Embed Size (px)
Citation preview

140
Chapter 6
Stress Removal and In Vitro Wound Healing Activity of
Peptides C2 and E1
6.1 INTRODUCTION
ROS are required by living systems to maintain a number of functions including
signal transduction, cell adhesion and wound healing (Park et al 2011). When the delicate
balance between the ROS creation and depletion is hampered due to excess ROS or faulty
detoxification mechanism, oxidative stress ensues. Excess ROS acts through multiple
mechanisms to cause stress: oxidizes cellular macromolecules leading to membrane
damage, enzyme dysfunction and metabolic flux hamper and impaired DNA repair,
resulting in mutagenesis or cell death (Lum and Roebuck, 2001).
During wound repair, reactive radicals created by neutrophils in the second phase
of wound healing serves as signals for other group of cells to initiate steps for the third
and fourth phase of wound healing. However, in case of persistent wounds or the
presence of foreign elements near a wound site, the resultant uncontrolled stress, ROS or
otherwise, can lead to deleterious effects. Although very low levels of free radicals can
actually increase the integrin attachment to ECM, the effect is reversed at higher levels.
Excess ROS displays a multitude of effects including altered integrin sub-unit gene
expression, consequently leading to improper connectivity with the ECM and eventually,
cell death (Lamari et al 2007; Mian et al 2008). Oxidative stress also results in
reorganization of actin filaments through FAK, resulting in loss of cell-matrix
adhesiveness. This may eventually lead to cell loss, compromised tissue integrity,
activation of apoptotic pathways and pathologic consequences (Zhou et al 1999).
It is hypothesized that this cell loss in the presence of ROS stress can be
countered by an efficient cell adhesion. Thus, components of the matrix with an increased
cell adhesive ability could provide some form of protection to stress-exposed cells. The
increased cell survival, in turn can also have a substantial effect on cell movement during
in vivo stress-generating events like tissue remodelling and wound closure.
The physiological effects of oxidative stress in vitro can be mimicked by the use
of exogenously administered H2O2 and by heavy metals. ROS stress can be generated by
metals through two distinct mechanisms: With redox-active metals like Fe and Cr, a

141
Fenton-like reaction produces free radicals while redox-inactive toxic metals such as Hg
and Cd act through depletion of major antioxidant components leading to unquenched
free radicals (Ercal et al 2001). For this study, oxidative stress has been created via three
systems; subjecting cells to H2O2, a combination of Fe/ H2O2 and through heavy metals.
Two cryptic peptides isolated earlier; E1 (Chapter 3 and 5) and C2 (Chapter 5),
both displaying cell adhesion abilities was used as possible „stress-relieving‟ agents. The
present study was carried out to determine the cyto-protective and de-stressor activity of
the these peptides in countering stress generated by ROS along with the ability to affect
in vitro wound healing properties.
6.2 MATERIALS AND METHODS
T-flasks (Nunclon surface) were procured from Nunc, Roskilde, Denmark and
disposable culture dishes (35×10mm) were obtained from Fischer Scientific, Hanover
park, IL, USA. DMEM supplemented with 2mM glutamine and 10X antibiotic-
antimycotic solution were obtained from HiMedia, India. Foetal bovine serum and 10X
sterile filtered trypsin-EDTA solution was obtained from Sigma-Aldrich. Vero and HeLa
cell lines were procured from National Centre for Cell Science, Pune, India. All bench
work associated with cell lines, including peptide coating have been carried out inside a
class II bio-safety cabinet (Clean Air Systems, Chennai, India) for maintaining sanitized
conditions.
6.2.1 PEPTIDE ISOLATION AND COATING
The peptides E1 and C2 were isolated as described in Chapter 3 and 5
respectively. The purified peptides were coated onto the disposable dishes according to
standard procedures described in Chapter 5. A coating density of 0.507nmole cm-2
area,
known to display maximal cell adhesion has been used throughout this study.
6.2.2 CELL MAINTENANCE
Vero and HeLa cell lines were maintained as mentioned in section 5.2.1.2,
Chapter 5. For experiments, cells reaching 80% confluence were detached from T-flasks
with trypsin-EDTA, centrifuged and cell number enumerated by Neuber‟s chamber. The
viability was tested by trypan blue exclusion assay.

142
6.2.3 HYDROXYL RADICAL SCAVENGING ASSAY
The hydroxide radical scavenging assay was performed according to Zhang et al
(2010). A known quantity of peptides C2 and E1 were dissolved in 20mM KH2PO4-KOH
buffer, pH 7.5 in various dilutions ranging from 0.1-100mM. To 0.1ml of each solution,
the following reagents were added in order; 0.1ml of 1mM EDTA, 10mM H2O2, 75mM
2-deoxy-D-ribose, 2mM ascorbic acid and 1mM FeCl3. The reaction mixture was
incubated for 1h at 37°C and stopped by addition of 0.25ml of 20% TCA. For colour
development, 1ml of 1% TBA was added and the tubes with the reaction mixture were
placed in a boiling water bath for 15min. Absorbance was measured at 532nm after
cooling to room temperature. For blank, water was used instead of FeCl3 in a similar
reaction mixture. BSA and bovine tendon collagen were used as „negative control‟.
Collagen hydrolysate obtained after proteolysis was used as „test control‟ and BHT was
used as the „positive control‟. Assays were done in triplicates. Scavenging ability was
calculated from the following equation:
Radical scavenging activity (%) = 1001
Control
Test
Test is the absorbance of the test peptide samples and Control is the absorbance without
the peptides.
6.2.4 DE-STRESSOR ACTIVITY OF THE PEPTIDES
Stress was generated by five different stress-creating agents as listed in Table 6.1.
Concentrations of the stressors used were chosen from previously reported toxicity levels
(Levis and Majone 1979; Houot et al 2001; Hultberg et al 2001; Jungas et al 2002). Metal
stock solutions were prepared in deionized water and sterilized by filtration through
0.2µm filter. At the time of treatment, stock solutions were diluted in pre-warmed culture
medium to the final concentration required. For agents 4 and 5, stock H2O2 was stored in
4°C and diluted in 0.2M sterilized PBS before adding to the medium.
143
Table 6.1 List of agents used as stressors (*- concentration given in µM).
Agent Stressor Range of final concentrations (mM)
1 Cr (VI) 0 0.01 0.1 1 10 100
2 Fe (II) 0 0.01 0.1 1 10 100
3 Hg (II) 0 1.5625* 3.125
* 12.5
* 25
* 50
*
4 H2O2 0 2.5 5 10 25 100
5 Fenton‟s
reagent
Fe (II) 0 0.1 5 10 50 100
H2O2 0 0.01 0.1 1 10 50
Table 6.2 Experimental set-up for in vitro wound healing studies for three conditions;
in the absence of a stressor, in the presence of stressor and in the presence of both stressor
and de-stressor.
Set I
C2 coated E1 coated Collagen coated Uncoated
Set II
C2 coated +
H2O2
E1 coated +
H2O2
Collagen coated
+ H2O2
Uncoated +
H2O2
Set III
C2 coated +
E1 dissolved +
H2O2
E1 coated +
E1 dissolved +
H2O2
Collagen coated
+ E1 dissolved +
H2O2
Uncoated +
E1 dissolved
+ H2O2

144
6.2.4.1 ASSAYING DE-STRESSOR ACTIVITY IN COATED FORM
3.5×106
cells were seeded onto the coated dishes followed immediately by
exposing them to stressor agents 1-5 as per the final concentration given in Table 6.1.
The total volume was kept constant at 1.5ml. Cells seeded onto CCS dishes without
exposure to any stress was used as the control cell count. After 6h incubation, the adhered
cells were trypsinized and counted by haemocytometry.
6.2.4.2 ASSAYING DE-STRESSOR ACTIVITY IN DISSOLVED FORM
3.5×106
cells suspended in DMEM were seeded on CCS dishes and subjected to
the same level of stress as in section 6.2.4.1. Based upon activity of E1 in section 3.3.3.2,
Chapter 3, 0.0815µmoles (0.2mg ml-1
) of the active peptides E1 and C2 were solubilized
separately in 0.5ml medium and added to the cells. Care was taken to adjust the added
amount of stressors and cells such that the final concentration and volume to be the same
as before. After 6h incubation, adherent cells were counted and stressor concentration
ensuing 50% cell survival was calculated.
6.2.5 WOUND CLOSURE ASSAY
The wound closure assay was performed according to the protocols given by
Liang et al (2007). 3.5×105cells were seeded onto C2 and E1 coated dishes. An equal
number of cells were seeded onto collagen coated (positive control) and uncoated dishes
(negative controls). When cells reached 90-95% confluence, two parallel scratch wounds
of approximately 400µm diameter were made with a pipette tip in all the dishes. The
medium was replenished for all the dishes with certain changes as given in Table 6.2.
