Embed Size (px)
Citation preview

Epilepsy Research, 16 (1993) 1-9 0920-121 l/93/%06.00 0 1993 Elsevier Science Publishers B.V. All rights reserved
EPIRES 00595
Cerebellar neuronal activity correlates with spike and wave EEG patterns in the rat
Adam Kandel, Gyijrgy Buz&ki*
Center for Molecular and Behavioral Neuroscience, Rutgers, The State University of New Jersey,
197 University Avenue, Newark, NJ 07102. USA
(Received 28 January 1993; revision received 10 April 1993; accepted 26 April 1993)
Abstract
In this study we investigated the involvement of the cerebellum in high voltage spike-and-wave spindles, a rodent model of petit mal epilepsy. High voltage spindles, recorded epidurally from the sensorimotor neocortex, were correlated with single or multiple unit activity in the cerebellar cortex and deep cerebellar nuclei. The majority of neurons or neuronal groups in the cerebellum (77.9%) fired rhythmically and phase-locked with the high voltage spindles, either during the spike (43.2%; n =41) or during the wave (34.7%; n = 33) component of the high voltage spindle. Tremor of the head and neck musculature, recorded with an accelero-meter, occurred during the high vohage spindle in approximately half of the rats. When present, rhythmic movement occurred predominantly during the wave phase of the high voltage spindle. The remaining half of the rats did not show tremor during high voltage spindles but, nevertheless, had cerebellar units that burst during the spike or wave phase of the high voltage spindle. These latter results demonstrate that phase-locked bursting of cerebellar units during high voltage spindle is independent of rhythmic movement. The findings suggest that rhythmic output from the cerebellum may contribute to the maintenance of generalized petit ma1 seizures.
Key words: Petit ma1 epilepsy; Cerebellum; Unit activity; (Rat)
1. Introduction
High voltage spike-and-wave spindles (HVS), which appear during immobility and sleep in var- ious strains of rodents, represent an animal model for the spike-and-wave patterns found in children with petit ma1 epilepsy [22,30,31]. These cortical events are thought to be initiated in the thalamus [4,27]. Rhythmic thalamic neuronal network bursts precede cortical HVS patterns and the ventral anterior and the ventral lateral (VA/VL) thalamic nuclei have been implicated as the regions where
*Correspon~ng author. Tel.: (201) 648-1080 ext. 3131; Fax: (201) 648-1588.
these oscillations are initiated in man and in ro- dent models for petit ma1 epilepsy [4,5,14,27].
A major input to the ventrobasal complex (VA/ VL nuclei) of the thalamus originates from the cer- ebellum [16]. Most of the thalamofugal efferents from the cerebellum project directly onto thalamo- cortical cells via the superior cerebellar peduncle [1,33]. These efferents originate solely from the deep cerebellar nuclei: the dentate, interpositus and fastigial nuclei. The dentate and interpositus nuclei form topographical connections with the VAjVL thalamic nuclei while the fastigial nucleus is thought to have connections predominantly with limbic structures (amygdala and hippocampus) [1,12,15].

2
Given the intimate relationship between the cere- bellar output and the ventrobasal region of the thalamus, we sought to investigate the possible in- volvement of cerebellar neuronal activity in HVS. This was accomplished by recording extracellularly from isolated neurons in the cerebellar cortex and deep cerebellar nuclei before and during HVS. Part of these findings have been reported in abstract form [17].
2. Ex~ri~ent~ procedures
2. I. Surgery
Twenty three male and female rats, approxi- mately 6 months of age (200400 g), were used in this study. Fifteen of them were of the WAG/Rij strain from Dr. Gilles van Luijtelaar’s colony (University of Nijmegen, The Netherlands) as well as from Harlan Sprague-Dawley of The Nether- lands. Another eight rats were the Fz descendants of the Brown-Norway (BN) and Fischer 344 (F344) inbred strains. This latter group was cho- sen because HVS is often associated with tremor of the vibrissae and head in these rats [28]. The animals were housed in individual cages, given ad libitum food and water and maintained on a 12: 12 1ight:dark cycle. They were anesthetized with a mixture of ketamine (25 mg/ml), xylazine (1.3 mg/ ml) and acepromazine (0.24 mg/ml) at a dosage of 4 ml/kg of weight. Following anesthesia animals were placed in a Kopf stereotaxic instrument. For recording cortical field potentials (EEG) four stainless steel epidural electrodes were placed bilat- erally in the skull (AP = 0.0 mm; L = f4.5 mm; AP = 2.5; L = +2.5 mm; relative to bregma). An approximately 3 mm diameter piece of the skull over the cerebellum was excised with the center of the hole at AP = 11.5 mm and L = 0.5 mm (relative to bregma). This area is above the largest extent of the deep cerebellar nuclei [23]. A plastic microelectrode drive holder was placed above the hole and fixed to the skull with dental acrylic. Stainless steel screw electrodes just anterior and lateral to the microdrive holder served as ground and indifferent electrodes, respectively.
