Embed Size (px)
Citation preview

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Feb. 2007, p. 1296–1307 Vol. 73, No. 40099-2240/07/$08.00�0 doi:10.1128/AEM.01888-06Copyright © 2007, American Society for Microbiology. All Rights Reserved.
Characterization of a Large, Stable, High-Copy-Number StreptomycesPlasmid That Requires Stability and Transfer Functions for
Heterologous Polyketide Overproduction�
Ryan Fong,1* Zhihao Hu,2 C. Richard Hutchinson,2 Jianqiang Huang,3Stanley Cohen,3 and Camilla Kao1
Department of Chemical Engineering, Stanford University, Stanford, California 943051; Kosan Biosciences, Inc., Hayward,California 945452; and Department of Genetics, Stanford University, Stanford, California 943053
Received 8 August 2006/Accepted 24 November 2006
A major limitation to improving small-molecule pharmaceutical production in streptomycetes is the inabilityof high-copy-number plasmids to tolerate large biosynthetic gene cluster inserts. A recent finding has overcomethis barrier. In 2003, Hu et al. discovered a stable, high-copy-number, 81-kb plasmid that significantly elevatedproduction of the polyketide precursor to the antibiotic erythromycin in a heterologous Streptomyces host(J. Ind. Microbiol. Biotechnol. 30:516–522, 2003). Here, we have identified mechanisms by which this SCP2*-derived plasmid achieves increased levels of metabolite production and examined how the 45-bp deletionmutation in the plasmid replication origin increased plasmid copy number. A plasmid intramycelial transfergene, spd, and a partition gene, parAB, enhance metabolite production by increasing the stable inheritance oflarge plasmids containing biosynthetic genes. Additionally, high product titers required both activator (actII-ORF4) and biosynthetic genes (eryA) at high copy numbers. DNA gel shift experiments revealed that the 45-bpdeletion abolished replication protein (RepI) binding to a plasmid site which, in part, supports an iteron modelfor plasmid replication and copy number control. Using the new information, we constructed a large high-copy-number plasmid capable of overproducing the polyketide 6-deoxyerythronolide B. However, this plasmidwas unstable over multiple culture generations, suggesting that other SCP2* genes may be required forlong-term, stable plasmid inheritance.
Streptomycetes produce many complex small molecules withtherapeutic value. Streptomyces coelicolor CH999 effectivelyexpresses heterologous gene clusters and hybrid genes for newnatural products (38, 39). The vectors usually consist of theSCP2*-based plasmid pRM5 and its derivatives, carrying thedivergent actI/actIII promoter pair from the endogenous acti-norhodin biosynthetic gene cluster (41). These native promot-ers are activated by the pathway-specific actII-ORF4 geneproduct (1, 11, 13). Low-copy-number vectors derived from the31.3-kb SCP2* (5) can stably replicate large DNA inserts (�30kb) through multiple culture generations (35). However, likewild-type streptomycete strains, recombinants carrying suchvectors typically synthesize natural products in small quantities(�1 mg per liter of fermentation medium). When these re-combinants are engineered to make “unnatural natural prod-ucts” (39), the titers are typically reduced even further, pre-senting a continuing challenge to improve productivity for drugdevelopment.
The use of a multicopy plasmid vector is one method ofoverexpressing biosynthetic gene clusters. However, existinghigh-copy-number Streptomyces plasmids tolerate large (�30-kb) DNA insertions poorly (46). To date, available high- ormedium-copy-number vectors, such as those derived frompIJ101 (8.8 kb, 50 to 300 copies per chromosome) (24, 26),pJV1 (11.1 kb, 150 copies per chromosome) (42), and pSG5
(12.2 kb, 20 to 50 copies per chromosome) (36), have lowplasmid stability and structural integrity when carrying largeinserts. However, Hu et al. recently discovered a plasmid,SCP2@, that breaks this trend with its high copy number,stability, and tolerance of large DNA inserts (17). This newplasmid represents a breakthrough for the expression of largegene clusters in Streptomyces hosts.
Hu et al. found SCP2@ through the discovery of a plasmid-containing CH999 variant that spontaneously produced up to25-fold-higher yields of the erythromycin precursor 6-deoxy-erythronolide B (6-dEB) (17). The plasmid in this strain,pJRJ2, contains the eryA genes encoding the 6-deoxyerythro-nolide B synthase (DEBS) (21) with a mutation in the firstcondensation active site (DEBS1 KS1 null) to allow uptake ofchemically altered substrates for polyketide biosynthesis (20).However, preparations of the plasmid DNA from the high-producing strain revealed that pJRJ2 recombined with a plas-mid similar to the naturally occurring SCP2*, designatedSCP2@. The 81-kb cointegrate plasmid, named pSmall, wasdetermined to have a copy number of ca. 100 to 125 copies perchromosome (17). In contrast, pJRJ2 and other SCP2*-derivedvectors replicate at ca. one to five copies per chromosome. Thisincrease in plasmid copy number was linked to the observedincrease in product titers. Sequencing of SCP2@ revealed a45-base-pair deletion in its replicon relative to the SCP2* se-quence (accession no. NC_003904). This mutation may be re-sponsible for the increase in plasmid copy number.
In this work, we extended the two main findings of Hu et al.that the 45-bp deletion increased plasmid copy number andthat the properties of a newly described SCP2@-derived plas-
* Corresponding author. Present address: 318 Campus Drive, RoomW-300, Stanford, CA 94305. Phone: (650) 736-0548. Fax: (650) 723-9780. E-mail: [email protected].
� Published ahead of print on 1 December 2006.
1296
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 19
Nov
embe
r 20
21 b
y 69
.65.
92.5
1.

mid called pBoost (see below) represent a technological ad-vance for high metabolite production (17). We discovered thatlarge plasmids containing only the minimal high-copy repliconwere unstable. To identify genes required for plasmid stabilityand high product titers and to progress towards a small cloningvector amenable to standard ligation protocols, we constructeda series of plasmids with different subsets of SCP2* genes. A50-kb plasmid with the high-copy replicon, the partition geneparAB, an intramycelial transfer gene, spd, a region containingsix putative open reading frames (ORFs) from SCP2*, and theeryA polyketide synthase genes could replicate and generatehigh product titers. However, a stability assay showed that thisplasmid was lost from the mycelium population over time,suggesting that other SCP2* regions were required for long-term plasmid maintenance. In addition, gene titration experi-ments determined that a copy number of the activator genehigher than those of biosynthetic genes increased product ti-ters by increasing transcript levels of the biosynthetic genes.Finally, we found that the 45-bp deletion abolished binding ofa plasmid replication protein to the plasmid DNA, supportingan iteron model for plasmid replication.
MATERIALS AND METHODS
Replication origin DNA sequencing. The primers for replication origin PCRand DNA sequencing are as follows: 5�-TGCCTACGGCCTGCAAGGTG and5�-CGCGCGCCACCTCGTCGGCT.
Cloning using in-gel ligation. One to 2 micrograms of plasmid DNA wasdigested after 12 to 16 h using 10 to 40 units of restriction enzyme (New EnglandBiolabs, Beverly, MA). Gel electrophoresis of the digested DNA used 0.8%SeaPlaque GTG agarose (catalog no. 50111; Cambrex Bio Science Rockland,Inc., Rockland, ME), 10 �g/ml crystal violet, and Tris-acetate-EDTA buffer with0.1 mM EDTA. DNA bands were excised, melted at 70°C, cooled to 37°C, andadded to a 40-�l total ligation volume with 1 unit of T4 DNA ligase (InvitrogenLife Technologies, Carlsbad, CA). The reaction mixture was incubated at roomtemperature for 12 to 16 h and melted at 70°C for transformation into chemical-competent Escherichia coli XL1-Blue. Cells were plated onto Luria-Bertani (LB)medium with a drug concentration of 100 mg/liter carbenicillin, 50 mg/literkanamycin, or 50 mg/liter apramycin. Artificial DNA linkers used to join twononcomplementary ends of a digested plasmid were annealed in 10 mM Tris-Clbuffer, pH 8.5 (QIAGEN buffer EB), with a 100 nM concentration of eacholigonucleotide, by using a GeneAmp PCR System 9700 (Applied Biosystems,Foster City, CA) PCR machine set to 98°C for 45 s and decreasing 0.5°C every30 s until 4°C and added directly to a ligation reaction.
Stability and transfer gene cloning. Stability and transfer genes were clonedfrom SCP2* by using the following primers: spd, 5�-TCT AGA TGG CTT GACGCG GCT and 5�-TCT AGA CCT GAC CGA CCT TCG G; traA, 5�-TCT AGAGGT GCG GGG TCA GGA and 5�-TCT AGA GCA GCA CCA GGA GCC;mrpA, 5�-ACT AGT ATG GGG TAG CCG TCC G and 5�-ACT AGT GTTGCC AGA TGC TGA GC; and parAB, 5�-AAG CTT GGG CTT ACG CGTCGT and 5�-AAG CTT CCG TAC CGT ACC GGC T. These primers werelocated upstream and downstream of the genes’ start and stop codons, respec-tively, of these distances (in base pairs): spd, 100 and 22; traA, 119 and 14; mrpA,136 and 18; and parAB, 76 and 4.
Isolation of plasmid constructs from E. coli. Plasmids smaller than 40 kb wereisolated using the QIAprep Spin miniprep kit (catalog no. 27106; QIAGEN Inc.,Valencia, CA) following the manufacturer’s instructions and eluted with bufferEB (10 mM Tris-Cl, pH 8.5) at 70°C. For plasmids �40 kb, the TENS phenol-chloroform E. coli large plasmid preparation was used. TENS solution contained10 mM Tris-HCl, pH 7.5, 1 mM EDTA, 0.1 N NaOH, and 0.5% sodium dodecylsulfate (SDS). After 12 to 16 h of growth in liquid culture, 1.5 ml of cells waspelleted, lysed with 600 �l of TENS solution, and precipitated with 320 �l of 3.0M sodium acetate (pH 5.2). The supernatant was extracted with 250 �l phenol-chloroform-isoamyl alcohol and then 400 �l chloroform, then precipitated withisopropanol, and washed twice with 70% ethanol. The DNA pellet was resus-pended in 40 to 60 �l of buffer EB containing RNase H (24 �g/ml).
