Embed Size (px)
Citation preview

REDUCTION OF Cr(VI)
35
CHAPTER 3: REDUCTION OF Cr(VI)
3.1 Introduction:
The reduction of highly toxic and mobile Cr(VI) to less toxic, less mobile Cr(III) is likely to be
useful for remediation of Cr(VI) contaminated soils and waters. Owing to the high toxicity of
Cr(VI) and disadvantages associated with conventional chemical methods of Cr(VI) removal, focus
of environmentalists has been shifted towards development of biological methods for detoxification
of Cr(VI) (Timmes and Pieper, 1999). Microbial mediated reduction of Cr(VI) to Cr(III) was first
reported by Romanenko and Korenkoc (1977) and there after number of Cr(VI) reducing bacterial
strains have been isolated and studied (Ackerley et al., 2004; Camargo et al., 2004; Elongovan et
al., 2008).
The earliest reports associated with isolation and characterization of Cr(VI) reducing bacteria were
of anaerobic bacterium, Enterobacter cloacae isolated from industrial wastewater. E. cloacae was
found to use chromate/dichromate as terminal electron acceptor during its anaerobic growth (Wang
et al., 1989; 1990). Anaerobic reduction of Cr(VI) has been subsequently described in several
organisms such as Shewanella oneidensis, Escherichia coli and several sulphate reducing bacteria
like Desulfovibrio vulgaris and Desulfovibrio fructosovorans (Lowe et al., 2003; Mclean and
Beveridge, 2001; Shen and Wang, 1993; Michel et al., 2001).
In addition to anaerobic Cr(VI) reducing microorganisms, a number of bacteria belonging to genera
Bacillus, Ochrobactrum, Pseudomonas, Escherichia and Parracoccus have been reported for their
Cr(VI) reduction ability under aerobic condition (Sultan and Hasnain, 2007; Hussein et al., 2005;
Garbisu et al., 1998). The Cr(VI) reducing micro-organisms can be categorized into aerobic or
anaerobic depending on their ability to reduce Cr(VI) under aerobic or anaerobic conditions.
Anaerobic bacteria mainly use Cr(VI) as terminal electron acceptor or reduce Cr(VI) in the
periplasmic space by hydrogenase or reduced cytochrome (Figure 3.1). For example, in D.
vulgaris and D. fructosovorans involvement of Cyt C3 and Ni-Fe dehydrogenase in Cr(VI)
reduction has been reported by Chardin et al. (2003) and Michel et al. (2001).
Several oxidoreductases with unrelated metabolic functions have been reported to catalyse Cr(VI)
reduction in bacteria. Examples of such enzymes include, quinone reductases, iron reductases, iron
reductases, nitro-reductases, flavin reductases and several NADH/NAD(P)H dependent reductases
of unknown function (Clark, 1994; Gonzalez et al., 2005; Gonzalez et al., 2003; Kwak et al., 2003;
Mazoch et al., 2004; Suziki et al., 1992). Chromate reductases from P. putida and E. coli have been
characterized in detail (Ackerley et al., 2004).

REDUCTION OF Cr(VI)
36
On the basis of sequence homology; the aerobic chromate reductases characterized till date has been
categorized into two main classes i.e
Class I chromate reductases
Class II chromate reductases
Figure 3.1 Mechanism of enzymatic Cr(VI) reduction under aerobic and anaerobic conditions
(Cheng and Gu, 2007). (MR: membrane chromate reductase; SR: Soluble chromate reductase).
Class I chromate reductase: Two of the most commonly studied class I enzyme include ChrR
(chromate reductase) of P. putida and YieF of E.coli (Accession number and 6058522, respectively)
(Ackerley et al., 2004b). Both these enzymes are dimeric flavoproteins of mass 50 KDa containing
signature sequences of the NADH_dh2 protein family (LFVTPEYNXXXXXXLKNAIDXXS)
representating putative flavin binding quinone reductases. The finding that ChrR and YieF are
members of flavin binding quinone reductase is consistent with the proposal made by Ishibashi et
al. (1990) that chromate reduction might be a secondary property of oxido-reductases with different
primary roles. With all these similarities, the two enzymes differ from each other in the way the
Cr(VI) is reduced to Cr(III).
Generally NADH_dh2 protein family consists of proteins displaying two electron reduction of
various electrophiles but ChrR is an one electron reducer enzyme which leads to the formation of
Cr(V) as intermediate. In contrast, enzyme YieF catalyses the direct reduction of Cr(VI) to Cr(III)

REDUCTION OF Cr(VI)
37
through transfer of four electrons. Out of four electrons, three electrons are consumed in reducing
Cr(VI) to Cr(III) whereas one electron is transferred to molecular oxygen, subsequently generating
H2O2. Hence YieF provide E. coli an effective mechanism of Cr(VI) reduction by forming lower
quantity of reactive oxygen species (Ackerley et al., 2004b).
Class II chromate reductase: The class II chromate reductases posses nitro-reductase activity and
bear no sequence homology to class I enzymes. The only member of the class II family include,
NfsA protein of E. coli (Accession number: 730007). NfsA is an obligate two electron reducer of
nitro compounds and quinones. However, the Cr(VI) reduction by NfsA involves a one electron
transfer and so more than 25% of the available electrons are consumed in ROS production and
Cr(V) is formed (Ackerley et al. 2004a).
Impact and application of redox mediators in bacterial biotransformation of contaminants
Redox mediators also referred as electron shuttle are organic molecules that can reversibly be
oxidized and reduced therefore conferring the capacity to serve as an electron carrier in multiple
redox reactions (Zee and Cervantes, 2009). Redox mediators are capable of transferring electrons in
redox reactions between a wide variety of both organic and inorganic compounds (Burgos et al.,
2003; Liu et al., 2009; Lovley et al., 1998; Maithreepala and Doong, 2009; Wang et al., 2009). The
most commonly studied redox mediators which support reductive biotransformation of various
contaminants include humic acid and their quinoid analogues (Lovley et al., 1998; Santos et al.,
2004a,b
).
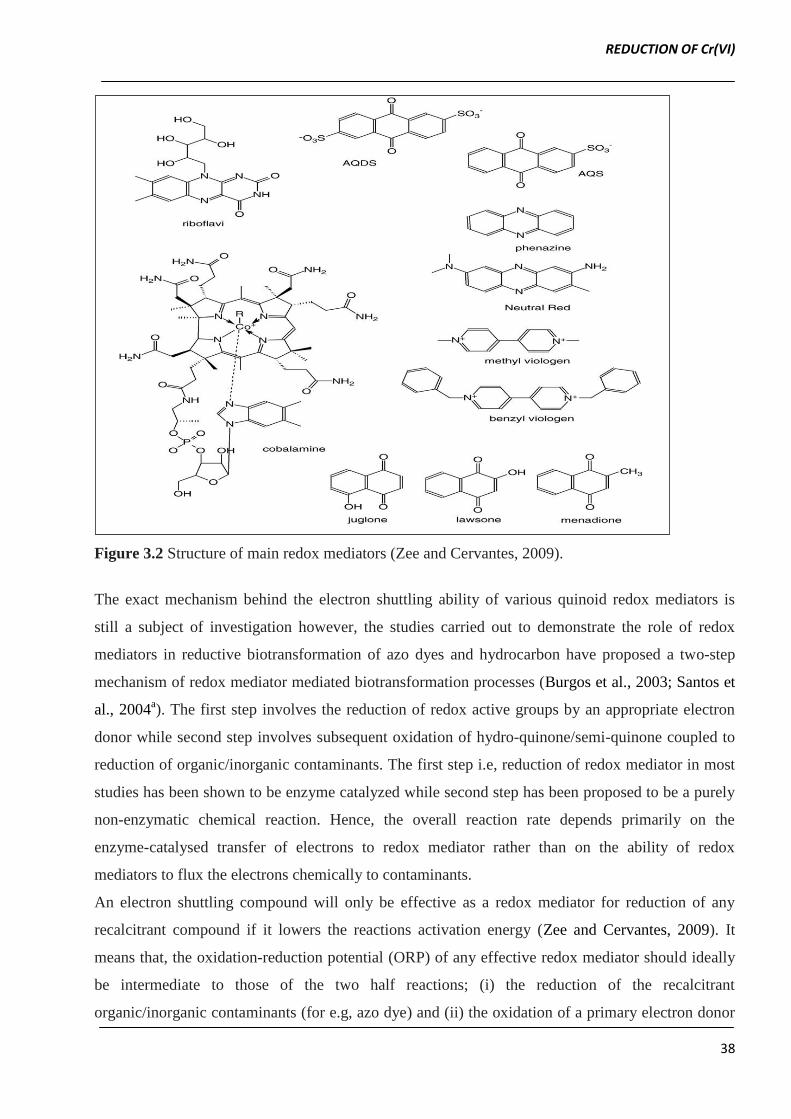
Figure 3.2 shows some of the redox mediators reported for their role in reductive biotransformation
of various organics and inorganic contaminants (e.g azo dye, hydrocarbons and several metal ions)
(Bond and Lovley 2002; Ling et al., 2009; Liu et al., 2009).
Among all these redox mediators, flavin based compounds such as flavin adenide mononucleotide
(FMN) and riboflavin as well as quinone based compounds such as anthra quinone sulphonate
(AQS), anthra quinone disulphonate (AQDS) and lawsone have been extensively investigated for
their application in degradation of environmental pollutants (Zee and Cervantes, 2009).
The term quinone collectively refers to organic structures that occur in three oxidation states and are
linked by one electron redox reactions. The three oxidation states are:
i. Hydroquinone with three protonation levels is the fully reduced form;
ii. The p-semiquinone radical with two protonation levels is an intermediate oxidation state;
iii. p-benzoquinone with one protonation level is the fully oxidized form.
REDUCTION OF Cr(VI)
38
Figure 3.2 Structure of main redox mediators (Zee and Cervantes, 2009).
The exact mechanism behind the electron shuttling ability of various quinoid redox mediators is
still a subject of investigation however, the studies carried out to demonstrate the role of redox
mediators in reductive biotransformation of azo dyes and hydrocarbon have proposed a two-step
mechanism of redox mediator mediated biotransformation processes (Burgos et al., 2003; Santos et
al., 2004a). The first step involves the reduction of redox active groups by an appropriate electron
donor while second step involves subsequent oxidation of hydro-quinone/semi-quinone coupled to
reduction of organic/inorganic contaminants. The first step i.e, reduction of redox mediator in most
studies has been shown to be enzyme catalyzed while second step has been proposed to be a purely
non-enzymatic chemical reaction. Hence, the overall reaction rate depends primarily on the
enzyme-catalysed transfer of electrons to redox mediator rather than on the ability of redox
mediators to flux the electrons chemically to contaminants.
An electron shuttling compound will only be effective as a redox mediator for reduction of any
recalcitrant compound if it lowers the reactions activation energy (Zee and Cervantes, 2009). It
means that, the oxidation-reduction potential (ORP) of any effective redox mediator should ideally
be intermediate to those of the two half reactions; (i) the reduction of the recalcitrant
organic/inorganic contaminants (for e.g, azo dye) and (ii) the oxidation of a primary electron donor
REDUCTION OF Cr(VI)
39
(NADH). Other wise the redox mediator will be ineffective or less effective in mediating the
reductive transformation of target pollutant.
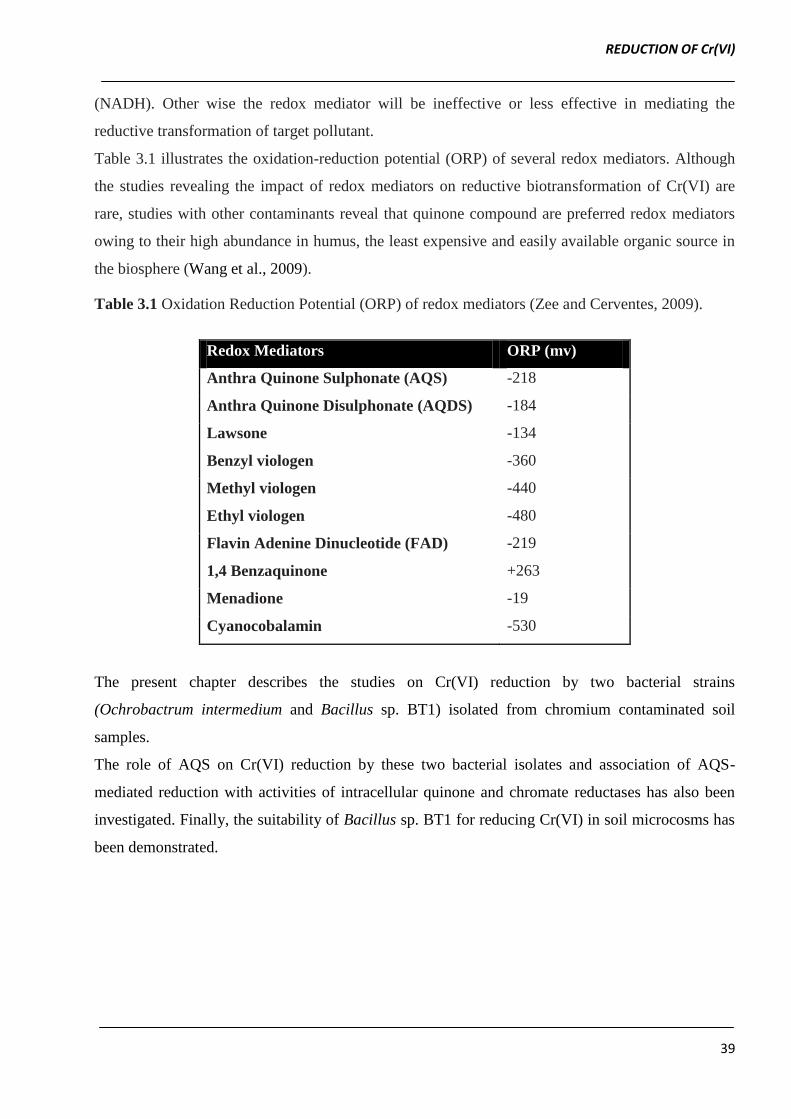
Table 3.1 illustrates the oxidation-reduction potential (ORP) of several redox mediators. Although
the studies revealing the impact of redox mediators on reductive biotransformation of Cr(VI) are
rare, studies with other contaminants reveal that quinone compound are preferred redox mediators
owing to their high abundance in humus, the least expensive and easily available organic source in
the biosphere (Wang et al., 2009).
Table 3.1 Oxidation Reduction Potential (ORP) of redox mediators (Zee and Cerventes, 2009).
Redox Mediators ORP (mv)
Anthra Quinone Sulphonate (AQS) -218
Anthra Quinone Disulphonate (AQDS) -184
Lawsone -134
Benzyl viologen -360
Methyl viologen -440
Ethyl viologen -480
Flavin Adenine Dinucleotide (FAD) -219
1,4 Benzaquinone +263
Menadione -19
Cyanocobalamin -530
The present chapter describes the studies on Cr(VI) reduction by two bacterial strains
(Ochrobactrum intermedium and Bacillus sp. BT1) isolated from chromium contaminated soil
samples.
The role of AQS on Cr(VI) reduction by these two bacterial isolates and association of AQS-
mediated reduction with activities of intracellular quinone and chromate reductases has also been
investigated. Finally, the suitability of Bacillus sp. BT1 for reducing Cr(VI) in soil microcosms has
been demonstrated.

REDUCTION OF Cr(VI)
40
3.2 Materials and Methods
3.2.1 Chemicals
Luria Bertani (LB) broth and redox mediators (namely, Anthraquinone sulphonate, Ethyl viologen,
Benzyl viologen and Methyl viologen) were purchased from Hi-Media Chemicals, India. Diphenyl
carbazide and potassium dichromate (K2Cr2O7) were procured from Qualigens, India.
3.2.2 Cr(VI) reduction by bacterial cultures
The 250 mL Erlenmeyer flasks containing 100 mL Luria Bertani broth amended with Cr(VI) (100
to 500 mg/L) were inoculated with overnight grown cells of bacterial culture (1.0 OD600 nm).
Uninoculated controls were used to determine abiotic Cr(VI) reduction. The inoculated cultures
along with un-inoculated controls were incubated and 1.0 mL sample was withdrawn at regular time
intervals to monitor growth and Cr(VI) reduction.
3.2.3 Effect of physico-chemical conditions on Cr(VI) reduction efficiency of bacterial isolate
Effect of different parameters including Cr(VI) concentration (100 to 500 mg/L), temperature (25 to
40 ºC), medium pH (5.5 to 9.0) and metal ions (Cu2+
, Co2+
, As2+
, Ni2+
) at concentration of 50 mg/L
on Cr(VI) reduction by bacterial cultures was investigated. Experiments were performed in 250 mL
Erlenmeyer flasks containing 100 mL LB broth amended with 100 mg Cr(VI)/L or as mentioned in
text. The desired initial pH of medium was adjusted using 1 N NaOH/ HCl. The overnight grown
culture of bacterial isolate was used as inoculum and added to experimental media so as to obtain an
initial cell density of 0.05 (600 nm). In each experiment, the residual Cr(VI) and bacterial growth
was monitored as described above in previous section.
3.2.4 Mediated Cr(VI) reduction by bacterial cultures in batch mode
Bacterial culture was inoculated in 150 mL Cr(VI) (100 to 500 mg/L) amended Luria-Bertani broth
augmented with AQS as redox mediator (1 mM). Cr(VI) reduction was monitored at regular time
interval in 1 mL sample withdrawn aseptically. Control experiments were performed in the same
manner except that no AQS was added to Luria Bertani broth. Un-inoculated media served as
abiotic controls for corresponding experiment.
The Cr(VI) reduction by bacterial culture in presence of different concentration of AQS was studied
wherein, Cr(VI) (100 mg/L) amended Luria Bertani broth was supplemented with AQS in the
concentration range of 0 to 5.0 mM. Cr(VI) reduction was monitored from samples withdrawn at
different time intervals.

REDUCTION OF Cr(VI)
41
3.2.5 Effect of temperature on AQS mediated Cr(VI) reduction
The Cr(VI) amended LB broth supplemented either with or with out 1 mM AQS was inoculated
with overnight grown bacterial culture and incubated at various temperatures (25 to 35 °C). The
samples were withdrawn at regular time interval to monitor residual Cr(VI).
The activation energy of the AQS mediated and non-mediated Cr(VI) reduction by bacterial culture
was calculated by employing Arrhenius equation as follows
ln k = −Ea/R T + lnAo ……………………………………………………………………(3.1)
Where k is the first order rate constant (h-1
), Ea is activation energy and Ao is constant. The value of
Ea can be determined from the Slope (-Ea/R) of ln k versus 1/T plot (Santos et al., 2004).
3.2.6 Repeated cycles of AQS mediated Cr(VI) reduction in fed batch mode
200 mL of Luria Bertani broth supplemented with or without AQS (1mM) was spiked with 100 mg
Cr(VI)/L. The Cr(VI) reduction was monitored immediately after inoculation of bacterial culture.
The samples were withdrawn and Cr(VI) was estimated. On complete reduction of Cr(VI), the
medium was spiked with another aliquot of Cr(VI) and its reduction was monitored. The cycles of
Cr(VI) spiking and reduction were repeated till Cr(VI) reduction ceased.
3.2.7 Preparation of cell free lysate
Bacterial culture was grown in 200 mL Luria Bertani broth for 24 h at 30 C. The cell pellet
obtained upon centrifugation (8,603 g for 15 min) was resuspended in 3 mL of phosphate buffer
(100 mM, pH: 7.0). The resuspended cells were disrupted by sonication for 15 min (Sonics &
Materials, Inc., USA) in cold conditions. The resultant homogenate was centrifuged at 8,603 g for
30 min at 4 ºC to remove cell debris and the clear supernatant was used as cell free lysate for
enzyme assays.
3.2.8 Characterization of quinone and chromate reductase
The influence of pH, temperature and substrate concentration on chromate and quinone reductase
activity of bacterial culture was assessed.
For determination of optimal pH, the quinone and chromate reductase activity was assayed at 30 °C
using 50 mM sodium acetate buffer (pH: 5.0 to 5.5), potassium phosphate buffer (pH: 6.0 to 7.5)
and tris-HCl buffer (7.5 to 8.5).
Optimal temperature was determined wherein the chromate and quinone reductase activity was
assayed at optimum pH with varying temperatures (20 °C to 50 °C) using temperature controlled
thermostatic cuvette holder (ELICO DL 198 biospectrophotometer, Hyderabad, India).