2.2. Physiological testing
At least 3 days after surgery animals were placed
in clear plastic testing chambers where recording sessions took place. A movable microelectrode drive for recording unit activity was placed in the drive holder. Microelectrodes (l-3 MQ tungsten wire) were inserted eccentrically in the microelec- trode drive, which enabled us to record from multi- ple tracks from within the same opening. Unit ac- tivity and neocortical EEG was amplified and fil- tered (Grass model 12). The filters for unit activity were set between 500 Hz and 10 kHz and for the EEG between 1 Hz and 1 kHz. Tremor and move- ment were recorded with an electrothermomecha- nical film (ETMF) accelero-meter (supplied from the University of Kuopio, Finland). All animals were able to move freely at all times. Physiological data were sampled at 10 kHz and recorded on a high capacity optical disk. Once a cell or a group of cells was well isolated, with better than 3:l sig- nal/background ratio, recording was initiated. In these studies no effort was made to positively iden- tify the different cerebellar cell types. The relation- ship between unit firing and HVS was assessed by computing the cross-correlation between these two events using the peak of the spike component of HVS as a reference event. Once testing was com- pleted the animals were perfused with the micro- electrode still in the cerebellum. Perfusion was made transcardially with physiological saline fol- lowed by 4% paraformaldehyde. Brains were re- moved and postfixed in paraformaldehyde and were sectioned at 40 mm on a freezing microtome and stained with cresyl violet to verify electrode placement.
3. Results
3.1. Relationship between cerebellar unit firing and
neocor tical H VS
A total of 95 units (multi or single) from the cerebellar cortex and deep cerebellar nuclei were isolated and recorded during HVS. Typical exam- ples are illustrated in Fig. 1. With the exception of putative Purkinje cells, it was not possible to iden- tify the various cell types in the cerebellum. Puta- tive Purkinje cells could be positively identified by the spontaneous occurrence of complex spiking patterns (Fig. 1E) [9]. Interestingly, neurons with complex spikes were rare (less than 10%) either
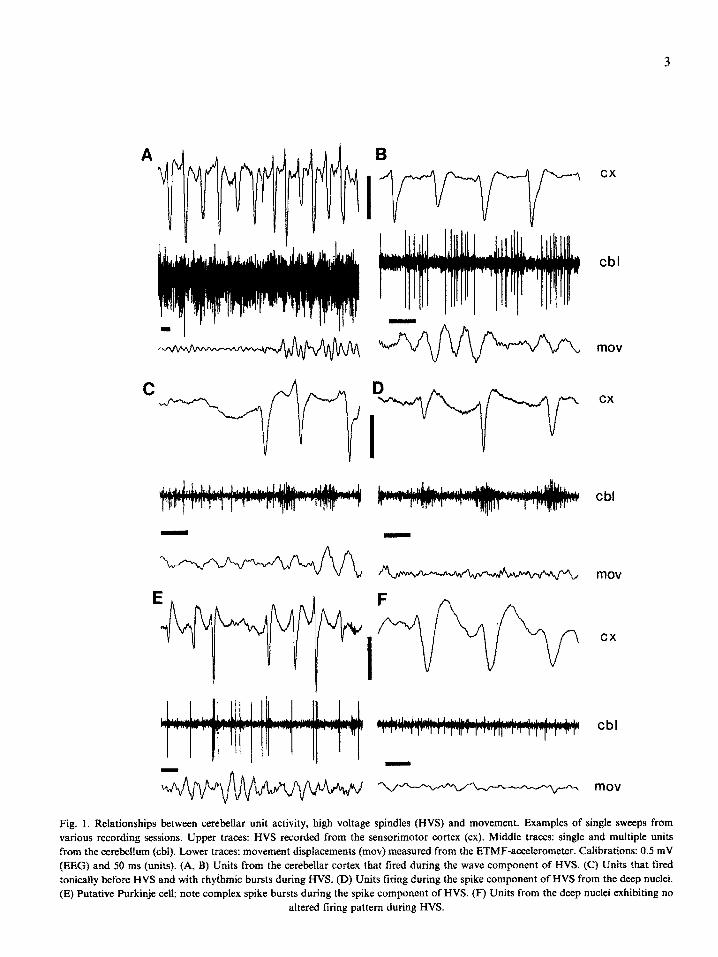
3
A ii-v-v-
Fig. 1. Relationships between cerebellar unit activity, high voltage spindles (HVS) and movement. Examples of single sweeps from
cx
cbl
mov
cbl
mov
cx
cbl
mov
various recording sessions. Upper traces: HVS recorded from the sensorimotor cortex (cx). Middle traces: single and multiple units from the cerebellum (cbl). Lower traces: movement displacements (mov) measured from the ETMF-accelerometer. Calibrations: 0.5 mV (EEG) and SO ms (units). (A, B) Units from the cerebellar cortex that fired during the wave component of HVS. (C) Units that fired tonically before HVS and with rhyihmic bursts during HVS. (D) Units tiring during the spike component of HVS from the deep nuclei. (E) Putative Purkinje cell; note complex spike bursts during the spike component of HVS. (F) Units from the deep nuclei exhibiting no
altered tiring pattern during HVS.
