Embed Size (px)
Citation preview

Gene 519 (2013) 318–325
Contents lists available at SciVerse ScienceDirect
Gene
j ourna l homepage: www.e lsev ie r .com/ locate /gene
Characterization of Cherax quadricarinatus prohibitin and its potentialrole in spermatogenesis
Di-An Fang a,b,1, Yang Wang b,1, Juan Wang b,1, Li-Hua Liu b, Qun Wang b,⁎a Scientific Observing and Experimental Station of Fishery Resources and Environment in the Changjiang River, Freshwater Fisheries Research Center, Wuxi, Shanshui Road 9, 214081, Chinab School of Life Science, East China Normal University, 3663 North Zhongshan Road, Shanghai 200062, China
Abbreviations: PHB, prohibitin; IM, innermembranes;prohibitin; RT-qPCR, real-time quantitative RT-PCR;ML, mTaylor–Thornton; PVDF, polyvinylidene difluoride; Iphosphate-buffered saline; FITC, fluorescein isothiocyanamicroscopy; TBS, Tris buffered saline; H&E, hematoxyliprohibitin/flotillin/HflK/C; mtDNA, mitochondrial DNA.⁎ Corresponding author. Tel.: +86 21 6223 2429; fax
E-mail address: [email protected] (Q. Wang).1 The first three authors contribute equally to the wo
0378-1119/$ – see front matter © 2013 Elsevier B.V. Alhttp://dx.doi.org/10.1016/j.gene.2013.02.003
a b s t r a c t
a r t i c l e i n f oArticle history:Accepted 6 February 2013Available online 24 February 2013
Keywords:Cherax quadricarinatusPHBSpermatogenesis
Prohibitin (PHB) proteins have diverse functions, such as cellular signaling, transcriptional control and mito-chondrial biogenesis. In this study, we characterized PHB gene and its protein expression in Cheraxquadricarinatus. PHB cDNA comprises 1472 nucleotides with an open reading frame of 828 bp, which encodes275 amino acid residues. The highest transcript levels were found during the spermatogonial developmentalphase, with the lowest levels detected during the resting phase in the reproductive cycle. Western blot anal-ysis revealed that PHB is an approximately 30 kDa protein, and occurs in a number of unexpected isoforms,ranging from 30 kDa to greater than 180 kDa in the testes of different developmental phases, which may bethe ubiquitinated substrates. The strongest immunolabeling signal was found in spermatogonia, with lowerlevels of staining in secondary spermatocytes, and weak or absent expression in mature sperm. Immunogoldelectron microscopy results confirmed the localization of PHB in the inner mitochondrial membranes. Theresults showed that PHB is a substrate protein for spermatogenesis, with a potential reproductive functioninvolving sperm ubiquitination in invertebrates.
© 2013 Elsevier B.V. All rights reserved.
1. Introduction
Prohibitin (PHB) is an evolutionally conserved protein belonging to aBand-7 or PHB domain family (Merkwirth and Langer, 2009; Mishra etal., 2006) and mainly localized in mitochondria (Artal-Sanz andTavernarakis, 2009; Ikonen et al., 1995; Terashima et al., 1994;Thompson et al., 1999). Two highly homologous forms of this proteinexist termed PHB1 and PHB2 (Mishra et al., 2006). Besides the initiallyproposed role in cell cycle progression (Cummins, 1998; Mishra et al.,2006; Nijtmans et al., 2000; Nuell et al., 1991; Piper et al., 2002;Sharma and Qadri, 2004; Theiss et al., 2007), PHBs have also been impli-cated in transcriptional regulation (Sun et al., 2004), sister chromatidcohesion regulation (Schmittgen and Livak, 2008), cellular signaling(Mishra et al., 2006; Rajalingam et al., 2005), apoptosis (Fusaro et al.,
Cq-PHB, Cherax quadricarinatusaximum likelihood; JTT, Jones–F, immunofluorescence; PBS,te; IEM, immunogold electronn and eosin; S SPFH, tomatin/
: +86 21 6223 3754.
rk.
l rights reserved.
2003) and mitochondrial biogenesis (Artal-Sanz and Tavernarakis,2009; Berger and Yaffe, 1998; Nijtmans et al., 2000). PHB1 mRNA andprotein were reported to be expressed differentially during germ celldevelopment. Subsequently, it was suggested that PHB1, a conserved30 kDa component of the inner mitochondrial membrane, is alsoexpressed as an unusual, high-molecular-mass isoform in mammalianspermatozoa (Sutovsky, 2003). However, little is known regarding thefunctional significance of PHB1 in the male germ cell in invertebrateanimals.
The mitochondrion is a complex sub-cellular organ present in thecytoplasm of all animal and plant cells (Sanz et al., 2003). Several stud-ies have indicated an important role for mitochondria in spermatogen-esis and fertility (Sharma and Qadri, 2004; Shivaji et al., 2009).Mitochondria are a double-membrane structure consisting of outerand inner membranes (IM) separated by an inter-membrane space.The IM forms numerous folds inside the mitochondrial matrix knownas “cristae” and is unusually high in protein content. PHB1 and PHB2 as-sociate to form a macromolecular structure of approximately 1 MDa atthe mitochondrial IM (Nijtmans et al., 2002).The PHB complex hasbeen implicated in regulation of membrane protein degradation bythe mitochondrial m-AAA protease (Langer, 2000). Ubiquitination is aversatile and universal mechanism for protein recycling, through whichmisfolded or aged proteins are tagged for degradation by the covalentattachment of one or more molecules of ubiquitin (Sutovsky, 2003).Mitochondrial DNA mutations have been reported in mitochondrial

Table 1Sequences of primers.
