Embed Size (px)
Citation preview

312 (2004) 319–332
www.elsevier.com/locate/jembe
Chaetognaths: a useful model for studying heat
shock proteins. Effect of wound healing
S. Jean, L. De Jong, X. Moreau*
Laboratoire de Biologie Animale (Plancton), ER bBiodiversite et Environnement Q, case 18,
Universite de Provence, 13331 Marseille Cedex3, France
Received 23 February 2004; received in revised form 27 June 2004; accepted 20 July 2004
Abstract
The pattern of expression of Heat Shock Protein 70 (Hsp70), a highly conserved cellular protein
chaperone, was investigated in Chaetognatha, a very important phylum of marine worms, which play
a major role in marine food webs. The in toto distribution of Hsp70-like immunoreactivity was
assessed in both intact and experimentally wounded specimens of Spadella cephaloptera Busch,
1851 that have been cut transversally just above the seminal vesicles. In intact animals, the ciliary
organs, the corona ciliata, the coronal nerve and the spermatocytes express Hsp70 proteins. The
kinetic of the expression pattern has been followed during the wound healing from the lesion to 5
days after. The Hsp70 immunoreactivity was observed according to time in the cerebral and
suboesophageal ganglia and in the plexuses of the hood, the mouth, the neck and the tail region. At
the wound level, the time-dependent Hsp70 expression was detected in the epidermal cells and along
numerous muscle fibres of the tail region. Five days after the tail section, two Hsp70 immunoreactive
areas were observed on both sides of the healed wound that correspond to the sites of formation of
two new seminal vesicles. It is suggested that, in Chaetognaths, Hsp70 may be implicated in the
regulation of several cellular processes especially at the level of the spermatocytes in intact and
wounded specimens, and of the nervous system and muscular apparatus during the wound healing. It
appears that chaetognaths are a good model as indicator of stress responses for experimental studies
at the level of a whole organism.
D 2004 Elsevier B.V. All rights reserved.
Keywords: Chaetognaths; Heat shock proteins; Spadella cephaloptera; Wound
* Corresponding author. Fax: +33 4 91 10 62 65.
0022-0981/$ -
doi:10.1016/j.
E-mail add
Journal of Experimental Marine Biology and Ecology
see front matter D 2004 Elsevier B.V. All rights reserved.
jembe.2004.07.009
ress: [email protected] (X. Moreau).

S. Jean et al. / J. Exp. Mar. Biol. Ecol. 312 (2004) 319–332320
1. Introduction
Chaetognaths, also called arrow worms, are marine organisms that constitute a small
phylum of very abundant animals. They play an important role in marine food webs
(Feigenbaum, 1991) and most of them are planktonic.
For many experimental studies, the chaetognath appears as a model animal for the
following reasons. Indeed, this taxon includes small species; their body is transparent
allowing in toto observations especially convenient for immunolabeling. They have a
simple body plane and lack digestive glands, circulatory apparatus, respiratory and
excretory organs (Duvert et al., 2000 for review). Lastly, benthic species such as Spadella
cephaloptera Busch, 1851 are easily kept alive in laboratory conditions. In natural
environments, living specimens lacking part of their body or showing obvious scars are
not infrequent in some regions of the world (Burfield, 1927; Pierce, 1951; Duvert et al.,
2000).
A previous experimental work (Duvert et al., 2000) was conducted to study the
histological and cytological aspects of wound healing in Chaetognatha. This model
organism appears to be particularly resistant in such experiments. The authors have
shown that specimens could survive for at least 30 days after a decapitation and that, in
such conditions, they are still able to mate. Moreover, after their amputation, the
seminal vesicles are formed in about a week. In fact, chaetognaths lacking head or tail
are able to restore the organization of their bodies by re-establishing their body
cavities. For these reasons, the chaetognath S. cephaloptera has been retained to
investigate some processes involved in the cicatrisation and regeneration of tissues
during wound healing. The cutting of a part of the animal generates an immediate
stress for the organism and the injured tissues. This stressful experiment is postulated
to influence the expression of a category of very important proteins, the heat shock
proteins (Hsp).
Hsp are a family of highly conserved cellular protein chaperones identified in a wide
range of organisms from microorganisms to humans. Constitutively expressed, they play
an important role in fundamental cellular processes such as folding, secretion and
degradation of other proteins (Mayer and Bukau, 1998). They act through cycles of
substrate binding and release governed by ATP binding and hydrolysis (Bukau and
Horwich, 1998; Hartl and Hayer-Hartl, 2002). Most of these proteins, especially the Hsp70
subfamily, are stress-inducible and accumulate in cells upon exposures to various kinds of
chemical and physical stressors. Increased expression of Hsp70 has been shown to be
protective in many cultured cells and animals tissues (Kiang and Tsokos, 1998; Nollen et
al., 1999; Kalmar et al., 2002). This protection is mediated by the capacity of Hsp70 to
prevent inappropriate protein aggregation, to facilitate the restoration of the function of the
renaturated proteins, and to mediate transport of irreversible damaged proteins to
degradative organelles and proteasomes. Thus, Hsp70 proteins allow cells to adapt to
abrupt or gradual changes of their environment and to survive in some cases to lethal
conditions.
The purpose of the present study is to describe the pattern of expression of Hsp70 in the
chaetognath S. cephaloptera, and to assess the effect of wound healing on its expression.
