Embed Size (px)
Citation preview

CHAPTER 6: Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
68
CHAPTER : 6
Studies on Plant Growth Promoting
Activity of Cyanobacterial Isolates

CHAPTER 6: Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
69
6.1 INTRODUCTION
The importance of agriculture in the socio-economic fabric of India can be
realized from the fact that the livelihood of majority of the country population
depends on agriculture. In India agriculture is facing many constraints, one is,
need of environmental friendly alternative of chemical fertilizers and second is
gradually depletion of nutrient supply in Indian agriculture soil. The use of
Cyanobacteria as PGPR can fulfill these criteria and ensure viability of farming
activity.
While working on the cyanobacteria of Indian paddy fields, Gupta and Lata
(1964) observed that cyanobacteria accelerated seed germination and promoted
seedling growth. In addition, they also observed that both yield and the quality of
the grains was improved because they were richer in proteins. It seems very likely
that the beneficial effect of the algae on the rice crop may not be restricted to their
capacity to fix atmospheric nitrogen alone, especially in fields where nitrogen-
fixing algae may not be present in appreciable quantities. It was assumed that
active substances are gibberellin (GA) type. A gibberellin-like substance has been
isolated from the cyanobacterium Phormidium foveolarum and this is active in
GA-bioassays (Gupta et al 1973). Growth-promoting substances were detected
by the effect of extracts of N. muscorum on seedlings of Panicum miliaceum. The
height of millet plants as well as their dry weight were also increased by all the
extracts (Caire et al., 1976). Others authors have reported that vitamins,
aminoacids and polypeptides benefit plant growth. Information about
cyanobacterial biomass or their substances being incorporated to other plants
different to rice is scarce (Halperin et al., 1981).
With the increasing prices of fertilizers and associated problems of global
warming and environmental pollution, there is a need for identifying organic
options for increasing food quality and yields. Soil fertility is diminishing
gradually due to soil erosions, accumulation of salts and other toxic elements,
water logging and un-balanced nutrient compensation. Organic wastes and bio-
fertilizers are the alternate sources to meet the nutrient requirement of crops and

CHAPTER 6: Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
70
to bridge those gaps. Farming regions that emphasized heavy chemical
application led to adverse environmental, agricultural and health consequences.
Many efforts are being exercised to combat the adverse consequences of chemical
farming.
Cyanobacterial biofertilizers have been reported to be very useful in ameliorating
various physico-chemical properties of marginal soils and the EPS produced by
the cyabobacteria seems to play an important role (Nisha et al., 2007). Since
salinity is a major constraint to crop growth and germination is the most
syensitive and decisive stage for successful crop establishment (Soltani et al.,
2006), it would be worthwhile to study whether EPS application is useful in
promoting seed germination in the presence of salts in the medium.
In some filamentous cyanobacteria nitrogen fixing heterocysts are formed.
Heterocysts are terminally differentiated cells whose interior becomes anaerobic,
mainly as a consequence of respiration, allowing the oxygen- sensitive process of
nitrogen fixation to continue. The regulation of dinitrogen fixation has been
extensively studied in the heterocyst system (Böhme, 1998).
The distribution of cyanobacteria in the soil depends on the soil pH, electrical
conductivity, and exchangeable sodium. Amongst these, pH is the most important
factor determining the Cyanobacterial diversity. Under laboratory conditions, the
optimal pH for cyanobacterial growth ranges from 7.5–10, with a lower limit of
6.5–7.0. Although infrequent at pHs below 6.0, cyanobacteria’s ability to grow at
diverse pH and modify their own environment.
A number of studies have demonstrated the growth promotion activity of
cyanobacterial extracts on plant regeneration and plantlet formation.
Cyanobacteria have potential to produce many metabolites which includes the
phytohormones (IAAs, cytokinin and gibberillin-like compounds) and
ironchelators (schizokinen, anachelin and synechobactins) and exhibit profound
effect on the productivity of the ecosystem. (Yadav, et. al., 2011). As plant
growth promoting agents, advantages of cyanobacteria over expensive synthetic
phytohormones include broader spectrum of activity and optimum levels of