Primers Sequence Code
Degenerate primers for target PHBForward primer 5′-ACCATGNCYTCHGGGTRCATMGA-3′ DP-RReverse primer 5′-GGBGATGAGYTNTTNACYGACA-3′ DP-F
Gene-specific primers for PHB cloningGene-specific primer pairs forRACE
5′-GAGTTCTCCAGCATCATACTGAGCCA-3′ Gp5-15′-GGAATGTCCCTGTAAACACTGGCAGT-3′ Gp3-15′-AGGGTGGTCTGGTTGCTTGGTAG-3′ Gp5-25′-TATCTATTACTCATCTGACCTTCGGC-3′ Gp3-2
RT-PCR primersPHB 5′ primer 5′-TTGCCGAAGGTCAGATGAGTAATA-3′ RT-RPHB 3′ primer 5′-GGAATGTCCCTGTAAACACTGGC-3′ RT-F
qRT-PCR primersPHB 5′ primer 5′-TGAAGCATCACCATCAGCACTAAT-3′ Real-RPHB 3′ primer 5′-TATCTATTACTCATCTGACCTTCGGC-3′ Real-F
18S rRNA primers for RT-PCR and qRT-PCR18S rRNA reverse 5′-GGAGGTAGTGACGAAAAATAACG -3′ 18S-R18S rRNA forward 5′-GGAGCGTGACAGTAAGCACCATCGG -3′ 18S-F
319D.-A. Fang et al. / Gene 519 (2013) 318–325
respiration defects giving rise to meiotic arrest and abnormalities insperm morphology (Ankel-Simons and Cummins, 1996; Cummins,1998; Merkwirth et al., 2008; Spiropoulos et al., 2002).
Cherax quadricarinatus is a native species in the north of Queens-land (Australia) and the southeast of Papua New Guinea (Jones,1997). It is an important cultured species in aquaculture that hasbeen reared for human consumption since 1985. Knowledge of thereproductive biology of male C. quadricarinatus is mainly limited tothe anatomy of the reproductive system and the effects of nutritionor neuroendocrine regulation on fecundity (López Greco and LoNostro, 2008). Hence, a better understanding of the molecular mecha-nisms of spermatogenesis has become an immediate research priority.PHB functions as a reproduction-related and mitochondrial membraneprotein, PHB cDNA and protein had been characterized from yeast(Tatsuta et al., 2005), Pneumocystis carinii (Narasimhan et al., 1997),Drosophila (Eveleth and Marsh, 1986), mouse (Terashima et al., 1994),rat (Altus et al., 1995; Choongkittaworn et al., 1993) and human (Satoet al., 1992), but no studies have been conducted in the Decapoda,Parastacidae.
The main objectives of this present study were: (1) to clone the PHBgene from C. quadricarinatus (designated Cq-PHB); (2) to investigate thetissue distribution and temporal mRNA expression of Cq-PHB in testesduring spermatogenesis, and (3) to locate Cq-PHB in testes and spermmitochondria. Insight into the PHB gene and its expression pattern inspermatogenesis is important for understanding the molecular mecha-nisms of crayfish which may be conducive to the development of repro-duction biology. In combination with the identification of its localizationin testes and sperm mitochondria, the information reported here mayfacilitate further investigations of inducible PHB in the freshwater cray-fish and inform further investigation of its mitochondrial inheritance ininvertebrates.
2. Materials and methods
2.1. Animals
Fifteen male C. quadricarinatus (70–110 g) were purchased fromShanghai Jinshan aquaculture farm during May (spring), July (sum-mer) September (autumn) and December (winter) in 2010, andthen placed in an ice bath for 3 to 5 min until lightly anesthetizedprior to sacrifice. Various tissues were dissected, immediately frozenin liquid nitrogen and stored at −80 °C.
2.2. Isolation of total RNA and cDNA synthesis
Total RNA was extracted using Trizol reagent (RNA Extraction Kit,CA, USA) according to the protocol provided by the manufacturer. Theconcentration and quality of total RNA were estimated by spectro-photometry (absorbance at 260 nm) and agarose gel electrophoresis,respectively. Total RNA (2 μg) isolated from testis was reverse tran-scribed using the SMART™ cDNA kit (Clonetech, CA, USA) for cDNAcloning and using the Prime Script™ RT-PCR Kit (TaKaRa, Japan) forsemi-quantitative RT-PCR (RT-PCR) analysis or the PrimeScriptReal-time PCR Kit (TaKaRa, Japan) for real-time quantitative RT-PCR(RT-qPCR) analysis.
2.3. PHB gene isolation
Target fragments of cDNAs encoding PHB were amplified with thedegenerate primers DP1 and DP2. All primers were synthesized atShanghai Invitrogen Biotech Co., Ltd. (Shanghai, China) and are listedin Table 1. The obtained target sequences were verified and analyzedfor similarity with other known PHB sequences using BLAST pro-grams (Altschul et al., 1997). The cDNA for 3′ or 5′ RACE was ampli-fied using the kit recommended reaction system and two pairs ofgene-specific primers (Gp5-1, Gp5-2, Gp3-1, and Gp3-2; Table 1),
which were designed based on the obtained cDNA partial sequenceof Cq-PHB. The full-length Cq-PHB cDNA was obtained using theRACE cDNA amplification kit according to the protocol provided bythe manufacturer (Bioscience Clontech, CA, USA).
2.4. Alignment of multiple sequences and phylogenetic analysis
Multiple alignments of the full-length PHB sequencewere comparedwith PHB sequences from other species. Amino acid sequences of vari-ous crustacean species were retrieved from the NCBI GenBank and ana-lyzed using the ClustalW 2.0Multiple Alignment program. Phylogeneticanalyses were conducted using maximum likelihood (ML) approach(Holmes, 2003; Huelsenbeck and Rannala, 1997). It is known to be ro-bust to a number of systematic biases of phylogenetic reconstruction.A ML phylogenetic tree was constructed using MEGA software version5.0 (Holmes, 2003; Tamura et al., 2011). Jones–Taylor–Thornton (JTT)model was used as substitution model for phylogenetic tree construc-tion (Jones et al., 1992). Reliability of nodes was estimated byML boot-strap percentages (BPML) (Sullivan, 2005) obtained after 1000 pseudoreplications, using the previously estimated ML parameters.