10mM H2O2 was added in set I, 10mM H2O2 along with E1 in dissolved form at a
concentration of 0.0815µmole/ml (based upon activity of E1 in section 6.2.4.2) was
added in set II and set III was incubated „as is‟. Images of a fixed area of the wound were
taken at regular intervals over the course of 24h. Image analysis was done by ImageJ
software (http://rsbweb.nih.gov/ij/) from the National Institute of Health, USA.
The wound-edge positions of the cells were averaged by digitally drawing lines. The area
of the wound was calculated as a rectangle, whose length was taken to be the length of
the wound in focus and breadth the average distance between the two advancing edges of
the wound. The decrease in this rectangular area with time was calculated by measuring
the decreasing distance between the advancing edges. Finally, the total was subtracted

145
from the original area to determine the area covered and expressed in % (Valster et al
2005).
6.2.6 STATISTICAL ANALYSIS
The assays were done in triplicates and activities reported as mean ± standard
deviation. Duplicate dishes were used for the wound healing migration assay. Larger
datasets were analyzed for statistical significance using one way and two-way ANOVA.
Comparison between two groups was accomplished by post-hoc Tukey‟s test and
student‟s t-test. P values less than 0.05 were considered significant.
6.3 RESULTS AND DISCUSSIONS
6.3.1 HYDROXYL RADICAL SCAVENGING ASSAY
E1 has already been confirmed in Chapter 3 as a moderately strong antioxidative
agent with radical scavenging, metal chelation and reductive ability. Since most ROS
stress generates •OH radicals, it was necessary to check for E1‟s ability to scavenge
•OH.
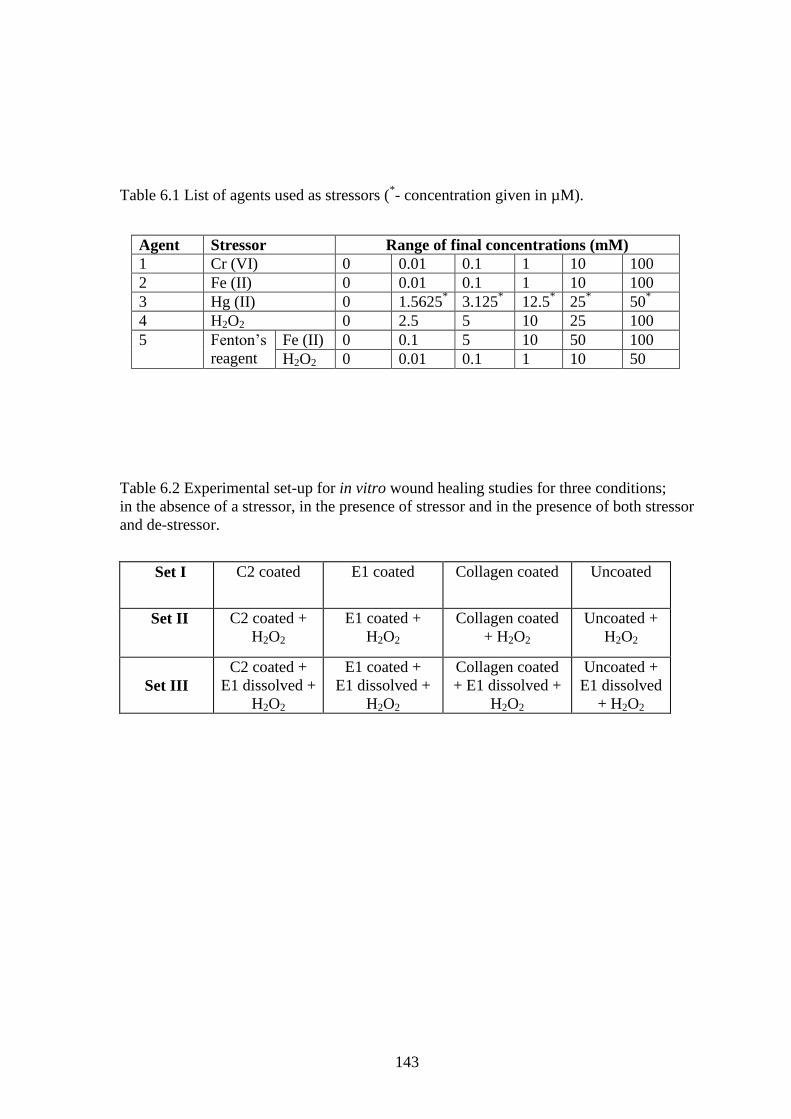
As displayed in Fig. 6.1, BHT, used as positive control, displayed highest activity
followed by E1 and the hydrolysate. The activities of BHT, the peptides and collagen