For a better understanding of the location of the regions and body structures that express
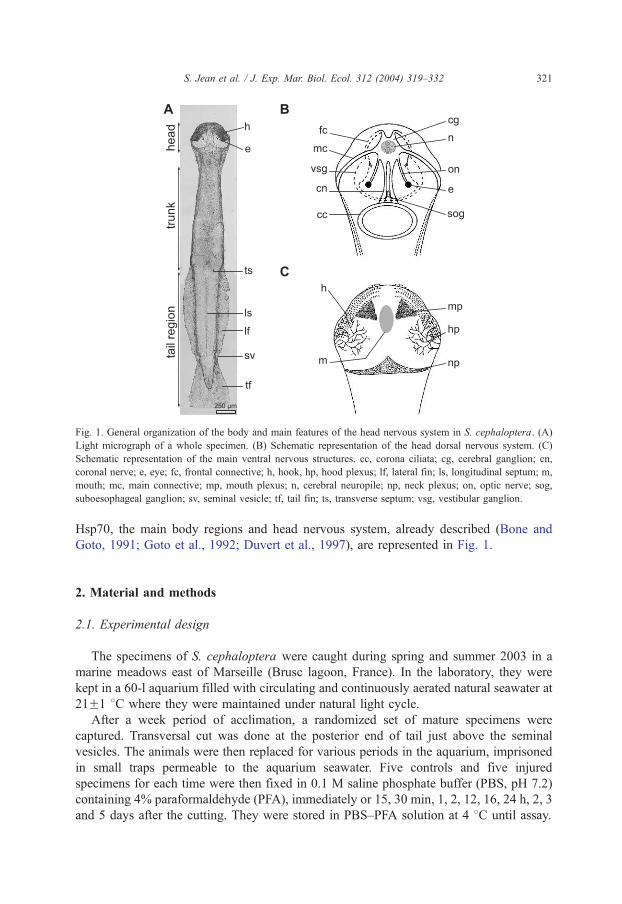
Fig. 1. General organization of the body and main features of the head nervous system in S. cephaloptera. (A)
Light micrograph of a whole specimen. (B) Schematic representation of the head dorsal nervous system. (C)
Schematic representation of the main ventral nervous structures. cc, corona ciliata; cg, cerebral ganglion; cn,
coronal nerve; e, eye; fc, frontal connective; h, hook, hp, hood plexus; lf, lateral fin; ls, longitudinal septum; m,
mouth; mc, main connective; mp, mouth plexus; n, cerebral neuropile; np, neck plexus; on, optic nerve; sog,
suboesophageal ganglion; sv, seminal vesicle; tf, tail fin; ts, transverse septum; vsg, vestibular ganglion.
S. Jean et al. / J. Exp. Mar. Biol. Ecol. 312 (2004) 319–332 321
Hsp70, the main body regions and head nervous system, already described (Bone and
Goto, 1991; Goto et al., 1992; Duvert et al., 1997), are represented in Fig. 1.
2. Material and methods
2.1. Experimental design
The specimens of S. cephaloptera were caught during spring and summer 2003 in a
marine meadows east of Marseille (Brusc lagoon, France). In the laboratory, they were
kept in a 60-l aquarium filled with circulating and continuously aerated natural seawater at
21F1 8C where they were maintained under natural light cycle.
After a week period of acclimation, a randomized set of mature specimens were
captured. Transversal cut was done at the posterior end of tail just above the seminal
vesicles. The animals were then replaced for various periods in the aquarium, imprisoned
in small traps permeable to the aquarium seawater. Five controls and five injured
specimens for each time were then fixed in 0.1 M saline phosphate buffer (PBS, pH 7.2)
containing 4% paraformaldehyde (PFA), immediately or 15, 30 min, 1, 2, 12, 16, 24 h, 2, 3
and 5 days after the cutting. They were stored in PBS–PFA solution at 4 8C until assay.

S. Jean et al. / J. Exp. Mar. Biol. Ecol. 312 (2004) 319–332322
2.2. Immunohistochemistry
Specimens were rinsed in PBS containing 1% Triton X-100 during 2 days. To saturate
nonspecific sites, entire animals were then pre-incubated for 35 min at room temperature
in 0.1 M PBS containing 1% Triton X-100, 10% goat serum and 2% bovine serum
albumin (PBS-T buffer). Animals were incubated overnight at 4 8C in a dilution (1:2000)
of a primary antibody against Hsp70 raised in mouse (Sigma, Saint Quentin-Fallavier,
France) in PBS-T buffer. This antibody reacts against both the constitutive and the
inducible form of Hsp70. Specimens were then rinsed three times and were incubated for
an hour, in the dark and at room temperature, with a secondary antibody (1:200), goat anti-
mouse IgG TRITC-conjugated (Sigma) diluted in PBS-T buffer. After rinsing in PBS for
2–3 h at room temperature, animals were mounted in a medium containing antifading
(Gel/MountR, Bibmeda, Foster City, CA, USA). Preparations were observed under a
fluorescence microscope (Leica M1560).
3. Results
3.1. Intact specimens
Fig. 2A shows the pattern of expression of Hsp70-like proteins in intact adult
specimens. A positive reaction is observed at the level of the corona ciliata and along the
coronal nerve (Fig. 2B). In the cerebral ganglion, a strong spot-like labeling is present near
the origin of the coronal nerves (Fig. 2C). Aweak labeled network in the epidermis is also
observed and it intensifies near by the corona ciliata (Fig. 2D) and ciliary organs (Fig. 2E),
these latter being distributed all along the body from the head to the tip of the tail (Fig.
2A). Sensory fibres that connect the ciliary organs are Hsp70-like immunoreactive (Fig.
2F). In the tail region, behind the transverse septum, and on both side of the longitudinal
septum, spermatocytes exhibit Hsp70 labeling (Fig. 2G,H). Content of the seminal vesicles
are not immunoreactive (Fig. 2I,J).
3.2. Wounded specimens
During wound healing, the immunoreactive structures observed on intact specimens are
still labeled. However, other labeled structures appear in some parts of the cerebral
nervous system and at the level of the wound. The following results show the
modifications of the expression pattern of Hsp70 in these body regions according to
time after tail section.