CHAPTER 6: Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
71
biologically active molecules, which are needed for normal plant development in-
vivo or in-vitro (Sergeeva et. al. 2002, and Prasanna et. al. 2010).
Microbial inoculation is known to play a major role in improving soil fertility and
plant growth/yields; however, most of the published literature on this aspect has
focused on cereals or legumes or important cash crops such as cotton or
fruits/vegetables (Bashan,1998; Balser et al., 2001; Roger et al., 1993;
Prasanna et al., 2013,; Stefan et al., 2013).
Statistical analysis confirm that there is a significant difference in plant height,
root length, number of leaf, fresh and dry weight of root, leaf and stem in treated
plants as compared to control. Venkataraman & Neelakantan (1967) showed
that the production of growth substances and vitamins by the algae may be partly
responsible for the greater plant growth and yield.
The capacity for biosynthesis of growth promoting substances such as auxins,
amino acids, sugars and vitamins (Vitamin B12, Folic acid, Nicotinic acid and
Pantothenic acid) also can enhance plant growth. The other reason that can
suggest for increased plant growth by using cyanobacterial extract is that, the
growth of BGA in soil seems to influence the physical and chemical properties of
soil.
The review of literatures showed that, there are only a few studies on similar
subjects, especially on unicellular Cyanobacterial culture as biofertilizer crops;
however results of heterocystous cyanobacteria as biofertilizer had been
investigated. The results obtained in this work showed that pre-soaking seeds by
Cyanobacterial culture accelerates seed germination and seedling height (Fig. 8.3
& 8.4). Previously, Nanda et al. (1991) showed that, pre-soaking of pumpkin and
cucumber seeds in Westiellopsis prolific extract can accelerate seed germination
and spraying extracts of this cyanobacterium to emerged seedling during their
subsequent cultivation led to significant increase in growth and development of
both crops.

CHAPTER 6: Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
72
A. Cyanobacteria and Mung bean (Vigna radiata or Phaseolus aureus)
Mung beans are cheap, reliable and easy to germinate, and offer a useful way to
look at the germination process. Mung beans are legumes (members of the
Fabaceae family), and are most commonly used in the India for growing
nutritious bean sprouts. Mung beans are annuals, growing up to about 1m in
height. The first flowers appear seven to eight weeks after planting and the crop
reaches maturity in 12 to 14 weeks. The mung bean plant comes originally from
India, but is now widespread throughout the tropics. Mung Beans can germinate
within 48-72 hrs.
Agronomic potential of biofilmed preparations of selected cyanobacterial strains
has been investigated. The formulations were prepared using paddy straw
compost:vermiculite (1:1) as carrier and tested as inoculants in mungbean and
soybean. The effects of the formulations were evaluated in terms of
microbiological, nutrient availability, and plant biometric parameters. The
Trichoderma viride–Bradyrhizobium biofilm exhibited 20–45% enhancement in
fresh/dry weight of plants over other microbial treatments, while the T. viride–
Azotobacter biofilm exhibited highest dehydrogenase activity in the soil and
nitrogen fixation. This study highlights the promise of cyanobacterial inoculants
and biofilmed biofertilizers as promising inputs for integrated nutrient
management strategies in agriculture. (Prasanna, R et al. 2014)
Soil and Fertilization
Mung bean performs best in fertile, well-drained sandy loam soil with a pH
between 6.2 and 7.2 and will suffer in poorly-drained, heavy soils. Plants in
alkaline soils will display symptoms of nutrient deficiencies. Nitrogen fertilizer is
unnecessary, though it may encourage early growth and faster establishment.
Mung bean has the same nutrient requirements as other legumes. A soil test is the
best way to determine phosphorus and potassium requirements. In fields or
gardens where mung beans are planted for the first time, a nitrogen-fixing
Rhizobium bacteria specifically for mung beans should be applied to the seeds or
planting area.

CHAPTER 6: Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
73
Irrigation
The deep-rooted mung beans are adapted to warm, dry climates and grow best
with only three to five deep waterings during the growing season. If the soil is
adequately moist at the time of planting, the first watering is not needed until
about 20 days after planting. Irrigation timing and frequency needs vary
depending on humidity, rainfall and winds. A regimen with only a few, deep
waterings limits vegetative growth and encourages seed production. The mung
bean requires adequate water between blooming and pod fill.
Planting and Spacing
Mung bean planting should occur early enough that harvest will occur before the
rainy season and bloom or pod fill will occur before the hottest, driest part of
summer and late enough that all danger of frost has passed and soil temperatures
are above about 60 degrees Fahrenheit. Two plantings annually, one in spring and
another in fall, are often possible in warmer regions. Tilling or cultivating the top
several inches of soil breaks up the ground and controls weeds. Mung bean seeds
germinate best when planted 1 to 2 inches deep in moist soil. One seed every 3
inches in rows 18 to 24 inches apart provides an adequate yield.
Process of germination occurs in different stages. Some steps of seed germination are as follows: • Seed absorbs water and seed coat gets burst. It is the first sign of germination.
There is an activation of enzymes, increase in respiration and plant cells get
duplicated. A chain of chemical changes starts which leads to development of
plant embryo.
• Chemical energy stored in the form of starch is converted to sugar, which is
used during germination process. Soon, embryo gets enlarged and seed coat
burst opens.
• Growing plant emerges out. Tip of root first emerges and helps to anchor the
seed in place. It also allows embryo to absorb minerals and water from soil.
• Some seeds require special treatment of temperature, light or moisture to start
germination.

