Embed Size (px)
Citation preview

32
CHAPTER 1
Collection, extraction of compounds and evaluation of bioactivity profile of
chosen marine sponge and holothuria
l.1. INTRODUCTION
Sponges
Marine sponges are the simplest of the multicellular invertebrate animals. Sponges are
exclusively aquatic and found in the deepest oceans to the edge of the sea and survive in a variety
of circumstances including places where there is scarce light as well as in cold or warm water.
Approximately 5000 different species of sponges are recorded all over the world. Among them
150 occur in freshwater and 17 sponges are of commercial importance. In India a total of 486
species of sponges have been identified. In the Gulf of Mannar and Palk Bay (India) 275 species
of sponges have been recorded (Thomas et al., 1998).
The rapid development of the pharmaceutical market has brought about a bloom of
information regarding various metabolites native to the sponges. Recent technologies introduced
to produce novel products from marine sponges have fantastic contribution to human healthcare
products. Some of the purified natural products from marine sponges exhibit antitumour, antiviral
and pharmacological activities. Over 60.0% of potentially useful bioactive compounds discovered
so far from living organisms have been obtained from marine fauna, of them 70% are reported
from sponges (Johannes et al. 2011). Apoptosis inducing compounds were isolated from marine
sponge have the potential to be developed into targeted cancer therapies. There are numerous
cytotoxic metabolites that have been isolated from marine sponge. Cytotoxic activity is regarded
as the first indicator in identifying anticancer drugs. However, increasing evidence has shown that
cell death can be induced via three different mechanisms: apoptosis, autophagy and oncosis (Wei
et al., 2010). Marine sponges are sources of numerous novel sterols, particularly in terms of
unique side chain structures such as those with high degree of alkylation and unusual function
groups. Steroids with a cyclopropane ring at C-25 and C-26 are relatively rare and first reported

33
from the sponges, namely Petrosia ficiformis2 and Halichondria sp. Some of these analogs
exhibit biological activities including anti-inflammation and cytotoxicity (Bakkestuen et al.,
2005). The sponges of the genus Ianthella (order Verongida, family Ianthellidae) contained
several bromotyrosine-derived metabolites, such as the ianthesines, purealin and others, which
were mainly isolated from their polar extracts (Ciminiello et al., 1996).
The different species of Aplysina (Demospongiae) can be distinguished by the lack of
terpenes, the presence of carotenoids and by a series of brominated compounds derived from
tyrosine and of sterols in moderately high content (Erpenbeck et al., 2002). The
bromometabolites, which are considered chemical markers of this order, have generally been
extracted with low-polarity solvents and are soluble in either dichloromethane or chloroform.
Some of these compounds present in Aplysina sp. possessed in vitro cytotoxic activities against
human tumor cells (Okamoto et al ., 2000). Oxozoline derivatives with cytotoxic activities have
been found in Aplysina collected on the Virgin Islands (Friedrich et al., 2001). Among these 2-
oxazoline derivatives are also bromotyrosine compounds known as fistularin-1 and -2, and a bis-
2-oxazoline derivative containing a dibromophenol ring. These substances have also been isolated
from other Caribbean sponges (Hentschel et al., 2002). Other more polar and nonbrominated
secondary metabolites, such as aplysillin A and some tryptophan-derived substances have been
isolated from Aplysina species. Bromotyrosine metabolites have represented one-fourth of the
crude extract of the Mediterranean sponge, Aplysina aerophoba. A novel acidic glycogen was
isolated from the Brazilian sponge Alysina fulva (Rodriguez and Pina, 1993).
Holothurians
The marine holothurians are spiny skinned invertebrates, which form important
commercial group among the echinoderms. Holothurians have been harvested for years from the
near shore areas in temperate and tropical seas for the preparation of highly priced product beche-
de –mer. In India, about 200 species of holothurians are known of which 75 species are from
shallow waters within 20m depths. They are considered to be good sources of novel metabolites

34
exhibiting different biological activities. They contain antimicrobial and antitumour activities
(Cuvillier, 2002). According to James (1994) a great potential exists for the extraction of valuable
bioactive compounds from the sea cucumbers in the Indian coast. A new immunomodulatory lead
Cumaside is a complex of monosulfated triterpene glycosides from the sea cucumber Cucumaria
japonica which possesses cytotoxic activity against Ehrlich carcinoma cells. Cumaside has an
antitumor activity against different forms of experimental mouse Ehrlich carcinoma in vivo both
independently and in combination with cytostatics. The highest effect occured at a treatment once
a day for 7 days before the tumour inoculation followed by Cumaside treatment once a day for 7
days. Prophylactic treatment with Cumaside and subsequent therapeutic application of 5-
fluorouracil suppressed the tumor growth by 43.0%. (Aminin et al., 2010). Sphingoid base
composition of cerebrosides from sea cucumber Stichopus variegates exhibited cytotoxicity
against human colon cancer cell and induced apoptosis. They are major constituents of c-17,c-19
alkyl chain and 1-3 double bonds (Sugwara et al., 2006).
In the present study, attempts were made to find out the bioactive compounds from marine
invertebrates such as the marine sponge and holothuria. The sponge Sigmadocia pumila ledenfeld
was collected from Kanyakumari coast. The species is widely distributed from the Red sea to the
Pacific Ocean through the Indian Ocean. The other sample used for the present work was the sea
cucumber Holothuria atra (The Black Sea Cucumber). The characteristic feature of this species is
a long black body with a few spots. The species was collected from the sandy bottom and often
partially covered with sand particles. The active bioactive compounds were biochemically
characterized and compounds were identified.

35
1.2. MATERIALS AND METHODS
1.2.1. Collection, identification and extract preparation from the marine sponge, Sigmadocia
pumila
Specimens of the marine sponge Sigmadocia pumila were collected from the coast of
Kanyakumari by “by-catch method” during active fishing season as per the method of Selvin and
Lipton (2004). The sponges entangled in the fishing nets were detached at a depth ranging from
10 to 25m while fishing and were segregated after completion of fishing in the morning hours.
They were examined for attached algae and other organisms and they were carefully removed.
Details of colour, shape, texture, residency, form and other characteristic features were noted
during the time of collection. The sponge of interest S. pumila was transferred directly to new
wide mouth plastic containers containing sea water to prevent contact of sponge tissue with air.
Another set required for bioactivity studies was transferred to bottles containing methanol. The
details of taxonomic scientific classification of the marine sponge S. pumila are given below:
Sponges
Phylum : Porifera Grant
Class : Demospongiae Sollas
Order : Haplosclerida Topsent
Family : Adociidae de Laubenfels
Genus : Sigmadocia de Laubenfels
Species : pumila (Ledenfeld)
Sigmadocia pumila
For the preparation of crude extracts, the method of Bergman et al. (1992) was followed.
Sponges weighing 250g were cut into small pieces and extracted thrice with distilled methanol
and the pooled organic solution was filtered through Whatman No.1 filter paper fitted in a
Buchner funnel using suction. Solvents were removed by rotary vacuum evaporator (Buchi-type)
Fig.1 Marine Sponge
Sigmadocia pumila