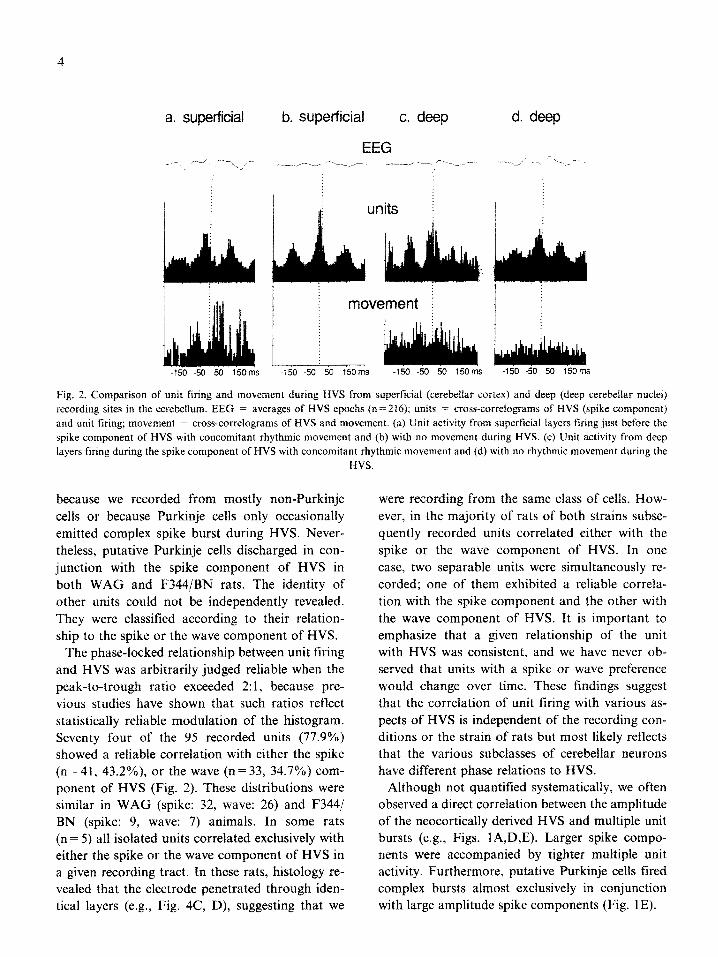
a. superficial b. superficial c. deep
EEG
i movement i
L -150 -50 50 150ms -150 -50 50 150ms -150 -50 50 150ms -150 -50 50 150ms
d. deep
____ ._ -. __-
Fig. 2. Comparison of unit tiring and movement during HVS from superficial (cerebellar cortex) and deep (deep cerebellar nuclei)
recording sites in the cerebellum. EEG = averages of HVS epochs (n = 216): units = cross-correlo~ams of HVS (spike component)
and unit tiring; movement = cross-correlograms of HVS and movement. (a) Unit activity from superficial layers firing just before the
spike component of HVS with concomitant rhythmic movement and (b) with no movement during HVS. (c) Unit activity from deep
layers tiring during the spike component of HVS with concomitant rhythmic movement and (d) with no rhythmic movement during the
HVS.
because we recorded from mostly non-Purkinje cells or because Purkinje cells only occasionally emitted complex spike burst during HVS. Never- theless, putative Purkinje cells discharged in con- junction with the spike component of HVS in both WAG and F344/BN rats. The identity of other units could not be independently revealed. They were classified according to their relation- ship to the spike or the wave component of HVS.