CHAPTER 6: Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
74
B. Cyanobacteria and Wheat
Wheat is an agriculturally important crop and requires high doses of fertilizers.
However, little work has been done on the supplementary input of cyanobacteria
asbiofertilizers to this crop, and available information is scarce. A study of the
application of vermicompost, farmyard manure, and biofertilizers (cyanobacteria
and Azotobacter) in different combinations with chemical fertilizers (N40P30K30)
had promising results for wheat crops (var. HD 2687). The application of
vermicompost in combination with cyanobacteria brings about significant
increases in nitrogenase activity, while Azotobacter + cyanobacteria (+N40P30K30)
treatment gives the highest values of chlorophyll. The addition of vermicompost
and farmyard manure (+N40P30K30) enhances cyanobacterial abundance and
diversity. Nostoc,Anabaena, Calothrix, Oscillatoria, and Phormidium are the
dominant genera in wheat-crop soil (Prasanna et al., 2008b).
Cyanobacteria are known to liberate substantial quantities of extracellular
nitrogenous compounds into the medium. Physiological attributes of a set of
cyanobacterial strains (Calothrix ghosei, Westiellopsis, Hapalosiphon intricatus,
and Nostoc sp.) isolated from the rhizosphere of wheat (var. HD 2687) were
analyzed by Karthikeyan et al. (2009). The concentrated culture filtrates of these
strains enhance wheat-seed germination percentages, and the radicle and
coleoptiles lengths. Thin layer chromatography analyses of the filtrates revealed
the presence of several amino acids such as histidine, and auxin-like compounds.
Co-culturing experiments with selected Cyanobacterial strains recorded
significant enhancements in plant chlorophyll.
6.2 Seed Vigor Index
Seed vigor is the sum total of those properties of seed which determine the level
of activity and performance of the seed or seed lot during germination and
seedling emergence. Although differences in physiological attributes of seed lots
can be demonstrated in the laboratory, it was suggested that the term should be
used to illustrate the performance of seed were shown in the field. The seed lot
showing the higher seed vigor index is considered to be more vigorous Seedling

CHAPTER 6: Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
75
vigor index was calculated following modified method of Abdul-Baki &
Anderson.
Vigor Index = Seedling length (mm) x Germination %
Importance of Seed Vigor Testing
Vigor testing does not only measure the percentage of viable seed in a sample, it
also reflects the ability of those seeds to produce normal seedlings under less than
optimum or adverse growing conditions similar to those which may occur in the
field. Seeds may be classified as viable in a germination test which provides
optimum temperature, moisture and light conditions to the growing seedlings;
however, they may not be capable of continuing growth and completing their life
cycle under a wide range of field conditions. Generally, seeds start to lose vigor
before they lose their ability to germinate; therefore vigor testing is an important
practice in seed production programs.
Testing for vigor becomes more important for carryover seeds, especially if seeds
were stored under unknown conditions or under unfavorable storage conditions.
Seed vigor testing is also used as indicator of the storage potential of a seed lot
and in ranking various seed lots with different qualities.
The biological basis of the seed vigor concept
It has been established that the conditions of seed development, maturation,
storage and aging influence seed vigor. Seeds developed under moisture stress,
nutrient deficiency, extreme temperatures, etc. often result in light, shriveled seed
or collectively called poor-vigor seed. Preharvest environment of high humidity
and warm temperatures can also cause loss in seed viability and vigor. Seed
mechanical damage, whether induced by harvesting or conditioning equipment, as
well as improper storage conditions are among the factors that adversely affect
seed vigor. In addition, genetic factors such as hard-seededness, resistance to
diseases, and seed chemical composition influence the expression of seed vigor.

