Embed Size (px)
Citation preview

Brain, Behavior, and Immunity 15, 227–234 (2001)
doi:10.1006/brbi.2000.0594, available online at http://www.idealibrary.com on
Changes in Immune Cell Counts and Interleukin (IL)-1β Productionin Humans after a Somnogenically Active Growth Hormone-
Releasing Hormone (GHRH) Administration
Lisa Marshall,* Boris Perras,† Horst L. Fehm,† and Jan Born*
*Clinical Neuroendocrinology and †Department of Internal Medicine, Medical University ofLuebeck, D-23538 Luebeck, Germany
Growth hormone-releasing hormone (GHRH) has been shown to enhance slow-wavesleep (SWS) and non-rapid eye movement sleep in animals and humans. In animals thesomnogenic effect of interleukin (IL)-1β appears to be mediated by GHRH. Neuroimmuno-logical interactions in sleep are most frequently studied in humans by sleep deprivationor by cytokine administration. The present study, in contrast, investigates in humans theeffect of enhanced sleep through GHRH administration on selected immune parameters.Results reveal that a single intravenous bolus of 50 µg GHRH which enhanced SWS stage4 in the first half of the night suppressed circulating suppressor T cell (CD31/CD81)numbers, with a similar tendency for B cells (CD191) and suppressed mitogen-stimulatedIL-1β production. When the same amount of GHRH was administered distributed acrossfive repetitive boluses of 10 µg GHRH within 1 h, neither corresponding sleep nor immuneparameters were changed significantly compared to placebo. These data suggest that GHRHcan modulate immune functions through brain mechanisms which are also involved in theregulation of sleep. 2001 Academic Press
Key Words: growth hormone-releasing hormone (GHRH); non-rapid eye movement(NREM) sleep; slow-wave sleep (SWS); immune cells; cytokines; interleukin (IL)-1β.
INTRODUCTION
Sleep is defined by changes in the brain’s electroencephalographic activity and isaccompanied by specific endocrine and immunological regulatory mechanisms. Amost prominent factor of sleep is a sharp rise in activation of the growth hormone-releasing hormone–growth hormone (GHRH-GH) somatotropic axis occurring inconjunction with pronounced phases of slow-wave sleep (SWS) in the beginning ofsleep. This activation includes the hypothalamic release of GHRH which also stimu-lates the release of GH from the pituitary. In humans and animals GHRH administra-tion has been shown to increase both SWS and GH release (reviewed in van Cauter,Plat, & Copinschi, 1998). The present study represents an attempt to clarify by theadministration of GHRH whether this neuroendocrine pattern (in the beginning ofsleep) has particular regulatory functions on immune parameters. This approach con-trasts with previous studies in which immunological functions of sleep were investi-gated primarily using sleep deprivation. Sleep deprivation has been found to enhancethe mitogen-stimulated production of interleukin (IL)-1β and TNFα (Born & Hansen,1997; Spath-Schwalbe, Porzsolt, Born, & Fehm, 1992; Uthgenannt, Schoolmann, Pie-trowsky, Fehm, & Born, 1995), whereby effects of sleep deprivation on plasma IL-1were less consistent (Moldofsky, Lue, Eisen, Keystone, & Gorczynski, 1986; Moldof-sky, Lue, Davidson, & Gorczynski, 1989; Dinges, Douglas, Hamarman, Zaugg, &Kapoor, 1995). IL-2 and the ability of T cells to produce IL-2, on the other hand,were reduced after sleep deprivation (Uthgenannt et al., 1995; Irwin, McClintick,
227
0889-1593/01 $35.00Copyright 2001 by Academic Press
All rights of reproduction in any form reserved.

228 MARSHALL ET AL.
Costlow, Fortner, White, & Gillin, 1996; Born, Lange, Hansen, Molle, & Fehm,1997). Effects of cytokines on the central nervous sleep process have also been inves-tigated in humans (Born & Spath-Schwalbe, 1997); however, cytokine administrationin healthy subjects is limited. Thus, the present study investigates immunologicaleffects after enhancing specific sleep-associated neuroendocrine activity, by GHRHadministration. The question is, to what extent GHRH induced alterations are oppo-site to those after sleep deprivation.