2.5. PHB mRNA transcript expression patterns
Quantitative analysis of tissue- and testes developmental cycle-dependent mRNA expression was conducted via Real-time RT-qPCR.First-strand cDNA was prepared as described in Section 2.2. RT-qPCR primers (Q-R and Q-F, Table 1) were designed based on thecloned Cq-PHB cDNA to produce a 310 bp amplicon. PCR reactionswere performed according to the PrimeScript Real-time PCR Kitprotocol. The primers 18S-F and 18S-R were designed based on thesequence of C. quadricarinatus 18S rRNA (AF235966) to amplify a232 bp fragment. Samples were run in triplicate and normalizedto the selected control gene, 18S rRNA. All PCR reactions were per-formed in triplicate using extracted RNA (pooled) of the same con-centration. Cq-PHB expression levels were calculated by the 2−ΔΔCt
comparative CT method (Colebatch et al., 2002). Mean and standarddeviation values were calculated from triplicate experiments, andpresented as fold differences in expression relative to 18S rRNA.Data were analyzed using the CFX Manager™ software version 1.0.

320 D.-A. Fang et al. / Gene 519 (2013) 318–325
2.6. Preparation of anti-PHB antibody
The purified recombinant Cq-PHB protein was prepared using apreviously described method (Zhou et al., 2010) and was then usedto raise antibodies in a New Zealand white rabbit. Approximately1 mg of purified PHB was emulsified in Freund's complete adjuvantand injected subcutaneously at multiple sites. Animals were subse-quently boosted with of 0.5 mg antigen mixed with Freund's incom-plete adjuvant delivered on in three separate injections at intervalsof 2 weeks (total, 1.5 mg antigen). One week after the final booster,blood was collected and serum was prepared. Serum from the samerabbit collected prior to immunization was used as a control. An in-crease the titers of peptide-specific antibodies was verified by ELISA.The anti-sera were aliquoted and stored at −80 °C.
2.7. Western blot analysis
Testes tissues were homogenized in SDS sample buffer supplementedwith 1 mM PMSF. Proteins were resolved on a 12.5% polyacrylamide gel
Fig. 1. Nucleotide sequence and deduced amino acid sequence of the C. quadricarinatusunderlined and the coiled-coil domain is shown in bold. The CCAAT box is shown in the re
under reducing conditions and electroblotted onto a polyvinylidenedifluoride (PVDF) membrane. The membrane was treated in blockingsolution (Roche, China) and incubated with primary antibody at 1:1000for 1 h at room temperature. After washing with Tris-buffered saline(20 mM Tris–HCl, 0.9% NaCl, pH 7.4) containing 0.1% Tween-20, themembrane was incubated with a second antibody (goat anti-rabbitserum) labeled with alkaline phosphatase (Bio-Rad) at 1:10,000 for 1 h.Detection was performed using NBT/BCIP (Roche) as a substrate.
2.8. Immunofluorescence (IF)
Freshly dissected testes were fixed in 4% paraformaldehyde andembedded in OCT compound (Sakura Finetek, Torrance, CA). Thecryosections (8 μm in thickness) were mounted on glass slides,washed in phosphate-buffered saline (PBS), and immersed in 3%BSA for 1 h to block non-specific binding. These slides were then in-cubated with primary antibodies against PHB at dilutions of 1:100for 18 h at 4 °C, washed twice in PBS/Tween-20 solution, incubatedwith a fluorescein isothiocyanate (FITC)- conjugated secondary
full-length cDNA. The transmembrane domain is shown in gray, the PHB domain isctangle.
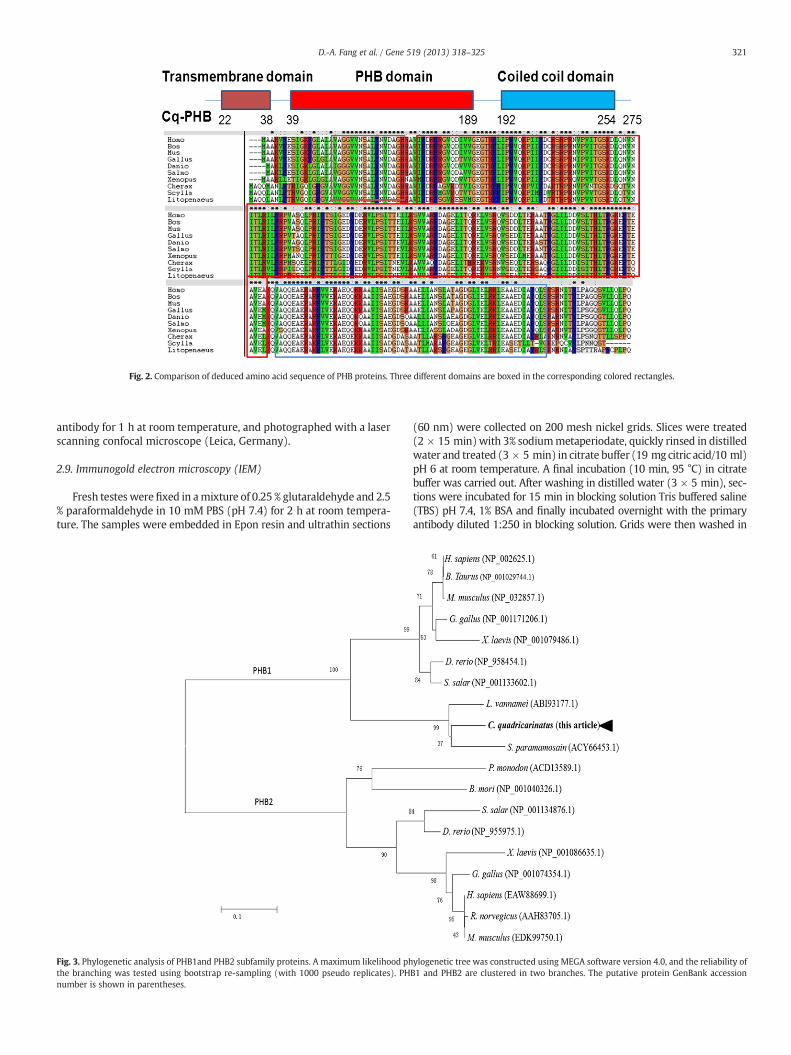
Fig. 2. Comparison of deduced amino acid sequence of PHB proteins. Three different domains are boxed in the corresponding colored rectangles.
321D.-A. Fang et al. / Gene 519 (2013) 318–325
antibody for 1 h at room temperature, and photographed with a laserscanning confocal microscope (Leica, Germany).