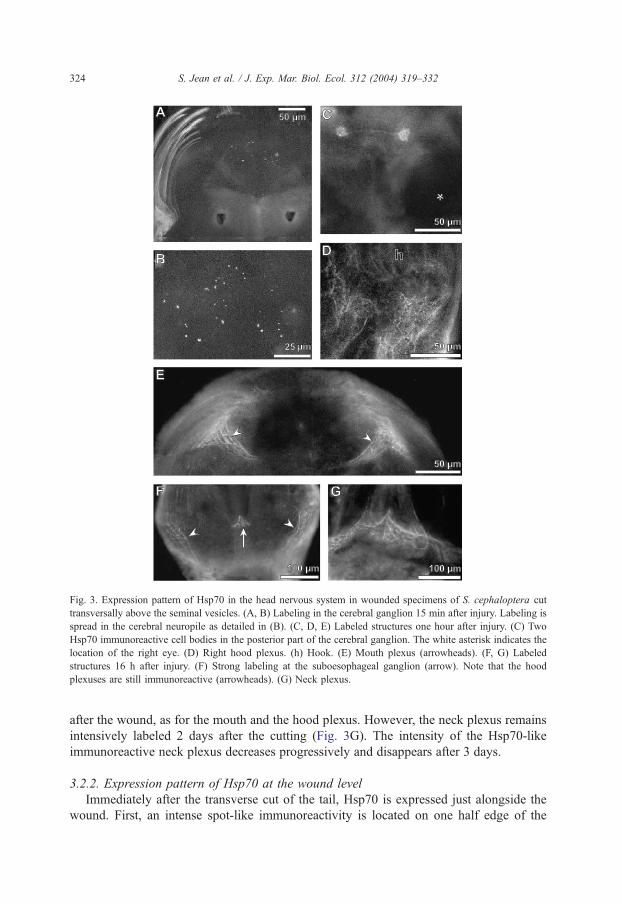
3.2.1. Expression pattern of Hsp70 in the cerebral nervous system
The first labeling appears in the cerebral ganglion 15 min after injury (Fig. 3A). It spreads
in the cerebral neuropile (Fig. 3B) and is observed until 30 min. At 30 min, the labeling is
concentrated in two symmetrical areas that correspond to cell bodies in the posterior part of
the cerebral ganglion (Fig. 3C). At the same time, the hood and the mouth plexuses are
immunoreactive (Fig. 3D,E). Two hours after injury, the cell bodies are no more reactive
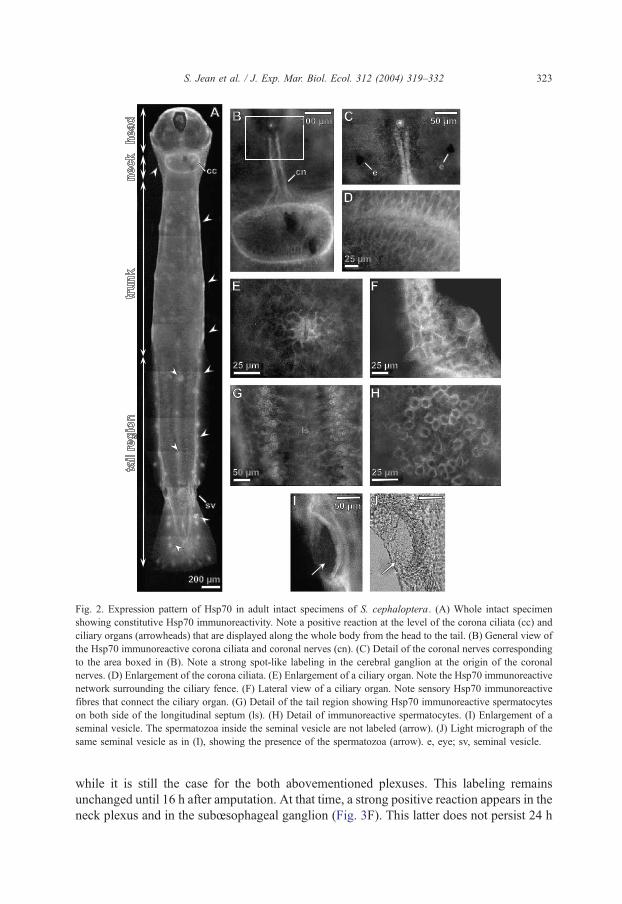
Fig. 2. Expression pattern of Hsp70 in adult intact specimens of S. cephaloptera. (A) Whole intact specimen
showing constitutive Hsp70 immunoreactivity. Note a positive reaction at the level of the corona ciliata (cc) and
ciliary organs (arrowheads) that are displayed along the whole body from the head to the tail. (B) General view of
the Hsp70 immunoreactive corona ciliata and coronal nerves (cn). (C) Detail of the coronal nerves corresponding
to the area boxed in (B). Note a strong spot-like labeling in the cerebral ganglion at the origin of the coronal
nerves. (D) Enlargement of the corona ciliata. (E) Enlargement of a ciliary organ. Note the Hsp70 immunoreactive
network surrounding the ciliary fence. (F) Lateral view of a ciliary organ. Note sensory Hsp70 immunoreactive
fibres that connect the ciliary organ. (G) Detail of the tail region showing Hsp70 immunoreactive spermatocytes
on both side of the longitudinal septum (ls). (H) Detail of immunoreactive spermatocytes. (I) Enlargement of a
seminal vesicle. The spermatozoa inside the seminal vesicle are not labeled (arrow). (J) Light micrograph of the
same seminal vesicle as in (I), showing the presence of the spermatozoa (arrow). e, eye; sv, seminal vesicle.