CHAPTER 6: Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
76
Methods of measuring seed vigor
The general strategy of determining seed vigor is to measure some aspects of seed
deterioration or weaknesses, which is inversely proportional to seed
vigor.
Cold test, accelerated aging test, electric conductivity test, seedling vigor
classification, and seedling growth rate are among the tests that are used to
measure seed vigor. In addition, the tetrazolium (TZ test) can be used as a vigor
test by classifying the pattern of stained seeds into high, medium and low quality.
The AOSA Seed Vigor Testing Handbook is a good source of information on
seed vigor testing. Below is a brief description for some of the most common seed
vigor tests that are used for various crops including corn, soybean, field beans,
peas, grasses, vegetable seeds, and other crops.
a) Electric Conductivity Test
This test measures the integrity of cell membranes, which is correlated with seed
vigor. It is well established that this test is useful for garden beans and peas. It has
been also reported that the conductivity test results are significantly correlated
with field emergence for corn, and soybean. As seeds lose vigor, nutrients exude
from their membranes, and so low quality seeds leak electrolytes such as amino
acids, organic acids while high quality seeds contains their nutrients within well
structured membranes. Therefore, seeds with higher conductivity measurement
are indication of low quality seeds as vice versa.

CHAPTER 6: Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
77
6.2 MATERIAL & METHODS
6.2.1 Seed Selection
Seeds were selected based on seed vigour index. The electrical conductivity test is
used to measure veed vigour index. Two cultivas were selected for the
conductivity test.
6.2.2 Germination Test.
Seeds of Vigna radiata and Triticum aestivum were surface sterilized with 5%
sodium hypochlorite for 8 min, then rinsed with distilled water several times to
remove any trace of sodium hypochlorite. The seeds were classified into two
groups. Then seeds were incubated with cyanobacterial isolates under shaking
condition for 2 hours. Water treated seeds were used as control.
Control and bacterized seeds (10 seeds per pot) were sown in sterilized pot
containing autoclaved soil (approximately 100 g). All pots were labeled properly.
The pots were kept at 25 ± 2°C, 60% relative humidity, under 8 hours/16 hours
dark/light photoperiod. Pots were observed and watered regularly during this
period. After 10 days, plants were harvested and their growth parameters were
analyzed.
At the end of the experiment total fresh weight, length of shoots and roots per
plant were determined.
6.2.3 sterile soil
The soil was sterilized twice by autoclaving, through intermittent sterilization for
2 h at 121.4 °C and 15 lbs pressure, on two consecutive days (to destroy spores
and avoid or minimize contamination). The physico-chemical properties of soil
should be done. Plastic pots of 6 inch size were employed in the study.
6.2.4 Plant Inoculation Experiments
Seeds of Vigna radiata and Triticum aestivum were surface sterilized with 5%
sodium hypochlorite for 8 min, then rinsed with distilled water several times to
remove any trace of sodium hypochlorite. The seeds were classified into two

CHAPTER 6: Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
78
groups. Then seeds were incubated with cyanobacterial isolates under shaking
condition for 2 hours. Water treated seeds were used as control.
Control and bacterized seeds (8 seeds per pot) were sown in sterilized pot
containing autoclaved soil (approximately 100 g). All pots were labeled properly.
The pots were kept at 25 ± 2°C, 60% relative humidity, under 8 hours/16 hours
dark/light photoperiod. Pots were observed and watered regularly during this
period. After 10 days, plants were harvested and their growth parameters were
analyzed.At the end of the experiment total fresh weight, length of shoots and
roots per plant were determined.
6.2.5 Growth Analysis
A. Growth Analysis: Above ground parts
Plant height:
Measurements of plant height were taken at the crop maturity stage in
three replicates of five plants each. Plant height was measured from the soil line
to shoot tip. Plant height was measured by placing the plant on a centimeter scale.
Plant population was uniform at the time of maturity of crop.
Dry matter production
At maturity fifteen plants (five plants from each replicate) were randomly
selected and the data on component part dry weights (leaf, root and shoot) were
recorded. Leaf, stem and root were dried at 60 ºC for 72 hours. Pods were
separated and the total numbers of branches were recorded. Dry weights of seed
components were recorded after drying at 35–40 ºC for 10 days.
Growth Analysis: Below ground parts
Root length:
The measurements of root length were done in plants at 10 days. Roots
were taken out carefully, washed, and measured against a cm scale.
Root fresh and dry weight
Plant roots with nodules at each sampling were washed and dried on filter
paper and weighed for the fresh weight. For dry weight roots were dried at 60oC
for 72 hours and weighed.