2.9. Immunogold electron microscopy (IEM)
Fresh testes were fixed in amixture of 0.25 % glutaraldehyde and 2.5% paraformaldehyde in 10 mM PBS (pH 7.4) for 2 h at room tempera-ture. The samples were embedded in Epon resin and ultrathin sections
Fig. 3. Phylogenetic analysis of PHB1and PHB2 subfamily proteins. A maximum likelihood phthe branching was tested using bootstrap re-sampling (with 1000 pseudo replicates). PHnumber is shown in parentheses.
(60 nm) were collected on 200 mesh nickel grids. Slices were treated(2 × 15 min)with 3% sodiummetaperiodate, quickly rinsed in distilledwater and treated (3 × 5 min) in citrate buffer (19 mgcitric acid/10 ml)pH 6 at room temperature. A final incubation (10 min, 95 °C) in citratebuffer was carried out. After washing in distilled water (3 × 5 min), sec-tions were incubated for 15 min in blocking solution Tris buffered saline(TBS) pH 7.4, 1% BSA and finally incubated overnight with the primaryantibody diluted 1:250 in blocking solution. Grids were then washed in
ylogenetic tree was constructed using MEGA software version 4.0, and the reliability ofB1 and PHB2 are clustered in two branches. The putative protein GenBank accession

Fig. 4. Expression of Cq-PHB in C. quadricarinatus adult tissues. 1: Brain, 2: gills,3: heart, 4: hemocytes, 5: hepatopancreas, 6: testes, 7: stomach, 8: muscle.
Fig. 5. Temporal expression of Cq-PHB in the testes developmental cycle. I: Absence phase,II: developmental phase, III: multiplication phase, IV: mature phase, V: resting phase.
322 D.-A. Fang et al. / Gene 519 (2013) 318–325
TBS (5 × 2 min), in TBS 0.5% BSA (5 × 2 min) and finally in TBS 1% BSA(15 min). Immunocomplexes were visualized by incubation (1 h) withgoat anti-rabbit immunoglobulins conjugated to colloidal gold (20 nmdi-ameter) diluted 1:200 in TBS. After a repeat series of washings, sectionswere stained with uranyl acetate for 15 min and analyzed using a Leo912 Zeiss electron microscope. Controls were incubated with primaryantibody preabsorbed with 10-6 M of the corresponding peptide (1 Vpeptide/10 V TBS). Negatives were scanned by Umax Powerlook 3000flat-bed scanner (Umax Technologies, CA, USA) and edited with AdobePhotoshop CS 3.0.
2.10. Statistical analysis
A multiple comparisons (Duncan's) test was used to compare sig-nificant differences in Cq-PHB gene expression between control andtested samples using SPSS15.0 software. P b 0.05 was considered sta-tistically significant.
3. Results
3.1. Characterization of Cq-PHB
Using degenerate PCR amplification, a cDNA fragment of 302 bpwas obtained (Fig. 1; GenBank accession number: JN168787) thatshowed 95% similarity to PHB of Litopenaeus vannamei and Scyllaparamamosain. Full-length cDNA of Cq-PHB was 1472 bp in lengthand contained a predicted open reading frame (ORF) of 828 bp,beginning with a methionine codon at position 161 and ending witha TGA termination codon at position 987. The 3′-untranslatedregion was 485 bp in length and contained a polyadenylation signal(AATAAA) and a poly (A) tail of 23 bp. The complete sequence wasbeen deposited in GenBank (accession number: HM345982). Theencoded 275 amino acid polypeptide had a calculated molecularmass of 30.3 kDa and a predicted isoelectric point of 6.37.
Using the InterPro search database, the primary structure of PHBprotein was shown to lack motifs typical of signal sequences, nuclearlocalization signals, ATP-binding sites or transcription factors. Com-parisons with PHB genes from other species revealed eight highlyconserved regions, four of which corresponded to binding sites ofknown transcriptional control proteins (CCAAT box, ‘SV40’ sites andtwo Spl sites; Fig. 2).
Blast analysis revealed that Cq-PHB shared high similaritywith otherPHBs from other species, including L. vannamei (88%), S. paramamosain(86%), Drosophila melanogaster (74%), Danio rerio (73%), Gallus gallus(72%), Salmo salar (71%), Bos taurus (71%), Mus musculus (71%), Homosapiens (71%) and Sus scrofa (71%). Furthermore, Cq-PHB shared >60%identity with PHBs from certain other species. The phylogeny of thesePHBs is shown in Fig. 3. The phylogenetic tree contained two distinctbranches, where C. quadricarinatus PHB belongs to PHB1 subfamilyand clusters with other ectotherms, while mammals, birds and fishformed a separate cluster.
3.2. Expression patterns of Cq-PHB transcripts
The mRNA transcript of Cq-PHB was expressed universally in allthe tissues investigated, including the brain, gills, heart, hemocytes,hepatopancreas, testes, stomach and muscles (Fig. 4). Expressionwas greater in the brain and testes, with lower levels in the othertissues.
Temporal expression of Cq-PHB in the testes during the crayfishreproductive cycle was measured using RT-qPCR with18S rRNA act-ing as the internal control. A single peak was observed for bothCq-PHB and 18S rRNA genes at the corresponding melting tempera-ture in the dissociation curves, indicating that the PCR specificallyamplified a single gene. The temporal expression of the Cq-PHBmRNA transcripts is presented in Fig. 5. During the developmental
cycle in the testes, expression of Cq-PHBmRNAwas upregulated dur-ing the developmental phase, and high expression was maintainedduring the spermatogonial multiplication period. Subsequently, ex-pression was downregulated during the mature sperm stage, andlower expression levels were found during the resting phase (P b 0.05).
3.3. Western blot analysis of testes proteins
Anti-PHB and anti-serum antibodies also recognized components ina crude extract of adult C. quadricarinatus testes. Probing of proteinextracts transferred to a nitrocellulose membrane with anti-PHB re-vealed a series of ladder bands characteristic of Cq-PHB. Anti-PHB prob-ing produced bands of approximately 30 kDa to 180 kDa. Cq-PHBprotein expression was higher in the developmental andmultiplicationphases comparedwith the absence andmature phases. No protein com-ponents of the C. quadricarinatus extracts were detected using the con-trol serum from the pre-immunized rabbit (Fig. 6).