were significantly different (p<0.004) at a confidence level of 95% based on ANOVA. At
100nmole, E1 displayed a scavenging activity of 58.7%, whereas C2 achieved only 15%.
The activity of the hydrolysate was probably due to the numerous small constituent
charged peptides. However, it was lower than that of E1 (p<0.01) probably due to the
number of active sites being less when compared to that of E1. The results confirmed E1
to be a potent ROS stress releiver in the dissolved form with the ability to quench •OH.
146
0
10
20
30
40
50
60
70
80
90
BHT C2 E1 Hydro Coll
Samples
% s
caven
gin
g a
cti
vit
y 1 nmole
10 nmole
100 nmole
Fig. 6.1 Hydroxyl radical scavenging activity of C2, E1 and the hydrolysate (Hydro).
BHT and collagen (Coll) were used as positive and negative controls, respectively.
6.3.2 DE-STRESSOR ACTIVITIES OF PEPTIDES
Stress is a condition in which the homeostasis maintained in living systems fail,
giving rise to diverse pathological and physiological consequences. The efficacy of the
peptides in relieving oxidative stress generated through H2O2 and heavy metals has been
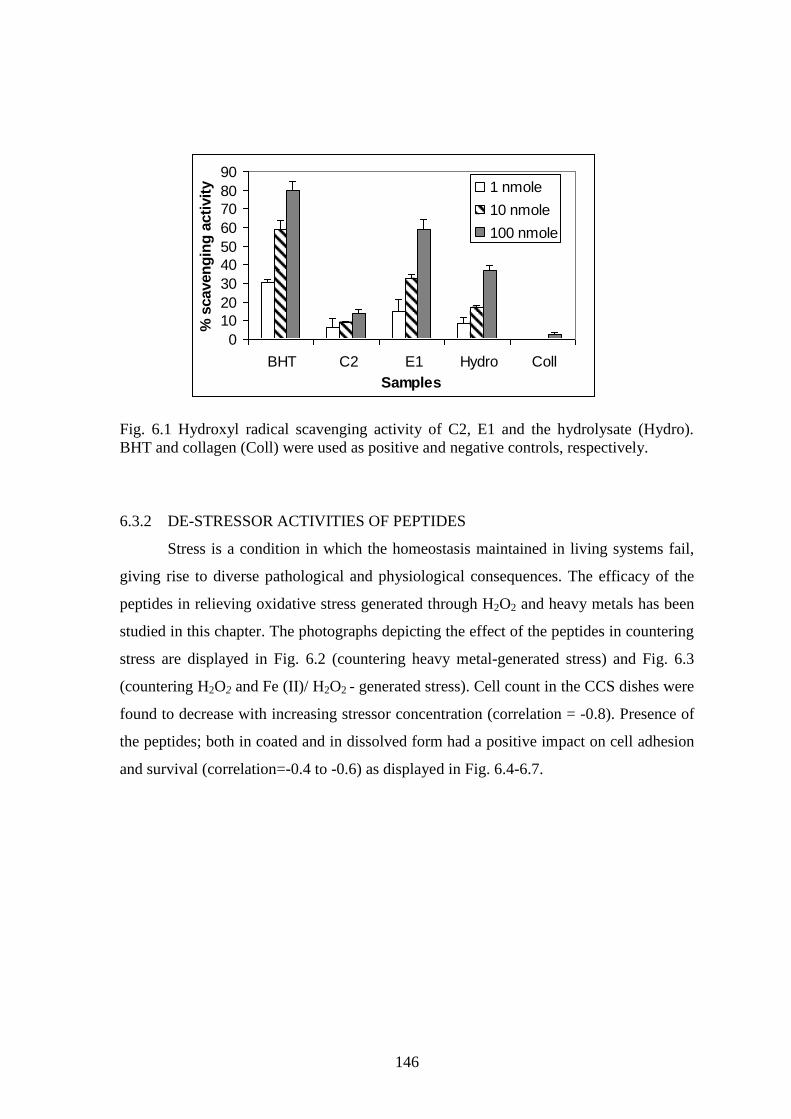
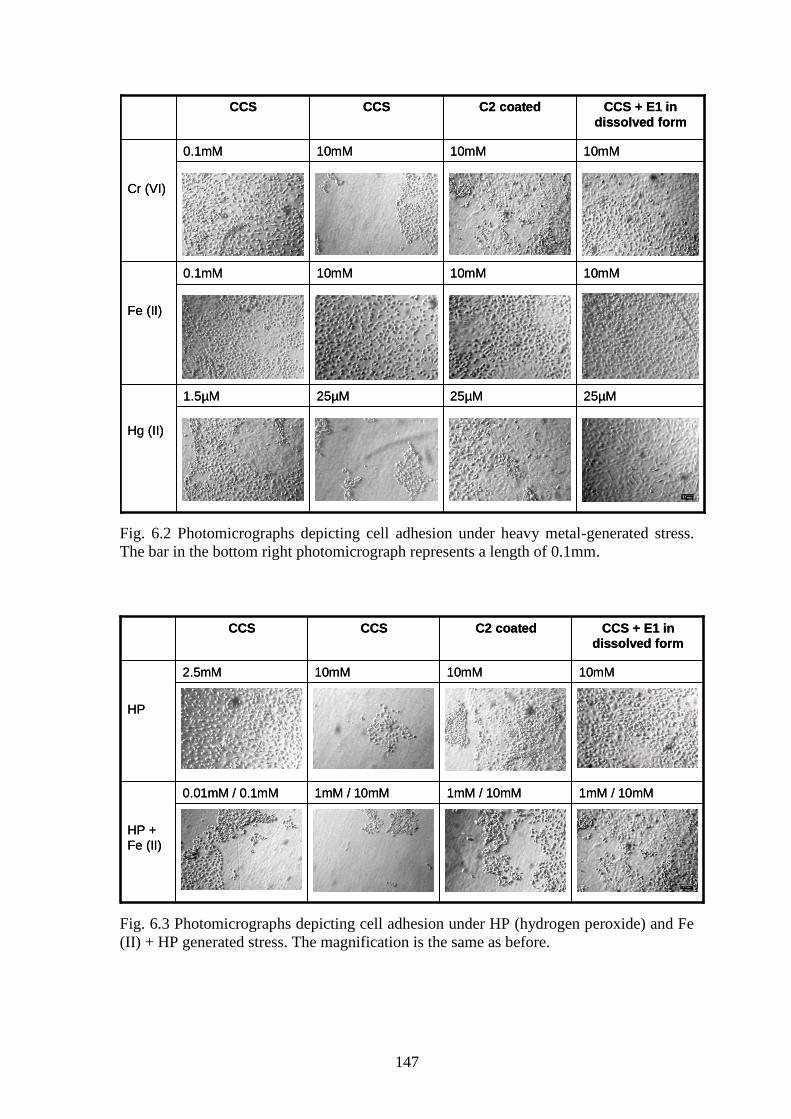
studied in this chapter. The photographs depicting the effect of the peptides in countering
stress are displayed in Fig. 6.2 (countering heavy metal-generated stress) and Fig. 6.3
(countering H2O2 and Fe (II)/ H2O2 - generated stress). Cell count in the CCS dishes were
found to decrease with increasing stressor concentration (correlation = -0.8). Presence of
the peptides; both in coated and in dissolved form had a positive impact on cell adhesion
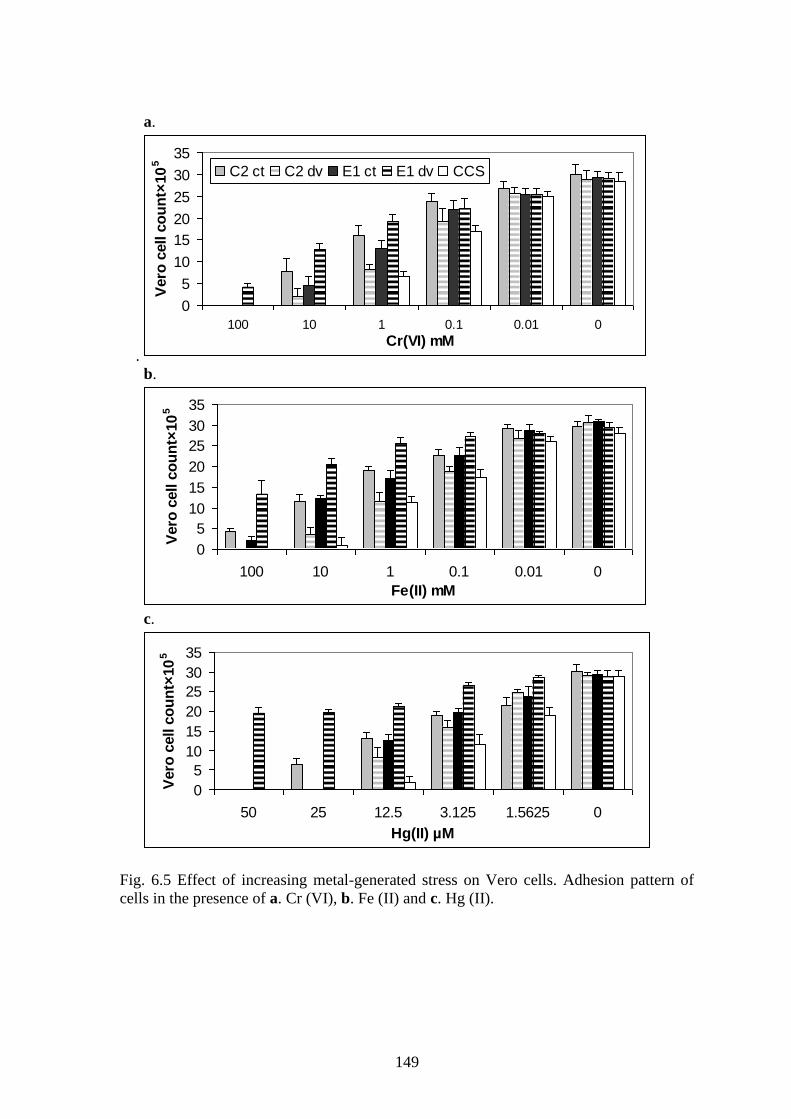
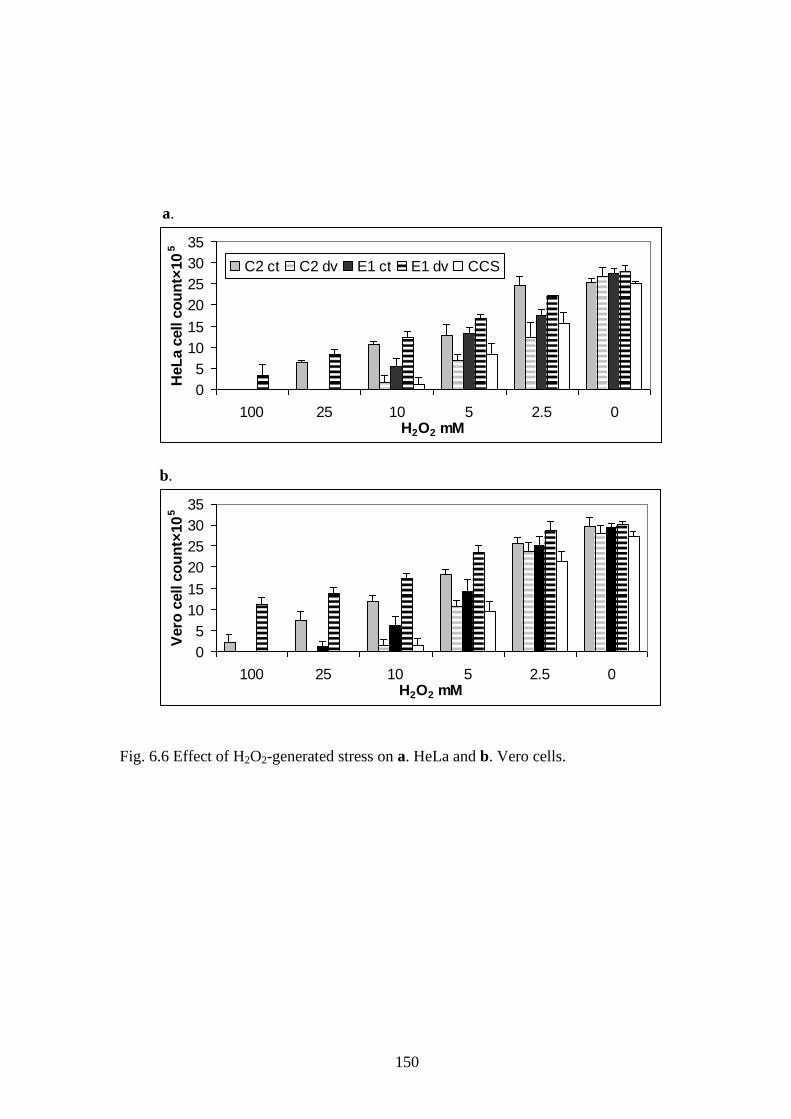
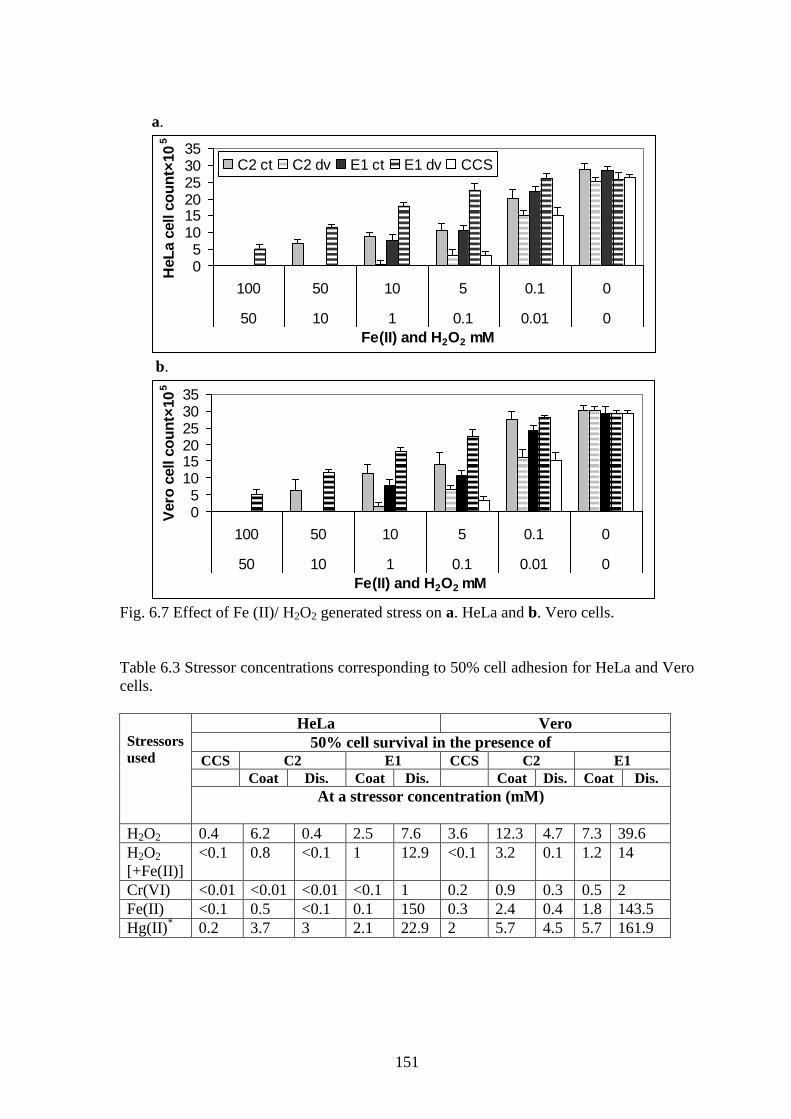
and survival (correlation=-0.4 to -0.6) as displayed in Fig. 6.4-6.7.