CHAPTER 6: Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
79
6.3 RESULT AND DISCUSSION
Seed cultivar Mung-2 and wheat-1 is better quality of seeds as they have
comparatively less conductivity value.
Type of seed Electrical conductivity mean Mung-1 0.5167 Mung-2 0.3467
Wheat-1 0.16 Wheat-2 0.23
Table 6.1 Result of electrical conductivity test
Cyanobacteria play a key role in improving growth of many plants when applied
as biofertilizers. This evidence was clearly appeared in growth criteria of wheat
and mung seeds as represented in Table 6.2 to 6.7. The growth profile of wheat
and mung plants, in terms of plant height and biomass (fresh and dry weight)
were enhanced by administration of cyanobacterial extracts (Fig 6.1 to Fig 6.7).
Figure 6.1 Comparison of growth between Cyanobacterization PBJ1 of the Mung Seed with control
Visual observation of wheat plants and comparison with control treatments
illustrated the significant improvement brought about by cyanobacterial
inoculation with the seed, with distinctly greener leaves and better plant
development (Fig. 6.1). Plant biometrical parameters, especially fresh and dry
weight were significantly higher in cyanobacterial PBG1 treated seeds.
Mung Control Mung Experimental

CHAPTER 6: Studies on
Parameter after 10 days
Germination %
Shoot Length in cm
Root Length in cm
No of leaves
Plant fresh weight in (mg)
Weight of fresh
Weight of dry root (
Weight of fresh
Weight of dry
Table 6.2 Effect of Cyanobacterium PBJ1 on Growth Parameters of Mung
Figure 6.2 Comparative study of effect of
0
20
40
60
80
100
120
140
160
Effect of Cyanobacterium PBJ1 on
Growth Parameters of Mung
Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
80
Parameter after 10 days Control Experimental
Germination % 70 ± 2.6
Length in cm 10.1± 0.13
Root Length in cm 9.1± 0.12
7.33± 1.52
Plant fresh weight in (mg) 101 ± 0.29
fresh root (mg) 63.6 ± 0.5
Weight of dry root (mg) 8.1 ± 0.20
Weight of fresh shoot (mg) 41.7 ± 0.29
Weight of dry Shoot (mg) 3.6 ± 0.3
Effect of Cyanobacterium PBJ1 on Growth Parameters of Mung at 10 Days after sowing
Comparative study of effect of Cyanobacterium PBJ1 inoculation vis a vis controls in the Mung
Effect of Cyanobacterium PBJ1 on
Growth Parameters of Mung
Control
Experimental
Plant Growth Promoting Activit of Cyanobacterial Isolates
Experimental
94 ± 1.6
17.4± 0.11
13.7± 0.11
12.00±1.73
150 ± 0.12
93.6± 0.11
11 ± 0.21
63.6 ± 0.32
4.5 ± 0.5
Effect of Cyanobacterium PBJ1 on Growth Parameters of Mung
Cyanobacterium PBJ1 inoculation
Effect of Cyanobacterium PBJ1 on
Control
Experimental

CHAPTER 6: Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
81
Data presented in the table 6.2, 6.3, 6.5 and 6.6 indicates the effect of
Cyanobacterial treatment on growth of mung and wheat seedlings in terms of root
and shoot length. The treatments with PBG1 culture showed significant
stimulatory effect on the length of roots and shoots.
Treatment of mung seeds with cyanobacteria PBJ1 significantly effect all growth
parameters.There was 20 % increase in germination % upon cyanobacterization
of seed with isolate PBJ1.
Days Control
(Shoot length in cm)
Experimental (Shoot length in cm)
2 1.7± 0.12 3.7± 0.10
4 3.6± 0.14 8.7± 0.14
6 5.3± 0.11 10.9± 0.10
8 6.9± 0.15 13.7± 0.13
10 10.1± 0.13 17.4± 0.11
Table 6.3 Effect of Cyanobacterium PBJ1 on Shoot length of Mung
Fig 6.3 Effect of Cyanobacterium PBJ1 on Shoot length of Mung
0
5
10
15
20
25
30
1 2 3 4 5
Cm
Effect of Cyanobacterium PBJ1 on Shoot
length of Mung
Experiment
al (Shoot
length in
cm)
Control
(Shoot
length in
cm)
2 4 6 8 10
No. of Days