3.4. Localization of Cq-PHB
Localization of the Cq-PHBproteinwas studied by IF and IEM.Wholesections of testes stained with hematoxylin and eosin (H&E) and withanti-PHB immunolabeling (counterstained with H&E) are shown inFigs. 7A and B. Positive immunoreactive signals (green fluorescence)for the PHB protein were detected in primary spermatocytes (I),secondary spermatocytes (II) and spermatids (III) of normal maturetestes. Within the testes, the strongest signals for Cq-PHB were foundin primary spermatocytes, with lower positive signals in secondaryspermatocytes, and weak or absent signals in spermatids or maturesperm (Fig. 7. D, III). Moreover, the Cq-PHB protein was concentratedmainly in the cytoplasm of developmental sperm cells. No positive sig-nals were detected in the negative control, which was incubated withpre-immune rabbit serum (Fig. 7B).
Prohibitin was found mainly on the mitochondrial inner membraneof the spermatozoa,with some labeling black particles in thematrix and
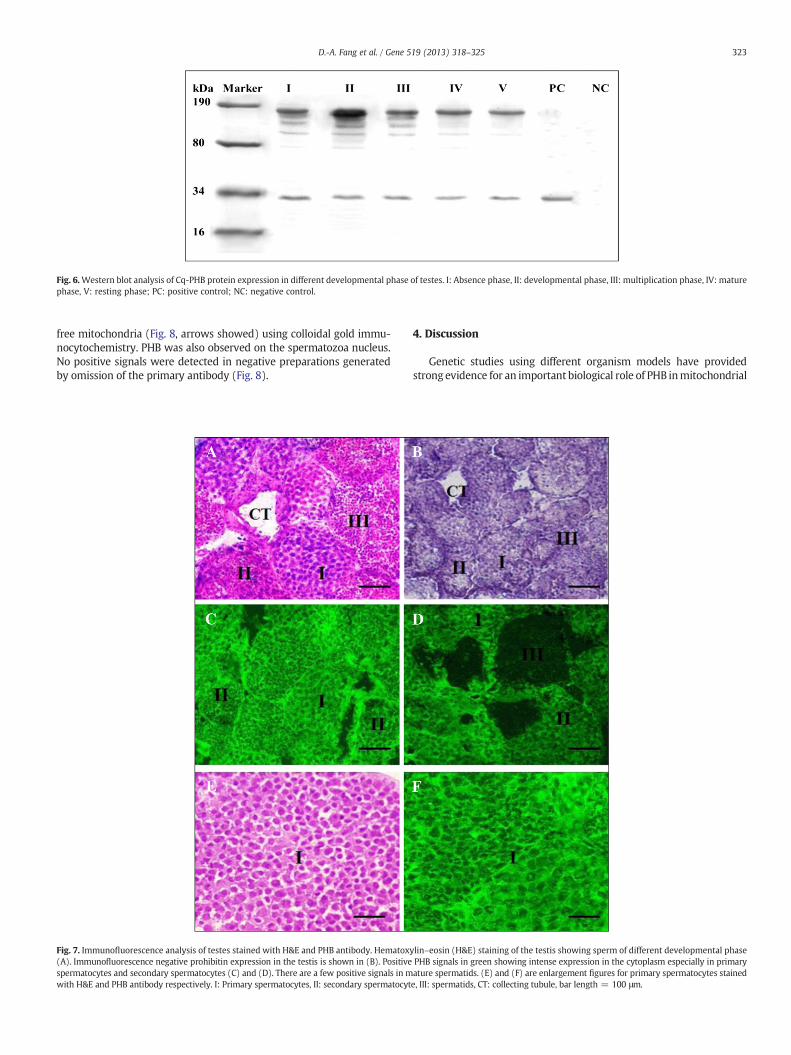
Fig. 6.Western blot analysis of Cq-PHB protein expression in different developmental phase of testes. I: Absence phase, II: developmental phase, III: multiplication phase, IV: maturephase, V: resting phase; PC: positive control; NC: negative control.
323D.-A. Fang et al. / Gene 519 (2013) 318–325
free mitochondria (Fig. 8, arrows showed) using colloidal gold immu-nocytochemistry. PHB was also observed on the spermatozoa nucleus.No positive signals were detected in negative preparations generatedby omission of the primary antibody (Fig. 8).
Fig. 7. Immunofluorescence analysis of testes stained with H&E and PHB antibody. Hematoxy(A). Immunofluorescence negative prohibitin expression in the testis is shown in (B). Positivespermatocytes and secondary spermatocytes (C) and (D). There are a few positive signals in mwith H&E and PHB antibody respectively. I: Primary spermatocytes, II: secondary spermatocyt
4. Discussion
Genetic studies using different organism models have providedstrong evidence for an important biological role of PHB inmitochondrial
lin–eosin (H&E) staining of the testis showing sperm of different developmental phasePHB signals in green showing intense expression in the cytoplasm especially in primaryature spermatids. (E) and (F) are enlargement figures for primary spermatocytes stainede, III: spermatids, CT: collecting tubule, bar length = 100 μm.