METHODS
Subjects and Procedure
Data of 12 healthy nonsmoking men (aged 22 to 36 years), free of medication,entered the study. Four subjects of the initial 16 were excluded on grounds of non-compliance or incomplete immunological data. A regular nighttime/sleep–daytime/wake schedule was maintained prior to participation. On the days before experimentalnights subjects were instructed to get up before 0800 h, not to take any naps, andnot to participate in any exerting physical activity. Studies took place in a sleeplaboratory at the Medical University of Lubeck, and the experimental protocol wasapproved by the University’s Ethics Committee.
On experimental nights subjects were assigned to bed at 2000 h and remainedin bed until 0900 h the next morning. Polysomnographic recordings were obtainedcontinuously from 2300 h (lights off) until 0700 h (awakening). Each subject partici-pated in three experimental nights, separated by at least 7 days according to a double-blind within-subject crossover design. On one night subjects received a single 50-µg GHRH (Ferring GmbH, Kiel, Germany) bolus at 2200 h: ‘‘Single GHRH.’’ Onanother night the same total dose of GHRH was given as five intravenous bolusesof 10 µg GHRH at 2145, 2200, 2215, 2230, and 2245 h: ‘‘Repetitive GHRH.’’ Thelatter mode of GHRH administration was introduced with the intent of obtaininga differential effect on central nervous sleep coincident with comparable pituitarystimulation of GH release. On the placebo night saline solution (0.9% NaCl) wasinjected. For blood sampling and substance administration, respectively, indwellingcatheters were inserted in antecubital veins of the forearm. Peripheral effects ofGHRH administration were also assessed by measuring serum insulin-like growthfactor (IGF)-1 concentration.
Data Reduction and Analysis
Blood cell counts and cytokines (IL-1β, IL-2, IL-6) and IGF-I serum concentrationwere determined in samples drawn at 2100, 2400, 0300, 0600, and 0900 h. Bloodsamples for plasma GH and cortisol analyses were drawn at 15- and 30-min intervals,respectively. Samples for blood cell count analysis were collected in EDTA tubes andstained at 1000 h on the following day. Leukocyte and erythrocyte counts, leukocytedifferential counts, platelets, and hematocrit were determined automatically by aTechnicon counter (Technicon H3 RTC, Bayer, Germany). Lymphocyte subsets weredetermined by flow cytometry (FACScan, Becton Dickinson, San Jose, CA) fromthe same samples: T cells (CD31; Leu-4/FITC), B cells (CD191; Leu-12/PE), Thelper cells (CD41; Leu-3a/FITC), T suppressor cells (CD31/CD81; Leu-2a/PE),activated T cells (anti-HLA-DR1/PE), and natural killer cells (CD31/CD161;Leu-4/11c/FITC and CD561; Leu-19/PE). Measurement of activated T cells ex-

SYSTEMIC GHRH ON THE IMMUNE SYSTEM 229
cluded B cell and monocyte populations. Antibodies were purchased from BectonDickinson (Heidelberg, Germany).
