Embed Size (px)
Citation preview

ResearchPap
er
New Biotechnology �Volume 28, Number 6 �October 2011 RESEARCH PAPER
Characterization of a fungistaticsubstance produced by Aspergillus flavusisolated from soil and its significance innatureYen-Ting Chen, Mei-Ju Lin, Ching-Hui Yang and Wen-Hsiung Ko
Department of Plant Pathology, National Chung Hsing University, Taichung, Taiwan
A fungus capable of using vegetable tissues for multiplication in soil was isolated and identified as
Aspergillus flavus based on morphological characteristics and sequence similarity of ITS and 28S. When
grown in liquid medium prepared from the same vegetable tissues used in soil amendment, the isolate of
A. flavus produced a substance capable of preventing disease development of black leaf spot of mustard
cabbage caused by Alternaria brassicicola and inhibiting the germination of A. brassicicola conidia. The
inhibitory substance was fungistatic, and was very stable under high temperature and high or low pH
value. It was soluble in ethanol or methanol, moderately soluble in water, and insoluble in acetone, ethyl
acetate or ether. The inhibitor is not a protein and has no charges on its molecule. This is the first
discovery of the production of a fungistatic substance by this deleterious fungus. Results from this study
suggest the possession of a strong competitive saprophytic ability by A. flavus, which in turn may
explain the widespread occurrence of this fungus in soils. Production of a fungistatic substance when A.
flavus was grown in medium prepared from vegetable tissues suggests the importance of antibiotic
production in its competitive saprophytic colonization of organic matters in soils.
IntroductionAspergillus flavus is an important deleterious fungus due to its
production of aflatoxins especially aflatoxin B1 which is the most
toxic and most potent naturally occurring carcinogen character-
ized [1,2]. It is also a common cause of invasive and non-invasive
aspergillosis in human and animals, and an opportunistic patho-
gen of many agricultural crops such as corn, cotton, peanuts as
well as tree nuts [3–5]. The fungus is found throughout the world
in soils including agricultural, forest, orchard, grassland and wet-
land soils [6,7]. It was also isolated from virgin as well as cultivated
desert soils [8–10]. The widespread occurrence of A. flavus in soil
has been associated with its ability to colonize plant debris in soil
[11–13]. However, little is known about the competitive sapro-
phytic ability [14] of this fungus in soils.
During our recent research, microorganisms capable of utilizing
vegetable tissues in soil for multiplication were selectively isolated.
Broth prepared from the same vegetable tissues was used to culti-
Corresponding author: Ko, W.-H. ([email protected])
1871-6784/$ - see front matter � 2011 Elsevier B.V. All rights reserved. doi:10.1016/j.nbt.2011.02.005
vate these organisms. Triturated cultures were screened for ability
to control black leaf spot of mustard cabbage caused by Alternaria
brassicicola. The culture of a fungus, which was subsequently
identified as A. flavus, was found to be very effective in controlling
the disease and its extract was strongly inhibitory to the pathogen.
This is the first discovery of the production of a fungistatic sub-
stance by this important fungus.
In this report, we describe the isolation and identification of A.
flavus from soil, the production and characterization of the fungi-
static substance produced by this fungus and the significance of
the discovery in the explanation of its strong competitive sapro-
phytic ability in soil.
Materials and methodsIsolation of soil microorganismsSoil samples collected from six locations in central Taiwan were
taken from a depth of 0–10 cm, sifted and moistened to about 65%
water-holding capacity. Media for selective isolation of fungi,
bacteria and actinomycetes were prepared as previously described
www.elsevier.com/locate/nbt 679

RESEARCH PAPER New Biotechnology �Volume 28, Number 6 �October 2011
Research
Pap
er
[15]. Approximately 1.3 g soil was mixed with 100 ml sterile dis-
tilled water in an Omni mixer chamber at 5000 rpm for 30 s. The
suspension was diluted to 10�4, 10�5 and 10�6 for fungi, 10�5, 10�6
and 10�7 for actinomycetes and 10�6, 10�7 and 10�8 for bacteria to
determine the dilution needed to obtain soil suspension essen-
tially free of each group of microorganisms. A 1-ml aliquot of
diluted soil suspension was mixed with 20 ml of a molten selective
medium at 458C in a Petri plate. Five plates were used for each
treatment.
