Embed Size (px)
Citation preview

System. App!. Microbio!. 21, 50-64 (1998) SYSTEIVlATIC AND _©_G_us_ta_v _Fis_ch_e_f \C_er_la_g ________________ APPLIED MICROBIOLOGY
Characterization of Atypical Aeromonas salmonicida by Different Methods
B. AUSTIN\ D. A. AUSTIN\ 1. DALSGAAR02, B. K. GUOMUNosOOTTIR3, s. H0IE\]. M. THORN TONS, ]. L. LARSEN6,
B. O'HICI5 and R. POWELL5
1 Department of Biological Sciences, Heriot-Watt University, Edinburgh, Scotland 2 Fish Disease Laboratory, Danish Institute of Fisheries Research, Denmark 3 Institute for Experimental Pathology, University of Iceland, Iceland 4 Department of Fish Health, Central Veterinary Laboratory, Oslo, Norway 5 Department of Microbiology, University College Galway, Ireland 6 Department of Veterinary Microbiology, Laboratory of Fish Diseases, Royal Veterinary and Agricultural University, Frederiksberg,
Denmark
Received September 10, 1997
Summary
Fifty two isolates of atypical Aeromonas salmonicida, recovered from a wide range of hosts and geographical locations, were heterogeneous in terms of molecular and phenotypic characteristics, and represented taxa which could not be accommodated by the current classification of four subspecies. Generally, there was incongruence between the molecular (PCR, RAPD and ribotyping) and phenotypic methods in terms of cluster membership. By PCR, 6 groups were described of which Group 1 encompassed 12 isolates including the type strain of A. salmonicida subsp. smithia. Group 2 accommodated 23 isolates including the reference cultures of subspecies achromogenes and masoucida. The named culture of Haemophilus piscium was recovered in Group 6. By ribotyping and RAPD, the reference cultures were recovered in separate groups. All methods pointed to the uniqueness of subspecies smithia. Most isolates contained 2-6 plasmids, of 2.3 to 150 kb in length. Nevertheless, all isolates possessed certain key characteristics, including Gram-negativity, and the absence of motility.
Key words: Aeromonas salmonicida - Fish isolates - Fish pathogens - Identification - Taxonomy -Characterization - Typing methods
Introduction
By definition, Aeromonas salmonicida comprises fermentative, Gram-negative, nonmotile, nonencapsulated coccobacilli, which produce catalase and oxidase, and grow optimally at 22-25 °C (POPOFF, 1984). Four subspecies (achromogenes, masoucida, salmonicida and smithia) have been described (AUSTIN et a!., 1989; CARNAHAN and ALTWEGG, 1996). However, some authors have suggested different delineations within the species (McCARTHY and ROBERTS, 1980; BELLAND and TRUST, 1988), and new isolates that do not fit into the existing classification have been frequently reported (AUSTIN and AUSTIN, 1993; WIKLUND et. al., 1994; HIRVELA-KoSKI et a!., 1994). An increasing number of atypical isolates have been documented, usually on the basis of reduced or slow pigmentation (PEDERSEN et a!., 1994), slow growth (PEDERSEN et a!., 1994), nutritional fastidiousness (often for blood products; AUSTIN and ADAMS, 1996), growth at elevated temperatures (AUSTIN, 1993) or oxidase-negativity (PEDERSEN et a!.,
1994; WIKLUND et a!., 1994). Such isolates have been inevitably associated with ulcerations in cyprinids (e.g. goldfish ulcer disease and carp erythrodermatitis; AUSTIN and AUSTIN, 1993) and marine fish (NAKATSUGAWA, 1994; WIKLUND, 1995; WIKLUND and DALSGAARD, 1995; LARSEN and PEDERSEN, 1996). Infections attributed to atypical A. salmonicida are of increasing economic importance to aquaculture, worldwide (AUSTIN and AUSTIN, 1993). Furthermore, infections of wild fish stocks have been reported (WIKLUND and BYLUND, 1993).
Currently, it is unclear whether or not the so-called atypical isolates should be included in the existing subspecies or in as yet undescribed groupings within A. salmonicida. Moreover, it remains to be determined whether Haemophilus piscium, named by SNIESZKO et a!. (1950) as the causal agent of ulcer disease, should be reclassified with atypical A. salmonicida, as originally suggested by PATERSON et a!. (1980).

This study has sought to examine the taxonomic status of a collection of atypical isolates of A. salmonicida, using a range of genotypic and phenotypic methods. Inherent in the analysis was a secondary objective of evaluating the potential of the different methods as useful and informative utilitarian systems for dealing with the increased number of atypical isolates being reported. In recent years, nucleotide sequences of the genome of typical A. salmonicida subsp. salmonicida, which causes furunculosis in salmonids (AUSTIN and AUSTIN, 1993), have been determined including the gene encoding for the extracellular A-layer protein (GUSTAFSON et aI., 1992), the serine protease gene (WHITBY et aI., 1992), the glycerophospholipid: cholesterol acetyltransferase (GCAT) gene (NERLAND, 1996), and the gene encoding the 16S rRNA transcript (MARTINEZ-MuRCIA et aI., 1992). Based on these data, A. salmonicida - specific detection tests using the polymerase chain reaction (PCR) have been described using PCR primers designed to target the A-layer protein (GUSTAFSON et aI., 1992) and 16S rDNA (H0lE et aI., 1996). The use of PCR targeting several genes within the genome of A. salmonicida subsp. salmonicida, on atypical strains, might provide both a useful taxonomic and diagnostic tool. In total, this study used PCR, ribotyping, random amplification of polymorphic DNA (RAPD), plasmid profiling, biochemical analysis, examination of extracellular products (ECP), and pathogencity with the twin objectives of clarifying current A. salmonicida taxonomy and identifying useful diagnostic tests for this taxon and its sub-taxa.
Materials and Methods
Examination of cultures: Fifty two cultures conforming with the general description of A. salmonicida (POPOFF, 1984) but regarded as comprising atypical isolates, obtained from 26 species of fish in 13 countries, plus the type strains of A. salmonicida subspecies achromogenes, masoucida, salmonicida and smithia, and a reference culture of Haemophilus piscium were examined (Table 1). These were maintained in 15-25% (w/v) glycerol at -70, -80 or -140°C, and as plate cultures at room temperature on tryptone soya agar (TSA; Oxoid, Basingstoke, England), brain heart infusion agar (BHIA; Oxoid) and blood agar base (BA; Gibco, Paisley, Scotland) with 5% (w/v) blood (bovine or sheep). Routinely, purity and authenticity were checked by the examination of Gram-stained smears (HUCKER and CONN, 1923), the absence of motility in wet preparations examined at magnifications of x400 and xl000, and whole cell agglutination reactions using polyclonal antiserum to A. salmonicida subsp. salmonicida (titre = 1:8192) produced in a female white New Zealand rabbit.
Preparation of bacterial DNA for PCR: The bacteria were grown aerobically at 22°C in 5 ml volumes of heart infusion broth (HIB; Difco, Michigan, USA) from stock cultures kept frozen at -70°C. One ml of bacterial culture was centrifuged at 12000xg for 2 min. The pellet was resuspended in 100 pi PCR buffer [50 mM KCl, 10 mM Tris-HCI pH 8.0,2.5 mM MgCI2,
0.1 % (w/v) gelatin, 0.45% (w/v) NONITED P40, 0.45% (w/v) Tween 20] (HIGUCHI, 1989) and 2 pi of proteinase K (Sigma, Missouri, USA) [2.5% (w/v) in H20]. The samples were placed in a heating block (Dri-Block, Techne Inc., Princeton Nj, USA) at 56°C for 1 h, and subsequently ultrasonicated in a water-
Atypical Aeromonas salmonicida 51
bath (Transsonic, Elma, Germany) for 10 min at the same temperature. Another 2 pi of proteinase K were added to the samples, followed by further incubation at 56°C for 1 h. The temperature was increased to 95°C for 10 min, and the samples were subsequently cooled on ice. Centrifugation was at 12000xg for 10 s. Volumes (5 pi) of the supernatants were used as the template for PCR amplification.
PCR: All the primers were synthesized at a commerciallaboratory. The protein A (AP) and 16S rDNA primers, including their amplification conditions, have been described by GUSTAFSON et al. (1992) and H0lE et al. (1996), respectively. The serine protease (Sprot) and GCAT primers were designed on the basis of published sequences, using the computer program OUGO© (WHITBY et aI., 1992; NERLAND, 1996). The primer sequences were: Sprot, 5'-CGG CAC ACC GCA CCG AAG TA-3' and 5'GGC TCC CAT CCC TCC AGA CC-3', GCAT, 5'-ACC ACC TGC TTC ACA TCA AT-3' and 5'-CAA ACG GCT TCC ACA CAT AG-3'. The length of the PCR products of the Sprot and GCAT primers were 1009 and 773 bp, respectively.
Samples were subjected to PCR in a total volume of 50 pI. Each sample (5 pi) was added to 45 pi of a mixture of PCR reagents, yielding a final concentration of 10 mM Tris-HCI pH 8.3, 50 mM KCl, 0.01 % gelatin, 200 pM of each of the deoxyribonucleosides, 1 pM of each primer and IU of thermostable DNA polymerase (Taq DNA polymerase, Boehringer Mannheim, Germany). The Mg-concentration for the 16S rDNA primers was 3.0 mM, whereas the Mg-concentration for the AP, Sprot and GCAT primers was 2.0 mM. Each PCR set up included a negative control sample of distilled water and a positive control sample of A. salmonicida subsp. salmonicida DNA dissolved in water. PCR amplification was performed in a thermocycler (Biometra Biomedizinische Analytik GmbH, Gottingen, Germany) by initial denaturation at 95°C for 3 min, then 30 cycles of 90 s of denaturation at 95°C, 1 min of primer annealing at temperatures varying with primer sets, and 90 s of extension at 72°C. After the last cycle, 7 min at 72 °C was used. The annealing temperatures for the 16S rDNA, AP, Sprot and GCAT primers were 54, 57, 65 and 55°C, respectively. The PCR products (8 pi) were loaded on horizontal 1.0% agarose gels, and subjected to electrophoresis for 35 min at 100 V in Tris-borate buffer (SAMBROOK et aI., 1989). Gels were stained with ethidium bromide, and photographed under UV-light transillumination. A 123 base pair DNA ladder (Gibco BRL, Life Technologies Inc., MD, USA) was included as a molecular weight standard on each gel.
Ribotyping: Bacteria were grown in a shaking incubator for 48-72 h at 20°C in Veal Infusion Broth (VIB; Difco). DNA was extracted from pellets, as described by PEDERSEN and LARSEN (1993), and cleaved for >3 h at 37°C with HindIIl, EeaRI and PstI (Promega, Madison, Wisconsin, USA). Restriction fragments were separated electrophoretically through 0.8% (w/v) agarose (Litex, Denmark) in Tris-acetate buffer (40 mM Tris, 1 mM EDTA buffer; pH 8.0) for 18-20 h at a voltage of 25 V, before transfer to nylon membranes (Magna Nylon, 0.45, Frisinette, Denmark) using a VacuGene System (Pharmacia, Uppsala, Sweden). The DNA was fixed (30 min at 80°C) before hybridization overnight at 56 °C with a digoxigenin labelled DNA probe complementary to 16S and 23S rRNA. After blocking, the membranes were incubated at room temperature in a shaking incubator for 30 min with alkaline phosphatase labelled anti-digoxigenin. Signal development was performed in 0.1 M Tris HCI, 0.1 M NaCI, 50 mM MgCI2; pH 9.5, after addition of nitro blue tetrazolium (Boehringer) and 5-bromo-4-chloro-3-indolyl phosphate (Boehringer) as substrate. Purpleblue bands appeared within a time span of a few minutes to one hour. A 1 kb DNA ladder (Gibco BRL) was used as a molecular size standard.