REDUCTION OF Cr(VI)
42
3.2.9 Cr(VI) bioremediation experiment: soil microcosm studies
The soil used for microcosm studies, was collected from agricultural field. Immediately after
sample collection, the soil was sieved to remove gravel and plant residues. 20 g of dry soil was
taken in each 100 mL Erlenmeyer flasks for microcosm experiments. The flasks were plugged with
non-adsorbent cotton and sterilized for three consecutive days by autoclaving (121 ºC, 30 minutes).
The cells of isolate BT1 grown overnight in LB were washed with sterile distilled water in order to
remove any media particles and subsequently resuspended in distilled water to be used as inoculum
for microcosm experiments. The inoculum was added to soil amended with 100 mg Cr(VI)/ kg soil
to achieve a cell density of 104
cfu/g soil. The moisture content of soil in microcosm experiment
was maintained at 50 %.
Abiotic control was kept in order to verify all the non-biological Cr(VI) reduction. Both un-
inoculated and inoculated soil microcosms were vigorously mixed and incubated at 30 ºC for 15
days under aseptic conditions. The water content of the soil was maintained at 50 % level by
weighing the microcosms and adding sterile distilled water to compensate for any weight loss.
3.2.10 Extraction of Cr(VI) and measurement of bacterial growth in soil microcosms.
Each of the soil microcosms was sampled by removing soil sample (0.5 g) with a sterile spatula on
day 3rd
, 6th
, 9th
, 12th
and 15th
. For extraction of Cr(VI), 0.5 g of soil sample was suspended in 5 mL
of distilled water, vortexed thoroughly followed by centrifugation (8,603, 10 minutes, room
temperature). The supernatant was used for Cr(VI) estimation. Likewise, the 0.5 g of soil
resuspended in 5 mL of distilled water was mixed thoroughly and allowed to settle down. Finally,
standard 10-fold serial dilution plate count method was carried out to monitor the total colony
forming units (cfu/g soil) of isolate BT1.
3.2.11 Influence of various factors on Cr(VI) reduction efficiency of isolate BT1 in soil
microcosms.
The efficiency of isolate BT1 was investigated using soils spiked with two different concentrations
of Cr(VI) (viz. 100 and 300 mg/kg soil) and residual Cr(VI) at different time intervals was
monitored as described above.
Effect of malt extract amendment (0.05 to 1.0 %, w/w) on Cr (VI) reduction by isolate BT1 was
evaluated.
3.2.11 Seed germination assay
The seed germination tests were carried out in petriplates by employing two types of test seeds
[mustard (Brassica nigra), variety type: western seed research and mungbean seeds (Vigna