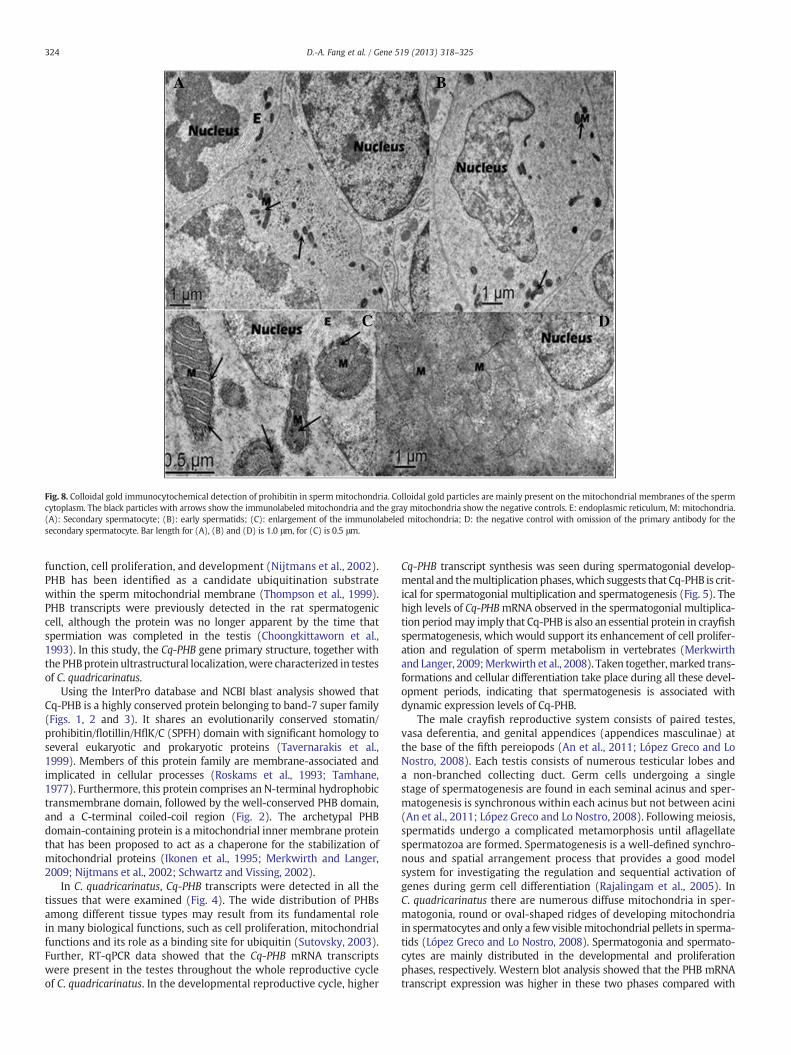
Fig. 8. Colloidal gold immunocytochemical detection of prohibitin in spermmitochondria. Colloidal gold particles are mainly present on the mitochondrial membranes of the spermcytoplasm. The black particles with arrows show the immunolabeled mitochondria and the gray mitochondria show the negative controls. E: endoplasmic reticulum, M: mitochondria.(A): Secondary spermatocyte; (B): early spermatids; (C): enlargement of the immunolabeled mitochondria; D: the negative control with omission of the primary antibody for thesecondary spermatocyte. Bar length for (A), (B) and (D) is 1.0 μm, for (C) is 0.5 μm.
324 D.-A. Fang et al. / Gene 519 (2013) 318–325
function, cell proliferation, and development (Nijtmans et al., 2002).PHB has been identified as a candidate ubiquitination substratewithin the sperm mitochondrial membrane (Thompson et al., 1999).PHB transcripts were previously detected in the rat spermatogeniccell, although the protein was no longer apparent by the time thatspermiation was completed in the testis (Choongkittaworn et al.,1993). In this study, the Cq-PHB gene primary structure, together withthe PHBprotein ultrastructural localization,were characterized in testesof C. quadricarinatus.
Using the InterPro database and NCBI blast analysis showed thatCq-PHB is a highly conserved protein belonging to band-7 super family(Figs. 1, 2 and 3). It shares an evolutionarily conserved stomatin/prohibitin/flotillin/HflK/C (SPFH) domain with significant homology toseveral eukaryotic and prokaryotic proteins (Tavernarakis et al.,1999). Members of this protein family are membrane-associated andimplicated in cellular processes (Roskams et al., 1993; Tamhane,1977). Furthermore, this protein comprises an N-terminal hydrophobictransmembrane domain, followed by the well-conserved PHB domain,and a C-terminal coiled-coil region (Fig. 2). The archetypal PHBdomain-containing protein is a mitochondrial inner membrane proteinthat has been proposed to act as a chaperone for the stabilization ofmitochondrial proteins (Ikonen et al., 1995; Merkwirth and Langer,2009; Nijtmans et al., 2002; Schwartz and Vissing, 2002).
In C. quadricarinatus, Cq-PHB transcripts were detected in all thetissues that were examined (Fig. 4). The wide distribution of PHBsamong different tissue types may result from its fundamental rolein many biological functions, such as cell proliferation, mitochondrialfunctions and its role as a binding site for ubiquitin (Sutovsky, 2003).Further, RT-qPCR data showed that the Cq-PHB mRNA transcriptswere present in the testes throughout the whole reproductive cycleof C. quadricarinatus. In the developmental reproductive cycle, higher
Cq-PHB transcript synthesis was seen during spermatogonial develop-mental and themultiplication phases,which suggests that Cq-PHB is crit-ical for spermatogonial multiplication and spermatogenesis (Fig. 5). Thehigh levels of Cq-PHBmRNA observed in the spermatogonial multiplica-tion periodmay imply that Cq-PHB is also an essential protein in crayfishspermatogenesis, which would support its enhancement of cell prolifer-ation and regulation of sperm metabolism in vertebrates (Merkwirthand Langer, 2009;Merkwirth et al., 2008). Taken together,marked trans-formations and cellular differentiation take place during all these devel-opment periods, indicating that spermatogenesis is associated withdynamic expression levels of Cq-PHB.