36
under reduced pressure so as to get the crude methanol extract. The concentrated crude extract
was collected in airtight plastic containers and kept in the refrigerator for further use.
1.2.2. Collection, identification and extract preparation of Sea cucumber (Holothuria atra )
Holothuria atra specimens with a size range of 10 to 30 cm in length and 30 to 180 g
weight were collected from fishing nets operated off Kanyakumari (8° 03' and 8° 35' of the north
Latitudes and 77° 15' and 77° 36' of the east longitudes).
The details of taxonomic Scientific classification of the Sea cucumber Holothuria atra are given
below
Sea cucumber
Phylum : Echinodermata
Class : Holothuroidea
Order : Aspidochirotida
Family : Holothuriidae
Genus : Holothuria
Species : atra
Holothuria atra
Extract preparation from sea cucumbers:
Immediately upon collection, Holothuria atra specimens were dissected to remove the
internal organs and packed using ice and kept at –80°C for extraction. The skin portion was
peeled off and stored in methanol in separate containers. The biologically active compounds were
extracted based on their polarity using methanol an organic solvent as per the method given by
Rashid et al. (2001) with appropriate modifications. About 200g of frozen samples were
homogenized with deionized water and methanol. The mixture was continuously stirred in the
dark at 4°C for 24 h. Then it was centrifuged at 5000 rpm for 15 minutes. The supernatant was
collected and filtered. The extracts thus collected were freeze-dried and kept at -80 °C, while the
Sea Cucumber
Holothuria atra

37
insoluble solid materials were re-extracted with methanol (100%). The organic extracts were
combined and the solvents were removed by rotary evaporation at 40°C under low pressure to
avoid degradation of compounds.
1.2.3. Antibacterial activity:
The antibacterial activities of the methanol extracts of sponge Sigmadocia pumila and sea
cucumber Holothuria atra were determined by the standard agar well diffusion assay using
Muller Hinton agar (Hi Media) (Perez et al ., 1990). The bacterial cultures were obtained from the
microbial type culture collection and gene bank (MTCC), Institute for Microbial Technology,
Chandigarh, India. The Gram positive bacterial strains included: Bacillus thuringiensis MTCC
4714, Enterococci faecalis MTCC 439, Listeria monocytogenes MTCC 1143, Staphylococcus
aureus MTCC 737 and Proteus vulgaris MTCC 426. The Gram negative strains such as the
E.coli MTCC 443, Klebsiella pneumonia MTCC 109, Pseudomonas putida MTCC 1688 and
Serratia liquefaciens MTCC 3039 were used in the test. Wells of 6.0 mm diameter were punched
using sterilized cork borer. Cultures of each microbial pathogenic strain were swabbed with sterile
cotton on the surface of medium. The crude extracts were suspended in methanol at
concentrations (1, 5, and 10 %) of 1mg/ml for antimicrobial studies. Sponge Sigmadocia pumila
and sea cucumber Holothuria atra extracts were tested with different aliquots 50,100,150 µl in
each well. The plates were incubated for 24 hrs at 370C and solvent control was performed in each
case. Areas of inhibited microbial growth were observed as clear zone around the well after 24
hours in bacteria. Antimicrobial activity was measured as diameter of zone of inhibition,
excluding the well diameter (Li et al., 2005).
1.2.4. Antifungal activity:
Antifungal activity of the crude extract of marine sponge Sigmadocia pumila and sea
cucumber Holothuria atra was determined by using the standard method. Fungal isolates were
obtained from the microbial type culture collection and gene bank (MTCC), Institute for
Microbial Technology, Chandigarh, India. Isolates such as Trichoderma viride MTCC 2047,

38
Fuzarium sp MTCC 284, Aspergillus niger MTCC 1344, Candida albicans MTCC 183,
Penicillium chrysogenum MTCC 5108 and Aspergillus flavis MTCC 873 were used for the
experiment. The fungal cultures were maintained in 0.2% dextrose medium at 5.6 pH and the
optical density 0.10 at 530 nm was adjusted using spectrophotometer. Each fungal inoculum was
applied on plate and evenly spread on Potato dextrose agar using a sterile swab. To each well, 100
µl of the samples were added and the solvent was used as the control. Agar diffusion assay was
followed to evaluate the antimicrobial activity. The Petri plates were incubated at 30°C for 2 days.
At the end of the 48 hrs, inhibition zones formed in the medium were measured in millimeters
(Selvin and Lipton, 2001).
1.2.5. Larvicidal activity:
The larvicidal activity of methanolic extracts of marine sponge and sea cucumber
species was evaluated against the third instar stage of Aedes egyptii. The egg cards of Aedes
egyptii were obtained from the Centre for Research in Medical Entomology, Indian Council of
Medical Research, Madurai. The susceptibility or resistance of the mosquito larvae to the selected
concentrations of the extracts was carried by adopting standard bioassay protocol given by WHO
(1981). The crude extract was tested to determine the larval bio-efficacy by diluting the original
extract to 1.0 to 5.5% levels. The bioassays were performed at a room temperature of 27 ± 1°C by
exposing 20 larvae in a final volume of 250 ml water in 500 ml glass beaker with minimum of
four replicates for each concentration. Simultaneously, control groups were also maintained in
beakers without addition of extract but with the solvent alone. Observations were made after 12,
24, 36, 48, and 60 hrs of treatment for larvicidal activity. Based on the percentage of mortality the
LC50 values of the extracts were determined using Probit Scale Analysis (Newman and Crag,
2004).
1.2.6. Brine shrimp cytotoxicity:
The brine shrimp (Artemiasalina) lethality assay was performed as described by Meyer
et al. (1982). For the bioassay, the dried cysts of 1g were allowed to hatch in a beaker filled with

39
filtered seawater (32 ppt) under constant aeration. After 48h, the phototropic brown coloured
nauplii were siphoned out using a glass pipette. The stock extracts were prepared at the
concentration of 5mg/ml. The nauplii were counted against an illuminated background and ten
nauplii were transferred to each cavity cup containing 2 ml of filtered seawater dissolved with
varying dilutions of sponge and the sea cucumber extract ranging from 0.1% to 1% from the
stock. To ensure that the mortality observed in the bioassay could be attributed to bioactive
compounds and not due to starvation, control was maintained without adding the sponge extract.
The larvae were not fed during the experiment, as the newly hatched brine shrimp nauplii can
survive for up to 48 h without food due to the presence of yolk-sac (Lewis, 1995). The cavity cups
were maintained under constant illumination. Experiments were carried out in triplicates to get
statistically significant results and the mean value was recorded as mortality after 24 h. Larvae
were considered dead if they did not exhibit any internal or external movement during several
seconds of observation. Based on the percent mortality, LC50 value of the extract was determined
using the probit scale analysis.
1.2.7. Hemolytic activity:
The hemolytic activity of the crude methanolic extracts of marine sponge S. pumila and
sea cucumber species H. atra on human red blood cells (RBC) was tested by a micro-hemolytic
method given by Chen et al. (1997). Human “B” positive blood was obtained, from the Vivek
Institute of Laboratory Medicine, Nagercoil, (Kanyakumari District, Tamilnadu) in EDTA
solution (2.7 g in 100 ml of distilled water) as an anticoagulant at 5% of the blood volume and
brought to the laboratory. Then 1% erythrocyte suspension was prepared by adding 99 ml of PBS,
pH 7.4, to 1 ml of packed RBC. The micro-hemolytic test was performed on 96-well U-bottom
microtiter plates. Serial two fold dilutions of the extracts were carried out in 100 µl of PBS, pH
7.4. For the control set 100µl of distilled water was added, to 3% RBC suspension. This served as
the positive control and an equal amount of PBS, pH 7.4, acted as the negative control. The plate
was gently shaken and allowed to stand for two hours at room temperature. Uniform red colour