Tab
le 1
. B
acte
rial
cul
ture
s
Lab
. N
ame
as r
ecei
ved
ref.
no.
AS
6 at
ypic
al A
. sa
lmon
icid
a
AS
24
atyp
ical
A.
salm
onic
ida
AS
54
atyp
ical
A.
salm
onic
ida
AS
109
atyp
ical
A.
salm
onic
ida
AS
l12
at
ypic
al A
. sa
lmon
icid
a
AS
128
atyp
ical
A.
salm
onic
ida
AS
130
atyp
ical
A.
salm
onic
ida
AS
132
atyp
ical
A.
salm
onic
ida
AS
176
atyp
ical
A.
salm
onic
ida
AS
203
atyp
ical
A.
salm
onic
ida
AS
204
atyp
ical
A.
salm
onic
ida
AS
208
A.
salm
onic
ida
subs
p.
smit
hia
AS
16
atyp
ical
A.
salm
onic
ida
AS
28
atyp
ical
A.
salm
onic
ida
AS
46
atyp
ical
A.
salm
onic
ida
AS
47
atyp
ical
A.
salm
onic
ida
AS
62
atyp
ical
A.
salm
onic
ida
AS
78
atyp
ical
A.
salm
onic
ida
AS
81
atyp
ical
A.
salm
onic
ida
AS
94
atyp
ical
A.
salm
onic
ida
AS
I02
at
ypic
al A
. sa
lmon
icid
a
AS
I03
at
ypic
al A
. sa
lmon
icid
a
AS
121
atyp
ical
A.
salm
onic
ida
AS
135
atyp
ical
A.
salm
onic
ida
AS
145
atyp
ical
A.
salm
onic
ida
AS
163
atyp
ical
A.
salm
onic
ida
AS
167
atyp
ical
A.
salm
onic
ida
AS
18
2
atyp
ical
A.
salm
onic
ida
AS
185
atyp
ical
A.
salm
onic
ida
AS
200
atyp
ical
A.
salm
onic
ida
AS
201
atyp
ical
A.
salm
onic
ida
AS
217
A.
salm
onic
ida
sub
sp.
achr
omog
enes
Col
lect
iona
/
stra
in n
o.
OLI
VIE
R;
F1
54
2-2
OLI
VIE
R;
Fin
-7
86
09
02
12
0
WIK
LUN
D;
2F
7-1
90
(T
6)
WIK
LUN
D;
8F
12
-38
11
(T
19)
93
07
19
-1/3
0
92
07
20
-2/4
MK
I03
51
27
/88
V1
79
CC
M4
10
3T
OLI
VIE
R;
87
48
0
OLI
VIE
R;
3.1
11
88
09
02
22
1
88
09
02
77
8
88
09
03
37
7
93
09
00
91
4
93
09
01
39
2
87
06
26
-1/1
a
84
10
18
-1/2
84
11
16
-3/1
92
05
01
-3/1
86
06
25
-4/3
M2
83
/89
POH
JAN
VIR
TA;
TP
-9
POH
JAN
VIR
TA;
TP
-121
18
4/7
6
M2
83
/89
BEN
EDIK
TSD
OTT
IR; T
5/9
2
GR
OM
AN
; B
A17
4
AT
CC
l92
6F
Sou
rce
and
/or
cou
ntr
y
of
orig
in
Atl
anti
c sa
lmo
n,
No
rway
bro
wn
tro
ut,
Fin
lan
d
tro
ut,
No
rway
flou
nder
, F
inla
nd
flou
nder
, F
inla
nd
dab
, Ic
elan
d
turb
ot,
Den
mar
k
turb
ot,
No
rway
Arc
tic
char
, F
inla
nd
wol
ffis
h, S
cotl
and
wra
sse,
Sco
tlan
d
roac
h,
En
gla
nd
Atl
anti
c sa
lmo
n,
Can
ada
gold
fish
, U
SA
char
, N
orw
ay
turb
ot,
No
rway
Atl
anti
c sa
lmo
n,
No
rway
cod
, N
orw
ay
wra
sse,
No
rway
blen
ny,
Den
mar
k
sand
eel,
Den
mar
k
sand
eel,
Den
mar
k
dab
, G
erm
an B
ight
Atl
anti
c sa
lmo
n,
Far
oes
Atl
anti
c sa
lmo
n,
Icel
and
gray
ling
, F
inla
nd
wh
ite
fish
, F
inla
nd
gold
fish
, It
aly
Atl
anti
c sa
lmo
n,
Icel
and
whi
ting
, Ic
elan
d
wol
ffis
h, C
anad
a
sea
tro
ut,
Sco
tlan
d
PC
R
Rib
og
rou
p
type
1 6
1 6
1 6
1 9
1 9
1 2
5
1 1
0
1 1
0
1 1
7
1 6
1 1 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2
15
2 18
29
13 3
14
3 3 8 8 8
11 3 3
12
12
31 3 3 6 3
Ph
eno
n
1 1 3 1 6 6 6 6 5 2 2 4 7 5 5 7 6 5 1 1 5 1 6 7 5 2 7 5 5 5 3 5
Pro
teas
e g
rou
p
2 Sa
5b
4 2 5b
4 2 5b
2 2 1 2 3 5b
2 3 2 3 2 2 2 4 2 2 5b
5b
5b
2 2 2 2
Pla
smid
co
nte
nts
(kb
)
8.1
,27
,36
,45
,96
5.8
,10
,45
, 1
50
8.1
,36
6.4
,21
,27
,54
13
,27
,64
4.7
,64
10
, 7
4
5.8
,10
,27
, 7
4
10
,27
,15
0
10
, 1
30
5.8
,10
, 1
30
6.4
, 7
4,
15
0
64
5.8
,27
,45
,85
13
,85
4.7
,5.2
,21
,45
,11
0
6.4
,74
4.7
,5,2
,54
,11
0
Pat
hoge
nici
ty
+ (
R)
+ (
A)
+ (
T)
+(A
,R)
+(A
,R.T
)
+ (
A,R
)
+ (
A,R
, T
)
+(A
,G.R
)
+(G
)
+(A
,G,R
)
+(A
,R)
+(A
,G,R
)
+ (
A,R
)
5.2
,6.4
,10
,15
0
5.2
,8.1
,36
,74
,13
0
5.8
,8.1
,27
,36
,45
,11
0
+ (
A,
R,
T)
5.8
,8.1
,27
,36
,64
,11
0
-
5.8
,54
,64
+
(A
, R
)
4.7
,5.2
,54
,96
4.7
,5.2
,27
,54
,96
27
,11
0
45
,64
5.8
,27
,45
, 6
4
4.7
,5.2
,54
,96
27
,96
10
,13
0
4.7
,21
+(A
,G,R
)
+(A
,G,R
)
+(A
,R)
+ (
A,
G,
R,
T)
+(A
,G,R
)
+(A
,G,R
)
+(A
,G,R
)
+(A
,G,T
)
+ (
A)
v,
N s:o > c: :3 z ~ ~