The male crayfish reproductive system consists of paired testes,vasa deferentia, and genital appendices (appendices masculinae) atthe base of the fifth pereiopods (An et al., 2011; López Greco and LoNostro, 2008). Each testis consists of numerous testicular lobes anda non-branched collecting duct. Germ cells undergoing a singlestage of spermatogenesis are found in each seminal acinus and sper-matogenesis is synchronous within each acinus but not between acini(An et al., 2011; López Greco and Lo Nostro, 2008). Following meiosis,spermatids undergo a complicated metamorphosis until aflagellatespermatozoa are formed. Spermatogenesis is a well-defined synchro-nous and spatial arrangement process that provides a good modelsystem for investigating the regulation and sequential activation ofgenes during germ cell differentiation (Rajalingam et al., 2005). InC. quadricarinatus there are numerous diffuse mitochondria in sper-matogonia, round or oval-shaped ridges of developing mitochondriain spermatocytes and only a few visiblemitochondrial pellets in sperma-tids (López Greco and Lo Nostro, 2008). Spermatogonia and spermato-cytes are mainly distributed in the developmental and proliferationphases, respectively. Western blot analysis showed that the PHB mRNAtranscript expression was higher in these two phases compared with

325D.-A. Fang et al. / Gene 519 (2013) 318–325
other phases. Interestingly, the immunoblot displayed a pattern consis-tent with speculated polyubiquitinated ladders (Fig. 6). It is possiblethat PHB exists in different molecular weight forms of macromolecularcomplexes through the spermatogenesis process. It also indicated thatthe different levels of ubiquitination occur in the process of spermato-genesis. PHB protein acts as an ubiquitin binding site (Figs. 7 and 8).Ubiquitinated proteins are degraded in the proteasome, thus suggestinga relationship between PHB and protein degradation. This then leads tomitochondrial degradation, suggesting that PHB is related to the mater-nal inheritance of mtDNA. This result is consistent with a previousstudy on bull and rhesus spermatozoa (Sutovsky, 2003; Tatsuta et al.,2005). Thus, these data indicate that PHB has a similar biological functionin crustaceans and other invertebrate animals.
This study demonstrated that PHB is a mitochondrial protein lo-calized in the inner membrane and that it exhibits dynamic changesin spermatogenesis. It is also known that mitochondria exhibit similardynamic changes during spermatogenesis. Thus, it can be speculatedthat PHB may serve as a molecular biomarker of mitochondria. How-ever, the crustacean sperm is immotile and has less mitochondriathan vertebrates, whether a similar mitochondrial inheritance mech-anism in the crustacean and invertebrates exists or not needs furtherresearches to confirm.
5. Conclusion
In the present study, we characterized the molecular structure,expression pattern and some functions of the novel mitochondrial-related gene PHB in the crayfish C. quadricarinatus. This provides an ini-tial step towards understanding ubiquitination during spermatogenesisand the mitochondrial inheritance mechanism in aquatic invertebrateanimals. The results showed that Cq-PHB regulates spermatogenesis,and may explain the molecular mechanisms underlying ubiquitinationof the crayfish sperm. Given the apparent new function for PHB identi-fied in this investigation, namely, the maintenance of mtDNA, degrada-tion of PHB by the ubiquitin–proteasome pathway could facilitate thedegradation of paternalmtDNA and enhance the inheritance ofmaternalmtDNA. Further research is required to elucidate the exact mechanismby which Cq-PHB maintains mtDNA and the biological significance ofthis regulation.
Acknowledgments
This studywas supported by grants from theNational Natural ScienceFoundation of China (No. 30972241) and the National Infrastructure ofFishery Germplasm Resources.
References
Altschul, S.F., et al., 1997. Gapped BLAST and PSI-BLAST: a new generation of proteindatabase search programs. Nucleic Acids Res. 25, 3389–3402.
Altus, M.S., et al., 1995. Regions of evolutionary conservation between the rat andhuman prohibitin-encoding genes. Gene 158, 291–294.
An, C.-G., Weng, X.-L., Xu, Y.-Z., Fan, Y.-J., Zhao, Y.-L., 2011. Histological and ultrastruc-tural studies on the male reproductive system and spermatogenesis in the red clawcrayfish, Cherax quadricarinatus. J. Crustac. Biol. 31, 223–230.
Ankel-Simons, F., Cummins, J.M., 1996. Misconceptions about mitochondria andmammalian fertilization: implications for theories on human evolution. Proc. Natl.Acad. Sci. 93, 13859–13863.
Artal-Sanz, M., Tavernarakis, N., 2009. Prohibitin and mitochondrial biology. TrendsEndocrinol. Metab. 20, 394–401.
Berger, K.H., Yaffe, M.P., 1998. Prohibitin family members interact genetically withmitochondrial inheritance components in Saccharomyces cerevisiae. Mol. Cell.Biol. 18, 4043–4052.
Choongkittaworn, N.M., Kim, K.H., Danner, D.B., Griswold, M.D., 1993. Expression ofprohibitin in rat seminiferous epithelium. Biol. Reprod. 49, 300–310.
Colebatch, G., Cooper, P., East, P., 2002. cDNA cloning of a salivary chymotrypsin-likeprotease and the identification of six additional cDNAs encoding putative digestiveproteases from the green mirid, Creontiades dilutus (Hemiptera: Miridae). InsectBiochem. Mol. Biol. 32, 1065–1075.
Cummins, J., 1998. Mitochondrial DNA in mammalian reproduction. Rev. Reprod. 3,172–182.
Eveleth, D.D., Marsh, J.L., 1986. Sequence and expression of the Cc gene, a member ofthe dopa decarboxylase gene cluster of Drosophila: possible translational regula-tion. Nucleic Acids Res. 14, 6169–6183.
Fusaro, G., Dasgupta, P., Rastogi, S., Joshi, B., Chellappan, S., 2003. Prohibitin induces thetranscriptional activity of p53 and is exported from the nucleus upon apoptotic sig-naling. J. Biol. Chem. 278, 47853–47861.
Holmes, S., 2003. Statistics for phylogenetic trees. Theor. Popul. Biol. 63, 17–32.Huelsenbeck, J.P., Rannala, B., 1997. Phylogenetic methods come of age: testing hy-
potheses in an evolutionary context. Science 276, 227–232.Ikonen, E., Fiedler, K., Parton, R.G., Simons, K., 1995. Prohibitin, an antiproliferative pro-
tein, is localized to mitochondria. FEBS Lett. 358, 273–277.Jones, C.M., 1997. The biology and aquaculture potential of the tropical freshwater frayfish
Cherax quadricarinatus. Department of Primary Industries, Queensland, Australia.Jones, D.T., Taylor, W.R., Thornton, J.M., 1992. The rapid generation of mutation data
matrices from protein sequences. Comput. Appl. Biosci.: CABIOS 8, 275–282.Langer, T., 2000. AAA proteases: cellular machines for degrading membrane proteins.
Trends Biochem. Sci. 25, 247–251.López Greco, L.S., Lo Nostro, F.L., 2008. Structural changes in the spermatophore of the
freshwater ‘red claw’ crayfish Cherax quadricarinatus (Von Martens, 1898)(Decapoda, Parastacidae). Acta Zool. 89, 149–155.
