Embed Size (px)
Citation preview

Chapter 8 Keratinolytic protease production by Bacillus amyloliquefaciens 6B using feather meal as substrate and application of feather hydrolysate as biocontrol (antifungal) as well as organic nitrogen input for agricultural soil This part has been communicated as: Anjali Bose, Shabnam Pathan, Khyati Pathak and Haresh Keharia (2012). Keratinolytic protease production by Bacillus amyloliquefaciens 6B using feather meal as substrate and application of feather hydrolysate as soil input. Waste & Biomass Valorization.

Feather as substrate for keratinase production Chapter 8
156
8.1. Introduction
Low commercial value keratinous wastes of animal origin such as nails, hair,
feathers, horns, hoofs, beak etc., which are generated in huge amount from poultry and
cattle slaughter houses, are hard-to-degrade. Considering the fact that feathers constitute
nearly 8.5% of the total weight of a chicken, the amount of solid waste generated
annually in India from approximately 823.5 million chickens, with an average
individual body weight of 2 kg, would be approximately 140 million kg (FAO 2002).
Currently, the waste feathers are either disposed by dumping or incineration which can
lead to serious health hazards and environmental pollution (Deydier et al 2005).
Alternately, they are transformed into animal feed supplement by harsh hydrothermal
and chemical treatment that result in loss of nutritionally essential amino acids, such as
methionine, lysine, and tryptophan, and formation of non-nutritive amino acids, such as
lysinoalanine and lanthionine (Dalev et al 1997). Consequently, microbial degradation
of such wastes along with production of commercially important enzymes (especially
keratinase) and other value-added products such as nutrient-rich animal feed or fertilizer
(Onifade et al 1998, Brandelli 2008, Jeong et al 2010, Lateef et al 2010) is desirable.
Contemporary to this problem, is the development of new fertilizers using natural
materials. In the same context, feather hydrolysate resulting from the bioconversion of
keratinous wastes being rich in nitrogen and inexpensive could be used as a potential
fertilizer for soil amendment (Gupta and Ramnani 2006), serving the dual purpose of
improving plant growth and stimulating microbial activity in soil (Vasileva-Tonkova et
al 2009). On the other hand, inadequate effectiveness of chemical control and
increasing restrictions on the use of fungicide/ pesticides have given increasing impetus
to biological control as an alternative tool for disease management in agriculture
(Raaijmakers et al 2002). Hence, microorganisms capable of degrading keratinous
waste as well as exhibiting antagonism against soil borne plant pathogens hold
prospects of dual benefit, waste utilization along with production of biocontrol
metabolites. Most of investigations reported in literature on keratinolytic
microorganisms have been directed toward keratinase production for increasing the
digestibility of feathers as an animal feed (Onifade et al 1998, Odetallah et al 2003,
Grazziotin et al 2006, Fakhfakh et al 2011). However, there is only one report on the
isolation and characterization of keratinolytic Stenotrophomonas maltophilia R13 with

Feather as substrate for keratinase production Chapter 8
157
plant growth-promoting activity (Jeong et al 2010). To the best of our knowledge, there
is no report on keratinolytic Bacillus sp. exhibiting biocontrol activity.
From this perspective, the poultry soil isolate Bacillus amyloliquefaciens 6B
(BA6B) was investigated for: its ability to grow on and efficiently degrade whole
feathers, production of keratinolytic protease and antifungal metabolites. Additionally,
the enzyme and the antifungal metabolites produced by BA6B were characterized.
Further, the fermentation broth with the bacterial cells and hydrolyzed feather was
investigated as nitrogenous soil input.
8.2. Materials and Methods
8.2.1. Materials
Bushnell and Haas Medium (BHM), Luria Bertani Broth, gelatin, yeast extract
and bovine serum albumin (BSA, protein standard) were obtained from Hi-Media,
India. Hammerstein casein was procured from Sisco Research Laboratories, India.
Chicken feathers were procured from a local poultry (Karamsad, Anand, Gujarat, India),
washed with tap water and air dried. The feathers were then ground to get feather meal
which was added to basal salt medium as a substrate for enzyme production. All other
solvents and chemicals used during the experiment were of analytical grade.
Phytopathogenic fungi namely, Aspergillus niger MTCC 16404, Aspergillus fumigatus
MTCC 343, Aspergillus parasiticus NCIM 898, Trichosporon sp. NCIM 1110,
Fusarium oxysporum NCIM 1072 were obtained from National Collection of Industrial
Microorganisms (NCIM), National Chemical Laboratory, Pune, India and Microbial
Type Culture Collection (MTCC), Institute of Microbial Technology, Chandigarh,
India.
8.2.2. Isolation and identification of keratin hydrolysing bacterium
Feather degrading bacteria were isolated upon enrichment of the poultry soil
(Karamsad, Gujarat) with whole feathers for 15 days. Upon incubation a 10% (w/v) soil
suspension was prepared in sterile distilled water and 100 µL aliquots of serially diluted
soil suspension were spread on 1% casein agar plates and incubated at 37°C for 48 to 72
h. Individual colonies exhibiting clear zones around them resulting from casein
hydrolysis were picked and purified by repeated sub-culturing on Luria Bertani agar
plate. The pure cultures were further screened for their ability to degrade feather on

Feather as substrate for keratinase production Chapter 8
158
feather meal agar (BHM supplemented with 1% w/v feather meal; FMA) plates. The
bacterial isolates exhibiting feather degradation on FMA plates were then investigated
for native feather degradation. The isolate exhibiting the highest keratinolytic activity
was then selected for further studies.
The identity of the selected bacterial isolate designated as 6B was determined on
the basis of 16S rRNA gene sequence analysis. The genomic DNA was extracted as
described by Ausubel et al (1997) and the 16S rRNA gene was amplified and sequenced
as described in Chapter 2. Further, the sequence has been submitted to Genbank at
NCBI.
8.2.3. Organic solvent tolerance of bacterial isolate 6B
The bacterium 6B was spot inoculated on casein agar plates using sterile tooth
picks upon which each plate was flooded with 7 mL of different solvents (iso-octane, n-
heptane, n-hexane, cyclohexane, xylene, toluene, benzene, chloroform, ethyl acetate and
methanol) and incubated at 37°C in air tight canister in order to prevent the evaporation
of solvent from the plates. Upon incubation, plates were observed for growth as well as
clear zone around the colony in the presence of solvents.
The stability of keratinolytic protease in organic solvents was investigated by
mixing equal volume of solvent and culture supernatant (crude enzyme) in screw cap
test tubes and incubated in shaking water bath (50 rpm) at 37 C. After 24 h of
incubation residual protease activity was assayed in the aqueous phase.
8.2.4. Profile of keratinolytic protease production by bacterial isolate 6B
The 500 mL Erlenmeyer flasks containing 200 mL feather meal broth (BHM
amended with 0.2% w/v feather meal; FMB) was inoculated with an overnight grown
culture of 6B to obtain an initial culture density (O.D.600nm) of 0.05 and incubated on
orbital shaker (150 rpm) at 37 C. The 2 mL aliquots were withdrawn after 3, 6, 9, 12,
24, 36 and 48 h of incubation, and monitored for growth and protease activity.
8.2.5. Medium optimization for keratinolytic protease production by bacterial isolate
6B
Various sugars (fructose, galactose, glucose, lactose, maltose, mannitol, sucrose
and xylose; 0.5% w/v) were supplemented individually to FMB, in order to investigate

Feather as substrate for keratinase production Chapter 8
159
their effect as co-carbon sources on the enzyme production by 6B. The influence of
varying concentrations (0.1% to 2%, w/v) of feather meal on the keratinolytic enzyme
production was also investigated.
8.2.6. Concentration of enzyme
The fermentation broth was harvested after 12 h of incubation and subjected to
centrifugation at 4°C and 12,310 ×g for 15 min. The supernatant was collected and
subjected to 40-80% ammonium sulphate saturation and kept overnight at 20°C to allow
protein precipitation. The precipitates were then harvested by centrifugation at 4°C and
27,540 ×g for 20 min. The pellet thus obtained was resuspended in minimum volume of
50 mM Tris-Cl buffer (pH 8.0) to allow solubilization of proteins and dialyzed against
same buffer overnight at 4°C.
The dialyzed enzyme preparation was then analyzed by Native and SDS-
polyacrylamide gel (10%, w/v) electrophoresis according to the method of Laemmli
(1970). One part of gel was subjected to activity staining in order to visualize protease
band and the other part was subjected to silver staining (Sambrook and Russell 2001) in
order to visualize protein bands. For activity staining, the native gel was transferred
onto a casein agar slab (1% casein and 1% (w/v) agar in 50 mM Tris-Cl buffer pH, 8.0)
and incubated at 37°C till transparent bands of protease activity appeared due to casein
hydrolysis in the agar slab. In case of SDS-PAGE, after electrophoresis, SDS was
removed by washing the gel for 15 min with 2.5% Triton X-100 and then twice for 10
min each with distilled water. Then, the gel was subjected to activity staining as
described above. Relative molecular mass (Mr) was estimated by comparison with
molecular mass standards (3.5 to 97.4 kDa) (Bangalore Genie, Bangalore, India).
8.2.7. Characterization of crude keratinolytic protease from isolate 6B
The keratinolytic protease activity was determined at different temperatures viz.,
30, 40, 50, 60, 70, 80°C in presence of 50 mM Tris-Cl buffer, pH 8.0. The
thermostability of enzyme was determined by pre-incubating the enzyme at 50 and
60°C in shaking water bath for varying time intervals upto 120 min and then assayed for
residual activity at 50ºC, which was expressed as percentage of initial activity.
The activity of keratinolytic protease was monitored over a pH range of 5.0 to
11.0 at 37°C. Sodium acetate buffer (50 mM) for pH 5.0-5.5, sodium phosphate buffer

