~ ~i ,~i~i '{ilfl ) .................. ,~,,,,~,~J~'~
i:i:!:8: i ~i, . " i~iiiiiii~i; . . . . ,)i~ ~ ~. ~, ~.
"i I 'i:.~ . . . . '
I .... i' ....,;!. 'i , . . . . ":",., .~:5'" ' ~ . i~
F

166
Several retrogradely labeled NC neurons were located ipsilaterally among orthodromi- cally labeled terminal fibers (Fig. 2B) or outside of the terminal plexus in the FN and in the medial part of the posterior IN (Fig. 3F). Most of the HRP-positive neurons in the FN were of fusiform small size or of multipolar medium size (Fig. 2B). Those neurons found in the posterior IN were medium- to large-size mono- and multipolar cells. Retrogradely labeled cells were observed also in the contralateral FN and posterior IN (Table II). Few clusters of neurons were seen in areas of the FN corresponding to those seen on the ipsilateral side. Also, few scattered neurons were observed in the medial end of the posterior IN, closely adjoining the FN. HRP-labeled neurons were observed very rarely in the lateral cerebellar nucleus (LN). When these single neurons were seen, they were very lightly labeled.
Injections in 2 other cases (R-28 and R-15) (Fig. 1) covered most of the left half of the vermis in lobule Via, but additional HRP labeling was observed in the rostral folia of VIb and in the most caudal folia of lobule V, respectively. In both these cases the distribution of CN fibers resembled partly that in the former animal. However, an additional terminal plexus was formed in the caudal and in the dorsal parts of the FNm, respectively, reflecting spread of HRP into the adjacent vermal folia of lobule Via.
Injections in the posterior half of lobule VI HRP was injected in the vermal folia VIb, c of 3 rats (R-5, R-21 and R-24). In 3 other
animals, HRP was deposited in the most caudal folia of lobule VIc and the anterior folia of lobule VII; these observed as controls (R-18, R-1 and R-12). The organization of CN fibers of lobule VIb, c is illustrated with Rat 21, in which HRP was placed in the left half of the vermal folia VIb, c (Fig. 4).
HRP covered most of the left vermis extending over the lateral border of the vermis, but did not cross the midline (Fig. 1). The most medial part of the ipsilateral vermis was involved in the injection, but part of the FN, close to the midline, was still devoid of anterogradely labeled fibers (Fig. 5A). In the medial planes of the FN, a well localized small plexus was found in the dorsocaudal part of the FNcm (Fig. 5B). More laterally, anterogradely labeled fibers entered the FN caudally and dorsocaudally and formed a larger terminal plexus in the dorsocaudal part of the caudomedial subdivision of the FN (FNcm) and in the caudal part of the FNm (Fig. 5C-E). In the lateral part of the vermis, P-cell axons were seen in the folial white matter, curving around the dip caudally and dorsally (Fig. 4). These fibers terminated in the caudal end of the FNm and contributed some input to caudoventral areas of the dlp (Fig. 5E). From the most lateral part of the vermis, HRP-labeled fibers were found to form a small terminal plexus in the medial part of the posterior IN (Fig. 5F). Few fibers were also seen curving around the dip, dorsally and caudally, and passing through the lateral FN and the adjacent medial part of the posterior IN, on their way to the juxtarestiform body. In contrast to the anterior half of lobule VI, there was a marked decrease in the number of HRP-labeled passing fibers which coursed between the FN and posterior IN. Only a few terminal fibers were found in the rostral part of the spinal and the superior vestibular nuclei and in the dorsal part of the Deiters' nucleus.
Several retrogradely labeled NC neurons were found among the stained CN fibers of just rostral to these (Fig. 4). These NC cells were fusiform or multipolar neurons of small to medium size. They were seen bilaterally in the same areas of the FN, although the majority of the cells were located on the ipsilateral side (Table II). Multipolar cells of a larger size were also seen in the most medial part of the posterior IN within the CN terminal fibers or just dorsal to those. In some occasions a few single labeled neurons were also found in the LN.
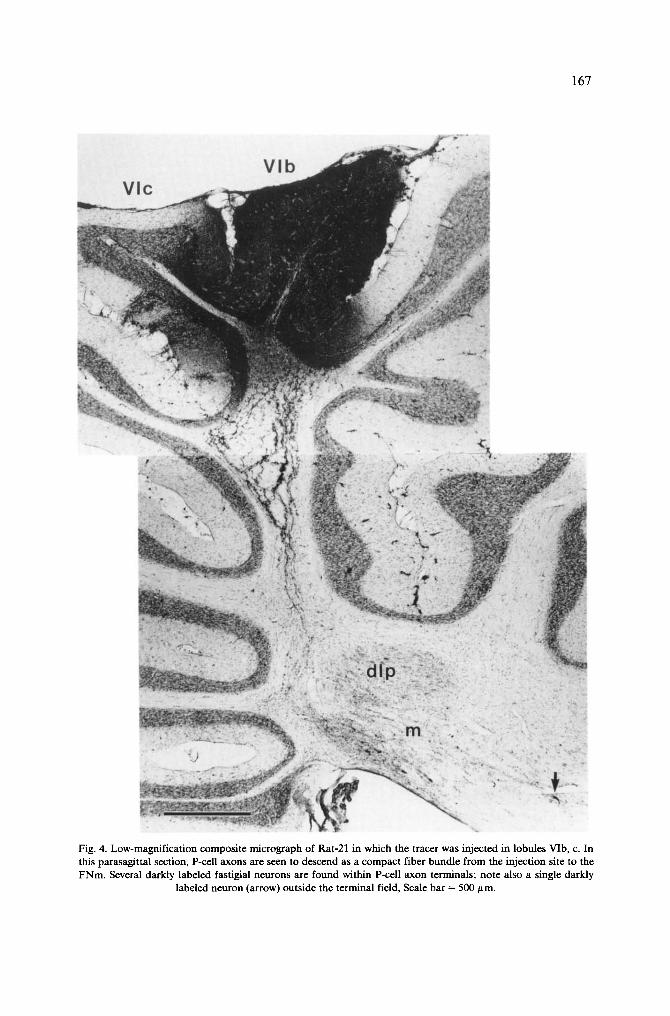
167
Fig. 4. Low-magnification composite micrograph of Rat-21 in which the tracer was injected in lobules VIb, c. In this parasagittal section, P-cell axons are seen to descend as a compact fiber bundle from the injection site to the FNm. Several darkly labeled fastigial neurons are found within P-cell axon terminals; note also a single darkly
labeled neuron (arrow) outside the terminal field, Scale bar = 500 ttm.