Introduction of plasmids into Streptomyces by transformation. Nonmethylatedplasmid DNA was isolated from transformants of E. coli GM (dam dcm) or
ET12567 (dam dcm hsdM) strains (37). Plasmids were then introduced intoStreptomyces coelicolor by using protoplast transformation, grown, and preservedas described previously (25).
Quantification of polyketide production. 6-dEB macrolide product quantifi-cation of the supernatant from CH999 liquid fermentations was carried out usinga high-performance liquid chromatography (HPLC) assay with known standardconcentrations of 6-dEB as described previously (33).
Total DNA isolation from Streptomyces coelicolor. A total of 750 to 1,000 �l ofmycelium from liquid culture was pelleted, washed with 1 ml of lysis buffer (15%sucrose, 25 mM Tris-HCl, 25 mM EDTA), and incubated for 45 min at 37°C in450 �l lysis buffer containing 5 mg/ml lysozyme (Sigma-Aldrich Inc., St. Louis,MO) and mixed frequently. Fifty-five microliters of 10% SDS and 0.1 mg ofproteinase K (Sigma-Aldrich) were added and incubated at 50°C for 5 min.Eighty-five microliters of 5 M NaCl and 450 �l distilled water were added, mixed,and extracted once with 350 �l buffer-saturated phenol, twice with 400 �l phenol-chloroform-isoamyl alcohol, and once with 400 �l chloroform. Nucleic acids werepelleted by adding 1 volume of isopropanol, washed twice with 1 ml 70% ethanol,and resuspended in 40 to 60 �l buffer EB containing RNase H (24 �g/ml).
Qualitative measurement of plasmid copy number using total DNA digestion.Forty micrograms of total DNA was digested with 20 units of restriction endo-nuclease (New England Biolabs) that would yield a characteristic plasmid DNAbanding pattern upon agarose gel electrophoresis. Samples containing high-copy-number plasmids produced bands visible over the background of thegenomic DNA smear. These bands were absent in the low-copy-number plasmidDNA samples.
Quantitative measurement of plasmid copy number using reverse transcrip-tion (RT)-PCR. Plasmid copy number relative to the chromosome was calculatedusing the following equation: plasmid copy number � 2(Tc_genome � Tc_plasmid),where Tc_genome and Tc_plasmid are the cycle numbers at which fluorescenceis detectable at a given threshold above noise (32). The following primers wereused for genomic DNA: thiC, 5�-ACC GCG GAG AAC ACG GAC AC and5�-CGG ACA TCG GTG TCG ACG AG; dnaB, 5�-ATG GAG ATC CGC GCCAAG TG and 5�-AGC TGG GAG AGC GCG ATG AC; and bldD, 5�-CCGCCG AAG CTG GTC CTG and 5�-CCG AGG GCG ACT GGT CGT AG. Thefollowing primers were used for plasmid DNA: actII-ORF4, 5�-GCT GCG GCTTTT TGG AAT GC and 5�-CGC CGG AGA TTC CGA TAC GA; repI, 5�-GCACCA GTT CGA CGG GAA GG and 5�-TTC CGG CAG GTG TCC TTG CT;tsr, 5�-AAA TGT CGC CAT CCG CCT TG and 5�-GAG CGT CGG GGA TCATCC TG; SCP2 ori #1, 5�-TCC CGA CTG ATG CCA CCT GA and 5�-CCTCCC TGG TGC GGG TGA T; SCP2 ori #2, 5�-CGG CGC GTC TCC CAG GTand 5�-TCG AGG ACC GGC GCA TC; pIJ101-rep-ori-1, 5�-CCT CGG CATCGC TCC GTA CT and 5�-CCA AGT CAC ACC AGC CCC AAG; pIJ101-rep-ori-2, 5�-CCT CTC GGG CTC TCC CCA TC and 5�-CCA CAC ACC GGGCAA ACG; aphII #1, 5�-GCT CTG ATG CCG CCG TGT TC and 5�-CGCCCA ATA GCA GCC AGT CC; and aphII #2, 5�-TGC CCA TTC GAC CACCAA GC and 5�-TAT TCG GCA AGC AGG CAT CG. The PCR was preparedusing the iQ SYBR green supermix (catalog no. 170-8880; Bio-Rad Laboratories,Hercules, CA) following the manufacturer’s instructions with 0.6 pmol/�l prim-ers and 0.24 ng/�l total DNA template and performed on the iCycler thermalcycler and real-time PCR detection system (Bio-Rad), using the SYBR-490 filterfor fluorescence detection (measured at the end of each cycle). The PCR con-ditions were 2 min 45 s at 95°C, followed by 40 cycles of 20 s at 95°C, 20 s at 64°C,and 30 s at 72°C. The analysis of each total DNA sample was performed intriplicate.
Colony plating assay for plasmid stability. Mycelium was grown in 10 ml oftryptic soy broth with the appropriate antibiotic selection for 24 to 36 h. Twohundred microliters of the mycelium was added to 10 ml fresh tryptic soy brothwithout antibiotic for the next “growth cycle.” Fifty microliters of the first culturewas vortexed for 1 min with small glass beads, mixed with 1 ml of 10% sucrosesolution, of which 5 �l was plated onto a nonselective R5 agar plate, and thenincubated overnight at 30°C. Single colonies (50 to 100) were replica plated ontoplates with and without antibiotic and incubated at 30°C for 2 days. Colonieswere then examined for growth and counted. Plasmid stability was expressed asa percentage of colonies that grew on both plates compared to colonies that grewonly on the nonselective plate. This procedure was repeated for subsequentgrowth cycles.
Microarray analysis of transcript levels. Strains harboring pKOS011-26 andpKOS011-26* were grown in rich medium, and mRNA samples were harvestedafter 16, 20, 24, 40, 64, and 88 h. Microarray procedures were followed aspreviously reported (18, 23). Data were analyzed using the Stanford MicroarrayDatabase (http://www-genome.stanford.edu/microarray) (2).
Plasmid cloning for actII-ORF4 and eryA relative copy number study. pRF20was created using a synthetic oligonucleotide linker with the sequence PacI-RBS-
VOL. 73, 2007 CHARACTERIZATION OF A LARGE HIGH-COPY-NUMBER PLASMID 1297
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 19
Nov
embe
r 20
21 b
y 69
.65.
92.5
1.

NdeI-BglII-SpeI-XbaI-EcoRI cloned into the vector portion of pKOS011-26digested with PacI-EcoRI. pRF20 digested with KpnI-EcoRV and ligated withthe synthetic linker KpnI-EcoRI-PstI-PacI-EcoRV produced pRF21. pRF21 di-gested with SalI and religated to eliminate actII-ORF4 produced pRF22. TheNdeI fragment from pRF22, now lacking actII-ORF4, was swapped with the NdeIfragment of pKOS011-26 to produce pRF23. pRF26 was created by first remov-ing an EcoRI site near the E. coli origin from pBoost to produce pRF8. aphII(neomycin phosphotransferase) added to pRF8 produced pRF15. actII-ORF4cloned into pRF15 produced pRF26. pRF152 was created by first inserting aphIIinto pWHM3 (44) at the ClaI-SstI site to produce pRF150. actII-ORF4 was theninserted into the EcoRI site of pRF150 to produce pRF152.
RT-PCR analysis of transcript levels. Total RNA samples were purified fromliquid culture using an RNeasy plant mini kit (catalog no. 74903; QIAGEN)following the manufacturer’s instructions. First-strand cDNA was synthesizedfrom 200 ng of the total RNA using SuperScript II (catalog no. 18064-014;Invitrogen) following the manufacturer’s instructions. To allow for the high GCcontent of Streptomyces, we used a deoxynucleoside triphosphate mix of 10 mMdGTP, 10 mM dCTP, 4 mM dATP, 4 mM dTTP, and random hexamers with72% GC content. Twenty units of RNaseOUT (catalog no. 10777-019; Invitro-gen) were also added to each reaction. RT-PCR was performed as describedabove using 1% of the first-strand reaction as the DNA template. 16S rRNAserved as the endogenous control. The following primers were used: repI 1,5�-ACG TCC TCG CCC AGC TCC TC and 5�-GCA CCA GTT CGA CGGGAA GG; repI 2, 5�-GGG TAG CCG GGA TGG TCT T and CGC GAT CCGGCT CTT ACC AG; repII 1, 5�-CGA GCG TCG GTG TTG CTG AC and GAACCT CCT CGT GCG GAC CTC; repII 2, 5�-GGT CCG CAC GAG GAG GTTCA and CTG TCC GCC GCT CAG CAC T; and 16S rRNA, 5�-CGA CGC AACGCG AAG AAC CT and 5�-TGC TGG CAA CAC GGG ACA AG.
Fluorescent EMSA. The fluorescent electrophoretic gel mobility shift assay(EMSA) protocol using Cy3 dCTP was developed by Lum (34). To harvest totalcrude Streptomyces proteins, 1 ml of mycelium from liquid culture was pelletedand washed twice in 500 �l ice-cold TA buffer (10 mM Tris, pH 7.5, 10 mMMgCl2, 1 mM EDTA, 1 mM dithiothreitol, 0.1% Triton X-100, 10% glycerol, 50mM NaCl), resuspended in 500 �l TA buffer with 1� complete protease inhib-itor cocktail (catalog no. 1697498; Roche Ltd., Basel, Switzerland), and sonicatedon ice using a model 450 digital sonifier (Branson Ultrasonics Corp., Danbury,CT) with 40% amplitude (four cycles of 10 s each with 0.5 s on and 1.0 s off eachcycle, with cooling on ice for 1 minute between cycles). The mixture was centri-fuged for 15 min at 13,200 rpm, the supernatant recovered, and the proteinconcentration measured using the method of Bradford (6) on a SpectraMax Plus384 spectrophotometer (Molecular Devices Corp., Sunnyvale, CA). RepI proteinwas produced in E. coli strain BL21 using the pET21-b vector (Novagen Inc.,Madison, WI) and repI PCR primers: 5�-CAT ATG GCC CTG GTC AAC ATGG and 5�-CTC GAG TCG CGT CTC TCC TGC. BL21 total crude protein
extract containing RepI was isolated as described above for Streptomyces. Thefollowing primers were used to generate probes 1 (247 bp) and 1-M (257 bp)using pKOS011-26 template: 5�-AGCCCGAGGGAGTACAGG and 5�-GTGGTGACCTGTTAGTTTCCTCT. The following primers were used to generateprobes 2 (202 bp) and 2-M (212 bp) using pKOS011-26* template: 5�-ACTCGCGCCTCTCCCAT and 5�-CTCTTGACCTGGTAAAACGCG. An extensiontime of 30 min at 72°C was added at the end of the PCR for tail addition of 3�adenines. The PCR product was fluorescently end labeled using terminal de-oxynucleotide transferase (New England Biolabs) and Cy3 dCTP (AmershamPharmacia Biotech, Piscataway, NJ) following the manufacturer’s instructions. Atotal of 1.5 �g of a 100-bp DNA ladder (New England Biolabs) was also labeled.Following end labeling, samples were washed with 400 �l Tris-EDTA bufferusing Microcon 10 filters (Millipore, Bedford, MA) and concentrated to 20 �l.Crude protein extract and Cy3-labeled DNA probe (�100 ng) were allowed tointeract in a 20-�l volume with 10 mM Tris, 5 mM MgCl2, 60 mM KCl, 50 mMEDTA, 10 mM dithiothreitol, 10% glycerol, and 1 �g poly(dI-dC) and wereincubated in ice for 10 to 15 min. Gel electrophoresis of the reaction was thencarried out using a 5% polyacrylamide Tris-borate-EDTA gel (Bio-Rad), andimages were taken on a Typhoon model 9410 variable-mode fluorescence imager(Amersham).