CHAPTER 6: Studies on
Days (Root length in cm)
2
4
6
8
10
Table 6.4 Effect of Cyanobacterium PBJ
Fig 6.4 Effect of Cyanobacterium PBJ1 on
Figure 6.5 Comparison
0
5
10
15
20
25
1
Cm
Effect of Cyanobacterium PBJ1 on Root
Wheat Control
2
Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
82
Control
(Root length in cm)
Experimental
(Root length in cm)
2.0± 0.11 3.2± 0.12
3.2± 0.13 4.7± 0.14
5.6 ± 0.14 6.4± 0.10
6.3± 0.15 9.7± 0.12
9.1± 0.12 13.7± 0.11
Effect of Cyanobacterium PBJ1 on Root length of Mung
Effect of Cyanobacterium PBJ1 on Root length of Mung
ison of growth between Cyanobacterization PBJ1 of the wheat Seed with control
2 3 4 5
Effect of Cyanobacterium PBJ1 on Root
length of Mung
Experiment
al (Root
length in
cm)
Control
(Root length
in cm)
Wheat Control Wheat Experimental
2 4 6 8 10
No. of Days
Plant Growth Promoting Activit of Cyanobacterial Isolates
on Root length of Mung
length of Mung
between Cyanobacterization PBJ1 of the
Effect of Cyanobacterium PBJ1 on Root
Experiment
al (Root
length in
cm)
Control
(Root length
in cm)
Wheat Experimental
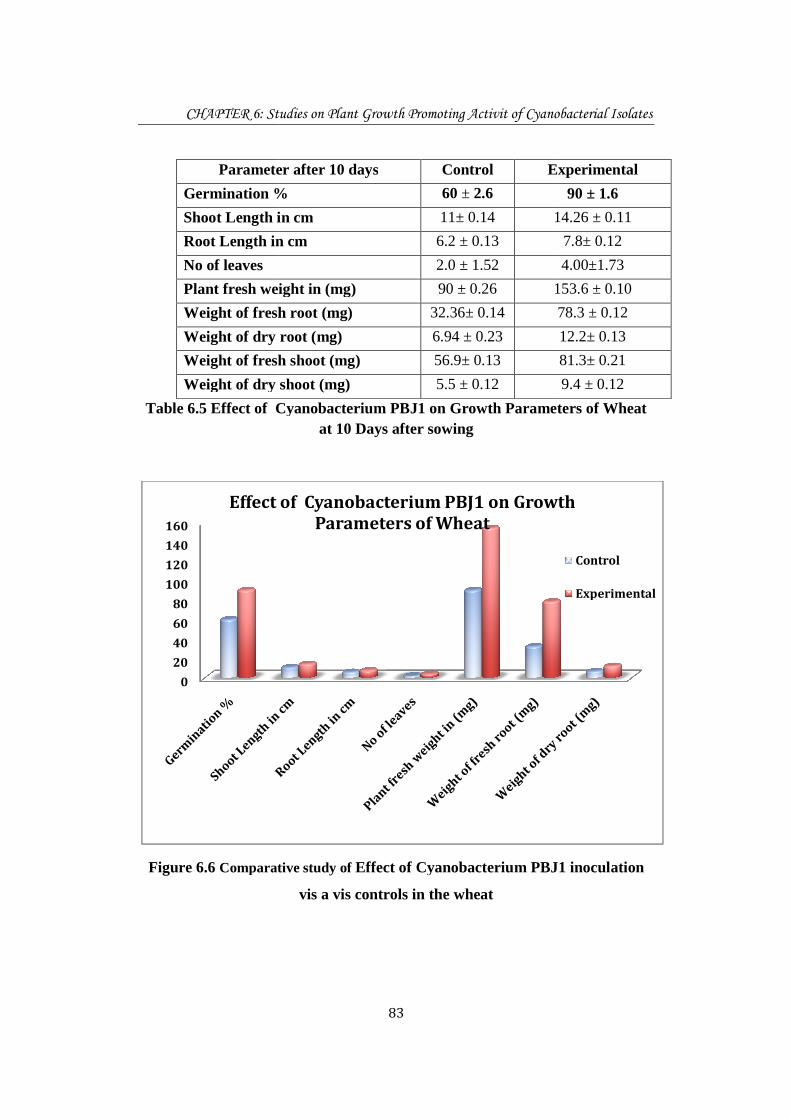
CHAPTER 6: Studies on
Parameter after 10 days
Germination %
Shoot Length in cm
Root Length in cm
No of leaves
Plant fresh weight in (mg)
Weight of fresh
Weight of dry root (
Weight of fresh
Weight of dry
Table 6.5 Effect of Cyanobacterium PBJ1 on Growth
Figure 6.6 Comparative
0
20
40
60
80
100
120
140
160
Effect of Cyanobacterium PBJ1 on Growth
Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
83
Parameter after 10 days Control Experimental
Germination % 60 ± 2.6
Shoot Length in cm 11± 0.14 14.26 ± 0.11
Root Length in cm 6.2 ± 0.13
2.0 ± 1.52
Plant fresh weight in (mg) 90 ± 0.26 153.6 ± 0.10
fresh root (mg) 32.36± 0.14
Weight of dry root (mg) 6.94 ± 0.23
Weight of fresh shoot (mg) 56.9± 0.13
Weight of dry shoot (mg) 5.5 ± 0.12
Effect of Cyanobacterium PBJ1 on Growth Parameters of Wheatat 10 Days after sowing
Comparative study of Effect of Cyanobacterium PBJ1 inoculation
vis a vis controls in the wheat
Effect of Cyanobacterium PBJ1 on Growth
Parameters of Wheat
Plant Growth Promoting Activit of Cyanobacterial Isolates
Experimental
90 ± 1.6
14.26 ± 0.11
7.8± 0.12
4.00±1.73
153.6 ± 0.10
78.3 ± 0.12
12.2± 0.13
81.3± 0.21
9.4 ± 0.12
Parameters of Wheat
Effect of Cyanobacterium PBJ1 inoculation
Effect of Cyanobacterium PBJ1 on Growth
Control
Experimental