AS
218
A.
salm
onic
ida
subs
p.
AT
CC
27
01
3T
m
aso
u s
alm
on
, Jap
an
2 4
3 1
13
,85
ma
sou
cid
a
AS
222
aty
pic
al A
. sa
lmon
icid
a B
ERN
OTH
; R
N 8
5:1
92
3-A
go
ldfi
sh,
Au
stra
lia
2 7
7 5
b
5.8
,16
,36
,64
AS
224
atyp
ical
A.
salm
onic
ida
BER
NO
TH;
WN
87
:44
2-E
go
ldfi
sh,
Aus
tral
ia
2 7
7 5
b
5.8
,27
,64
+
(G)
AS4
at
yp
ical
A.
salm
onic
ida
OLI
VIE
R;
813
77
A
tlan
tic
cod,
Can
ada
3 6
7 5
a 5
.8,1
0,4
5,1
50
AS
42
atyp
ical
A.
salm
onic
ida
89
09
02
40
3
Atl
anti
c sa
lmo
n,
No
rway
3
6 1
2 8
.1,2
7,4
5,1
50
+
(A,R
, T
)
AS4
8 at
yp
ical
A.
salm
onic
ida
90
09
00
11
5
wra
sse,
No
rway
3
28
2 5
b
5.2
,6.4
, 10
, 1
50
AS
69
aty
pic
al A
. sa
lmon
icid
a 8
70
90
23
03
A
tlan
tic
salm
on
, N
orw
ay
3 3
7 3
5.2
,6.4
,10
, 1
50
+
(A)
AS
74
aty
pic
al A
. sa
lmon
icid
a 9
20
90
17
77
sp
ott
ed w
olff
ish,
No
rway
3
6 4
2 5
.2,
16
,85
+
(A
,G,
R,
T)
AS
83
atyp
ical
A.
salm
onic
ida
93
09
01
50
5
wra
sse,
No
rway
3
3 1
3 5
.2,6
.4,
10
, 1
50
+
(A
,R)
AS
89
aty
pic
al A
. sa
lmon
icid
a C
AR
SON
; 9
40
90
10
74
fl
ound
er,
Tas
man
ia
3 2
6
4 5
b
6.4
,74
+
(A,R
, T
)
AS
93
aty
pic
al A
. sa
lmon
icid
a 92
1113
-1I1
Oa
plai
ce,
Den
mar
k
3 16
6
4 7
,27
,36
,74
,96
+
(R)
AS
219
A.
salm
onic
ida
subs
p.
AT
CC
14
17
41
bro
ok
tro
ut,
USA
3
5 3
4 5
.2,7
,85
salm
onic
ida
AS
I0
aty
pic
al A
. sa
lmon
icid
a O
LIV
IER
; M
T-3
73
Atl
anti
c sa
lmon
, S
cotl
and
4
3 5
2 4
.7,5
.2,2
1,2
7,1
10
+
(A
, G
,R,
T)
AS5
1 at
yp
ical
A.
salm
onic
ida
76
09
00
85
6
tro
ut,
No
rway
4
30
5
5b
2
.3,4
5,
64,
74
+
(A
,R)
AS
59
aty
pic
al A
. sa
lmon
icid
a 9
00
90
34
41
ch
ar,
No
rway
4
22
1
5b
1
3,2
7,3
6,8
5
+ (
A,R
, T
)
AS
122
aty
pic
al A
. sa
lmon
icid
a 9
20
50
1-3
/3
dab
, D
enm
ark
4
11
6 4
5.8
,54
,74
AS
19
atyp
ical
A.
salm
onic
ida
OLI
VIE
R;
91
54
9
eel,
Can
ada
5 21
7
5b
+
(A
)
AS
30
atyp
ical
A.
salm
onic
ida
OLI
VIE
R;
V23
4/81
ca
rp,
Th
e N
eth
erla
nd
s 5
19
7 5
a 1
6,3
6,4
5,6
4,1
50
+
(G)
AS
I08
at
ypic
al A
. sa
lmon
icid
a W
IKLU
ND
; 11
411
(Tl)
fl
ound
er,
Fin
lan
d
5 9
6 4
6.4
,21
, 36
, 4
5
AS
209
atyp
ical
A.
salm
onic
ida
wol
ffis
h, S
cotl
and
5 2
7
5 4
5.2
, 7,
54
+
(A
,R,
T)
AS
221
atyp
ical
A.
salm
onic
ida
AF
HR
L 1
go
ldfi
sh,
Aus
tral
ia
5 7
7 5
b
13
,13
0
+ (
A)
AS1
at
ypic
al A
. sa
lmon
icid
a O
LIV
IER
; S
AB
-l
sabl
e fi
sh,
Can
ada
6 2
0
4 1
5.2
,5.8
AS
60
aty
pic
al A
. sa
lmon
icid
a 9
00
90
27
17
tu
rbo
t, N
orw
ay
6 2
4
3 4
5.2
,10
+
(A,R
,T)
AS
92
aty
pic
al A
. sa
lmon
icid
a 9
21
13
-l/3
a pl
aice
, D
enm
ark
6
23
6
5a
4.7
,21
,36
,74
AS
239
Ha
emo
ph
ilu
s pi
sciu
m
NC
IMB
19
52
U
SA
6 1
5 5
4,
74
1 ty
pe s
trai
n.
d S
ourc
es:
AF
HR
L,
Au
stra
lian
Fis
h H
ealt
h R
esea
rch
Lab
ora
tory
; A
TC
C,
Am
eric
an T
ype
Cu
ltu
re C
olle
ctio
n, R
ockv
ille
, M
ary
lan
d;
CC
M,
Cze
cho
slo
vak
ian
Co
llec
tio
n o
f M
i-cr
oo
rgan
ism
s, B
rno,
Cze
chos
lova
kia;
NC
IMB
, N
atio
nal
Col
lect
ion
of
Indu
stri
al a
nd
Mar
ine
Bac
teri
a, A
berd
een,
Sco
tlan
d; D
r. E
BEN
EDIK
TSD
OTT
IR,
Inst
itut
e o
f B
iolo
gy,
Uni
vers
i-ty
of
Icel
and;
Dr.
E.-
M.
BER
NO
TH C
SIR
O A
ustr
alia
n A
nim
al H
ealt
h L
abor
ator
y, G
eelo
ng,
Vic
tori
a, A
ustr
alia
; D
r. J.
CA
RSO
N,
Mo
un
t P
leas
ant
Lab
ora
tori
es,
Kin
gs M
ead
ow
s, T
as-
man
ia,
Aus
tral
ia;
Dr.
G.
OLI
VIE
R,
Dep
artm
ent
of
Fis
heri
es a
nd
Oce
ans,
Hal
ifax
Fis
heri
es R
esea
rch
Lab
ora
tory
, H
alif
ax,
Can
ada;
Dr.
D.
GR
OM
AN
, A
tlan
tic
Vet
erin
ary
Col
lege
, U
nive
rsit
y o
f P
rinc
e E
dw
ard
s Is
land
, C
anad
a; D
r. T
. PO
HJA
NV
IRTA
, N
atio
nal
Vet
erin
ary
and
Fo
od
Res
earc
h In
stit
ute,
Reg
iona
l L
abo
rato
ry,
Ku
op
io,
Fin
land
; D
r. T
. W
IKLU
ND
, In
-st
itut
e o
f P
aras
itol
ogy,
Ab
o,
Fin
land
. A
, G
, R
an
d T
co
rres
po
nd
to
Atl
anti
c sa
lmon
, go
ldfi
sh,
rain
bo
w t
rou
t an
d t
urb
ot,
res
pect
ivel
y.
2;- '<
~ n ~
>- '" .... o ;:: o ;:s
;::, '" '" ;::, ~
o ;:s ;:;. ~
v.
w

54 B. AUSTIN et al.
Genomic DNA extraction for RAPD analysis: Cells were harvested from 10 ml cultures by centrifugation (Centrifuge 5416, Eppendorf, Germany), and genomic DNA extraction was performed using a modification of a previously described method (AusuBEL et aI., 1992). Briefly, the bacterial pellets were resuspended and washed in 1 ml of sterile distilled water, followed by centrifugation for 5 min at 9650xg (MSE Micro Centaur, Sanyo, UK), and resuspension in 400 pi TE buffer (10 mM Tris HCI, 1 mM EDTA, pH 8.0). The cells were lysed by the addition of 200 pI of lysozyme (10 mg/ml) (Boehringer) and incubation at 37°C for 60 min. Then, the preparation was incubated for 10 min with 40 pI of proteinase K (10 mg/ml) (Boehringer) at room temperature, followed by the addition of sodium dodecyl sulphate (SDS) to a final concentration of 1.0%, and incubation at room temperature until the preparation was clear. For many of the atypical strains, the preparation was further incubated at 60°C for 5 min to ensure complete lysis. Eighty pI of 0.5 M EDTA were added, mixed by gentle agitation, and the solution was deproteinised by sequential phenol and chloroform-isoamyl alcohol (24:1 voVvol) extraction. The genomic DNA was precipitated in ethanol and resuspended in TE buffer. The DNA concentration was estimated by visual comparison with standard DNA size markers after electrophoresis through a 1 % agarose (Bio/Gene Ltd., Cambridge, England) TAE gel stained with 0.5 pg/ml ethidium bromide (Sigma).
RAPD analysis: The previously described RAPD primer, H3, 5'-CGCGCCGG-3', was used (CAETANO-ANOLLES et aI., 1991). Amplification was performed using a hot bonnet thermocycler (Omnigene TR3 CM220, Hybaid Ltd., Middlesex, England) with no overlay of mineral oil. The 50 pI reaction mixture contained lOx NH4 buffer (Bioline U.K.), 4.0 mM MgCI2, 200 pM of dNTPs (Pharmacia), 2 U Taq DNA Polymerase (Bioline), 60-80 pmol of primer and 20 ng of genomic DNA. Negative control reaction mixtures lacking DNA template were also included. Amplification conditions were 35 cycles of 10 s at 94°C, 30 s at 37°C, 60 s at 72°C, and a final extension of 5 min at 72 dc. Five pI aliquots of the amplification products were analysed by electrophoresis through 1 % agarose (Bio/Gene), 1 % NuSieve (FMC Bioproducts, Rockland, USA) TAE gels stained with 0.5 pg/ml ethidium bromide and visualized over UV using the Bio-Profil imaging system (Vilber Lourmat, Marne La Vallee, France). The 100 bp DNA ladder (Gibco BRL) standard DNA size markers were included in each electrophoresis.
RAPD cluster analysis: The lengths of the RAPD generated DNA fragments were determined using the Bio-Gene software package (Vilber Lourmat) on the Bio-Profil imaging system. Each isolate was then visually coded for the presence (coded 1) or absence (coded 0) of each DNA fragment and combined into one data matrix which was then analysed using the inverse of the Jaccard (SJ) coefficient as previously described (MORGAN et aI., 1993). The similarity dendrograms were generated using the unweighted average linkage (UPGMA) clustering method of the PHYLIP phylogeny inferance package (FELSENSTEIN, 1993).
Phenotypic tests: Cultures, grown in VIE for 48 h, were used to inoculate the test media. When carry-over of nutrient might influence the results, cells from the top of colonies on BA with 5% (w/v) bovine blood were used. Unless indicated otherwise, the methods and media were taken from DALSGAARD et al. (in press). The strains were incubated at 20°C, and the tests were read daily during an incubation period of 14 days. The following biochemical tests were employed: cytochrome oxidase, catalase, glucose fermentation, gas from glucose, arginine dihydrolase, lysine decarboxylase, ornithine decarboxylase, indole production, aesculin hydrolysis, acid production from sucrose, salicin, sorbitol, cellobiose and glycerol, and susceptibility to
ampicillin and cephalothin. In addition, haemolysin production was assessed on bovine blood agar plates, and pigment production on TSA.
Cultures were also examined by the API 20NE and API ZYM rapid identification systems (BioMerieux, Marcy-I'Etoile, France). Thus, inocula were prepared from 24-28 h cultures incubated at 20°C on TSA. Cells were harvested in 0.9% (w/v) sodium chloride, and the suspensions adjusted with fresh saline to achieve a standard opacity equivalent to McFarland No.3. The API 20NE rapid identification systems were inoculated according to the manufacturers recommendations, with incubation at 25°C for 24 and 48 h.
Preparation of extracellular products (ECP) and whole cells: The cellophane overlay method, as described by GUDMUNDSDOTTIR (1996), using BHIA supplemented with 5% (w/v) newborn calf serum (Gibco) at 22°C for 72-144 h, was used for cultivation of the bacterial isolates. The bacteria and ECPs were washed-off the cellophane overlay with minimal amounts of phosphate buffered saline (PBS, pH 7.2). Following centrifugation (2000xg for 30 min), the supernatant (ECP) was filtered (Millipore 0.22 pm porosity filters) and stored at -80°C, until required. Cells were killed with formalin [to a final concentration of 2% (w/v) formaldehyde] and incubated at 22°C for 24 h. Killed cell suspensions were washed three times in PBS, resuspended in fresh PBS and sonicated for 5 min at an amplitude of 14-18 pm. The protein concentration was measured with the BioRad protein assay, using bovine serum albumin as a standard.
Detection of A-layer protein and LPS O-antigen by immunoblotting: The SDS-PAGE separation of whole cell preparations (containing Img protein/ml) for the detection of A-layer protein and the same preparations digested by proteinase K for detection of lipopolysaccharide (LPS) was carried out using the Mini-Protean II system (BioRad), according to the manufacturer's instructions. Ten pI samples, heated for 10 min at 100°C in 5% (w/v) mercaptoethanol in 125 mM Tris buffer, pH 6.8, containing 2 % SDS, were applied to a 4.5% stacking gel and a 14% separation gel. Molecular weight markers were prestained low range standards from BioRad. Proteins separated by SDS-PAGE were transferred to a nitrocellulose membrane by electrophoresis at 100 V and 250 rnA for 90 min. The buffer used for transfer contained 25 mM Tris, 192 mM glycine and 20% (v/v) methanol, pH 8.8. Following transfer, the membranes were blocked (1 hat 22°C) with 1 % BSA, and 1 % normal pig serum in PBS containing 0.1 % Tween-20 (PBS-T). Primary antibodies, as well as the alkaline phosphatase labelled murine IgG, were diluted in PBS-T containing 0.2% BSA and 0.2% pig serum. Incubation with primary antibody was at 4 °C overnight and with the conjugate for 1 h at room temperature. The blots were washed extensively in PBS-T between each step. Monoclonal murine antibodies (provided by S. Espelid, FORUT, Tromso, Norway) to the A-layer protein and to the O-polysaccharide component of LPS of A. salmonicida subsp. salmonicida, diluted 1:1000, were used as primary antibodies. Immunoreactive bands were visualised by incubating membranes in 0.1 M carbonate buffer (pH 9.8) containing 0.3 mg/ml of p-nitro blue tetrazolium chloride (NBT; Sigma) and 0.15 mg/ml of 5-bromo-4-chloro-3 indolyl phosphate toluidine salt (BCIP; Sigma).
Analysis of ECP activities: • Haemolytic titration: Haemolytic activity of ECP against
horse and salmon red blood cells (RBC) was estimated, as previously described (GUDMUNDSDOTTIR et aI., 1990). One hundred pI two-fold serial dilutions of ECP in PBS were added to 100 pI of 1 % (v/v) RBC suspension in PBS, and incubated for 20 h at 22°C. PBS was used instead of the enzyme solution as a nega-