147
25µM25µM25µM1.5µM
10mM10mM10mM0.1mM
10mM10mM10mM0.1mM
Hg (II)
Fe (II)
Cr (VI)
CCS + E1 in
dissolved form
C2 coatedCCSCCS
25µM25µM25µM1.5µM
10mM10mM10mM0.1mM
10mM10mM10mM0.1mM
Hg (II)
Fe (II)
Cr (VI)
CCS + E1 in
dissolved form
C2 coatedCCSCCS
Fig. 6.2 Photomicrographs depicting cell adhesion under heavy metal-generated stress.
The bar in the bottom right photomicrograph represents a length of 0.1mm.
1mM / 10mM1mM / 10mM1mM / 10mM0.01mM / 0.1mM
10mM10mM10mM2.5mM
HP +
Fe (II)
HP
CCS + E1 in
dissolved form
C2 coatedCCSCCS
1mM / 10mM1mM / 10mM1mM / 10mM0.01mM / 0.1mM
10mM10mM10mM2.5mM
HP +
Fe (II)
HP
CCS + E1 in
dissolved form
C2 coatedCCSCCS
Fig. 6.3 Photomicrographs depicting cell adhesion under HP (hydrogen peroxide) and Fe
(II) + HP generated stress. The magnification is the same as before.
148
a.
0
5
10
15
20
25
30
35
100 10 1 0.1 0.01 0
Cr(VI) mM
HeL
a c
ell c
ou
nt×
10
5
C2 ct C2 dv E1 ct E1 dv CCS
b.
0
5
10
15
20
25
30
35
100 10 1 0.1 0.01 0Fe (II) mM
HeL
a c
ell c
ou
nt×
10
5
c.
0
5
10
15
20
25
30
35
50 25 12.5 3.125 1.5625 0
Hg(II) µM
HeL
a C
ell c
ou
nt×
10
5
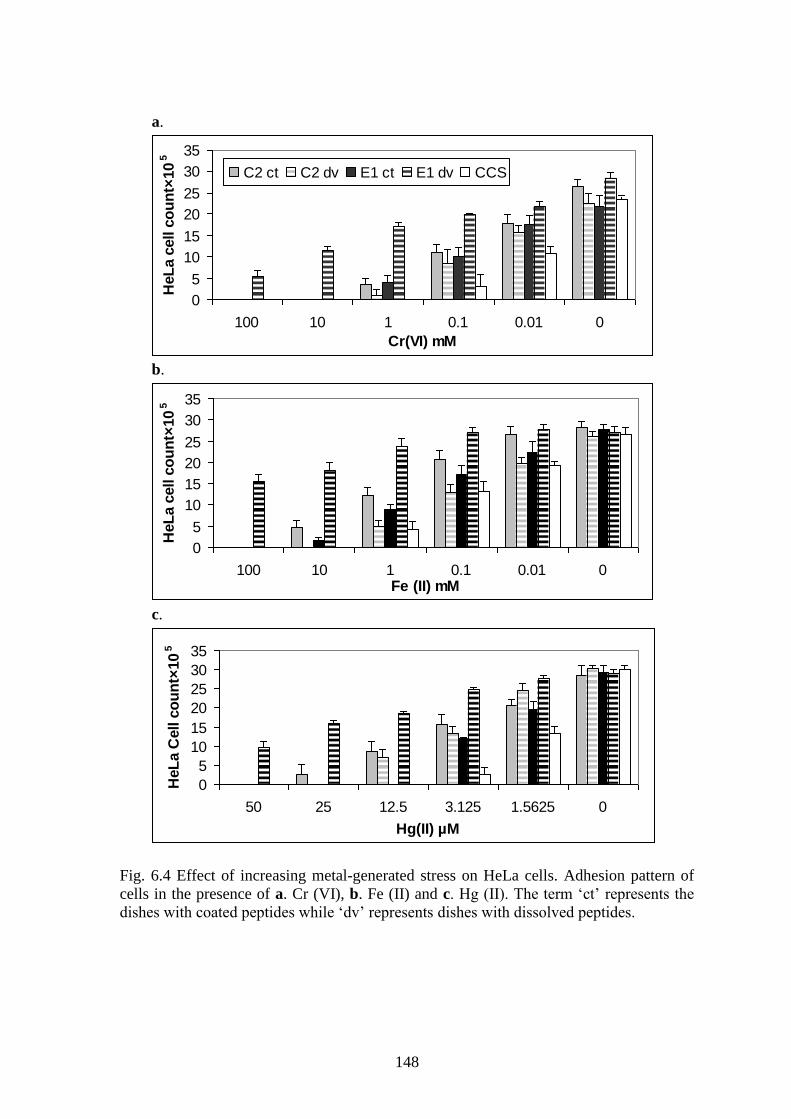
Fig. 6.4 Effect of increasing metal-generated stress on HeLa cells. Adhesion pattern of
cells in the presence of a. Cr (VI), b. Fe (II) and c. Hg (II). The term „ct‟ represents the
dishes with coated peptides while „dv‟ represents dishes with dissolved peptides.
149
a.
.
0
5
10
15
20
25
30
35
100 10 1 0.1 0.01 0
Cr(VI) mM
Vero
cell c
ou
nt×
10
5
C2 ct C2 dv E1 ct E1 dv CCS
b.
0
5
10
15
20
25
30
35
100 10 1 0.1 0.01 0
Fe(II) mM
Vero
cell c
ou
nt×
10
5
c.
0
5
10
15
20
25
30
35
50 25 12.5 3.125 1.5625 0
Hg(II) µM
Vero
cell c
ou
nt×
10
5
Fig. 6.5 Effect of increasing metal-generated stress on Vero cells. Adhesion pattern of
cells in the presence of a. Cr (VI), b. Fe (II) and c. Hg (II).
150
a.
0
5
10
15
20
25
30
35
100 25 10 5 2.5 0H2O2 mM
HeL
a c
ell c
ou
nt×
10
5
C2 ct C2 dv E1 ct E1 dv CCS
b.
0
5
10
15
20
25
30
35
100 25 10 5 2.5 0H2O2 mM
Vero
cell c
ou
nt×
10
5
Fig. 6.6 Effect of H2O2-generated stress on a. HeLa and b. Vero cells.
151
a.
05
101520253035
100 50 10 5 0.1 0
50 10 1 0.1 0.01 0
Fe(II) and H2O2 mM
HeL
a c
ell c
ou
nt×
10
5C2 ct C2 dv E1 ct E1 dv CCS
b.