Feather as substrate for keratinase production Chapter 8
160
(50 mM) for pH in range of 6.0 to 7.5, Tris-Cl buffer (50 mM) for pH 8.0 to 8.5 and
glycine-NaOH (50 mM) for pH in range of 9.0 to 11.0 were used to adjust the pH of the
reaction mixture. The pH stability of the enzyme was estimated by pre-incubating the
enzyme in buffers of various pH for 1 h and then assayed for protease activity at
optimum pH.
To determine the effect of various additives viz., metal ions (Ca2+
, Cu2+
, Fe2+
,
Mg2+
, Mn2+
, Hg2+
, Zn2+
; 5 mM), enzyme inhibitors (EDTA and PMSF; 5 mM),
surfactants (SDS, CTAB, Triton X-100, Tween 80; 1% w/v), and bleach-oxidant (H2O2;
1% v/v); the enzyme was pre-incubated with these additives for 1 h at 37°C and then
assayed using casein as a substrate. The enzyme activity was expressed as % activity
relative to control (without any of the aforementioned additives) which was considered
as 100%.
8.2.8. In vitro screening for antifungal activity
A co-culture technique (Trivedi et al 2008) was used to test the ability of
bacterium 6B to inhibit the growth of several phytopathogenic fungi (A. niger MTCC
16404, A. parasiticus NCIM 898, A. fumigatus MTCC 343, Trichosporon sp. NCIM
1110 and F. oxysporum NCIM 1072). The bacterium was spot inoculated on one side of
PDA plate and incubated at 37 C for 48 h after which each fungal strain was inoculated
individually, next to the bacterial culture and the plates were incubated at 27°C up to 5 d
for monitoring antifungal activity.
The antifungal activity of the culture supernatant harvested from stationary-phase
culture of isolate 6B was bioassayed against the above mentioned fungi using agar well
diffusion method. Briefly, the culture supernatant (100 µL) was inoculated in the center
of PDA plates which had been inoculated previously with test fungus. After the plates
were incubated at 28°C for 5 d, the inhibition of fungal growth was observed around the
well .The control plate was prepared by growing each test fungus with uninoculated
feather medium.
Further the antifungal metabolites from culture broth were precipitated by
lowering the pH of cell free supernatant upto 2.0 using (6M) HCl. The precipitate thus
obtained was solubilized in methanol, which was then analyzed by ESI-MS. Mass
spectra was recorded in positive ion mode using Esquire 3000 plus mass spectrometer
(Bruker Daltonics, Germany) consisting of two octopole followed by an ion trap.

Feather as substrate for keratinase production Chapter 8
161
Helium was used as collision gas for collision induced dissociation (CID) experiment
and the data obtained was processed using Esquire data analysis software, version 3.1.
8.2.9. Use of the feather hydrolysate as a bioactive agricultural nitrogen input (30 d
plant growth assay).
Pot study was carried out in triplicates to compare the ability of feather
hydrolysate and the reference commercial fertilizer as agriculture input and its effect on
the growth of Vigna radiate var. meha (mung). The seeds (5 seeds/test pot) were
planted in 6 liters plastic pots with 4 kg garden soil. The feather hydrolysate
(concentrated five times using rotary evaporator at 60°C; 153 mL of feather
hydrolysates/kg soil) and the fertilizer (625 mg of fertilizer/kg soil) were diluted with
tap water and poured onto the top of the soil to obtain the desired concentration of
soluble nitrogen (100 mg of N/kg soil). All the pots were watered regularly with tap
water to replace the evaporation loss and after 30 days of sowing, all plants were
carefully uprooted and washed. Various growth parameters such as plant height, plant
fresh weight, root length and root fresh weight were recorded.
8.2.10. Analytical procedures: enzyme activity
The enzyme activity was determined as per the modified method of Khardenavis
et al (2009). In brief, 2 mL of reaction mixture containing 1 mL of 1% (w/v) casein
(acc. to Hammerstein) and 1 mL of appropriately diluted enzyme in 50 mM Tris-Cl
buffer (pH 8.0) was incubated for 20 min at 50°C. The reaction was stopped by
precipitation of the residual substrate with 2 mL 10% (w/v) trichloroacetic acid. A
control was run in parallel in which the enzyme was added after addition of
trichloroacetic acid. The reaction mixture was then centrifuged at 13,201 ×g for 10 min
and supernatant was assayed for tyrosine by reading its absorbance was read at 280 nm
(absorption maxima for tyrosine) against the assay blank, using UV-Visible
spectrophotometer (Thermo Electron corporation, HEλIOS α). The standard curve of
tyrosine was prepared by reading absorbance of different dilutions of 500 µg
tyrosine/mL stock. One unit of protease activity was defined as the amount of enzyme
required to liberate 1 μg of tyrosine/min.mL under specified assay conditions. The total
protein was estimated according to the method of Lowry et al (1951) using bovine
serum albumin (fraction V) as standard.
Feather as substrate for keratinase production Chapter 8
161.1
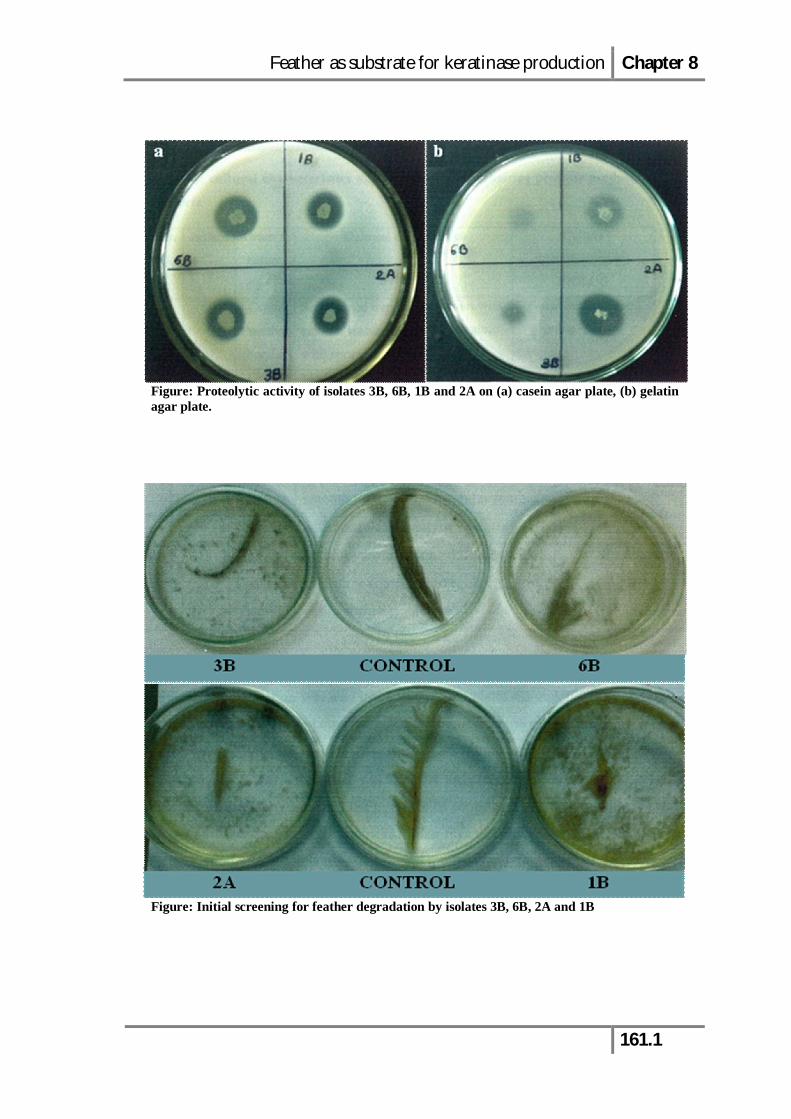
Figure: Proteolytic activity of isolates 3B, 6B, 1B and 2A on (a) casein agar plate, (b) gelatin agar plate.
Figure: Initial screening for feather degradation by isolates 3B, 6B, 2A and 1B