tive contro!' One unit of haemolytic activity (HU) was defined as the dilution causing 50% haemolysis.
• GCAT assay: GCAT activity was detected by the method described by LEE and ELLIS (1990). Phospholipase activity was determined by applying 20 rl samples into wells cut in 1 % (w/v) agarose in PBS containing 1 % L-a lecithin (Sigma) and incubating at 22°C for 48 h. The appearance of transparent zones around a well in the agar indicated a positive phospholipase reaction. GCAT activity of ECP was determined as a positive phospholipase reaction together with pronounced haemolytic activity.
• Caseinase assay: Caseinase activity of ECP was determined after GUDMUNDSDOTTIR (1996) using azocasein (Sigma) as a substrate. Fifty rl of enzyme sample were incubated with 450 rl of 1 % (w/v) azocasein in 0.06 molll phosphate buffer (pH 7.2) at 22 °C for 60 min. In reagent blanks, PBS was used instead of enzyme sample. The reaction was stopped by adding 0.5 ml of 10% (w/v) trichloroacetic acid (TCA). After 30 min, the precipitate was pelleted by centrifugation, and a 0.5 ml aliquot of the supernatant added to 0.5 ml of 1 molll NaOH. Released azodye was measured spectrophotometrically at 450 nm (A45o) against a reagent blank. The assay was performed in triplicate. One caseinase unit was defined as an increase of 0.001 in A450 under the assay conditions.
Gelatinase assay: Gelatinase detection was performed by a radial diffusion method, as described by GUDMUNDSD(lTTIR (1996), using 3% gelatin (Difco) in 1 % agarose gels. Twenty rl volumes of ECP were placed in 4 mm diameter wells cut into the gels, and incubated at 22 °C for 20 h. After incubation, the plates were immersed in a saturated ammonium sulphate solution at 70°C to precipitate unhydrolysed gelatin. The diameter of the zone of proteolysis around the inoculated wells was measured against a black background. Proteolytic activity was determined from a standard curve using trypsin (bovine pancreas type Ill, Sigma). One unit of gelatinase activity was defined as equivalent to that of 1 rg trypsin.
• Protease inhibition: Solutions of 0.025 M phenyl methyl sulphonyl fluoride (PMSF) in ethanol and 0.05 M 1 10-phenanthroline (OPA) in ethanol were freshly prepared. Equal volumes of enzyme and inhibitor solutions were mixed and incubated at 22°C for 10 min prior to proteolytic assay. Controls contained only solvent. The inhibition was deemed positive if > 1 0% of the activity was reduced by treatment.
• Substrate SDS-PAGE: Analysis of proteases by substrate gel electrophoresis was performed according to a modified procedure of HEUSSEN and DOWDLE (1980). Electrophoresis was carried out using 14% gels supplemented with 0.1 % substrate from 10% stock solutions, prepared in distilled water. No substrate was added to the stacking gels. After electrophoresis, the remaining steps were carried out at room temperature. In order to remove SDS, the gels were incubated for 1 h in 2.5% Triton X-100 in 20 mM Tris-HCI, pH 8.0, with one change after 30 min. Then, the gels were incubated for 4 h in 20 mM TrisHCl,2 mM CaCI2, pH 8.0, with one change after 30 min. Gels were stained with Coomassie Brilliant Blue (Pharmacia) and destained until bands of clearing appeared at the migration distance of the respective proteases. Prestained SDS-PAGE standards (BioRad) were used to estimate the apparent molecular weight (AMW) of the proteases (HEUSSEN and DOWDLE, 1980).
Analyses of phenetic data: Phenetic data were compared by the simple matching (SS\1) and SJ coefficients, and by Euclidean Distance, with clustering by UPGMA and Wards method (SNEATH and SaKAL, 1973; PRIEST and AUSTIN, 1993), using NTSYS (Applied Biostatistics, USA) and STATISTICA (StatSoft, USA).
Plasmid profiling: Cultures, grown for 48-72 h at 20°C in VIB, were used for the determination of plasmid profiles. Plas-
Atypical Aeromonas salmonicida 55
mid DNA was extracted from the bacteria by the method of KADO and LIu (1981). Plasmids were separated by gel electrophoresis at 80 V for 3 h in 0.8% agarose gels (SeaKem GTG; FMC, Bio Products, USA) in TAE buffer, at pH 8.0. The gels were stained in 2 rg/ml of ethidium bromide (Sigma) and photographed under ultraviolet light. The size of the plasmids was determined by the length of migration, according to the method of ROCHELLE et a!. (1985). All cultures were examined for plasmids at least twice.
Fish pathogenicity experiments: Bacterial cultures were grown overnight at 22°C in tryptone soya broth (Oxoid), centrifuged at 5,000xg for 10 min and resuspended in 10 ml volumes of 0.9% (w/v) saline to approximately 108 cells/m!. Ten fold dilutions were prepared to 102 cells/m!. Atlantic salmon (Salmo salar), goldfish (Carassius auratus), rainbow trout (Oncorhynchus mykiss) and turbot (Scophthalmus maximus) were used to assess pathogenicity, as follows:
Groups of 10 fish were infected by intraperitoneal (i.p.) or intramuscular (i.m.) injection with 0.1 ml volumes of the washed bacterial suspensions to achieve doses of 102-106
cellslfish. The infected animals were maintained for up to 14 days in covered polypropylene tanks supplied with dechlorinated, aerated static freshwater (the water was changed daily) or seawater, as appropriate, at a temperature of _15°C. Dead and moribund fish were removed, and subjected to standard bacteriological and pathological examination (AUSTIN and AUSTIN, 1989). Any survivors at the end of the experiment were sacrificed and examined, as above. The disease signs were recorded, and attempts made to recover and confirm the identity of the pathogen by the methods described by AUSTIN and AUSTIN (1993). The Lethal Dose 50% (LDso) was determined from mortality patterns using the statistical methods described in WARDLAW (1985).
Test reproducibility: Generally, the tests were carried out twice, and were repeated in the advent of discrepancies. Phenotypic tests of doubtful reliability were not included in the computer analyses .
Results
peR Groups
All the isolates were ascribed to 6 PCR groups (Table 1), as follows: Group 1 (with 12 isolates including the type strain of A. salmonicida subsp. smithia AS208) was defined on the basis of possession of PCR products generated from all the primer-sets, of sizes consistent with the expected fragment lengths. Group 2, with 23 isolates including subspecies achromogenes (AS217) and masoucida (AS218), was characterised by the presence of PCR products from the 16S rDNA, GCAT and AP primer-sets. Group 3 contained 9 isolates including A. salmonicida subsp. salmonicida (AS219), that amplified PCR products using the 16S rDNA, GCAT and Sprot primer-sets. Group 4 included 4 isolates that amplified PCR products using 16S rDNA and GCAT primer-sets. Group 5 comprised 5 isolates that amplified PCR products with only the 16S rDNA and AP primer-sets; and Group 6 (4 isolates including the named culture of H. piscium AS239) produced PCR products solely with the 16S rDNA primer-set. Generally, each PCR group included isolates from a variety of freshwater and marine fish, and a diverse range of geographical locations (Table 1).