Vegetables including fruit of tomato (Lycopersicon esculentum),
tubers of sweet potato (Ipomoea batatas), and leaves and stems of
spinach (Spinacia oleracea), ong choy (Ipomoea aquatica) and com-
mon purslane (Portulaca oleracea) were purchased from local mar-
ket and chopped into small pieces. For soil amendment, about
500 g soil was mixed with 4% of each chopped vegetable in a 1000-
ml bottle and incubated at 248C for at least two weeks before use.
To isolate microorganisms with ability to utilize amended nutri-
ents for multiplication, suspension of amended soil was diluted to
the concentration pre-determined for each group of microorgan-
isms and plated on the selective medium as described above. After
incubation at 248C for seven days, colonies appeared were indi-
vidually transferred to 10% V-8 agar (10% V-8 juice, 0.02% CaCO3
and 2% agar) plates.
Culture of isolated microorganisms in liquid mediumLiquid medium was prepared by triturating 4 g each of the five
chopped vegetables in 100 ml water in an Omni mixer at 4000 rpm
for 3 min and dispensing 50 ml broth in a 250-ml flask. After
autoclaving, each flask was inoculated with two loopfuls of bac-
terium, or a piece (ca. 4 � 5 � 3 mm) of actinomycete or fungus
agar culture. Inoculated flasks were incubated on a shaker for two
weeks. After incubation, cultures were separately ground in an
Omni mixer at 4000 rpm for 1 min before being used to test their
ability to reduce disease incidence.
Inoculum preparationA. brassicicola (Schw.) Wiltshire (isolate Aba-31) was grown on 10%
V-8 agar at 248C under light for four to six days for the production
of conidia. A conidial suspension was prepared by placing two
pieces of culture blocks (ca. 5 � 5 � 3 mm) in 5 ml sterile distilled
water in a test tube and by agitating the test tube for 30 s with a
Vortex mixer. The concentration of conidia was adjusted to 3
conidia ml�1 with a Pipetman microliter pipette (West Coast Scien-
tific, Oakland, CA) [16].
Disease control assay of liquid culturesSeeds of mustard cabbage [Brassica juncea (L.) Coss.] were grown in
8-cm pots containing a mixture of peat moss and vermiculite (9:1,
v/v). Two leaves of a four-week-old plant were sprayed to run off
with liquid culture of a test microorganism daily three times before
inoculation on the fourth day.
Each mustard cabbage leaf was inoculated with five 2-ml drops of
conidial suspension of A. brassicicola along the edge of the leaf, and
a 10-ml drop of molten agar consisting of 1% agar and 1% V-8 juice
at 608C was added to each inoculum drop to fix the inoculum on
the target site [17]. Leaves of mustard cabbage sprayed with liquid
medium were inoculated with conidia of A. brassicicola and used as
controls. Inoculated plants were placed in moist chambers and
680 www.elsevier.com/locate/nbt
kept in the greenhouse. The number and the size of lesions that
developed at the inoculated sites were recorded three days after
inoculation. Two leaves were used for each treatment, and all the
experiments were repeated at least twice.
DNA extractionThe DNA of A. flavus isolate V5F-13 was extracted from 0.1 g three-
day-old mycelia grown on cellophane placed on PDA [18] by the
plant genomic DNA extraction kit (GeneMark Technology Co.,
Taichung, Taiwan). The ITS region was amplified with primers ITS1
and ITS4 [19]. PCR was performed in a 50 ml volume reaction
containing 2 ml DNA, 1 pmol of upstream and downstream pri-
mers and 2.5 units of SuperTaq polymerase (Protech Technology
Enterprise Co., Ltd, Taipei, Taiwan) with buffer system recom-
mended by the manufacturer. Cycling conditions of PCR were:
initial denaturation at 948C for 2 min, 30 cycles at 948C for 30 s,
558C for 30 s, 728C for 1 min, and a final elongation at 728C for
6 min. The PCR product was analyzed by electrophoresis in a 1.2%
agarose gel. In the same manner the large ribosomal subunit 28S
was analyzed with primer pairs LROR and LR7 [20]. The annealing
temperature was changed to 508C.
BioassayA 10-ml aliquot of conidial suspension (2 � 104 spores ml�1) of A.
brassicicola was mixed with 10 ml of culture or extract in a cavity of
a sterile eight-cavity slide. Slides with spores were kept moist by
placing each on an L-shaped glass rod in a 9-cm Petri plate
containing 10 ml sterile distilled water. Germination was recorded
after incubation at 288C for four hours, and 100 spores were
counted in each of three replicates.