S. Jean et al. / J. Exp. Mar. Biol. Ecol. 312 (2004) 319–332 323
while it is still the case for the both abovementioned plexuses. This labeling remains
unchanged until 16 h after amputation. At that time, a strong positive reaction appears in the
neck plexus and in the subKsophageal ganglion (Fig. 3F). This latter does not persist 24 h
2525
Fig. 3. Expression pattern of Hsp70 in the head nervous system in wounded specimens of S. cephaloptera cut
transversally above the seminal vesicles. (A, B) Labeling in the cerebral ganglion 15 min after injury. Labeling is
spread in the cerebral neuropile as detailed in (B). (C, D, E) Labeled structures one hour after injury. (C) Two
Hsp70 immunoreactive cell bodies in the posterior part of the cerebral ganglion. The white asterisk indicates the
location of the right eye. (D) Right hood plexus. (h) Hook. (E) Mouth plexus (arrowheads). (F, G) Labeled
structures 16 h after injury. (F) Strong labeling at the suboesophageal ganglion (arrow). Note that the hood
plexuses are still immunoreactive (arrowheads). (G) Neck plexus.
S. Jean et al. / J. Exp. Mar. Biol. Ecol. 312 (2004) 319–332324
after the wound, as for the mouth and the hood plexus. However, the neck plexus remains
intensively labeled 2 days after the cutting (Fig. 3G). The intensity of the Hsp70-like
immunoreactive neck plexus decreases progressively and disappears after 3 days.
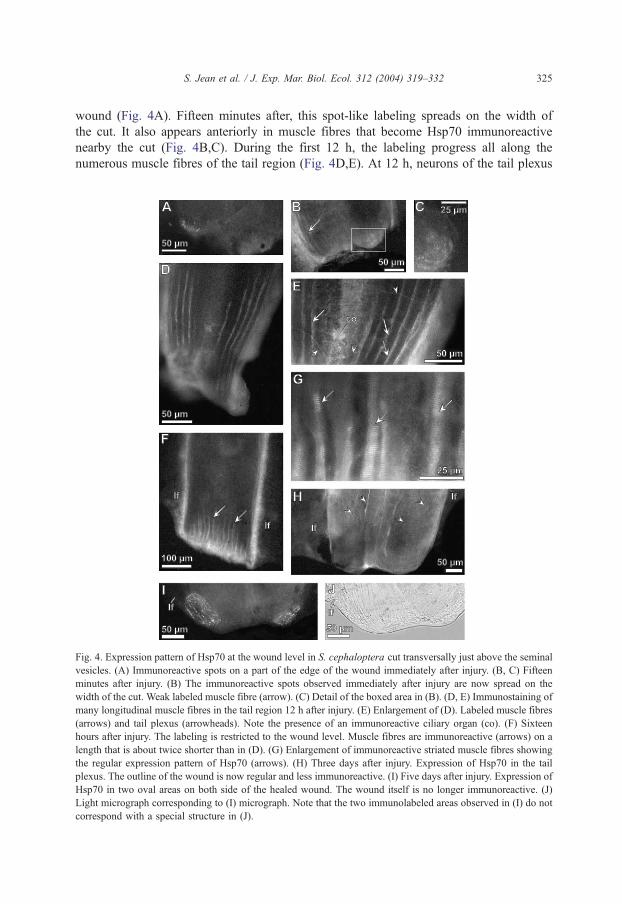
3.2.2. Expression pattern of Hsp70 at the wound level
Immediately after the transverse cut of the tail, Hsp70 is expressed just alongside the
wound. First, an intense spot-like immunoreactivity is located on one half edge of the
S. Jean et al. / J. Exp. Mar. Biol. Ecol. 312 (2004) 319–332 325
wound (Fig. 4A). Fifteen minutes after, this spot-like labeling spreads on the width of
the cut. It also appears anteriorly in muscle fibres that become Hsp70 immunoreactive
nearby the cut (Fig. 4B,C). During the first 12 h, the labeling progress all along the
numerous muscle fibres of the tail region (Fig. 4D,E). At 12 h, neurons of the tail plexus
Fig. 4. Expression pattern of Hsp70 at the wound level in S. cephaloptera cut transversally just above the seminal
vesicles. (A) Immunoreactive spots on a part of the edge of the wound immediately after injury. (B, C) Fifteen
minutes after injury. (B) The immunoreactive spots observed immediately after injury are now spread on the
width of the cut. Weak labeled muscle fibre (arrow). (C) Detail of the boxed area in (B). (D, E) Immunostaining of
many longitudinal muscle fibres in the tail region 12 h after injury. (E) Enlargement of (D). Labeled muscle fibres
(arrows) and tail plexus (arrowheads). Note the presence of an immunoreactive ciliary organ (co). (F) Sixteen
hours after injury. The labeling is restricted to the wound level. Muscle fibres are immunoreactive (arrows) on a
length that is about twice shorter than in (D). (G) Enlargement of immunoreactive striated muscle fibres showing
the regular expression pattern of Hsp70 (arrows). (H) Three days after injury. Expression of Hsp70 in the tail
plexus. The outline of the wound is now regular and less immunoreactive. (I) Five days after injury. Expression of
Hsp70 in two oval areas on both side of the healed wound. The wound itself is no longer immunoreactive. (J)
Light micrograph corresponding to (I) micrograph. Note that the two immunolabeled areas observed in (I) do not
correspond with a special structure in (J).