12 k
b 10
kb
8 kb
6 kb
kb
2 kb
kb
o 1
2 3
5 6
8 9
10
11
12
13
1 15
16
17
18
19
20 2
1 22
23
24
25
26 2
7 28
29
30
3
Fig.
1.
Rib
otyp
e p
atte
rns
of
A.
salm
onic
ida.
Ch
rom
oso
mal
DN
A w
as d
iges
ted
wit
h H
indI
II.
Lan
e 0,
1 kb
DN
A l
add
er.
Th
e 31
dif
fere
nt p
atte
rns
are
in t
he f
ollo
win
g la
nes:
1,
AS
239;
2,
AS
208;
3,
AS
217;
4,
AS
218;
S,
AS
219;
6,
AS
6; 7
, A
S22
1; 8
, A
SI0
2;
9, A
S10
9;
10,
AS
130;
11,
AS
122;
12
, A
S16
7; 1
3,
AS
46;
14,
AS
62;
IS,
AS
204;
16,
AS
93;
17,
AS
176;
18
, A
S16
; 19
, A
S30
; 2
0,
AS
1; 2
1,
AS
19;
22
, A
SS9;
23,
AS
92
;24
, A
S60
; 2S
, A
S12
8;
26,
AS
89;
27,
AS
209;
28
, A
S48
; 2
9,
AS
28;
30
, A
SSl;
31
, A
S18
2.

Ribotyping
There was little relationship between the results of PCR grouping and ribotyping. The ribotyping divided the isolates into approximately 30 ribotypes, which were similar for the three enzymes used. Using the enzyme HindIII, 31 ribotypes were defined of which ribotypes 1, 2,4,5 and 13-31 accommodated single isolates (Table 1; Fig. 1). Of these, ribotypes 1,2,4 and 5 comprised exclusively the reference cultures of H. piscium (AS239) and A. salmonicida subsp. smithia (AS208), masoucida (AS218) and salmonicida (AS219), respectively. The membership of some ribotypes traversed PCR group boundaries. For example, ribotype 3, which was the most common, included A. salmonicida subsp. achromogenes (AS217) and 10 isolates from PCR groups 2, 3 and 4 (Table 1). Similarly, ribotype 6, with 8 isolates, comprised cultures from PCR groups 1,2 and 3. Many ribotypes contained isolates from restricted geographical locations. Thus, ribotypes 7, 8, 10, 11 and 12 accommodated isolates from Australia, Denmark, Scandinavia, German Bight/Denmark and Finland, respectively. Overall, all the isolates possessed 3 or 4 bands at -2 kb and two bands of <5 and <6 kb, respectively. The ribotype of A. salmonicida subsp. smithia was more distinguishable than the other reference strains.
RAPD analysis
Fifty two isolates were analysed using RAPD with the C oligodeoxynucleotide DNA primer, H3. Cultures ASl, AS89, AS130, AS185 and AS224 could not be exam-ined. Using the RAPD conditions described, the H3 primer amplified 75 DNA fragments that could be scored for similarity between the strains. Fourteen RAPD DNA fragments were unique to individual iso-lates, whereas 61 DNA fragments were shared by two or more cultures. Fig. 2A and B shows the RAPD profiles amplified from 35 isolates. Nineteen cultures produced an identical RAPD profile with at least one other strain. AS51 and AS59, both from PCR Group 4 (Fig. 2B, lanes 3 and 4) were identical to one another, as were AS 122 (PCR Group 4), ASI0 (PCR Group 4) and AS19 (PCR Group 5) (Fig. 2B, lanes 6, 7 and 8). The other pairs of isolates producing similar RAPD profiles were ASI08 and AS221 (both of PCR Group 5) (Fig. 2B, lanes 9 and
Fig. 2. RAPD profiles of atypical A. salmonicida strains. A, Lanes: 1, 100 bp DNA molecular size markers; 2, blank; 3, AS135; 4, AS145; 5, AS47; 6, AS62; 7, AS121; 8, AS200; 9, AS163, 10, AS167; 11, AS182; 12, AS201; 13, AS16; 14, AS28; 15, AS42; 16, AS48; 17, AS74; 18, AS93; 19, AS4; 20,100 bp DNA molecular size markers. B, Lanes: 1, 100 bp DNA molecular size markers; 2, AS54; 3, AS51; 4, AS59; 5, AS78; 6, AS122;7,AS10;8,AS19;9,AS108; 10, AS209; 11, AS221; 12, AS30; 13, AS60; 14, AS69; 15, AS83; 16, AS92; 17, AS102; 18, AS176; 19, AS94; 20, 100 bp DNA molecular size markers. C, UPGMA dendrogram describing genetic similarity based on these RAPD profiles. The scale bar denotes a relative difference of 1 % in branch lengths.
.......-
0.01
-
Atypical Aeromonas salmonicida 57
~ ,...--~ LrC ~
y
----r-
A5218
A5176
A5102
A5221
A5103
A59)
A5203
AS239
AS209
A5135
A5219
A5217
A52111
A551
A578
A592
A5222
A554
A519
A5208
A516
A560
A5204
A52!l)
AS42
A54 A547
AS28
ASI~3
AS)~7
A524
A581
A5m
A5~9
A594
AS8]
A5145
AS62 ASII:!
A~
A5121
58 B. AUSTIN et al.
11), AS30 (PCR Group 5) and AS60 (PCR Group 6) (Fig. 2B, lanes 12 and 13), AS42 and AS 74 (both of PCR Group 3) (Fig. 2A, lanes 15 and 17), AS28 (PCR Group 2) and AS48 (PCR Group 3) (Fig. 2A, lanes 14 and 16), AS167 and AS182 (both of PCR Group 2) (Fig. 2A, lanes 10 and 11), AS6 and AS112 (both of PCR Group 1), and AS46 (PCR Group 2) and ASI09 (PCR Group 1). One isolate, AS128, produced no RAPD profile. The lengths of the RAPD fragments resolved in the agarose gels were determined and a numerical matrix and dendrogram describing genetic similarity were constructed. Note: for pairs or triplets of isolates showing identical RAPD profiles, only one culture was analysed in this fashion.
Fig. 2C shows the dendrogram produced from the similarity matrix. It is readily apparent that atypical A. salmonicida comprise a genetically heterogenous collection as seen by RAPD, with negligible relationships to PCR groups or ribotypes. Along with the isolates that were represented in the similarity analysis by one strain only, only two other pairs of cultures, AS239 (H. piscium; PCR Group 6) and AS209 (PCR Group 5), and AS163 and AS167 (both of PCR Group 2) showed a close genetic similarity, being 95% and 92% similar to one another respectively. The majority of the isolates (i.e. 33 isolates) examined by RAPD showed a similarity of between 80% (e.g. the distance at the node joining strains AS69 and AS94, Fig. 2C) and 88% (e.g. the distance at the node joining strains AS200 and AS42, Fig. 2C). Three cultures, AS121, AS46 and AS112 showed less than 70% similarity to all other isolates as determined by RAPD.
Phenotypic data
Many characteristics were not used in the computer analyses because of problems with reproducibility (gas from glucose, lysine and ornithine decarboxylase) or absence of discriminatory value (glucose fermentation,
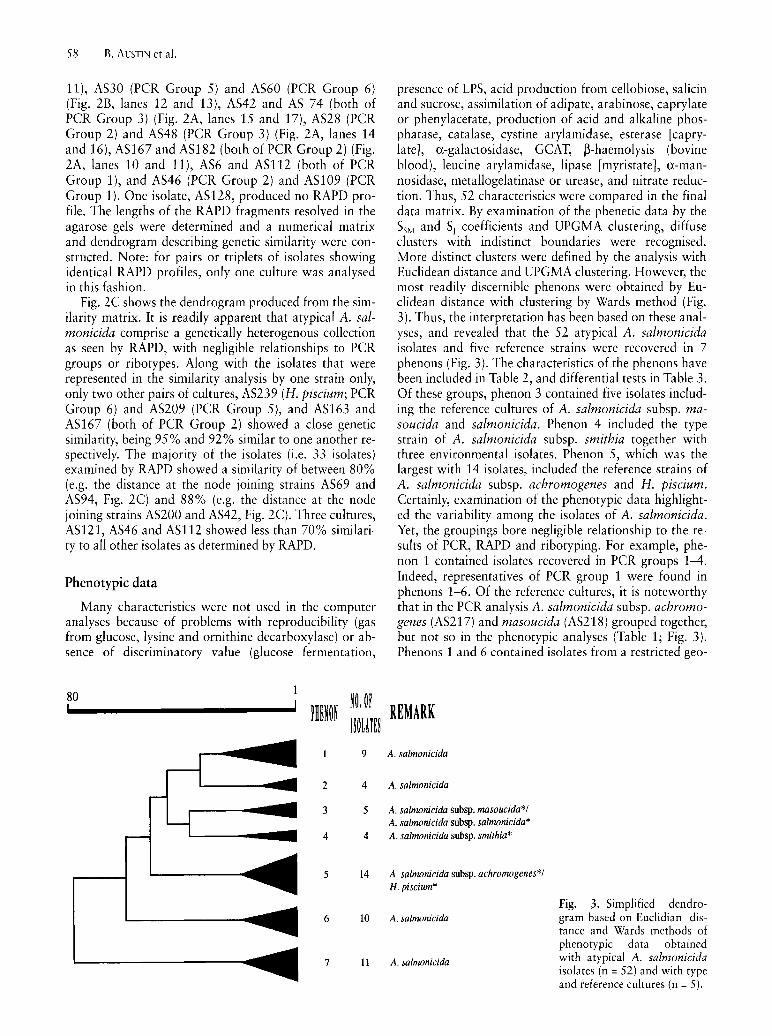
80 ~ij, ijf mm I!ijlm! 9
2 4
3 5
4 4
5 14
6 10
7 11
presence of LPS, acid production from cellobiose, salicin and sucrose, assimilation of adipate, arabinose, caprylate or phenylacetate, production of acid and alkaline phosphatase, catalase, cystine arylamidase, esterase [caprylate], a-galactosidase, GCAT, ~-haemolysis (bovine blood), leucine arylamidase, lipase [myristate], a-mannosidase, metallogelatinase or urease, and nitrate reduction. Thus, 52 characteristics were compared in the final data matrix. By examination of the phenetic data by the SSM and S, coefficients and UPGMA clustering, diffuse clusters with indistinct boundaries were recognised. More distinct clusters were defined by the analysis with Euclidean distance and UPGMA clustering. However, the most readily discernible phenons were obtained by Euclidean distance with clustering by Wards method (Fig. 3). Thus, the interpretation has been based on these analyses, and revealed that the 52 atypical A. salmonicida isolates and five reference strains were recovered in 7 phenons (Fig. 3). The characteristics of the phenons have been included in Table 2, and differential tests in Table 3. Of these groups, phenon 3 contained five isolates including the reference cultures of A. salmonicida subsp. masoucida and salmonicida. Phenon 4 included the type strain of A. salmonicida subsp. smithia together with three environmental isolates. Ph en on 5, which was the largest with 14 isolates, included the reference strains of A. salmonicida subsp. achromogenes and H. piscium. Certainly, examination of the phenotypic data highlighted the variability among the isolates of A. salmonicida. Yet, the groupings bore negligible relationship to the results of PCR, RAPD and ribotyping. For example, phenon 1 contained isolates recovered in PCR groups 1-4. Indeed, representatives of PCR group 1 were found in phenons 1-6. Of the reference cultures, it is noteworthy that in the PCR analysis A. salmonicida subsp. achromogenes (AS217) and masoucida (AS218) grouped together, but not so in the phenotypic analyses (Table 1; Fig. 3). Phenons 1 and 6 contained isolates from a restricted geo-
REMARK
A. salnwnicida
A. salmonicida
A. salmonicida subsp. masoucida*! A. salmonicida subsp. salmonicida* A. salmonicida subsp. smithia*
A. salmonicida subsp. achronwgenes*! H. piscium*
Fig. 3. Simplified dendro-A. salmonicida gram based on Euclidian dis-
tance and Wards methods of phenotypic data obtained
A. salmonicida with atypical A. salmonicida isolates (n = 52) and with type and reference cultures (n = 5).