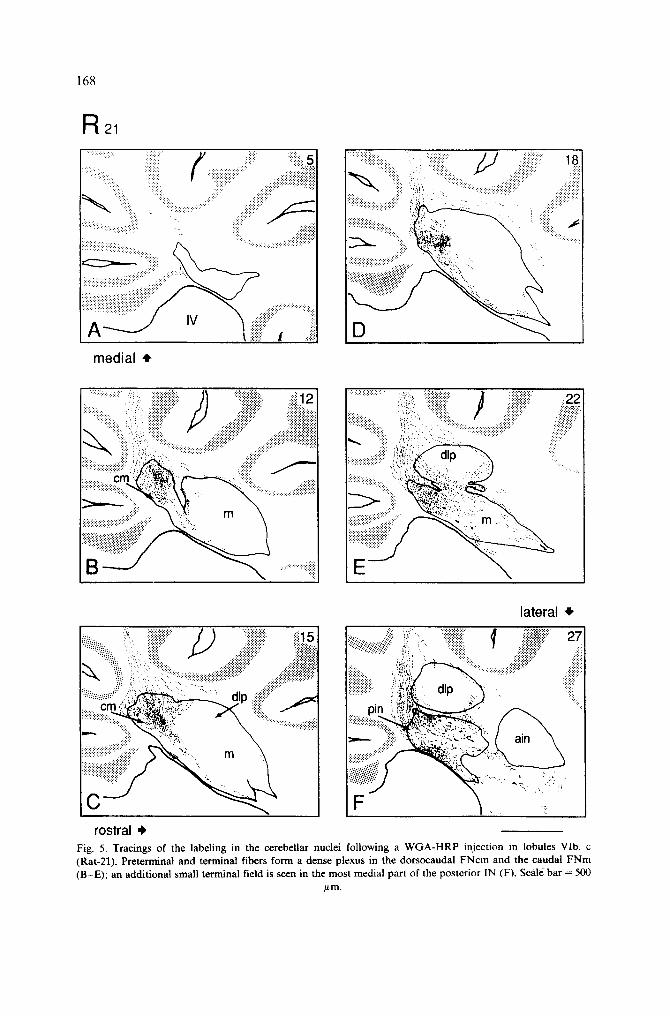
168
R21
I
: . : , . . . .
medial 4,
........ ~ ............ ~:~ i; iii!iiiiiiiiiiiiiiii: / ::~!iiii!iiiiii:: ::i::ii12 .......... ~:':~ ............ ::;~!":i!;; ~:iiiiiiiiiiii; ~': ~ ;iiiii~ii! ;~; ..~.~
: ;
lateral ~,
~ .... ~ ~ / ~ .... ~ ~51 i ~ i ! ~ ~!i~i ~ ii~ 27] :i~ ~ i ........................ ~iii!! ~?
rostral ,I, Fig. 5. Tracings of the labeling in the cerebellar nuclei following a W G A - H R P injection in lobules VIb, c (Rat-21). Preterminal and terminal fibers form a dense plexus in the dorsocaudal F N c m and the caudal F N m (B-E); an additional small terminal field is seen in the most medial part of the posterior IN (F), Scale bar = 500
p,m.
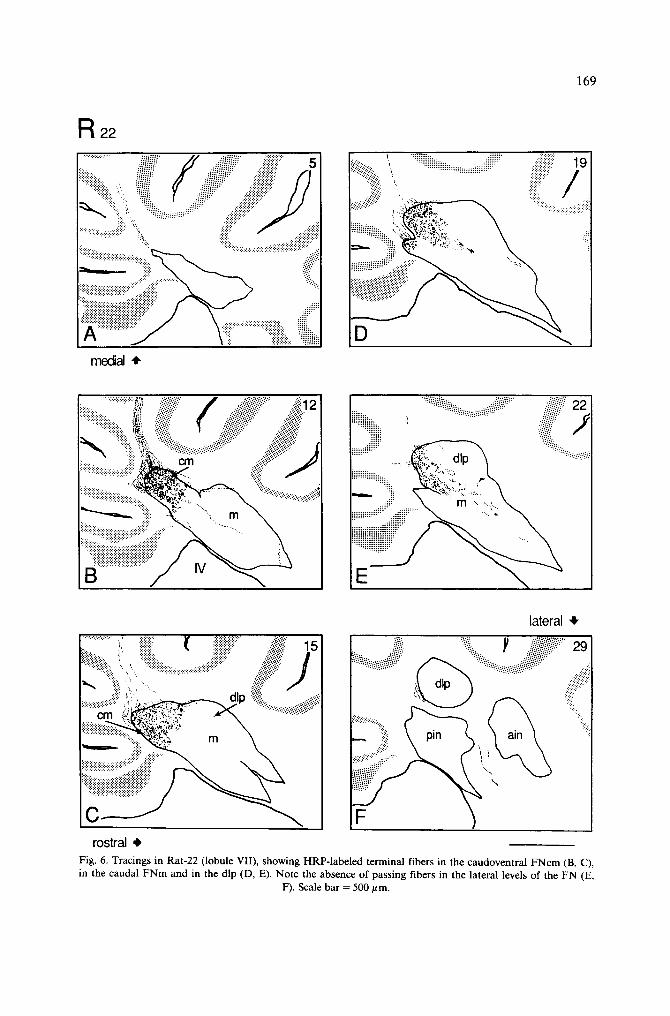
169
R 22
::,::.:i.i.ii ................. ~ ~::iiii!i~ ........................... ~:~:~:.~:.~:.~:.::.:~j~::
medal 4"
~iii~ii~:::~ ", i i ~ "~ i i ! ! i i i ; i i ~ ........... ~iiii! ii~,: ii!i~iiii::~::::~ji:iii:il/::::~:;~i~i~i;~ !: 19
: : : : : : : : : : . ',r.i ~',C.~ "~ :~ '- : - ' : . ' : : .," :5:::.:~"" ~t , ~:':'-.'.:':9. . . . . " . . . .