The phase-locked relationship between unit firing and HVS was arbitrarily judged reliable when the peak-to-trough ratio exceeded 2:1, because pre- vious studies have shown that such ratios reflect statistically reliable modulation of the histogram. Seventy four of the 95 recorded units (77.9%) showed a reliable correlation with either the spike (n=41, 43.2%), or the wave (n=33, 34.7%) com- ponent of HVS (Fig. 2). These distributions were similar in WAG (spike: 32, wave: 26) and F344/ BN (spike: 9, wave: 7) animals. In some rats (n = 5) all isolated units correlated exclusively with either the spike or the wave component of HVS in a given recording tract. In these rats, histology re- vealed that the electrode penetrated through iden- tical layers (e.g., Fig. 4C, D), suggesting that we
were recording from the same class of cells. How- ever, in the majority of rats of both strains subse- quently recorded units correlated either with the spike or the wave component of HVS. In one case, two separable units were simultaneously re- corded; one of them exhibited a reliable correla- tion with the spike component and the other with the wave component of HVS. It is important to emphasize that a given relationship of the unit with HVS was consistent, and we have never ob- served that units with a spike or wave preference would change over time. These findings suggest that the correlation of unit firing with various as- pects of HVS is independent of the recording con- ditions or the strain of rats but most likely reflects that the various subclasses of cerebellar neurons have different phase relations to HVS.
Although not quantified systematicaily, we often observed a direct correlation between the amplitude of the neocortically derived HVS and multiple unit bursts (e.g., Figs. lA,D,E). Larger spike compo- nents were accompanied by tighter multiple unit activity, Furthermore, putative Purkinje cells fired complex bursts almost exclusively in conjunction with large amplitude spike components (Fig. 1 E).

0
.I
.2
-2
4
-5
-6
. : . :
t . . . .
.
i
. : . . i .
: .
.
Spike Wave Other
Fig. 3. Scatter plot of unit activity as a function of phase of
HVS: spike or wave component. Ordinate: depth from the dor-
sal surface of the cerebellar cortex. Spike = unit(s) that signifi-
cantly fired during the spike component of HVS; wave =
unit(s) that significantly fired during the wave component of
HVS; other = units that failed to show phase-locking to HVS.
Units recorded from 0 to 2.5 mm correspond approximately to
the cerebellar cortex and units from 4.0 to 5.5 correspond to the
deep cerebellar nuclei. The area without units (2.54.0) corre-
sponds to white matter [23].
Units that correlated with either the spike or wave component did not appear to cluster in any particular region of the cerebellar cortex or the deep cerebellar nuclei (Fig. 3.). Reconstruction of the sites of recorded units from all animals re- vealed two major areas of cellular activity (Fig. 3). The upper region (O-2.5 mm) roughly corresponds to units that were recorded from the cerebellar cor- tex while the ventral region (45.5 mm) corre- sponds to units that were recorded from the deep cerebellar nuclei [23].
Electrode placements were verified by cresyl vio- let staining (Fig. 4). Again, this figure supports the observation that all regions that we recorded from
5
contain neurons which correlate with either the spike or the wave component of HVS. Units with no particular relationship with HVS were inter- spersed among these neurons.
3.2. Phase-locking of tremor, HVS and cerebellar activity
Visible rhythmic movements of the vibrissae, head and trunk (tremor) were rarely present in rats of the WAG strain but occurred frequently in the F2 descendants of the Brown-Norway and Fischer 344 strains [28]. HVS-associated tremor was recorded as a change of linear velocity by the ETMF-accelerator device. During analysis the analog signal was rectified. Movement was de- fined by a threshold value set above the irregular deflections recorded during immobility epochs without HVS. The digital output pulses of the movement-detector program were cross-correlated with HVS.
Tremor was present and correlated with HVS in 49 cases (59.7%). Of these, tremor movements oc- curred during the wave component of HVS (n = 43; 87.7%). In the remaining 46 cases (40.3%) move- ments of the head and neck musculature were either not present or occurred in an irregular man- ner. More frequently, movement, when present, occurred shortly after the onset of HVS and disap- peared during the latter part of the HVS epoch. These irregular, transient movements did not cor- relate with any phase of HVS (Fig. 2b,d). These findings therefore provide evidence that peripher- ally observable tremor was not a prerequisite for the HVS-associated rhythmic discharge of cerebel- lar neurons.