Extraction of inhibitory substanceEach flask of liquid culture of A. flavus was tested for the ability to
inhibit spore germination of A. brassicicola to assure the presence
of the inhibitory substance before freeze drying. One gram of dry
powder, obtained from approximately 50 ml of culture, was
extracted with 50 ml of water, ethanol, methanol, acetone, ethyl
acetate or ether in a 250-ml flask by shaking on a shaker for 24
hours. The mixture was centrifuged at 1500 � g for 5 min to obtain
clear extract. For bioassay and characterization of the inhibitory
substance, 10 ml extract was evaporated to 2 ml followed by the
addition of 2 ml water and evaporation to 2 ml again. For control,
water was similarly extracted with each solvent and tested for the
ability to inhibit spore germination.
Characterization of the inhibitorTo study the ability of different adsorptive materials to remove the
inhibitor from the culture extract of A. flavus, 5 g of Diaion SK1B
cation exchange resins (equivalent to Amberlite 1R-120), Diaion
SA 12A anion exchange resins (equivalent to Amberlite 1RA-420;
Tai-Young Chemical Co., Kaohsiung, Taiwan) or activated char-
coal (Sigma–Aldrich) was washed with 50 ml of distilled water
three times by shaking over a six-hour period to remove possible
inhibitory substances [21]. Ten milliliter extract was shaken with
1 g cation exchange resins, anion exchange resins or activated
charcoal in a 150-ml flask at 100 strokes min�1 for 24 hours and
filtered through a Whatman no. 1 filter paper. The filtrates were
then used for germination tests.
New Biotechnology �Volume 28, Number 6 �October 2011 RESEARCH PAPER
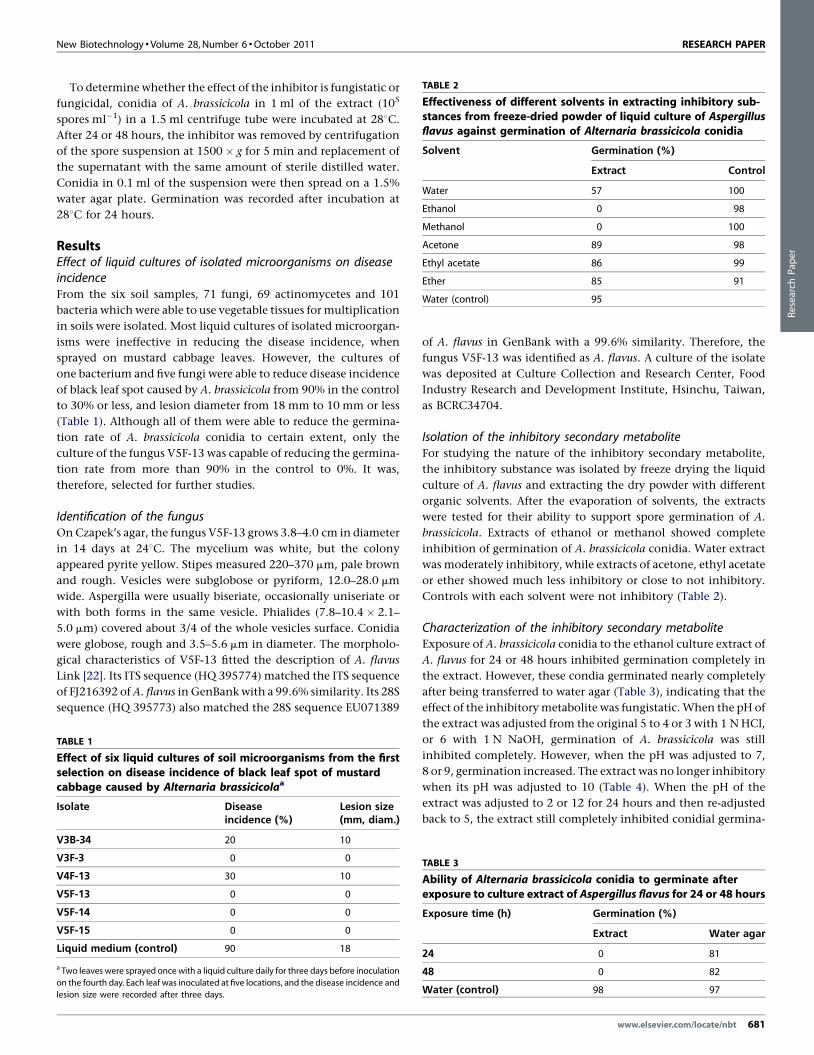
TABLE 2
Effectiveness of different solvents in extracting inhibitory sub-stances from freeze-dried powder of liquid culture of Aspergillusflavus against germination of Alternaria brassicicola conidia
Solvent Germination (%)
Extract Control
Water 57 100
Ethanol 0 98
Methanol 0 100
Acetone 89 98
Ethyl acetate 86 99
Ether 85 91
Water (control) 95
ResearchPap
er
To determine whether the effect of the inhibitor is fungistatic or
fungicidal, conidia of A. brassicicola in 1 ml of the extract (105
spores ml�1) in a 1.5 ml centrifuge tube were incubated at 288C.