Production of IL-1β and IL-2 were assessed after in vitro mitogen stimulation sinceplasma concentrations are normally close to the detection threshold. Aliquots of 50µl of blood were resuspended under laminar air flow in 400 µl of RPMI 1640 medium(containing 2 mM glutamine, 100 U/ml penicillin, and 100 µg/ml streptomycin; Ser-omed, Berlin, Germany). For stimulation of IL-1β 0.5 µg of the mitogen lipopolysac-charide (Sigma Chemical Co., St. Louis, MO) from Escherichia coli was added anddissolved in 50 µl of a medium containing 80% RPMI and 20% sterile water (1 µg/ml). For stimulation of IL-2, 2.5 µg phytohemagglutinin (Murex, Dartford, UK) wasadded and dissolved in 50 µl of a medium containing 50% RPMI and 50% sterilewater (5 µg/ml). Samples were stimulated in duplicate and incubated for 48 h at37°C with 5% CO2 in humidified air. Supernatants were harvested and stored at270°C until assay. Cytokine levels were measured by enzyme immunoassay kits(R&D Systems, Minneapolis, MN). Assay sensitivities were 0.3 pg/ml for IL-1β,6.0 pg/ml for IL-2, and 0.094 pg/ml for IL-6. The intra- and interassay coefficientsof variations were all ,10%. The concentrations of GH and cortisol in plasma weremeasured by a commercial radioimmunoassay (DPC Biermann, Bad Nauheim, Ger-many) and enzyme immunoassay kits (Boeringer Mannheim, Immunodiagnostica,Mannheim, Germany), respectively. IGF-I serum concentrations were measured by acommercial enzyme immunoassay (Diagnostic Systems Laboratories, Webster, TX);intra- and inter-assay coefficients of variation were each below 7%). Polysomno-graphic data were analyzed according to the criteria of Rechtschaffen and Kales(1968).
Statistical analysis relied on analysis of covariance variance (ANCOVA) with arepeated measures factor representing the three experimental conditions. Values at2100 h served as covariate. Pairwise contrasts specified significant treatment effects.Results are given in means 6 SEM. A p value ,0.05 was considered significant.
RESULTS
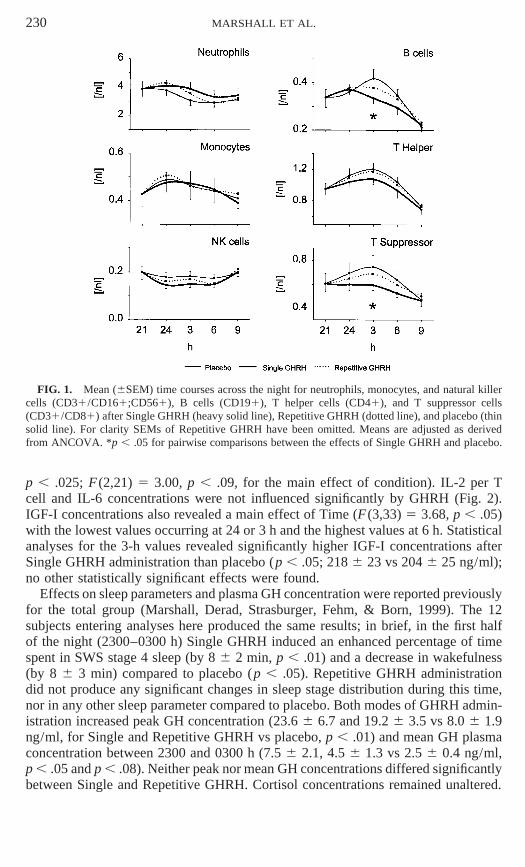
Across the night typical changes in the numbers of leukocytes, erythrocytes, neu-trophilic granulocytes, lymphocytes, and monocytes; in hemoglobin and hematocrit;and in all investigated cell subsets were found reflecting part of the circadian rhythm(p , 0.05). Figure 1 depicts the time courses for selected cell types and significanteffects of treatment. Effects on immune cells were most pronounced at 0300 h. Sig-nificance was reached at this time for T suppressor cells (F (2,21) 5 6.24, p , .01;F (2,21) 5 3.80, p , .05, for the main effect across all times) and B cell and totalT cell numbers (F (2,21) 5 4.30, p , .05 and F (2,21) 5 3.62, p , .05, respectively).The single GHRH bolus significantly reduced the number of T suppressor cells at0300 h compared to placebo (p , .025). Pairwise comparisons of B cells again indi-cated reduced cell numbers after Single GHRH compared to placebo (p 5 .05). ForT cells a corresponding suppression after Single GHRH approached significance(p , .08). Counts of neutrophils, monocytes, and natural killer cells were not in-fluenced by GHRH.