Feather as substrate for keratinase production Chapter 8
162
8.1. Results and Discussion
8.1.1. Isolation and identification of keratin hydrolysing bacterium
Thirteen proteolytic bacterial strains isolated from the poultry farm soil sample
were screened for feather degradation on FMA plates. The growth of bacterial isolates
on FMA indicated their ability to utilize feather keratin as a sole carbon source for
growth. Amongst all, four isolates designated as 1B, 2A, 3B and 6B were selected on
the basis of their growth on FMA and were then screened for their ability to degrade
native feather. Among the four isolates, 6B exhibited complete degradation of intact
feather within 24 h of incubation at 37°C (Fig. 8.1).
Figure 8.1 Feather degradation by isolate designated 6B
The isolate 6B was found to be a Gram positive, spore-forming bacillus. The
nucleotide sequence of 16S rRNA gene of this isolate exhibited 100% sequence
homology with the partial sequence of Bacillus amyloliquefaciens DJFZ40 (GU568197)
16S rRNA gene. The 1,354 bp sequence of isolate 6B (Appendix 3) identified as B.
amyloliquefaciens has been submitted to NCBI GenBank (JQ904625).
8.1.2. Organic solvent tolerance of B. amyloliquefaciens 6B (BA6B)
The solvent tolerance of BA6B was determined by its ability to grow on casein
agar plates flooded with different solvents (iso-octane log Pow = 4.5, n-heptane log Pow
= 4.39, n-hexane log Pow = 3.86, cyclohexane log Pow = 3.2, xylene log Pow = 3.1,
toluene log Pow = 2.64, benzene log Pow = 2.13, chloroform log Pow = 2.0, ethyl acetate
Pow = 0.68 and methanol log Pow = -0.764) during incubation. The BA6B exhibited
growth as well as protease production in presence of all the solvents except ethyl acetate
Feather as substrate for keratinase production Chapter 8
163
and methanol, with maximum zone of hydrolysis on plates flooded with iso-octane
(Table 8.1).
Table 8.1: Organic solvent tolerance of B. amyloliquefaciens 6B and its protease.
Organic solvents log Pow Zone/Growth ratio Relative enzyme activity (%)
None (Control) 4.3 100
iso-Octane 4.5 4.5 99
n-Heptane 4.39 4.1 94
n-Hexane 3.86 3.6 85
Cyclohexane 3.2 2.6 82
Xylene 3.1 2.8 70
Toluene 2.64 2.4 46
Benzene 2.13 1.6 39
Chloroform 2.0 1.4 22
Ethyl acetate 0.68 ND 7
Methanol -0.76 ND 4
In general, organic solvents with a log Pow between 1.5 and 4.0 are extremely
toxic to microorganisms and other living cells because they partition preferentially in
the cytoplasmic membrane, disorganizing its structure and impairing vital functions
(Inoue and Horikoshi 1991). However, solvent toxicity not only depends on the
inherent toxicity of the solvent but also on the intrinsic tolerance of the bacterial species
and strains (Sikkema et al 1995). There exits few protease producing Bacillus sp. that
can grow in presence of toxic organic solvents such as DMSO, methanol, toluene,
xylene, cyclohexane, hexane and iso-octane (Gorbel et al 2003, Sareen and Mishra
2008, Fang et al 2009, Li et al 2009). The solvent tolerance in such microorganisms has
been attributed to the alteration in cell morphology, composition of the membrane fatty
acids and/or presence of active efflux pumps which maintain minimal sub toxic
concentration of organic solvents within cells (Torres et al 2011).
The stability of crude enzyme from BA6B was determined in presence of all the
above organic solvents at 50% (v/v) concentration upto 24 h (Table 8.1). The enzyme
was found stable in presence of non-polar (high log Pow value) solvents, retaining more
than 80% activity, whereas, the enzyme activity was significantly inhibited in presence
of solvents with lower log Pow value such as chloroform, ethyl acetate and methanol.
Low log Pow value corresponds to greater polarity and toxicity of the solvent to the
enzyme (Inoue and Horikoshi 1991). Similar inactivation of keratinase from B. subtilis
KD-N2 in presence of polar solvents was reported by Cai et al (2008). In contradiction
to our observations, keratinase from Kocuria rosea (Bernal et al 2006),
Feather as substrate for keratinase production Chapter 8
164
Fervidobacterium islandicum (Nam et al 2002) and Chryseobacterium sp. kr6 (Riffle et
al 2003) have been reported to be stable in presence of even polar organic solvents.
8.1.3. Production profile of the extracellular keratinolytic protease by BA6B in
batch culture
The isolate BA6B was able to produce keratinolytic protease in medium
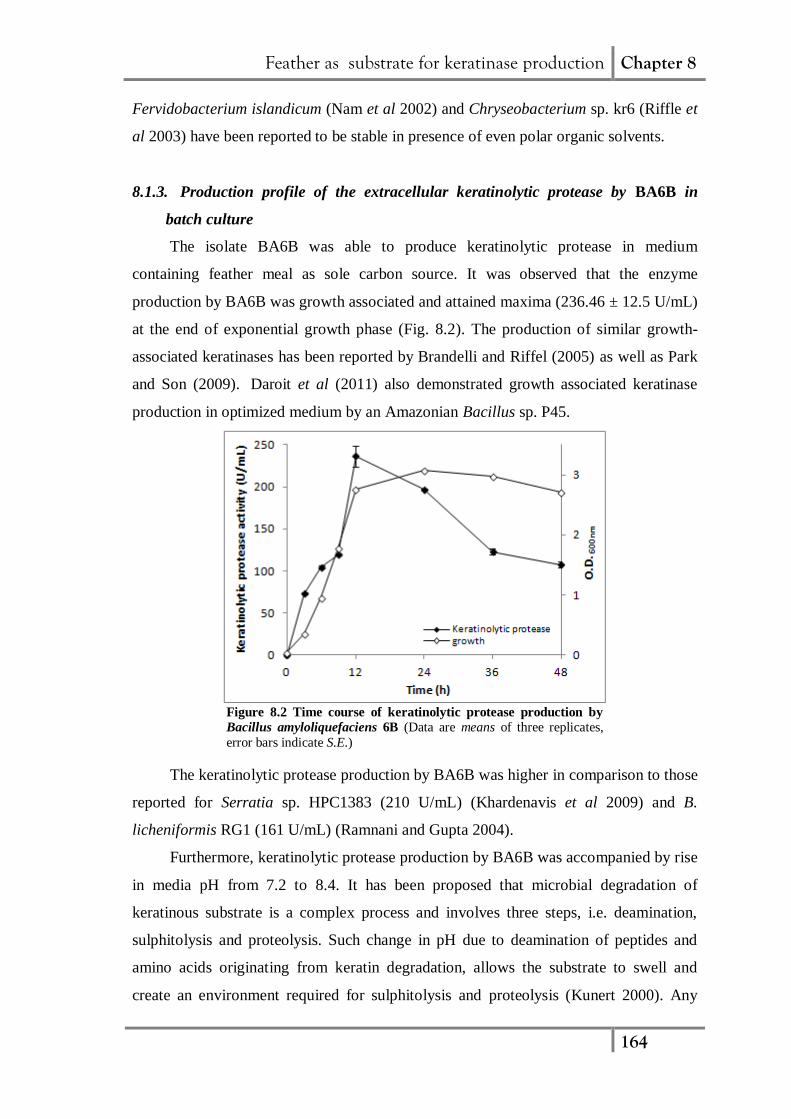
containing feather meal as sole carbon source. It was observed that the enzyme
production by BA6B was growth associated and attained maxima (236.46 ± 12.5 U/mL)
at the end of exponential growth phase (Fig. 8.2). The production of similar growth-
associated keratinases has been reported by Brandelli and Riffel (2005) as well as Park
and Son (2009). Daroit et al (2011) also demonstrated growth associated keratinase
production in optimized medium by an Amazonian Bacillus sp. P45.
Figure 8.2 Time course of keratinolytic protease production by
Bacillus amyloliquefaciens 6B (Data are means of three replicates,
error bars indicate S.E.)
The keratinolytic protease production by BA6B was higher in comparison to those
reported for Serratia sp. HPC1383 (210 U/mL) (Khardenavis et al 2009) and B.
licheniformis RG1 (161 U/mL) (Ramnani and Gupta 2004).
Furthermore, keratinolytic protease production by BA6B was accompanied by rise
in media pH from 7.2 to 8.4. It has been proposed that microbial degradation of
keratinous substrate is a complex process and involves three steps, i.e. deamination,
sulphitolysis and proteolysis. Such change in pH due to deamination of peptides and
amino acids originating from keratin degradation, allows the substrate to swell and
create an environment required for sulphitolysis and proteolysis (Kunert 2000). Any
Feather as substrate for keratinase production Chapter 8
165
further increase in pH was limited by the volatility of ammonia, which escapes from
culture broth in gaseous form. Similar increase in medium pH from neutral to alkaline,
have been reported during keratinase production by several other feather-degrading
bacteria (Park and Son 2009, Lateef et al 2010) and fungi (Kaul and Sumbali 1999).
8.1.4. Optimization of media components and culture conditions for BA6B
keratinolytic protease production
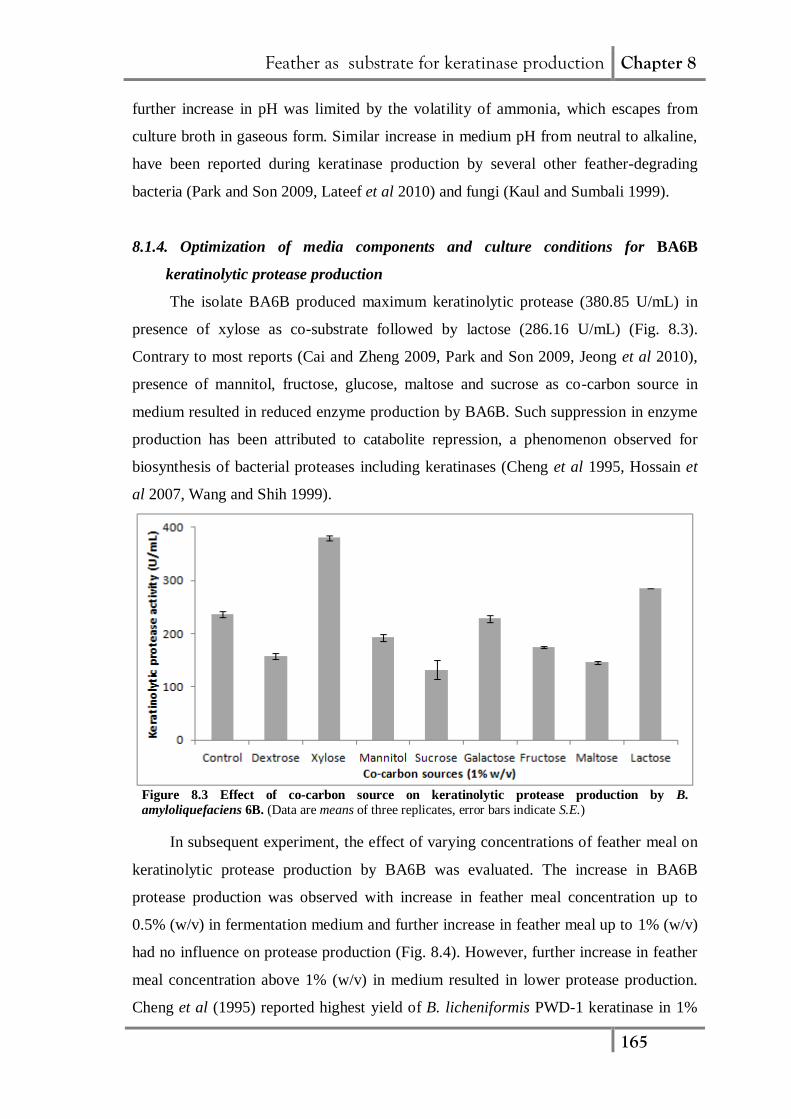
The isolate BA6B produced maximum keratinolytic protease (380.85 U/mL) in
presence of xylose as co-substrate followed by lactose (286.16 U/mL) (Fig. 8.3).
Contrary to most reports (Cai and Zheng 2009, Park and Son 2009, Jeong et al 2010),
presence of mannitol, fructose, glucose, maltose and sucrose as co-carbon source in
medium resulted in reduced enzyme production by BA6B. Such suppression in enzyme
production has been attributed to catabolite repression, a phenomenon observed for