: . . . . ~; i ~'h.. c m ::....'.:.~::."
..... .,:,,~ ~ "-, . "% . . . . ., ~ ~::::::::::::::::::::::::::::::::::::-.':,ii!::.. '~''" , . . , . . . . "":':~'::"':':':" . . . . . . ":':~"~
.ii ................ ,!, :!iiiili~,i! ' If i~;,',~:,!i~ ':: . . ~ 151
rostral,
lateral ,e,
Fig. 6. Tracings in Rat-22 (lobule VII), showing HRP-labeled terminal fibers in the caudoventral FNcm (B, C), in the caudal FNm and in the dip (D, E). Note the absence of passing fibers in the lateral levels of the FN (E,
F). Scale bar = 500/~m.
1 7 0
R 2o
~!::.::.::,:.~..........S ii',',i',',i:,;,i ~/ iii~;i'~i',ili',i'~iii~,!!ii ;~': ::,,ii~,',,,';;;;ii,~!i!: 5 iii~,:::.:!~'~;!~!i~:'i~iii!i:;, :iiiiiiii i;::ii!i::!: .... iiii!iiiiiiiiilill ~;
media l i
7
:!!i23
....... ' ":: :ii!!!!
i :
? : ,
•!!i,i yliii r ~ i ! ~' .... .:~iii!iii~ ~:: ~o
:,:!~.::,~,'~::~!.~!:~, ,:!!,~: '~:: ~":-"..:..? :.::::::.ii~ ............... ~ ............ ~:i:!ii: >: C I T I " " ; " "":~,,i; 5.,.., "" ' : ' : ' : '" '"
:::::::::::::::::::::::::::::::::::::::: "¢.... " - . ~.,"
!.'.'~> ~.... ,~:::..?~'.;: • .....
. . . . ::.~..<~:~:,.::...,.'...~/~.. .... ::!!i~: ~. \:~,~
:::::::::::::::::::::::::::::::::::::::: '::..
~..'.:':.::..':..:.~
'i:ii!
.... :.:,,,. ,.:..',~ ~:-~:::" ~ : :
rostral ,i~
la tera l ~,
" ~~ ~:! ...... " " ~ ~ . ~ . . . . ~ i ~ :!~ 28
~'.'.::,:..:.'.::.':.<.<..',;...,:. ~ , . , . .
.il) it " : .i "
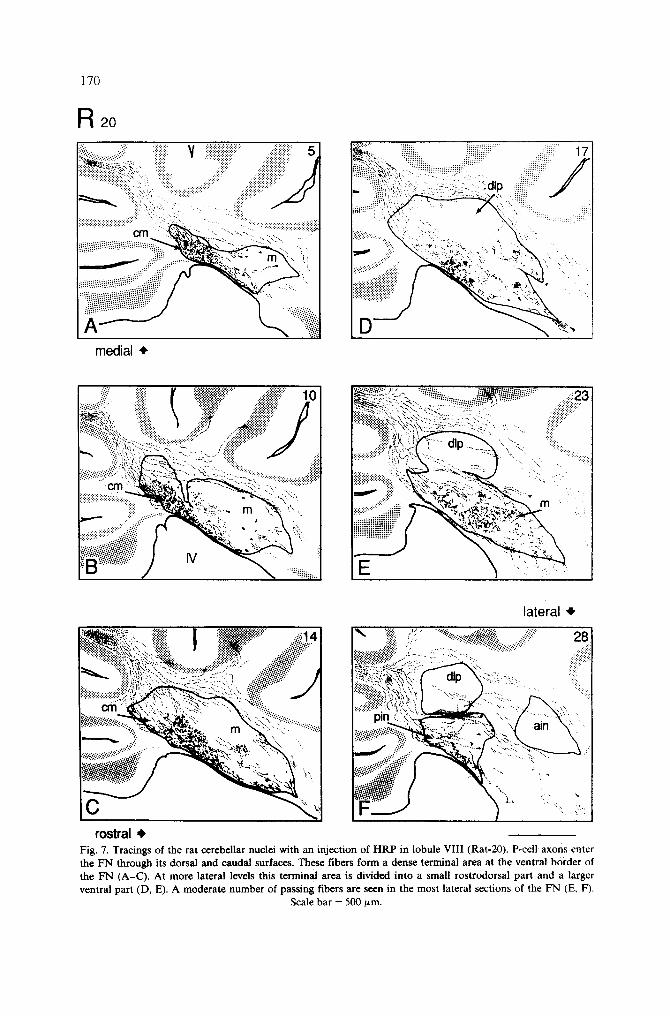
Fig. 7. Tracings of the rat cerebellar nuclei with an injection of HRP in lobule VIII (Rat-20). P-cell axons enter the FN through its dorsal and caudal surfaces. These fibers form a dense terminal area at the ventral border of the FN (A-C) . At more lateral levels tiffs terminal area is divided into a small rostrodorsal part and a larger ventral part (D, E). A moderate number of passing fibers are seen in the most lateral sections of the FN (E, F).
Scale bar = 500 #m.

171
Injections in the vermal lobule VII HRP was successfully deposited in lobule VII in 3 rats (R-19, R-22 and R-26). In 3
other animals, HRP encroached upon caudal portions of lobule VI. These 3 rats served as controls, demonstrating a transitional pattern from lobules VI to VII (Table I, R-18, R-1 and R-12).