Microarray data accession number. The microarray data discussed in thispublication have been deposited into the NCBI Gene Expression Omnibus(GEO; http://www.ncbi.nlm.nih.gov/geo/) and are accessible through GEO Seriesaccession number GSE6096.
RESULTS
Copy numbers of SCP2@-derived plasmids. pKOS011-26(39) and its high-copy-number variant, pKOS011-26*, repre-sent the standard low- and high-copy-number plasmids, respec-tively, for this work (Table 1). pKOS011-26 lacks mutations inits active-site domains, which eliminates the substrate feedingrequirement for 6-dEB synthesis. pKOS011-26* was created bycointegration of pKOS011-26 with SCP2@.
The 45-bp deletion was detected in plasmid replicons byusing PCR and DNA sequencing. Copy numbers were mea-sured over 3-day time courses using quantitative real-timePCR (see Materials and Methods). 6-dEB titers were mea-sured using HPLC after 6- or 7-day fermentations. Table 2summarizes these data for relevant SCP2*-derived plasmids.
TABLE 1. Plasmids used in this study
Plasmid Description Reference or source
SCP2* High-fertility variant of low-copy Streptomyces plasmid SCP2 5SCP2@ SCP2* containing 45-bp deletion in replication origin 17pBoost SCP2@ with pBR322-derived replicon and apramycin resistance 17pJRJ2 pRM5 derivative containing eryA with KS1° mutation 20pJRJ2 pJRJ2 containing 45-bp deletion in replication origin Kosan BiosciencespKOS011-26 pRM5 derivative containing functional eryA 39pKOS011-26* Cointegrate of SCP2@ and pKOS011-26 Kosan BiosciencespKOS159-10 pSET152 derivative with C31 integrase, actII-ORF4, and eryA Kosan BiosciencespRM5 Low-copy SCP2*-derived shuttle vector 38pSmall Cointegrate of SCP2@ and pJRJ2 17pRF26 pBoost containing actII-ORF4 and neomycin resistance This studypRF62 Minimal low-copy SCP2* replicon in pBR322-derived vector This studypRF63 Minimal high-copy SCP2@ replicon in pBR322-derived vector This studypRF89 pBoost lacking EcoRI fragment that contains SCP2.01 to SCP2.12c This studypRF103 Shuttle vector with SCP2@ replicon, actII-ORF4, and actI promoter This studypRF104 Same as pRF103 but contains apramycin marker in reverse direction This studypRF121-pRF126 Various pRF127 derivatives (see Fig. 2) This studypRF127 pRF104 containing eryA; unstable in Streptomyces This studypRF128 Direct ligation of pBoost with actII-ORF4, actI promoter, and eryA This studypRF152 pIJ101 derivative containing actII-ORF4 This studypRF160 pJRJ2 containing functional eryA This studypRF169 pRF160 with parAB under actIII promoter; high production This study
1298 FONG ET AL. APPL. ENVIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 19
Nov
embe
r 20
21 b
y 69
.65.
92.5
1.
All high-copy-number plasmids possess the 45-bp deletion,whereas the low-copy-number plasmids contain the wild-typesequence. pKOS011-26* had a threefold-higher plasmid copynumber and a threefold-higher titer than pKOS011-26, show-ing that an elevated copy number correlates with elevatedtiters. However, the high-copy-number plasmid pJRJ2, whicharose by resolution of the cointegrate plasmid pSmall, gener-ated low titers (Z. Hu, personal communication). It containsthe 45-bp deletion but is otherwise identical to pJRJ2. ThatpJRJ2 produced low titers revealed the insufficiency of the45-bp deletion to generate high product titers and thereforethe requirement of other SCP2@ genes.
The replication region sequencing data reflect an extra G,relative to the published SCP2* sequence (accession no.NC_003904), 619 bp upstream of the repI gene. This extra basewas found in both the high- and low-copy-number plasmids.We assumed it had no effects on plasmid replication or simplyreflected an error in the published sequence.
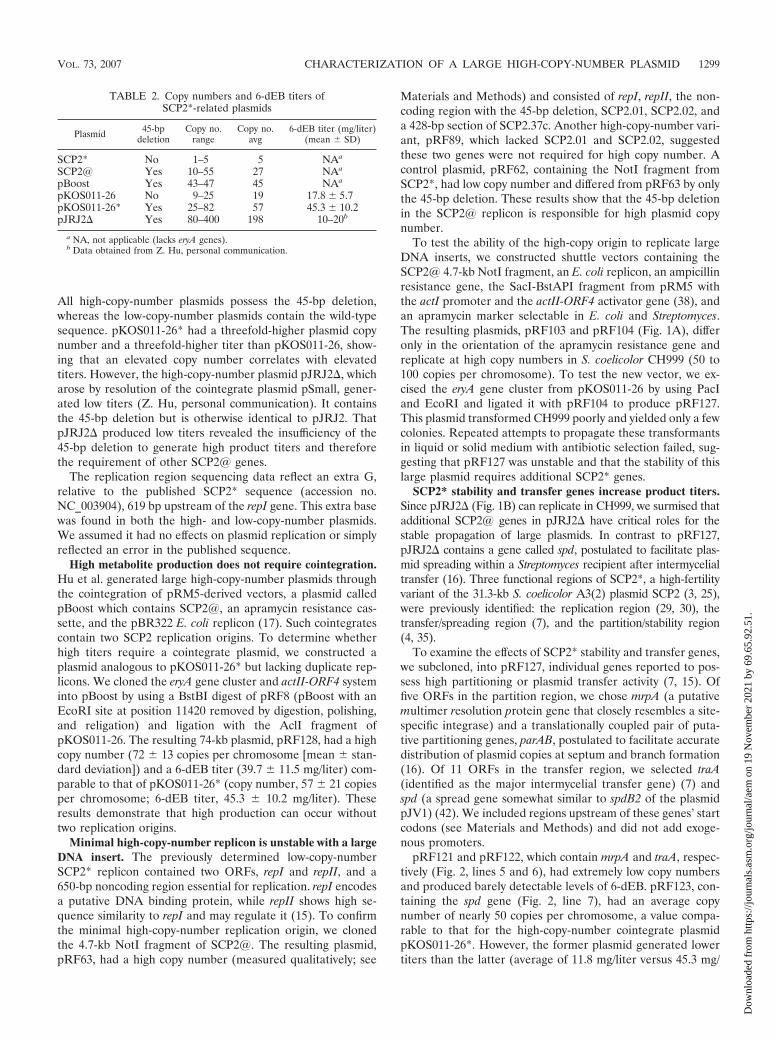
High metabolite production does not require cointegration.Hu et al. generated large high-copy-number plasmids throughthe cointegration of pRM5-derived vectors, a plasmid calledpBoost which contains SCP2@, an apramycin resistance cas-sette, and the pBR322 E. coli replicon (17). Such cointegratescontain two SCP2 replication origins. To determine whetherhigh titers require a cointegrate plasmid, we constructed aplasmid analogous to pKOS011-26* but lacking duplicate rep-licons. We cloned the eryA gene cluster and actII-ORF4 systeminto pBoost by using a BstBI digest of pRF8 (pBoost with anEcoRI site at position 11420 removed by digestion, polishing,and religation) and ligation with the AclI fragment ofpKOS011-26. The resulting 74-kb plasmid, pRF128, had a highcopy number (72 � 13 copies per chromosome [mean � stan-dard deviation]) and a 6-dEB titer (39.7 � 11.5 mg/liter) com-parable to that of pKOS011-26* (copy number, 57 � 21 copiesper chromosome; 6-dEB titer, 45.3 � 10.2 mg/liter). Theseresults demonstrate that high production can occur withouttwo replication origins.
Minimal high-copy-number replicon is unstable with a largeDNA insert. The previously determined low-copy-numberSCP2* replicon contained two ORFs, repI and repII, and a650-bp noncoding region essential for replication. repI encodesa putative DNA binding protein, while repII shows high se-quence similarity to repI and may regulate it (15). To confirmthe minimal high-copy-number replication origin, we clonedthe 4.7-kb NotI fragment of SCP2@. The resulting plasmid,pRF63, had a high copy number (measured qualitatively; see
Materials and Methods) and consisted of repI, repII, the non-coding region with the 45-bp deletion, SCP2.01, SCP2.02, anda 428-bp section of SCP2.37c. Another high-copy-number vari-ant, pRF89, which lacked SCP2.01 and SCP2.02, suggestedthese two genes were not required for high copy number. Acontrol plasmid, pRF62, containing the NotI fragment fromSCP2*, had low copy number and differed from pRF63 by onlythe 45-bp deletion. These results show that the 45-bp deletionin the SCP2@ replicon is responsible for high plasmid copynumber.