biosynthesis of bacterial proteases including keratinases (Cheng et al 1995, Hossain et
al 2007, Wang and Shih 1999).
Figure 8.3 Effect of co-carbon source on keratinolytic protease production by B.
amyloliquefaciens 6B. (Data are means of three replicates, error bars indicate S.E.)
In subsequent experiment, the effect of varying concentrations of feather meal on
keratinolytic protease production by BA6B was evaluated. The increase in BA6B
protease production was observed with increase in feather meal concentration up to
0.5% (w/v) in fermentation medium and further increase in feather meal up to 1% (w/v)
had no influence on protease production (Fig. 8.4). However, further increase in feather
meal concentration above 1% (w/v) in medium resulted in lower protease production.
Cheng et al (1995) reported highest yield of B. licheniformis PWD-1 keratinase in 1%
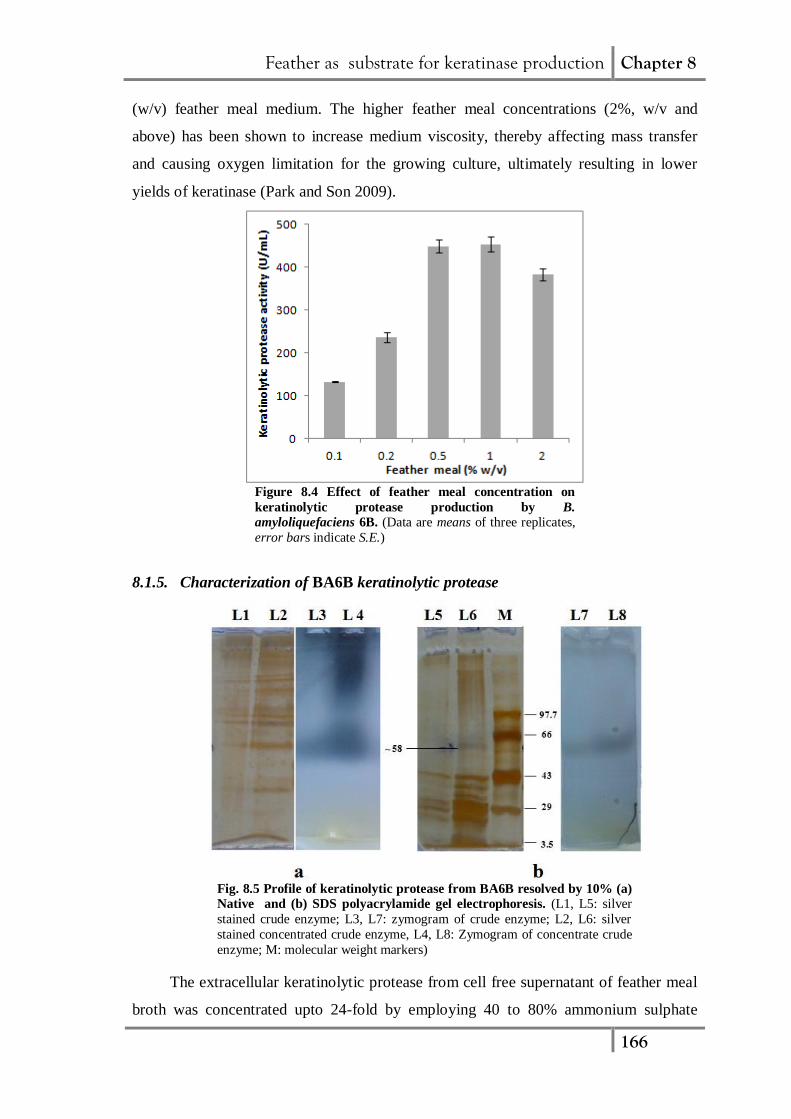
Feather as substrate for keratinase production Chapter 8
166
(w/v) feather meal medium. The higher feather meal concentrations (2%, w/v and
above) has been shown to increase medium viscosity, thereby affecting mass transfer
and causing oxygen limitation for the growing culture, ultimately resulting in lower
yields of keratinase (Park and Son 2009).
Figure 8.4 Effect of feather meal concentration on
keratinolytic protease production by B.
amyloliquefaciens 6B. (Data are means of three replicates,
error bars indicate S.E.)
8.1.5. Characterization of BA6B keratinolytic protease
Fig. 8.5 Profile of keratinolytic protease from BA6B resolved by 10% (a)
Native and (b) SDS polyacrylamide gel electrophoresis. (L1, L5: silver
stained crude enzyme; L3, L7: zymogram of crude enzyme; L2, L6: silver
stained concentrated crude enzyme, L4, L8: Zymogram of concentrate crude
enzyme; M: molecular weight markers)
The extracellular keratinolytic protease from cell free supernatant of feather meal
broth was concentrated upto 24-fold by employing 40 to 80% ammonium sulphate
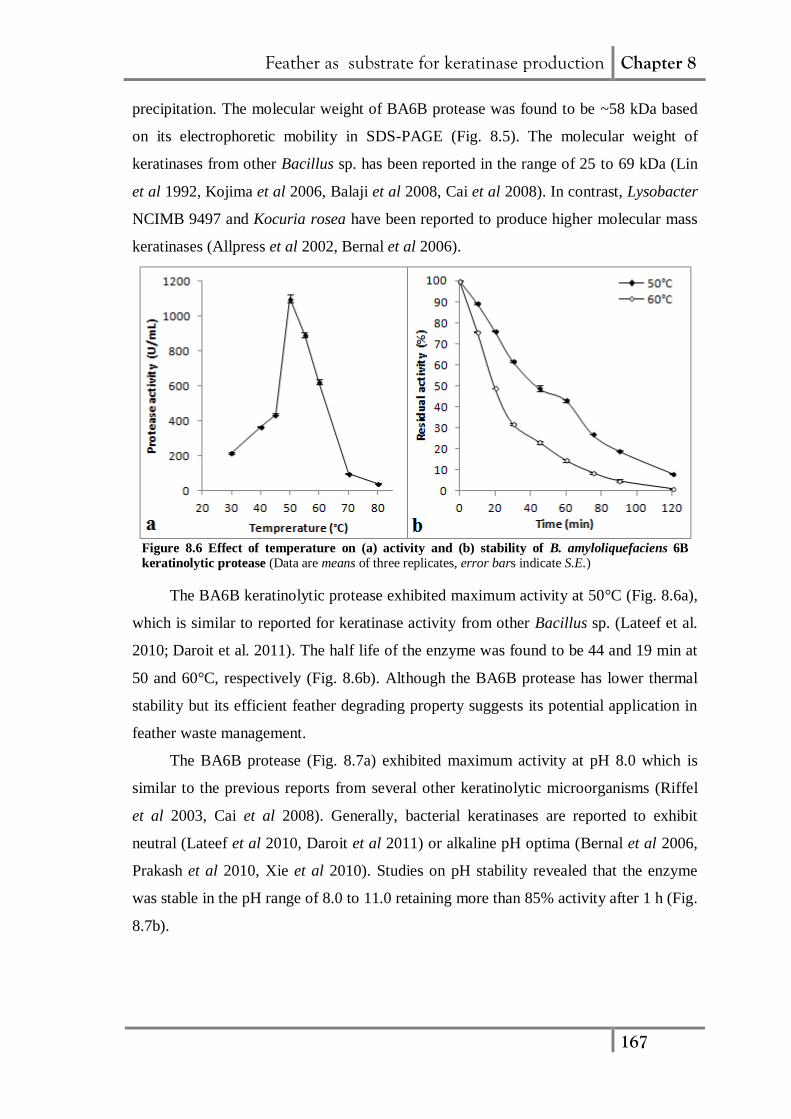
Feather as substrate for keratinase production Chapter 8
167
precipitation. The molecular weight of BA6B protease was found to be ~58 kDa based
on its electrophoretic mobility in SDS-PAGE (Fig. 8.5). The molecular weight of
keratinases from other Bacillus sp. has been reported in the range of 25 to 69 kDa (Lin
et al 1992, Kojima et al 2006, Balaji et al 2008, Cai et al 2008). In contrast, Lysobacter
NCIMB 9497 and Kocuria rosea have been reported to produce higher molecular mass
keratinases (Allpress et al 2002, Bernal et al 2006).
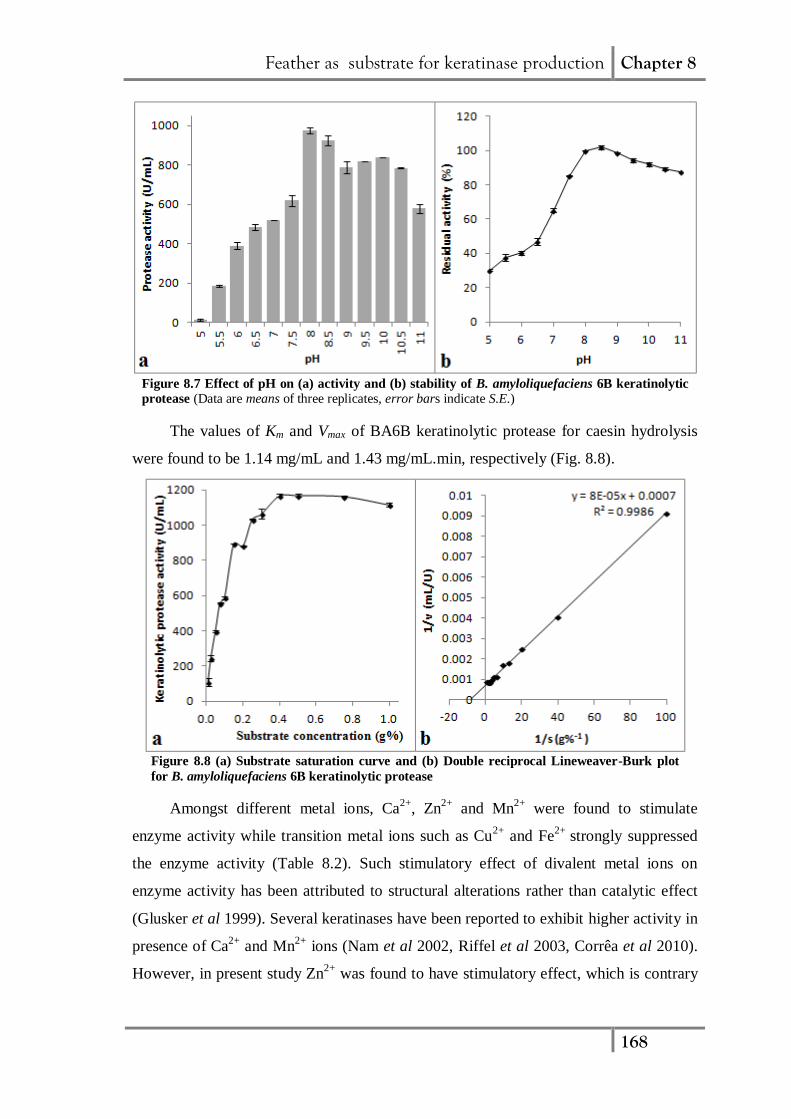
Figure 8.6 Effect of temperature on (a) activity and (b) stability of B. amyloliquefaciens 6B
keratinolytic protease (Data are means of three replicates, error bars indicate S.E.)
The BA6B keratinolytic protease exhibited maximum activity at 50°C (Fig. 8.6a),
which is similar to reported for keratinase activity from other Bacillus sp. (Lateef et al.
2010; Daroit et al. 2011). The half life of the enzyme was found to be 44 and 19 min at
50 and 60°C, respectively (Fig. 8.6b). Although the BA6B protease has lower thermal
stability but its efficient feather degrading property suggests its potential application in
feather waste management.
The BA6B protease (Fig. 8.7a) exhibited maximum activity at pH 8.0 which is
similar to the previous reports from several other keratinolytic microorganisms (Riffel
et al 2003, Cai et al 2008). Generally, bacterial keratinases are reported to exhibit
neutral (Lateef et al 2010, Daroit et al 2011) or alkaline pH optima (Bernal et al 2006,
Prakash et al 2010, Xie et al 2010). Studies on pH stability revealed that the enzyme
was stable in the pH range of 8.0 to 11.0 retaining more than 85% activity after 1 h (Fig.
8.7b).
Feather as substrate for keratinase production Chapter 8
168
Figure 8.7 Effect of pH on (a) activity and (b) stability of B. amyloliquefaciens 6B keratinolytic
protease (Data are means of three replicates, error bars indicate S.E.)
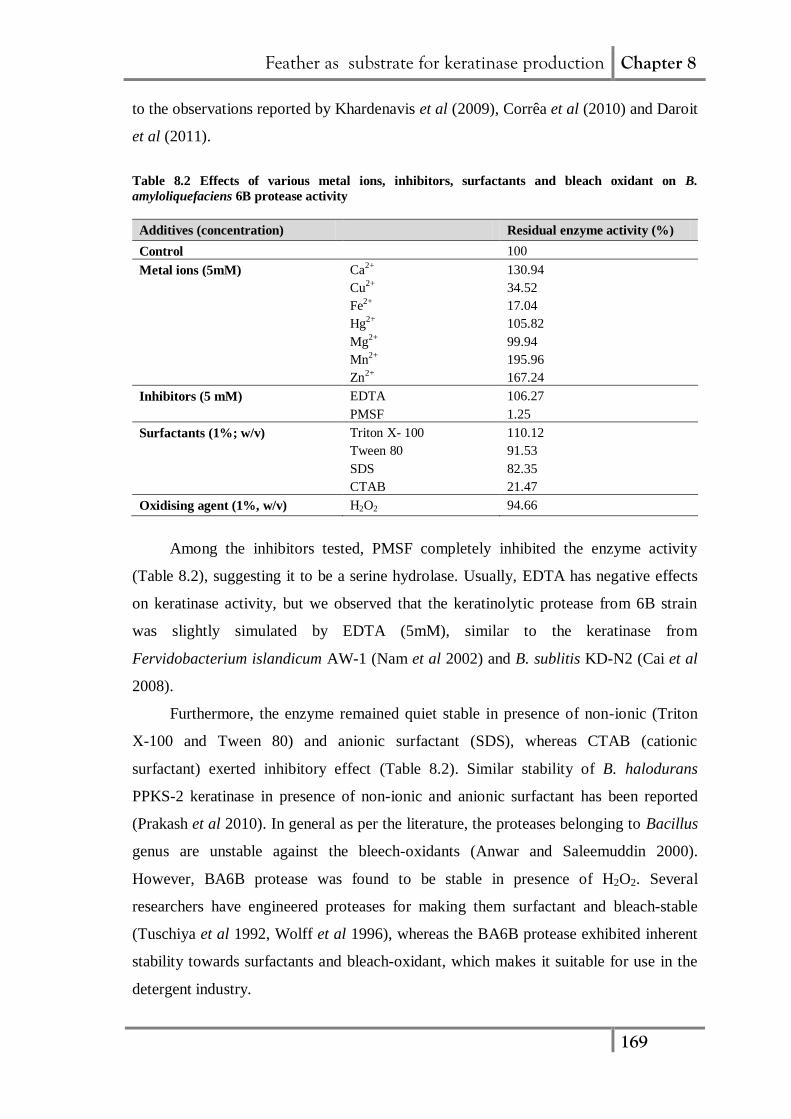
The values of Km and Vmax of BA6B keratinolytic protease for caesin hydrolysis
were found to be 1.14 mg/mL and 1.43 mg/mL.min, respectively (Fig. 8.8).
Figure 8.8 (a) Substrate saturation curve and (b) Double reciprocal Lineweaver-Burk plot
for B. amyloliquefaciens 6B keratinolytic protease
Amongst different metal ions, Ca2+
, Zn2+
and Mn2+
were found to stimulate
enzyme activity while transition metal ions such as Cu2+
and Fe2+
strongly suppressed
the enzyme activity (Table 8.2). Such stimulatory effect of divalent metal ions on
enzyme activity has been attributed to structural alterations rather than catalytic effect
(Glusker et al 1999). Several keratinases have been reported to exhibit higher activity in
presence of Ca2+
and Mn2+
ions (Nam et al 2002, Riffel et al 2003, Corrêa et al 2010).
However, in present study Zn2+
was found to have stimulatory effect, which is contrary
Feather as substrate for keratinase production Chapter 8
169
to the observations reported by Khardenavis et al (2009), Corrêa et al (2010) and Daroit
et al (2011).
Table 8.2 Effects of various metal ions, inhibitors, surfactants and bleach oxidant on B.
amyloliquefaciens 6B protease activity
Additives (concentration) Residual enzyme activity (%)
Control 100
Metal ions (5mM) Ca2+ 130.94
Cu2+ 34.52
Fe2+ 17.04
Hg2+ 105.82
Mg2+ 99.94
Mn2+ 195.96
Zn2+ 167.24
Inhibitors (5 mM) EDTA 106.27
PMSF 1.25
Surfactants (1%; w/v) Triton X- 100 110.12
Tween 80 91.53
SDS 82.35
CTAB 21.47
Oxidising agent (1%, w/v) H2O2 94.66
Among the inhibitors tested, PMSF completely inhibited the enzyme activity
(Table 8.2), suggesting it to be a serine hydrolase. Usually, EDTA has negative effects
on keratinase activity, but we observed that the keratinolytic protease from 6B strain
was slightly simulated by EDTA (5mM), similar to the keratinase from
Fervidobacterium islandicum AW-1 (Nam et al 2002) and B. sublitis KD-N2 (Cai et al
2008).
Furthermore, the enzyme remained quiet stable in presence of non-ionic (Triton
X-100 and Tween 80) and anionic surfactant (SDS), whereas CTAB (cationic
surfactant) exerted inhibitory effect (Table 8.2). Similar stability of B. halodurans
PPKS-2 keratinase in presence of non-ionic and anionic surfactant has been reported
(Prakash et al 2010). In general as per the literature, the proteases belonging to Bacillus
genus are unstable against the bleech-oxidants (Anwar and Saleemuddin 2000).
However, BA6B protease was found to be stable in presence of H2O2. Several
researchers have engineered proteases for making them surfactant and bleach-stable
(Tuschiya et al 1992, Wolff et al 1996), whereas the BA6B protease exhibited inherent
stability towards surfactants and bleach-oxidant, which makes it suitable for use in the
detergent industry.