40
suspension in the wells was considered positive hemolysis while sedimentation on the bottom was
considered lack of hemolysis. The percentage of the hemolytic cells was calculated using the
formula
% Hemolysis = Absorbance of sample –Absorbance of blank X 100
Highest Absorbance for positive control
1.2.8. Nitro Blue Tetrazoliun assay (NBT assay)
The invitro immunomodulation study was done on both the crude methanolic extracts of S.
pumila and H. atra . Phagocytic index (PI) in percentage was assessed by NBT reduction assay as
per the method described by Park et al., (1968). In five test tubes the reaction mixture consisted
of leucocytes suspension (0.4ml), endotoxin activated plasma as standard (0.1ml) PBS solution
at 0.1ml was used as control. To each test tube 0.1 ml different concentrations of test samples
(25, 50, 100, 200 μg/ml) were added and 5%s NBT solution (0.8ml) was added and incubated at
370 C in water bath with shaker for 30 minutes and reaction was stopped with cold PBS (Ph 7.2).
Then it was centrifuged at 1000 rpm for five minutes. After discarding the supernatant, a drop of
PBS was added and gently the cells were re suspended. A drop of this reaction mixture was
spread on a clean glass slide,dried and fixed in methanol for two minutes, stained with 0.8%
aqueous safranin for 2 minutes. The smear was washed, dried and mounted. NBT Positive cells
represented by dark blue colour were counted under 100X in a Nikon (E200) binocular
microscope.

41
1.3. RESULTS
Antibacterial activity
The trend of antibacterial activity is given in Fig 1.1. In the agar well diffusion method,
the methanol extracts of Sigmadocia pumila exhibited high inhibitory activity towards human
pathogens of Gram negative organisms such as E. coli, Klebsiella pneumoniae, Pseudomonas
putida, Serratia liquefaciens and Staphylococcus aureus. They were not active against the Gram
positive organisms. The Fig 1.2. represents the effect of antibacterial activity of the methanolic
crude extracts of the sea cucumber Holothuria atra. It showed effective inhibitory action against
the Gram negative organisms such as Klebsiella pneumoniae, Serratia liquefaciens, E. coli, and
Staphylococcus aureus and in Gram positive organism such as Listeria monocytogenes. The
remaining organisms were found to be less active.
Table 1.1 denotes the inhibitory zone diameter formed in the Gram negative organisms
using the Sigmadocia pumila crude methanolic extracts diluted at various concentrations. In
E.coli the zone formed at 50µl was 2mm and at 150 µl it was 16mm diameter. Klebsiella
Pneumoniae formed 14mm zone at 100 µl concentration of extract, Pseudomonas putida formed
the zone of 15mm at 100 µl of extract. In Serratia liquefaciens 16mm zone was formed at 50 µl
concentration of the extract. In Staphylococcus aureus 2mm and 12mm diameter zones were
formed at 50 and 100 µl of extract. In the rest of the concentrations the extract was not effective.
Table 1.2.states the inhibitory zones formed in the Gram positive and Gram negative
organisms using the methanolic crude extracts of Holothuria atra at various dilutions. In Listeria
monocytogenes the zone of inhibition formed at 150 µl was 14mm. In E.coli the zone formed at
50µl was 2mm and at 100 µl it was 16mm diameter. Klebsiella Pneumoniae formed the zone of
3mm and 14 mm diameter size at the concentrations of 50 and 100 µl. Serratia liquefaciens
showed the zone of inhibition as 12mm and 4mm diameter size in 100 and 150 µl concentrations.

42
Finally in Staphylococcus aureus the zone of inhibition found at 100 and 150 µl was 5mm and
16mm diameter size.
Antifungal activity
The results of anifungal activity of the methanolic crude extracts of the sea cucumber H.
atra are summarized in Fig 1.3. It showed effective antifungal property. It was found to be
effective on various fungal organisms such as Trichoderma viride, Aspergillus niger, Candida
albicans, Penicillium chrysogenum and Aspergillus flavis. Data given in Fig 1.4 suggest the
antifungal activity of the methanolic extratcts of S. pumila showing moderate effects on some
organisms such as Candida albicans and Penicillium chrysogenum and no activity was seen in
organisms such as Trichoderma viride, Aspergillus niger, Aspergillus flavis and Fusarium. In
Candida albicans the diameter of the zone was 6mm at 50 µl and 10mm at 100 µl. In Penicillium
chrysogenum the inhibitory zone measured was 3mm and 6mm for 50 and 100 µl.
In respect of the methanol extracts of sea cucumber H. atra, effective antifungal activity
was recorded towards Trichoderma viride, Aspergillus niger and Aspergillus flavis with diameter
of 16, 14 and 12 mm respectively in Table 1.3. In Candida albicans and Penicillium chrysogenum
the inhibitory zone diameter was 18 and 14 mm at 100 µl concentration of extract.
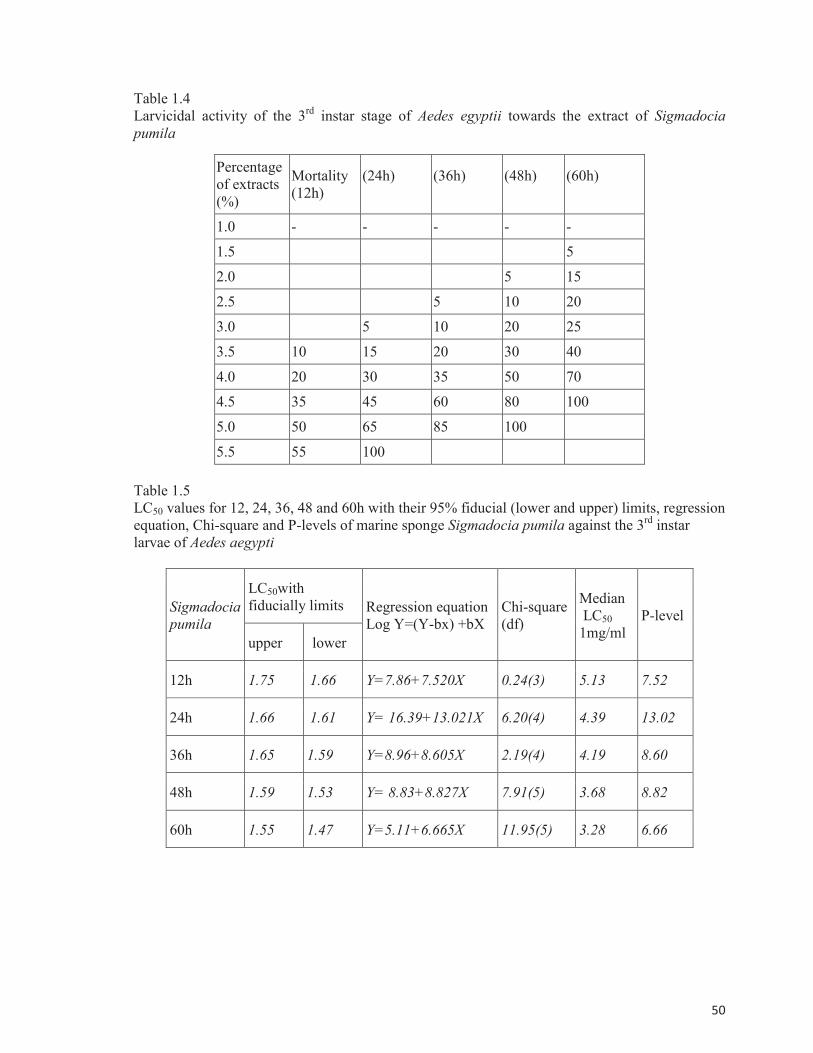
Larvicidal activity
Larvicidal potentials using the extracts of marine sponge S. pumila based on the mortality
of the third instar stage of Aedes egyptii are depicted in Table 1.4. The mortality rate was
calculated at different intervals of time from 12, 24, 36, 48 and 60 h. The sponge extracts
produced 100% mortality during 24 h at 5.5% concentration of the extracts and 100% mortality
achieved during 48h at 5.5% concentration of extracts and 100% mortality was obtained at 60 h
with 4.5% level of the extracts The results of LC50 values which were evaluated based on the
potency of the extracts at various concentrations form 1.0 to 5.5% and at different intervals of
time from 12, 24, 36, 48 and 60 h using the mortality rate against the third instar stage of Aedes
aegypti are presented in Table 1.5.