To test the ability of the high-copy origin to replicate largeDNA inserts, we constructed shuttle vectors containing theSCP2@ 4.7-kb NotI fragment, an E. coli replicon, an ampicillinresistance gene, the SacI-BstAPI fragment from pRM5 withthe actI promoter and the actII-ORF4 activator gene (38), andan apramycin marker selectable in E. coli and Streptomyces.The resulting plasmids, pRF103 and pRF104 (Fig. 1A), differonly in the orientation of the apramycin resistance gene andreplicate at high copy numbers in S. coelicolor CH999 (50 to100 copies per chromosome). To test the new vector, we ex-cised the eryA gene cluster from pKOS011-26 by using PacIand EcoRI and ligated it with pRF104 to produce pRF127.This plasmid transformed CH999 poorly and yielded only a fewcolonies. Repeated attempts to propagate these transformantsin liquid or solid medium with antibiotic selection failed, sug-gesting that pRF127 was unstable and that the stability of thislarge plasmid requires additional SCP2* genes.
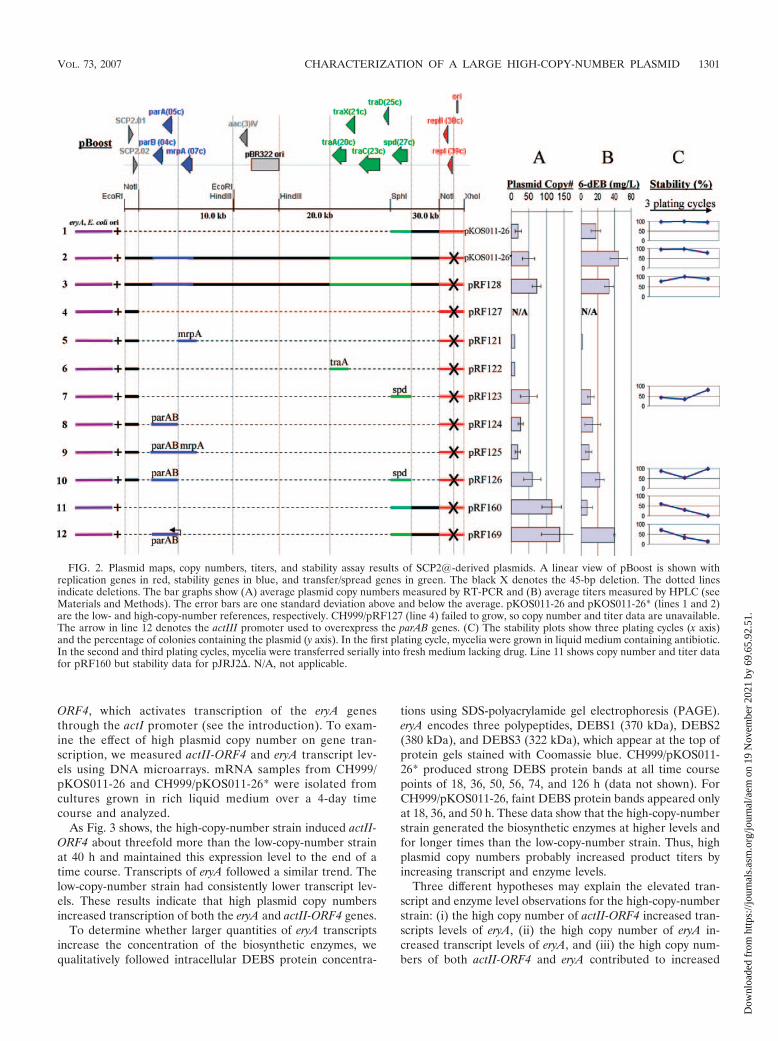
SCP2* stability and transfer genes increase product titers.Since pJRJ2 (Fig. 1B) can replicate in CH999, we surmised thatadditional SCP2@ genes in pJRJ2 have critical roles for thestable propagation of large plasmids. In contrast to pRF127,pJRJ2 contains a gene called spd, postulated to facilitate plas-mid spreading within a Streptomyces recipient after intermycelialtransfer (16). Three functional regions of SCP2*, a high-fertilityvariant of the 31.3-kb S. coelicolor A3(2) plasmid SCP2 (3, 25),were previously identified: the replication region (29, 30), thetransfer/spreading region (7), and the partition/stability region(4, 35).
To examine the effects of SCP2* stability and transfer genes,we subcloned, into pRF127, individual genes reported to pos-sess high partitioning or plasmid transfer activity (7, 15). Offive ORFs in the partition region, we chose mrpA (a putativemultimer resolution protein gene that closely resembles a site-specific integrase) and a translationally coupled pair of puta-tive partitioning genes, parAB, postulated to facilitate accuratedistribution of plasmid copies at septum and branch formation(16). Of 11 ORFs in the transfer region, we selected traA(identified as the major intermycelial transfer gene) (7) andspd (a spread gene somewhat similar to spdB2 of the plasmidpJV1) (42). We included regions upstream of these genes’ startcodons (see Materials and Methods) and did not add exoge-nous promoters.
pRF121 and pRF122, which contain mrpA and traA, respec-tively (Fig. 2, lines 5 and 6), had extremely low copy numbersand produced barely detectable levels of 6-dEB. pRF123, con-taining the spd gene (Fig. 2, line 7), had an average copynumber of nearly 50 copies per chromosome, a value compa-rable to that for the high-copy-number cointegrate plasmidpKOS011-26*. However, the former plasmid generated lowertiters than the latter (average of 11.8 mg/liter versus 45.3 mg/
TABLE 2. Copy numbers and 6-dEB titers ofSCP2*-related plasmids
Plasmid 45-bpdeletion
Copy no.range
Copy no.avg
6-dEB titer (mg/liter)(mean � SD)
SCP2* No 1–5 5 NAa
SCP2@ Yes 10–55 27 NAa
pBoost Yes 43–47 45 NAa
pKOS011-26 No 9–25 19 17.8 � 5.7pKOS011-26* Yes 25–82 57 45.3 � 10.2pJRJ2 Yes 80–400 198 10–20b
a NA, not applicable (lacks eryA genes).b Data obtained from Z. Hu, personal communication.
VOL. 73, 2007 CHARACTERIZATION OF A LARGE HIGH-COPY-NUMBER PLASMID 1299
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 19
Nov
embe
r 20
21 b
y 69
.65.
92.5
1.

liter, respectively). pRF124, which contains parAB (Fig. 2, line8), had a copy number half that of pRF123, but the two plas-mids generated nearly equal 6-dEB titers. pRF125, containingboth parAB and mrpA (Fig. 2, line 9), had a copy number andproduct titer similar to those of the plasmid with parAB alone(pRF124). pRF126, which contains both spd and parAB (Fig. 2,line 10), had a high copy number but a low 6-dEB titer, likepRF123. However, pRF126 generated titers twofold higherthan those of pRF123 (22.5 mg/liter in CH999/pRF126 versus11.8 mg/liter in CH999/pRF123).
These results counter our original hypothesis that high plas-mid copy number is the cause of high 6-dEB titers. In addition,pRF123 and pRF126 had the same high copy number butgenerated different amounts of product. Of the genes tested,the spd and parAB genes together caused the largest increasein plasmid copy number and product titer.
pJRJ2 had significantly higher copy number than pRF123and contained six putative ORFs between the minimal high-copy origin and spd that pRF123 lacked. Only one putativegene in this region, SCP2.35, bears any similarity to knowngenes and is thought to be a transfer gene regulator (15). SincepJRJ2 was already available as another variant containingSCP2* genes to be tested, and for consistency with our otherplasmids, we replaced the active-site mutation in pJRJ2 withwild-type eryA sequence to generate pRF160 (Fig. 2, line 11),which could produce 6-dEB without substrate feeding. pRF160had a copy number more than twofold higher than that ofpRF123 but generated only half the product titer. Based onour finding that parAB genes enhanced product titers, weadded these genes to pRF160 under the control of the actIIIpromoter, thereby overexpressing these genes. The resultingplasmid, pRF169 (Fig. 2, line 12), had nearly the same copynumber as pRF160 (�115 copies per chromosome). However,pRF169 generated 39.8 � 0.4 mg/liter of 6-dEB, which greatly
surpassed that of pRF160 and approached that of the originalhigh-copy-number cointegrate pKOS011-26* (45.3 � 10.2 mg/liter). Thus, overexpression of the parAB genes over a very-high-plasmid-copy-number background increased product ti-ters significantly.
Plasmid stability analysis. To probe how the spd, parA, andparB genes enhance copy number and metabolite production, weexamined plasmid distribution in the mycelium population byusing a colony plating assay. Since Streptomyces grows as mycelialclumps, liquid cultures were vortexed vigorously with glass beadsto break up these clumps before being plated for single colonies.Stability values were expressed as a percentage of colonies thatcontained the plasmid. Figure 2C shows plasmid stability dataover three plating cycles. The low-copy-number pKOS011-26showed high stability over three plating cycles, maintaining a100% distribution in the mycelium population. The high-copy-number, high-producing plasmids pKOS011-26* and pRF128also remained well distributed. In contrast, the low-producingplasmid pJRJ2 (data shown with pRF160 in Fig. 2, line 11) hadan average stability of 30% over the three plating cycles and waslost completely by the third cycle. pRF123 and pRF126, withaverage stabilities of 52% and 79%, respectively, had higher titersthan pRF160 (pJRJ2) but lower titers than pKOS011-26*. Thehigher stability of pRF126 may explain its titers being twofoldhigher than those of pRF123. Additionally, the high-producingplasmid pRF169 (average stability of 40%) was better distributedthan pJRJ2 (30%). In general, greater plasmid stability in themycelium population correlated with higher product titers.
Effect of actII-ORF4 and eryA gene dosage on metaboliteproduction. That pKOS011-26* led to improved metaboliteproduction is consistent with the generally accepted notionthat gene transcription increases with higher copy numbersof the biosynthetic genes. pRM5-related plasmids contain apathway-specific transcriptional regulator, encoded by actII-
FIG. 1. (A) High-copy-number shuttle vector pRF104. SCP2.37 is truncated (see text). The section containing actII-ORF4, actIII, the divergentactI/III promoter, the �-lactamase marker (bla), and the E. coli replication origin is from pRM5. (B) pJRJ2. “ori SCP2@” denotes the contiguousSphI-XhoI portion from SCP2@ with replication genes repI and repII and transfer gene spd. Six putative genes lie between repII and spd (notshown). The three eryA genes are shown between PacI and EcoRI.