S. Jean et al. / J. Exp. Mar. Biol. Ecol. 312 (2004) 319–332326
express Hsp70 (Fig. 4E). After 16 h, the labeling of the muscle fibres is restricted to the
wound level and is no more visible anteriorly in the tail region (Fig. 4F). In the wound
area, the expression pattern of Hsp70 in striated muscle fibres displays a scale-like
aspect (Fig. 4G). It becomes weaker from 1 to 5 days after the tail section (Fig. 4H),
which allows to set off the Hsp70 immunoreactive tail plexus. During this period, the
outlines of the wound, which had a rough aspect, become regular and smooth. Five days
after the operation, two strong Hsp70-like immunoreactive areas are localized on both
side of the healed wound (Fig. 4I). This unusual labeling does not seem to correspond
with a peculiar structure when animals are observed under transmitted light microscope
(Fig. 4J).
4. Discussion
Only a few studies on multicellular eukaryotes have reported the expression pattern of
Hsp proteins in intact tissues and organisms (Feder and Hofmann, 1999 for review).
Moreover, most of works dealing with Hsp70 expression have been performed under
thermal stress conditions (Feder and Hofmann, 1999 for review). For the first time, the
constitutive expression of Hsp70 proteins in non-stressed and entire animals is described
here using a new animal model, a chaetognath. Furthermore, owing to the remarkable
healing capacity of the chaetognath S. cephaloptera (Duvert et al., 2000), the expression
pattern of Hsp70 following a lesion is observed in the present study.
4.1. Distribution of Hsp70 in control animals
4.1.1. Nervous system and sensitive organs
The immunohistochemical investigations reveal the presence of Hsp70 in sensitive
organs consisting of fans of elongated cilia arranged all along the body. The ciliated
cells constitute sensory cells that receive synapses at their bases (Welsh and Storch,
1983). Cilia and sensory cells are involved in the response to short range vibrations,
chiefly to deformation by water movements in the transverse plane to the axis of the
organ. Hsp70 is also expressed in the corona ciliata. In S. cephaloptera, this organ is
elliptical and lies across the neck where an inner ring of secretory cells is surrounded by
a single ring of ciliated cells. The function of the ciliary loop remains enigmatic.
Malakhov and Frid (1984) have first described this organ at the ultrastructural level and
have shown the presence of primary receptor cells with branching cilia. To a certain
extent, as the corona ciliata is linked to the coronal nerve, which is itself connected to
the cerebral ganglion, this former may have sensitive functions. Indeed, increasing body
of evidence shows that cilia also play prominent roles as sensory organelles (Sloboda,
2002). However, on the basis of the present results, it is difficult to hypothesize the
Hsp70 function in the ciliary organs of chaetognaths. Cilia and flagella are dynamic cell
structures presenting a continuous transport of axonemal particles between them and the
cytoplasm. It has been shown that, in green microalgae, in protozoa and in sea urchin
embryos, several Hsp70 family members are associated with microtubule nucleation,
tubulin transport and/or axoneme assembly (Bloch and Johnson, 1995; Williams and

S. Jean et al. / J. Exp. Mar. Biol. Ecol. 312 (2004) 319–332 327
Nelsen, 1997; Stephens, 1997). Therefore, these molecular chaperones, also involved in
protein turnover, play an important role in the structure and the biogenesis of cilia
(Stephens and Lemieux, 1999). In chaetognaths, the intense Hsp70-like immunoreac-
tivity of the ciliary organs and the corona ciliata may be explained by at least two non-
exclusive hypotheses. First, the cilia of these two structures that are exposed to the
external environment might be frequently damaged and so, it can be assumed that an
intense protein turnover occurs. A second hypothesis could be linked to the presence of
nervous terminals in the ciliary organs and in the corona ciliata. Hsp70 immunoreactivity
in the coronal nerve strengthens this hypothesis. Indeed, the constitutive form of Hsp70
plays a critical role in the regulation of the neurotransmitter release at the level of the
synaptic contacts (Zinsmaier and Bronk, 2001). This process leads to repeated cycles of
exocytosis and endocytosis. It also requires that the participating components are rapidly
recycled and that they sustain an optimal conformation despite repeated use. Hsp70
proteins are involved in the process of exocytosis (Zinsmaier et al., 1994;
Mastrogiacomo et al., 1994; Bronk et al., 2001) and take part in the synaptic vesicle
recycling (Morgan et al., 2001).
4.1.2. Testes
Hsp70 proteins were also observed in circulating spermatocytes in the general cavity
of the tail region. In these animals, the spermatogenesis occurs according to a special
mode. Indeed, the testes lie in the tail cavities close to the transverse septum. Clusters of
spermatogonia continuously depart from the testes and continue their maturation while
floating in the liquid which fills the cKlomic cavities; different stages of spermato-
genesis co-exist in these tail cavities but immature spermatocytes predominate. They are
in constant motion in each tail cavity, owing to the ciliated cells that cover the median
septum (Ghirardelli, 1968). The Hsp70 protein was found to be essential for
spermatogenic cell development especially during the meiotic phase of spermatogenesis
as male mice defective for the hsp70 gene are infertile (Eddy, 1999). Indeed, Hsp70 is
required for the assembly of the maturation promoting factor which is needed for the
G2-M transition in the primary spermatocytes. In this review, it was also specified that
homologues of Hsp70 are present in the testes of mammals, birds, amphibians and
fishes, suggesting that the role of this spermatogenic cell chaperone is conserved across
phyla. In Drosophila, it seems that other molecular chaperones (Hsp23, Hsp27, Hsp60
and Hsp90) are involved in the spermatogenesis process (Michaud et al., 1997; Yue et
al., 1999; Timakov and Zhang, 2001). In this invertebrate, the synthesis of Hsp70 in
testes is observed only under heat shock conditions (Michaud et al., 1997).
Alternatively, the finding of Hsp70-like proteins in the chaetognath spermatocytes
supports the necessity of this protein during the spermatogenesis as described for
vertebrates (Eddy, 1999).