43
The larvicidal effect of the extracts of sea cucumber Holothuria atra based on the
mortality of the third instar stage of Aedes egyptii is given in Table 1.6. The mortality rate was
calculated at different intervals of time from 12, 24, 36, 48 and 60 h. The extracts produced 100%
mortality during 24 h at 5.0% concentration of the extracts and 100% mortality achieved during
48 h at 4.5% concentration of extracts and 100% mortality was obtained at 60 h with 4.0 % conc
of the extracts. Table 1.7 shows the LC50 values calculated based on the potency of the extracts at
various concentrations form 1.0% to 5.5% and at different intervals of time such as 12, 24, 36, 48
and 60 h using the mortality rate against the third instar stage of Aedes aegypti. The medium level
of larvicidal effects revealed that the bioactive sponge crude extracts possessed highly significant
larvicidal activities with LC50 value of 0.43% in 24 h. It was also observed that the crude extracts
of the sea cucumber H. atra also had medium level of larvicidal effects with less toxic effects
with LC50 value of 0.58% in 24h with less minimum effects compared to sponges.
Determination of brine shrimp cytotoxicity range
The results from cytotoxicity studies showed the presence of bioactive compounds in the
tested sponge extracts and sea cucumber extracts. The crude extracts of the sponge S. pumila
exhibited high toxicity against Artemia salina nauplii followed with the moderate toxic effect of
sea cucumber H. atra. Results of Artemia cytotoxicity bioassay of marine sponge S. pumila are
depicted in Table 1.8. The sponge extracts were added at various concentrations of 0.1, 0.2, 0.4,
0.6, 0.8 and 1.0%. Results indicated 50% mortality at 0.4% concentration of the extracts and
100% mortality happened at the higher dose of 10.0% of the extracts during 24 h interval.
Results given in Table 1.9 indicate that the extracts of sea cucumber H. atra were active
against the Artemia naupli and showed cytotoxicity profile of various concentrations at 0.1, 0.2,
0.4, 0.6, 0.8, and 1.0%. Mortality increased linearly with increase of concentration of every
extract in 24 h. The median lethal dose indicated that the sponge S. pumila produced 50%
mortality at 0.45% concentration. The LC50 values of the sea cucumber H. atra showed the toxic
effect at 0.52% concentration.

44
Table 1.10 shows the hemolytic activity using the methanolic crude extracts of S. pumila.
The optical density value for the extract at (5mg/ml) concentration was 0.492. The calculated
hemolytic activity was 96.47%.
Strong hemolytic activity of H. atra in its crude methanolic extract is described in Table
1.11 .The optical density value of the sample was 0.550 at the highest concentration of extract of
1mg/ml. The positive hemolysis was measured by uniform red colour suspension and a button
formation at the bottom of wells showed negative hemolysis. The calculated hemolytic activity
was 100%.
A comparative analysis of the hemolytic activity of S. pumila and H. atra is depicted in
Fig 1.5. Though both samples had hemolytic activity, maximum effect was shown by Holothuria
atra.
In the in vitro model of Qualitative NBT test significant immunostimulant activity of the
sponge S. pumila was found at a dose of 50% (Table 1.12). The methanolic fractions of the
sponge had significantly increased the phagocytic function of human neutrophils, when compared
with control and increased the movement of neutrophils towards the foreign body which is the
most important step in the phagocytosis. The effect of the H. atra found to be moderate. In this, it
showed less effect at higher concentrations. The comparative analysis of the immuno modulatory
activity invitro of both the crude extracts of marine sponge S. pumila and sea cucumber H. atra is
depicted in Fig 1.6. The graph showed maximum phagocytic activity in the extracts of the sponge
S. pumila.
The presence of NBT positive cells at 100x magnification in blue colour was due to the
effect of the methanolic crude extracts of marine sponge S. pumila (Fig 1.7). It was assumed that
the sponge extracts had significant immunomodulatory activity.

45
a.Escherichia coli b. Klebsiella pneumoniae
c.Pseudomonas putida d. Serratia liquefaciens
e. Staphylococcus aureus
Fig 1.1. (a-e) Antibacterial activity of the methanolic crude extracts of the sponge Sigmadocia
pumila in well diffusion

46
a. Escherichia coli b. Klebsiella pneumoniae
c.Listeria monocytogenes d. Serratia liquefaciens
e. Staphylococcus aureus
Fig 1.2. (a-e) Antibacterial activity of the methanolic crude extracts of the Sea cucumber
Holothuria atra in well diffusion

47
a.Aspergillus flavis b. Aspergilus niger
c. Trichoderma viride d. Candida albicans
e. Penicillium chrysogenum
Fig 1.3. (a-e) Antifungal activity of the methanolic crude extracts of the Sea cucumber Holothuria
atra

48
a. Candida albicans b. Penicillium chrysogenum
Fig 1.4. (a and b) Antifungal activity of the methanolic crude extracts of the marine sponge
Sigmadocia pumila