1300 FONG ET AL. APPL. ENVIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 19
Nov
embe
r 20
21 b
y 69
.65.
92.5
1.
ORF4, which activates transcription of the eryA genesthrough the actI promoter (see the introduction). To exam-ine the effect of high plasmid copy number on gene tran-scription, we measured actII-ORF4 and eryA transcript lev-els using DNA microarrays. mRNA samples from CH999/pKOS011-26 and CH999/pKOS011-26* were isolated fromcultures grown in rich liquid medium over a 4-day timecourse and analyzed.
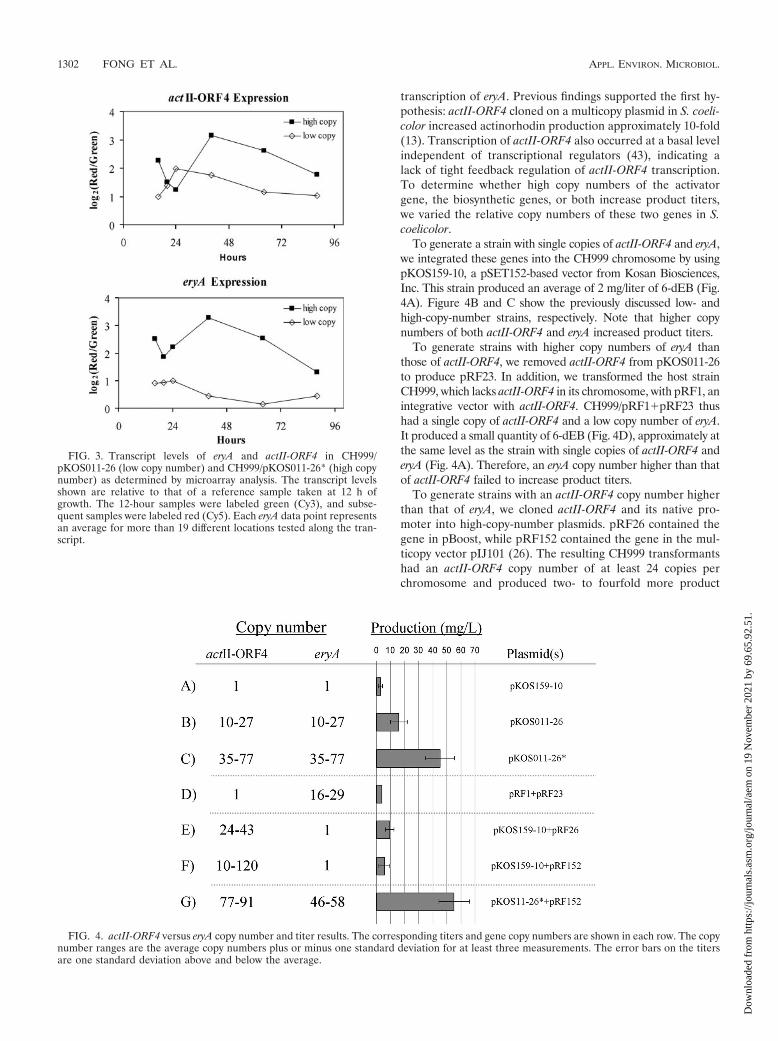
As Fig. 3 shows, the high-copy-number strain induced actII-ORF4 about threefold more than the low-copy-number strainat 40 h and maintained this expression level to the end of atime course. Transcripts of eryA followed a similar trend. Thelow-copy-number strain had consistently lower transcript lev-els. These results indicate that high plasmid copy numbersincreased transcription of both the eryA and actII-ORF4 genes.
To determine whether larger quantities of eryA transcriptsincrease the concentration of the biosynthetic enzymes, wequalitatively followed intracellular DEBS protein concentra-
tions using SDS-polyacrylamide gel electrophoresis (PAGE).eryA encodes three polypeptides, DEBS1 (370 kDa), DEBS2(380 kDa), and DEBS3 (322 kDa), which appear at the top ofprotein gels stained with Coomassie blue. CH999/pKOS011-26* produced strong DEBS protein bands at all time coursepoints of 18, 36, 50, 56, 74, and 126 h (data not shown). ForCH999/pKOS011-26, faint DEBS protein bands appeared onlyat 18, 36, and 50 h. These data show that the high-copy-numberstrain generated the biosynthetic enzymes at higher levels andfor longer times than the low-copy-number strain. Thus, highplasmid copy numbers probably increased product titers byincreasing transcript and enzyme levels.
Three different hypotheses may explain the elevated tran-script and enzyme level observations for the high-copy-numberstrain: (i) the high copy number of actII-ORF4 increased tran-scripts levels of eryA, (ii) the high copy number of eryA in-creased transcript levels of eryA, and (iii) the high copy num-bers of both actII-ORF4 and eryA contributed to increased
FIG. 2. Plasmid maps, copy numbers, titers, and stability assay results of SCP2@-derived plasmids. A linear view of pBoost is shown withreplication genes in red, stability genes in blue, and transfer/spread genes in green. The black X denotes the 45-bp deletion. The dotted linesindicate deletions. The bar graphs show (A) average plasmid copy numbers measured by RT-PCR and (B) average titers measured by HPLC (seeMaterials and Methods). The error bars are one standard deviation above and below the average. pKOS011-26 and pKOS011-26* (lines 1 and 2)are the low- and high-copy-number references, respectively. CH999/pRF127 (line 4) failed to grow, so copy number and titer data are unavailable.The arrow in line 12 denotes the actIII promoter used to overexpress the parAB genes. (C) The stability plots show three plating cycles (x axis)and the percentage of colonies containing the plasmid (y axis). In the first plating cycle, mycelia were grown in liquid medium containing antibiotic.In the second and third plating cycles, mycelia were transferred serially into fresh medium lacking drug. Line 11 shows copy number and titer datafor pRF160 but stability data for pJRJ2. N/A, not applicable.
VOL. 73, 2007 CHARACTERIZATION OF A LARGE HIGH-COPY-NUMBER PLASMID 1301
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 19
Nov
embe
r 20
21 b
y 69
.65.
92.5
1.
transcription of eryA. Previous findings supported the first hy-pothesis: actII-ORF4 cloned on a multicopy plasmid in S. coeli-color increased actinorhodin production approximately 10-fold(13). Transcription of actII-ORF4 also occurred at a basal levelindependent of transcriptional regulators (43), indicating alack of tight feedback regulation of actII-ORF4 transcription.To determine whether high copy numbers of the activatorgene, the biosynthetic genes, or both increase product titers,we varied the relative copy numbers of these two genes in S.coelicolor.
To generate a strain with single copies of actII-ORF4 and eryA,we integrated these genes into the CH999 chromosome by usingpKOS159-10, a pSET152-based vector from Kosan Biosciences,Inc. This strain produced an average of 2 mg/liter of 6-dEB (Fig.4A). Figure 4B and C show the previously discussed low- andhigh-copy-number strains, respectively. Note that higher copynumbers of both actII-ORF4 and eryA increased product titers.
To generate strains with higher copy numbers of eryA thanthose of actII-ORF4, we removed actII-ORF4 from pKOS011-26to produce pRF23. In addition, we transformed the host strainCH999, which lacks actII-ORF4 in its chromosome, with pRF1, anintegrative vector with actII-ORF4. CH999/pRF1�pRF23 thushad a single copy of actII-ORF4 and a low copy number of eryA.It produced a small quantity of 6-dEB (Fig. 4D), approximately atthe same level as the strain with single copies of actII-ORF4 anderyA (Fig. 4A). Therefore, an eryA copy number higher than thatof actII-ORF4 failed to increase product titers.
To generate strains with an actII-ORF4 copy number higherthan that of eryA, we cloned actII-ORF4 and its native pro-moter into high-copy-number plasmids. pRF26 contained thegene in pBoost, while pRF152 contained the gene in the mul-ticopy vector pIJ101 (26). The resulting CH999 transformantshad an actII-ORF4 copy number of at least 24 copies perchromosome and produced two- to fourfold more product
FIG. 3. Transcript levels of eryA and actII-ORF4 in CH999/pKOS011-26 (low copy number) and CH999/pKOS011-26* (high copynumber) as determined by microarray analysis. The transcript levelsshown are relative to that of a reference sample taken at 12 h ofgrowth. The 12-hour samples were labeled green (Cy3), and subse-quent samples were labeled red (Cy5). Each eryA data point representsan average for more than 19 different locations tested along the tran-script.
FIG. 4. actII-ORF4 versus eryA copy number and titer results. The corresponding titers and gene copy numbers are shown in each row. The copynumber ranges are the average copy numbers plus or minus one standard deviation for at least three measurements. The error bars on the titersare one standard deviation above and below the average.
1302 FONG ET AL. APPL. ENVIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 19
Nov
embe
r 20
21 b
y 69
.65.
92.5
1.
(Fig. 4E and F) than the strain with single copies of the genes(Fig. 4A). However, these strains had titers approximately5-fold lower than that of the original high-copy-number strain(Fig. 4C) and about 1.5-fold lower than that of the originallow-copy-number strain (Fig. 4B).
Together, these results show that high metabolite produc-tion requires elevated gene doses of both actII-ORF4 and eryA.Additionally, the data suggest that actII-ORF4 copy numbermay limit production. To test whether actII-ORF4 limits pro-duction in strains with multiple copies of eryA, we transformedCH999/pKOS011-26* with pRF152, generating an actII-ORF4copy number slightly higher than that of eryA (Fig. 4G). Theresulting strain produced approximately 10 mg/liter moreproduct than the original high-copy-number strain, indicatingthat further titer increases can arise with an activator genedosage slightly higher than those of the biosynthetic genes.