CHAPTER 6: Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
84
Cyanobacterization of wheat seeds with isolate PBJ1 singnificantly enhanced the
growth parameters in the seedlings. There is 30 % increase in germination % in
cyanobacterized seed as compared to controls.
Days Control (Shoot length in cm)
Experimental (Shoot length in cm)
2 3± 0.12 4.5± 0.12
4 4.7± 0.18 5.1± 0.14
6 5.4± 0.11 7.5± 0.10
8 6.8± 0.15 10.5± 0.13
10 11± 0.14 14.26 ± 0.11
Table 6.6 Effect of Cyanobacterium PBJ2 on Shoot length of Wheat
Table 6.7 Effect of Cyanobacterium PBJ2 on Shoot length of Wheat
Days Control
(Root length in cm)
Sample
(Root length in cm)
2 1.5 ± 0.10 2.5± 0.14
4 2.5± 0.12 4.0± 0.11
6 3.6± 0.11 4.5 ± 0.12
8 4.2 ± 0.09 5.8 ± 0.10
10 6.2 ± 0.13 7.8± 0.12
Table 6.7 Effect of Cyanobacterium PBJ2 on Root length of Wheat
0
5
10
15
20
25
30
1 2 3 4 5
Cm
Effect of Cyanobacterium PBJ2 on Shoot length
of Wheat
Experimental
(Shoot length
in cm)
Control
(Shoot length
in cm)
2 4 6 8 10
No. of Days

CHAPTER 6: Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
85
Other growth parameters such as shoot length, root length, number of leaves, dry
and fresh weight of the seedlings were also positively effected by bacterization of
seed.
Table 6.8 Effect of Cyanobacterium PBJ2 on Root length of Wheat
Cyanobacteria produce bioactive compounds including plant growth regulators
that influence the physiological and biochemical profile of inoculated plants. The
objective of the present investigation was to study the effect of cyanobacterial
species PBJ1 on growth of mung and wheat parameters.Seeds presoaked in the
culture extract of cyanobacterial species PBJ1 shawed enhanced the germination
percentage, vegetative growth as well as chlorophyll content. These effects were
statistically proved significant. In all the treatments, the germination of wheat and
mung seeds were growing faster compared to the control.
0
2
4
6
8
10
12
14
16
1 2 3 4 5
Cm
s
Effect of Cyanobacterium PBJ2 on Root length
of Wheat
Sample (Root
length in cm)
Control (Root
length in cm)
2 4 6 8 10
No. of Days

CHAPTER 6: Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
86
6.4 REFERENCES
1. Abdul Baki, A.A. and Anderson, JD (1973) Vigor determinations in
soybean seed multiple criteria. Crop Sci.13:630–633.
2. Adam, M. S., (1999).The promotive effect of the cyanobacterium Nostoc
muscorum on the growth of some crop plants. Acta Microbiol. Polonica.
48: 163-171
3. Bashan, Y., (1998). Inoculants of plant growth-promoting bacteria for use
in agriculture. Biotechnol. Adv. 16, 729–770.
4. Balser, T.C., Kinzig, A.P., Firestone, M.K., (2001). Linking soil microbial
com-munities and ecosystem functioning. In: The Functional
Consequences of Biodiversity: Empirical Progress and Theoretical
Extensions. Princeton University Press, Princeton, NJ, 265–293.
5. Bohme, H. (1998). Regulation of nitrogen fixation in heterocyst-forming
cyanobacteria.Trends Plant Sci. 3 (9): 346-351.
6. Caire, G.; Storni de Cano, M.; Zaccaro de Mulé, M.C.; Palma, R.M. and
Colombo, K. 1997. Exopolysaccharides of Nostoc muscorum Ag.
(Cyanobacteria) in the aggregation of soil particles. J. Appl. Phycol. 9:
249-253.
7. Elias, S. G., and L. O. Copeland. (1997). Evaluation of seed vigor tests for
canola. Seed Technol. 19(1) 78-87.
8. Gupta, A.B.; Agarwal, P.R. 1973. Extraction, isolation and bioassay of a
gibberellin-like substance from Phormidium foveolarum. Ann. Bot.
37(152): 737-741.
9. Halperin, D.R.; Cano, M.; Zaccaro de Mulé‚ M.C.; Caire,G. (1992).
Diazotrophic Cyanobacteria from Argentine paddy fields. Int. J. Exp. Bot
(Phyton) 53: 135-142.
10. Karthikeyan, N., Prasanna, R.,(2009) Physiological characterization and
electron microscopic investigations of cyanobacteria associated with
wheat rhizosphere. Folia Microbiologica, 54, 43–51.