49
Table 1.1
Antibacterial activities in the sponge Sigmadocia pumila extract (Agar well diffusion method)
Bacterial strains
Inhibition zone diameter (in mm)
1%
50µl
5%
100 µl
10%
150 µl
E. coli 2 NA 16
Klebsiella pneumonia NA 14 NA
Pseudomonas putida NA 15 NA
Serratia liquefaciens 16 NA NA
Staphylococcus aureus 2 12 NA
Table 1.2
Antibacterial activities in Sea cucumber Holothuria atra extract (Agar well diffusion method)
Bacterial strains
Inhibition Zone diameter (in mm)
1%
50µl
5%
100 µl
10%
150 µl
Listeria monocytogenes NA NA 14
E. coli 2 16 NA
Klebsiella pneumonia 3 14 NA
Serratia liquefaciens NA 12 4
Staphylococcus aureus NA 5 16
NA: No Activity
Table 1.3
Antifungal activity in Holothuria atra (well diffusion method)
Fungal strains Inhibition Zone diameter
(in mm)
Aspergillu niger 14
Aspergillus flavis 12
Trichoderma viride 16
Fusarium spp NA
Candida albicans 18
Penicillium chrysogenum 14
NA: No Activity
50
Table 1.4
Larvicidal activity of the 3rd
instar stage of Aedes egyptii towards the extract of Sigmadocia
pumila
Percentage
of extracts
(%)
Mortality
(12h)
(24h)
(36h)
(48h)
(60h)
1.0 - - - - -
1.5 5
2.0 5 15
2.5 5 10 20
3.0 5 10 20 25
3.5 10 15 20 30 40
4.0 20 30 35 50 70
4.5 35 45 60 80 100
5.0 50 65 85 100
5.5 55 100
Table 1.5
LC50 values for 12, 24, 36, 48 and 60h with their 95% fiducial (lower and upper) limits, regression
equation, Chi-square and P-levels of marine sponge Sigmadocia pumila against the 3rd
instar
larvae of Aedes aegypti
Sigmadocia
pumila
LC50with
fiducially limits Regression equation
Log Y=(Y-bx) +bX
Chi-square
(df)
Median
LC50
1mg/ml
P-level
upper lower
12h 1.75 1.66 Y=7.86+7.520X 0.24(3) 5.13 7.52
24h 1.66 1.61 Y= 16.39+13.021X 6.20(4) 4.39 13.02
36h 1.65 1.59 Y=8.96+8.605X 2.19(4) 4.19 8.60
48h 1.59 1.53 Y= 8.83+8.827X 7.91(5) 3.68 8.82
60h 1.55 1.47 Y=5.11+6.665X 11.95(5) 3.28 6.66

51
Table 1.6
Larvicidal activity of the 3rd
instar stage of Aedes egyptii towards the extract of sea cucumber
Holothuria atra
Percentage
of extract
(%)
Mortality
(12h)
(24h)
(36h)
(48h)
(60h)
0.5
1.0 5
1.5 5 15
2.0 5 20 40
2.5 5 10 30 45
3.0 10 15 20 45 60
3.5 30 35 45 65 80
4.0 35 45 60 80 100
4.5 50 65 85 100
5.0 65 100
Table 1.7
LC50 values for 12, 24, 36, 48 and 60h with their 95% fiducial (lower and upper) limits, regression
equation, Chi-square and P-levels of Sea cucumber Holothuria atra against the 3rd
instar larvae
of Aedes aegypti
Holothuria
atra
LC50with
fiducial limits Regression equation
Log Y=(Y-bx) +bX
Chi-square
(df)
Median
LC50
1mg/ml
P-level
upper lower
12h 1.68 1.60 Y=6.53 +7.006X 0.60(3) 4.42 7.0
24h 1.61 1.55 Y=12.95+11.313X 6.43(4) 3.86 11.31
36h 1.59 1.52 Y=6.65+7.474X 1.71(4) 3.61 7.47
48h 1.50 1.42 Y=5.40+7.106X 4.46(5) 2.90 7.10
60h 1.42 1.32 Y=2.73+5.614X 5.76(5) 2.38 5.61

52
Table 1.8
Mortality of brine shrimp exposed for 24 h to different concentrations of methanol extract from
sponge Sigmadocia pumila (± represents standard deviation)
Conc of extracts (%) Mortality (24 h) (%)
Control Nil
0.1 20 ±0.10
0.2 30 ±0.24
0.4 50 ±1.52
0.6 70 ±2.20
0.8 90 ±3.16
1.0 100 ±0.0
Table 1.9
Mortality of brine shrimp exposed for 24 h to different concentrations of methanol extract from
Sea cucumber Holothuria atra (± represents standard deviation)
Conc of extracts (%) Mortality (24 h) (%)
Control Nil
0.1 10 ±0.12
0.2 30 ±1.30
0.4 50 ±1.72
0.6 70 ±2.16
0.8 80 ±2.34
1.0 98 ±0.0
LC50 (% extract) value of methanolic crude extracts to Artemia salina
Samples LC50 ( in % extract)
Sigmadocia pumila 0.45
Holothuria atra 0.52

53
Ab
sorb
ance
at
54
1n
m
mm
Table 1.10
Hemolytic activity of Sigmadocia pumila
Sigmadocia pumila Absorbance at 541nm
Blank 0.164
Sample (1mg/ml) 0.492
Positive control 0.340
Negative control 0.265
% Hemolysis = 96.47%
Table 1.11
Hemolytic activity of Holothuria atra
Holothuria atra jaeger Absorbance at 541nm
Blank 0.152
Sample (5mg/ml) 0.550
Positive control 0.340
Negative control 0.242
% Hemolysis = 100.75%
Fig.1.5. Hemolytic activity using the methanolic crude extracts of Sigmadocia pumila and
Holothuria atra

54
Per
cen
tag
e N
BT
Po
siti
ve
Cel
ls
Table 1.12
Effect of the methanolic crude extracts of Sigmadocia pumila and Holothuria atra jaeger using
qualitative nitroblue tetrazolium (NBT) test
Groups Conc. of extract
1mg/ml
% NBT positive
cells
Control 23
Endotoxin activated
plasma (standard)
10% 85
Sigmadocia pumila 10%
25%
50%
50%
35%
51%
63%
72%
Holothuria atra 10%
25%
50%
50%
26%
34%
46%
52%
Fig.1.6.Percentage of positive cells obtained from NBT test using the methanolic extracts of
Sigmadocia pumila and Holothuria atra
0
10
20
30
40
50
60
70
80
90
100
Standard Sigmadocia pumila Holothuria atra control

55
Fig 1.7. NBT Positive cells in the extract of Sigmadocia pumila (100x magnification)

56
1.4. DISCUSSION
The Bioactivity studies of S. pumila and H. atra extracts were evaluated through means of
in vitro experiments. A large number of compounds exhibiting antimicrobial activity with
potential biomedical application have been isolated from marine sponges. Organic extracts
tended to be more active than the aqueous extracts. The strongest antimicrobial activity was found
in the organic extracts from the sponges of the genus Ircinia and Verongula (Martinez et al.,
1997).
Antibacterial activity
The sponge Sigmadocia pumila belongs to the class Demospongiae. It showed effective
antibacterial activity against the Gram negative organisms. The sponge class Demospongiae is
known for producing the largest number and diversity of secondary metabolites isolated from
marine invertebrates. There is a worldwide interest in marine natural products as one of the few de
novo sources of drug discovery (McConnell et al., 1994). In the present study, the extracts of the
sponge S. pumila and H. atra showed antimicrobial action against the Gram negative organisms
effectively. Organisms such as E.coli, Pseudomonas putida and Staphylococcus aureus were
inhibited effectively by the extracts. Similar results were reported by Manilal et al., (2010).
Marine sponges form close associations with a wide variety of microorganisms which act as a rich
source of biologically active secondary metabolites. Sponges associated Streptomyces were
cultivable to develop marine-derived products are a promising source for the discovery of novel
bioactive compounds.
S. pumila collected from the south east coast of India showed antimicrobial properties. It
could be due to the presence of active compounds such as terpenes, sterols, phenols etc. Similarly
the crude extracts of Caribbean sponges Ircinia campassa, Verongula rigida, Agelas conifera and
Discus oxeata were found to have ichthyotoxic, cytotoxic and antimicrobial activities (Zea et al.,
1986).