4. Discussion
All rats used in this study contained cerebellar units that displayed rhythmic bursting patterns that were phase-locked to either the spike or the wave component of HVS. Some animals with HVS-associated rhythmic tiring of cerebellar neu- rons also exhibited rhythmic movements (tremor). However, in several rats HVS was not accompa- nied by tremor of the vibrissae or the head/neck muscles, although cerebellar units were phase- locked to either the spike or wave component of
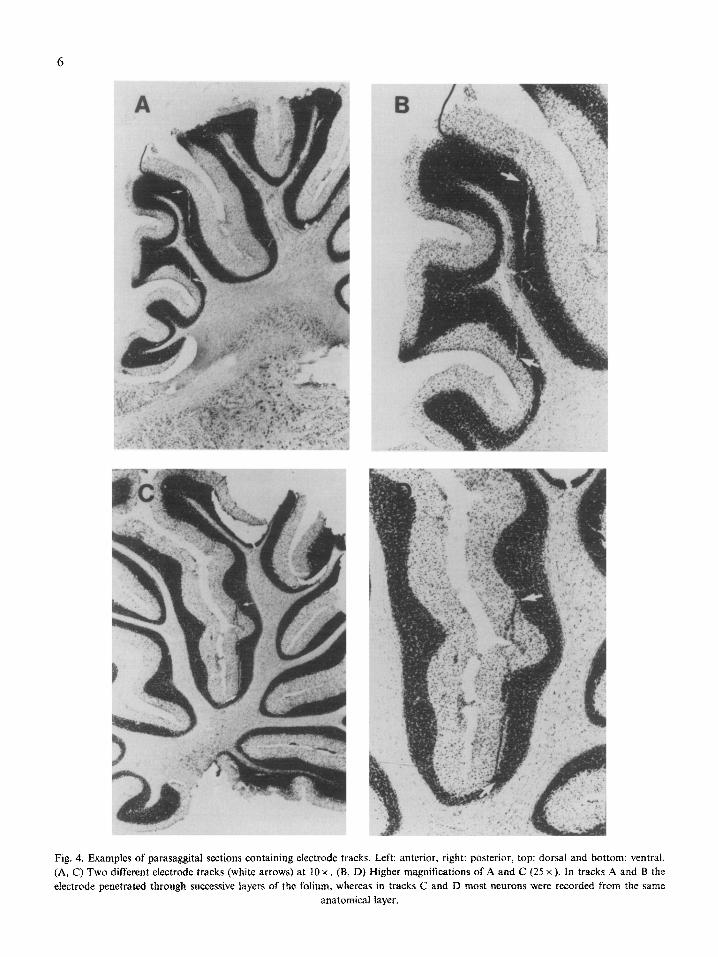
Fig. 4. Examples of parasaggital sections containing electrode tracks. Left: anterior, right: posterior, top: dorsal and bottom: ventrat. (A, C) Two different electrode tracks (white arrows) at 10 x . (B, D) Higher magnifications of A and C (25 x ). In tracks A and B the electrode penetrated through successive layers of the foiium, whereas in tracks C and D most neurons were recorded from the same
anatomical layer.

7
HVS. These findings suggest that neuronal pat- terns in the cerebellum may play a role in the main- tenance of neocortical spike-and-wave activity in petit ma1 epilepsy.
During HVS neurons from a variety of structures are entrained into rhythmic network activity, in- cluding all parts of the neocortex, thalamus, cau- date nucleus, entopeduncular nucleus, basal fore- brain and the locus coeruleus [2,4]. A current view of the generation of neocortically recorded spike- and-wave patterns is that the rhythmic oscillation emerges from the thalamus as a result of the inter- play between the intrinsic properties of individual neurons and the thalamocortical network 16,271. The initiation zone of the emergent thalamic oscil- lation, the ventrobasal thalamic region (VA/VL nuclei), is the main target of cerebellar efferents [l]. Based on the anatomical circuitry alone, one can postulate that rhythmic output from the cere- bellum should have an important role in the main- tenance and/or frequency modification of thalamic oscillations underlying HVS.