After 24 or 48 hours, the inhibitor was removed by centrifugation
of the spore suspension at 1500 � g for 5 min and replacement of
the supernatant with the same amount of sterile distilled water.
Conidia in 0.1 ml of the suspension were then spread on a 1.5%
water agar plate. Germination was recorded after incubation at
288C for 24 hours.
ResultsEffect of liquid cultures of isolated microorganisms on diseaseincidenceFrom the six soil samples, 71 fungi, 69 actinomycetes and 101
bacteria which were able to use vegetable tissues for multiplication
in soils were isolated. Most liquid cultures of isolated microorgan-
isms were ineffective in reducing the disease incidence, when
sprayed on mustard cabbage leaves. However, the cultures of
one bacterium and five fungi were able to reduce disease incidence
of black leaf spot caused by A. brassicicola from 90% in the control
to 30% or less, and lesion diameter from 18 mm to 10 mm or less
(Table 1). Although all of them were able to reduce the germina-
tion rate of A. brassicicola conidia to certain extent, only the
culture of the fungus V5F-13 was capable of reducing the germina-
tion rate from more than 90% in the control to 0%. It was,
therefore, selected for further studies.
Identification of the fungusOn Czapek’s agar, the fungus V5F-13 grows 3.8–4.0 cm in diameter
in 14 days at 248C. The mycelium was white, but the colony
appeared pyrite yellow. Stipes measured 220–370 mm, pale brown
and rough. Vesicles were subglobose or pyriform, 12.0–28.0 mm
wide. Aspergilla were usually biseriate, occasionally uniseriate or
with both forms in the same vesicle. Phialides (7.8–10.4 � 2.1–
5.0 mm) covered about 3/4 of the whole vesicles surface. Conidia
were globose, rough and 3.5–5.6 mm in diameter. The morpholo-
gical characteristics of V5F-13 fitted the description of A. flavus
Link [22]. Its ITS sequence (HQ 395774) matched the ITS sequence
of FJ216392 of A. flavus in GenBank with a 99.6% similarity. Its 28S
sequence (HQ 395773) also matched the 28S sequence EU071389
TABLE 1
Effect of six liquid cultures of soil microorganisms from the firstselection on disease incidence of black leaf spot of mustardcabbage caused by Alternaria brassicicolaa
Isolate Diseaseincidence (%)
Lesion size(mm, diam.)
V3B-34 20 10
V3F-3 0 0
V4F-13 30 10
V5F-13 0 0
V5F-14 0 0
V5F-15 0 0
Liquid medium (control) 90 18
a Two leaves were sprayed once with a liquid culture daily for three days before inoculation
on the fourth day. Each leaf was inoculated at five locations, and the disease incidence and
lesion size were recorded after three days.
of A. flavus in GenBank with a 99.6% similarity. Therefore, the
fungus V5F-13 was identified as A. flavus. A culture of the isolate
was deposited at Culture Collection and Research Center, Food
Industry Research and Development Institute, Hsinchu, Taiwan,
as BCRC34704.
Isolation of the inhibitory secondary metaboliteFor studying the nature of the inhibitory secondary metabolite,
the inhibitory substance was isolated by freeze drying the liquid
culture of A. flavus and extracting the dry powder with different
organic solvents. After the evaporation of solvents, the extracts
were tested for their ability to support spore germination of A.
brassicicola. Extracts of ethanol or methanol showed complete
inhibition of germination of A. brassicicola conidia. Water extract
was moderately inhibitory, while extracts of acetone, ethyl acetate
or ether showed much less inhibitory or close to not inhibitory.
Controls with each solvent were not inhibitory (Table 2).