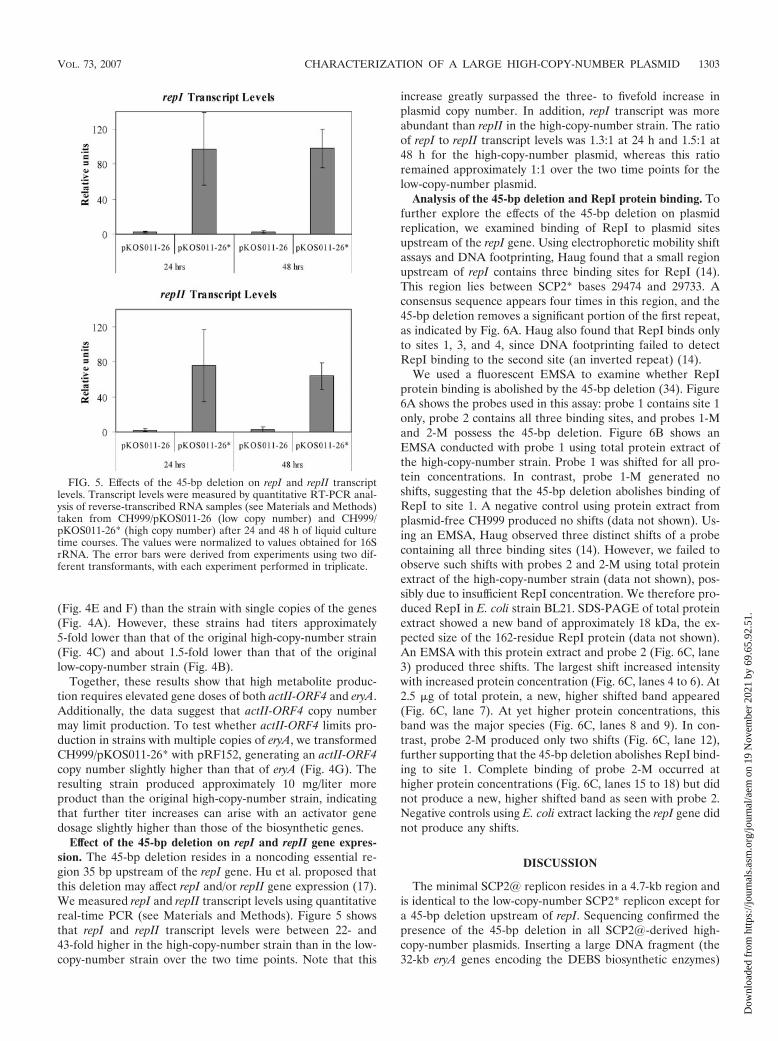
Effect of the 45-bp deletion on repI and repII gene expres-sion. The 45-bp deletion resides in a noncoding essential re-gion 35 bp upstream of the repI gene. Hu et al. proposed thatthis deletion may affect repI and/or repII gene expression (17).We measured repI and repII transcript levels using quantitativereal-time PCR (see Materials and Methods). Figure 5 showsthat repI and repII transcript levels were between 22- and43-fold higher in the high-copy-number strain than in the low-copy-number strain over the two time points. Note that this
increase greatly surpassed the three- to fivefold increase inplasmid copy number. In addition, repI transcript was moreabundant than repII in the high-copy-number strain. The ratioof repI to repII transcript levels was 1.3:1 at 24 h and 1.5:1 at48 h for the high-copy-number plasmid, whereas this ratioremained approximately 1:1 over the two time points for thelow-copy-number plasmid.
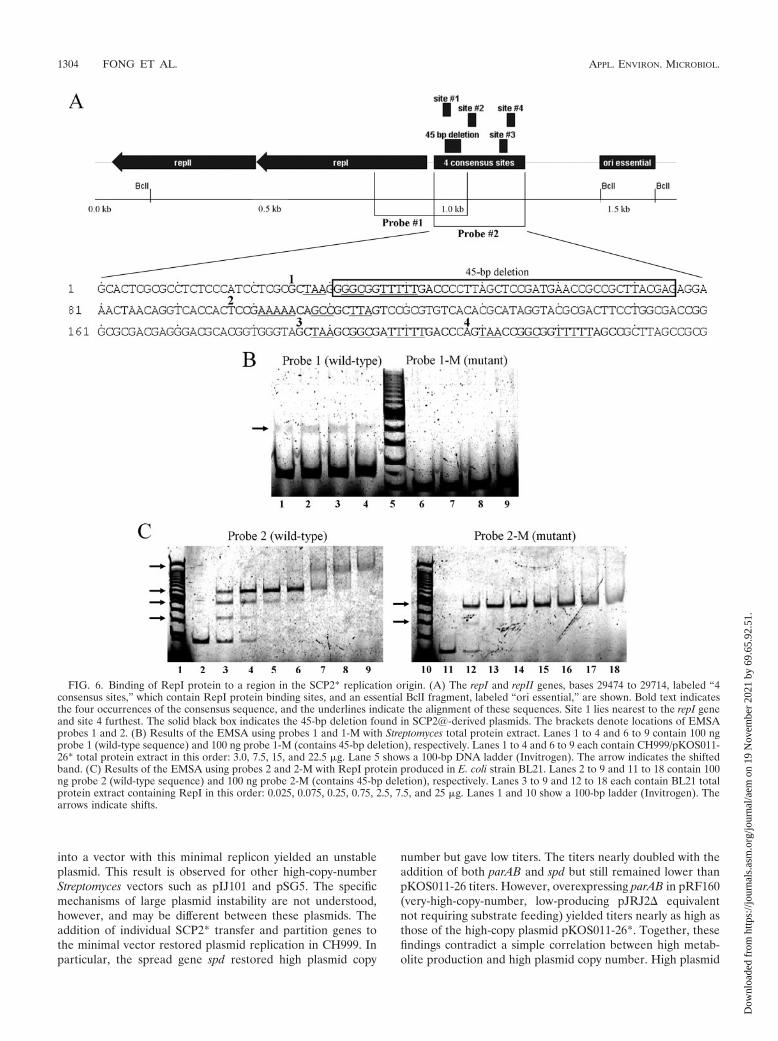
Analysis of the 45-bp deletion and RepI protein binding. Tofurther explore the effects of the 45-bp deletion on plasmidreplication, we examined binding of RepI to plasmid sitesupstream of the repI gene. Using electrophoretic mobility shiftassays and DNA footprinting, Haug found that a small regionupstream of repI contains three binding sites for RepI (14).This region lies between SCP2* bases 29474 and 29733. Aconsensus sequence appears four times in this region, and the45-bp deletion removes a significant portion of the first repeat,as indicated by Fig. 6A. Haug also found that RepI binds onlyto sites 1, 3, and 4, since DNA footprinting failed to detectRepI binding to the second site (an inverted repeat) (14).
We used a fluorescent EMSA to examine whether RepIprotein binding is abolished by the 45-bp deletion (34). Figure6A shows the probes used in this assay: probe 1 contains site 1only, probe 2 contains all three binding sites, and probes 1-Mand 2-M possess the 45-bp deletion. Figure 6B shows anEMSA conducted with probe 1 using total protein extract ofthe high-copy-number strain. Probe 1 was shifted for all pro-tein concentrations. In contrast, probe 1-M generated noshifts, suggesting that the 45-bp deletion abolishes binding ofRepI to site 1. A negative control using protein extract fromplasmid-free CH999 produced no shifts (data not shown). Us-ing an EMSA, Haug observed three distinct shifts of a probecontaining all three binding sites (14). However, we failed toobserve such shifts with probes 2 and 2-M using total proteinextract of the high-copy-number strain (data not shown), pos-sibly due to insufficient RepI concentration. We therefore pro-duced RepI in E. coli strain BL21. SDS-PAGE of total proteinextract showed a new band of approximately 18 kDa, the ex-pected size of the 162-residue RepI protein (data not shown).An EMSA with this protein extract and probe 2 (Fig. 6C, lane3) produced three shifts. The largest shift increased intensitywith increased protein concentration (Fig. 6C, lanes 4 to 6). At2.5 �g of total protein, a new, higher shifted band appeared(Fig. 6C, lane 7). At yet higher protein concentrations, thisband was the major species (Fig. 6C, lanes 8 and 9). In con-trast, probe 2-M produced only two shifts (Fig. 6C, lane 12),further supporting that the 45-bp deletion abolishes RepI bind-ing to site 1. Complete binding of probe 2-M occurred athigher protein concentrations (Fig. 6C, lanes 15 to 18) but didnot produce a new, higher shifted band as seen with probe 2.Negative controls using E. coli extract lacking the repI gene didnot produce any shifts.
DISCUSSION
The minimal SCP2@ replicon resides in a 4.7-kb region andis identical to the low-copy-number SCP2* replicon except fora 45-bp deletion upstream of repI. Sequencing confirmed thepresence of the 45-bp deletion in all SCP2@-derived high-copy-number plasmids. Inserting a large DNA fragment (the32-kb eryA genes encoding the DEBS biosynthetic enzymes)
FIG. 5. Effects of the 45-bp deletion on repI and repII transcriptlevels. Transcript levels were measured by quantitative RT-PCR anal-ysis of reverse-transcribed RNA samples (see Materials and Methods)taken from CH999/pKOS011-26 (low copy number) and CH999/pKOS011-26* (high copy number) after 24 and 48 h of liquid culturetime courses. The values were normalized to values obtained for 16SrRNA. The error bars were derived from experiments using two dif-ferent transformants, with each experiment performed in triplicate.
VOL. 73, 2007 CHARACTERIZATION OF A LARGE HIGH-COPY-NUMBER PLASMID 1303
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 19
Nov
embe
r 20
21 b
y 69
.65.