Table 2. Characteristics of the phenons
Phenon:
1 2 3 4 No. of isolates: 9 4 5 4
Character:
Colonial characteristics: A-layer 44 25 80 25 brown diffusible pigment 88 75 80 50
Assimilation of: N -acety l-glucosamine 11 0 80 25 citrate 0 0 0 50 gluconate 0 0 60 75 glucose 88 75 100 100 malate 22 0 0 50 maltose 88 50 10 75 mannitol 11 50 100 75 mannose 55 50 20 75
Degradation of: aesculin 77 75 100 25 blood (Atlantic salmon) 22 25 80 25 blood (horse) 11 0 40 25 Production of:
N-acetyl-~-glucosaminidase 88 100 100 25 acid from glycerol 55 25 100 75 acid from glucose 77 75 100 25 acid from sorbitol 0 75 20 0 arginine dihydrolase 0 0 80 25 chemotrypsin 55 100 100 0 esterase (butyrate) 100 100 60 75 ~-galactosidase 0 50 80 100 a-glucosidase 11 0 20 100 ~-glucosidase 77 25 60 50 indole 88 100 60 50 metallocaseinase 55 50 20 25 phosphoamidase 22 100 100 100 senne casemase 55 50 80 75 serine gelatinase 11 0 60 50 valine arylamidase 11 100 40 25
Resistance to: ampicillin 77 75 60 75 cephalothin 88 75 60 50
Table 3. Differential characteristics of the phenons
Phenon:
1 2 3
Assimilation of N-acetyl- + glucosamine
Assimilation of maltose + D Degradation of aesculin D D + Production of esterase (butyrate) + + D Production of a-glucosidase Production of valine arylamidase + D
+, - and D correspond to ~80, ~20 and 21-79% or positive responses, respectively.
4
D
D D D + D
Atypical Aeromonas salmonicida
5 14
93 71
0 0 0
78 50 0
21 7
7 36 36
78 86 21
0 14 57 93 71 0 0
78 64 71 64 7 7
86 71
5
+
6 10
50 10
20 10 0
50 10 0
10 10
100 50 40
30 30 0 0 0
30 90 40 0
80 0
20 80 70 50 0
20 40
6
+ +
59
7 11
81 54
0 0 0 0 0 0 0 0
18 36 36
10 9 0 9
27 9
18 63 0 9
90 45
100 27 0 0
81 27
7

60 B. AUSTIN et al.
graphical area, i.e. Northern Europe. Moreover, the isolates were overwhelmingly from marine fish. In contrast, the other phenons included representatives from diverse geographical locations and both freshwater and marine fish species.
Protease groups
The 52 fresh isolates were assigned to 5 protease groups (see Table 1) according to substrate specificity, sensitivity to phenyl methyl sulphonyl fluoride (PMSF) and 1, 10-phenanthroline (OPA) inhibition and the pro-
3 4 5 6
Fig. 4. Casein SDS-PAGE analysis comparing ECPs of four A. salmonicida strains, representative of protease groups 1-4 in Table l. Bands of clearing indicate the presence of proteases. Lane 1, protease group 2; Lane 2, protease group 1; Lane 3, protease group 4; Lane 4, MW, molecular weight marker proteins (values in kDa); Lane 5, protease group 3; Lane 6, AsaP1 (a toxic metallo-caseinase isolated from strain A. salmonicida subsp. achromogenes, strain 265-87).
A
file detected in substrate SDS-PAGE of their ECPs. The type strains for subspecies masoucida and smithia together with one fresh isolate from protease group 1 produced a caseinolytic and gelatinolytic PMSF sensitive proteinase with the same molecular weight as the purified PI protease. Caseinolytic activity was not inhibited by OPA, but some of the gelatinolytic activity was affected. Gelatinolytic components with both high and low molecular weight were also visible in the ECPs of protease group 1. The type strain for A. salmonicida subsp. achromogenes and 24 of the fresh isolates in protease group 2 and 3 produced the AsaPI caseinase with a molecular weight of -20 kDa (Fig. 4), but gelatinolytic acitvity of that size was not detected (Fig. 5). A caseinolytic and gelatinolytic component with a molecular weight of -30 kDa was produced by all these isolates (Fig. 4 and 5). Also, the strains belonging to protease group 2 secreted caseinolytic activity that was sensitive to PMSF inhibition, but this activity was always relatively weak. A weak caseinolytic component with molecular weights of -50 kDa was detected in casein SDS-PAGE of ECPs from protease group 2. Highly gelati no lytic bands with molecular weights of 35 kDa and above were seen in the ECPs of all isolates from protease groups 2 and 3. PMSF sensitivity of the gelatinolytic activity could not be detected. The type strains for A. salmonicida subsp. salmonicida and 9 fresh isolates in protease group 4 produced the PI protease, but other proteases were not detected. No measurable caseinase activity was detected in ECPs from the 19 isolates in protease group 5. The division of the strains into protease groups Sa and 5b was based on different gelatinase profiles and the quantity of the gelatinase production. Thus the isolates in protease group 5b were low gelatinase producers showing only 2-3 faint bands in gelatin substrate gels. However, the composition of the protease groups again traversed those of PCR, RAPD, ribotype and phenon (Table 1).
Fig. 5. Gelatin SDS-PAGE analysis comparing ECPs of 8 A. salmonicida strains representative of protease groups 1-5 in Table l. Bands of clearing indicate the presence of proteases. (A) Lane 1, protease group 2; Lane 2, protease group 4; Lane 3, MW, molecular
weight marker proteins (values in kDa); Lane 4 protease group Sa; Lane 5, protease group 1; Lane 6, protease group 1; Lane 7, protease group 3; (B) Lane 8, protease group Sa; Lane 9, MW markers; Lane 10, protease group 5b.

The named culture of H. piscium grew poorly, and did not produce any caseinase or gelatinase, and was negative for haemolytic activity.
Plasmid profiles
The majority (56/57 = 98%) of the isolates carried plasmids (Table 1). Only one isolate, ASI9, was totally devoid of plasmids (Table 1). Apart from ASI6, which contained a single plasmid of 64 kb, the other isolates carried 2-6 plasmids of 2.3 to 150 kb (Table 1). There was no relationship in terms of plasmid size and PCR groupings, neither was there any association between possession of a particular plasmid and pathogenicity (Table 1). Indeed, ASI9, which lacked plasmids, was pathogenic to Atlantic salmon. Interestingly, H. piscium (AS239) shared its two plasmids with AS122 (Table 1).
Pathogenicity
The majority (45/57 = 79%) of the isolates were pathogenic to fish at a dose of :::::106 cellslfish, as determined by the development of overt clinical signs of disease and eventual mortalities. In these cases, pure growth corresponding to the infective organism was recovered from the diseased animals. Representatives of all PCR and protease groups and phenons were pathogenic (Table 1), with LDso doses varying from 7.4xl03 cells/ fish (AS74) to 6.2xl0s cellslfish (AS135) for Atlantic salmon and rainbow trout, and from d03 (AS203) to 2.4xl0s (AS74) for turbot. For AS60 (which was recovered from turbot) the LDso dose was less for turbot (dOS ) than salmonids (>107 ). Overall, Atlantic salmon and rainbow trout were most susceptible to the isolates. In contrast, turbot was the most resistant species, with only 12 isolates causing pathological changes. Generally following infection via the i.p. route, the pathology was consistent with septicaemia, namely the development of gastro-enteritis, ascites, haemorrhaging in the musculature, and liquefaction of the internal organs, especially the kidney. After administration by the i.m. route, haemorrhaging and muscle liquefaction leading to abscesses and ulcers developed around and extending from the injection site. Although many isolates caused disease in two or more fish species, there was some evidence of host specificity. For example, AS28, AS30 and AS224, which were recovered from cyprinids, attacked only goldfish (Table 1). Similarly, the Norwegian isolates AS132 (from turbot), AS54 (from trout) and AS69 (from salmon) were pathogenic to only turbot, rainbow trout and Atlantic salmon, respectively.
Discussion
In agreement with previous studies (WIKLUND et al., 1994; HIRVELA-KoSKI et al., 1994), it is apparent that atypical A. salmonicida represent a diverse group of organisms, that do not fit into the existing classification of the four validly described subspecies (AUSTIN et al.,
Atypical Aeromonas salmonicida 61
1989). Moreover as with other taxonomic studies (e.g. AUSTIN et al., 1997), the results obtained by the different molecular and phenotypic methods were not congruent, and group membership was not consistent. However, the influence of the different methods of analyses of the data obtained in this study is largely unknown (PRIEST and AUSTIN, 1993). Overall, the results of the PCR, but not RAPD or ribotyping, supported a previously held notion that subspecies achromogenes and masoucida could be combined (MCCARTHY and ROBERTS, 1980; BELLAND and TRUST, 1988). Yet from the phenotypic data, there could be a case for amalgamating subspecies masoucida and salmonicida.
Similar to the views of PATERSON et al. (1980) and BELLAND and TRUST (1988), it seems likely that strains regarded as belonging to H. piscium should be included with A. salmonicida. Indeed, it could be argued from the phenotypic data that H. piscium is synonymous with A. salmonicida subsp. achromogenes, although this view is not supported from molecular studies, i.e. ribotyping, RAPD and PCR grouping. Moreover, there was similarity in the plasmid profile between H. piscium and one of the atypical isolates of A. salmonicida (ASI22). Unfortunately, an authentic type strain of H. piscium was never deposited in a culture collection (SNIESZKO et al., 1950). Therefore, further discussion on the taxonomic status of this organism is inappropriate.
Certainly, the diversity in traits of atypical A. salmonicida has been well recognised previously (see AUSTIN and AUSTIN, 1993). Yet, this study has highlighted the variability among the isolates labelled as 'atypical'. The diversity in ribotypes, i.e. 31 ribotypes from 57 isolates, extends previous findings (PEDERSEN et al., 1996). However, the apparent distinction between the named reference strains (achromogenes, masoucida, salmonicida and smithia) and the 'atypical' isolates indicates a greater heterogeneity in the species than hitherto realised. This variability, particularly in terms of the genetic analyses employed, has been consistent throughout all facets of the study. The discriminatory power of plasmid profiling was generally low. Therefore, it is likely that plasmid profiles are of minor value for identifyingltyping atypical A. salmonicida. The diverse array of plasmids, associated with the isolates of this study, reflects the plasmid heterogeneity found in other studies (e.g. PEDERSEN et al., 1996). DNA profiling using the RAPD technique is claimed to be a very useful method for discriminating among strains of a species (BERG et al., 1994). One previous study describing RAPD analysis of 13 A. salmonicida subsp. salmonicida isolates from the U.K., U.S.A. and Japan found identical RAPD profiles among all the typical A. salmonicida cultures examined (MIYATA et al., 1995), whereas a second RAPD study of typical A. salmonicida also found very homogenous profiles with few polymorphic loci (HANNINEN et al., 1995). In contrast, this study of atypical A. salmonicida isolates produced a different picture showing a great genetic heterogeneity among the cultures. The RAPD technique is known to be subject to variation based on technical parameters such as primer sequence, amplification conditions and template concentration. In