HRP was placed in lobule VII in rat R-22. The enzyme was located entirely on the left side, the effective injection site extending from the midline to the entire left vermis (Fig. 1). Although the midsagittal plane was involved in the injection, the most medial sections of the FN were devoid of terminal fibers (Fig. 6A). In these sections, a few preterminal fibers were found caudal to the FN, but they disappeared before entering the nucleus. In slightly more lateral planes, a compact sheet of P-cell axons was found descending from the injection site towards the FN in the caudal half of the deep folial white matter (Fig. 6B). Similar parallel bundles of fibers could be followed from lobule VII to the FN in its medial and lateral planes, except the most medial plane. HRP-labeled CN terminal fibers entered the FN caudally. They formed a dense terminal plexus in the caudal half of the FNcm (Fig. 6B, C). The heaviest terminal labeling was located in the ventral part of the FNcm (Fig. 6B). In more lateral planes, the terminal plexus extended to the caudal end of the FNm and the dlp region (Fig. 6D, E). In all these sections the terminal fibers in FNm and dip were far less numerous than those in the FNcm. Interestingly, a few lightly stained CN fibers were also found in the small-cell region of the posterior IN (Table I). There was also a conspicuous deficiency of HRP-labeled fibers which enter the juxtarestiform body either directly or via the FN (Fig. 6F).
Retrogradely labeled NC neurons were found throughout the mediolateral extent of the FN. These neurons were of small to medium size, fusiform or multipolar cells, which were mainly located among the labeled terminal CN fibers or dorsal to those. This NC projection was bilateral and symmetrical, but the number of retrogradely labeled cells was relatively small as compared to the density of anterogradely labeled CN fibers (Table II).
The distribution of CN fibers in rats R-19 and R-26 resembled that in the former animal. However, the effective cortical labelling in the rat R-19 was confined bilaterally across the midsagittal plane. Anterogradely stained terminal plexi were found bilaterally in the FN, showing a symmetrical pattern. In this case, HRP granules were less concentrated in the terminal fields of the FN when compared to the former animal. On the other hand, the number of retrogradely labeled NC neurons was much higher in this latter case.
Injections in vermal lobule VI I I Three rats were injected in lobule VIII (R-20, R-23 and R-27). All these cases showed
very similar CN and NC projections. The pattern of CN projections from lobule VIII is illustrated with rat R-20. In this case, HRP was placed to the left of the midsagittal plane.
The effective injection covered an area which extended from the lateral vermis across the midline (Fig. 1). In the most medial planes of the ipsilateral FN, a compact bundle of labeled fibers could be traced from the injection site through the deep white matter. These preterminal fibers entered the FN from its dorsocaudal end and formed a dense terminal plexus in the rostroventral half of the FNcm (Fig. 7A). In slightly more lateral planes, the terminal area shifted more rostrally and P-cell axons formed a plexus along the caudo- ventral border of the FNm (Fig. 7C). This area of the terminal plexus was shaped like an irregular triangle, the apex pointing towards the center of the nucleus. Laterally, the plexus was divided into a small rostrodorsal and a larger ventral part (Fig. 7D). In the most lateral planes of the vermis, a few fibers terminated in the ventral border of the

172
posterior IN in its most medial planes (Fig. 7F). In the lateral sections of the FN, bundle~ of HRP-labeled passing fibers were seen, on the ipsilateral side (Fig. 7E). They curved dorsally around the dlp or they coursed ventrally and rostrally through the nucleus. All these bundles of fibers continued towards the vestibular complex via the juxtarestiform body. A moderate number of terminal fibers was seen in the dorsal part of Deiters' nucleus and in the rostrodorsal part of the superior vestibular nucleus.
Numerous retrogradely labeled NC neurons were found in the FN and in the posterior IN (Fig. 7A-F). A few lightly stained neurons were also seen in the LN. Retrogradely labeled small- to medium-sized, fusiform on multipolar, neurons were found throughout the mediolateral extent of the FN bilaterally. On the ipsilateral side, these NC neurons were mainly located among the stained terminal CN fibers or just rostral to these (Fig. 7A-F) . The majority of labeled neurons were located in the FNm, reflecting the major CN projection. A larger number of retrogradely labeled medium- to large-sized neurons, mono- and multipolar, were also found scattered in the medial planes of the posterior IN (Fig. 7F), although the CN projection to this area was small. Both NC projections from the FN and the posterior IN were bilateral and symmetrical. In addition, a few labeled cells were present in the medial planes of the anterior IN, even though no terminal or preterminal CN fibers were seen around this area.