92.5
1.
into a vector with this minimal replicon yielded an unstableplasmid. This result is observed for other high-copy-numberStreptomyces vectors such as pIJ101 and pSG5. The specificmechanisms of large plasmid instability are not understood,however, and may be different between these plasmids. Theaddition of individual SCP2* transfer and partition genes tothe minimal vector restored plasmid replication in CH999. Inparticular, the spread gene spd restored high plasmid copy
number but gave low titers. The titers nearly doubled with theaddition of both parAB and spd but still remained lower thanpKOS011-26 titers. However, overexpressing parAB in pRF160(very-high-copy-number, low-producing pJRJ2 equivalentnot requiring substrate feeding) yielded titers nearly as high asthose of the high-copy plasmid pKOS011-26*. Together, thesefindings contradict a simple correlation between high metab-olite production and high plasmid copy number. High plasmid
FIG. 6. Binding of RepI protein to a region in the SCP2* replication origin. (A) The repI and repII genes, bases 29474 to 29714, labeled “4consensus sites,” which contain RepI protein binding sites, and an essential BclI fragment, labeled “ori essential,” are shown. Bold text indicatesthe four occurrences of the consensus sequence, and the underlines indicate the alignment of these sequences. Site 1 lies nearest to the repI geneand site 4 furthest. The solid black box indicates the 45-bp deletion found in SCP2@-derived plasmids. The brackets denote locations of EMSAprobes 1 and 2. (B) Results of the EMSA using probes 1 and 1-M with Streptomyces total protein extract. Lanes 1 to 4 and 6 to 9 contain 100 ngprobe 1 (wild-type sequence) and 100 ng probe 1-M (contains 45-bp deletion), respectively. Lanes 1 to 4 and 6 to 9 each contain CH999/pKOS011-26* total protein extract in this order: 3.0, 7.5, 15, and 22.5 �g. Lane 5 shows a 100-bp DNA ladder (Invitrogen). The arrow indicates the shiftedband. (C) Results of the EMSA using probes 2 and 2-M with RepI protein produced in E. coli strain BL21. Lanes 2 to 9 and 11 to 18 contain 100ng probe 2 (wild-type sequence) and 100 ng probe 2-M (contains 45-bp deletion), respectively. Lanes 3 to 9 and 12 to 18 each contain BL21 totalprotein extract containing RepI in this order: 0.025, 0.075, 0.25, 0.75, 2.5, 7.5, and 25 �g. Lanes 1 and 10 show a 100-bp ladder (Invitrogen). Thearrows indicate shifts.
1304 FONG ET AL. APPL. ENVIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 19
Nov
embe
r 20
21 b
y 69
.65.
92.5
1.

copy numbers appear to be necessary, but not sufficient, forhigh product titers.
Plasmid partition and transfer genes appear to increaseproduct titers by aiding the distribution of high-copy-num-ber plasmids through the mycelium population. A stabilityassay showed that high-producing plasmids were more sta-ble than low-producing plasmids. For example, pKOS011-26* was retained in nearly 100% of the population afterthree plating cycles, whereas pJRJ2 disappeared by thethird plating cycle. When pJRJ2 also contained parAB, theresulting high-producing plasmid, pRF169, remained in14% of the population after three plating cycles. Interest-ingly, the low-copy-number, low-producing pKOS011-26had stability similar to that of the high-copy-number, high-producing pKOS011-26* (near 100%). This observation in-dicates that copy number, and not plasmid stability, limitsthe product titers of low-copy-number plasmids.
Plasmid partition and transfer genes may increase producttiters by different mechanisms. The spd gene product is thoughtto transfer DNA between connected compartments withinStreptomyces hyphae (16). This function may increase in im-portance if passive distribution of plasmids during growth de-creases with increasing plasmid size. The parAB gene productsmay also facilitate the distribution of large plasmids at septumformation and into side branches. Additionally, these proteinsmay physically alter the structure of DNA to make large plas-mids more accessible to transcription. Plasmid replication,gene transcription, and partitioning functions all involve DNAsupercoiling (10, 31). A high replication rate may increaseplasmid supercoiling and hinder transcription of the biosyn-thetic genes, an effect that may be alleviated by parAB.
Plasmid distributions within the mycelium population haveimplications for the copy number in an individual compart-ment. For example, pRF123 and pRF126 had similar copynumbers of about 50 copies per chromosome. At the firstplating cycle of the stability assay, pRF123 existed in 44% ofthe population whereas pRF126 existed in 88%. Thus, if oneconsiders the copy number only in a plasmid-containing my-celial compartment, pRF123 must be on average twice thecopy number of pRF126. pRF126 differs from pRF123 only inthe presence of parAB, suggesting that parAB decreases plas-mid copy number while concomitantly increasing plasmid dis-tribution. This coupling of plasmid partitioning and replicationhas been observed with the E. coli plasmid pSC101 (19). Thus,product titers may depend critically on the fraction of hyphaein the fermentation that are productive. A technique such asfluorescence microscopy might determine plasmid concentra-tion in specific mycelial compartments and may help to confirmthis hypothesis.
DNA microarray analysis showed that high-producingstrains have significantly higher mRNA levels of the actII-ORF4 activator gene and eryA biosynthetic genes. SDS-PAGEshowed that these strains maintain the biosynthetic enzymesencoded by eryA at higher levels and for longer times. Thus,high product titers appear to arise from increased eryA expres-sion and higher enzyme concentrations. Experiments that var-ied the relative copy numbers of the activator and biosyntheticgenes showed that, in stable plasmids, both actII-ORF4 anderyA must replicate at high copy numbers to generate hightiters. The actII-ORF4 gene product appears to limit produc-
tion, since a ratio of actII-ORF4 to eryA greater than 1:1 in-creased product titers by at least 22%. However, actII-ORF4existed at a copy number 47% higher than those of the bio-synthetic genes in this higher-producing strain. This observa-tion may reflect a nonlinear relationship between plasmid copynumbers and product titers or a yield limit for the biosynthesisof the foreign natural product.
Our results possibly support an iteron mechanism of copynumber control for the SCP2@ replicon. The plasmids regu-lated by this mechanism contain repeated sequences in theirreplicons that bind to a plasmid-specific replication initiatorprotein (8). The arrangement of binding sites in the SCP2*replicon resembles that in other replicons controlled by iter-ons. The replication protein initiates replication at low plasmidconcentrations. At high concentrations of both the plasmid andreplication protein, the proteins physically couple the plasmidsto block replication. Previous studies have shown that the de-letion of iteron sequences increases plasmid copy number (27,28, 40). The 45-bp deletion in SCP2@ removed one of threedirect repeats of a consensus sequence in the replicon andabolished binding of RepI to that site. Furthermore, high RepIconcentrations produced a new, extremely high EMSA shiftwith a wild-type probe but failed to do so with a mutant probe.The additional shift may reflect coupled probes. Loss of RepIbinding may also reduce transcriptional control of the repIgene. Autoregulation of the rep gene has been observed foriteron plasmids such as R6K (12), P1 (9), and pSC101 (45, 47).The site removed by the 45-bp deletion resides 35 bp upstreamof repI, and the high-copy-number strain expressed repI andrepII at a level more than 10-fold higher than that of thelow-copy-number strain. Higher repI expression may increaseRepI concentrations and therefore replication rates and plas-mid copy numbers. Studies to delete other consensus sites,detect plasmid multimers, and detect plasmid coupling by RepIremain to be conducted to further determine whether theiteron model applies to SCP2@ replication.
Previously reported data suggest that RepII serves as a re-pressor of RepI (29, 30). The high-copy-number strain ex-pressed repI at a level 1.3- to 1.5-fold higher than that of repII,even though these two genes’ adjacent coding sequences sug-gest cotranscription (15). In contrast, the low-copy-numberstrain expressed these two genes at nearly the same level.These observations suggest an additional layer of copy numberregulation that involves altered concentrations of RepI andRepII. For example, smaller amounts of RepII or largeramounts of RepI may increase plasmid replication rates.
A small, practical cloning vector derived from the high-copy-number plasmid pBoost would aid efforts to overexpress entireantibiotic gene clusters in Streptomyces strains. In this work, wehave identified a minimal high-copy replicon consisting of tworeplication genes, repI and repII, and a 45-bp deletion up-stream of the repI gene. We have found that an intramycelialtransfer gene, spd, and overexpression of a partition gene,parAB, increase the stability of a large plasmid with the mini-mal replicon. We also found that high metabolite productionrequires six putative genes located between repII and spd. Fur-ther study of these genes may reveal their roles in elevating thecopy number of pJRJ2 so strongly or why the plasmid isunstable over time. An additional difference between thepRF127-derived plasmids and the pJRJ2-derived plasmids is
VOL. 73, 2007 CHARACTERIZATION OF A LARGE HIGH-COPY-NUMBER PLASMID 1305
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 19
Nov
embe
r 20
21 b
y 69
.65.
92.5
1.

the SCP2.01 and SCP2.02 genes. These two putative trans-posases are present identically in two copies on the Streptomy-ces genome. Based on this observation, and the results for ourlarge plasmid variant pRF89, SCP2.01 and SCP2.02 do notappear to be required in cis for plasmid stability or high pro-duction. The resulting high-producing plasmid, pRF169, stilldisappears from the mycelium population over several cellplating cycles, indicating that full plasmid stability requiresadditional SCP2* genes. This work has generated severalplasmids with useful experimental properties. For example,pRF103 and pRF63, which contain the minimal high-copy rep-licon, have successfully overexpressed single genes in S. coeli-color (22). These plasmids can tolerate a 2.1-kb insert withoutthe loss of stability, suggesting that they have some practicalutility in the laboratory.
ACKNOWLEDGMENTS
This work was supported in part by NSF grant BES-0093900-001 andNIH grant GM65470-02 to C.K. and NIH grant AI-08619 to S.C.
We greatly appreciate Sir David Hopwood for his valuable com-ments on the manuscript. We especially thank Josef Altenbuchner forkindly sharing his SCP2* replicon study results with us, without whichour own studies would not have been possible. We also thank JonathanVroom for his help with the EMSAs, Christine Miller for her advice onproject experiments, Kai Bao for his advice on the stability assays, andAmy Lum for her comments on the manuscript.
REFERENCES
1. Arias, P., M. A. Fernandez-Moreno, and F. Malpartida. 1999. Characteriza-tion of the pathway-specific positive transcriptional regulator for actinor-hodin biosynthesis in Streptomyces coelicolor A3(2) as a DNA-bindingprotein. J. Bacteriol. 181:6958–6968.
2. Ball, C. A., I. A. Awad, J. Demeter, J. Gollub, J. M. Hebert, T. Hernandez-Boussard, H. Jin, J. C. Matese, M. Nitzberg, F. Wymore, Z. K. Zachariah,P. O. Brown, and G. Sherlock. 2005. The Stanford Microarray Databaseaccommodates additional microarray platforms and data formats. NucleicAcids Res. 33:D580–D582.
3. Bibb, M., R. F. Freeman, and D. A. Hopwood. 1977. Physical and geneticalcharacterisation of a second sex factor, SCP2, for Streptomyces coelicolor.Mol. Gen. Genet. 154:155–166.
4. Bibb, M., J. L. Schottel, and S. N. Cohen. 1980. A DNA cloning system forinterspecies gene transfer in antibiotic-producing Streptomyces. Nature 284:526–531.