62 B. AUSTIN et al.
this study, care was taken to avoid these problems and the RAPD profiles were reproducible. Therefore, it is likely that the diversity among the atypical strains found using the RAPD technique correlated with the diversity found with the other methods. The majority of the atypical isolates showed a RAPD-based similarity of between 80% and 90% with their most similar neighbour, but the most distant 11 divisions among the 52 isolates occurred at less than 80% similarity. The A. salmonicida reference strains (salmonicida, achromogenes, masoucida and smithia) were scattered throughout the similarity analysis with no close relationships to any of the atypical isolates. The ribotyping, plasmid profiling and RAPD data provided evidence that atypical A. salmonicida represents a far greater genetically heterogenous taxon that its typical A. salmonicida counterpart.
In agreement with other views (see AUSTIN and AUSTIN, 1993), some of the isolates were more fastidious than typical A. salmonicida. Also, there was evidence of oxidase-negativity in 5 isolates from flounder and turbot, as reported by other workers (e.g. PEDERSEN et a!., 1994; WIKLUND et a!. 1994). Among the phenotypic traits, there was variability in the production of the brown diffusible pigment, which was nevertheless produced by isolates from all phenons. Interestingly, pigment production is usually associated with A. salmonicida subsp. salmonicida (POPOFF, 1984); atypical isolates being often regarded as slow or non-pigmenting (AUSTIN and AUSTIN, 1993). A complication is that there have been some reports of typical isolates that do not produce pigment (e.g. WIKLUND et a!. 1993).
The data from this study supports a view that there is less variation between atypical and typical isolates regarding the sharing of cell-associated virulence factors, such as the A-layer protein, outer membrane proteins and the LPS component, as compared to the exotoxins (CHART et a!., 1984; EVENBERG et a!', 1985; LEE and ELLIS, 1990; GUDMUNDSDOTTIR et a!., 1990; AUSTIN and AUSTIN, 1993; TORANZO and BARJA, 1993; HIRST and ELLIS, 1994). Currently, there is clear evidence that a 70 kDa serine protease, PI, is one of the major exotoxins of typical A. salmonicida strains (FYFE et a!., 1986; ELLIS, 1991), and also that a 20 kDa metallo-caseinase, AsaP1, is the major exotoxin of a group of atypical strains including the reference strain for A. salmonicida subsp. achromogenes, NCIMB1110 (GUDMUNDSDOTTIR, 1996; GUDMUNDSDOTTIR and MAGNADOTTIR, 1997; GUNNLAUGSDOTTIR and GUDMUNDSDOTTIR, 1997). In the present study, the isolates did not produce both the PI and AsaPI proteases. The PI protease was detected in the reference cultures of subspecies masoucida, salmonicida and smithia and 9 of the fresh isolates, originating from 6 different fish species from 6 different geographical 10-cations. The AsaPI protease was detected in the reference culture for A. salmonicida subsp. achromogenes and 24 fresh isolates from 10 species of fish in the Nordic countries, Scotland and Canada. A total of 19 strains did not produce either of the proteolytic exotoxins. Overall the protease profiles for the atypical isolates were in agreement with GUDMUNDSDOTTIR (1996).
The pathogenicity of A. salmonicida for fish, especially salmonids, is well established (AUSTIN and AUSTIN, 1993). The increasing recognition of atypical isolates causing disease, notably skin lesions/ulceration, in marine fish (e.g. WIKLUND and DALSGAARD, 1995; LARSEN and PEDERSEN, 1996) is borne out from the results of this study. In contrast with the findings of WIKLUND (1995), some of the atypical isolates from marine fish adversely affected salmonids and therefore may pose a risk to aquaculture. It may be argued that the apparently nonpathogenic isolates could well cause disease in other fish species under less favourable environmental conditions (= stress) (AUSTIN and AUSTIN, 1993).
Further work is needed to clarify the precise taxonomic position of atypical A. salmonicida strains and their relationship to other taxa in the genus Aeromonas. The current view from this study is that these atypical strains warrant description as new Aeromonas subspecies and possibly in separate Aeromonas species. It is ironic that this suggestion for new taxa reflects the earlier views of SMITH (1963), who recommended that A. salmonicida should be transferred to the newly described genus Necromonas, as N. salmonicida. In addition, she proposed a new species, N. achromogenes, which subsequently became recognised as A. salmonicida subsp. achromogenes. This classification was not supported on the basis of DNA relatedness (e.g. MAcINNES et a!., 1979); consequently the genus Necromonas was not accepted. Finally, the heterogeneity found among the atypical isolates and the lack of correlation between the typing methods used in this study emphasises the problem of defining useful diagnostic tests to deal with the increasing number of reports of atypical A. salmonicida isolates. In this study, perhaps only the PCR analysis in terms of ease, speed and number of groups identified, i.e. 6, provided a possible candidate utilitarian diagnostic system, but a greater number of isolates need to be examined. It seems likely that in the short term, atypical isolates of A. salmonicida must continue to be identified and grouped together simply on the basis of slow and/or fastidious growth.
Acknowledgements Financial support was provided by CEC Contract No: AIR3-
CT94-1884. We are grateful to colleagues for the gift of cultures. B. GUNNLAUGSD6TTIR, M.-B. H0]GAARD, K. KAAS and H. SIGURJ6NSD6TTIR are acknowledged for excellent technical assistance.
References
AUSTIN, B.: Recovery of 'atypical' isolates of Aeromonas salmonicida, which grow at 37°C, from ulcerated nonsalmonids in England. J. Fish Dis. 16, 165-168 (1993).
AUSTIN, B., ADAMS, c.: 8. Fish pathogens, pp. 197-243. In: The Genus Aeromonas (B. AUSTIN, ALTWEGG, M., GOSLING, P. ]., JOSEPH, S., eds.) Chichester, England, John Wiley & Sons (1996).
AUSTIN, B., AUSTIN, D. A.: Methods for the Microbiological Examination of Fish and Shellfish. Chichester, England, Ellis Horwood (1989).