CHAPTER 6: Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
87
11. Karthikeyan, N., Prasanna, R., Lata., and Kaushik, B.D., (2007).
Evaluating the potential of plant growth promoting cyanobacteria as
inoculants for wheat. European Journal of Soil Biology, 43, 23–30.
12. Kim, S. H., Z. R. Choe, J. H. Kang, L. O. Copeland, and S. G. Elias.
(1994) Multiple seed vigor indices to predict field emergence and
performance of barley. Seed Sci. &Technol., 22:59-68.
13. Nanda, B., Tripathy, S.K. & Padhi, S., (1991). Effect of algalization on
seed germination of vegetable crops. World Journal of Microbiology
and Biotechnology. 7: 622–623.
14. Nisha R., A. Kaushik and C.P. Kaushik. (2007) Effect of indigenous
Cyanobacterial application on structural stability and productivity of an
organically poor semi-arid soil. Geoderma, 138, 49-56.
15. Prasanna, R., Triveni, S., Bidyarani, N., Babu, S., Yadav, K., Saxena, A.
K. (2014). Evaluating the efficacy of cyanobacterial formulations and
biofilmed inoculants for leguminous crops. Archives of Agronomy and
Soil Science, 60(3), 349-3
16. Prasanna R, Sood A, Jaiswal P, Nayak S,Gupta V, Chaudhary V, Joshi M
and Natarajan C, (2010).Rediscovering Cyanobacteria as valuable sources
of bioactive compounds. Appl. Biochem Micro. 46: 119-134.
17. Roger, P.A., Zimmerman, W.J., Lumpkin, T.A., (1993). Microbiological
management of wetland rice fields. In: Metting, B. (Ed.), Soil Microbial
Ecology- Applicationin Agricultural and Environment al
Management. Marcel Dekker, New York,.417–455.
18. Sergeeva E, Liaimer A and Bergman B, (2002). Evidence for production
of the phytohormone indole-3-acetic acid by cyanobacteria. Planta, 215:
229-238.
19. Singh, U.P., Sharma, B.K., Singh, D.P., (2003). Effect of plant growth
promoting rhi-zobacteria and culture filtrate of Sclerotium rolfsii on
phenolic and salicylic acidcontents in chickpea. Curr. Microbiol. 46,
131–140.

CHAPTER 6: Studies on Plant Growth Promoting Activit of Cyanobacterial Isolates
88
20. Soltani, A., M. Gholipoor and E. Zeinali. (2006). Seed reserve utilization
and seedling growth of wheat as affected by drought and salinity.
Environ. Exp.Bot., 55, 195-200.
21. Stefan, M., Munteanu, N., Stoleru, V., Mihasan, M., Hritcu, L., (2013).
Seed inoculationwith Plant growth promoting rhizobacteria enhances
photosynthesis and yieldof runner bean (Phaseolus coccineus L). Sci.
Hortic. 151, 22–29.
22. Venkataraman, G.S. & Neelakantan, S. (1967). Effect of the cellular
constituents of the nitrogen fixing blue-green algae Cylindrospermum
muscicola on the root growth of rice seedlings. Journal of General and
Applied Microbiology 13: 53–61.
23. Wang, S. M., Wang, Q. L., Li, S. H. and Zhang, J. R., (1991). A study of
treatment of spring wheat with growth promoting substances from
nitrogen-fixing blue green algae. Acta Hydrobiol. Sin. 15: 45-52.
24. Yadav, S., Sinha, R. P., Tyagi, M. B., Kumar, A.,(2011) Cyanobacterial
secondary metabolites.Intr. J. of Pharma and Biosciences. 2: 144-167.





