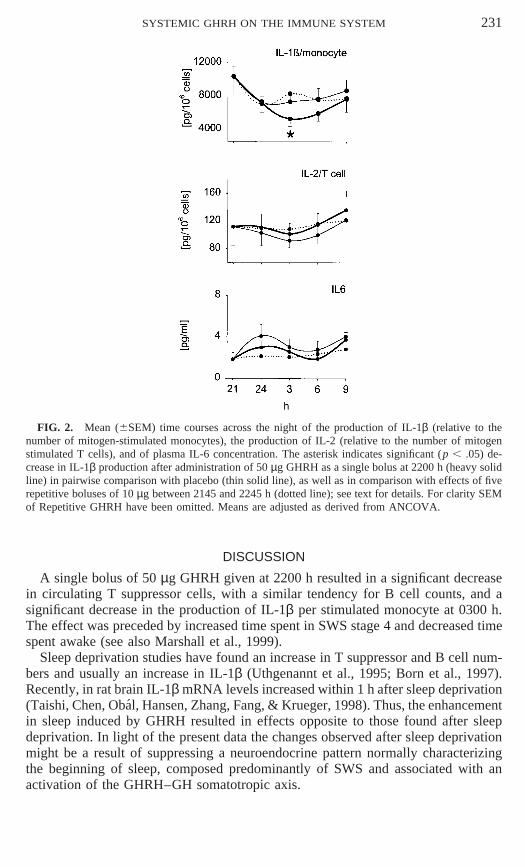
Concurrently, with the suppression of cell counts, Single GHRH decreased produc-tion of IL-1β per monocyte; again the focus of this effect was at 0300 h (Fig. 2,p , .025, in pairwise comparison with placebo). The decrease in IL-1β after thesingle bolus also reached significance when compared to Repetitive GHRH (Fig. 2,
230 MARSHALL ET AL.
FIG. 1. Mean (6SEM) time courses across the night for neutrophils, monocytes, and natural killercells (CD31/CD161;CD561), B cells (CD191), T helper cells (CD41), and T suppressor cells(CD31/CD81) after Single GHRH (heavy solid line), Repetitive GHRH (dotted line), and placebo (thinsolid line). For clarity SEMs of Repetitive GHRH have been omitted. Means are adjusted as derivedfrom ANCOVA. *p , .05 for pairwise comparisons between the effects of Single GHRH and placebo.
p , .025; F (2,21) 5 3.00, p , .09, for the main effect of condition). IL-2 per Tcell and IL-6 concentrations were not influenced significantly by GHRH (Fig. 2).IGF-I concentrations also revealed a main effect of Time (F (3,33) 5 3.68, p , .05)with the lowest values occurring at 24 or 3 h and the highest values at 6 h. Statisticalanalyses for the 3-h values revealed significantly higher IGF-I concentrations afterSingle GHRH administration than placebo (p , .05; 218 6 23 vs 204 6 25 ng/ml);no other statistically significant effects were found.
Effects on sleep parameters and plasma GH concentration were reported previouslyfor the total group (Marshall, Derad, Strasburger, Fehm, & Born, 1999). The 12subjects entering analyses here produced the same results; in brief, in the first halfof the night (2300–0300 h) Single GHRH induced an enhanced percentage of timespent in SWS stage 4 sleep (by 8 6 2 min, p , .01) and a decrease in wakefulness(by 8 6 3 min) compared to placebo (p , .05). Repetitive GHRH administrationdid not produce any significant changes in sleep stage distribution during this time,nor in any other sleep parameter compared to placebo. Both modes of GHRH admin-istration increased peak GH concentration (23.6 6 6.7 and 19.2 6 3.5 vs 8.0 6 1.9ng/ml, for Single and Repetitive GHRH vs placebo, p , .01) and mean GH plasmaconcentration between 2300 and 0300 h (7.5 6 2.1, 4.5 6 1.3 vs 2.5 6 0.4 ng/ml,p , .05 and p , .08). Neither peak nor mean GH concentrations differed significantlybetween Single and Repetitive GHRH. Cortisol concentrations remained unaltered.