REDUCTION OF Cr(VI)
43
radiata), variety type: K-851-certified]. In order to determine toxicity of Cr(VI), 15 seeds were
placed in each petriplate containing whattman filter paper-1 soaked with pure Cr(VI) solution (50,
100, 200 or 300 mg/L). The plates were incubated at 30 ºC for 48 h and monitored for seed
germination.
In order to determine effectiveness of Cr(VI) bioremediation by isolate BT1 in soil microcosm,
seeds germination experiments were performed using the soil extract obtained from microcosm
experiments. The soil extract was prepared by suspending 10 g of soil from each microcosm system
in 10 mL of distilled water, followed by vortexing and subsequently filtering through whattman
number 1.0 filter paper. The seed germination experiments using microcosm soil extracts were
performed as described above. In all seed germination experiments, seeds sterilized with 0.1 %
(w/v) HgCl2 solution were employed.
3.2.12 Analytical Methods
3.2.12.1 Enzyme assays
The quinone and chromate reductase were assayed as described previously by Puzon et al. (2002).
Briefly, quinone and chromate reductase activity was assayed spectrophotometrically at constant
temperature of 30 ºC following the oxidation of NADH at 340 nm (molar absorption coefficient
6.22 mM-1
cm-1
). The reaction was initiated by addition of cell free lysate to reaction mixture
containing 50 mM phosphate buffer (pH 6.0) and 0.1 mM substrate (Lawsone as quinone reductase
substrate and K2Cr2O7 as chromate reductase substrate).
One unit of enzyme activity was defined as the amount of enzyme required for oxidation of 1 µmole
of NADH per min under standard assay conditions (Santos et al., 2004a).
3.2.12.2 Quantification of growth and Cr(VI)
Bacterial growth was monitored turbidometrically. The cell pellet obtained upon centrifugation of
1.0 mL of culture was resuspended in 1.0 mL distilled water and its absorbance was measured at
600 nm.
The Cr(VI) concentration in the cell-free supernatant was measured using the Diphenylcarbazide
(DPC) reagent (Ishibashi et al., 1990). Briefly, the hexavalent chromium containing samples (in the
range of 1.0 to 10 µg) were acidified by adding 330 µL of 6N sulfuric acid. To this acidified
solution of the hexavalent chromium, DPC was added at concentration of 0.25 % and final volume
was made upto 10 mL. The mixture was incubated for 10 min at room temperature and the colour of
DPC:Cr(VI) complex was measured by reading the absorbance at 540 nm.
REDUCTION OF Cr(VI)
44
3.2.12.3 Protein estimation
The protein concentration of the cell-free extract was estimated using Folin-phenol reagent by
reading absorbance at 720 nm, following the principle of Lowry et al. (1951).
3.3 Results and Discussion
Part A: Reduction of hexavalent chromium by Ochrobactrum intermedium BCR400 isolated
from a chromium-contaminated soil
A hexavalent chromium reducing bacterial strain designated as BCR400 was isolated from the
Cr(VI) contaminated soil collected from the landfill site of chemical industry, Vadodara, Gujarat,
India.
On the basis of the carbon substrate utilization pattern (employing GN2 as well as GENIII plates of
BioLog, USA) and 16s rDNA sequence, isolate BCR400 was identified as O. intermedium with
similarity index of 0.74 and 99% probability (Accession number: JN033212).
O. intermedium belongs to ά–proteobacteria subclass and representatives of this taxa have been
isolated previously from chromium contaminated soils, by several researchers (He et al., 2009;
Ozdemir et al., 2003; Sultan and Hasnain, 2007; Thacker and Madamwar, 2005). O. intermedium
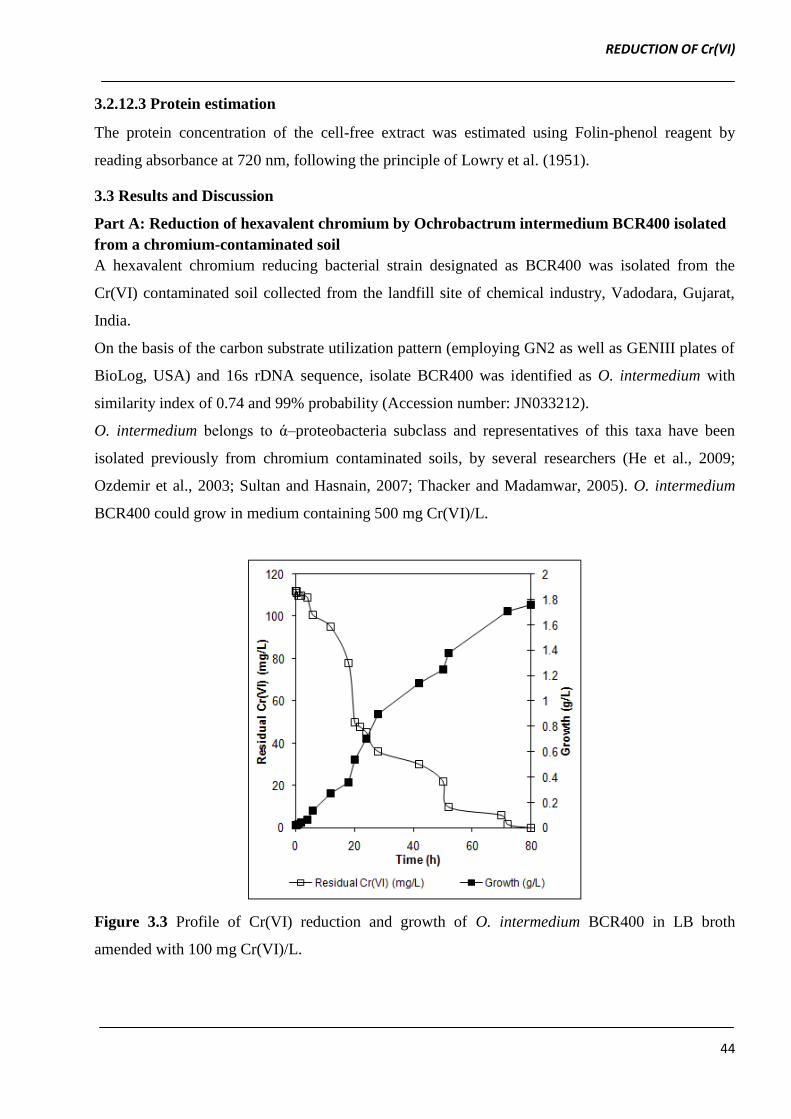
BCR400 could grow in medium containing 500 mg Cr(VI)/L.
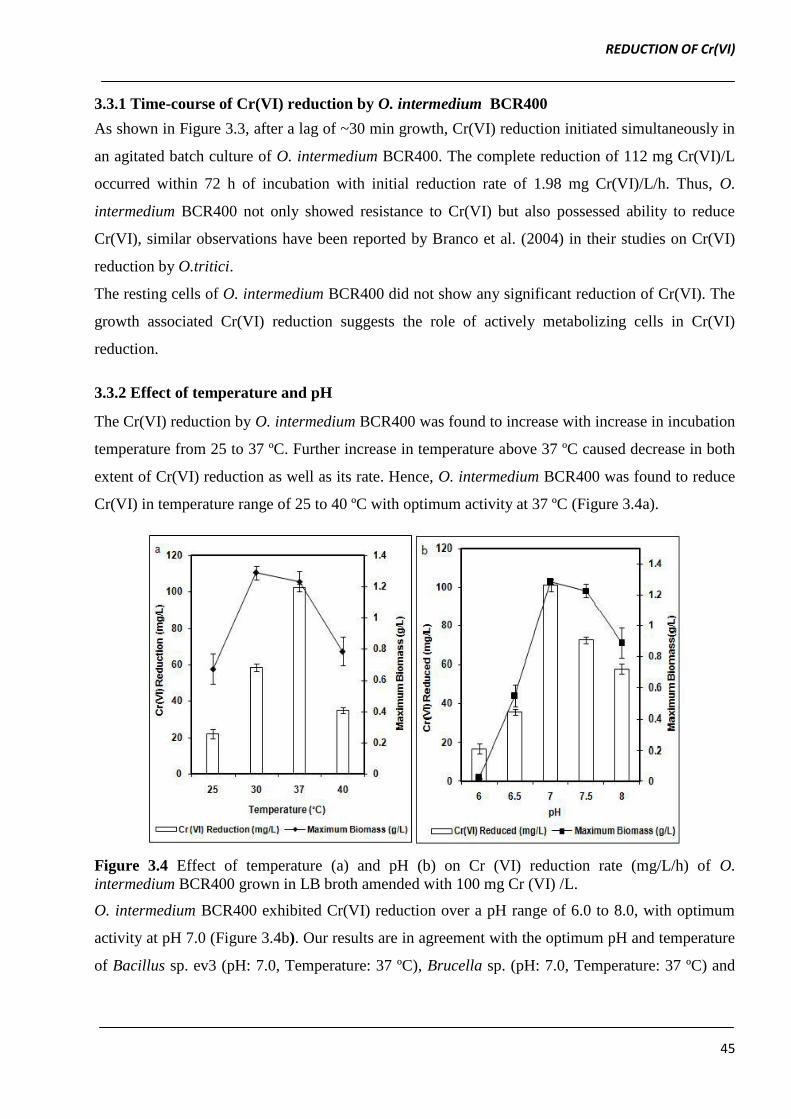
Figure 3.3 Profile of Cr(VI) reduction and growth of O. intermedium BCR400 in LB broth
amended with 100 mg Cr(VI)/L.
REDUCTION OF Cr(VI)
45
3.3.1 Time-course of Cr(VI) reduction by O. intermedium BCR400
As shown in Figure 3.3, after a lag of ~30 min growth, Cr(VI) reduction initiated simultaneously in
an agitated batch culture of O. intermedium BCR400. The complete reduction of 112 mg Cr(VI)/L
occurred within 72 h of incubation with initial reduction rate of 1.98 mg Cr(VI)/L/h. Thus, O.
intermedium BCR400 not only showed resistance to Cr(VI) but also possessed ability to reduce
Cr(VI), similar observations have been reported by Branco et al. (2004) in their studies on Cr(VI)
reduction by O.tritici.
The resting cells of O. intermedium BCR400 did not show any significant reduction of Cr(VI). The
growth associated Cr(VI) reduction suggests the role of actively metabolizing cells in Cr(VI)
reduction.
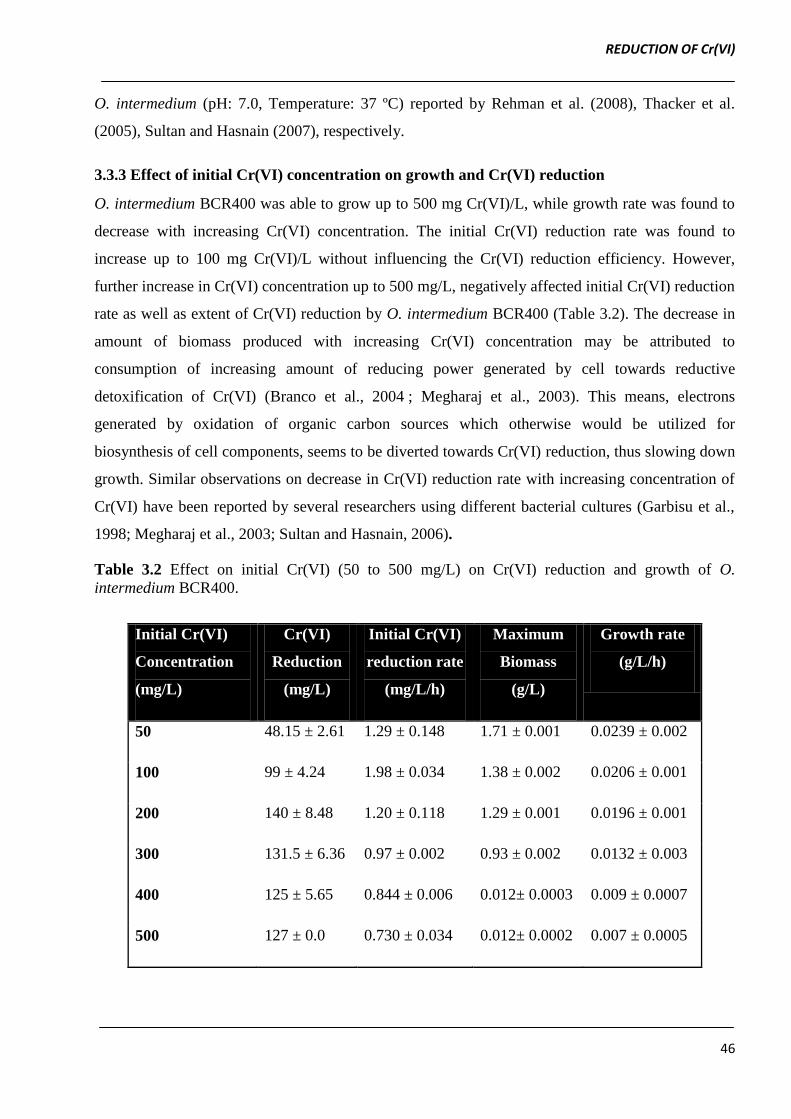
3.3.2 Effect of temperature and pH
The Cr(VI) reduction by O. intermedium BCR400 was found to increase with increase in incubation
temperature from 25 to 37 ºC. Further increase in temperature above 37 ºC caused decrease in both
extent of Cr(VI) reduction as well as its rate. Hence, O. intermedium BCR400 was found to reduce
Cr(VI) in temperature range of 25 to 40 ºC with optimum activity at 37 ºC (Figure 3.4a).
Figure 3.4 Effect of temperature (a) and pH (b) on Cr (VI) reduction rate (mg/L/h) of O.
intermedium BCR400 grown in LB broth amended with 100 mg Cr (VI) /L.
O. intermedium BCR400 exhibited Cr(VI) reduction over a pH range of 6.0 to 8.0, with optimum
activity at pH 7.0 (Figure 3.4b). Our results are in agreement with the optimum pH and temperature
of Bacillus sp. ev3 (pH: 7.0, Temperature: 37 ºC), Brucella sp. (pH: 7.0, Temperature: 37 ºC) and
REDUCTION OF Cr(VI)
46
O. intermedium (pH: 7.0, Temperature: 37 ºC) reported by Rehman et al. (2008), Thacker et al.
(2005), Sultan and Hasnain (2007), respectively.
3.3.3 Effect of initial Cr(VI) concentration on growth and Cr(VI) reduction
O. intermedium BCR400 was able to grow up to 500 mg Cr(VI)/L, while growth rate was found to
decrease with increasing Cr(VI) concentration. The initial Cr(VI) reduction rate was found to
increase up to 100 mg Cr(VI)/L without influencing the Cr(VI) reduction efficiency. However,
further increase in Cr(VI) concentration up to 500 mg/L, negatively affected initial Cr(VI) reduction
rate as well as extent of Cr(VI) reduction by O. intermedium BCR400 (Table 3.2). The decrease in
amount of biomass produced with increasing Cr(VI) concentration may be attributed to
consumption of increasing amount of reducing power generated by cell towards reductive
detoxification of Cr(VI) (Branco et al., 2004 ; Megharaj et al., 2003). This means, electrons
generated by oxidation of organic carbon sources which otherwise would be utilized for
biosynthesis of cell components, seems to be diverted towards Cr(VI) reduction, thus slowing down
growth. Similar observations on decrease in Cr(VI) reduction rate with increasing concentration of
Cr(VI) have been reported by several researchers using different bacterial cultures (Garbisu et al.,
1998; Megharaj et al., 2003; Sultan and Hasnain, 2006).
Table 3.2 Effect on initial Cr(VI) (50 to 500 mg/L) on Cr(VI) reduction and growth of O.
intermedium BCR400.
Initial Cr(VI)
Concentration
(mg/L)
Cr(VI)
Reduction
(mg/L)
Initial Cr(VI)
reduction rate
(mg/L/h)
Maximum
Biomass
(g/L)
Growth rate
(g/L/h)
50 48.15 ± 2.61 1.29 ± 0.148 1.71 ± 0.001 0.0239 ± 0.002
100 99 ± 4.24 1.98 ± 0.034 1.38 ± 0.002 0.0206 ± 0.001
200 140 ± 8.48 1.20 ± 0.118 1.29 ± 0.001 0.0196 ± 0.001
300 131.5 ± 6.36 0.97 ± 0.002 0.93 ± 0.002 0.0132 ± 0.003
400 125 ± 5.65 0.844 ± 0.006 0.012± 0.0003 0.009 ± 0.0007
500 127 ± 0.0 0.730 ± 0.034 0.012± 0.0002 0.007 ± 0.0005
REDUCTION OF Cr(VI)
47
3.3.4 Effect of metal ions on Cr(VI) reduction by O. intermedium BCR400
The industrial wastewaters would contain various other heavy metals apart from Cr(VI), which
though at lower concentration might influence the activity of chromium reducing bacteria (Agrawal
et al., 2008). Thus, Cr(VI) reduction by O. intermedium BCR400 was studied in presence of other
metallic ions at concentration of 50 mg/L. Table 3.3 shows that Cr(VI) reduction increases slightly
in presence of copper (50 mg/L) whereas it decreases in presence of same concentration of cobalt,
arsenic, nickel, zinc and cadmium. The, level of inhibition was different with different metal ions;
cadmium was found to exert maximum inhibitory effect followed by zinc, nickel, cobalt, arsenate at
50 mg/L concentration. Inhibition of Cr(VI) reduction by O. intermedium in presence of metal ions
like Zn2+
, Cd2+
and Pb2+
has been reported by Sultan and Hasnain (2007) however it is interesting to
note they also observed increase in Cr(VI) reduction in presence of metal ions like Cu2+
, Co2+
, Mn2+
and Ni2+
. The main mechanism behind stimulatory effect of certain metals on Cr(VI) reduction is
not clear how ever it may be attributed to their role as cofactors for reductases involved in Cr(VI)
transformation.
Table 3.3 Maximum Cr (VI) reduction and Cr (VI) reduction rate of O. intemedium BCR400 in LB
broth amended with 100 mg/L Cr (VI) supplemented with co-metal ions at concentration of 50
mg/L.
Co-Metal ions
50 (mg/L)
Maximum Cr(VI)
Reduction (mg/L)
Cr(VI) Reduction
Rate (mg/L/h)
None 95.5±0.707 1.98±0.014
Copper (Cu2+
) 100±0.707 2.09±0.014
Cobalt (Co2+
) 65±4.24 1.35±0.088
Arsenic (As5+
) 65±5.65 1.35±0.117
Nickel (Ni2+
) 59.5±3.5 1.23±0.073
Zinc (Zn2+
) 54.5±2.12 1.13±0.044
Cadmium (Cd2+
) 44±2.82 0.91±0.058
3.3.5 AQS-mediated reduction of Cr(VI) by O. intermedium BCR400
Inclusion of low molecular weight redox mediator along with the metal is being currently
considered in order to enhance efficiency of metal reduction by micro-organisms (Bond and Lovley,
2002; Lovley et al., 1998; Lovley et al., 1999). In the present study, O. intermedium BCR400
exhibited higher initial Cr(VI) reduction rate in presence of 1.0 mM AQS, ethyl and methyl
REDUCTION OF Cr(VI)
48
viologen. In presence of 1.0 mM AQS, O. intermedium BCR400 reduced Cr(VI) at 1.4 fold higher
rate (2.88 mg Cr(VI)/L/h) than in absence of any redox mediators (1.98 mg Cr(VI)/L/h).
AQS has been reported as powerful mediator for reductive biotransformation of several organic
recalcitrants (Liu et al. 2010; Zee and Cervantes 2009). Although ethyl and methyl viologen also
enhanced Cr(VI) reduction rate, they were found to inhibit growth of O. intermedium BCR400
(Table 3.4). Benzyl viologen severely inhibited the growth of O. intermedium BCR400.
Quinones are known for their potential role as redox centres in humic acid and therefore the Cr(VI)
reduction rates in presence of AQS may attributed to their electron shuttling ability between O.
intermedium BCR400 and Cr(VI). Zee and Cervantes (2009) suggested that the oxidation reduction
potential of any mediator should not be much lower than -0.320 V, which is the lowest oxidation
reduction potential of cofactor (NADPH) in the cell otherwise it would not be reduced significantly.
Hence, the transfer of electrons from AQS to Cr(VI) seems to be thermodynamically favourable
phenomenon as the standard redox potential (E°) of AQS is -0.218 V which is significantly lower
than the redox potential of CrO42-
(1.28 V) and higher than -0.320 V (Fredrickson et al., 2006).
According to the above hypothesis the poor mediating ability of ethyl viologen and methyl viologen
may be explained by their much lower oxidation reduction potential (- 0.480 and – 0.440 V
respectively) than – 0.320 V.
Table 3.4 Effect of redox mediators on Cr(VI) reduction and growth of O. intermedium BCR400.
Redox mediators
(1.0 mM)
Cr(VI)
Reduction
(mg/L)
Initial Cr(VI)
reduction
rate (mg/L/h)
Maximum
Biomass
(g/L)
Growth rate
(g/L/h)
None 101 1.98 1.01 0.038
AQS 101 2.88 1.35 0.051
Ethyl Viologen 102 2.12 0.825 0.022
Methyl Viologen 103 2.14 0.768 0.021
Benzyl Viologen 65.52 0.91 0.045 0.0012
Furthermore, O. intermedium BCR400 exhibited higher initial Cr(VI) reduction rate in presence of
1.0 mM AQS at all Cr(VI) concentrations tested (Table 3.5). The initial Cr(VI) reduction rates in
batch cultures of O. intermedium BCR400 were found to be 1.29, 1.98, 1.20, 0.97, 0.844 and 0.73
mg Cr(VI)/L/h corresponding to the initial Cr(VI) concentration of 50, 100, 200, 300, 400 and 500
REDUCTION OF Cr(VI)
49
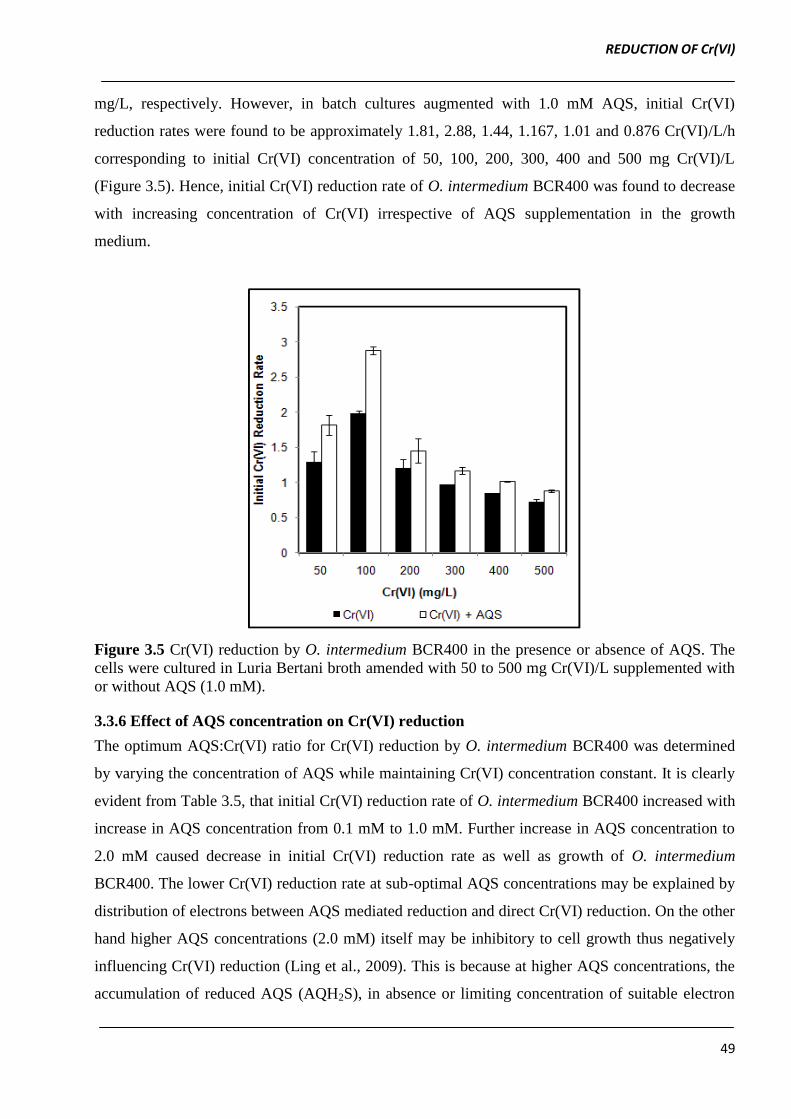
mg/L, respectively. However, in batch cultures augmented with 1.0 mM AQS, initial Cr(VI)
reduction rates were found to be approximately 1.81, 2.88, 1.44, 1.167, 1.01 and 0.876 Cr(VI)/L/h
corresponding to initial Cr(VI) concentration of 50, 100, 200, 300, 400 and 500 mg Cr(VI)/L
(Figure 3.5). Hence, initial Cr(VI) reduction rate of O. intermedium BCR400 was found to decrease
with increasing concentration of Cr(VI) irrespective of AQS supplementation in the growth
medium.
Figure 3.5 Cr(VI) reduction by O. intermedium BCR400 in the presence or absence of AQS. The
cells were cultured in Luria Bertani broth amended with 50 to 500 mg Cr(VI)/L supplemented with
or without AQS (1.0 mM).
3.3.6 Effect of AQS concentration on Cr(VI) reduction
The optimum AQS:Cr(VI) ratio for Cr(VI) reduction by O. intermedium BCR400 was determined
by varying the concentration of AQS while maintaining Cr(VI) concentration constant. It is clearly
evident from Table 3.5, that initial Cr(VI) reduction rate of O. intermedium BCR400 increased with
increase in AQS concentration from 0.1 mM to 1.0 mM. Further increase in AQS concentration to
2.0 mM caused decrease in initial Cr(VI) reduction rate as well as growth of O. intermedium
BCR400. The lower Cr(VI) reduction rate at sub-optimal AQS concentrations may be explained by
distribution of electrons between AQS mediated reduction and direct Cr(VI) reduction. On the other
hand higher AQS concentrations (2.0 mM) itself may be inhibitory to cell growth thus negatively
influencing Cr(VI) reduction (Ling et al., 2009). This is because at higher AQS concentrations, the
accumulation of reduced AQS (AQH2S), in absence or limiting concentration of suitable electron
REDUCTION OF Cr(VI)
50
acceptor, would transfer electrons to O2, resulting in generation of ROS and thereby causing cell
death (Sedlacek and Kucera, 2010). The maximum reduction rate of Cr(VI) as well as growth of O.
intermedium BCR400 at optimal AQS (1.0 mM) in medium containing 100 mg Cr(VI)/L may be
due to efficient relay of electrons from primary electron donor to AQS to Cr(VI) mediated through
action of reductases. Rau et al. (2002) also observed highest reduction of azo dye amaranth in
presence of 1.0 mM AQS.
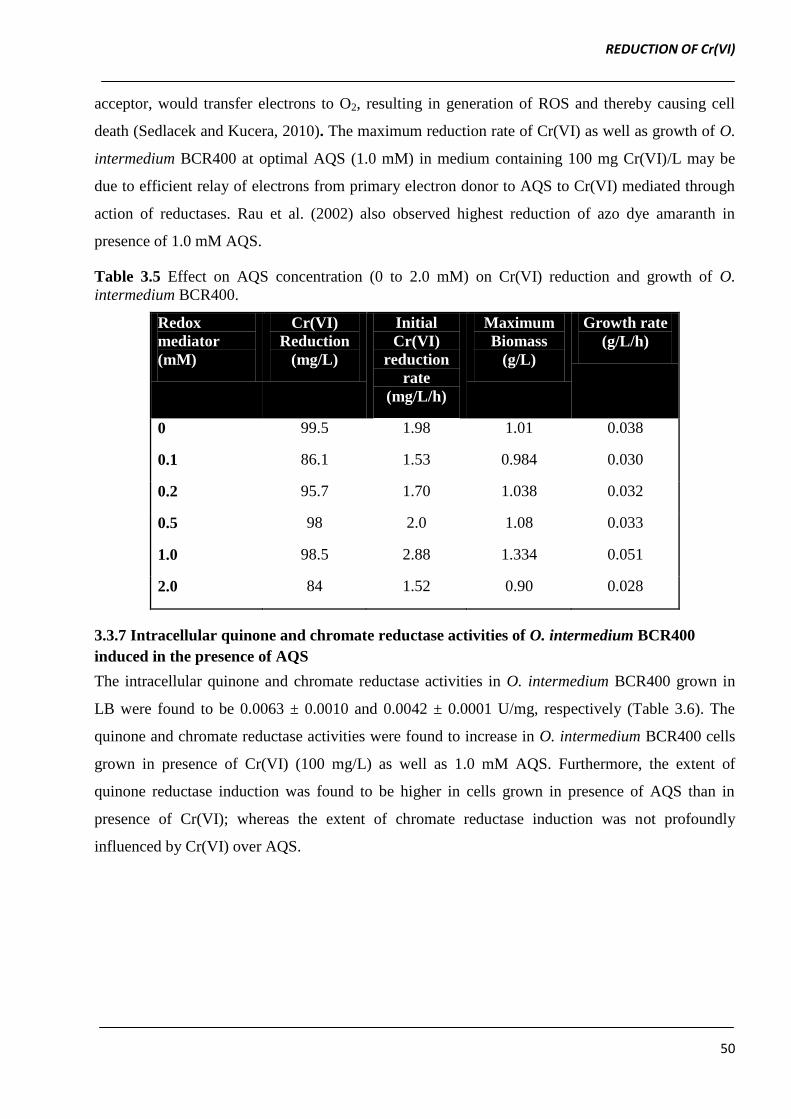
Table 3.5 Effect on AQS concentration (0 to 2.0 mM) on Cr(VI) reduction and growth of O.
intermedium BCR400.
Redox
mediator
(mM)
Cr(VI)
Reduction
(mg/L)
Initial
Cr(VI)
reduction
rate
(mg/L/h)
Maximum
Biomass
(g/L)
Growth rate
(g/L/h)
0 99.5 1.98 1.01 0.038
0.1 86.1 1.53 0.984 0.030
0.2 95.7 1.70 1.038 0.032
0.5 98 2.0 1.08 0.033
1.0 98.5 2.88 1.334 0.051
2.0 84 1.52 0.90 0.028
3.3.7 Intracellular quinone and chromate reductase activities of O. intermedium BCR400
induced in the presence of AQS
The intracellular quinone and chromate reductase activities in O. intermedium BCR400 grown in
LB were found to be 0.0063 ± 0.0010 and 0.0042 ± 0.0001 U/mg, respectively (Table 3.6). The
quinone and chromate reductase activities were found to increase in O. intermedium BCR400 cells
grown in presence of Cr(VI) (100 mg/L) as well as 1.0 mM AQS. Furthermore, the extent of
quinone reductase induction was found to be higher in cells grown in presence of AQS than in
presence of Cr(VI); whereas the extent of chromate reductase induction was not profoundly
influenced by Cr(VI) over AQS.
REDUCTION OF Cr(VI)
51
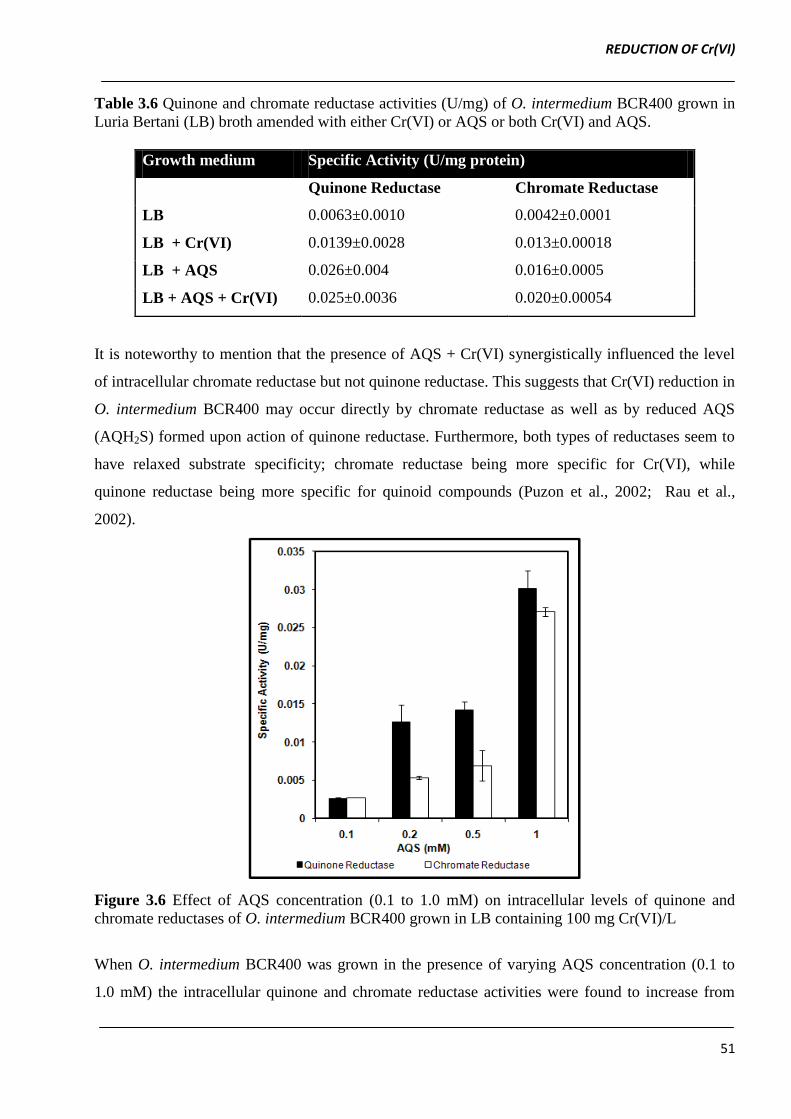
Table 3.6 Quinone and chromate reductase activities (U/mg) of O. intermedium BCR400 grown in
Luria Bertani (LB) broth amended with either Cr(VI) or AQS or both Cr(VI) and AQS.
Growth medium Specific Activity (U/mg protein)
Quinone Reductase Chromate Reductase
LB 0.0063±0.0010 0.0042±0.0001
LB + Cr(VI) 0.0139±0.0028 0.013±0.00018
LB + AQS 0.026±0.004 0.016±0.0005
LB + AQS + Cr(VI) 0.025±0.0036 0.020±0.00054
It is noteworthy to mention that the presence of AQS + Cr(VI) synergistically influenced the level
of intracellular chromate reductase but not quinone reductase. This suggests that Cr(VI) reduction in
O. intermedium BCR400 may occur directly by chromate reductase as well as by reduced AQS
(AQH2S) formed upon action of quinone reductase. Furthermore, both types of reductases seem to
have relaxed substrate specificity; chromate reductase being more specific for Cr(VI), while
quinone reductase being more specific for quinoid compounds (Puzon et al., 2002; Rau et al.,
2002).
Figure 3.6 Effect of AQS concentration (0.1 to 1.0 mM) on intracellular levels of quinone and
chromate reductases of O. intermedium BCR400 grown in LB containing 100 mg Cr(VI)/L
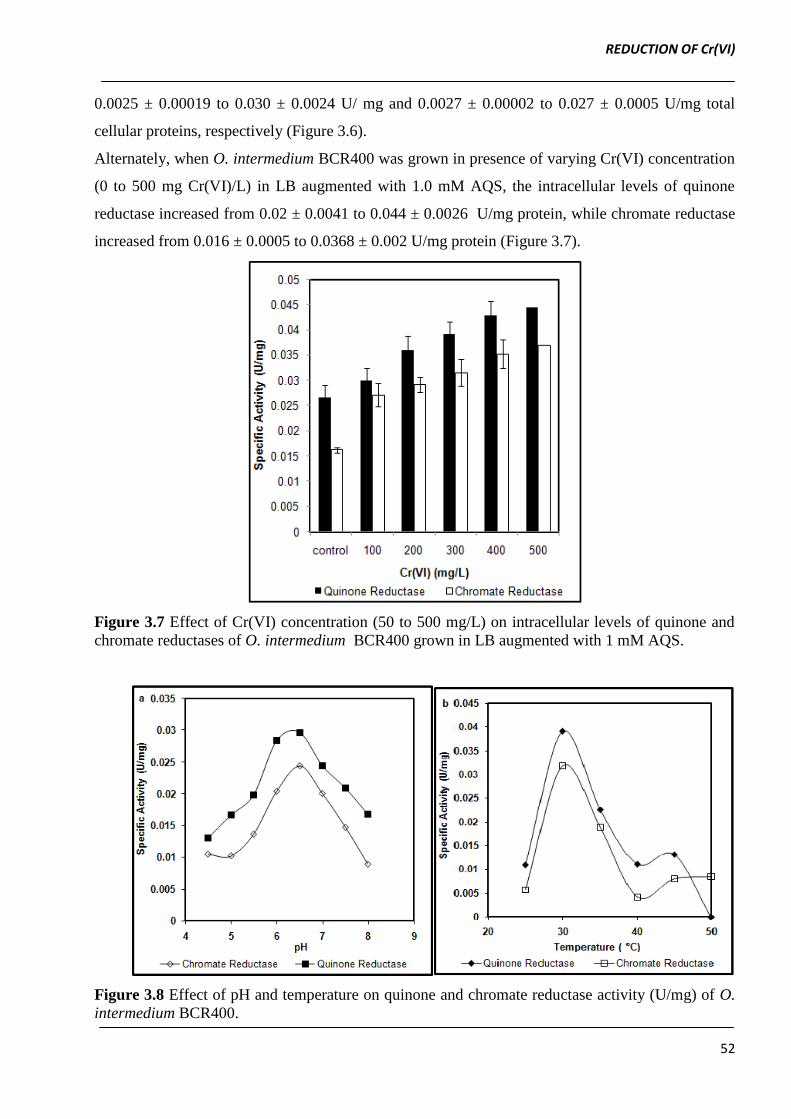
When O. intermedium BCR400 was grown in the presence of varying AQS concentration (0.1 to
1.0 mM) the intracellular quinone and chromate reductase activities were found to increase from
REDUCTION OF Cr(VI)
52
0.0025 ± 0.00019 to 0.030 ± 0.0024 U/ mg and 0.0027 ± 0.00002 to 0.027 ± 0.0005 U/mg total
cellular proteins, respectively (Figure 3.6).
Alternately, when O. intermedium BCR400 was grown in presence of varying Cr(VI) concentration
(0 to 500 mg Cr(VI)/L) in LB augmented with 1.0 mM AQS, the intracellular levels of quinone
reductase increased from 0.02 ± 0.0041 to 0.044 ± 0.0026 U/mg protein, while chromate reductase
increased from 0.016 ± 0.0005 to 0.0368 ± 0.002 U/mg protein (Figure 3.7).
Figure 3.7 Effect of Cr(VI) concentration (50 to 500 mg/L) on intracellular levels of quinone and
chromate reductases of O. intermedium BCR400 grown in LB augmented with 1 mM AQS.
Figure 3.8 Effect of pH and temperature on quinone and chromate reductase activity (U/mg) of O.
intermedium BCR400.
REDUCTION OF Cr(VI)
53
A strong and non linear positive co-relation between the concentration of AQS/Cr(VI) and activities
of both the enzymes (γ > + 90) was observed. This suggests the role of AQS in induction of quinone
and chromate reductases in O. intermedium BCR400.
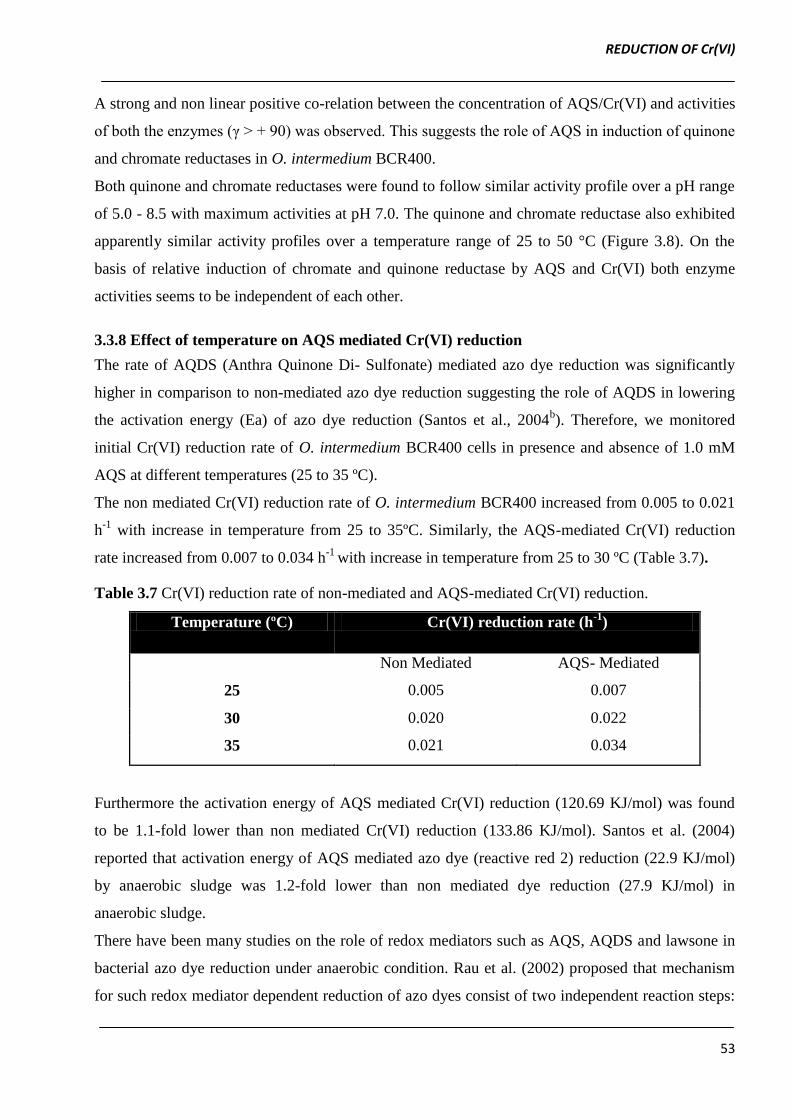
Both quinone and chromate reductases were found to follow similar activity profile over a pH range
of 5.0 - 8.5 with maximum activities at pH 7.0. The quinone and chromate reductase also exhibited
apparently similar activity profiles over a temperature range of 25 to 50 °C (Figure 3.8). On the
basis of relative induction of chromate and quinone reductase by AQS and Cr(VI) both enzyme
activities seems to be independent of each other.
3.3.8 Effect of temperature on AQS mediated Cr(VI) reduction
The rate of AQDS (Anthra Quinone Di- Sulfonate) mediated azo dye reduction was significantly
higher in comparison to non-mediated azo dye reduction suggesting the role of AQDS in lowering
the activation energy (Ea) of azo dye reduction (Santos et al., 2004b). Therefore, we monitored
initial Cr(VI) reduction rate of O. intermedium BCR400 cells in presence and absence of 1.0 mM
AQS at different temperatures (25 to 35 ºC).
The non mediated Cr(VI) reduction rate of O. intermedium BCR400 increased from 0.005 to 0.021
h-1
with increase in temperature from 25 to 35ºC. Similarly, the AQS-mediated Cr(VI) reduction
rate increased from 0.007 to 0.034 h-1
with increase in temperature from 25 to 30 ºC (Table 3.7).
Table 3.7 Cr(VI) reduction rate of non-mediated and AQS-mediated Cr(VI) reduction.
Temperature (ºC) Cr(VI) reduction rate (h-1
)
Non Mediated AQS- Mediated
25 0.005 0.007
30 0.020 0.022
35 0.021 0.034
Furthermore the activation energy of AQS mediated Cr(VI) reduction (120.69 KJ/mol) was found
to be 1.1-fold lower than non mediated Cr(VI) reduction (133.86 KJ/mol). Santos et al. (2004)
reported that activation energy of AQS mediated azo dye (reactive red 2) reduction (22.9 KJ/mol)
by anaerobic sludge was 1.2-fold lower than non mediated dye reduction (27.9 KJ/mol) in
anaerobic sludge.
There have been many studies on the role of redox mediators such as AQS, AQDS and lawsone in
bacterial azo dye reduction under anaerobic condition. Rau et al. (2002) proposed that mechanism
for such redox mediator dependent reduction of azo dyes consist of two independent reaction steps:

REDUCTION OF Cr(VI)
54
First, the quinones are enzymatically reduced to the corresponding hydroquinone (Ling et al., 2009;
Rau and Stolz, 2003) and second, the hydroquinones cleave the azo dyes in purely chemical
reaction. Following the proposed mechanism by Rau et al. (2002), Ling et al. (2009) suggested that
entire reaction rate depends on enzymatic reduction of redox mediators. Accordingly, the
effectiveness of any mediator depends on the presence of membrane bound or intracellular
reductase with mediator reducing ability.
Part B: Reduction of Cr(VI) in liquid media and soil microcosms by Bacillus sp BT1 isolated
from chromium contaminated soil of tanning industry
A hexavalent chromium resistant bacterial strain designated as BT1 was isolated from the Cr(VI)
contaminated soil collected from the vicinity of tannery industry, Ranipet, Tamilnadu, India. The
carbon substrate utilization pattern of BT1 (using Biolog identification system) and 16s rDNA
sequencing revealed that BT1 was closely related to Bacillus sp. (Accession number: JN093017).
The Bacillus sp. have been reported for their occurrence in metal stressed environment as well as
ability to reduce Cr(VI) (Camargo et al., 2004; Campos et al., 1995; Elangovan et al., 2006; Liu et
al., 2006). Bacillus sp. BT1 showed similar Cr(VI) reduction under both agitated as well as static
growth conditions (Data not shown). Hence, all the experiments were carried out under static
growth conditions.
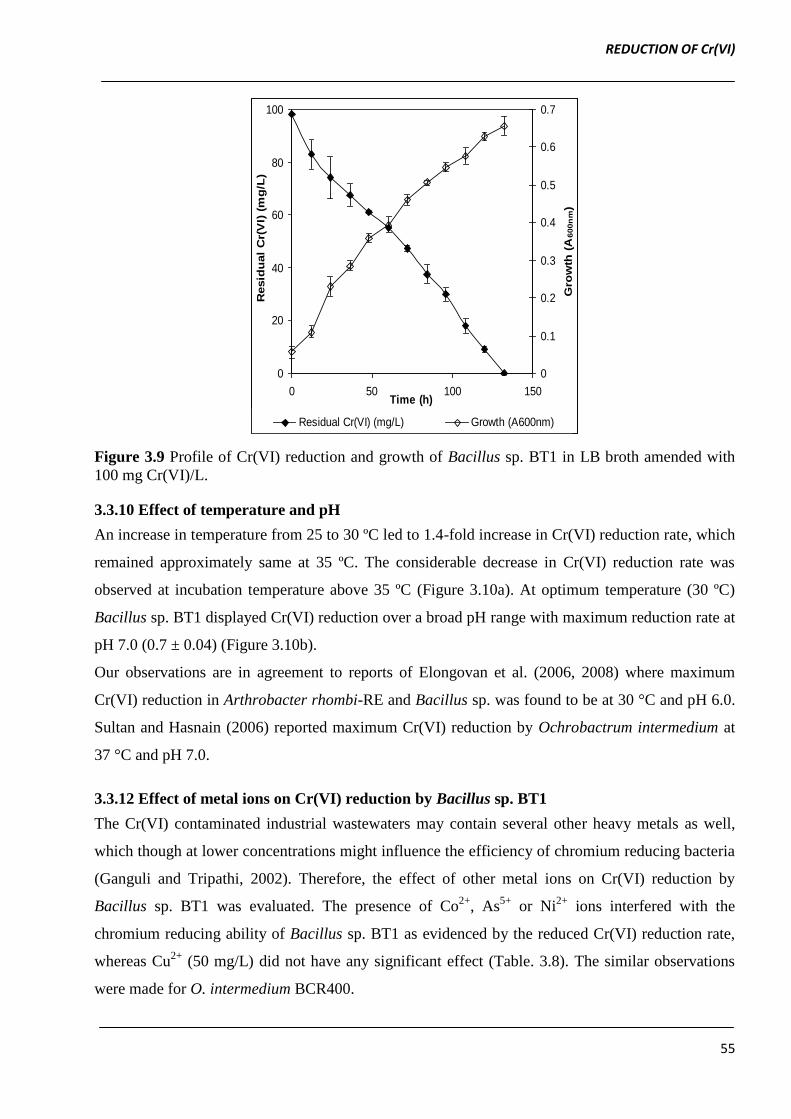
3.3.9 Time-course of Cr(VI) reduction by Bacillus sp. BT1
Growth as well as Cr(VI) reduction was monitored at initial Cr(VI) concentration of 100 mg/L at
pH 7.0 and 30 °C under static condition. The Cr(VI) reduction by Bacillus sp. BT1 was found to be
growth associated as reduction efficiency increased with increase in the growth of culture. The
complete reduction of 100 mg Cr(VI)/L was observed after 132 h of incubation at 30 °C (Figure
3.9). Liu et al. (2006) also demonstrated that there exists relationship between growth and Cr(VI)
reduction as indicated by difference in the growth curve pattern corresponding to various
concentration of Cr(VI).
REDUCTION OF Cr(VI)
55
0
20
40
60
80
100
0 50 100 150Time (h)
Resid
ual
Cr(
VI)
(m
g/L
)
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
Gro
wth
(A
600n
m)
Residual Cr(VI) (mg/L) Growth (A600nm)
Figure 3.9 Profile of Cr(VI) reduction and growth of Bacillus sp. BT1 in LB broth amended with
100 mg Cr(VI)/L.
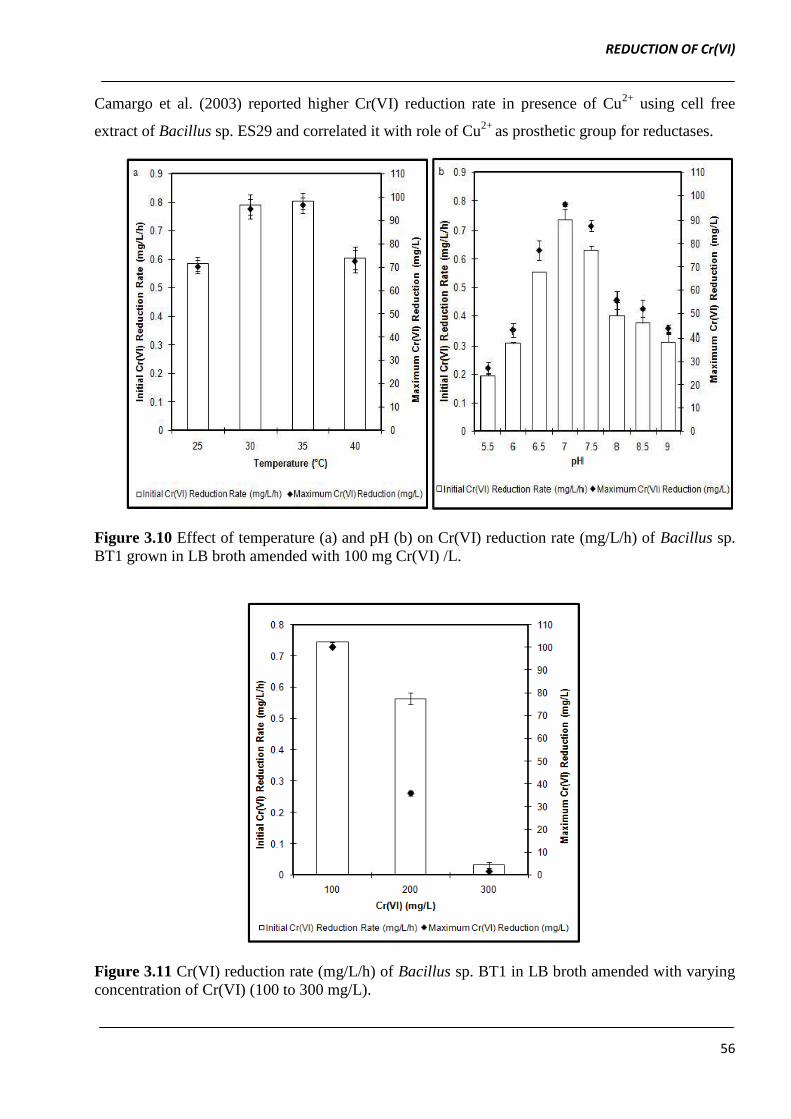
3.3.10 Effect of temperature and pH
An increase in temperature from 25 to 30 ºC led to 1.4-fold increase in Cr(VI) reduction rate, which
remained approximately same at 35 ºC. The considerable decrease in Cr(VI) reduction rate was
observed at incubation temperature above 35 ºC (Figure 3.10a). At optimum temperature (30 ºC)
Bacillus sp. BT1 displayed Cr(VI) reduction over a broad pH range with maximum reduction rate at
pH 7.0 (0.7 ± 0.04) (Figure 3.10b).
Our observations are in agreement to reports of Elongovan et al. (2006, 2008) where maximum
Cr(VI) reduction in Arthrobacter rhombi-RE and Bacillus sp. was found to be at 30 °C and pH 6.0.
Sultan and Hasnain (2006) reported maximum Cr(VI) reduction by Ochrobactrum intermedium at
37 °C and pH 7.0.
3.3.12 Effect of metal ions on Cr(VI) reduction by Bacillus sp. BT1
The Cr(VI) contaminated industrial wastewaters may contain several other heavy metals as well,
which though at lower concentrations might influence the efficiency of chromium reducing bacteria
(Ganguli and Tripathi, 2002). Therefore, the effect of other metal ions on Cr(VI) reduction by
Bacillus sp. BT1 was evaluated. The presence of Co2+
, As5+
or Ni2+
ions interfered with the
chromium reducing ability of Bacillus sp. BT1 as evidenced by the reduced Cr(VI) reduction rate,
whereas Cu2+
(50 mg/L) did not have any significant effect (Table. 3.8). The similar observations
were made for O. intermedium BCR400.
REDUCTION OF Cr(VI)
56
Camargo et al. (2003) reported higher Cr(VI) reduction rate in presence of Cu2+
using cell free
extract of Bacillus sp. ES29 and correlated it with role of Cu2+
as prosthetic group for reductases.
Figure 3.10 Effect of temperature (a) and pH (b) on Cr(VI) reduction rate (mg/L/h) of Bacillus sp.
BT1 grown in LB broth amended with 100 mg Cr(VI) /L.
Figure 3.11 Cr(VI) reduction rate (mg/L/h) of Bacillus sp. BT1 in LB broth amended with varying
concentration of Cr(VI) (100 to 300 mg/L).
REDUCTION OF Cr(VI)
57
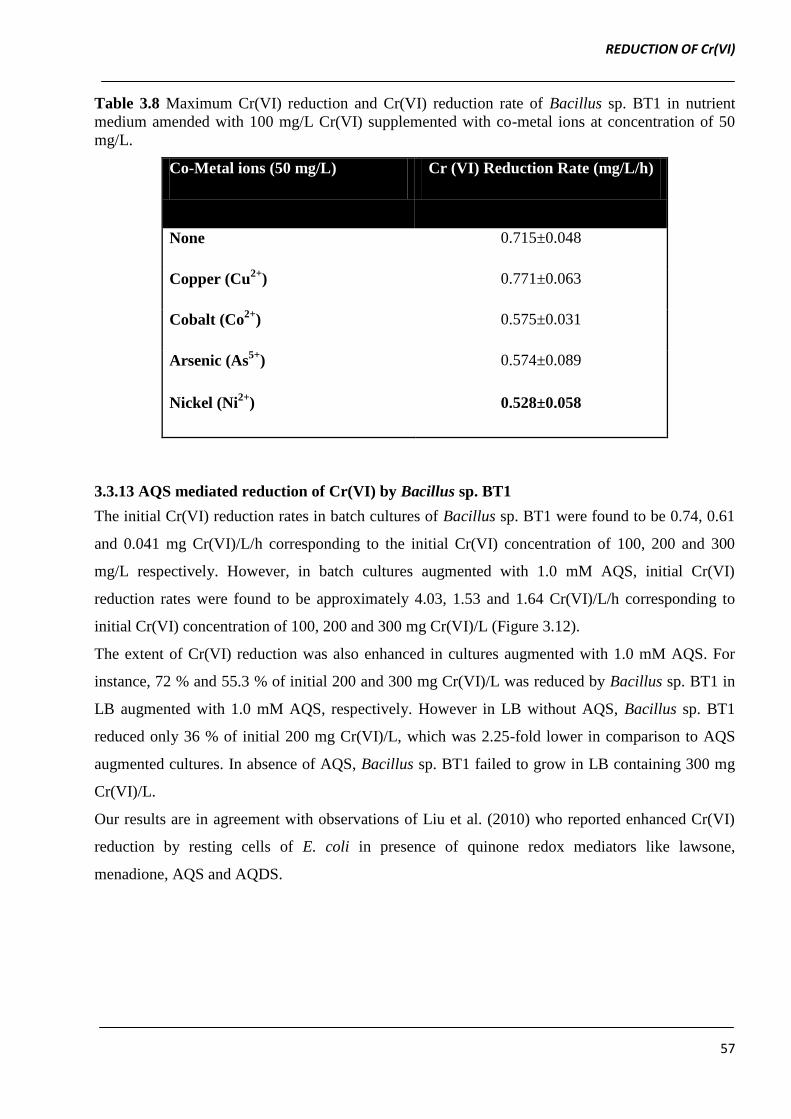
Table 3.8 Maximum Cr(VI) reduction and Cr(VI) reduction rate of Bacillus sp. BT1 in nutrient
medium amended with 100 mg/L Cr(VI) supplemented with co-metal ions at concentration of 50
mg/L.
Co-Metal ions (50 mg/L) Cr (VI) Reduction Rate (mg/L/h)
None 0.715±0.048
Copper (Cu2+
) 0.771±0.063
Cobalt (Co2+
) 0.575±0.031
Arsenic (As5+
) 0.574±0.089
Nickel (Ni2+
) 0.528±0.058
3.3.13 AQS mediated reduction of Cr(VI) by Bacillus sp. BT1
The initial Cr(VI) reduction rates in batch cultures of Bacillus sp. BT1 were found to be 0.74, 0.61
and 0.041 mg Cr(VI)/L/h corresponding to the initial Cr(VI) concentration of 100, 200 and 300
mg/L respectively. However, in batch cultures augmented with 1.0 mM AQS, initial Cr(VI)
reduction rates were found to be approximately 4.03, 1.53 and 1.64 Cr(VI)/L/h corresponding to
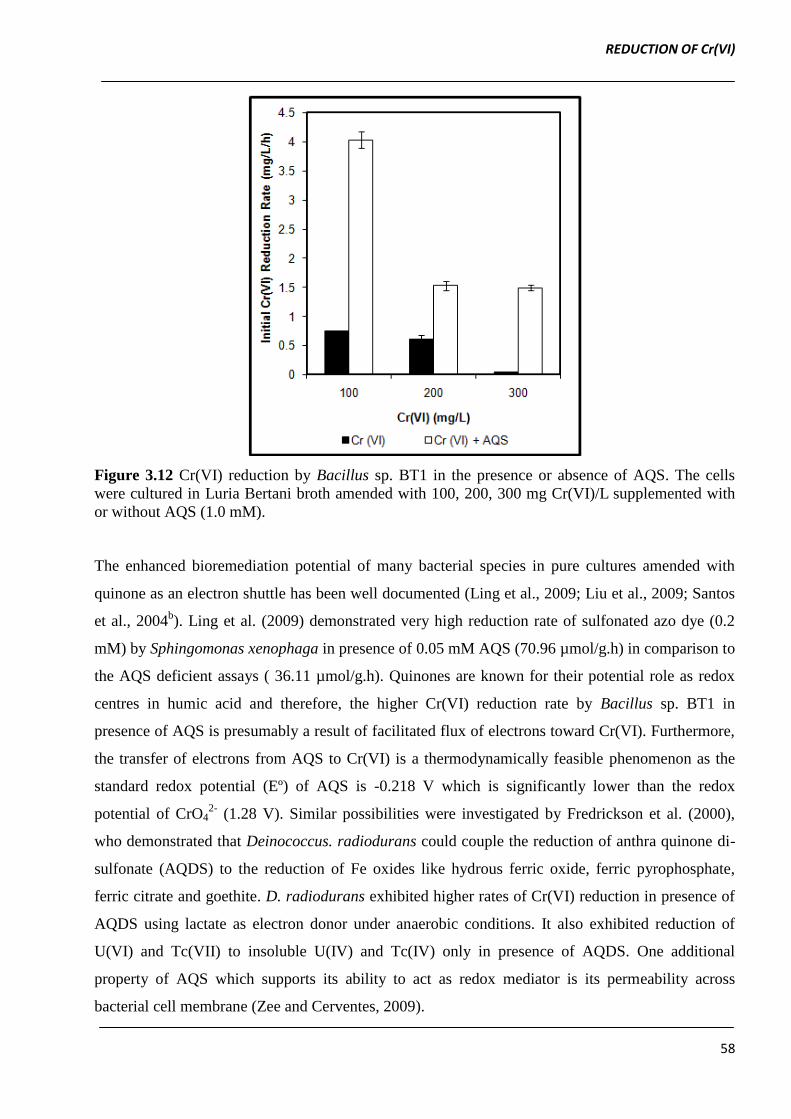
initial Cr(VI) concentration of 100, 200 and 300 mg Cr(VI)/L (Figure 3.12).
The extent of Cr(VI) reduction was also enhanced in cultures augmented with 1.0 mM AQS. For
instance, 72 % and 55.3 % of initial 200 and 300 mg Cr(VI)/L was reduced by Bacillus sp. BT1 in
LB augmented with 1.0 mM AQS, respectively. However in LB without AQS, Bacillus sp. BT1
reduced only 36 % of initial 200 mg Cr(VI)/L, which was 2.25-fold lower in comparison to AQS
augmented cultures. In absence of AQS, Bacillus sp. BT1 failed to grow in LB containing 300 mg
Cr(VI)/L.
Our results are in agreement with observations of Liu et al. (2010) who reported enhanced Cr(VI)
reduction by resting cells of E. coli in presence of quinone redox mediators like lawsone,
menadione, AQS and AQDS.
REDUCTION OF Cr(VI)
58
Figure 3.12 Cr(VI) reduction by Bacillus sp. BT1 in the presence or absence of AQS. The cells
were cultured in Luria Bertani broth amended with 100, 200, 300 mg Cr(VI)/L supplemented with
or without AQS (1.0 mM).
The enhanced bioremediation potential of many bacterial species in pure cultures amended with
quinone as an electron shuttle has been well documented (Ling et al., 2009; Liu et al., 2009; Santos
et al., 2004b). Ling et al. (2009) demonstrated very high reduction rate of sulfonated azo dye (0.2
mM) by Sphingomonas xenophaga in presence of 0.05 mM AQS (70.96 µmol/g.h) in comparison to
the AQS deficient assays ( 36.11 µmol/g.h). Quinones are known for their potential role as redox
centres in humic acid and therefore, the higher Cr(VI) reduction rate by Bacillus sp. BT1 in
presence of AQS is presumably a result of facilitated flux of electrons toward Cr(VI). Furthermore,
the transfer of electrons from AQS to Cr(VI) is a thermodynamically feasible phenomenon as the
standard redox potential (Eº) of AQS is -0.218 V which is significantly lower than the redox
potential of CrO42-
(1.28 V). Similar possibilities were investigated by Fredrickson et al. (2000),
who demonstrated that Deinococcus. radiodurans could couple the reduction of anthra quinone di-
sulfonate (AQDS) to the reduction of Fe oxides like hydrous ferric oxide, ferric pyrophosphate,
ferric citrate and goethite. D. radiodurans exhibited higher rates of Cr(VI) reduction in presence of
AQDS using lactate as electron donor under anaerobic conditions. It also exhibited reduction of
U(VI) and Tc(VII) to insoluble U(IV) and Tc(IV) only in presence of AQDS. One additional
property of AQS which supports its ability to act as redox mediator is its permeability across
bacterial cell membrane (Zee and Cerventes, 2009).
REDUCTION OF Cr(VI)
59
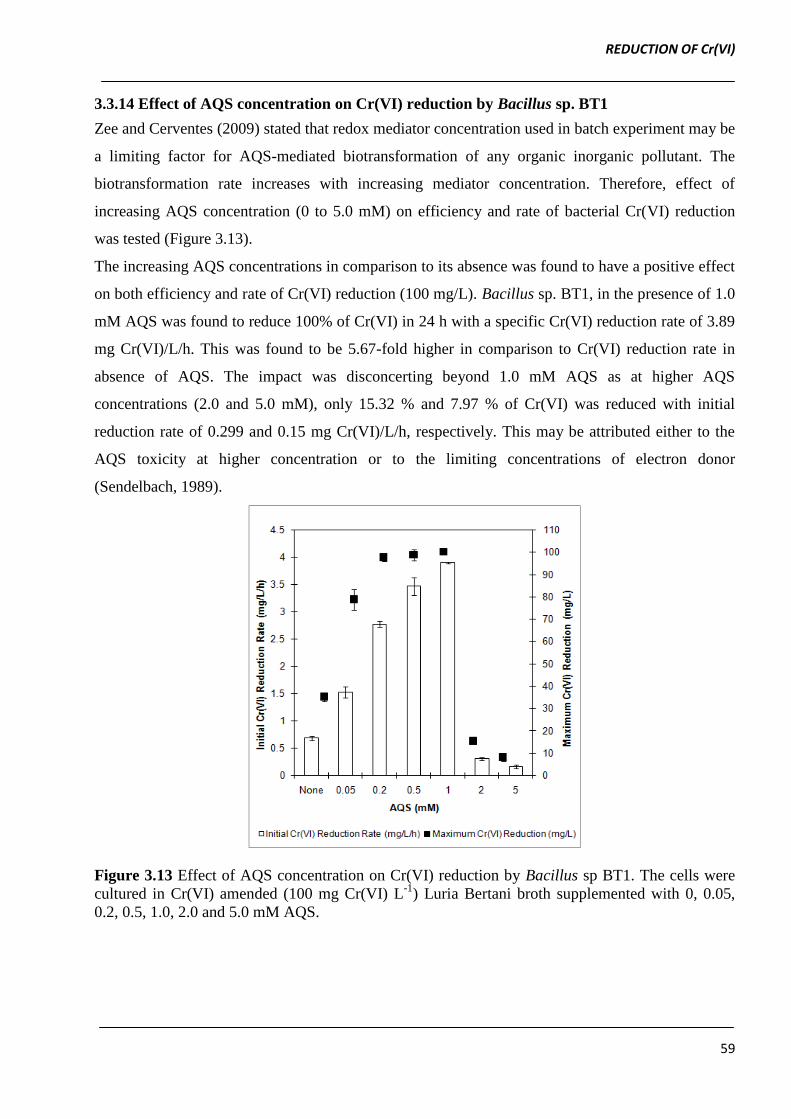
3.3.14 Effect of AQS concentration on Cr(VI) reduction by Bacillus sp. BT1
Zee and Cerventes (2009) stated that redox mediator concentration used in batch experiment may be
a limiting factor for AQS-mediated biotransformation of any organic inorganic pollutant. The
biotransformation rate increases with increasing mediator concentration. Therefore, effect of
increasing AQS concentration (0 to 5.0 mM) on efficiency and rate of bacterial Cr(VI) reduction
was tested (Figure 3.13).
The increasing AQS concentrations in comparison to its absence was found to have a positive effect
on both efficiency and rate of Cr(VI) reduction (100 mg/L). Bacillus sp. BT1, in the presence of 1.0
mM AQS was found to reduce 100% of Cr(VI) in 24 h with a specific Cr(VI) reduction rate of 3.89
mg Cr(VI)/L/h. This was found to be 5.67-fold higher in comparison to Cr(VI) reduction rate in
absence of AQS. The impact was disconcerting beyond 1.0 mM AQS as at higher AQS
concentrations (2.0 and 5.0 mM), only 15.32 % and 7.97 % of Cr(VI) was reduced with initial
reduction rate of 0.299 and 0.15 mg Cr(VI)/L/h, respectively. This may be attributed either to the
AQS toxicity at higher concentration or to the limiting concentrations of electron donor
(Sendelbach, 1989).
Figure 3.13 Effect of AQS concentration on Cr(VI) reduction by Bacillus sp BT1. The cells were
cultured in Cr(VI) amended (100 mg Cr(VI) L-1
) Luria Bertani broth supplemented with 0, 0.05,
0.2, 0.5, 1.0, 2.0 and 5.0 mM AQS.
REDUCTION OF Cr(VI)
60
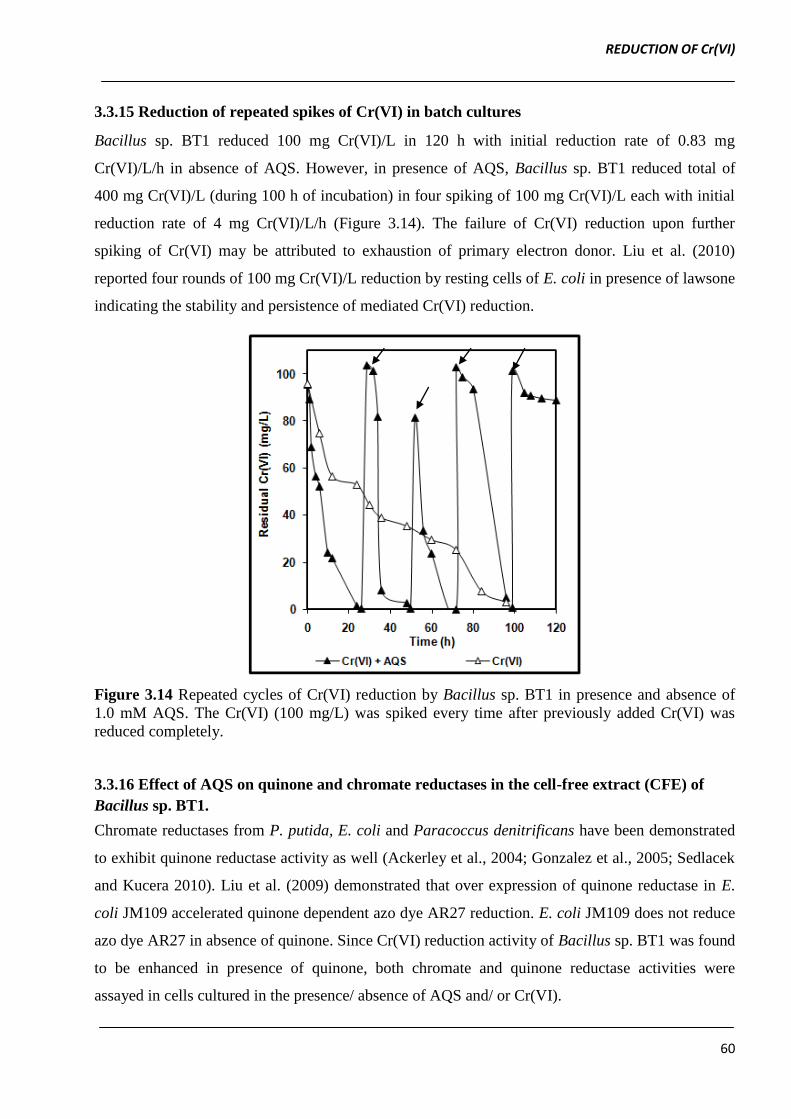
3.3.15 Reduction of repeated spikes of Cr(VI) in batch cultures
Bacillus sp. BT1 reduced 100 mg Cr(VI)/L in 120 h with initial reduction rate of 0.83 mg
Cr(VI)/L/h in absence of AQS. However, in presence of AQS, Bacillus sp. BT1 reduced total of
400 mg Cr(VI)/L (during 100 h of incubation) in four spiking of 100 mg Cr(VI)/L each with initial
reduction rate of 4 mg Cr(VI)/L/h (Figure 3.14). The failure of Cr(VI) reduction upon further
spiking of Cr(VI) may be attributed to exhaustion of primary electron donor. Liu et al. (2010)
reported four rounds of 100 mg Cr(VI)/L reduction by resting cells of E. coli in presence of lawsone
indicating the stability and persistence of mediated Cr(VI) reduction.
Figure 3.14 Repeated cycles of Cr(VI) reduction by Bacillus sp. BT1 in presence and absence of
1.0 mM AQS. The Cr(VI) (100 mg/L) was spiked every time after previously added Cr(VI) was
reduced completely.
3.3.16 Effect of AQS on quinone and chromate reductases in the cell-free extract (CFE) of
Bacillus sp. BT1.
Chromate reductases from P. putida, E. coli and Paracoccus denitrificans have been demonstrated
to exhibit quinone reductase activity as well (Ackerley et al., 2004; Gonzalez et al., 2005; Sedlacek
and Kucera 2010). Liu et al. (2009) demonstrated that over expression of quinone reductase in E.
coli JM109 accelerated quinone dependent azo dye AR27 reduction. E. coli JM109 does not reduce
azo dye AR27 in absence of quinone. Since Cr(VI) reduction activity of Bacillus sp. BT1 was found
to be enhanced in presence of quinone, both chromate and quinone reductase activities were
assayed in cells cultured in the presence/ absence of AQS and/ or Cr(VI).
REDUCTION OF Cr(VI)
61
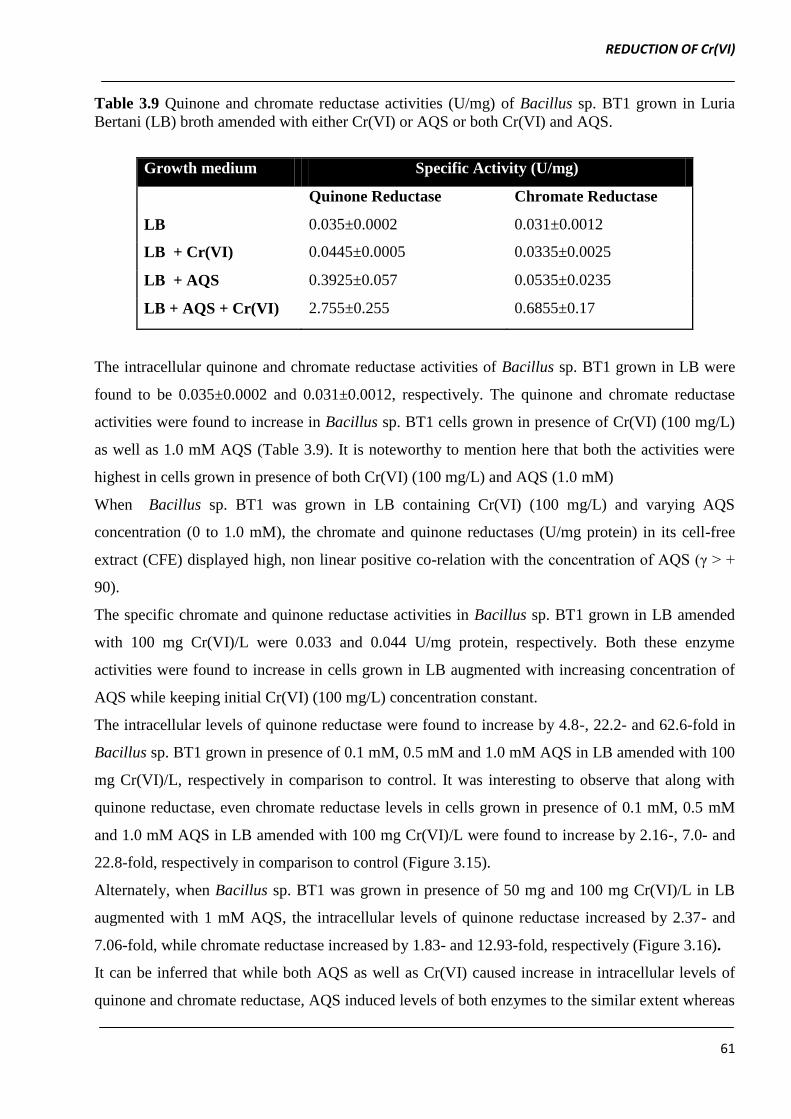
Table 3.9 Quinone and chromate reductase activities (U/mg) of Bacillus sp. BT1 grown in Luria
Bertani (LB) broth amended with either Cr(VI) or AQS or both Cr(VI) and AQS.
Growth medium Specific Activity (U/mg)
Quinone Reductase Chromate Reductase
LB 0.035±0.0002 0.031±0.0012
LB + Cr(VI) 0.0445±0.0005 0.0335±0.0025
LB + AQS 0.3925±0.057 0.0535±0.0235
LB + AQS + Cr(VI) 2.755±0.255 0.6855±0.17
The intracellular quinone and chromate reductase activities of Bacillus sp. BT1 grown in LB were
found to be 0.035±0.0002 and 0.031±0.0012, respectively. The quinone and chromate reductase
activities were found to increase in Bacillus sp. BT1 cells grown in presence of Cr(VI) (100 mg/L)
as well as 1.0 mM AQS (Table 3.9). It is noteworthy to mention here that both the activities were
highest in cells grown in presence of both Cr(VI) (100 mg/L) and AQS (1.0 mM)
When Bacillus sp. BT1 was grown in LB containing Cr(VI) (100 mg/L) and varying AQS
concentration (0 to 1.0 mM), the chromate and quinone reductases (U/mg protein) in its cell-free
extract (CFE) displayed high, non linear positive co-relation with the concentration of AQS (γ > +
90).
The specific chromate and quinone reductase activities in Bacillus sp. BT1 grown in LB amended
with 100 mg Cr(VI)/L were 0.033 and 0.044 U/mg protein, respectively. Both these enzyme
activities were found to increase in cells grown in LB augmented with increasing concentration of
AQS while keeping initial Cr(VI) (100 mg/L) concentration constant.
The intracellular levels of quinone reductase were found to increase by 4.8-, 22.2- and 62.6-fold in
Bacillus sp. BT1 grown in presence of 0.1 mM, 0.5 mM and 1.0 mM AQS in LB amended with 100
mg Cr(VI)/L, respectively in comparison to control. It was interesting to observe that along with
quinone reductase, even chromate reductase levels in cells grown in presence of 0.1 mM, 0.5 mM
and 1.0 mM AQS in LB amended with 100 mg Cr(VI)/L were found to increase by 2.16-, 7.0- and
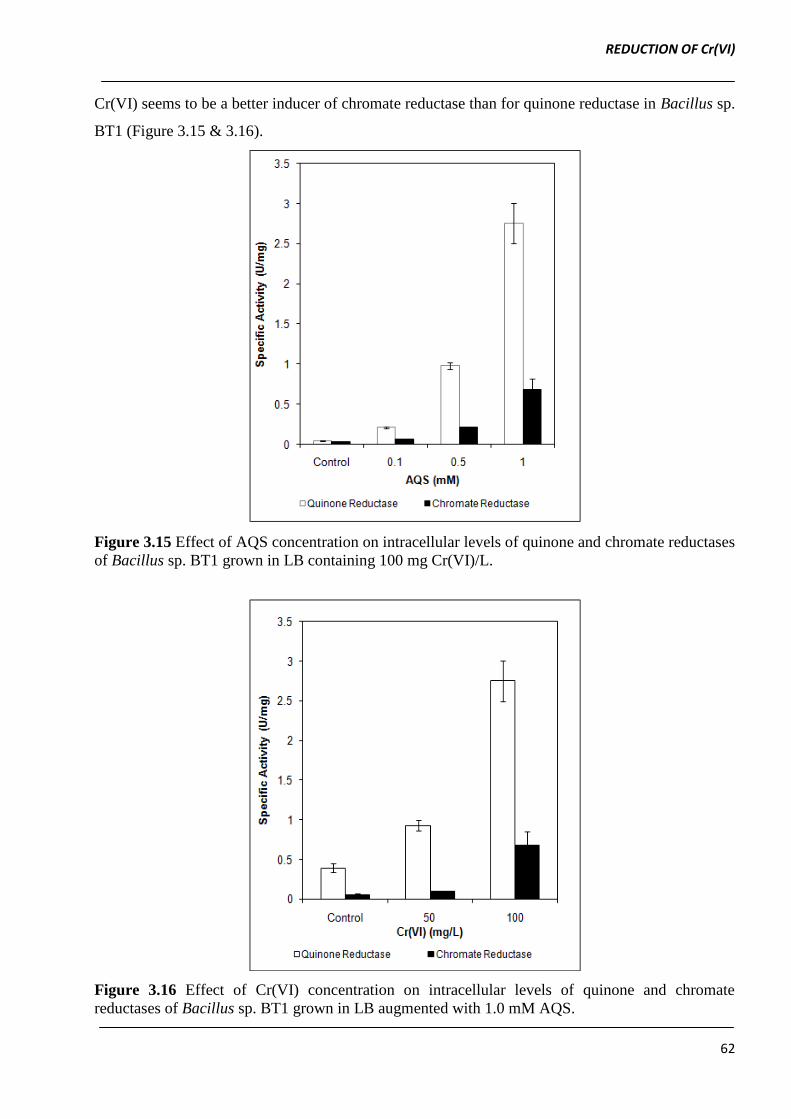
22.8-fold, respectively in comparison to control (Figure 3.15).
Alternately, when Bacillus sp. BT1 was grown in presence of 50 mg and 100 mg Cr(VI)/L in LB
augmented with 1 mM AQS, the intracellular levels of quinone reductase increased by 2.37- and
7.06-fold, while chromate reductase increased by 1.83- and 12.93-fold, respectively (Figure 3.16).
It can be inferred that while both AQS as well as Cr(VI) caused increase in intracellular levels of
quinone and chromate reductase, AQS induced levels of both enzymes to the similar extent whereas
REDUCTION OF Cr(VI)
62
Cr(VI) seems to be a better inducer of chromate reductase than for quinone reductase in Bacillus sp.
BT1 (Figure 3.15 & 3.16).
Figure 3.15 Effect of AQS concentration on intracellular levels of quinone and chromate reductases
of Bacillus sp. BT1 grown in LB containing 100 mg Cr(VI)/L.
Figure 3.16 Effect of Cr(VI) concentration on intracellular levels of quinone and chromate
reductases of Bacillus sp. BT1 grown in LB augmented with 1.0 mM AQS.
REDUCTION OF Cr(VI)
63
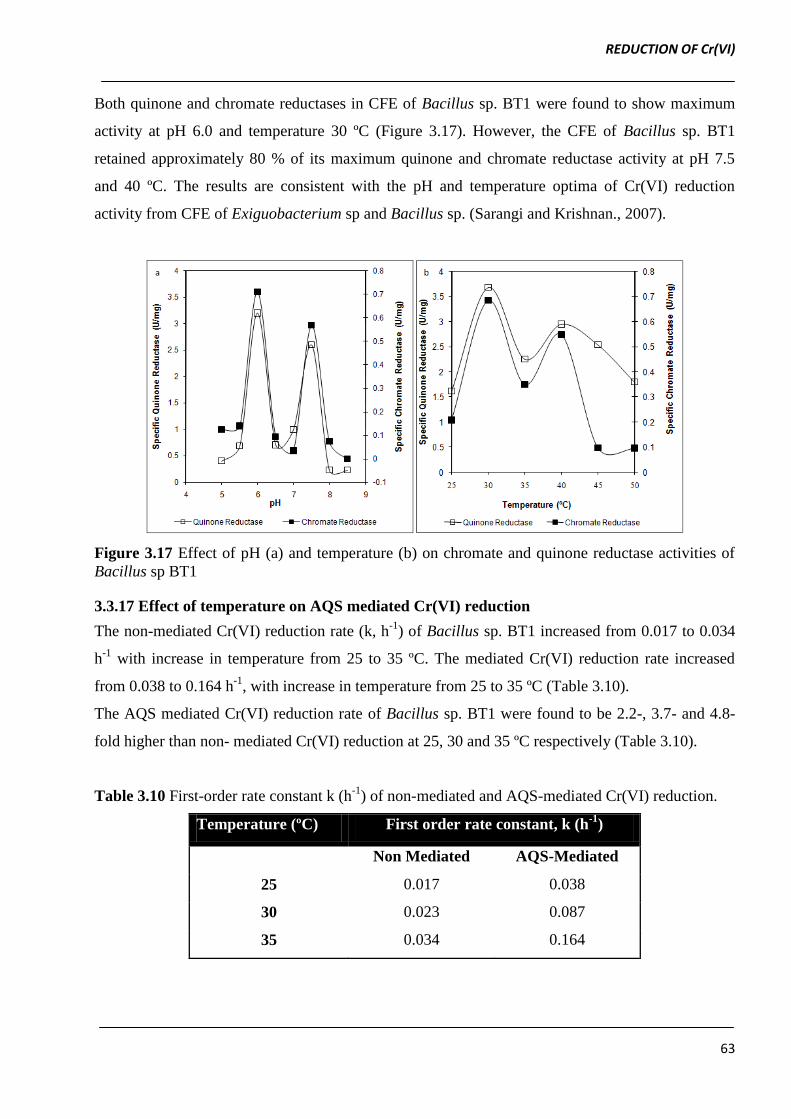
Both quinone and chromate reductases in CFE of Bacillus sp. BT1 were found to show maximum
activity at pH 6.0 and temperature 30 ºC (Figure 3.17). However, the CFE of Bacillus sp. BT1
retained approximately 80 % of its maximum quinone and chromate reductase activity at pH 7.5
and 40 ºC. The results are consistent with the pH and temperature optima of Cr(VI) reduction
activity from CFE of Exiguobacterium sp and Bacillus sp. (Sarangi and Krishnan., 2007).
Figure 3.17 Effect of pH (a) and temperature (b) on chromate and quinone reductase activities of
Bacillus sp BT1
3.3.17 Effect of temperature on AQS mediated Cr(VI) reduction
The non-mediated Cr(VI) reduction rate (k, h-1
) of Bacillus sp. BT1 increased from 0.017 to 0.034
h-1
with increase in temperature from 25 to 35 ºC. The mediated Cr(VI) reduction rate increased
from 0.038 to 0.164 h-1
, with increase in temperature from 25 to 35 ºC (Table 3.10).
The AQS mediated Cr(VI) reduction rate of Bacillus sp. BT1 were found to be 2.2-, 3.7- and 4.8-
fold higher than non- mediated Cr(VI) reduction at 25, 30 and 35 ºC respectively (Table 3.10).
Table 3.10 First-order rate constant k (h-1
) of non-mediated and AQS-mediated Cr(VI) reduction.
Temperature (ºC) First order rate constant, k (h-1
)
Non Mediated AQS-Mediated
25 0.017 0.038
30 0.023 0.087
35 0.034 0.164
REDUCTION OF Cr(VI)
64
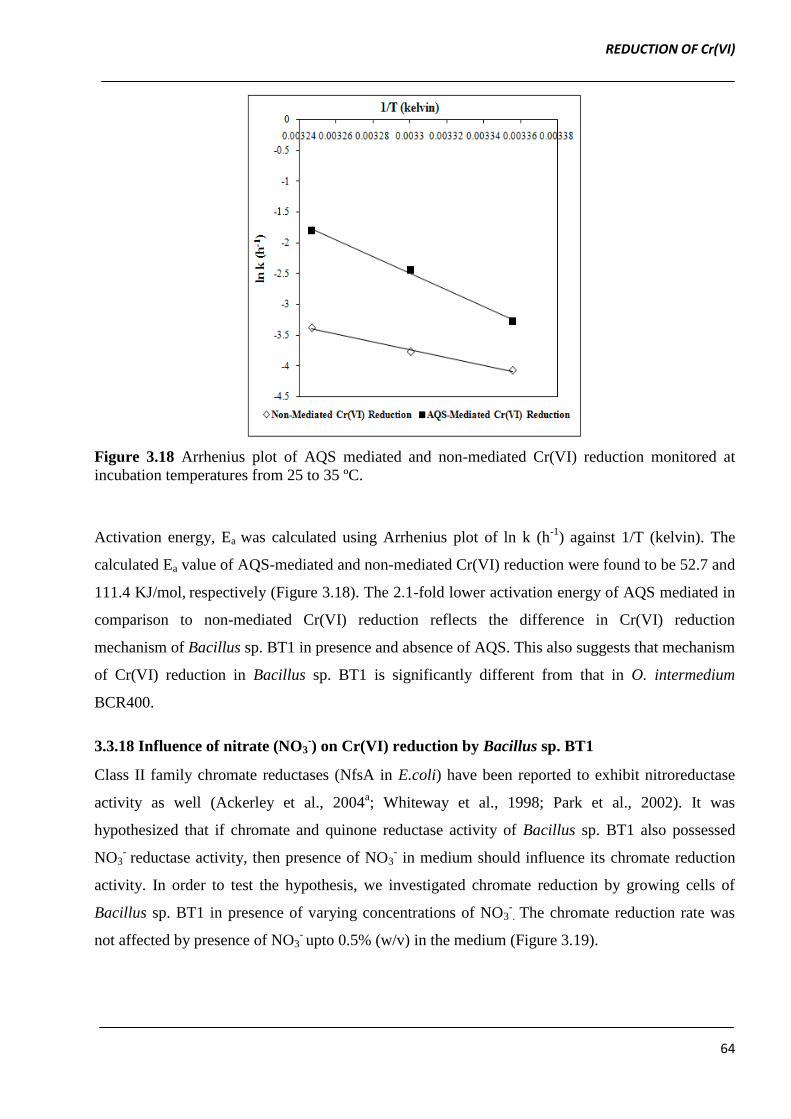
Figure 3.18 Arrhenius plot of AQS mediated and non-mediated Cr(VI) reduction monitored at
incubation temperatures from 25 to 35 ºC.
Activation energy, Ea was calculated using Arrhenius plot of ln k (h-1
) against 1/T (kelvin). The
calculated Ea value of AQS-mediated and non-mediated Cr(VI) reduction were found to be 52.7 and
111.4 KJ/mol, respectively (Figure 3.18). The 2.1-fold lower activation energy of AQS mediated in
comparison to non-mediated Cr(VI) reduction reflects the difference in Cr(VI) reduction
mechanism of Bacillus sp. BT1 in presence and absence of AQS. This also suggests that mechanism
of Cr(VI) reduction in Bacillus sp. BT1 is significantly different from that in O. intermedium
BCR400.
3.3.18 Influence of nitrate (NO3-) on Cr(VI) reduction by Bacillus sp. BT1
Class II family chromate reductases (NfsA in E.coli) have been reported to exhibit nitroreductase
activity as well (Ackerley et al., 2004a; Whiteway et al., 1998; Park et al., 2002). It was
hypothesized that if chromate and quinone reductase activity of Bacillus sp. BT1 also possessed
NO3- reductase activity, then presence of NO3
- in medium should influence its chromate reduction
activity. In order to test the hypothesis, we investigated chromate reduction by growing cells of
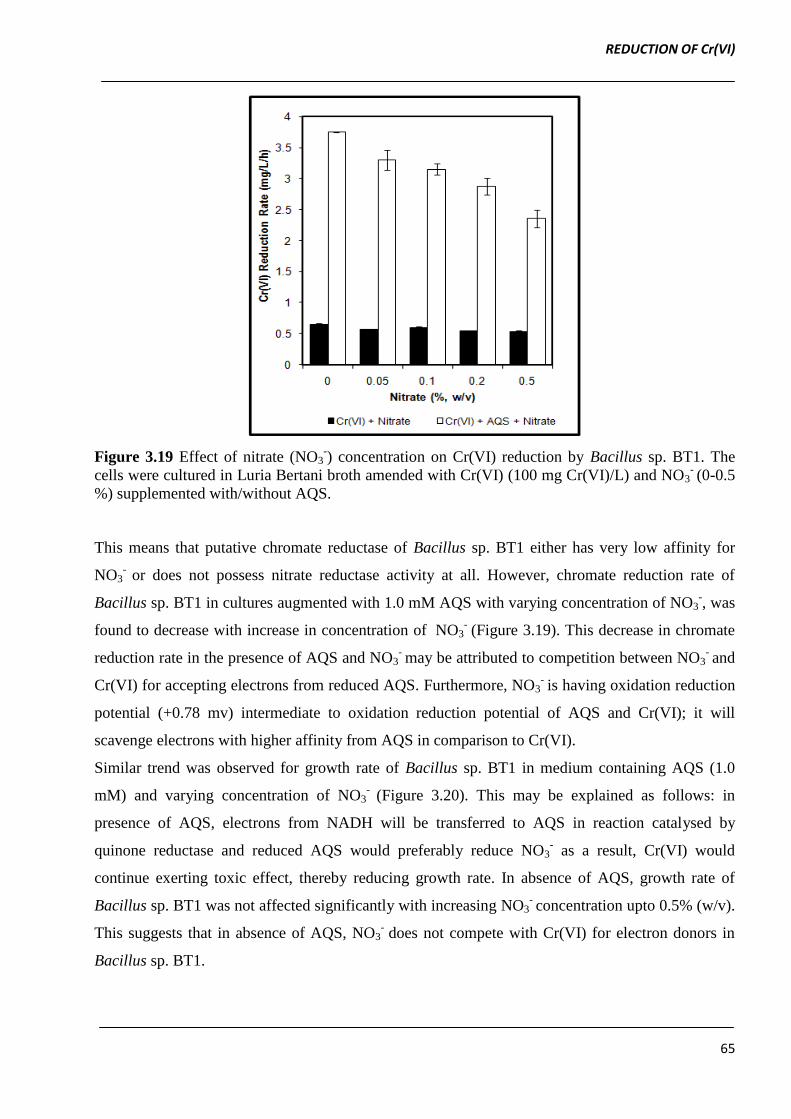
Bacillus sp. BT1 in presence of varying concentrations of NO3-. The chromate reduction rate was
not affected by presence of NO3- upto 0.5% (w/v) in the medium (Figure 3.19).
REDUCTION OF Cr(VI)
65
Figure 3.19 Effect of nitrate (NO3-) concentration on Cr(VI) reduction by Bacillus sp. BT1. The
cells were cultured in Luria Bertani broth amended with Cr(VI) (100 mg Cr(VI)/L) and NO3- (0-0.5
%) supplemented with/without AQS.
This means that putative chromate reductase of Bacillus sp. BT1 either has very low affinity for
NO3- or does not possess nitrate reductase activity at all. However, chromate reduction rate of
Bacillus sp. BT1 in cultures augmented with 1.0 mM AQS with varying concentration of NO3-, was
found to decrease with increase in concentration of NO3- (Figure 3.19). This decrease in chromate
reduction rate in the presence of AQS and NO3- may be attributed to competition between NO3
- and
Cr(VI) for accepting electrons from reduced AQS. Furthermore, NO3- is having oxidation reduction
potential (+0.78 mv) intermediate to oxidation reduction potential of AQS and Cr(VI); it will
scavenge electrons with higher affinity from AQS in comparison to Cr(VI).
Similar trend was observed for growth rate of Bacillus sp. BT1 in medium containing AQS (1.0
mM) and varying concentration of NO3- (Figure 3.20). This may be explained as follows: in
presence of AQS, electrons from NADH will be transferred to AQS in reaction catalysed by
quinone reductase and reduced AQS would preferably reduce NO3- as a result, Cr(VI) would
continue exerting toxic effect, thereby reducing growth rate. In absence of AQS, growth rate of
Bacillus sp. BT1 was not affected significantly with increasing NO3- concentration upto 0.5% (w/v).
This suggests that in absence of AQS, NO3- does not compete with Cr(VI) for electron donors in
Bacillus sp. BT1.
REDUCTION OF Cr(VI)
66
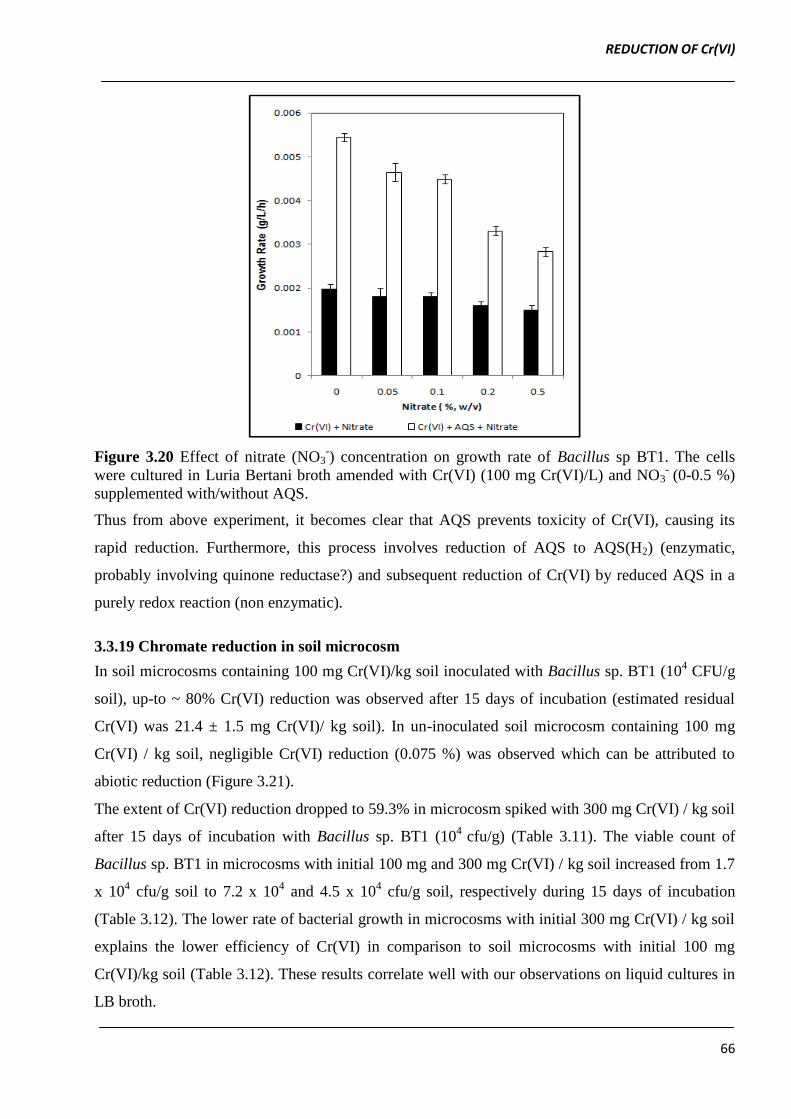
Figure 3.20 Effect of nitrate (NO3-) concentration on growth rate of Bacillus sp BT1. The cells
were cultured in Luria Bertani broth amended with Cr(VI) (100 mg Cr(VI)/L) and NO3- (0-0.5 %)
supplemented with/without AQS.
Thus from above experiment, it becomes clear that AQS prevents toxicity of Cr(VI), causing its
rapid reduction. Furthermore, this process involves reduction of AQS to AQS(H2) (enzymatic,
probably involving quinone reductase?) and subsequent reduction of Cr(VI) by reduced AQS in a
purely redox reaction (non enzymatic).
3.3.19 Chromate reduction in soil microcosm
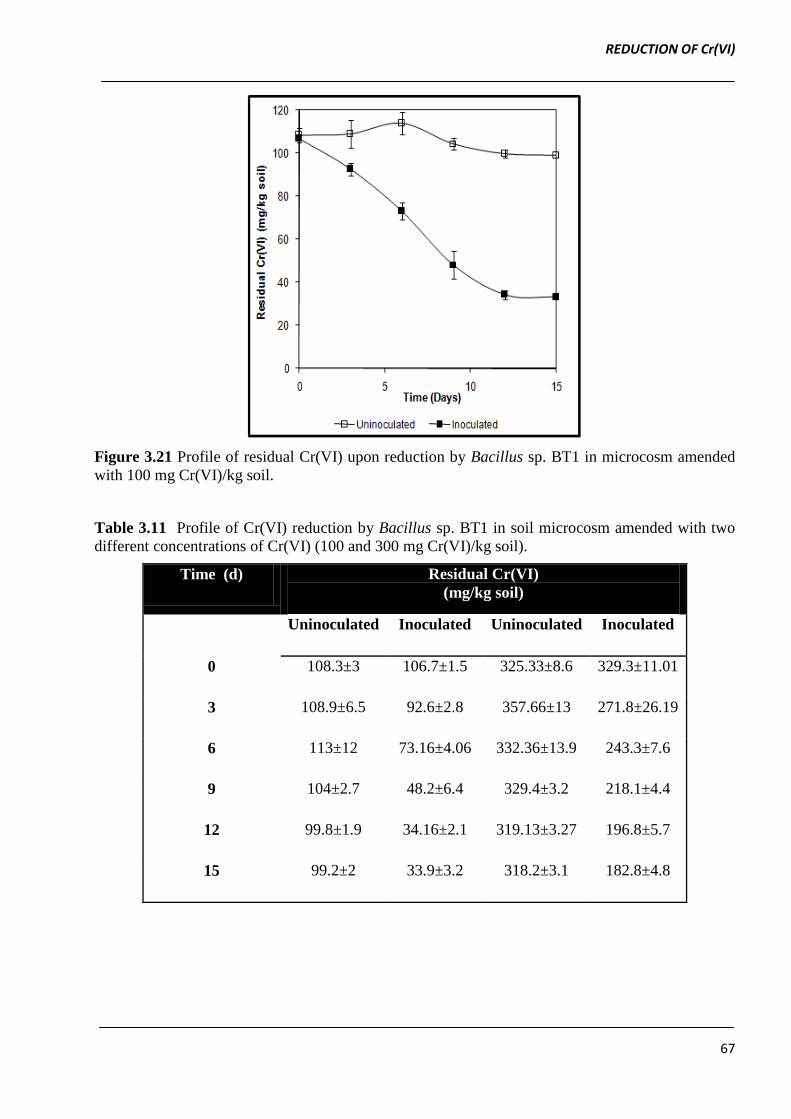
In soil microcosms containing 100 mg Cr(VI)/kg soil inoculated with Bacillus sp. BT1 (104 CFU/g
soil), up-to ~ 80% Cr(VI) reduction was observed after 15 days of incubation (estimated residual
Cr(VI) was 21.4 ± 1.5 mg Cr(VI)/ kg soil). In un-inoculated soil microcosm containing 100 mg
Cr(VI) / kg soil, negligible Cr(VI) reduction (0.075 %) was observed which can be attributed to
abiotic reduction (Figure 3.21).
The extent of Cr(VI) reduction dropped to 59.3% in microcosm spiked with 300 mg Cr(VI) / kg soil
after 15 days of incubation with Bacillus sp. BT1 (104
cfu/g) (Table 3.11). The viable count of
Bacillus sp. BT1 in microcosms with initial 100 mg and 300 mg Cr(VI) / kg soil increased from 1.7
x 104 cfu/g soil to 7.2 x 10
4 and 4.5 x 10
4 cfu/g soil, respectively during 15 days of incubation
(Table 3.12). The lower rate of bacterial growth in microcosms with initial 300 mg Cr(VI) / kg soil
explains the lower efficiency of Cr(VI) in comparison to soil microcosms with initial 100 mg
Cr(VI)/kg soil (Table 3.12). These results correlate well with our observations on liquid cultures in
LB broth.
REDUCTION OF Cr(VI)
67
Figure 3.21 Profile of residual Cr(VI) upon reduction by Bacillus sp. BT1 in microcosm amended
with 100 mg Cr(VI)/kg soil.
Table 3.11 Profile of Cr(VI) reduction by Bacillus sp. BT1 in soil microcosm amended with two
different concentrations of Cr(VI) (100 and 300 mg Cr(VI)/kg soil).
Time (d) Residual Cr(VI)
(mg/kg soil)
Uninoculated Inoculated Uninoculated Inoculated
0 108.3±3 106.7±1.5 325.33±8.6 329.3±11.01
3 108.9±6.5 92.6±2.8 357.66±13 271.8±26.19
6 113±12 73.16±4.06 332.36±13.9 243.3±7.6
9 104±2.7 48.2±6.4 329.4±3.2 218.1±4.4
12 99.8±1.9 34.16±2.1 319.13±3.27 196.8±5.7
15 99.2±2 33.9±3.2 318.2±3.1 182.8±4.8
REDUCTION OF Cr(VI)
68
Table 3.12 Influence of varying Cr(VI) concentration on growth (cfu/g soil) of Bacillus sp. BT1 in
soil microcosms amended with either 100 or 300 mg Cr(VI)/kg soil.
Time (d) Cr(VI) mg/kg soil
100 300
0 1.7 X 104
1.9 X 104
3 1.7 X 104 1.9 X 10
4
9 3.9 X 104 2.1 X 10
4
12 5.4 X 104 3.3 X 10
4
15 7.2 X 104
4.5 X 104
Since, Cr(VI) reduction in Bacillus sp. BT1 was found to be associated with growth we attempted
to stimulate growth by supplementing soil microcosms with malt extract and thereby enhance the
Cr(VI) reduction efficiency. The malt extract was added to mimic nutrients in plant root exudates
and thus simulating microcosm experiment more closer to field conditions (Ellis, 2004). The
augmentation of soil microcosms with malt extract above 5 g/kg soil stimulated growth of Bacillus
sp. BT1 as well as its Cr(VI) reduction efficiency (Table 3.13).
Table 3.13 Influence of varying malt extract concentration on Cr(VI) reduction by Bacillus sp.
BT1 and its growth (cfu/g soil) in soil microcosms amended with 100 mg Cr(VI)/kg soil.
Time
(d)
Malt Extract concentration in microcosms (%, w/w)
0.05 0.1 0.2 0.5 1.0
Cr(VI),
mg/kg
soil
CFU/g
soil
Cr(VI),
mg/kg
soil
CFU/g
soil
Cr(VI),
mg/kg
soil
CFU/g
soil
Cr(VI),
mg/kg
soil
CFU/g
soil
Cr(VI),
mg/kg
soil
CFU/g
soil
0 102.96 1.0x104
102.9 1.4x104 100 1.2x10
4 104.24 1.7x10
4 101.46 1.4x10
4
5 97.7 3.2x104 95.08 8.5x10
4 90 8.6x10
4 93.97 8.9x10
4 86.16 9.1x10
4
10 79.99 8.5x104 67.7 9.4x10
4 52.7 1.0x10
5 52.57 1.2x10
5 33.54 1.2x10
5
15 40.7 1.3x105 27.9 1.5x10
5 17.46 1.6x10
5 9.01 1.7x10
5 8.05 1.7x10
5
REDUCTION OF Cr(VI)
69
3.3.20 Seed germination assay
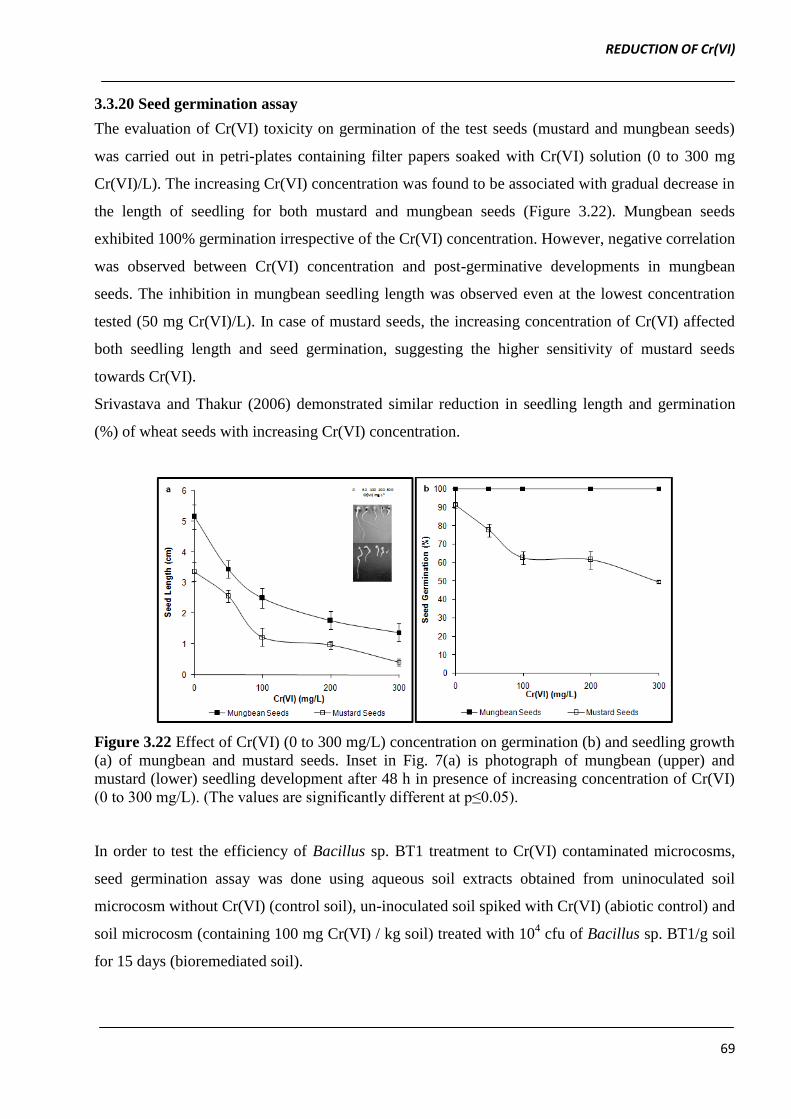
The evaluation of Cr(VI) toxicity on germination of the test seeds (mustard and mungbean seeds)
was carried out in petri-plates containing filter papers soaked with Cr(VI) solution (0 to 300 mg
Cr(VI)/L). The increasing Cr(VI) concentration was found to be associated with gradual decrease in
the length of seedling for both mustard and mungbean seeds (Figure 3.22). Mungbean seeds
exhibited 100% germination irrespective of the Cr(VI) concentration. However, negative correlation
was observed between Cr(VI) concentration and post-germinative developments in mungbean
seeds. The inhibition in mungbean seedling length was observed even at the lowest concentration
tested (50 mg Cr(VI)/L). In case of mustard seeds, the increasing concentration of Cr(VI) affected
both seedling length and seed germination, suggesting the higher sensitivity of mustard seeds
towards Cr(VI).
Srivastava and Thakur (2006) demonstrated similar reduction in seedling length and germination
(%) of wheat seeds with increasing Cr(VI) concentration.
Figure 3.22 Effect of Cr(VI) (0 to 300 mg/L) concentration on germination (b) and seedling growth
(a) of mungbean and mustard seeds. Inset in Fig. 7(a) is photograph of mungbean (upper) and
mustard (lower) seedling development after 48 h in presence of increasing concentration of Cr(VI)
(0 to 300 mg/L). (The values are significantly different at p≤0.05).
In order to test the efficiency of Bacillus sp. BT1 treatment to Cr(VI) contaminated microcosms,
seed germination assay was done using aqueous soil extracts obtained from uninoculated soil
microcosm without Cr(VI) (control soil), un-inoculated soil spiked with Cr(VI) (abiotic control) and
soil microcosm (containing 100 mg Cr(VI) / kg soil) treated with 104 cfu of Bacillus sp. BT1/g soil
for 15 days (bioremediated soil).
REDUCTION OF Cr(VI)
70
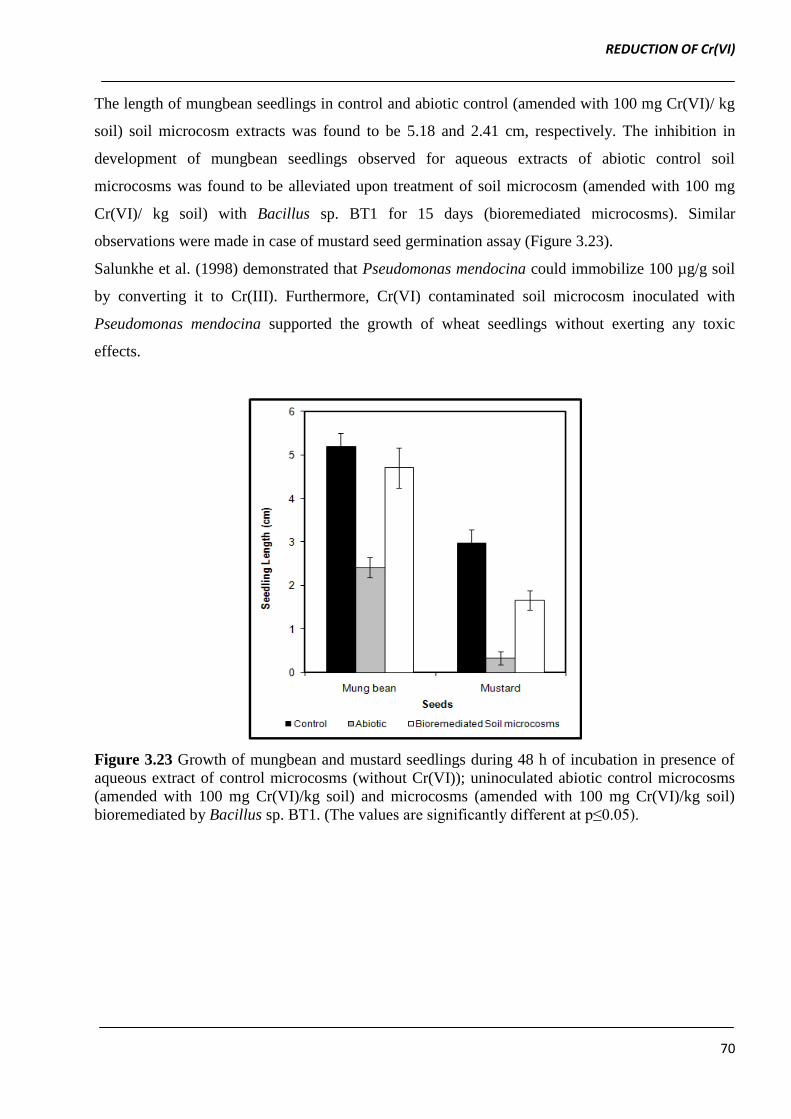
The length of mungbean seedlings in control and abiotic control (amended with 100 mg Cr(VI)/ kg
soil) soil microcosm extracts was found to be 5.18 and 2.41 cm, respectively. The inhibition in
development of mungbean seedlings observed for aqueous extracts of abiotic control soil
microcosms was found to be alleviated upon treatment of soil microcosm (amended with 100 mg
Cr(VI)/ kg soil) with Bacillus sp. BT1 for 15 days (bioremediated microcosms). Similar
observations were made in case of mustard seed germination assay (Figure 3.23).
Salunkhe et al. (1998) demonstrated that Pseudomonas mendocina could immobilize 100 µg/g soil
by converting it to Cr(III). Furthermore, Cr(VI) contaminated soil microcosm inoculated with
Pseudomonas mendocina supported the growth of wheat seedlings without exerting any toxic
effects.
Figure 3.23 Growth of mungbean and mustard seedlings during 48 h of incubation in presence of
aqueous extract of control microcosms (without Cr(VI)); uninoculated abiotic control microcosms
(amended with 100 mg Cr(VI)/kg soil) and microcosms (amended with 100 mg Cr(VI)/kg soil)
bioremediated by Bacillus sp. BT1. (The values are significantly different at p≤0.05).