4.2. Wounded specimens
4.2.1. The cerebral nervous system
Immediately after the tail section and during the wound healing process, the expression
pattern of Hsp70 was modified, not only at wound level but also in the cerebral nervous

S. Jean et al. / J. Exp. Mar. Biol. Ecol. 312 (2004) 319–332328
system. In the cerebral nervous system, the expression of Hsp70 was early and transiently
observed in the neuropile and later in some cell bodies of the cerebral ganglion. Therefore,
it can be assumed that the cerebral ganglion is involved in the regulation of the response to
the injury. This phenomenon may reveal an intensification of the neuronal activity, on the
one hand at the level of the synapses in the neuropile and, on the other hand, by an
activation of cell signaling pathways in which the formation of protein complexes and
their regulation are Hsp70-dependent. The hood, the mouth and the neck plexuses were
also immunoreactive according to time from half an hour to 3 days after the injury. These
plexus networks present numerous varicosities where Hsp70 may be over expressed in
wounded animals. However, the reasons why these plexuses, which innerve the mouth and
other head elements, are immunoreactive remain enigmatic in the wounded animals. In this
context, the labeling of the tail plexus that is frequently observed in this experiment is
more comprehensive because the plexus is partly damaged by the tail section, and so,
Hsp70 are needed to avoid protein breakdown and allow the folding of newly synthesized
proteins.
4.2.2. The tail region
Chaetognaths are built around a large hydroskeleton that is limited by a strong
connective tissue, the basement membrane, which is covered outside by a stratified
epidermis. On the internal face of the basement membrane, four large longitudinal muscle
quadrants in the trunk and in the tail maintain the body architecture and allow
locomotion. In the tail region, the transverse section of the tail affects the body
organization of chaetognaths. Immediately after the cut, a spot-like Hsp70 immunor-
eactivity was observed at the edges of the wound. The superficial localization of this
immunostaining suggests that the induction of Hsp70 occurs in epidermal cells where
these proteins play their role of molecular chaperones and mediate the cellular protection.
The wound opens the body cavity to the external environment, so animals must face
rapidly to preserve their binternal milieuQ and to survive. The sequential wound healing of
beheaded S. cephaloptera has been analyzed at the histological and the ultrastructural
levels (Duvert et al., 2000). This study has shown that, first the muscles contract rapidly
to physically close the wound and isolate the general cavity from the seawater. Then, the
muscular layer of the body wall is reshaped to close up the wound whilst a bclotQ, ahypothesized gel-like substance, is produced to increase the mechanical resistance of the
body wall. In the present study, an Hsp70-immunoreactivity in circulating spermatocytes
is observed as early as 30 min after the section of the tail region (data not shown)
indicating that the wound closure has occurred subsequently to muscular contraction.
Besides, a large and unlabeled area is also frequently visible on both side of the
longitudinal caudal septum slightly anteriorly to the wound (Fig. 4H). This dark region
may correspond to the bclotQ situated in the aqueous phase of the general cavity. This plug
does not exhibit a cellular/tissular organisation as demonstrated by Duvert et al. (2000).
Therefore, the cytosolic Hsp70 protein is not expressed in the body regions where the
bclotQ is present.The fibres of the muscular apparatus are strongly Hsp70 immunoreactive during 24 h
in wounded animals as compared with intact specimens where this typical labeling has
never been observed. This early and remarkable induction of Hsp70 expression in the

S. Jean et al. / J. Exp. Mar. Biol. Ecol. 312 (2004) 319–332 329
contractile apparatus may be achieved by a wide range of elaborate regulatory strategies
at both transcriptional and post-transcriptional levels as described in Drosophila
melanogaster and mammals (Huang et al., 2001; Kaarniranta et al., 1998). The
production of Hsp70 in skeletal muscles of mammals has been observed during and after
a high contractile activity (Neufer et al., 1998). The Hsp70 proteins are supposed to
influence the excitation–contraction coupling process, either directly or indirectly in
these muscles (Nosek et al., 2000). However, little is known about the relationships
between the level of expressed Hsp70 within skeletal muscles and their functions, in
particular concerning their response to the occurrence of the damaged myofibres and to
an acute contraction. High-intensity muscular contraction has been shown to produce
oxidative stress and to generate damaged proteins (Essig and Nosek, 1997). Therefore,
the increase of Hsp70 synthesis may prevent these detrimental effects of fatiguing
stimulation and may enhance an adaptation to stress (Liu et al., 2004; McArdle et al.,
2004). Finally, these proteins have also been detected during skeletal muscle
regeneration suggesting an additional function in the remodeling response of muscle
tissue (Duguez et al., 2003). Thus, in chaetognaths, it can be assumed that the presence
of Hsp70 in muscle fibres a few minutes after the tail section until 24 h may result from
the multiple processes mentioned above in relation with their contraction and
regeneration.
Five days after amputation, as the wound healing was fulfilled, two labeled areas
were visible on the lateral posterior edge of the scar in one of the specimens. The
location and shape of the labeled areas, and the delay after which the Hsp70 appears are
in agreement with the results of Duvert et al. (2000). These authors have described the
formation of the seminal vesicles in the place of the observed immunoreactive areas a
week after the tail cutting. These structures were not detected in the present study when
the chaetognaths were observed under a transmitted light microscope. Therefore, it can
be assumed that Hsp70 expression precedes the formation and the maturation of the
seminal vesicles of the injured specimen cut off above the vesicles. Although the
biological mechanism is unknown, Hsp70 may be involved in the regulation of these
phenomena.
It can be concluded that the multiple functions of the constitutive form of Hsp70,
already described in diverse organisms, are illustrated here in the Chaetognatha. These
functions imply several specific co-chaperones, also highly conserved, that participate
in vesicle secretion and recycling, protein transport, and in the regulation of the
assembly and/or disassembly of protein complexes as reviewed by Young et al.