Feather as substrate for keratinase production Chapter 8
170
8.1.6. In-vitro screening for antifungal activity
The genus Bacillus includes species considered particularly promising as
biocontrol agents against plant pathogens (Ongena and Jacques 2008). Thus, the
bacterium BA6B was investigated for its biocontrol potential.
Figure 8.9 Antifungal activity of Bacillus amyloliquefaciens 6B against (a) Aspergillus parasiticus
NCIM 898, (b) Aspergillus niger MTCC 16404, (c) Aspergillus fumigatus MTCC 343, (d) Fusarium
oxysporum NCIM 1072, (d) Trichosporon sp. NCIM 1110 and (e) Chrysosporium indicum.
Table 8.3: Antifungal activity of Bacillus amyloliquefaciens 6B culture supernatant against fungal
phytopathogens
Test organism Inhibition Zone (mm)
Control (uninoculated broth) -
Aspergilus niger MTCC 16404 14
Aspergilus parasiticus NCIM 898 19
Aspergilllus fumigates MTCC 343 12
Fusarium oxysporum NCIM 1072 21
Trichosporon sp. NCIM 1110 17
In dual culture assays, the bacterium BA6B was found to inhibit all the test fungi
(A. niger MTCC 16404, A. fumigatus MTCC 343, A. parasiticus NCIM 898,
Trichosporon sp. NCIM 1110, F. oxysporum NCIM 1072) on PDA plates at 25°C (Fig.
8.9). Interestingly, BA6B grown in feather meal medium also inhibited mycelial growth
of all the fungi tested (Table 8.3) with maximum antagonism against F. oxysporum
(1072).
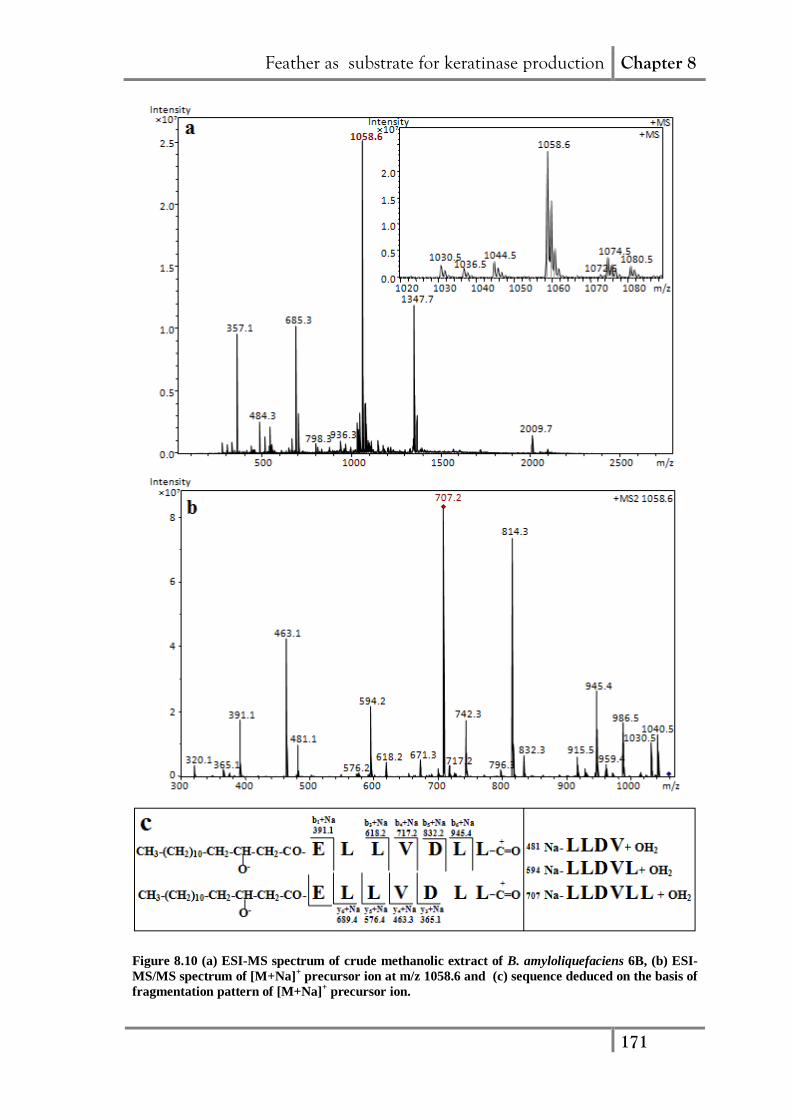
Feather as substrate for keratinase production Chapter 8
171
Figure 8.10 (a) ESI-MS spectrum of crude methanolic extract of B. amyloliquefaciens 6B, (b) ESI-
MS/MS spectrum of [M+Na]+ precursor ion at m/z 1058.6 and (c) sequence deduced on the basis of
fragmentation pattern of [M+Na]+ precursor ion.