5. Bibb, M. J., and D. A. Hopwood. 1981. Genetic studies of the fertility plasmidSCP2 and its SCP2* variants in Streptomyces coelicolor A3(2). J. Gen. Mi-crobiol. 126:427–442.
6. Bradford, M. M. 1976. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dye bind-ing. Anal. Biochem. 72:248–254.
7. Brolle, D. F., H. Pape, D. A. Hopwood, and T. Kieser. 1993. Analysis of thetransfer region of the Streptomyces plasmid SCP2. Mol. Microbiol. 10:157–170.
8. Chattoraj, D. K. 2000. Control of plasmid DNA replication by iterons: nolonger paradoxical. Mol. Microbiol. 37:467–476.
9. Chattoraj, D. K., K. M. Snyder, and A. L. Abeles. 1985. P1 plasmid replica-tion: multiple functions of RepA protein at the origin. Proc. Natl. Acad. Sci.USA 82:2588–2592.
10. Conley, D. L., and S. N. Cohen. 1995. Effects of the pSC101 partition (par)locus on in vivo DNA supercoiling near the plasmid replication origin.Nucleic Acids Res. 23:701–707.
11. Fernandez-Moreno, M. A., J. L. Caballero, D. A. Hopwood, and F. Malpar-tida. 1991. The act cluster contains regulatory and antibiotic export genes,direct targets for translational control by the bldA tRNA gene of Streptomy-ces. Cell 66:769–780.
12. Filutowicz, M., G. Davis, A. Greener, and D. R. Helinski. 1985. Autorepres-sor properties of the pi-initiation protein encoded by plasmid R6K. NucleicAcids Res. 13:103–114.
13. Gramajo, H. C., E. Takano, and M. J. Bibb. 1993. Stationary-phase produc-tion of the antibiotic actinorhodin in Streptomyces coelicolor A3(2) is tran-scriptionally regulated. Mol. Microbiol. 7:837–845.
14. Haug, I. 2003. Ph.D. thesis. Streptomyces coelicolor A3(2) plasmid SCP2*:deductions from the complete sequence. University of Stuttgart, Stuttgart,Germany.
15. Haug, I., A. Weissenborn, D. Brolle, S. Bentley, T. Kieser, and J. Altenbuchner.
2003. Streptomyces coelicolor A3(2) plasmid SCP2*: deductions from the com-plete sequence. Microbiology 149:505–513.
16. Hopwood, D. A. 2006. Soil to genomics: the Streptomyces chromosome.Annu. Rev. Genet. 40:1–23.
17. Hu, Z., D. A. Hopwood, and C. R. Hutchinson. 2003. Enhanced heterologouspolyketide production in Streptomyces by exploiting plasmid co-integration.J. Ind. Microbiol. Biotechnol. 30:516–522.
18. Huang, J., C. J. Lih, K. H. Pan, and S. N. Cohen. 2001. Global analysis ofgrowth phase responsive gene expression and regulation of antibiotic bio-synthetic pathways in Streptomyces coelicolor using DNA microarrays. GenesDev. 15:3183–3192.
19. Ingmer, H., and S. N. Cohen. 1993. Excess intracellular concentration of thepSC101 RepA protein interferes with both plasmid DNA replication andpartitioning. J. Bacteriol. 175:7834–7841.
20. Jacobsen, J. R., C. R. Hutchinson, D. E. Cane, and C. Khosla. 1997. Pre-cursor-directed biosynthesis of erythromycin analogs by an engineeredpolyketide synthase. Science 277:367–369.
21. Kao, C. M., L. Katz, and C. Khosla. 1994. Engineered biosynthesis of acomplete macrolactone in a heterologous host. Science 265:509–512.
22. Karoonuthaisiri, N. 2004. Ph.D. thesis. Improving antibiotic production inStreptomyces coelicolor. Stanford University, Stanford, CA.
23. Karoonuthaisiri, N., D. Weaver, J. Huang, S. N. Cohen, and C. M. Kao.2005. Regional organization of gene expression in Streptomyces coelicolor.Gene. 353:53–66.
24. Kendall, K. J., and S. N. Cohen. 1988. Complete nucleotide sequence of theStreptomyces lividans plasmid pIJ101 and correlation of the sequence withgenetic properties. J. Bacteriol. 170:4634–4651.
25. Kieser, T., M. J. Bibb, M. J. Buttner, K. F. Chater, and D. A. Hopwood. 2000.Practical Streptomyces genetics. John Innes Foundation, Norwich, UnitedKingdom.
26. Kieser, T., D. A. Hopwood, H. M. Wright, and C. J. Thompson. 1982. pIJ101,a multi-copy broad host-range Streptomyces plasmid: functional analysis anddevelopment of DNA cloning vectors. Mol. Gen. Genet. 185:223–228.
27. Kittell, B. L., and D. R. Helinski. 1991. Iteron inhibition of plasmid RK2replication in vitro: evidence for intermolecular coupling of replication ori-gins as a mechanism for RK2 replication control. Proc. Natl. Acad. Sci. USA88:1389–1393.
28. Krishnan, B. R., and V. N. Iyer. 1990. IncN plasmid replicon. A deletion andsubcloning analysis. J. Mol. Biol. 213:777–788.
29. Larson, J. L., and C. L. Hershberger. 1986. The minimal replicon of astreptomycete plasmid produces an ultrahigh level of plasmid DNA. Plasmid15:199–209.
30. Larson, J. L., and C. L. Hershberger. 1984. Shuttle vectors for cloningrecombinant DNA in Escherichia coli and Streptomyces griseofuscus C581. J.Bacteriol. 157:314–317.
31. Liu, L. F., and J. C. Wang. 1987. Supercoiling of the DNA template duringtranscription. Proc. Natl. Acad. Sci. USA 84:7024–7027.
32. Livak, K. J., and T. D. Schmittgen. 2001. Analysis of relative gene expressiondata using real-time quantitative PCR and the 2(-Delta Delta C(T)) method.Methods 25:402–408.
33. Lombo, F., B. Pfeifer, T. Leaf, S. Ou, Y. S. Kim, D. E. Cane, P. Licari, andC. Khosla. 2001. Enhancing the atom economy of polyketide biosyntheticprocesses through metabolic engineering. Biotechnol. Prog. 17:612–617.
34. Lum, A. M. 2005. Ph.D. thesis. Reverse engineering industrial bacteria thatoverproduce antibiotics. Stanford University, Stanford, CA.
35. Lydiate, D. J., F. Malpartida, and D. A. Hopwood. 1985. The Streptomycesplasmid SCP2*: its functional analysis and development into useful cloningvectors. Gene 35:223–235.
36. Maas, R. M., J. Gotz, W. Wohlleben, and G. Muth. 1998. The conjugativeplasmid pSG5 from Streptomyces ghanaensis DSM 2932 differs in its transferfunctions from other Streptomyces rolling-circle-type plasmids. Microbiology144:2809–2817.
37. MacNeil, D. J., K. M. Gewain, C. L. Ruby, G. Dezeny, P. H. Gibbons, andT. MacNeil. 1992. Analysis of Streptomyces avermitilis genes required foravermectin biosynthesis utilizing a novel integration vector. Gene 111:61–68.
38. McDaniel, R., S. Ebert-Khosla, D. A. Hopwood, and C. Khosla. 1993. Engi-neered biosynthesis of novel polyketides. Science 262:1546–1550.
39. McDaniel, R., A. Thamchaipenet, C. Gustafsson, H. Fu, M. Betlach, and G.Ashley. 1999. Multiple genetic modifications of the erythromycin polyketidesynthase to produce a library of novel “unnatural” natural products. Proc.Natl. Acad. Sci. USA 96:1846–1851.
40. Park, K., E. Han, J. Paulsson, and D. K. Chattoraj. 2001. Origin pairing(�handcuffing’) as a mode of negative control of P1 plasmid copy number.EMBO J. 20:7323–7332.
41. Parro, V., D. A. Hopwood, F. Malpartida, and R. P. Mellado. 1991. Tran-scription of genes involved in the earliest steps of actinorhodin biosynthesisin Streptomyces coelicolor. Nucleic Acids Res. 19:2623–2627.
1306 FONG ET AL. APPL. ENVIRON. MICROBIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 19
Nov
embe
r 20
21 b
y 69
.65.
92.5
1.

42. Servin-Gonzalez, L., A. I. Sampieri, J. Cabello, L. Galvan, V. Juarez, andC. Castro. 1995. Sequence and functional analysis of the Streptomycesphaeochromogenes plasmid pJV1 reveals a modular organization of Strep-tomyces plasmids that replicate by rolling circle. Microbiology 141:2499–2510.
43. Uguru, G. C., K. E. Stephens, J. A. Stead, J. E. Towle, S. Baumberg, and K. J.McDowall. 2005. Transcriptional activation of the pathway-specific regulatorof the actinorhodin biosynthetic genes in Streptomyces coelicolor. Mol. Mi-crobiol. 58:131–150.
44. Vara, J., M. Lewandowska-Skarbek, Y. G. Wang, S. Donadio, and C. R.Hutchinson. 1989. Cloning of genes governing the deoxysugar portion of the
erythromycin biosynthesis pathway in Saccharopolyspora erythraea (Strepto-myces erythreus). J. Bacteriol. 171:5872–5881.
45. Vocke, C., and D. Bastia. 1985. The replication initiator protein of plasmidpSC101 is a transcriptional repressor of its own cistron. Proc. Natl. Acad. Sci.USA 82:2252–2256.
46. Xue, Q., G. Ashley, C. R. Hutchinson, and D. V. Santi. 1999. A multiplasmidapproach to preparing large libraries of polyketides. Proc. Natl. Acad. Sci.USA 96:11740–11745.
47. Yamaguchi, K., and Y. Masamune. 1985. Autogenous regulation of syn-thesis of the replication protein in plasmid pSC101. Mol. Gen. Genet.200:362–367.
VOL. 73, 2007 CHARACTERIZATION OF A LARGE HIGH-COPY-NUMBER PLASMID 1307
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/a
em o
n 19
Nov
embe
r 20
21 b
y 69
.65.
92.5
1.





