SYSTEMIC GHRH ON THE IMMUNE SYSTEM 231
FIG. 2. Mean (6SEM) time courses across the night of the production of IL-1β (relative to thenumber of mitogen-stimulated monocytes), the production of IL-2 (relative to the number of mitogenstimulated T cells), and of plasma IL-6 concentration. The asterisk indicates significant (p , .05) de-crease in IL-1β production after administration of 50 µg GHRH as a single bolus at 2200 h (heavy solidline) in pairwise comparison with placebo (thin solid line), as well as in comparison with effects of fiverepetitive boluses of 10 µg between 2145 and 2245 h (dotted line); see text for details. For clarity SEMof Repetitive GHRH have been omitted. Means are adjusted as derived from ANCOVA.
DISCUSSION
A single bolus of 50 µg GHRH given at 2200 h resulted in a significant decreasein circulating T suppressor cells, with a similar tendency for B cell counts, and asignificant decrease in the production of IL-1β per stimulated monocyte at 0300 h.The effect was preceded by increased time spent in SWS stage 4 and decreased timespent awake (see also Marshall et al., 1999).
Sleep deprivation studies have found an increase in T suppressor and B cell num-bers and usually an increase in IL-1β (Uthgenannt et al., 1995; Born et al., 1997).Recently, in rat brain IL-1β mRNA levels increased within 1 h after sleep deprivation(Taishi, Chen, Obal, Hansen, Zhang, Fang, & Krueger, 1998). Thus, the enhancementin sleep induced by GHRH resulted in effects opposite to those found after sleepdeprivation. In light of the present data the changes observed after sleep deprivationmight be a result of suppressing a neuroendocrine pattern normally characterizingthe beginning of sleep, composed predominantly of SWS and associated with anactivation of the GHRH–GH somatotropic axis.

232 MARSHALL ET AL.
However, it is also necessary to discuss possible effects of GHRH on immuneparameters as a consequence of a direct peripheral action merely coinciding with thecentral nervous effect on sleep. First, any effect of cortisol, mediating activity withinthe hypothalamic–pituitary–adrenocortical axis, can be excluded since mean concen-trations were the same for all conditions. Second, mediation of effects by a directstimulatory action on pituitary GH release also appears to be unlikely, consideringa comparable GH enhancement after both single bolus and repetitive GHRH adminis-trations. Moreover, GH at concentrations in vitro and in vivo comparable with thoseobserved here had no effect on the production of IL-1β (Kappel, Hansen, Diamant,Jorgensen, Gyhrs, & Pedersen, 1993; Kappel, Hansen, Diamant, & Pedersen, 1994).In another study serum IL-1β was increased 4 h after subcutaneous GH injection inGH-deficient children (Bozzola, De Amici, Zecca, Schimpff, & Rapaport, 1998). Ingeneral, if effective at all, many studies support a stimulatory rather than an inhibi-tory immunomodulatory effect of GH and IGF-I on production and release of IL-1β(Kelley, 1989; Wiedermann, Reinisch, & Braunsteiner, 1993; Murphy, Rui, & Longo,1995; Buul-Offers & Kooijman, 1998). Also, it should be noted that no significantchanges were found between conditions in circulating monocyte counts which repre-sent the main source of blood-borne IL-1β. Third, however, a direct effect of GHRHon circulating immune cells cannot be excluded. Mononuclear leukocytes possessspecific receptors for GHRH (Guarcello, Weigent, & Blalock, 1991), and GHRH hasbeen reported to modulate lymphocyte proliferation and natural killer cell activity(Weigent & Blalock, 1994). Furthermore, GHRH can induce increased plasma levelsof substances such as insulin, glucagon, and somatostatin which have been shownto influence cytokine production and to be entrained to the ultradian sleep cycle(Campos & Baumann, 1992; Kern, Offenheuser, Born & Fehm, 1996; Peluso, Petillo,Melone, Mazzarella, Ranieri & Tajana, 1996). The possible role of these factors inthe effects induced specifically by the single bolus administration of GHRH in thepresent study needs to be investigated systematically. Whether GHRH per se affectsproduction of IL-1 by monocytes is not known, to the best of our knowledge.