REDUCTION OF Cr(VI)
71
3.3 Summary
The physiological characterization of Cr(VI) reduction by O. intermedium and Bacillus sp. BT1
indicated that both the bacterial strains could reduce 100 mg Cr(VI)/L completely in Luria Bertani
broth at optimum pH 7.0 and temperature 30 to 37 ºC. The Cr(VI) reduction by both O.
intermedium and Bacillus sp. BT1 remained either un-affected or slightly affected in presence of
co-contaminant metal ions.
The efficiency and rate of Cr(VI) reduction of both the bacterial strains was enhanced in presence of
AQS. The higher efficiency of Cr(VI) reduction in presence of AQS was accompanied with higher
intracellular levels of quinone and chromate reductase.
Hence, a hypothetical model for AQS mediated two-step Cr(VI) reduction by O. intermedium
BCR400 and Bacillus sp. BT1 has been proposed. In the first-step, AQS may be reduced to AQH2S
(hydra-quinone) by quinone reductase. In the second-step, the AQH2S reduces Cr(VI) by 2e-
transfer, thereby reducing it to Cr(IV) in a purely redox chemical reaction. The direct reduction of
Cr(VI) to Cr(IV) by AQH2S would also prevent ROS generation, thereby exert protective effect on
the growth of O. intermedium BCR400. The further reduction of Cr(IV) to Cr(III) may be mediated
by single electron transfer catalysed by specific chromate reductase or non-specific reductases. The
model is based on several lines of evidences:
1. The supplementation of AQS to nutrient growth medium amended with Cr(VI), not only had
protective effect against toxicity due to Cr(VI) but also enhanced the rate of Cr(VI)
reduction by both O. intermedium BCR400 and Bacillus sp. BT1.
2. Increase in both quinone and chromate reductase activity (U/mg of total proteins) was
observed in cell free lysate prepared from O. intermedium BCR400 and Bacillus sp. BT1
grown in presence of either AQS or Cr(VI)+AQS.
3. The cell-free extract (CFE) of both the bacterial strains grown in presence of variable AQS
concentration (0 to 1mM) displayed high, non linear positive correlation between the
concentration of AQS and activities of both quinone reductase and chromate reductase (γ >
+ 90).
Finally, the soil microcosm studies with Bacillus sp. BT1 demonstrated its potential application in
bioremediation of Cr(VI) contaminated soils