Characterization of the inhibitory secondary metaboliteExposure of A. brassicicola conidia to the ethanol culture extract of
A. flavus for 24 or 48 hours inhibited germination completely in
the extract. However, these condia germinated nearly completely
after being transferred to water agar (Table 3), indicating that the
effect of the inhibitory metabolite was fungistatic. When the pH of
the extract was adjusted from the original 5 to 4 or 3 with 1 N HCl,
or 6 with 1 N NaOH, germination of A. brassicicola was still
inhibited completely. However, when the pH was adjusted to 7,
8 or 9, germination increased. The extract was no longer inhibitory
when its pH was adjusted to 10 (Table 4). When the pH of the
extract was adjusted to 2 or 12 for 24 hours and then re-adjusted
back to 5, the extract still completely inhibited conidial germina-
TABLE 3
Ability of Alternaria brassicicola conidia to germinate afterexposure to culture extract of Aspergillus flavus for 24 or 48 hours
Exposure time (h) Germination (%)
Extract Water agar
24 0 81
48 0 82
Water (control) 98 97
www.elsevier.com/locate/nbt 681
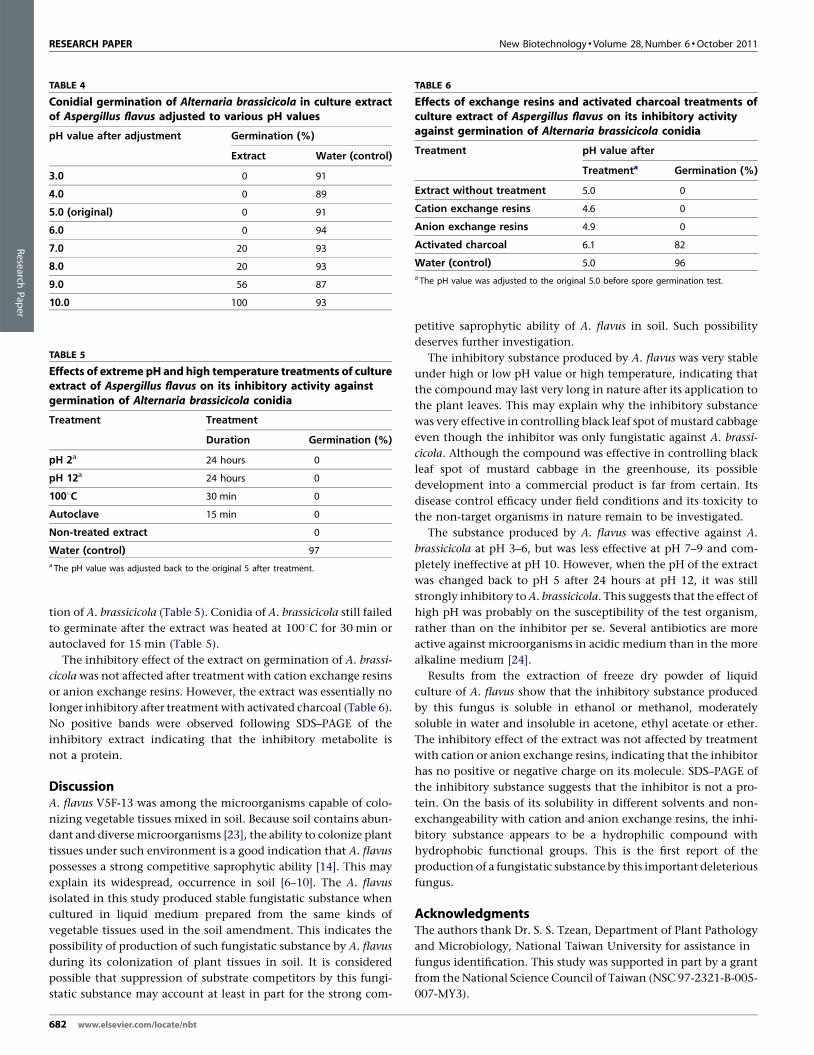
RESEARCH PAPER New Biotechnology �Volume 28, Number 6 �October 2011
TABLE 6
Effects of exchange resins and activated charcoal treatments ofculture extract of Aspergillus flavus on its inhibitory activityagainst germination of Alternaria brassicicola conidia
Treatment pH value after
Treatmenta Germination (%)
Extract without treatment 5.0 0
Cation exchange resins 4.6 0
Anion exchange resins 4.9 0
Activated charcoal 6.1 82
Water (control) 5.0 96a The pH value was adjusted to the original 5.0 before spore germination test.