05
101520253035
100 50 10 5 0.1 0
50 10 1 0.1 0.01 0
Fe(II) and H2O2 mM
Vero
cell c
ou
nt×
10
5
Fig. 6.7 Effect of Fe (II)/ H2O2 generated stress on a. HeLa and b. Vero cells.
Table 6.3 Stressor concentrations corresponding to 50% cell adhesion for HeLa and Vero
cells.
Stressors
used
HeLa Vero
50% cell survival in the presence of
CCS C2 E1 CCS C2 E1
Coat Dis. Coat Dis. Coat Dis. Coat Dis.
At a stressor concentration (mM)
H2O2 0.4 6.2 0.4 2.5 7.6 3.6 12.3 4.7 7.3 39.6
H2O2
[+Fe(II)]
<0.1
0.8 <0.1 1 12.9 <0.1 3.2 0.1 1.2 14
Cr(VI) <0.01 <0.01 <0.01 <0.1 1 0.2 0.9 0.3 0.5 2
Fe(II) <0.1 0.5 <0.1 0.1 150 0.3 2.4 0.4 1.8 143.5
Hg(II)* 0.2 3.7 3 2.1 22.9 2 5.7 4.5 5.7 161.9
152
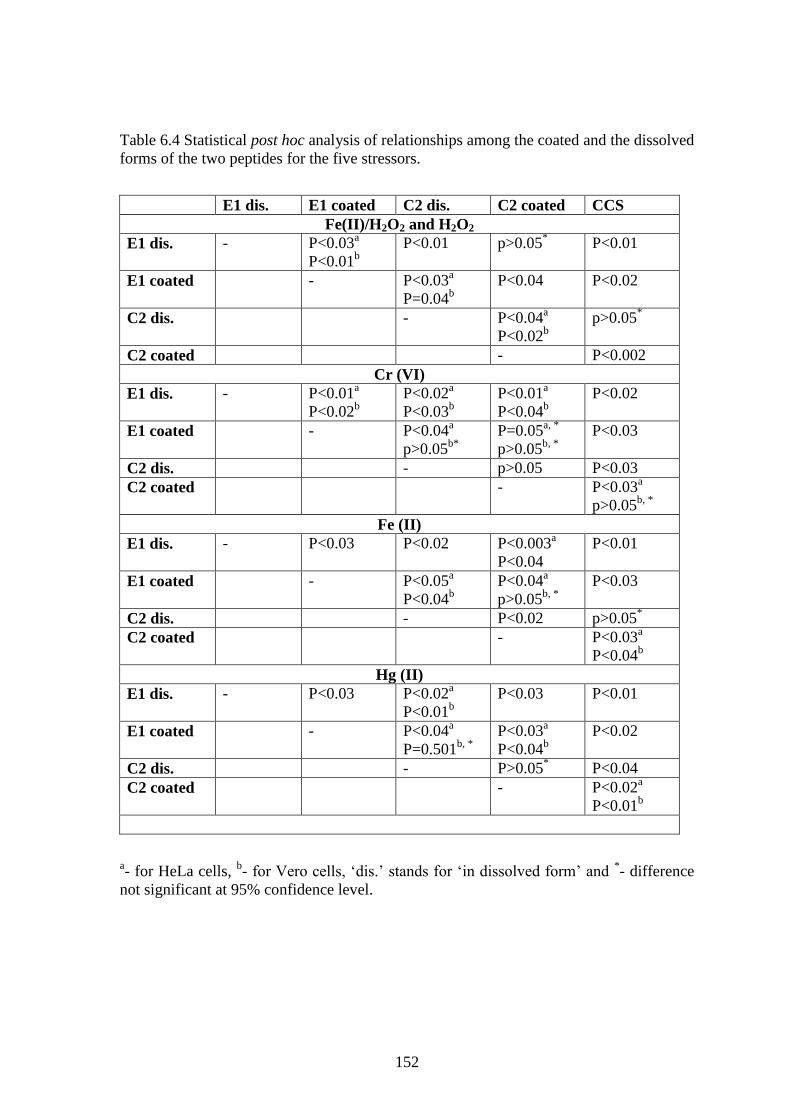
Table 6.4 Statistical post hoc analysis of relationships among the coated and the dissolved
forms of the two peptides for the five stressors.
E1 dis. E1 coated C2 dis. C2 coated CCS
Fe(II)/H2O2 and H2O2
E1 dis. - P<0.03a
P<0.01b
P<0.01 p>0.05
* P<0.01
E1 coated - P<0.03a
P=0.04b
P<0.04 P<0.02
C2 dis. - P<0.04a
P<0.02b
p>0.05*
C2 coated - P<0.002
Cr (VI)
E1 dis. - P<0.01a
P<0.02b
P<0.02a
P<0.03b
P<0.01a
P<0.04b
P<0.02
E1 coated - P<0.04a
p>0.05b*
P=0.05a, *
p>0.05b, *
P<0.03
C2 dis. - p>0.05 P<0.03
C2 coated - P<0.03a
p>0.05b, *
Fe (II)
E1 dis. - P<0.03 P<0.02 P<0.003a
P<0.04
P<0.01
E1 coated - P<0.05a
P<0.04b
P<0.04a
p>0.05b, *
P<0.03
C2 dis. - P<0.02 p>0.05*
C2 coated - P<0.03a
P<0.04b
Hg (II)
E1 dis. - P<0.03
P<0.02a
P<0.01b
P<0.03 P<0.01
E1 coated - P<0.04a
P=0.501b, *
P<0.03a
P<0.04b
P<0.02
C2 dis. - P>0.05* P<0.04
C2 coated - P<0.02a
P<0.01b
a- for HeLa cells,
b- for Vero cells, „dis.‟ stands for „in dissolved form‟ and
*- difference
not significant at 95% confidence level.

153
Table 6.3 lists the stressor concentrations in which 50% of the seeded cells
survived in presence of the peptides. Certain trends were clearly discernable. On an
average for both cell lines, C2-coated displayed cell survival at 10 times more stressor
concentration than CCS for redox-oriented stress and 5 times more for metal-oriented
stress. Similarly, E1-dissolved displayed cell survival at 70 times more stressor
concentration than CCS for redox-oriented stress, and 365 times more for metal-oriented
stress.
Combining the cell survival results and the statistical analysis given in Table 6.4,
some distinct patterns in peptide activity could be inferred, and they are listed below:
(i) The coated form of the peptides displayed a different activity pattern than
their dissolved form and activity depended on the stressor levels.
(ii) E1 in the coated form displayed higher cell survival for all stressor agents
when compared to CCS dishes, hinting that cryptic peptide coated dishes
could provide cytoprotection through adhesion.
(iii) E1 in dissolved form displayed better de-stressing/cytoprotection activity in
comparison to E1 in coated form, implying that E1 is a better antioxidant than
a cell adhesion/cytoprotective agent.
(iv) C2- coated displayed significantly better cell adhesion than E1-coated dishes
for most of the cases, implying C2 to be a comparatively better cell adhesion
agent.
(v) C2-dissolved dishes, for the most part, displayed very little cytoprotective
activity, reflective of the fact that C2 wasn‟t an antioxidative agent as reported
in section 6.3.1.
(vi) For Cr (VI) and Hg (II) generated stress, adherent cell counts were
significantly equal for C2-coated and C2-dissolvced. However, for stress
generated by Fe (II), H2O2 and Fe (II)/H2O2, C2-coated registered more live
cells than C2-dissolved. Cell survival on C2-coated dishes was significantly
better if the stressors generated pure ROS stress, rather than metal-generated
stress.

154
6.3.2.1 STRESS GENERATION BY THE AGENTS
Exogenously administered H2O2 induces cell death through apoptosis and
necrosis in a concentration-dependent manner (Choi et al 2009). H2O2 reacts with Fe2+
to
form Fe3+
, •OH radical, and
–OH ion (Fenton reaction), which can react further to form
the ferryl ion.
Fe2+
+ H2O2 → Fe3+
+ •OH +
–OH
→ Fe=O2+
+ H2O
Either species is a powerful oxidant and can cause massive damage to living tissue. H2O2
by itself can stimulate the upregulation and activity of redox-oriented enzymes such as
NOS and NADPH oxidase. This may lead to unregulated production of superoxide
radicals initiating further H2O2 leading to a vicious cycle (Gough and Cotter, 2011).
Superoxide anions can also have an effect on several signalling pathways; including Map
kinases, angiotensin II signalling and cell proliferation pathways. Additionally,
superoxide anions stimulates the release of intracellular Fe (II) from cellular stores
allowing for uncontrolled Fenton‟s reaction with H2O2 leading to hydroxyl ions.
Cr (VI) is a powerful oxidant and it can gain entry inside cells via sulphate
transport protein. Once inside, the reduction of Cr (VI) by human cytochrome b5 may
lead to the production both Cr (IV) and Cr (V). Either of the species can react with H2O2
to produce HO• radical by a Fenton-like reaction.