REDUCTION OF Cr(VI)
72
Reference
Ackerley Df., Gonzalez CF., Keyhan M., Blake R. and Matin A. 2004a. Mechanism of chromate
reduction by the Escherichia coli, NfsA, and the role of different chromate reductases in
minimizing oxidative stress during chromate reduction. Environmental Microbiology 6:851-
860.
Ackerley DF., Gonzalez CF., Park CH., Blake R., Keyhan M. and Matin A. 2004b. Chromate
reducing properties of soluble flavoproteins from Pseudomonas putida and Escherichia coli.
Applied and Environmental Microbiology. 70:873-882.
Agrawal A., Kumar V. and Pandey BD. 2008. Remediation options for the treatment of
electroplating and leather tanning effluent containing chromium-A Review. Mineral
Processing Extractive Metallurgy Reviews. 272:99-130.
Bond DR. and Lovley DR. 2002. Reduction of Fe(III) oxide by methanogens in the presence and
absence of extracellular quinones. Environmental Microbiology. 4:115-124.
Branco R., Alpoim MC. and Morais PV. 2004. Ochrobactrum tritici strain 5bvI1-characterization of
a Cr(VI) resistant and Cr(VI) reducing strain. Canadian Journal of Microbiology. 50:697-
703.
Burgos W., Fang Y., Royer RA., Yeh GT., Stone JJ., Jeonn BH. and Dempsey BA. 2003. Reaction
based modelling of quinone mediated bacterial iron(III) reduction. Geochemica et
Cosmochimica Acta. 67:2735-2748.
Camargo FAO., Okeke BC., Bento FM. and Frankenberger WT. 2004. Hexavalent chromium
reduction by immobilized cells and cell free-extracts of Bacillus sp. ES 29. Bioremediation
Journal. 8:23-30.
Camargo FAO., Okeke BC., Bento FM and Frankenberger WT. 2003. In-vitro reduction of
hexavalent chromium by a cell-free extract of Bacillus sp. ES 29 stimulated by Cu2+
.
Applied Microbiology and Biotechnology. 62:569-573.
Campos J., Martinez PM. and Cerventes C. 1995. Hexavalent chromium reduction by chromate
resistant Bacillus sp. strain. Antonie Van Leeuwenhoek. 68:203-208.
Cheung KH. and Gu JD. 2005. Chromate reduction by Bacillus megaterium TKW3 isolated from
marine sediments. World Journal of Microbiology and Biotechnology. 16:855-862.
Clark DP. 1994. Chromate reductase activity of Enterobacter aerogenes is induced by nitrite.
FEMS Microbiology Letters. 122:233-238.