TABLE 5
Effects of extreme pH and high temperature treatments of cultureextract of Aspergillus flavus on its inhibitory activity againstgermination of Alternaria brassicicola conidia
Treatment Treatment
Duration Germination (%)
pH 2a 24 hours 0
pH 12a 24 hours 0
1008C 30 min 0
Autoclave 15 min 0
Non-treated extract 0
Water (control) 97a The pH value was adjusted back to the original 5 after treatment.
TABLE 4
Conidial germination of Alternaria brassicicola in culture extractof Aspergillus flavus adjusted to various pH values
pH value after adjustment Germination (%)
Extract Water (control)
3.0 0 91
4.0 0 89
5.0 (original) 0 91
6.0 0 94
7.0 20 93
8.0 20 93
9.0 56 87
10.0 100 93
Research
Pap
er
tion of A. brassicicola (Table 5). Conidia of A. brassicicola still failed
to germinate after the extract was heated at 1008C for 30 min or
autoclaved for 15 min (Table 5).
The inhibitory effect of the extract on germination of A. brassi-
cicola was not affected after treatment with cation exchange resins
or anion exchange resins. However, the extract was essentially no
longer inhibitory after treatment with activated charcoal (Table 6).
No positive bands were observed following SDS–PAGE of the
inhibitory extract indicating that the inhibitory metabolite is
not a protein.
DiscussionA. flavus V5F-13 was among the microorganisms capable of colo-
nizing vegetable tissues mixed in soil. Because soil contains abun-
dant and diverse microorganisms [23], the ability to colonize plant
tissues under such environment is a good indication that A. flavus
possesses a strong competitive saprophytic ability [14]. This may
explain its widespread, occurrence in soil [6–10]. The A. flavus
isolated in this study produced stable fungistatic substance when
cultured in liquid medium prepared from the same kinds of
vegetable tissues used in the soil amendment. This indicates the
possibility of production of such fungistatic substance by A. flavus
during its colonization of plant tissues in soil. It is considered
possible that suppression of substrate competitors by this fungi-
static substance may account at least in part for the strong com-
682 www.elsevier.com/locate/nbt
petitive saprophytic ability of A. flavus in soil. Such possibility
deserves further investigation.
The inhibitory substance produced by A. flavus was very stable
under high or low pH value or high temperature, indicating that
the compound may last very long in nature after its application to
the plant leaves. This may explain why the inhibitory substance
was very effective in controlling black leaf spot of mustard cabbage
even though the inhibitor was only fungistatic against A. brassi-
cicola. Although the compound was effective in controlling black
leaf spot of mustard cabbage in the greenhouse, its possible
development into a commercial product is far from certain. Its
disease control efficacy under field conditions and its toxicity to
the non-target organisms in nature remain to be investigated.
The substance produced by A. flavus was effective against A.
brassicicola at pH 3–6, but was less effective at pH 7–9 and com-
pletely ineffective at pH 10. However, when the pH of the extract
was changed back to pH 5 after 24 hours at pH 12, it was still
strongly inhibitory to A. brassicicola. This suggests that the effect of
high pH was probably on the susceptibility of the test organism,
rather than on the inhibitor per se. Several antibiotics are more
active against microorganisms in acidic medium than in the more
alkaline medium [24].
Results from the extraction of freeze dry powder of liquid
culture of A. flavus show that the inhibitory substance produced
by this fungus is soluble in ethanol or methanol, moderately
soluble in water and insoluble in acetone, ethyl acetate or ether.
The inhibitory effect of the extract was not affected by treatment
with cation or anion exchange resins, indicating that the inhibitor
has no positive or negative charge on its molecule. SDS–PAGE of
the inhibitory substance suggests that the inhibitor is not a pro-
tein. On the basis of its solubility in different solvents and non-
exchangeability with cation and anion exchange resins, the inhi-
bitory substance appears to be a hydrophilic compound with
hydrophobic functional groups. This is the first report of the
production of a fungistatic substance by this important deleterious
fungus.
AcknowledgmentsThe authors thank Dr. S. S. Tzean, Department of Plant Pathology
and Microbiology, National Taiwan University for assistance in
fungus identification. This study was supported in part by a grant
from the National Science Council of Taiwan (NSC 97-2321-B-005-
007-MY3).