Cr4+
/Cr5+
+ H2O2 → Cr5+
/Cr6+
+ •OH+ HO
Cr (VI) can also be reduced to Cr (III), primarily by ascorbate in a manner that also leads
to hydroxyl and superoxide radical production. Cr (III) due to its low tendency to cross
the cell membrane gets trapped inside the cell, enabling it to form stable Cr (III)
complexes with DNA (Borthiry et al 2007; Nickens et al 2010).
Hg ions are known to generate stress through various mechanisms: (i) formation
of covalent bonds with cysteine residues of proteins. (ii) impairing electron transport in
the mitochondria leading to increase H2O2 and superoxide production, (iii) alteration in
Ca levels, leading to increase in enzyme activation and further ROS production and (iv)
increase in lipid peroxidation. Overall, the known increased toxicity of Hg salts when
compared to other metals, required the concentration ranges used for the assay to be
decreased to the µM level

155
6.3.2.2 DE-STRESSOR PATHWAYS
The results of this study confirmed that cell survival was achieved by the peptides
through two different mechanisms. One pathway involved scavenging of free radicals and
the other, through adhesion. Both the pathways are illustrated in Fig. 6.8
Fig. 6.8 Schematic representation of the two mechanisms through which C2 and E1
display cytoprotective effects. The dark arrows represent the pathways leading to cell
death in presence of stress. The red arrows and circles represent the action of the
peptides. E1 in the dissolved form (Ed) scavenges free radicals formed along with
chelation of Fe (II) and Cr (III) and possibly Hg (II). In the coated form, E1 and C2
maintain cell adhesion and possibly activate anti-apoptotic pathways.
6.3.2.2.1 DE-STRESSING THROUGH ROS NEUTRALIZATION
ROS generation by metals occurs through a common reaction;
Metal + prooxidant = Metal (oxidized) + reduced prooxidant + •OH…Eqn. (1)
Antioxidants like BHT and vitamin E generally disrupt this equilibrium by scavenging
the reactive radical formed but leaves chance for more radical production as the metal
responsible for radical production is not affected.
Antioxidant + •OH = Oxidised antioxidant + H2O…………… Eqn. (2)
Stress reduction activity of peptides such as E1 could be two-fold; E1 has been reported
to be a radical scavenger as well as a metal-chelator. Thus it can hamper both sides of the
equilibrium in Eqn. 1, effectively cutting off the metal from the pro-oxidant and by
Cr4+
/Cr5+
+ H2O2 → Cr5+
/Cr6+
+ •OH + OH
Fe2+
+ H2O2 → Fe3+
+ •OH +
–OH
→ Fe=O2+
+ H2O
ascorbate Cr
3+ +
•OH
Complex formation
with DNA
+
other biomolecules
Cr6+
Cytochrome b5
Hg2+ Disruption of ETC in
mitochondria
E1d
E1d
Uncontrolled reactions
leading to further ROS
production.
Loss of FAK
formation
Loss in cell
adhesion
C2, E1
Interaction
with integrin
Apoptosis/ Necrosis
? ?

156
scavenging the product •OH radicals. This form of efficient stress neutralization probably
ensures cell survival even at higher stressor concentrations. An indirect evidence of such
activity is reflected in the fact that Fe-induced toxicity is seen to harm the cells the least,
even though Fe (II) can interact with cellular H2O2 and is capable of inducing significant
damage.
There are two major factors influencing coordination of metal by peptides;
reactive state of the peptide amide group and the side chain residues with coordination
properties. The effect of side chain residues on the stability of peptide complexes have
been studied with a number of residues. The alcoholic-OH from S and T, -COO– from D
and E and NH3+ from Lys have been implicated in coordinate bond formation to metal
ions along with the free N- and C-terminal groups (Davies 2006). E1 displays the
following characteristics: (i) N-terminal G and a C-terminal Q, (ii) % occurrences of
amino acids are G (33.3%), E (8.3%), R, Q, T (5.6%) and K (2.8%). Presence of
hydrophilic residues E, Q, T, R and K were possibly responsible for metal chelation
property.
Most of the metal-peptides complex structures report a closed loop or ringed
structure with the COO– and amide N or the N –terminus satisfying the coordination sites
for the central metal ion (Davies, 2006). Fe (II) chelation is known to occur through the
carboxylate groups of E, which had a % occurrence of 8.3% in E1 (Lv et al 2009). The
peptide E1 could possibly have some role in Cr (III) chelation also. Crystal structures of
octahedral complexes of Cr (III) with peptides and amino acids have revealed a variety of
binding modes. Cr (III) has the ability to bond to carboxylate oxygen, amino nitrogen,
imidazole nitrogen and sulfhydryl donor atoms and is known to form stable complexes
with collagen via –COOH chelation from residues E and D (Covington and Covington
2009). Several studies have reported that peptides rich in E, D, G and C have been found
to be efficient Cr (III) chelators and are useful in Cr removal from portal circulation
(Bertini et al 2001; Dinakarpandian et al 2004). Possibly, the presence of excess amounts
of G along with a moderate amount of E could render peptide E1 with Cr (III)-chelating
properties. Complex formation with Hg (II) requires the presence of C and H residues
(Ngu-Schwemlein et al 2009). The absence of such residues in E1 would possibly restrain
it from direct Hg (II) chelation. Antioxidative agents supplied with chelating agents have
already been reported to be an effective treatment against acute metal toxicity.
Combinatorial therapies like treatment with antioxidants like NAC, lipoic acid, melatonin
and gossypin have shown considerable promise in decreasing clinical pathologies related

157
to ROS and heavy metal-generated stress (Flora and Pachauri 2010). NAC is known to
have free radical scavenging properties and the thiol groups provide chelation sites for
metal ions. Co-administration of several antioxidative/chelating agents along with
vitamins has been proven to display reversal of the stress-induced altered parameters
(Arrigo et al 2005; Marreilha dos Santos et al 2008).
Peptides offer a unique advantage. Presence of a variety of functional residues
allows a bioactive peptide to display high solubility in both aqueous and lipid medium
along with specificity of action. Collagen peptides isolated from fish skin collagen has
been found to offer cyto-protection to a wide array of cell lines including keratinocytes,
fibroblasts and macrophages (Ngo et al 2010; Kato et al 2011).
6.3.2.2.2 DE-STRESSING THROUGH CELL ADHESION
Integrin-ECM interaction has been found to be necessary for cell survival in the
event of stress. The dynamic cytoskeleton is an interconnected continuous network inside
cells capable of transmitting tension from ECM and other cells. A balance between
tensile filaments and resistive focal points and cellular junctions determines the overall
cell shape. An outside interference, like presence of excessive stress can lead to loss of
cell-matrix adhesion and a shift in the cellular balance of forces, resulting in changes in
cell shape and detachment from the surface (Lum and Roebuck, 2001). This leads to cell
division suspension and the activation of pro-apoptotic pathways ultimately leading to
cell death. Loss of cell-matrix attachment also leads to metabolic stress characterized by
reduced nutrient uptake, decreased entry of glucose into citric acid cycle, decreased ATP
production and further increase in ROS (Grassian et al 2011).
C2 in the coated form supported significantly higher cell adhesion and survival,
when compared to its dissolved form, indicating its effectiveness as a de-stressor peptide
working through cell adhesion. Overall, both C2- and E1-coated ensured significantly
higher cell survival when compared to CCS, emphasizing the role of cell adhesion in
countering stress.
It is a well accepted fact that suitable coating materials can influence the adhesion
rate by virtue of a firm cell-to-coated material interaction resulting in faster cell
spreading. Peptides C2 and E1, by virtue of their small size could supply a relatively
greater number of integrin recognition domains per cm2.
This would lead to better cell adhesion when compared to collagen, which would be
adsorbed on the dish surface in a random orientation due to its large size, resulting in

158
comparatively lesser number of integrin recognition sites per cm2. A firm ECM-integrin
attachment can increase integrin clustering, hastening up the focal adhesion sites
formation. Once FAS is formed, a number of cellular signals including anti apoptotic, cell
spreading and proliferation rate could be affected, all eventually leading to higher
survival rates. In the presence of low amounts of stress, the activity between E1 and C2 in
coated form is almost similar (p>0.05) but the difference widens in presence of higher
concentration of stressors, particularly so if the stressors are H2O2 and/or Fe (II) based
(p<0.002).
This study suggests that C2 and E1 can supply a more firm integrin-peptide
interaction resulting in increased cell adhesion, leading to cell survival even in the
presence of stressor agents. The predicament of C2 towards countering specifically redox
stress indicates that E1 and C2 may initiate different signalling patterns. RGD-containing
peptides like E1 interact with αVβ3, αVβ5, αVβ6 and αVβ8 subtypes of integrin receptor
while C2 is thought to interact with the collagen-binding integrin subtypes: α1β1, α2β1,
α10β1 and α11β1.