Considering that joint changes in immune parameters and SWS were selectivelyobserved after the single bolus of GHRH while after repetitive administration ofsmaller boluses neither sleep nor immune parameters were altered significantly, themost likely explanation for the suppressive action of GHRH on suppressor T and Bcell counts and on IL-1β production appears to be an effect of the peptide on centralnervous mechanisms. Such brain mechanisms might be commonly involved in pro-moting SWS and regulatory peripheral immune functions. This view is further sup-ported by the fact that immunological changes were most pronounced immediatelyfollowing the time range of distinctly enhanced SWS. The greater efficacy of a highbolus of GHRH than repetitive administration of smaller boluses has been previouslyexplained by the higher peak GHRH plasma level obtained after the single boluswhich facilitates passage of the peptide into the brain/cerebrospinal fluid by crossingthe blood–brain barrier or via circumventricular organs (Marshall et al., 1999).
Findings by Krueger and co-workers should be pointed out in this context whichreveal pronounced somnogenic effects of IL-1β in rats mediated by activation ofhypothalamic GHRH release (see Krueger, Obal, & Fang, 1999). On this backgroundthe present findings of an enhanced SWS stage 4 activity after GHRH followed bydecreased IL-1β production could hint at a negative feedback action of GHRH onmonokine production when the propensity for SWS dissipates.

SYSTEMIC GHRH ON THE IMMUNE SYSTEM 233
ACKNOWLEDGMENTS
We are very grateful to Nicole Boes and Thorsten Spielvogel for data collection and to SteffanieBaxmann, Ann-Kristin Jurs, and Anja Otterbein for further technical assistance. This study was supportedby a grant from the Deutsche Forschungsgemeinschaft to H.L.F. and J.B.
REFERENCES
Born, J., & Spath-Schwalbe, E. (1997). Effects of cytokines on human sleep. In G. Wieselmann (Ed.),Current update in psychoimmunology: Key topics in brain research, pp. 103–118. Springer-Verlag,Wien.
Born, J., Lange, T., Hansen, K., Molle, M., & Fehm, H. L. (1997). Effects of sleep and circadian rhythmon human circulating immune cells. J. Immunol. 158, 4454–4464.
Born, J., & Hansen, K. (1997). Dependence of human cytokine production and mononuclear cell subsetcounts on circadian rhythm and sleep. In A. E. Henneberg & W. P. Kaschka (Eds.), Immunologicalalterations in psychiatric diseases, Adv. Biol. Psychiatry, Vol. 18, pp. 18–31. Karger: Basel.
Bozzola, M., De Amici, M., Zecca, M., Schimpff, R. M., & Rapaport, R. (1998). Modulating effect ofhuman growth hormone on tumour necrosis factor-alpha and interleukin-1beta. Eur. J. Endocrinol.138, 640–643.
Buul-Offers, S. C., & Kooijman, R. (1998). The role of growth hormone and insulin-like growth factorsin the immune system. Cell. Mol. Life Sci. 54, 1083–1094.
Campos, S. P., & Baumann, H. (1992). Insulin is a prominent modulator of the cytokine-stimulatedexpression of acute-phase plasma protein genes. Mol. Cell. Biol. 12, 1789–1797.
Dinges, D. F., Douglas, S. D., Hamarman, S., Zaugg, L., & Kapoor, S. (1995). Sleep deprivation andhuman immune function. Adv. Neuroimmunol. 5, 97–110.
Guarcello, V., Weigent, D. A., & Blalock, J. E. (1991). Growth hormone releasing hormone receptorson thymocytes and splenocytes from rats. Cell. Immunol. 136, 291–302.
Irwin, M., McClintick, J., Costlow, C., Fortner, M., White, J., & Gillin, J. C. (1996). Partial night sleepdeprivation reduces natural killer and cellular immune responses in humans. FASEB J. 10, 643–653.
Kappel, M., Hansen, M. B., Diamant, M., Jorgensen, J. O. L., Gyhrs, A., & Pedersen, B. K. (1993).Effects of an acute bolus growth hormone infusion on the human immune system. Horm. Metab.Res. 25, 579–585.