Rhythmic inputs to the cerebellum during HVS may be conveyed by any of the several cerebellofu- gal systems. The two main sources of cerebellar afferents are the climbing fibers, which ascend from the inferior olive, and the mossy fibers origi- nating from the pontine nuclei [9,21]. The cerebral cortex projects indirectly to the cerebellum via the pontine nuclei through the corticopontocerebellar pathway. Importantly, these projections originate from cortical areas 4 and 6, the targets of the ven- trobasal thalamus [ 161. The inferior olive receives afferents from large areas of the cerebral cortex and it sends efferents as climbing fibers to the mo- lecular layer of the cerebellar cortex and to the deep cerebellar nuclei [29]. If the cerebellum is in fact synchronized from the cerebral cortex during epilepsy the climbing and mossy/parallel fiber sys- tems would be the most likely candidates by nature of their extensive connections throughout the cere- bellar cortex. The exclusive or predominant role of the olivocerebellar system is reduced by our obser- vation that putative Purkinje cells rarely displayed rhythmic complex spike patterns during HVS. Sti- mulation of the climbing fibers invariably induces complex spikes in Purkinje cells [Zl].
Some of the animals in this study displayed HVS-
associated tremor of the vibrissae and head/neck muscles. The objection that rhythmic cerebellar ac- tivity is caused solely by peripheral reafferentation can be, however, rejected on several grounds. First, rhythmic bursting units in the cerebellum were equally found in rats with and without tremor. Second, careful inspection of the original records in animals with tremor did not reveal a reliable relationship between the amplitude of tremor and multiple unit activity. Third, in cats with a convul- sive form of epilepsy cerebellar activity failed to reveal any correspondence to clonic seizures [lo]. In fact, a major puzzle of petit ma1 epilepsy is the remarkable lack of peripheral muscular activity in spite of the synchronous bursting of neurons in the motor cortex and other motor structures 161.
The modulatory role of the cerebellum in convul- sive forms of epilepsies has been implicated by pre- vious research [3,8,10,32]. Electrical stimulation of the cerebellar cortex as well as the superior cerebel- lar peduncle has been shown to inhibit ongoing seizures and lesions of the cerebellar cortex have increased the severity of seizure activity in rodent models of generalized epilepsy [7,32]. In humans, the cerebellum of some epileptic patients has shown pathological alterations, primarily of the Purkinje cell layer [3,X]. Similar changes have also been seen in animal models for non-convulsive as well as convulsive generalized epilepsies [l&24,25].
~europathological changes have also been re- ported in mutant mouse models of petit ma1 epi- lepsy. The tottering mouse has a slight ataxia, con- comitant with tremor and absence seizures [18,22]. In rats containing the autosomal recessive muta- tions zitter and tremor, seizures have been charac- terized and determined to include tonic convul- sions. Absence-like seizures occurring during im- mobility in these rats were associated with the on- set or the offset of tonic seizures [25]. The histology on these rats showed spongy degeneration of the cerebellum, specifically of granule and Purkinje cells, as well as neuronal degeneration in the thala- mus and the reticular formation [24,25].
Abnormal noradrenergic innervation of the cere- bellar cortex have been observed in several rodent strains with genetic mutations that have a high in- cidence of spike and wave discharges, including the tottering, reeler and staggerer mice [I 8,22,24].

8
Some of these changes include hyperinnervation of
the Purkinje cell layer and an increased sensitivity
of Purkinje cells to the neuromodulatory effects of
norepinepherine [ 11,13,20]. The abnormalities of
noradrenergic innervation may be especially im-
portant in the present context, since neurons in
the locus coeruleus fire phase-locked with HVS
(Berrige, Foote and Buzsaki, unpublished observa-
tions). The altered responsiveness of the cerebel-
lum arising from the widespread modulation of
the noradrenergic fibers may result in excessive
rhythmic output of the cerebellum. It may, there-
by, be causally related to the occurrence of exces-
sive thalamic rhythmicity underlying HVS in the
affected rodent strains.
This study has demonstrated that cerebellar neu-
rons are entrained in rhythmic bursts during HVS
in rats with a high incidence of spike-and-wave
patterns. Such correlations were reliably observed
in both WAG and F344/BN rats. Putative Purkinje
cells fired predominantly during the spike compo-
nent of HVS. It is logical to assume that neurons
that discharged on the spike and wave components
of HVS, respectively, were inhibited by each other.
However, such a statement requires independent
physiological identification of neurons in future
studies.
Excessive oscillation of the thalamic network
may result from a single or from several subtle
alterations, including ion channel or receptor
changes of thalamic neurons, increased inhibition
of thalamocortical cells by the extrapyramidal in-
put or alteration of cellular responsivity to cate-
cholamines [5,6]. The present findings suggest that
increased rhythmic excitation by the cerebellotha-
lamic pathway may also be an important factor.