57
The present study showed that the methanolic extracts of the sponge S. pumila showed
significant active results against the Escherichia coli and Serratia liquefaciens. The sponge
species of Clathria vulpina showed better antibacterial activity against pathogens particularly
Bacillus subtilis and Vibrio harveyi. It was confirmed that S. pumila had the ability to inhibit the
growth of E. coli and Staphylococcus aureus at lower concentrations of 50 µl and 100 µl of the
extracts (Schroder et al., 2003). S. pumila showed maximum antibacterial activitiy against the
Staphylococcus aureus. This could be due to the presence of aliphatic and aromatic compounds.
Cacospongionolide, B a new sesterpene, isolated from the sponge Lascispongia covernosa
showed antimicrobial activity, brine shrimp cytotoxicity and ichythyotoxicity (De Rosa et al.,
1995).
It was noted that the sponge S. pumila displayed high antibacterial activity against the
Gram negative organisms. S. pumila showed high antibacterial activity than H.atra. All extracts
from Ircinia felix showed considerable antimicrobial activity. The most active samples were the
butanol extracts from Ircinia strobilina, the acetone extract from Verongula gigantea, and the
butanol extract from Verongula rigida. The only extracts that showed considerable activity
towards Gram negative strain were those from Topsentia ophiraphidites and Aplysina archeri.
(Laport et al., 2009). The methanolic extracts of the sponge S. pumila and H. atra were more
active and exhibited antimicrobial and larvicidal activities. From the sponge Dysidea etheria the
antimicrobial activity was studied, against the yeast-Rhodoturula glutinous and fungi- Curvularia
lunata. Furano sesterterpene tetronic acid obtained from the sponge Latrunculia brevis exhibited
the antimicrobial activity (Venkateshwarlu et al., 2001).
The presence of glycosides in H. atra could act as the potent compound to induce the
antimicrobial activities. Similarly Galactocerebrosides in sea cucumbers and their metabolites
have been shown to possess significant functions on promoting the regulation of nerve cell on
regulating protein kinase C activities and on modulating the function of hormone receptors for
several antimicrobial activities (Kariya et al., 2002). The alcoholic extracts of H. atra were

58
inhibitory to human pathogens such as the Klebsiella pneumonia, E.coli, Listeria monocytogenes
and staphylococcus aureus. This could be due to the availability of compounds such as triterpene
glycosides and aglycones. Similarly Sea cucumber (Holothuria scabra) showed strong broad
spectrum antibacterial activity against both Gram positive and Gram negative bacteria such as the
E.coli, Staphylococcus sp and Proteus effectively (Nagaraj et al ., 2008).
The body wall extracts of H. atra showed antibacterial activity which could be due to the
availability of various natural products. It was identified by the agar well diffusion method.
Assay of antibacterial activity with purified lectin and cell free coelomic fluid of sea cucumber,
revealed that it exhibited strong antibacterial activity against both Gram positive and Gram
negative bacteria. It inhibited the growth of Staphylococcus sp and also inhibited effectively the
growth of other Gram negative Serratia sp., Proteus sp., Shigella sp., and E. coli (Tunkijjanukij
and Olafsen, 1998). More than 300 sphingolipids were identified to occur in nature on various
marine organisms while majority of them were reported from marine sponges (Wang et al., 2000).
Antifungal activity
The phenol compound from the marine sponge Dysidea herbacea had significant activity
against the human fungal pathogens Candida albicans and Aspergillus fumigatus which was
compared well with the clinically used antifungal amphotericin B (Sionov et al ., 2005). The S.
pumila extracts showed less antifungal activity than H. atra. It showed effective antifungal
activity against Candida albicans and Penicillium chrysogenum by using agar well diffusion
method.
From the sponge Psammo clemma sp. a potent antifungal agent such as
debromoclathrynamide A and clathrynamide A was obtained. From the sponge of Clathria sp. the
clathrynamide A. compound has significant inhibitory activity against Aspergillus niger and
Aspergillus flavis (Omura et al., 1990). The sponge S. pumila exhibited moderate antifungal
activity against the few organisms which could be due to the presence of compounds such as
carotenoids and alkaloids.

59
The antibacterial and antifungal activities of the S. pumila extracts suggest that it could also
have antitumour property. Marine sponge Aplysina cavernicola produced aeroplysinin and
aerthionin and other dibromo and dichlorotyrosine derivatives, with some antibiotic activity
against Bacillus subtilis and Proteus vulgaris (Pedpradab et al., 2010). From the marine sponge
Clathria sp. clathrinamide and debromoclathrynamide from Psammoclemma sp. acted as potent
antifungal substances (Ohta et al., 1993). The methanolic extract of S. pumila was used to
analyze the antifungal activity. Similarly the methanolic extract of the mairne sponge Haliclona
exigua showed promising antifungal activity against Candida albicans, Cryptococcus neofroman,
Aspergillus fumigatus, and Candida parapsilolis. The marine sponges Amphimedon viridis,
Neopetrosia sp. possessed antileishmanial activity. The Haliclona exigua, was active against
various fungal organisms (Lakshmi et al., 2010).
The methanol extract of S. pumila exhibited antimicrobial property. This could be due to
the presence of methanol which is a high polar solvent. Amade et al. (1982) tested the crude
ethanolic extracts of 15 species from Polynesia and they were tested against bacteria and fungai.
Among these sponges, 8 were completely inactive, 4 were very weak active and 7 were found to
be significantly active against some bacteria and fungi. Dendrilla sp. inhibited the growth of
Neisseria flava and Escherichia coli. Hyrtios eubamma inhibited the growth of Staphylococcus
aureus. The antimicrobial property of S. pumila could be also due to the presence of microbes in
it. As the sponge bioactive compounds could be synthesized due the microbial association.
Bacteria synthesize sponge-specific compounds either completely or in the form of precursors
completed subsequently by sponge metabolism. Antimicrobial and other biological activities of
associated bacteria play a significant ecological role in sponge-bacteria associations (Oclarit et al.,
1994).
The presence of enzymes and compounds such as alkaloids and ligustillides in H. atra
could induce the antimicrobial property. Previous studies reported that lysozyme-like antibacterial
factors against Gram positive bacteria could be wide-spread in the reproductive tissues of