(2003). Besides, the present study demonstrates that the considerable power of healing
of Chaetognatha requires both the constitutive and inducible forms of Hsp70. These
chaperone proteins contribute, at least in part, to restore the body wall allowing the
survival of the wounded animal. Moreover, they are involved in the sperm maturation
and in the seminal vesicle formation, i.e. two major steps of the reproductive process
of the species. The chaetognaths have already been recognized as a model animal in
phylogeny (Casanova et al., 2001), owing to both molecular data that place them at
the root of the triploblastic radiation (Telford and Holland, 1993, 1997; Papillon et
al., 2003) and their simple body plan permitting the easy observation of gene markers
(Yasuda et al., 1997; Takada et al., 2002). The present study shows that the

S. Jean et al. / J. Exp. Mar. Biol. Ecol. 312 (2004) 319–332330
chaetognath could also be, when associated with Hsp70 expression, a good model
as an indicator of abiotic and biotic stress responses at the level of the whole
organism.
Acknowledgements
The authors are indebted to Dr. J.-P. Casanova, who introduced us to the biology of
chaetognaths, and to whom this article is dedicated. We thank him for his valuable
comments and his critical suggestions for the manuscript. [SS]
References
Bloch, M., Johnson, K.A., 1995. Identification of a molecular chaperone in the eukaryotic flagellum and its
localization to the site of microtubule assembly. J. Cell. Sci. 108, 3541–3545.
Bone, Q., Goto, T., 1991. The nervous system. In: Bone, Q., Kapp, H., Pierrot-Bults, A.C. (Eds.), The Biology of
Chaetognaths. Oxford University Press, New York, pp. 18–31.
Bronk, P., Wenniger, J.J., Dawson-Scully, K., Guo, X., Hong, S., Atwood, H.L., Zinsmaier, K.E., 2001.
Drosophila Hsc70-4 is critical for neurotransmitter exocytosis in vivo. Neuron 30, 475–488.
Bukau, B., Horwich, A., 1998. The Hsp70 and Hsp60 chaperone machines. Cell 92, 351–366.
Burfield, S.T., 1927. Sagitta. L.M.B.C. Memoirs, 28, 1–104, 12 pls.
Casanova, J.-P., Duvert, M., Perez, Y., 2001. Phylogenetic interest of the bchaetognath modelQ. Mesogee 59,
27–31.
Duguez, S., Bihan, M.C., Gouttefangeas, D., Feasson, L., Freyssenet, D., 2003. Myogenic and nonmyogenic cells
differentially express proteinases, Hsc/Hsp70, and BAG-1 during skeletal muscle regeneration. Am. J.
Physiol: Endocrinol. Metab. 285, E206–E215.
Duvert, M., Savineau, J.-P., Campistron, G., Onteniente, B., 1997. Distribution and role of aspartate in the
nervous system of the chaetognath Sagitta. J. Comp. Neurol. 380, 485–494.
Duvert, M., Perez, Y., Casanova, J.-P., 2000. Wound healing and survival of beheaded chaetognaths. J. Mar. Biol.
Assoc. UK 80, 891–898.
Eddy, E.M., 1999. Role of heat shock protein Hsp70-2 in spermatogenesis. Rev. Reprod. 4, 23–30.
Essig, D.A., Nosek, T.M., 1997. Muscle fatigue and induction of stress protein genes: a dual function of reactive
oxygen species? Can. J. Appl. Physiol. 22, 409–428.
Feder, M.E., Hofmann, G.E., 1999. Heat-shock proteins, molecular chaperones, and the stress response:
evolutionary and ecological physiology. Annu. Rev. Physiol. 61, 243–282.
Feigenbaum, D., 1991. Food and feeding behaviour. In: Bone, Q., Kapp, H., Pierrot-Bults, A.C. (Eds.), The
Biology of Chaetognaths. Oxford University Press, New York, pp. 45–54.
Ghirardelli, E., 1968. Some aspects of the biology of the chaetognaths. Adv. Mar. Biol. 6, 271–375.
Goto, T., Katayama-Kumoi, Y., Tohyama, M., Yoshida, M., 1992. Distribution and development of the serotonin-
and RFamide-like immunoreactive neurons in the arrowworm Paraspadella gotoi (Chaetognatha). Cell
Tissue Res. 267, 215–222.
Hartl, F.U., Hayer-Hartl, M., 2002. Molecular chaperones in the cytosol: from nascent chain to folded protein.
Science 295, 1852–1858.
Huang, L., Mivechi, N.F., Moskophidis, D., 2001. Insights into regulation and function of the major stress-
induced hsp70 molecular chaperone in vivo: analysis of mice with targeted gene disruption of the hsp70.1 or
hsp70.3 gene. Mol. Cell. Biol. 21, 8575–8591.
Kaarniranta, K., Elo, M., Sironen, R., Lammi, M.J., Goldring, M.B., Eriksson, J.E., Sistonen, L., Helminen,
H.J., 1998. Hsp70 accumulation in chondrocytic cells exposed to high continuous hydrostatic pressure
coincides with mRNA stabilization rather than transcriptional activation. Proc. Natl. Acad. Sci. U. S. A. 95,
2319–2324.

S. Jean et al. / J. Exp. Mar. Biol. Ecol. 312 (2004) 319–332 331
Kalmar, B., Burnstock, G., Vrbova, G., Urbanics, R., Csermely, P., Greensmith, L., 2002. Upregulation of heat
shock proteins rescues motoneurones from axotomy-induced cell death in neonatal rats. Exp. Neurol. 176,
87–97.
Kiang, J.G., Tsokos, G.C., 1998. Heat shock protein 70 kDa: molecular biology, biochemistry, and physiology.
Pharmacol. Ther. 80, 183–201.