ROS can be lethal for cells causing apoptosis through caspase activation or
necrosis through poly-(ADP-Ribose) polymerase over-activation. ROS triggers apoptosis
through activation of sphingomyelinase-generating ceramide and alteration in
mitochondrial redox balance, resulting in release of cytochrome c from mitochondria,
Cytochrome c activates Apaf 1, consequently activating caspase 9 and leading to the
formation of apoptosome (Fiers et al 1999; Ha and Synder, 1999). Integrin-ligand
interactions are implicated in cellular resistance to apoptotic stimuli, particularly to
signals that activate the stress pathway or the mitochondrial pathway. Integrins preserve
cell viability in response to stress at several levels. Signalling by integrins regulates both
the expression and activity of several anti-apoptotic proteins. The ligation of integrins
α5β1 or αvβ3, the RGD-binding integrins has been reported to inhibit apoptosis through a
wide variety of stimulations. Two major pathways include the PI3-K/ PKB and the Raf/
MEK/ ERK signalling cascade. The downstream signalling leads to phosphorylation of
pro-apoptotic proteins followed by sequestering them to chaperones and activation of the
Bcl-2 family of anti-apoptotic proteins, resulting in inhibition of pore formation in the
mitochondria. Ligand-integrin-FAK formation leads to activation of Hsp 2, another anti-
apoptotic protein which inactivates caspases and prevents cytochrome c release. Last but
not the least, it results in the up-regulation of IAP (inhibitors of apoptotic pathway) class
of proteins, which performs a plethora of functions, including inactivating executioner

159
caspases and regulating cell division (Stupack and Cheresh, 2002; Gough and Cotter,
2011).
6.3.3 WOUND CLOSURE MONOLAYER ASSAY
The „scratch assay‟ is a direct and inexpensive process to study wound healing in
vitro. The assay is based on the fact that creation of an artificial „scratch‟ or a gap on a
fully confluent cell monolayer will cause the cells at the edges of the gap to move
towards each other for closing the gap and re-establishing contact. The assay was
conducted with Vero cell lines because of its faster doubling time in comparison to HeLa.
The photographs of wound closure under various circumstances are depicted in Fig. 6.9.
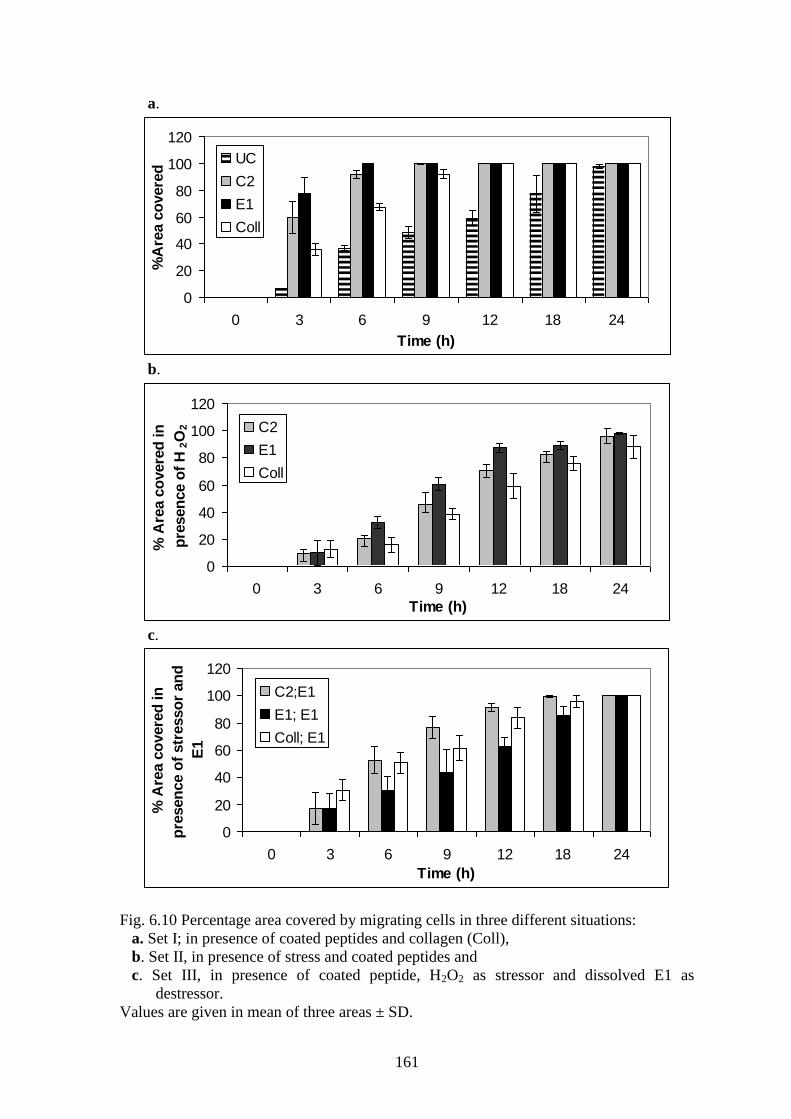
Cell migration was measured by calculation of %area covered by the inward moving cells
in a certain time period. In the absence of stress (Set I; Fig. 6.10a), the migration of the
cells was significantly faster on peptide coated dishes when compared to collagen
(p<0.02) and in general, gap closure on coated dishes were faster than uncoated dishes
(p<0.005).
Cells growing on E1 covered 77% of the area within 3h of the wound creation and
100% within 6h. C2 displayed 59% coverage at 3h and 99.6% in 9h. Cells growing on
collagen closed the wound completely within 12h whereas cells in uncoated dishes took
24h. In the 3rd
hour, area coverage in C2 and E1 was, on an average, 7 and 2 times more
than for uncoated and collagen coated dishes, respectively. In the presence of oxidative
stress (Set II; Fig. 6.10b), the uncoated dishes were unable to promote any cell adhesion.
Collagen coated dishes exhibited live, migrating cells but wound closure was partially
complete even after 24h. On the other hand, peptide coated dishes displayed live
migrating cells and even though a delay was observed, the gap was closed within 24h.
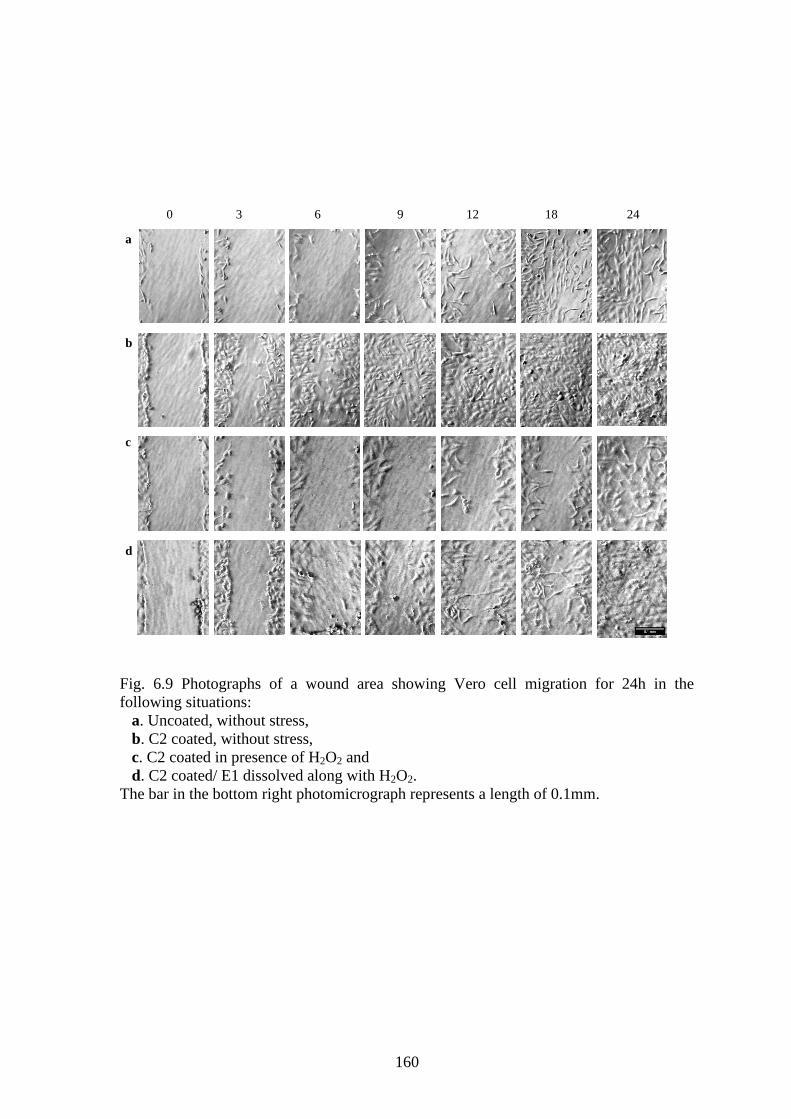
160
Fig. 6.9 Photographs of a wound area showing Vero cell migration for 24h in the
following situations:
a. Uncoated, without stress,
b. C2 coated, without stress,
c. C2 coated in presence of H2O2 and
d. C2 coated/ E1 dissolved along with H2O2.