60
echinoderms. These were detected in several other echinoderms such as Cucumaria frondosa
(Benkendor et al., 2001). Hence it is assumed that H. atra could also act as one of the sources
for lysozyme. It could be inferred that the extract from sea cucumber H. atra has got the ability to
induce the antifungal activity. This could be due to the presence of active compounds such as
terpenes, phenoilc compounds and saponins in the methanolic extract of H. atra. Similarly
Patagonicoside a novel antifungal disulfated triterpene glycoside from the sea cucumber Psolus
patagonicus acts against various human fungal pathogens (Haug et al., 2002).
The antifungal activity of the H. atra could be due to the presence of compounds such as
terpenes, alkaloids and saponins. Patagonicoside a bioactive novel antifungal disulphated
triterpene glycoside from the sea cucumber Psolus patagonicus was found to be active against the
pathogenic fungus Cladosporium cucumerinum (Ana et al., 2001). Fucosylated chondroitin
sulfates (FCSs) were isolated from four sea cucumbers Pearsonothuria graeffei, Stichopus
tremulus, Holothuria vagabunda, and Isostichopus badionotus. The detailed sequences of FCSs,
particularly their fucose branches, were characterized and compared (Chena et al., 2011).
The extracts of H. atra were more active against the fungal organisms such as the Aspergillus
niger, Aspergillus flavis and Candida albicans at low concentrations. Similarly the extracts of
Hippospongia communis and Ircinia variabilis have the capacity to inhibit the growth of Candida
tropicalis R2 (an amphotericin B and nystatin resistant strain) as well as other bacteria (Rifai et
al., 2005).
Triterpene glycosides from the sea cucumber Actinopyga lecanora a bioactive compound
showed effective anifungal activity against Aspergillus flavis, Candida albicans, Aspergillus niger
(Kumar et al., 2007). Hence it is proved that the sea cucumber H. atra showed effective
antifungal nature against the organisms such as Aspergillus flavis, Candida albicans, Aspergillus
niger and Penicillium chrysogenum. The antifungal activity of H. atra was found to be maximum
than S. pumila. It was also noted that the antifungal activity was maximum against Trichoderma
viridae and Penicillium chrysogenum. H. atra exhibited more antifungal activity than S. pumila. It

61
was observed from the body wall extracts of H. atra. In the sea cucumber, Cucumaria frondosa
high antibacterial activity was found in the 80% of the extracts obtained from the coelomocytes
and the body wall. It was mainly against the Gram-positive bacteria (Bryan et al., 1994).
Larvicidal activity
The crude extracts of S. pumila showed considerable larvicidal activity against Aedes
egyptii. It was observed from the LC50 values. The crude extracts from Clathria gargonoids and
Callyspongia diffusa have the larvicidal activity against Culex and insecticidal properties.
Sigmodosia carnosa has a higher toxic effect on Aedes egypti. Haliclona pigmentifera, Petrosia
similies are considered as good source, from this novel pestcidal molecules are obtained (Sujatha
and Joseph, 2011). From the current study, it could be inferred that S. pumila possessed highest
larvicidal activity against the 3rd
instar stage of Aedes egyptii and the exhibited high mortality
rate. Mosquitoes such as Aedes aegypti act as vectors of dengue, yellow fever and chikungunya
while Anopheles stephensi and Culex sp act as vectors of malaria and filariasis respectively. Most
of the mosquito control programmes target the larval stage in their breeding sites with larvicides,
as adulticides temporarily reduce the adult population only (Elhag et al., 2001). Reports on
larvicidal activity of sponge extracts are scanty. In India, marine sponges, Dendrilla nigra,
Clathria gorgonoides and Axinella donnani exhibited larvicidal potential against second instar
larvae of Culex sp (Selvin et al., 2004). From the current observation, the methanolic crude
extracts of S. pumila and the H. atra found were to be active marine invertebrates with bioactive
compounds showing larvicidal activities. The mosquito larvicidal activity of the sponge S. pumila
extracts showed the possibilities of reducing the growth of Aedes aegypti. Hence it could be
possible to prevent diseases caused by mosquitoes. Similarly the presence of compounds such as
terpenoids, alkaloids and phenolic compounds could induce the larvicidal activity. Manzamines
are the most promising antimalarial compound that has been discovered in many marine sponges
(Ang et al., 2000). Manzamine-type alkaloids isolated from the Philippine marine sponge
Xestospongia ashmorica showed larvicidal, antibacterial and cytotoxicity activities (Edrada et al.,

62
1996). The presence of larvicidal activity from S. pumila extracts could act as the possible source
for various biological activities. The methanolic crude extracts of the H. atra had larvicidal effect
on Aedes egyptii with the LC50 of 0.52%. Antimicrobial substances possibly the presence of
steroidal sapogenins revealed antibacterial and antifungal activities in the alcoholic extracts of
Holothuria scabra, and Actinopyga echinites. It inhibited the organisms such as the E. coli,
Klebsiella pneumonia, Pseudomonas aeruginosa and Staphylococcus aureus (Fuestani, 2004).
Holothurians contained chondroiton and glucosamine bioactive substances as important cartilage
building blocks with anti-inflammatory, anti-tumour activity and antifungal properties (Hamel
1997). The strong mosquito larvicidal activity was displayed on the body wall methanolic extracts
of Holothuria scabra and Holothuria argus. It was attributed due to the presence of steroids
(Indap and Thakur, 1995).
Brine shrimp cytotoxicity
The cytotoxic activity of the S. pumila extracts against the Artemia salina was found to be
maximum than H. atra. The brine shrimp lethality assay is considered to be one of the most useful
tools for the preliminary assessment of biotoxicity and bioassay with cytotoxic activity against
some human solid tumours. The antitumour activity of cell free extracts from sponge associate
Actinomycetes might be due to the presence of the active secondary metabolites such as alkaloids
and terpenoids (Selvin and Lipton, 2004).
The present investigation indicates that the S. pumila and H. atra crude extracts showed
significant LC50 values of cytotoxic effect against the Artemia salina. Studies made by Zhang et
al. (2003) revealed that more than 10% of the investigated marine sponge species exhibited
cytotoxic activity suggesting production of potential medicines for potent cytotoxic drugs.
Harada et al. (1997) explained that the sponges such as the Thalysias vulpina from the
Pacific ocean showed high activity on the brine shrimp cytotoxicity. The sponge Tedania
anhelans were active against the gram positive and gram negative organisms. S. pumila exhibited
high cytotoxic activity against Artemia salina with the LC50 value of 0.45%. It showed the