AUSTIN, B., AUSTIN, D. A.: Bacterial Fish Pathogens Disease of Farmed and Wild Fish, 2nd edn. Chichester, England, Ellis Horwood (1993).
AUSTIN, D. A., McINTOSH, D., AUSTIN, B.: Taxonomy of fish associated Aeromonas spp., with the description of Aeromonas salmonicida subsp. smithia subsp. nov. System. Appl. MicrobioI. 11,277-290 (1989).
AUSTIN, B., AUSTIN, D. A., BLANCH, A. R., CERDA, M., GRIMONT, E, GRIMONT, P. A. D., JOFRE, J., KOBLAVI, S., LARSEN, J. L., PEDERSEN, K., TIAINEN, T., VERDONCK, L., SWINGS, J.: A comparison of methods for the typing of fish-pathogenic Vibrio spp. System. Appl. Microbiol. 20,89-101 (1997).
AUSUBEL, EM., BRENT, R., KINGSTON, R. E., MOORE, D. D., SEIDMAN, J. G., SMITH, J. A., STRUHL, K.: Short protocols in molecular biology, 2nd. edn. New York, USA, John Wiley & Sons (1992).
BELLAND, R. J., TRUST, T. J.: DNA: DNA reassociation analysis of Aeromonas salmonicida. J. Gen. Microbiol. 134,307-315 (1988).
BERG, D. E., AKOPYANTS, N. S., KERSULYTE, D.: Fingerprinting microbial genomes using RAPD and AP-PCR method. Methods Mol. Cell. BioI. 5, 13-24 (1994).
CAETANO-ANOLLES, G., BASSAM, B. J., GRESSHOFF, P. M.: DNA amplification fingerprinting using very short arbitrary oligonucleotide primers. Biotechnol. 9,553-557 (1991).
CARNAHAN, A. M., ALTWEGG, M.: 1. Taxonomy, pp. 1-38. In: The Genus Aeromonas (B. AUSTIN, ALTWEGG, M., GOSLING, P. J., JOSEPH, S., eds.). Chichester, England, John Wiley & Sons (1996).
CHART, H., SHAW, D. H., ISHIGURO, E. E., TRUST, T. J.: Structural and immunochemical homogeneity of Aeromonas salmonicida lipopolysaccharide. J. Bacteriol. 158, 16-22 (1984).
DALSGAARD, I., GUDMUNDSDOTTIR, B. K., HELGASON, S., HOlE, S., THORESEN, O. E, WICHARDT, U.-P., WIKLUND, T.: Identification of atypical Aeromonas salmonicida: inter-laboratory evaluation and harmonization of methods. J. Appl. MicrobioI. (1997) (in press).
ELLIS, A. E.: An appraisal of the extracellular toxins of Aeromonas salmonicida ssp. salmonicida. J. Fish Dis. 14, 265-277 (1991).
EVENBERG, D., VERSLUIS, R., LUGTENBERG, B.: Biochemical and immunological characterization of the cell surface of the fish pathogenic bacterium Aeromonas salmonicida. Biochim. Biophys. Act. 815,233-244 (1985).
FELSENSTEIN, J.: PHYLIP: phyogeny inferance package. Distributed by author, University of Washington, Seattle (1993).
FYFE, L., FINLEY, A., COLEMAN, G., MUNRO, A. L. S.: A study of the pathological effect of isolated Aeromonas salmonicida extracellular protease on Atlantic salmon, Salmo salar L. J. Fish Dis. 9, 403-409 (1986).
GUDMUNDSDOTTIR, B. K., HASTINGS, T. S., ELLIS, A. E.: Isolation of a new toxic protease from a strain of Aeromonas salmonicida subspecies achromogenes. Dis. Aquat. Org. 9, 199-208 (1990).
GUDMUNDSDOTTlR, B. K.: Comparison of extracellular proteases produced by Aeromonas salmonicida strains, isolated from various fish species. J. Appl. Bacteriol. 80, 105-113 (1996).
GUDMUNDSDOTTIR, B. K., MAGNADOTTIR, B.: Protection of Atlantic salmon (Salmo salar L.) against an experimental infection of Aeromonas salmonicida ssp. achromogenes. Fish Shellfish lmmunol. 7,55-69 (1997).
GUNNLAUGSDOTTIR, B., GUDMUNDSDOTTlR, B. K.: Pathogenicity of atypical Aeromonas salmonicida in Atlantic salmon compared with protease production. J. Appl. Microbiol. (1997) (in press).
GUSTAFSON, C. E., THOMAS, C. J., TRUST, T. J.: Detection of Aeromonas salmonicida from fish by using polymerase chain
Atypical Aeromonas salmonicida 63
reaction amplification of the virulence surface array protein gene. Appl. Environ. Microbiol. 58, 3816-3825 (1992).
HANNINEN, M.-L., RID ELL, J., HIRVELA-KoSKI, V: Phenotypic and molecular characteristics of Aeromonas salmonicida subsp. salmonicida isolated in southern and northern Finland.J. Appl. Bacteriol. 79, 12-21 (1995).
HEUSSEN, c., DOWDLE, E. B.: Electrophoretic analysis of plasminogen activators in polyacrylamide gels containing sodium dodecyl sulfate and copolymerized substrates. Analyt. Biochem. 102, 196-202 (1980).
HIGUCHI, R.: Simple and rapid preparation of samples for PCR, pp. 31-38. In: PCR technology. Principles and applications for DNA amplification. (A. ERLICH, ed.) Stockton Press, New York (1989).
HIRS1~ I. D., ELLIS, A. E.: Iron-regulated outer membrane proteins of Aeromonas salmonicida are important protective antigens in Atlantic salmon against furunculosis. Fish Shellfish Immunol. 4, 29-45 (1994).
HIRVELA-KoSKl, V., KOSKI, P., NIIRANEN, H.: Biochemical properties and drug resistance of Aeromonas salmonicida in Finland. Dis. Aquat. Org. 20, 191-196 (1994).
HOlE, S., HEUM, M., THORESEN, O. E: Detection of Aeromonas salmonicida by polymerase chain reaction in Atlantic salmon vaccinated against furunculosis. Fish Shellfish Immunol. 6, 199-206 (1996).
HUCKER, G. J., CONN, H. J.: Methods of Gram staining. Tech. Bull. N. Y. Agric. Ex. Stn. No. 93 (1923).
KADO, C. T., LIU, S.-T.: Rapid procedure for detection and isolation of large and small plasmids. J. Bacteriol. 145, 1365-1373 (1981).
LARSEN, J. L., PEDERSEN, K.: Atypical Aeromonas salmonicida isolated from diseased turbot (Scophthalmus maximus). Acta Vet. Scand. 37, 139-146 (1996).
LEE, K.-K., ELLIS, A. E.: Glycerophospholipid: cholesterol acyltransferase complexed with LPS is a major lethal exotoxin and cytolysin of Aeromonas salmonicida: LPS stabilises and enhances toxicity of the enzyme. J. Bacteriol. 172, 5382-5393 (1990).
MARTINEZ-MuRCIA, A. J., BENLLOCH, S., COLLINS, M. D.: Phylo-. genetic interrelationships of members of the genera Aeromonas and Plesiomonas as determined by 16S Ribosomal DNA Sequencing: Lack of congruence with results of DNADNA hybridizations. Int. J. System. Bacteriol. 42, 412-421 (1992).
MCCARTHY, D. H., ROBERTS, R. J.: Furunculosis of fish - the present state of our knowledge, pp. 293-341. In: Advances in Aquatic Microbiology (DROOP, M. A., JANNASCH, H. W., eds). London, England, Academic Press (1980).
McINNES, J. I., TRUST, T. J., CROSA, J. H.: Deoxyribonucleic acid relationships among members of the genus Aeromonas. Can. J. Microbiol. 25, 579-586 (1979).
MIYATA, M., AOKI, T., INGLIS, V., YOSHIDA, T., ENDO, M.: RAPD analysis of Aeromonas salmonicida and Aeromonas hydrophila. J. Appl. Bacteriol. 79,181-185 (1995).
MORGAN, U. M., CONSTANTINE, C. c., GREENE, W. K., THOMPSON, R. C. A.: RAPD (random amplified polymorphic DNA) analysis of Giardia DNA and correlation with isoenzyme data. Trans. Royal Soc. Trop. Med. 87, 702-705 (1993).
NAKATSUGAWA, T.: Atypical Aeromonas salmonicida isolated from cultured shotted halibut. Fish Pathol. 29,193-198 (1994).
NERLAND, A. H.: The nucleotide sequence of the gene encoding GCAT from Aeromonas salmonicida subsp. salmonicida. J. Fish Dis. 19, 145-150 (1996).
PATERSON, W. D., DOUEY, D., DESAUTELS, D.: Relationship between selected strains of typical and atypical Aeromonas salmonicida, Aeromonas hydrophila and Haemophilus piscium. Can. J. Microbial. 26, 588-598 (1980).

64 B. AUSTIN et al.
PEDERSEN, K., LARSEN, J. L.: rRNA gene restriction patterns of Vibrio anguillarum serogroup 01. Dis. Aquat. Org. 16, 121-126 (1993).
PEDERSEN, K., KOFOD, H., DALSGAARD, I., LARSEN, J. L.: Isolation of oxidase-negative Aeromonas salmonicida from diseased turbot Scophthalmus maximus. Dis. Aquat. Org. 18, 149-154 (1994).
PEDERSEN, K., DALSGAARD, I., LARSEN, J. L.: Characterization of atypical Aeromonas salmonicida isolates by ribotyping and plasmid profiling. J. Appl. Bacteriol. 80,37-44 (1996).
POPOFF, M.: Genus III. Aeromonas Kluyver and Van Niel 1936, 398AL, pp. 545-548. In: Bergey's Manual of Systematic Bacteriology, Vol. 1 (N. R. KRIEG, HOLT, J. G., eds.). Baltimore, Williams & Wilkins (1984).
PRIEST, E G., AUSTIN, B.: Modern Bacterial Taxonomy, 2nd. edn. London, Chapman & Hall (1993).
ROCHELLE, P. A., FRY, J. c., DAY, M. J., BALE, M. J.: An accurate method for estimating sizes of small and large plasmids and DNA fragments by gel electrophoresis. J. Gen. Microbiol. 132,53-59 (1985) .
. SAMBROOK, ]., FRITSCH, E. E, MANIATIS, T. J.: Molecular cloning: A laboratory manual. 2nd ed. Cold Spring Harbor Laboratory Press, New York (1989).
SMITH, I. W.: The classification of Bacterium salmonicida. J. Gen. Microbiol. 33, 263-274 (1963).
SNEATH, P. H. A., SOKAL, R. R.: Numerical Taxonomy: The Principles and Practice of Numerical Classification. San Francisco, W. H. Freeman (1983).
SNIESZKO, S. E, GRIFFIN, P., FRIDDLE, S. B.: A new bacterium (Haemophilus piscium n. sp.) from ulcer diseases of trout. J. Bacteriol. 59,699-701 (1950).
TORANZO, A. E., BARJA, J. L.: Virulence factors of bacteria pathogenic for coldwater fish. Ann. Rev. Fish Dis. 3, 5-36 (1993).
WARDLAW, A. c.: Practical Statistics for Experimental Biologists. Chichester, England, John Wiley & Sons Limited (1985).
WHITBY, P. W., LANDON, M., COLEMAN, G.: The cloning and nucleotide sequence of the serine protease gene (aspA) of Aeromonas salmonicida ssp. salmonicida. FEMS Microbiol. Lett. 99,65-71 (1992).
WIKLUND, T.: Virulence of 'atypical' Aeromonas salmonicida isolated from ulcerated flounder Platichthys flesus. Dis. Aquat. Org. 21,145-150 (1995).
WIKLUND, T., BYLUND, G.: Skin ulcer disease of flounder Platichthys flesus in the northern Baltic Sea. Dis. Aquat. Org. 17,165-174 (1993).
WIKLUND, T., DALSGAARD, I.: Atypical Aeromonas salmonicida associated with ulcerated flatfish species in the Baltic Sea and the North Sea. J. Aquat. Animal Health 7, 218-224 (1995).
WIKLUND, T., DALSGAARD, 1., EROLA, E., OLIVIER, G.: Characteristics of 'atypical', cytochrome oxidase-negative Aeromonas salmonicida isolated from ulcterated flounders [Platichthys flesus (L.ll. J. Appl. Bacteriol. 76,511-520 (1994).
WIKLUND, T., LONNSTROM, L., NIIRANEN, H.: Aeromonas salmonicida ssp. salmonicida lacking pigment production, isolated from farmed salmonids in Finland. Dis. Aquat. Org. 15,219-223 (1993).
Corresponding author: Professor B. AUSTIN, Department of Biological Sciences, HeriotWatt University, Riccarton, Edinburgh EH14 4AS, Scotland (U.K.) Tel.: +44-131-451-3452; Fax: +44-131-451-3009; e-mail: [email protected]


![Aeromonas salmonicida proliferation and quorum … salmonicida proliferation and ... • Autoinducer-2 system – A. hydrophila – 4,5- ... J. Padra-08-04.ppt [Compatibility Mode]](https://img.pdfslide.net/doc/110x75/5ada57817f8b9aee348ca8d7/aeromonas-salmonicida-proliferation-and-quorum-salmonicida-proliferation-and.jpg)


