REDUCTION OF Cr(VI)
73
Ellis RJ. 2004. Artificial soil microcosms: a tool for studying microbial autecology under controlled
conditions. Journal of Microbiological Methods. 56:287-290.
Elongovan R., Abhipsa S., Rohit B., Ligy P. and Chandraraj K. 2006. Reduction of Cr(VI) by
Bacillus sp. Biotechnology Letters. 28:247-252.
Elongovan R., Philip L. and Chandraraj K. 2008. Hexavalent chromium reduction by free and
immobilized cell-free extract of Arthrobacter rhombi-RE. Applied Biochemistry and
Biotechnology. 160:81-97.
Fredrickson JK., Kostandarithes HM., Li SW., Plymale AE. and Daly MJ. 2006. Reduction of
Fe(III), Cr(VI), U(VI) and Tc(VII) by Deinococcus radiodurans R1. Applied and
Environmental Microbiology. 66:2006-2011.
Ganguli A. and Tripathi AK. 2002. Bioremediation of toxic chromium from electroplating effluent
by chromate reducing Pseudomonas aeroginosa A2Chr in two bioreactors. Applied
Microbiology and Biotechnology. 58:416-420.
Garbisu C., Alkorta I., Llama MJ. and Serra JL. 1998. Aerobic chromate reduction by Bacillus
subtilis. Biodegradation. 9:133-141.
Gonzalez CF., Ackerley DF., Lynch SV. and Matin A. 2005. ChrR, a soluble quinone reductase of
Pseudomonas putida that defends against H2O2. The Journal of Biological Chemistry.
280:22590-22595.
Gonzalez CF., Ackerley DF., Park CH. and Matin A. 2003. A soluble flavoprotein contributes to
chromate reduction and tolerance by Pseudomonas putida. Acta Biotechnologica. 23:233-
239.
He Z., Gao F., Sha T., Hu Y. and He C. 2009. Isolation and characterization of a Cr(VI)-reduction
Ochrobactrum sp. strain CSCr-3 from chromium landfill. Journal of Hazardous Materials.
163:869-873.
Hussein H., Farag S., Kandil K. and Moawad H. 2005. Tolerance and uptake of heavy metals by
Pseudomonads. Process Biochemistry. 40:955-961.
Ishibashi Y., Cerventes C. and Silver S. 1990. Chromium reduction in Pseudomonas putida.
Applied and Environmental Microbiology. 6:2268-2270.
Kwak YH., Lee DS. and Kim HB. 2003. Vibrio harveyi nitroreductase is also a chromate reductase.
Applied and Environmental Microbiology. 69:4390-4395.