Merkwirth, C., Langer, T., 2009. Prohibitin function within mitochondria: essential rolesfor cell proliferation and cristae morphogenesis. Biochim. Biophys. Acta (BBA)-Mol.Cell Res. 1793, 27–32.
Merkwirth, C., et al., 2008. Prohibitins control cell proliferation and apoptosis by regulatingOPA1-dependent cristae morphogenesis in mitochondria. Genes Dev. 22, 476–488.
Mishra, S., Murphy, L.C., Murphy, L.J., 2006. The prohibitins: emerging roles in diversefunctions. J. Cell Mol. Med. 10, 353–363.
Narasimhan, S., Armstrong, M., McClung, J.K., Richards, F.F., Spicer, E.K., 1997.Prohibitin, a putative negative control element present in Pneumocystis carinii.Infect. Immun. 65, 5125–5130.
Nijtmans, L.G.J., et al., 2000. Prohibitins act as a membrane-bound chaperone for thestabilization of mitochondrial proteins. EMBO J. 19, 2444–2451.
Nijtmans, L.G.J., Artal Sanz, M., Grivell, L.A., Coates, P.J., 2002. The mitochondrial PHBcomplex: roles in mitochondrial respiratory complex assembly, ageing and degen-erative disease. Cell Mol. Life Sci. 59, 143–155.
Nuell, M.J., et al., 1991. Prohibitin, an evolutionarily conserved intracellular protein thatblocks DNA synthesis in normal fibroblasts and HeLa cells. Mol. Cell. Biol. 11,1372–1381.
Piper, P.W., Jones, G.W., Bringloe, D., Harris, N., MacLean, M., Mollapour, M., 2002. Theshortened replicative life span of prohibitin mutants of yeast appears to be due todefective mitochondrial segregation in old mother cells. Aging Cell 1, 149–157.
Rajalingam, K., et al., 2005. Prohibitin is required for Ras-induced Raf-MEK-ERK activa-tion and epithelial cell migration. Nat. Cell Biol. 7, 837–843.
Roskams, A.J.I., et al., 1993. Cell cycle activity and expression of prohibitin mRNA. J. Cell.Physiol. 157, 289–295.
Sanz, M.A., et al., 2003. The mitochondrial prohibitin complex is essential for embryonic via-bility and germline function in Caenorhabditis elegans. J. Biol. Chem. 278, 32091–32099.
Sato, T., et al., 1992. The human prohibitin gene located on chromosome 17q21 is mu-tated in sporadic breast cancer. Cancer Res. 52, 1643–1646.
Schmittgen, T.D., Livak, K.J., 2008. Analyzing real-time PCR data by the comparative CTmethod. Nat. Protoc. 3, 1101–1108.
Schwartz, M., Vissing, J., 2002. Paternal inheritance of mitochondrial DNA. N. Engl. J.Med. 347, 576–580.
Sharma, A., Qadri, A., 2004. Vi polysaccharide of Salmonella typhi targets the prohibitinfamily of molecules in intestinal epithelial cells and suppresses early inflammatoryresponses. Proc. Natl. Acad. Sci. U. S. A. 101, 17492–17497.
Shivaji, S., Kota, V., Siva, A.B., 2009. The role of mitochondrial proteins in sperm capac-itation. J. Reprod. Immunol. 83, 14–18.
Spiropoulos, J., Turnbull, D.M., Chinnery, P.F., 2002. Can mitochondrial DNA mutationscause sperm dysfunction? Mol. Hum. Reprod. 8, 719–721.
Sullivan, J., 2005. Maximum-likelihood methods for phylogeny estimation. In: Elizabeth,A.Z., Eric, H.R. (Eds.), Methods in Enzymology. Academic Press, pp. 757–779.
Sun, L., Liu, L., Yang, X.-J., Wu, Z., 2004. Akt binds prohibitin 2 and relieves its repressionof MyoD and muscle differentiation. J. Cell Sci. 117, 3021–3029.
Sutovsky, P., 2003. Ubiquitin-dependent proteolysis in mammalian spermatogenesis,fertilization, and sperm quality control: killing three birds with one stone. Microsc.Res. Tech. 61, 88–102.
Tamhane, A.C., 1977. Multiple comparisons in model I one-way ANOVA with unequalvariances. Commun. Stat.-Theory Methods 6, 15–32.
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., Kumar, S., 2011. MEGA5:molecular evolutionary genetics analysis using maximum likelihood, evolutionarydistance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739.
Tatsuta, T., Model, K., Langer, T., 2005. Formation of membrane-bound ring complexesby prohibitins in mitochondria. Mol. Biol. Cell 16, 248–259.
Tavernarakis, N., Driscoll, M., Kyrpides, N.C., 1999. The SPFH domain: implicated in reg-ulating targeted protein turnover in stomatins and other membrane-associatedproteins. Trends Biochem. Sci. 24, 425–427.
Terashima, M., et al., 1994. The IgM antigen receptor of B lymphocytes is associatedwith prohibitin and a prohibitin-related protein. EMBO J. 13, 3782–3792.
Theiss, A.L., et al., 2007. Prohibitin protects against oxidative stress in intestinal epithelialcells. FASEB J. 21, 197–206.
Thompson, W.E., Powell, J.M., Whittaker, J.A., Sridaran, R., Thomas, K.H., 1999.Immunolocalization and expression of prohibitin, a mitochondrial associatedprotein within the rat ovaries. Anat. Rec. 256, 40–48.
Zhou, J., et al., 2010. Expression of HSP60 and HSP70 in white shrimp, Litopenaeusvannamei in response to bacterial challenge. J. Invertebr. Pathol. 103, 170–178.




















![Comparative Ultrastructure and Carbohydrate Composition of ... et...It has been suggested for two freshwater crayfish species, Procambarus clarkii and Cherax quadricarinatus [8,9],](https://img.pdfslide.net/doc/110x75/60e359f5e1e7bd32a1046092/comparative-ultrastructure-and-carbohydrate-composition-of-et-it-has-been.jpg)