DISCUSSION
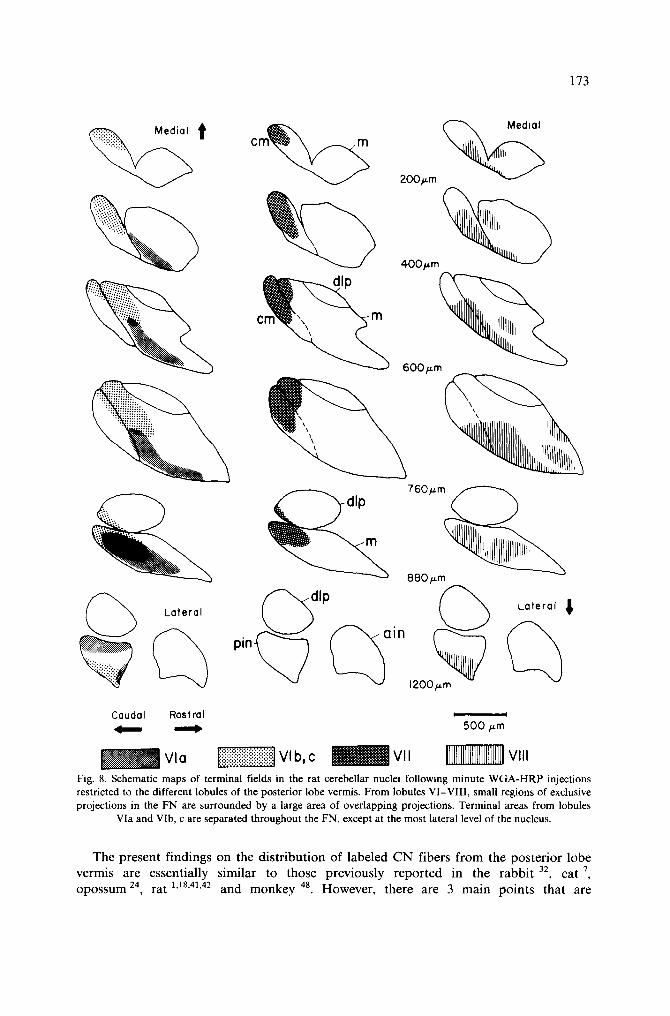
Three main points concerning CN fibers and NC cells of the rat posterior vermal lobules emerge from the present study: (1) CN fibers from vermal lobules VI-VIII terminate in the ipsilateral FN in a rostral-caudal-rostral sequence (Fig. 8), their terminal fields are relatively compact, and for each lobule there is a small region of exclusive projection surrounded by a larger zone, where P-cell axon terminals from two or more lobules overlap. (2) Neuronal somata within the cerebellar nuclei can be retrogradely labeled by cortical placement of HRP; for the large part CN and NC projections are reciprocally organized; retrogradely labeled NC cells are usually associated with ante- rogradely filled CN axons. (3) Some NC cells project to the contralateral cortex and, therefore, are not in the domain of the terminal area of the ipsilateral CN fibers.
Cerebellar corticonuclear (CN) projections Several investigators (see Ito 23 for review) have noted in mammals an orderly relation-
ship between the cerebellar cortex and the nuclei. Studies of myeloarchitecture of the cerebellum in cats 44, ferrets 45 and mice 28, an anterograde degeneration study of CN projections in rabbits 32, and HRP tracing studies in cats 46 and rats 4.5 have delineated 7 structurally distinct compartments (A, B, C 1, C 2, C 3, D 1, D2) separated from each other along the longitudinal sagittal axis and segmented along the transverse lobular axis. There is a general consensus in the recent literature that vermal zone A projects to the ipsilateral FN, and zone B to the ipsilateral Deiters' nucleus. In the cat an additional small x-zone has been recognized in lobule V of the anterior lobe 4o. This zone is known to project to the junctional region between the FN and posterior IN. Although these sagittal compart- ments seem to be a prominent feature of mammalian cerebellar architecture, another type of rostrocaudal topography in the CN projection has also been recognized. Courville and Diakiw 7 have shown in the cat that CN fibers arising from vermal lobules form precise and well-localized terminal areas in the FN, in a circumferential manner. Some degree of topography in this CN projection has also been seen in the rat 1,18,41-43 and in other species 15,20,24,32,48
173
~ Latera
c m ~ 200/~m
400/~m
, ~dlp pln~ ~ain
600/~m
760/J.m~
880p.m Latera~l
1200Fm Caudal Rostral ~ ~ 500/~m
Fig. 8. Schematic maps of terminal fields in the rat cerebellar nuclei following minute WGA-HRP injections restricted to the different lobules of the posterior lobe vermis. From lobules VI-VII1, small regions of exclusive projections in the FN are surrounded by a large area of overlapping projections. Terminal areas from lobules
Via and VIb, c are separated throughout the FN, except at the most lateral level of the nucleus.
T h e p r e sen t f i nd ings o n the d i s t r i b u t i o n of l abe l ed C N f ibers f r o m the pos te r io r lobe vermis are essen t ia l ly s imi la r to those p rev ious ly r e p o r t e d in the r a b b i t 32, cat 7, o p o s s u m 24, rat 1,18,41,42 a n d m o n k e y 48. However , there are 3 m a i n p o i n t s tha t are

174
noteworthy. First, the present study shows that the terminal areas in the FN of the P-cell fibers arising in the anterior (Via) and the posterior (VIb, c) folia of lobule VI are distinctly different. These terminal fields overlap only in the most lateral sections of the FN. This is different from the pattern seen following posterior vermis lesions in the cat 7.9.47 primate 15, rabbit 32, and rat ~,ls where terminal fields tend to be more coexistent and no distinction is made between the terminal fields of the sublobules of lobule VI. Second, CN fibers from lobules VIb, c and VII form dense terminal plexi in the caudal end of the FN. Medially, terminal fields are seen in the caudodorsal and caudoventral parts of the FNcm, respectively. More laterally, terminals are seen in the caudal part of the FNm and a few terminal fibers are also seen in the caudoventral part of the dlp. Compared with the recent literature on cats 7.9.47, rabbits 32 primates 15.20 and rats 1,12.18.41-43, the present results suggest that the terminal fields in the rat FN for posterior vermal lobules VI-VII I contain a more highly defined organization. The distribution of terminal fibers following small injections of W G A -H RP shows that small zones within the FN receive distinct projections from the individual vermal lobules (Fig. 8). In each lobule, there is a small region of exclusive projection surrounded by a zone, where P-cell axon terminals overlap with terminals from adjacent cortical regions. This projection pattern is similar to the organization demonstrated for the cat by Courville and Daikiw 7. Third, preterminal and terminal fibers from lobules Via and VIII form dense terminal plexi along the ventral border of the FN (Fig. 8). After a HRP injection into lobule VIII, anterogradely stained P-cell axons were seen to branch and end in two separate terminal plexi, one located ventral and the other in dorsal part of the FNm. This is concordant with that previously demonstrated in the cat 9. The present study shows that the ventral terminal field of lobule VIII and the terminal plexus of lobule Via are almost completely coexistent in the rat. This is different from the organization previously reported for the cat 7.9, primate 15 and rat 1.1s where terminal fields of lobules VI and VIII are located dorsocaudally and caudoventrally, respectively. Furthermore, Courville and Diakiw 7 described for the cat a topographical distribution of terminal fields in the FN, where there was extensive overlap only with the distributions of adjacent cortical regions.