Feather as substrate for keratinase production Chapter 8
172
Furthermore, to identify and characterize the bioactive compounds from BA6B,
the crude extract of the fermentation broth was subjected to ESI-mass spectrometry. The
peaks in ESI-MS spectra (Fig. 8.10a) at m/z, 1030.5, 1044.5, 1058.6 and 1072.6 differ in
mass by 14 Da and may be assigned as sodium adducts of C13-, C14-, C15- and C16-
fatty acid chain length variants of a same surfactin group, respectively (Vater et al.
2002). The peak at m/z 1058.6 was further subjected to MS2 analysis and on the basis of
fragment ions obtained, its sequence was deduced (Fig. 8.10b, c) The fragment ion at
m/z 1030.5 with a mass difference of 28 Da from the parent ion (m/z 1058.6) revealed
the carbonyl group coupled cleavage of lactone bond between fatty acid and C-terminal
end of peptide leading to the linearization of surfactin cyclic lipopeptide ring (Cavelier
et al 1999). The peak at m/z 707.2 in MS2 spectrum of m/z 1058.6 could be assigned as
sodiated form of internal fragment ion with molecular mass of 684 Da. The protonated
from of this fragment ion (m/z 685) has been reported to be ubiquitously detected in
MS2 spectra of surfactins [M+H]+ and is considered as characteristic marker ion for
identification of surfactins (Hue et al 2001). The sequence derived from the
fragmentation of m/z 1058.6 is: β-hydroxy fatty acid-Glu1-Leu
2-Leu
3-Val
4-Asp
5-Leu
6-
Leu7
(Fig. 8.10c). Similarly the peak at m/z, 1036.5 and 1075.4 could be assigned as
protonated [M+H]+ form and potassium adduct [M+K]
+of C15-surfactin with 1035 Da,
respectively. The m/z 1080.5 may be attributed to sodium adduct of C15-Iturin A
isoform (Pathak 2011).
Surfactin and iturin are the most commonly found antifungal lipopeptide
belonging to Bacillus genus (Ongena and Jacques 2008). Both the lipopeptides are
synthesized in a nonribosomal manner by large multienzyme complexes, exhibiting a
wide antimicrobial spectrum and exceptional surfactant activities (Ongena and Jacques
2008). They share a common cyclic structure consisting of either a β-amino (for iturin)
or β-hydroxy (for surfactin) fatty acid integrated into a peptide moiety. In addition,
surfactins show a strong synergistic action in combination with iturin (Maget-Dana et al
1992). It has also been reported that the lipopeptides also play a role to enhance
bacterial colonization on plant tissues and to induce plant resistance against pathogens
(Ongena and Jacques, 2008). Consequently, a great deal of interest has centered on the
use of such bioactive compounds in agriculture for eco-friendly and sustainable
suppression of crop diseases (Ongena and Jacques, 2008). Thus, the feather
Feather as substrate for keratinase production Chapter 8
173
hydrolysates resulting from the microbial conversion of feather keratin by BA6B may
represent a promising biocontrol agent against fungal phytopathogens.
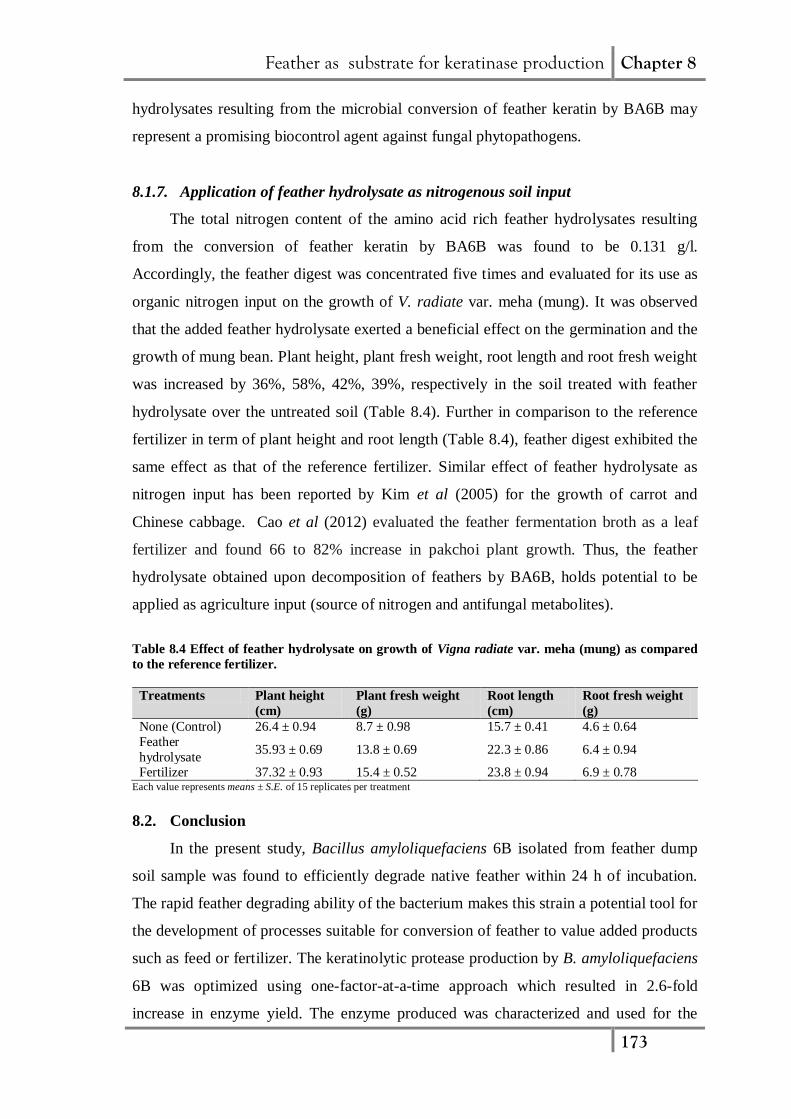
8.1.7. Application of feather hydrolysate as nitrogenous soil input
The total nitrogen content of the amino acid rich feather hydrolysates resulting
from the conversion of feather keratin by BA6B was found to be 0.131 g/l.
Accordingly, the feather digest was concentrated five times and evaluated for its use as
organic nitrogen input on the growth of V. radiate var. meha (mung). It was observed
that the added feather hydrolysate exerted a beneficial effect on the germination and the
growth of mung bean. Plant height, plant fresh weight, root length and root fresh weight
was increased by 36%, 58%, 42%, 39%, respectively in the soil treated with feather
hydrolysate over the untreated soil (Table 8.4). Further in comparison to the reference
fertilizer in term of plant height and root length (Table 8.4), feather digest exhibited the
same effect as that of the reference fertilizer. Similar effect of feather hydrolysate as
nitrogen input has been reported by Kim et al (2005) for the growth of carrot and
Chinese cabbage. Cao et al (2012) evaluated the feather fermentation broth as a leaf
fertilizer and found 66 to 82% increase in pakchoi plant growth. Thus, the feather
hydrolysate obtained upon decomposition of feathers by BA6B, holds potential to be
applied as agriculture input (source of nitrogen and antifungal metabolites).
Table 8.4 Effect of feather hydrolysate on growth of Vigna radiate var. meha (mung) as compared
to the reference fertilizer.
Treatments Plant height
(cm)
Plant fresh weight
(g)
Root length
(cm)
Root fresh weight
(g)
None (Control) 26.4 ± 0.94 8.7 ± 0.98 15.7 ± 0.41 4.6 ± 0.64
Feather
hydrolysate 35.93 ± 0.69 13.8 ± 0.69 22.3 ± 0.86 6.4 ± 0.94
Fertilizer 37.32 ± 0.93 15.4 ± 0.52 23.8 ± 0.94 6.9 ± 0.78 Each value represents means ± S.E. of 15 replicates per treatment
8.2. Conclusion
In the present study, Bacillus amyloliquefaciens 6B isolated from feather dump
soil sample was found to efficiently degrade native feather within 24 h of incubation.
The rapid feather degrading ability of the bacterium makes this strain a potential tool for
the development of processes suitable for conversion of feather to value added products
such as feed or fertilizer. The keratinolytic protease production by B. amyloliquefaciens
6B was optimized using one-factor-at-a-time approach which resulted in 2.6-fold
increase in enzyme yield. The enzyme produced was characterized and used for the