New Biotechnology �Volume 28, Number 6 �October 2011 RESEARCH PAPER
ResearchPap
er
References
1 Squire, R.A. (1981) Ranking animal carcinogens: a proposed regulatory approach.
Science 214, 877–880
2 Bennett, J.W. and Klich, M.A. (2003) Mycotoxins. Clin. Microbiol. Rev. 16, 497–516
3 Scheidegger, K.A. and Payne, G.A. (2003) Unlocking the secrets behind secondary
metabolism: a review of Aspergillus flavus from pathogenicity to functional
genomics. J. Toxicol-Toxin Rev. 22, 423–459
4 Yu, J. et al. (2005) Aspergillus flavus genomics: gateway to human and animal
health, food safety, and crop resistance to diseases. Rev. Iberoam. Mycol. 22, 194–
202
5 Hedayati, M.T. et al. (2007) Aspergillus flavus: human pathogen, allergen and
mycotoxin producer. Microbiology 153, 1677–1692
6 Klich, M.A. (2002) Biogeography of Aspergillus species in soil and litter. Mycologia
94, 21–27
7 Horn, B.W. (2003) Ecology and population biology of aflatoxigenic fungi in soil. J.
Toxicol-Toxin Rev. 22, 351–379
8 Ranzoni, F.V. (1968) Fungi isolated in culture from soils of the Sonoran Desert.
Mycologia 60, 356–371
9 Klich, M.A. (1998) Soil fungi of some low-altitude desert cotton fields and ability of
their extracts to inhibit Aspergillus flavus. Mycopathologia 142, 97–100
10 Abdel-Azeem, A.M. and Ibrahim, M.E. (2004) Diversity of terrophilous mycobiota
of Sinai. Egypt J. Biol. 6, 21–31
11 Griffin, G.J. and Garren, K.H. (1976) Colonization of rye green manure and peanut
fruit debris by Aspergillus flavus and Aspergillus niger group in field soils. Appl.
Environ. Microbiol. 32, 28–32
12 Abbas, H.K. et al. (2004) Spatial variability of Aspergillus flavus soil populations
under different crops and corn grain colonization and aflatoxins. Can. J. Bot. 82,
1768–1775
13 Abbas, H.K. et al. (2009) Ecology of Aspergillus flavus, regulation of aflatoxin
production, and management strategies to reduce aflatoxin contamination of
corn. Toxin Rev. 28, 142–153
14 Garrett, S.D. (1970) Pathogenic Root-Infecting Fungi. Cambridge University Press
15 Ko, W.H. et al. (2010) Activity and characterization of secondary metabolites
produced by a new microorganism for control of plant diseases. New Biotechnol. 27,
397–402
16 Ann, P.J. et al. (2010) Reevaluation of the report of the A2 mating type of
Phytophthora infestans on tomato in Taiwan. Bot. Stud. 51, 203–207
17 Xu, X.L. and Ko, W.H. (1998) A quantitative confined inoculation method for
studies of pathogenicity of fungi on plant. Bot. Bull. Acad. Sin. 39, 187–190
18 Wang, P.H. et al. (2010) Severe decline of wax apple trees caused by Fusarium solani
in northern Taiwan. Bot. Stud. 51, 75–80
19 White, T.J. et al. (1990) Amplification and direct sequencing of fungal ribosomal
RNA genes for phylogenetics. In PCR Protocols, a Guide to Methods and Applications
(Innis, M.A. et al. eds), pp. 315–322, Academic Press
20 Vilgalys, R. and Hester, M. (1990) Rapid genetic identification and mapping of
enzymatically amplified ribosomal DNA from several Cryptococcus species.
Bacteriology 172, 4238–4248
21 Ho, W.C. et al. (2007) Effect of oriental medicinal plant extracts on spore
germination of Alternaria brassicicola and nature of inhibitory substance from
speedweed. Plant Dis. 91, 1621–1624
22 Tzean, S.S. et al. (2005) Fungal Flora of Taiwan, vol. 4. Basidiomycota and Anamorphic
Fungi. Taiwan National Science Council
23 Alexander, M. (1977) Introduction to Soil Microbiology (2nd ed.), John Wiley
24 Poala, P. et al. (1970) Effect of pH of medium and size of inoculum on activity of
antibiotics against group D Streptococcus (Enterococcus). Appl. Microbiol. 19, 623–637
www.elsevier.com/locate/nbt 683





