Our final comment concerns the physiological
role of the cerebellar input in thalamic oscillation.
It is logical to assume that if the cerebellar feed-
back is important in the generation of spike-and-
wave (absence) epilepsy, then the cerebellothalamic
pathway would play a similar role in the mainte-
nance of physiological oscillations, such as sleep
spindles [19,26], and pathologic states, such as Par-
kinsonian tremor [6]. Such a reasoning is sup-
ported by previous observations that barbiturate
spindles can be converted to spike-and-wave pat-
terns in the penicillin model of absence epilepsy
[19]. Nevertheless, such a hypothesis requires the
demonstration of the entrainment of cerebellar
neurons by naturally occurring sleep spindles and
tremor, independent of the occurrence of necorti-
cal HVS.
Acknowledgements
The authors wish to thank Dr. Lasse Raisanen of
the University of Kuopio, Finland, for the kind
gift of the electrothermomechanical film (ETMF)
device with which our movement recordings were
made. The procedure for manufacturing ETMF is
held by Kari Khirjavainen (U.S. Patent, no.
4654546, 1987). We also thank Dr. J.L. Noebels
for discussions and Melissa Hsu for helping with
the histology. This work was supported by
NINDS (NS-27058, 28121) and the Whitehall
Foundation.
References
1
2
3
4
5
6
7
8
9
10
Asanuma, C., Thach, W.T. and Jones, E.G., Anatomical
evidence for segregated focal groupings of efferent cells and
their terminal ramification in the cerebellothalamic pathway
of the monkey, Bruin Res. Rev., 5 (1983) 2677269.
Avoli, M., Gloor, P., Kostopoulos, G. and Gotman, J., An
analysis of penicillin-induced generalized spike and wave
discharges using simultaneous recordings of cortical and
thalamic single neurons, J. Neurophysiol., 50 (1983) 819-
837.
Botez, M.I., Attig, E. and Vezina, J.L., Cerebellar atrophy
in epileptic patients, J. Can. Sci. Neurosci., 15 (1988) 299-
303.
Buzsaki, G., The thalamic clock: emergent network proper-
ties, Neuroscience, 41 (1991) 351-364.
Buzsaki, G., Kennedy, B., Solt, V.B. and Ziegler, M..
Noradrenergic control of thalamic oscillation: the role of
a-2 receptors, Eur. J. Neurosci.. 3 (1991) 222-229.
Buzsaki, G., Smith, A., Berger, S., Fisher, L.J. and Gage,
F.H., Petit ma1 epilepsy and Parkinsonian tremor: hypoth-
esis of a common pacemaker, Neuroscience, 36 (1990) l-14.
Cooke, P.M. and Snider, R.S., Some cerebellar influences
on electrically-induced cerebral seizures, Epilepsia, 4 (1955) 19-28.
Dam, M., The density and ultrastructure of the Purkinje
cells following DPH treatment in animals and man, Acta
Neural. Stand., 48 (Suppl. 49) (1972) I I-65.
Eccles, J.C., Ito, M. and Szentagothai, J., The Cerebelfum as
a Neuronal Machine, Springer-Verlag, New York, NY, 1967.
Fernindez-Guardiola, A., Manni, E., Wilson, J.H. and
Dow, R.S., Microelectrode recording of cerebellar and cere-
bral unit activity during convulsive afterdischarge, Exp.

9
Neurol., 6 (1962) 48-69. 11 Freedman, R., Hoffer, B.J., Woodward, D.J. and Puro, D.,
Interaction of norepinephrine with cerebellar activity evoked by mossy and climbing fibers, Exp. Neural., 55 (1977) 269- 288.
12 Heath, R.G. and Harper, J.W., Ascending projections of the cerebellar fastigial nucleus to the hippocampus, amygdala, and other temporal lobe sites: evoked potentials and histo- logical studies in monkeys and cats, Exp. Neural., 45 (1974) 268-287.
13 Hoffer, B.J., Siggins, G.R. and Bloom, F.E., Studies on norepinephrine-containing afferents to Purkinje cells of rat cerebellum. II. Sensitivity of Purkinje cells to norepinephrine and related substances administered by microiontophoresis, Bruin Res., 25 (1971) 523-534.