Feather as substrate for keratinase production Chapter 8
174
preparation of feather hydrolysate, which was evaluated as nitrogenous soil input for
plant growth promotion. In addition, the strain was found to produce antifungal
metabolites. Thus, the feather hydrolysate obtained upon degradation of feather by
BA6B bacterial culture, may be useful as nitrogenous input for agricultural soils as well
as biocontrol agent against fungal phytopathogens.

Feather as substrate for keratinase production Chapter 8
175
References
Allpress JD, Mountain G, Gowland PC (2002) Production, purification and
characterization of an extracellular keratinase from Lysobacter NCIMB 9497. Lett
Appl Microbiol 34: 337-342.
Anwar A, Saleemuddin M (2000) Alkaline protease from Spilosoma obliqua: potential
application in bioformulations. Biotechnol Appl Biochem 31: 85-89.
Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JA, Smith JG, Struhl DJ
(1997) Current Protocols in Molecular Biology Unit (24). New York: John Wiley
& Sons.
Balaji S, Senthil Kumar M, Karthikeyan R, Kumar R, Kirubanandan S, Sridhar R,
Sehgal PK (2008) Purification and characterization of an extracellular keratinase
from a hornmeal-degrading Bacillus subtilis MTCC 9102. World J Microbiol
Biotechnol 24: 2741-2745.
Bernal C, Cairo J, Coello N (2006) Purification and characterization of a novel
exocellular keratinase from Kocuria rosea. Enzyme Microb Technol 38: 49-54.
Brandelli A, Riffel A (2005) Production of an extracellular keratinase from
Chryseobacterium sp. growing on raw feathers. Electron J Biotechnol 8: 35-42.
Brandelli A (2008) Bacterial keratinases: Useful enzymes for bioprocessing
agroindustrial wastes and beyond. Food Bioprocess Technol 1: 105-116.
Cai CG, Zheng XD (2009) Medium optimization for keratinase production in hair
substrate by a new Bacillus subtilis KD-N2 using response surface methodology. J
Ind Microbiol Biotechnol 36: 875-883.
Cai CG, Lou BG, Zheng XD (2008) Purification and characterization of keratinase from
a new Bacillus subtilis strain. J Zhejiang Univ Sci B 9: 60-67.
Cao Z-J, Lu D, Luo L-S, Deng Y-X, Bian Y-G, Zhang X-Q, Zhou M-H (2012)
Composition analysis and application of degradation products of whole feathers
through a large scale of fermentation. Environ Sci Pollut Res 19: 2690-2696.
Cavelier F, Enjalbal JM, Roque M, Sanchez P, Aubagnac J-L (1999) Comparison of
collisionally dissociation mass spectra for the identification of cyclopeptides and
cyclodepsipeptides. Rapid Commun Mass Spectrom 13: 880-885.
Cheng SW, Hu HM, Shen SW, Takagi H, Asano M, Tsai YC (1995) Production and
characterization of keratinase of a feather-degrading Bacillus licheniformis PWD-
1. Biosci Biotechnol Biochem 59: 2239-2243.