There has been considerable discussion on whether there is a point-to-point representa- tion in cerebellar CN projections 47 Most authors have concluded that there is a certain degree of divergence in the projection, so that each point in the cerebellar cortex corresponds to a large area of a nucleus and, on that basis, the projection is considered to be rather diffusely arranged. The present findings provide new evidence on this question in rats. Small localized injections in the cortex of the posterior lobe vermis (e.g. rats R-25, R-21, R-22, R-20) are not followed by an intense distribution of HRP deposits over a small region in the FN as would be expected in a point-to-point arrangement, but rather by a diffuse distribution which may cover nearly the whole projection region of a given lobule. This is in agreement with the data of the morphology of the FN obtained in Golgi preparations 2.3. The P-cell axon terminals branch extensively and there is an intermin- gling of the axonal fields within the FN and the medial posterior IN. In addition to that principle of divergence, the present results indicate that there is at the same time a convergence of axons from one cortical region within one nuclear region, such that the axons terminals are, for the most part, intermingling with the terminals of adjacent P-cells, but also in some cases with distant P-cells (lobules Via and VIII). Hence, the anatomical arrangement shown here supports the existence of overlaps between the projections of distant cerebellar cortical regions, at least in the case of the posterior vermis of the rat.

175
Cerebellar nucleocortical (NC) projections Recent investigations using anterograde and retrograde tracing methods have shown
that cerebellar nuclear cells project to the overlying cortex 6,8,10,11,13,14,16,17,19,22,34-39. These NC fibers terminate as mossy fibers in the granular layer 27,34,36 and some of these fibers are collaterals of cerebellofugal axons 21.29.31,34,36,37.39
Following small injections in vermal lobules VI-VIII , we found retrogradely labeled cells in various parts of the ipsilateral cerebellar nuclei. HRP placements in single vermal lobules, covering the whole medial-to-lateral extent of the vermal cortex, resulted in labeling of cells in the FN, posterior IN, and occasionally in the LN, respectively. This medial NC projection in the rat is similar to that previously reported for the ca t 8,9.13.14.38, rat 31 and monkey 48
Tolbert et al. 38 found labeled cells in the LN, posterior IN, anterior IN, and FN following HRP injections in lobules VI and VII of macaque monkeys. In contrast, Yamada and Noda 48 reported some LN labeling only when the injection site in lobule VII involved the paravermal area. Also, Hess 22 reported that labeled cells were 'rarely seen' in the rat LN, posterior IN or anterior IN following HRP placements in lobule VII. An LN-to-vermis projection would represent an NC connection considerably out of proportion with the distribution of CN fibers arising from the posterior vermis (see Haines et al. 20 for review). Very little evidence for similar LN projection, following unilateral HRP injections in the vermal cortex, was seen in the present study. In all cases, single labeled cells seen in the LN had very-low-intensity enzyme deposits in their soma. If these few lightly labeled cells represent an LN-to-vermis projection in the rat, this connection is markedly less developed than that reported for macaque monkeys 38
The present results provide evidence for somewhat different contralateral NC projec- tion. Following vermal injections, labeled NC cells were present in the contralateral FN and the posterior IN, both in the absence of CN labeling. These data suggest that some FN and posterior IN neurons project t o the contralaterat vermis, which also receive projections from neurons located in symmetrical areas of the FN and posterior IN, on the ipsilateral side. Although there is a brief description by Gould and Graybiel 14 of a contralateral NC projection, convincing evidence of such a connection was first provided by Haines 16. He described retrogradely labeled cells in the contralateral posterior IN and the LN, following HRP injections in the paraflocculus and noted that they were fewer in number than, but topographically related to, the ipsilateral cell population. Subsequent studies 6,9,10,38 confirmed the existence of a contralateral NC projection. These data suggest that the posterior IN is a primary source of these fibers in addition to the FN. The LN contributes less and the anterior IN apparently is not involved. In our study the occurrence of labeled NC cells in the contralateral FN and the absence of CN labeling strongly suggest that these are contralateral fastigial projections. Likewise, labeled neu- rons found in the medial part of the posterior IN suggest that similar contralateral projection exists from the posterior IN to posterior vermis. Our data partially confirm previous observations 9,14.17.19.27.34 that the contralateral NC cell population is smaller than their ipsilateral counterpart, although in some cases ipsilateral and contralateral cells are almost equal in number (see, e.g., R-20 in Table II).
Corticonuclear (CN) - - nucleocortical (NC) topography The present study with minute HRP injections in the rat allowed a close comparison
between the location of labeled NC cells and the domain of filled CN fibers arising from the same cortical locus. In the rat most retrogradely labeled NC cells were located in or on the immediate periphery of labeled CN terminal fields, confirming previous observa-

176
t ions for cats and rats 6,~-1~. A l though some labeled N C cells were found outs ide CN terminal fields ( ips i la teral side), for the large par t in ject ions in vermal lobules resul ted in over lapp ing retro- and an te rograde label ing in the FN. Buisse re l -De lmas and Angau t ~' have repor ted that N C cells loca ted wi thin the CN te rmina l fields were a lways more s t rongly labe led than those loca ted outs ide the CN terminal fields. This was not conf i rmed in our data , since in tense ly labe led N C cells were seen bo th wi thin and outs ide the te rminal fields (see, e.g., Fig. 4). A n in teres t ing f inding in the present s tudy was the mode ra t e n u m b e r of N C cells found in the con t ra la te ra l FN. These cells were usual ly fewer in number than the c o m b i n e d ips i la tera l cell popu la t ion , but in some cases both cell counts were a lmost equal (see Tab le II). Con t ra l a t e ra l fast igial cells d id not receive P-cell f ibers f rom the vermis to which they projec ted , and appea red to be topograph ica l ly
re la ted to specific sublobules of the cerebel lar vermis on the oppos i t e side. A l though some evidence of a con t ra la te ra l N C pro jec t ion has been repor ted in the rat 6.22, a specific pa t t e rn has not yet been recognized.