14 Hunter, J. and Jasper, H.H., Effects of thalamic stimulation in unanaesthetised convulsions, Clin. Neurophysiol., 1 (1949) 272-298.
15 Iwata, K. and Snider, R.S., Cerebello-hippocampal influ- ences on the electroencephalogram, Electroenceph. Clin.
Neurophysiol., 11 (1959) 439446. 16 Jones, E.G., Functional sub~vision and synaptic organiza-
tion of the mamm~ian thalamus. In: R. Porter (Ed.), Neu- rophysiology ZV, University Park Press, Baltimore, MD, 1981, pp. 1733245.
17 Kandel, A. and Buzsaki, G., Unit recordings from the cere- bellum correlate with spike and wave cortical EEG patterns in the rat, Sot. Neurosci. Abstr., 18 (1992) 554.
18 Kaplan, B.J., Seyfried, T.N. and Glaser, G.H., Spontaneous polyspike discharges in an epileptic mutant mouse (totter- ing), Exp. Neuroi., 66 (1979) 577-586.
19 Kostopoulos. G., Gloor, P., Pellegrini, A. and Gotman, J., A study of the transition from spindles to spike and wave discharge in feline generalized penicillin epilepsy: microphy- siological features, Exp. Neurol., 73 (1981) 55-77.
20 Landis, S.C., Shoemaker, W.J., Schlumpf, M. and Bloom, F.E., Catecholamines in mutant mouse cerebellum: fluores- cence microscopic and chemical studies, Brain Res., 93 (1975) 253-266.
21 Llinas, R.R., Functional significance of the basic cerebellar circuit in motor coordination. In: Bloedel (Ed.), Cerebeflm
Funcfions, Springer-Verlag, Berlin, 1984, pp. 170-185. 22 Noebels, J.L., Isolating single genes of the inherited epilep-
sies, Ann. Neurol., 16 (Suppl.) (1984) Sl8-S21. 23 Paxinos, G. and Watson, C., The Rat Brain in Stereotactic
Coordinates, 2nd edn., Academic Press, San Diego, CA, 1986.
24 Rehm, S., Mehraein, P., Anzil, A.P. and Deerberg, F., A new rat mutant with defective overhairs and spongy degen- eration of the central nervous system: clinical pathological studies, Lob. Anim. Sci., 32 (1982) 70-73.
25 Serikawa, T. and Yamada, J., Epileptic seizures in rats homozygous for two mutations, zitter and tremor, J.
Hered., 77 (1986) 44-444. 26 Steriade, M. and Deschtnes, M., The thalamus as a neuro-
nal oscillator, Bruin Res. Rev., 8 (1984) 163. 27 Steriade, M. and Llinas, R.R., The functional states of the
thalamus and the associated neuronal interplay, Physiol.
Rev., 68 (1988) 649-742. 28 Urioste, R., Horvath, Z., Li, X.-G., Kandel, A., Pierre, E.,
Vadi, D., Hsu, M., Vadasz, C. and Buzsiki, G., Genetical analysis of neocortical-spike-and-wave patterns in inbred rat strains, Sot. Neurosci. Absfr., 17 (1991) 508.
29 van der Want, J.J.L., Wiklund, L., Guegan, M., Ruigrok, T. and Voogd, J., Anterograde tracing of the rat olivocerebel- lar system with PhuseoZus vulgaris leucoagglutinin (PHA-L). Demonstration of climbing fiber collateral innervation of the cerebellar nuclei, J. Camp. Neural., 288 (1989) I-18.
30 Van Luijtelaar, E.L.J.M. and Coenen, A.M.L., Two types of electrocortical paroxysms in an inbred strain of rats, Neuro- sci. Letc., 70 (1986) 393397.
31 Vergnes, M., Marescaux, C., Micheletti, G., Reis, J., Depau- lis, A., Rumbach, L. and Warter, J., Spontaneous paroxys- mal electroclinical patterns in rat: a model of generalized non-convulsive epilepsy, Neurosci. Letr., 33 (1982) 97-101.
32 Willott, J.F. and Urban, G.P., Paleocerebellar lesions en- hance audiogenic seizures in mice, Exp. Neural., 58 (1978)
575-577.
33 Yamamoto, T., Hassler, R., Huber, R., Wagner, A. and Sasaki, K., El~~ophysiologic studies on the pallido- and cere~llothalamic projections in squirrel monkeys (Suimiri sciureus), Exp. Bruin Res., 51 (1983) 77-87.





