Feather as substrate for keratinase production Chapter 8
176
Corrêa APF, Dariot DJ, Brandelli A (2010) Characterization of a keratinase produced
by Bacillus sp. P7 isolated from an Amazonian environment. Int Biodeterior
Biodegrad 64: 1-6.
Dalev P, Ivanov I, Liubomirova A (1997) Enzymatic modification of feather keratin
hydrolyzates with lysine aimed at increasing the biological value. J Sci Food
Agric 73: 242-244.
Daroit DJ, Corrêa APF, Brandelli A (2011) Production of keratinolytic proteases
through bioconversion of feather meal by the Amazonian bacterium Bacillus sp.
P45. Int Biodeterior Biodegrad 65: 45-51.
Deydier E, Guilet R, Sarda S, Sharrock P (2005) Physical and chemical characterization
of crude meat and bone meal combustion residue: waste or raw material? J Hazard
Mater 121: 141-148.
Fang Y, Liu S, Wang S, Lv M (2009) Isolation and screening of a novel extracellular
organic solvent-stable protease producer. Biochem Eng J 43: 212-215.
Fakhfakh N, Ktari N, Haddar A, Mnif IH, Dahmen I, Nasri M (2011) Total
solubilisation of the chicken feathers by fermentation with a keratinolytic
bacterium, Bacillus pumilus A1, and the production of protein hydrolysate with
high antioxidative activity. Process Biochem 46: 1731-1737.
FAO (2002) http://www.fao.org/docrep/004/AD452E/ad452e30.htm (retrieved on
28/05/2012)
Ghorbel B, Sellami-Kamoun A, Nasri M (2003) Stability studies of protease from
Bacillus cereus BG1. Enzyme Microb Technol 32: 513-518.
Glusker JP, Katz AK, Bock CW (1999) Metal ions in biological systems. The Rigaku J
16: 8-16.
Grazziotin A, Pimentel FA, de Jong EV, Brandelli A (2006) Nutritional improvement of
feather protein by treatment with microbial keratinase. Anim Feed Sci Technol
126: 135-144.
Gupta R, Ramnani P (2006) Microbial keratinase and their prospective application: an
overview. Appl Microbiol Biotechnol 70: 21-33.
Hossain MS, Azad AK, Abu Sayem SM, Mostafa G, Hoq MM (2007) Production and
partial characterization of feather-degrading keratinolytic serine protease from
Bacillus licheniformis MZK-3. J Biol Sci 7: 599-606.

Feather as substrate for keratinase production Chapter 8
177
Hue N, Serani L, Laprevote O (2001) Structural investigation of cyclic peptidolipids
from Bacillus subtilis by high energy tandem mass spectrometry. Rapid Commun
Mass Spectrom 15: 203-209.
Inoue A, Horikoshi K (1991) Estimation of solvent-tolerance of bacteria by the solvent
parameter logP. Journal Ferment Bioeng 71: 194-196.
Jeong J-H, Lee, O-M, Jeon Y-D, Kim J-D, Lee N-R, Lee C-Y, Son H-J (2010)
Production of keratinolytic enzyme by a newly isolated feather-degrading
Stenotrophomonas maltophilia that produces plant growth-promoting activity.
Process Biochem 45: 1738-1745.
Kaul S, Sumbali G (1999) Production of extracellular keratinases by keratinolytic
fungal species inhabiting feathers of living poultry birds (Gallus domesticus): a
comparison. Mycopathologia 146: 19-24.
Khardenavis AA, Kapley A, Purohit HJ (2009) Processing of poultry feathers by
alkaline keratin hydrolyzing enzyme from Serratia sp. HPC1383. Waste Manage
29: 1409-1415.
Kim JM, Choi YM, Suh HJ (2005) Preparation of feather digests as fertilizer with B.
pumilus KHS-1. J Microbiol Biotechnol 15: 472-476.
Kojima M, Kanai M, Tominaga M, Kitazume S, Inoue A, Horikoshi K (2006) Isolation
and characterization of a feather degrading enzyme from Bacillus pseudofirmus
FA30-01. Extremophiles 10: 229-235.
Kunert J (2000) Physiology of keratinophylic fungi. In: Kushwaha RKS, Guarro J
(Eds.) Biology of dermatophytes and other keratinophylic fungi. Bilbao: Rev
Iberoamericana de Micología, pp. 77-85.
Laemmli UK (1970) Cleavage of structural proteins during the assembly of the head of
bacteriophage T4. Nature 227: 680-685.
Lateef A, Oloke JK, Gueguim Kana EB, Sobowale BO, Ajao SO, Bello BY (2010)
Keratinolytic activities of a new feather-degrading isolate of Bacillus cereus LAU
08 isolated from Nigerian soil. Int Biodeterior Biodegrad 64: 162-165.
Li S, He B, Bai Z, Ouyang P (2009) A novel organic solvent-stable alkaline protease
from organic solvent-tolerant Bacillus licheniformis YP1A. J Mol Catal B 56: 85-
88.

Feather as substrate for keratinase production Chapter 8
178
Lin X, Lee CG, Casale ES, Shih JCH (1992) Purification and characterization of a
keratinase from a feather degrading Bacillus licheniformis strain. Appl Environ
Microbiol 58: 3271-3275.
Lowry OH, Rosebrough NJ, Farr AL, Randall RJ (1951) Protein measurement with the
Folin phenol reagent. The J Biol Chem 193: 265-275.
Maget-Dana R, Thimon L, Peypoux F, Ptak M (1992) Surfactin/iturin A interactions
may explain the synergistic effect of surfactin on the biological properties of iturin
A. Biochimie 74: 1047-1051.
Nam GW, Lee DW, Lee HS, Lee NJ, Kim BC, Choe EA, Hwang JK, Suhartono MT,
Pyun YR (2002) Native-feather degradation by Fervidobacterium islandicum
AW-1, a newly isolated keratinase-producing thermophilic anaerobe. Arch
Microbiol 178: 538-547.
Odetallah NH,Wang JJ, Garlich JD, Shih JCH (2003) Keratinase in starter diets
improves growth of broiler chicks. Poult Sci 82: 664-670.
Ongena M, Jacques P (2008) Bacillus lipopeptides: versatile weapons for plant disease
biocontrol. Trends Microbiol 16: 115-125.
Onifade AA, Al-Sane NA, Al-Musallam AA, Al-Zarban S (1998) A review: potentials
for biotechnological applications of keratin-degrading microorganisms and their
enzymes for nutritional improvement of feathers and other keratins as livestock
feed resources. Biores Technol 66: 1-11.
Park GT, Son HJ (2009) Keratinolytic activity of Bacillus megaterium F7-1, a feather
degrading mesophilic bacterium. Microbiol Res 164: 478-485.
Pathak KV (2011) Purification and characterization of antifungal compounds produced
by Banyan endophytic Bacilli. Ph. D. Thesis, Sardar Patel University, Vallabh
Vidynagar, India.
Prakash P, Jayalakshmi SK, Sreeramulu K (2010) Production of keratinases by free and
immobilized cells of Bacillus halodurans strain PPKS-2: partial charcaterization
and its application in feather degradation and dehairing of the goat skin. Appl
Biochem Biotechnol 160: 1909-1920.
Raaijmakers JM, Vlami M, Souza JT (2002) Antibiotic production by bacterial
biocontrol agents. Antonie Leeuwenhoek 81: 537-547.

Feather as substrate for keratinase production Chapter 8
179
Ramnani P, Gupta G (2004) Optimization of medium composition for keratinase
production on feather by Bacillus licheniformis RG1 using statistical methods
involving response surface methodology. Biotechnol Appl Biochem 40: 191-196.
Riffel A, Lucas FS, Heeb P, Brandelli A (2003) Characterization of a new keratinolytic
bacterium that completely degrades native feather keratin. Arch Microbiol 179:
258-265.
Sambrook J, Russell DW (2001) Molecular Cloning: A Laboratory Manual (3rd ed),
New York: Cold Spring Harbor Laboratory Press.
Sareen R, Mishra P (2008) Purification and characterization of organic solvent stable
protease from Bacillus licheniformis RSP-09-37. Appl Microbiol Biotechnol 79:
399-405.
Sikkema J, de Bont JAM, Poolman B (1995) Mechanisms of membrane toxicity of
hydrocarbons. Microbiol Rev 59: 201-222.
Torres S, Pandey A, Castro GR (2011) Organic solvent adaptation of Gram positive
bacteria: applications and biotechnological potentials. Biotechnol Adv 29: 442-52.
Trivedi P, Pandy A, Palni LM (2008) In vitro evaluation of antagonistic properties of
Pseudomonas corrugata. Microbiol Res 163: 329-336.
Tuschiya K, Nakamura Y, Sakashita H, Kimura T (1992) Purification and
characterization of a thermostable alkaline protease from alkalophilic
Thermoactinomyces sp. HS682. Biosci Biotechnol Biochem 56: 246-250.
Vasileva-Tonkova E, Gousterova A, Neshev G (2009) Ecologically safe method for
improved feather wastes biodegradation. Int Biodeterior Biodegrad 63: 1008-
1012.
Vater J, Kablitz B, Wilde C, Franke P, Mehta N, Cameotra SS (2002) Matrix-assisted
Laser Desorption Ionization-Time of Flight mass spectrometry of lipopeptide
biosurfactants in whole cells and culture filtrates of Bacillus subtilis C-1 isolated
from petroleum sludge. Appl Environ Microbiol 68: 6210-6219.
Wang JJ, Shih JCH (1999) Fermentation production of keratinase from Bacillus
licheniformis PWD-1 and a recombinant B. subtilis FDB-29. J Ind Microbiol
Biotechnol 2: 608-616.
Wolff AM, Showell MS, Venegas MG, Barnett BL, Wertz WC (1996) Laundry
performance of subtilisin proteases. In: Bott R, Betzel C (Eds.) Subtilisin
enzymes: practical protein engineering. New York: Plenum Press, pp. 113-120.

Feather as substrate for keratinase production Chapter 8
180
Xie F, Chao Y, Yang X, Yang J, Xue Z, Luo Y, Qian S (2010) Purification and
characterization of four keratinases produced by Streptomyces sp. strain 16 in
native human foot skin medium. Biores Technol 101: 344-350.





