The bar in the bottom right photomicrograph represents a length of 0.1mm.
0 3 6 9 12 18 24
a
b
c
d
161
a.
0
20
40
60
80
100
120
0 3 6 9 12 18 24
Time (h)
%A
rea c
overe
d UC
C2
E1
Coll
b.
0
20
40
60
80
100
120
0 3 6 9 12 18 24
Time (h)
% A
rea c
overe
d in
pre
sen
ce o
f H
2O
2 C2
E1
Coll
c.
0
20
40
60
80
100
120
0 3 6 9 12 18 24
Time (h)
% A
rea c
overe
d in
pre
sen
ce o
f str
esso
r an
d
E1
C2;E1
E1; E1
Coll; E1
Fig. 6.10 Percentage area covered by migrating cells in three different situations:
a. Set I; in presence of coated peptides and collagen (Coll),
b. Set II, in presence of stress and coated peptides and
c. Set III, in presence of coated peptide, H2O2 as stressor and dissolved E1 as
destressor.
Values are given in mean of three areas ± SD.

162
Overall, the oxidative stress had a significant impact on wound closure; both E1-
coated and C2-coated dishes responded in a similar manner (p=0.59) but still displayed
relatively faster cell migration and quicker area coverage than collagen-coated dishes
(p<0.01).
Wound closure is a multi-component synergistic effect. To mimic the various
synergistic effects of the peptides occurring in the in vitro model of wound healing, E1 in
the dissolved form was added as an additional de-stressor (Set III; Fig. 6.10c). This had a
significant effect on cell survival and movement; the gap closure rate for C2- and
collagen-coated dishes was found to increase (p<0.04) in comparison to Set II.
Interestingly, gap closure in C2-coated/ E1-dissolved dishes was found to be
significantly faster (p<0.05) than collagen-coated/ E1-dissolved dishes. This could be
attributed to the synergistic effects between the activity of E1 in dissolved form and C2 in
coated form. E1 in the dissolved form acted as an efficient ROS de-stressor, removing
free radicals whereas C2 in the coated form acted through integrin receptors to maintain
adhesion. Consequently, the combination of these two peptides resulted in faster wound
closure. However, gap closure rate in E1-coated/ E1-dissolved dishes was found to be
significantly lower (p<0.02) than collagen/E1 and C2/E1 combinations, probably arising
from an „excess mass‟ effect and the recoiling tendency of collagen peptides. Cryptic
peptides generally display a hyperbolic increase in bioactivity along with concentration
and excess amounts actually decrease activity. Possibly in the E1/E1 situation, the
dissolved E1 in the media was being deposited on the exposed wound surface, thus
increasing the amount of coated peptide and reducing activity connect to chapters. A
decrease of E1 in the medium could also lead to lower ROS scavenging, again leading to
reduced gap closure.
To summarize, the two conditions with which maximum wound closure could be
observed under oxidative stress, were:
(i) C2 coated/ E1 dissolved and
(ii) collagen coated/ E1 dissolved.

163
6.3.4 GAP CLOSURE BY „CONTACT-GUIDANCE‟ IN PRESENCE OF PEPTIDES
Collective cell movement is responsible for wound closure. Gap closure takes
place through the initiation of few „leader‟ cells that are closely followed by a group of
„followers‟. The „leader‟ is a cell at the edge of the wound that extends a lamellipodium
in order to meet a similar cell from the other side of the edge. The neighbouring cells that
accompany them lag slightly behind but they form the mass of the connecting bridge.
This process of gap closure, termed as „contact guidance‟ is possible only when cells can
interact with a coated material (Haga et al 2005).
As displayed in Fig 6.9b, c and e, cells from both sides start extending into the wound
area during the 3rd
hour while cells on the uncoated surface displayed a lack of such cell
movement (Fig. 6.9a).
Leader cells exhibit enhanced tension along the length of the advancing
outgrowth. This produces a forward tension leading to reorientation of stress fibers and
through cell–cell contacts, the leader cell pulls the followers into the wound area
followed by subsequent active movement of neighbouring cells (Omelchenko et al 2003).
The stress fiber arrangements in the leader cell are probably initiated through interaction
of integrin and the coated ligand. Peptides like RGD and C2, in this study, in the coated
form are capable of integrin clustering, consequently leading to the arrangement of FAK
and stress fiber formation (Kawakami et al 2001). This is schematically represented in
Fig. 6.11.
For set II, leader cells could be observed in the 6th
hour (Fig. 6.9c) reaching out their
lamellipedia, but collective cell movement was absent. However, it was notable that the
cells survived even in the presence of stress and this could probably be attributed to the
presence of the coated peptide. The expected loss in filament tension leading to shape
loss and detachment of the cells was also not observed. Instead, the cells maintained the
usual shape, indicative of the retained arrangement of the actin fibers due to interaction
with the coated peptides.
In the 12th
hour, the cells were found scattered and unable to follow the earlier „leader-
follower‟ nomenclature, probably due to the stress. However, cells were eventually able
to cover the wound after 24 h. The situation is reverted back for cells in the presence of
dissolved E1 (Set III). The faster gap closure observed in Set III could probably be due to
the synergistic effect of the coated C2 leading to firm adhesion/leader cell formation and
ROS scavenging activity of E1.
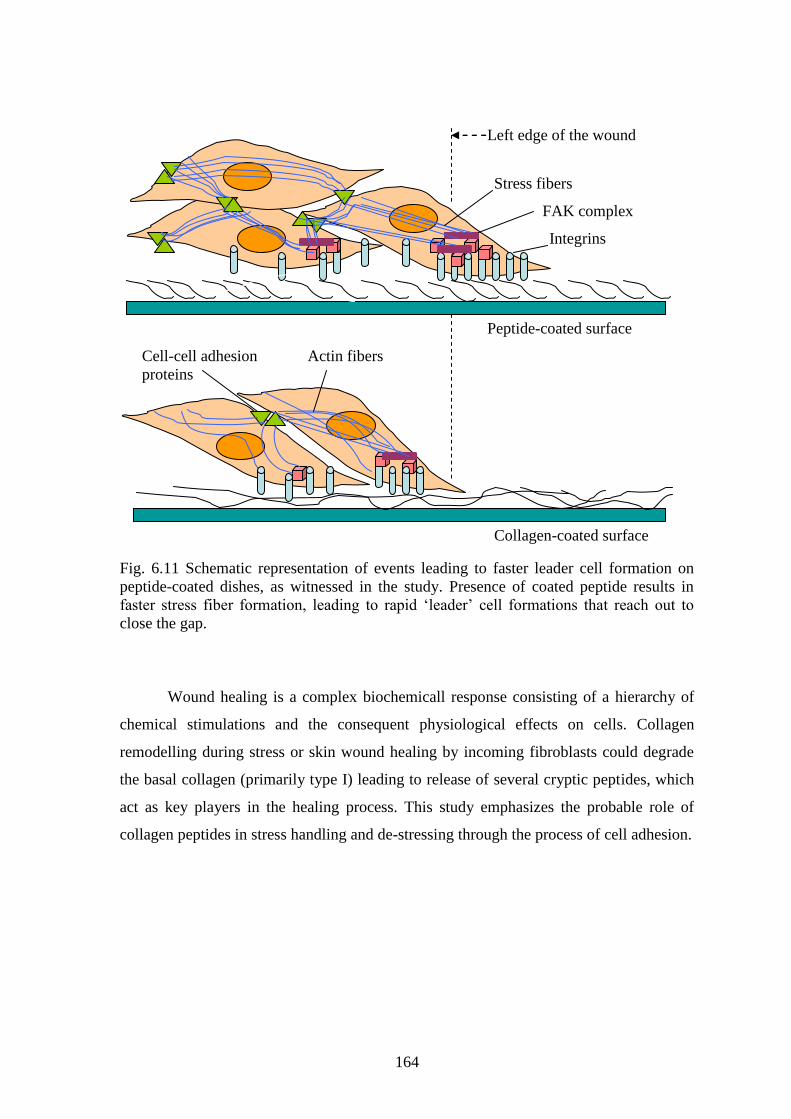
164
Fig. 6.11 Schematic representation of events leading to faster leader cell formation on
peptide-coated dishes, as witnessed in the study. Presence of coated peptide results in
faster stress fiber formation, leading to rapid „leader‟ cell formations that reach out to
close the gap.
Wound healing is a complex biochemicall response consisting of a hierarchy of
chemical stimulations and the consequent physiological effects on cells. Collagen
remodelling during stress or skin wound healing by incoming fibroblasts could degrade
the basal collagen (primarily type I) leading to release of several cryptic peptides, which
act as key players in the healing process. This study emphasizes the probable role of
collagen peptides in stress handling and de-stressing through the process of cell adhesion.
Left edge of the wound
FAK complex
Integrins
Cell-cell adhesion
proteins
Stress fibers
Actin fibers
Peptide-coated surface
Collagen-coated surface