Kappel, M., Hansen, M. B., Diamant, M., & Pedersen, B. K. (1994). In vitro effects of human growthhormone on the proliferative responses and cytokine production of blood mononuclear cells. Horm.Metab. Res. 26, 612–614.
Kelley, K. W. (1989). Growth hormone, lymphocytes and macrophages. Biochem. Pharmacol. 38, 705–713.
Kern, W., Offenheuser, S., Born, J., & Fehm, H. L. (1996). Entrainment of ultradian oscillations in thesecretion of insulin and glucagon to the nonrapid eye movement/rapid eye movement sleep rhythmin humans. J. Clin. Endocrinol. Metab. 81, 1541–1547.
Krueger, J. M., Obal, F., Jr., & Fang, J. (1999). Humoral regulation of physiological sleep: Cytokinesand GHRH. J. Sleep Res. 8(Suppl. 1), 53–59.
Marshall, L., Derad, I., Strasburger, C. J., Fehm, H. L., & Born, J. (1999). A determinant factor in theefficacy of GHRH administration in promoting sleep: high peak concentration versus recurrentincreasing slopes. Psychoneuroendocrinology 24, 363–370.
Moldofsky, H., Lue, F. A., Eisen, J., Keystone, E., & Gorczynski, R. M. (1986). The relationship ofinterleukin-1 and immune functions to sleep in humans. Psychosom. Med. 48, 309–318.
Moldofsky, H., Lue, F., Davidson, J., & Gorczynski, R. (1989). Effects of sleep deprivation on humanimmune functions. FASEB J. 3, 1972–1977.
Murphy, W. J., Rui, H., & Longo, D. L. (1995). Effects of growth hormone and prolactin immunedevelopment and function. Life Sci. 57, 1–14.
Peluso, G., Petillo, O., Melone, M. A., Mazzarella, G., Ranieri, M., & Tajana, G. F. (1996). Modulation

234 MARSHALL ET AL.
of cytokine production in activated human monocytes by somatostatin. Neuropeptides 30, 443–451.
Rechtschaffen, A., & Kales, A. (1968). A manual of standardized terminology, techniques and scoringsystem for sleep stages of human subjects, NIH Publication 204. U.S. Govt. Printing Office: Wash-ington, DC.
Spath-Schwalbe, E., Porzsolt, F., Born, J., & Fehm, H. L. (1992). Differences in stimulated cytokinerelease between sleep and sleep deprivation. In M. Freund, H. Link, R. Schmidt, & K. Welte. (Eds.),Cytokines in hemopoiesis, oncology, and AIDS, II, pp. 457–463. Springer-Verlag, Berlin.
Taishi, P., Chen, Z., Obal, F., Jr, Hansen, M. K., Zhang, J., Fang, J., & Krueger, J. M. (1998). Sleep-associated changes in interleukin-1beta mRNA in the brain. J. Interferon Cytokine. Res. 18, 793–798.
Uthgenannt, D., Schoolmann, D., Pietrowsky, R., Fehm, H. L., & Born, J. (1995). Effects of sleep onthe production of cytokines in humans. Psychosom. Med. 57, 97–104.
Van Cauter, E., Plat, L., & Copinschi, G. (1998). Interrelations between sleep and the somatotropic axis.Sleep 21, 553–566.
Weigent, D. A., & Blalock, J. E. (1994). Role of neuropeptides in the bidirectional communicationbetween the immune and neuroendocrine systems. In B. Scharrer, E. M. Smith, & G. B. Stefano(Eds.), Neuropeptides and immunoregulation, pp. 14–27. Springer-Verlag, Berlin.
Wiedermann, C. J., Reinisch, N., & Braunsteiner, H. (1993). Stimulation of monocyte chemotaxis byhuman growth hormone and its deactivation by somatostatin. Blood 82, 954–960.
Received September 28, 1999; published online February 2, 2001





