This s tudy was not des igned to address the ques t ion of whether or not P-cell f ibers make synapt ic contac t s with the fast igial neurons in a rec iprocal manner . However , an te rograde ly labe led thin f ibers with end swell ing were seen close to some soma ta or p rox ima l dendr i t es and in the neuropi l wi th in obvious range of the more dis ta l dendr i t es of labe led N C neurons. This obse rva t ion together wi th the repor t s of Ha ines and Pearson ~9, Die t r ichs and Wa lbe rg 10,1~ and Die t r ichs g suppor t the no t ion that these CN axons are making synap t ic contac t s on some par ts of the labe led N C neurons found wi thin their t e rmina l fields. Similar ly, those fast igial cells found ou t s ide the C N te rmina l fields on ipsi- and con t ra la te ra l sides can be cons ide red as non- rec ip roca l cells that most p r o b a b l y funct ion d i f ferent ly f rom their rec iproca l counterpar t s .
ACKNOWLEDGEMENTS
W e thank Mr. Jacque K u b l e y for p h o t o g r a p h i c assis tance. This work was s u p p o r t e d by the N a t i o n a l Ins t i tu tes of Heal th , g ran t EY04063.
REFERENCES
1 Armstrong, D.M. and Schild, R.F., An investigation of the cerebellar cortico-nuclear projections in the rat using an autoradiographic tracing method. I. Projections from the vermis, Brain Res., 141 (1978) 1-19.
2 Beitz, A.J. and Chan-Palay, V., The medial cerebellar nucleus in the rat: nuclear volume, cell number~ density and orientation, Neuroscience, 4 (1979) 31-45.
3 Beitz, A.J. and Chan-Palay, V., A Golgi analysis of neuronal organization in the medial cerebellar nucleus of the rat, Neuroscience, 4 (1979) 47-63.
4 Buisseret-Delmas, C., Sagittal organization of the olivocerebcllonuclear pathway in the rat. I. Connections with the nucleus fastigii and the nucleus vestibularis lateralis, Neurosci; Res., 5 (1988) 475-493.
5 Buisseret-Delmas, C., Sagittal organization of the olivocerebellonuclear pathway in the rat; II. Connections with the nucleus interpositus, Neurosci. Res., 5 (1988) 494-512.
6 Buisseret-Delmas, C., and Angaut, P., The cerebellar nucleocortical projections in the rat: a retrograde labelling study using horseradish peroxidase combined to a lectin, Neurosci. Lett., 84 (1988) 255-260.
7 Courville, J. and Diakiw, N., CerebeUar corticonuclear projection in the cat: the vermis of the anterior and posterior lobes, Brain Res., 110 (1976) 1-20.
8 Dietrichs, E., The cerebellar corticonuclear and nucleocortical projections in the cat as studied with anterograde and retrograde transport of horseradish peroxidase. III. The anterior lobe, Anat. EmbryoL, 162 (1981) 223-247.
9 Dietrichs, E., The cerebellar corticonuclear and nucleocortical projections in the cat as studied with anterograde and retrograde transport of horseradish peroxidase. V. The posterior lobe vermis and floccuto- nodular lobe, Anat. EmbryoL, 167 (1983) 449-462.

177
10 Dietrichs, E. and Walberg, F., The cerebellar corticonuclear and nucleocortical projections in the cat as studied with anterograde and retrograde transport of horseradish peroxidase. I. The paramedian lobule, Anat. Embryol., 158 (1979) 13-39.
11 Dietrichs, E. and Walberg, F., The cerebellar corticonuclear and nucleocortical projections in the cat as studied with anterograde and retrograde transport of horseradish peroxidase. II. Lobule simplex, crus I and II, Anat. Embryol., 161 (1980) 83-103.
12 Goodman, D.C., Hallet, R.E. and Welch, R.B., Patterns of localization in the cerebellar cortico-nuclear projections of the albino rat, J. Comp. Neurol., 121 (1963) 51-67.
13 Gould, B.B., The organization of afferents to the cerebellar cortex in the cat: projections from the deep cerebellar nuclei, J. Comp, Neurol., 184 (1979) 7-42.
14 Gould, B.B. and Graybiel, A.M., Afferents to the cerebellar cortex in the cat: Evidence for an intrinsic pathway leading from the deep nuclei to the cortex, Brain Res., 110 (1976) 601-611.
15 Haines, D.E., Cerebellar cortical efferents of the posterior lobe vernfis in a prosirnian primate (Galago) and the tree shrew (Tupaia), J. Comp. Neurol., 163 (1975) 21-40.
16 Haines, D.E., Contralateral nucleocortical cells of the paraflocculus of tree shrew (Tupaia glis), Neurosci. Lett., 8 (1978) 183-190.
17 Haines, D.E., HRP study of cerebellar corticonuclear-nucleocortical topography of the dorsal culminate lobule-lobule V- in a prosimian primate (Galago): with comments on nucleocortical cell types, J. Comp. Neurol., 282 (1989) 274-292.
18 Haines, D.E. and Koletar, S.L., Topography of cerebellar corticonuclear fibers of the albino rat: vermis of anterior and posterior lobes, Brain Behav. Evol., 16 (1979) 271-292.
19 Haines, D.E. and Pearson, J.C., Cerebellar corticonuclear-nucleocortical topography: a study of the tree shrew (Tupaia) paraflocculus, J. Comp. Neurol., 187 (1979) 745-758.
20 Haines, D.E., Patrick, G.W. and Satrulee, P., Organization of cerebellar corticonuclear fiber systems. In S.L. Palay and V. Chan-Palay (Eds.), The Cerebellum, New Vistas. Springer-Verlag, New York, 1982, pp. 320-371.