63
presence of cytotoxicity of the extracts. The brine shrimp larval mortality assay is widely accepted
as a convenient probe for potential in vitro cytotoxicity and pharmacological activity in marine
natural products (Carballo et al., 2002). The Indonesian sponge, Callyspongia pseudoreticulata
yielded diyne which was found to be toxic in the brine shrimp assay (Braekman et al., 2003).
Sponges of the class Demospongiae are known to produce the largest number and diversity
of secondary metabolites isolated from marine invertebrates, most of them with medically
relevant biological activities and important ecological roles (Faulkner, 2002b). In the present
observation, the S. pumila (Demospongiae) and the sea cucumber H. atra crude methanolic
extracts were active against the Artemia salina showing effective cytotoxicity.
Hemolytic activity
S. pumila showed haemolytic activity of 96% against RBC. Sepcic et al. (1997) described
significant levels of hemolytic activities of the sponge Reniera sarai extracts compared to the
moderate hemolytic activity of extracts from Saracotragus muscarum and Aplysina aerophoba.
Makarieva et al., (1983) reported that sterol derivatives from halichondriid sponges, namely
halistanol sulfates and sokotrasterol sulfates, possessed hemolytic activity. The crude methanolic
extracts of the marine sponge Callyspongia diffusa collected from the Mumbai coast was active
against the Escherichia coli, Pseudomonas aeruginosa and Vibrio cholera. It also induced
pronounced hemolysis on chicken erythrocytes (Boopathy et al ., 2009).
Sponges grouped under the class Demospongia are reported to be effective in producing the
diversity of secondary metabolites. The chloroform-methanol extract Haliclona panacea
(Demospongia) increased the acetylcholine esterase (AchE) activity. It also exhibited hemolytic
activity on chicken and human erythrocytes. At lower concentrations exhibited immune
stimulative effect, whereas at higher concentration it had immune suppressive effects (Newbold et
al., 1999). The S.pumila extracts showed maximum in vitro immunomodulatory effects than
H.atra. It showed increase in concentration of extracts showed high activity. A bioassay-guided
pharmacological screening study with sponge-derived extracts from specimens collected from the

64
coast of São Paulo State, on the south-eastern region of Brazil, recorded that the organic extracts
of Geodia corticostylifera was hemolytic on mice erythrocytes (Rangel et al. 2001).
Cariello and Zanetti (1999) studied the hemolytic activity of the sponge extracts Reniera
sarai. Another sponge Suberites domuncula showed the antimicrobial activities such as the
antibacterial and antifungal activities against the clinical pathogens. The sponge Sigmadocia
pumila extracts showed high activity on the human erythrocytes. The compounds from sponge
Topsentia sp showed strong hemolytic activity on fresh bovine erythrocytes. An aqueous extract
of Myrmekioderma styx exhibited the heamagglutinating activity. The Spongia sp and Spirastrella
sp released the spongistatins compound, which exhibited antiprolific activity (Kossuga et al.,
2008).
Tropical marine sponges exhibited various biological activities including hemolytic,
hemagglutinating, antibacterial and anti-acetyl cholinesterase activity. The hemolytic activity
exhibited from the sponge Pandaros acanthifolium showed effective amount of proteins. Ircinia
felix and Verongula rigida exhibited the strong antibacterial activity against both Gram positive
and Gram negative organisms. Then the sponge Myremekioderma styx showed the
hemagglutinating activity against human RBC (Hirata and Uemura, 1986). The peptide molecules
discovered from marine sponge are potent antibiotics. For example, Discodermis kiiensis produce
the discodermin as an antimicrobial peptide. Peptide was used in preparing antibiotics against
infectious human pathogen (Matsunaga et al., 1985). Secondary metabolites isolated from the
marine sponges, are considered to be an alternative source for vector control agents. It replaces
the existence and high toxic synthetic insecticides. The secondary metabolites were produced
through the association of sponge microorganisms. From Haliclona pigmentifera and Sigmadocia
carnosa sponges the secondary metabolites were produced and used to obtain novel pesticidal
molecules (Proksch et al., 2002). The major edible parts of sea cucumbers are the body walls,
which contain mainly collagen and acidic polysaccharides. They have anticoagulant and
haemolytic activities (Foneska and Mourao, 2006). The crude extracts of the H. atra showed high

65
hemolytic activity against the human erythrocytes. This could indicate the possibility of various
biological activities. The presence of triterpene glycosides is characteristic for most of sea
cucumbers and some sponges. Sea cucumber triterpene glycosides have lanostane type aglycones.
At milli- and micromolar concentrations sea cucumber glycosides show hemolytic, antifungal,
cytotoxic and other kinds of biological activities caused by membranotropic action of these
substances (Avilov et al., 2008).
H. atra body wall extracts showed hemolytic activity against the erythrocytes. Marine
invertebrates offer a source of potential antimicrobial drugs. Studies of antimicrobial mechanisms
and compounds of marine invertebrates may provide valuable information for new antibiotic
discoveries and give new insights into bioactive compounds (Mayer et al., 2007). Sea cucumbers
are echinoderms inhabitating relatively shallow coastal areas experience little or no currents. The
coelomic fluid of the holothuroid is known to contain naturally occurring factors exerting
hemagglutinating and hemolytic activity against various erythrocytes and microorganisms
(Olafsen et al., 1992).
NBT assay
The methanolic fractions of S. pumila have significantly increased in the intracellular
reduction of nitroblue tetrazolium dye to formazan by the neutrophils confirming the intracellular
killing and preserves the integrity of neutrophils. This was observed in NBT assay. Lectins
obtained from Holothuria scabra are involved in the processes such as the non-self recognition,
inflammation, cell-cell or cell-extracellular matrix interactions, fertilization, development and
regeneration (Saito et al., 1997). The immuno modulatory effects analysed in S. pumila extracts
was found to be maximum than H. atra. Even at low dose significant activity was seen. The
extracts of Halichondria panicea had immunostimulative effects at lower concentrations.
However, at higher concentrations, they exhibited immunosuppressive effects (Purushottama et
al., 2009). The H. atra extracts also showed immune modulatory activity which could be due to
the presence of compounds such as sulponamides and diketones. Cucumarioside derived from the

66
Sea cucumber Cucumaria japonica have potent immunomodulatory properties, and exhibited high
efficacy against E. coli, Proteusmirabilis, Neisseria meningitidis BT-2, Salmonellaminnesota,
Pertussis meningoencephalitis and Salmonella typhimurium (Sedov et al., 1990).
A variety of lectins have been isolated in the coelomic plasma and coelomocytes of marine
invertebrates specifically sea cucumbers. One of the roles of marine invertebrate lectins is to act
as the humoral factors in the defense mechanism, as the immunoglobulins in vertebrates. This is
suggested from some observations such as the activation of phagocytes by the binding of lectin to
foreign cell that is the opsonin activity (Hatakeyama et al., 1995). Therefore, the methanolic
extracts of S. pumila and sea cucumber H. atra have significant chemo attractant property.
The methanolic extracts of S. pumila and H. atra showed potent biological activities. The
solvent extracts and the compounds isolated from sea cucumbers have shown variety of biological
activities such as antifungal, anticancer, hemolytic, cytostatic, antioxidant and immuno-
modulatory effects (Dang et al., 2007). The presence of saponins, triterpene glycosides in sea
cucumbers have shown wide spectrum of biological activities such as the antifungal, hemolytic
and immunomodulatory activities (Kalinin et al., 1996). In vitro models such as the qualitative
nitroblue tetrazolium test showed significant immunostimulant activity for sponge S. pumila and
sea cucumber H. atra at a dose of 50% of the extract concentration. This investigation reveals the
methanolic fractions have significantly increased the phagocytic function of human neutrophils,
when compared with control and increase the movement of neutrophils towards the foreign body
which is the most important step in the phagocytosis.
The present investigation provides a comprehensive suggestion that the extracts of S. pumila
and the H. atra are sources for the bioactive compounds with antitumour and antiviral activity.





