Liu, Y., Lormes, W., Wang, L., Reissnecker, S., Steinacker, J.M., 2004. Different skeletal muscle Hsp70 responses
to high-intensity strength training and low-intensity endurance training. Eur. J. Appl. Physiol. 91, 330–335.
Malakhov, V.V., Frid, M.G., 1984. Structure of the ciliary loop and retrocerebral organ in Sagitta glacialis
(Chaetognatha). Dokl. Akad. Nauk SSSR 277, 763–765.
Mastrogiacomo, A., Parsons, S.M., Zampighi, G.A., Jenden, D.J., Umbach, J.A., Gundersen, C.B., 1994.
Cysteine string proteins: a potential link between synaptic vesicles and presynaptic Ca2+ channels. Science
263, 981–982.
Mayer, M.P., Bukau, B., 1998. Hsp70 chaperone systems: diversity of cellular functions and mechanism of action.
Biol. Chem. 379, 261–268.
McArdle, A., Dillmann, W.H., Mestril, R., Faulkner, J.A., Jackson, M.J., 2004. Overexpression of Hsp70 in
mouse skeletal muscle protects against muscle damage and age-related muscle dysfunction. FASEB J. 18,
355–357.
Michaud, S., Marin, R., Westwood, J.T., Tanguay, R.M., 1997. Cell-specific expression and heat-shock induction
of Hsps during spermatogenesis in Drosophila melanogaster. J. Cell. Sci. 110, 1989–1997.
Morgan, J.R., Prasad, K., Jin, S., Augustine, G.J., Lafer, E.M., 2001. Uncoating of clathrin-coated vesicles in
presynaptic terminals: roles for Hsc70 and auxilin. Neuron 32, 289–300.
Neufer, P.D., Ordway, G.A., Williams, R.S., 1998. Transient regulation of c-fos, alpha B-crystallin, and hsp70 in
muscle during recovery from contractile activity. Am. J. Physiol. 274, 341–346.
Nollen, E.A., Brunsting, J.F., Roelofsen, H., Weber, L.A., Kampinga, H.H., 1999. In vivo chaperone activity of
heat shock protein 70 and thermotolerance. Mol. Cell. Biol. 19, 2069–2079.
Nosek, T.M., Brotto, M.A., Essig, D.A., Mestril, R., Conover, R.C., Dillmann, W.H., Kolbeck, R.C., 2000.
Functional properties of skeletal muscle from transgenic animals with upregulated heat shock protein 70.
Physiol. Genomics 4, 25–33.
Papillon, D., Perez, Y., Fasano, L., Le Parco, Y., Caubit, X., 2003. Hox gene survey in the chaetognath Spadella
cephaloptera: evolutionary implications. Dev. Genes Evol. 213, 142–148.
Pierce, E.L., 1951. The chaetognatha of the west coast of Florida. Biol. Bull. Mar. Biol. Lab., Woods Hole 100,
206–228.
Sloboda, R.D., 2002. A healthy understanding of intraflagellar transport. Cell Motil. Cytoskelet. 52, 1–8.
Stephens, R.E., 1997. Synthesis and turnover of embryonic sea urchin ciliary proteins during selective inhibition
of tubulin synthesis and assembly. Mol. Biol. Cell 8, 2187–2198.
Stephens, R.E., Lemieux, N.A., 1999. Molecular chaperones in cilia and flagella: implications for protein
turnover. Cell Motil. Cytoskelet. 44, 274–283.
Takada, N., Goto, T., Satoh, N., 2002. Expression pattern of the brachyury gene in the arrow worm Paraspadella
gotoi (chaetognatha). Genesis 32, 240–245.
Telford, M.J., Holland, P.W.H., 1993. The phylogenetic affinities of the chaetognaths: a molecular analysis. Mol.
Biol. Evol. 10, 660–676.
Telford, M.J., Holland, P.W., 1997. Evolution of 28S ribosomal DNA in chaetognaths: duplicate genes and
molecular phylogeny. J. Mol. Evol. 44, 135–144.
Timakov, B., Zhang, P., 2001. The hsp60B gene of Drosophila melanogaster is essential for the spermatid
individualization process. Cell Stress Chaperones 6, 71–77.
Welsh, U., Storch, V., 1983. Fine structural and enzyme histochemical observations on the epidermis and the
sensory cells of Sagitta elegans (chaetognata). Zool. Anz. 210, 34–43.
Williams, N.E., Nelsen, E.M., 1997. Hsp70 and Hsp90 homologs are associated with tubulin in hetero-oligomeric
complexes, cilia and the cortex of Tetrahymena. J. Cell. Sci. 110, 1665–1672.
Yasuda, E., Goto, T., Makabe, K.W., Satoh, N., 1997. Expression of actin genes in the arrow worm Paraspadella
gotoi (chaetognatha). Zool. Sci. 14, 953–960.
Young, J.C., Barral, J.M., Ulrich Hartl, F., 2003. More than folding: localized functions of cytosolic chaperones.
Trends Biochem. Sci. 28, 541–547.

S. Jean et al. / J. Exp. Mar. Biol. Ecol. 312 (2004) 319–332332
Yue, L., Karr, T.L., Nathan, D.F., Swift, H., Srinivasan, S., Lindquist, S., 1999. Genetic analysis of viable Hsp90
alleles reveals a critical role in Drosophila spermatogenesis. Genetics 151, 1065–1079.
Zinsmaier, K.E., Bronk, P., 2001. Molecular chaperones and the regulation of neurotransmitter exocytosis.
Biochem. Pharmacol. 62, 1–11.
Zinsmaier, K.E., Eberle, K.K., Buchner, E., Walter, N., Benzer, S., 1994. Paralysis and early death in cysteine
string protein mutants of Drosophila. Science 263, 977–980.





