REDUCTION OF Cr(VI)
74
Ling J., Hong LU., Jiti Z. and Jing W. 2009. Quinone mediated decolourization of sulfonated azo
dyes by cells and cell extracts from Sphingomonas xenophaga. Journal of Environmental
Sciences. 21:503-508.
Liu YG., Xu WH., Zeng GM., Li X. and Gao H. 2006. Cr(VI) reduction by Bacillus sp. isolated
from chromium landfill. Process Biochemistry. 41: 1981-1986.
Liu G., Yang H., Wang J., Jin R., Zhou J. and Lv H. 2010. Enhanced chromate reduction by resting
Escherichia coli cells in the presence of quinone redox mediators. Bioresource Technology.
101:8127-8131.
Liu G., Zhou J., Wang J., Zhou M., Lu H. and Jin R. 2009. Acceleration of azo dye decolorization
by using quinone reductase activity of azoreductase and quinone redox mediator.
Bioresource Technology. 100:2781-2795.
Lovley DR., Fraga JL., Blunt-Harris EL., Hayes LA., Phillips EJP. and Coates JD. 1998. Humic
substance as a mediator for microbially catalyzed metal reduction. Acta Hydrochimica Et
Hydrobiologica. 26:152-157.
Lovley DR., Fraga JL., Coates JD. and Blunt-Harris EL. 1999. Humics as an electron donor for
anaerobic respiration. Environmental Microbiology. 1:89-98.
Lowe KL., Straube W., Little B. and Meehan J. 2003. Aerobic and anaerobic reduction of Cr(VI) by
Shewanella oneidensis: Effects of cationic metals, sorbing agents and mixed microbial
cultures. Acta Biotechnologica. 23:161-178.
Lowry OH., Rosebroough NJ., Farr AL. and Randall RJ. 1951. Protein measurements with folin
phenol reagents. The Journal of Biological Chemistry. 193:265-275.
Maithreepala RA. and Doong R. 2009. Transformation of carbon tetrachloride by biogenic iron
species in the presence of Geobacter sulfurreducens and electron shuttles. Journal of
Hazardous Materials. 164:337-344.
Mazoch J., Tesar R., Sedlacek V., Kucera I. and Turanek J. 2004. Isolation and biochemical
characterization of two soluble iron (III) reductases from Paracoccus denitrificans.
European Journal of Biochemistry. 271:553-562.
McLean J. and Beveridge TJ. 2001. Chromate reduction by a Pseudomonad isolated from a site
contaminated with chromate copper arsenate. Applied and Environmental Microbiology.
67:1076-1084.
Megharaj M., Avudainayagam S. and Naidu R. 2003. Toxicity of hexavalent chromium and its
reduction by bacteria isolated from soil contaminated with tannery waste. Current
Microbiology. 27:51-54.

REDUCTION OF Cr(VI)
75
Michel C., Brugna M., Aubert C., Bernadac A. and Brushi M. 2001. Enzymatic reduction of
chromate: comparative studies using sulphate reducing bacteria. Key role of polyheme
cytochrome c and hydrogenases. Applied Microbiology and Biotechnology. 55:95-100.
Ozdemir G., Ozturk T., Ceyhan N., Isler R. and Cosar T. 2003. Heavy metal biosorption by biomass
of Ochrobactrum anthropi producing exopolysaccharide in activated sludge. Bioresource
Technology. 90:71-74.
Park CH., Gonzalez CF., Ackerley DF., Keyhan M. and Matin A. 2002. Molecular engineering of
soluble bacterial proteins with chromate reductase activity. In Proceedings of the 1st
International Conference on Remediation of Contaminated Sediments, Venice, Italy, Vol.
III. Hinchee, R.E.,Porta, A., and Pellei M. (eds). Columbus, OH: Batelle Press, pp. 103–111.
Puzon GJ., Petersen JN., Roberts AG., Kramer DM. and Xun L. 2002. A bacterial flavin reductase
system reduces chromate to a soluble chromium (III)-NAD+ complex. Biochemical and
Biophysical Research Communications. 294:76-81.
Rau J., Knackmuss HJ. and Stolz A. 2002. Effects of different quinoid redox mediators on the
anaerobic reduction of azo dyes by bacteria. Environmental Science and Technology.
36:1497-1504.
Rau J. and Stolz A. 2003. Oxygen- insensitive nitroreductases NfsA and NfsB of Escherichia coli
function under anaerobic conditions as lawsone dependent azo reductases. Applied and
Environmental Microbiology. 69:3448-3455.
Rehman A., Zahoor A., Muneer B. and Hasnain S. 2008. Chromium tolerance and reduction
potential of a Bacillus sp. ev3 isolated from metal contaminated wastewater. Bulletin of
Environmental Contamination and Toxicology. 81:25-29.
Romanenko VI. and Korenkov VN. 1977. A pure culture of bacterial cells assimilating chromates
and bichromates as hydrogen acceptors when grown under anaerobic conditions.
Mikrobiologiya. 46:414-417.
Salunkhe PB., Dhakephalkar PK. and Paknikar KM. 1998. Bioremediation of hexavalent chromium
in soil microcosms. Biotechnology Letters. 20:749-751.
Santos ABD., Bisschops IAE., Cerventes FJ. and Lier JBV. 2004a. Effect of different redox
mediators during thermophillic azo dye reduction by anaerobic granular sludge and
comparative study between mesophillic (30 ºC) and thermophillic (55 ºC) treatments for
decolourization of textile wastewaters. Chemosphere. 55:1149-1157.
Santos ABD., Cervantes FJ. and Lier JB. 2004b. Azo dye reduction by thermophilic anaerobic
granular sludge and the impact of the redox mediator anthraquinone - 2, 6 - disulfonate

REDUCTION OF Cr(VI)
76
(AQDS) on the reductive biochemical transformation. Applied Microbiology and
Biotechnology. 64:62-69.
Sarangi A. and Krishnan C. 2008. Comparison of in vitro Cr(VI) reduction by CFEs of chromate
resistant bacteria isolated from chromate contaminated soil. Bioresource Technology.
99:4130-4137.
Sedlacek V. and Kucera I. 2010. Chromate reductase activity of the Paracoccus denitrificans ferric
reductase B (FerB) protein and its physiological relevance. Archives of Microbiology.
192:919-926.
Sendelbach LE. 1989. A review of the toxicology and carcinogenicity of anthraquinone derivatives.
Toxicology. 57:227-240.
Shen H. and Wang YT. 1993. Characterization of enzymatic reduction of hexavalent chromium by
Escherichia coli ATCC 33456. Applied and Environmental Microbiology. 59:3771-3777.
Srivastava S. and Thakur IS. 2006. Evaluation of bioremediation and detoxification potentiality of
Aspergillus niger for removal of hexavalent chromium in soil microcosm. Soil Biology
Biochemistry. 38: 1904-1911.
Sultan S. and Hasnain S. 2006. Characterization of an Ochrobactrum intermedium strain STCr-5
manifesting high level Cr(VI) resistance and reduction potential. Enzyme and Microbial
Technology. 39:833-888.
Sultan S. and Hasnain S. 2007. Reduction of toxic hexavalent chromium by Ochrobactrum
intermedium strain SDCr - 5 stimulated by heavy metals. Bioresource Technology. 98:340-
344.
Suzuki T., Miyata N., Horitsu H., Kawai K., Takamizawa K., Tai Y. and Okazaki M. 1992.
NAD(P)H-dependent chromium(VI) reductase of Pseudomonas ambigua G-1: a Cr(V)
intermediate is formed during the reduction of Cr(VI) to Cr(III). Journal of Bacteriology.
174:5340-5345.
Thacker U. and Madamwar D. 2005. Reduction of toxic chromium and partial localization of
chromium reductase activity in bacterial isolate DM1. World Journal of Microbiology and
Biotechnology. 21:891-899.
Timmis KN. and Pieper DH. 1999. Bacteria designed for bioremediation. Trends in Biotechnology.
17:200-204.
Wang PC., Mori T., Komoril K., Sasatsu M., Toda K. and Ohtake H. 1989. Isolation and
characterization of an Enterobacter cloacae strain that reduces hexavalent chromium under
anaerobic conditions. Applied and Environment Microbiology. 55:1665-1669.

REDUCTION OF Cr(VI)
77
Wang PC., Mori T., Toda K. and Ohtake H. 1990. Membrane-associated chromate reductase
activity from Enterobacter cloacae. Journal of Bacteriology. 172:1670-1672.
Wang Y., Wu C., Wang X. and Zhou S. 2009. The role of humic substances in the anaerobic
reductive dechlorination of 2,4-dichlorophenoxyacetic acid by Comamonas korensis strain
CY01. Journal of Hazardous Materials. 164:941-947.
Whiteway J., Koziarz P., Veall J., Sandhu N., Kumar P. and Hoecher B. 1998. Oxygen-insensitive
nitroreductases: analysis of the roles of nfsA and nfsB in development of resistance to 5-
nitrofuran derivatives in Escherichia coli. Journal of Bacteriology. 180:5529-5539.
Zee FPVD. and Cervantes FJ. 2009. Impact and application of electron shuttles on the redox (bio)
transformation of contaminants: A Review. Biotechnology Advances. 27:256-277.