21 Hamori, J., Mezey, E. and Szentagothai, J., Electron microscopic identification of cerebellar nucleo-cortical mossy terminals in the rat, Exp. Brain Res., 44 (1981) 97-100.
22 Hess, D.T., Cerebellar nucleo-cortical neurons projecting to the vermis of lobule VII in the rat, Brain Res., 248 (1982) 361-366.
23 Ito, M., The Cerebellum and Neural Control. Raven Press, New York, 1984. 24 Klinkhachorn, P.S., Haines, D.E. and Culberson, J.L., Cerebellar cortical efferent fibers in the North
American opossum, Didelphis virginiana. II. The posterior vermis, J. Comp. Neurol., 227 (1984) 439-451. 25 Korneliussen, H.K., On the morphology and subdivision of the cerebellar nuclei of the rat, J. Hirnforsch, 10
(1968) 109-122. 26 Langer, T., Fuchs, A.F., Chubb, M.C., Scudder, C.A. and Lisberger, S.G., Floccular efferents in the rhesus
macaque as revealed by autoradiography and horseradish peroxidase, J. Comp. Neurol., 235 (1985) 26-37. 27 Legendre, A. and Courville, J., Cerebellar nucleocortical projection with a survey of factors affecting the
transport of radioactive tracers, J. Comp. Neurol., 252 (1986) 392-403. 28 Marani, E., Topographic Enzyme Histochemistry of the Mammalian Cerebellum, 5'-Nucleotidase and
Acetylcholinesterase. Dissertation, University of Leiden, 1982. 29 McCrea, R.A., Bishop, G.A. and Kitai, S.T., Morphological and electrophysiological characteristics of
projection neurons in the nucleus interpositus of the cat cerebellum, J. Comp. Neurol., 181 (1978) 397-420. 30 Mesulam, M.M., Tetramethylbenzidine for horseradish peroxidase neurohistochemistry: a non-carcinogenic
blue reaction-product with superior sensitivity for visualizing neural afferents and efferents, J. Histochem. Cytochem., 26 (1978) 106-117.
31 Payne, J.N., The cerebellar nucleo-cortical projection in the rat studied by retrograde fluorescent double-la- belling method, Brain Res., 271 (1983) 141-144.
32 Van Rossum, J., Corticonuclear and Corticovestibular Projections of the Cerebellum. An Experimental Investi- gation of the Anterior lobe, the Simple Lobule and the Caudal Vermis in the Rabbit. Dissertation, University of Leiden, 1969.
33 Sugita, S., P~i~illysaho, J. and Noda, H., Topographical organization of the olivocerebellar projection upon the posterior vermis in the rat, Neurosci. Res., 7 (1989) 87-102.
34 Tolbert, D.L., The cerebellar nucleocortical pathway, Exp. Brain Res., Suppl., 6 (1982) 296-319. 35 Tolbert, D.L. and Bantli, H., An HRP and autoradiographic study of cerebellar corticonuclear-nucleocortical
reciprocity in the monkey, Exp. Brain Res., 36 (1979) 563-571. 36 Tolbert, D.L., Bantli, H. and Bloedel, J.R., Anatomical and physiological evidence for a cerebellar
nucleo-cortical projection in the cat, Neuroscience, 1 (1976) 205-217.

178
37 Tolbert, D.L., Bantli, H. and Bloedel, J.R., The intracerebellar nucleocorlical projection in a primate, l: W' Brain Res., 30 (1977) 425-434.
38 Tolbert, D.L., Bantli, H. and Bloedel, J.R., Organizational features ¢~f the cat and monkey cerebelklr nucleocortical projection, J. Comp. Neurol., 182 (1978) 39-56.
39 Tolbert, D.L., Bantli, H. and Bloedel, J.R., Multiple branching of cerebellar efferent projections in cats. t~ ~p Brain Res., 31 (1978) 305-316.
40 Trott, J.R. and Armstrong, D.M., The cerebellar corticonuclear projection from lobule Vb /c of the cat anterior lobe: a combined electrophysiological and autoradiographic study. II. Projections from the vermis. Exp. Brain Res., 68 (1987) 339-354.
41 Umetani, T., Topographic organization of the corticonuclear fibers from the tuber vermis and paramedian lobule in the albino rat. Brain Behav. Evol., 651 (1989).
42 Umetani, T. and Tabuchi, T., Topographic organization of the corticonuclear and corticovestibular projec- tions from the pyramis and copula pyramidis in the albino rat: an autoradiographic orthograde tracing study, Brain Behav. Evol., 32 (1988) 160-168.
43 Umetani, T., Tabuchi, T. and Ichimura, R., Cerebellar corticonuclear and corticovestibular fibers from the posterior lobe of the albino rat, with comments on zones, Brain Behav. Evol., 29 (1986) 54-67.
44 Voogd, J., The Cerebellum of the Cat. Dissertation, University of Leiden, 1964. 45 Voogd, J., The importance of fiber connections in the comparative anatomy of the mammalian cerebellum.
In R. Llinas (Ed.), Neurobiology of Cerebellar Evolution and Development. A.M.A., Chicago, 1969, pp. 493-541.
46 Voogd, J. and Bigar~., F., Topographical distribution of olivary and corticonuclear fibers in the cerebellum: a review. In J. Courville, C. de Montigny and Y. Lamarre (Eds.), The Inferior Olivary Nucleus: Anatomv and Physiology. Raven Press, New York. 1980, pp. 207-234.
47 Walberg, F. and Jansen, J., Cerebellar corticonuclear projection studied experimentally with silver impreg- nation methods, J. Hirnforsch., 6 (1964) 338-354.
48 Yamada, J. and Noda, H., Afferent and efferent connections of the oculomotor cerebellar vermis in the macaque monkey, J. Comp. Neurol., 265 (1987) 224-241.





























