Embed Size (px)
Citation preview

The Plant Cell, Vol. 8, 2033-2046, November 1996 O 1996 American Society of Plant Physiologists
Characterization of edsl, a Mutation in Arabidopsis Suppressing Resistance to Peronospora parasíiíca Specified by Severa1 Different RPP Genes
Jane E. Parker,a91i2 Eric B. Holub,b*l Louise N. Frost,' Andem Falk,' Nick D. Gunn,b and Michael J. Danielsa a Sainsbury Laboratory, Norwich Research Park, Colney Lane, Norwich NR4 7UH, United Kingdom b Horticultural Research lnternational, Wellesbourne CV34 9EF, United Kingdom
The interaction between Arabidopsis and the biotrophic oomycete Peronospora parasifica (downy mildew) provides an attractive model pathosystem to identify molecular components of the host that are required for genotype-specific rec- ognition of the parasite. These components are the so-called RPP genes (for Eesistance to E earasifica). Mutational analysis of the ecotype Wassilewskija (Ws-O) revealed an RPPnonspecific locus called EDSl (for gnhanced disease - susceptibility) that is required for the function of RPP genes on chromosomes 3 (RPPlIRPPl4 and R P P l 0 ) and 4 (RPPTP). Genetic analyses demonstrated that the eds7 mutation is recessive and is not a defective allele of any known RPP gene, mapping to the bottom arm of chromosome 3 (43 centimorgans below RPP7/RPP74). Phenotypically, the Ws-edsl mu- tant seedlings supported heavy sporulation by R parasitica isolates that are each diagnostic for one of the RPP genes in wild-type Ws-O; none of the isolates is capable of sporulating on wild-type Ws-O. Ws-edsl seedlings exhibited enhanced susceptibility to some R parasifica isolates when compared with a compatible wild-type ecotype, Columbia, and the eds l parenta1 ecotype, Ws-O. This was observed as earlier initiation of sporulation and elevated production of conidi- osporangia. Surprisingly, cotyledons of Ws-edsl also supported low sporulation by five isolates of R parasifica from Brassica oleracea. These isolates were unable to sporulate on >100 ecotypes of Arabidopsis, including wild-type Ws-O. An isolate of Albugo candida (white blister) from 6. oleracea also sporulated on Ws-edsl, but the mutant exhibited no alteration in phenotype when inoculated with several oomycete isolates from other host species. The bacterial resistance gene RPM7, conferring specific recognition of the avirulence gene avrB from Pseudomonas syringae pv glycinea, was not compromised in Ws-edsl plants. The mutant also retained full responsiveness to the chemical inducer of systemic acquired resistance, 2,6-dichloroisonicotinic acid; Ws-edsl seedlings treated with 2,6-dichloroisonicotinic acid became resistant to the Ws-O-compatible and Ws-O-incompatible R parasifica isolates Emwal and Noc02, respectively. In sum- mary, the EDSl gene appears to be a necessary component of the resistance response specified by several RPP genes and is likely to function upstream from the convergence of disease resistance pathways in Arabidopsis.
INTRODUCTION
Disease resistance in plants to microbial pathogens is often triggered by recognition of the invading pathogen by the host in a race-specific manner (Crute, 1985; Keen, 1990; Pryor and Ellis, 1993). In many cases, this is accompanied by localized host cell death at the site of infection (the so-called hypersen- sitive response [HR]) and a series of defense-related reactions, such as an oxidative burst, cell wall reinforcements, and the accumulation of pathogenesis-related proteins (Lamb, 1994). Although there is a strong correlation between the induction of these responses and the occurrence of disease resistance, their actual role in limiting pathogen growth is not clear.
J.E.P. and E.B.H. contributed equally to this work and are consid- ered first authors. * To whom correspondence should be addressed.
A key objective in plant pathology isto unravel the molecu- lar mechanisms underlying recognition specificity and the associated signaling events leading to disease resistance or susceptibility. The cloning of several race-specific disease re- sistance (R) genes from different dicotyledonous species provided some clues about their possible function and loca- tion in the plant cell (Staskawicz et al., 1995). Their deduced amino acid sequences revealed the presence of either leucine- rich repeats (Bent et al., 1994; Jones et al., 1994; Mindrinos et al., 1994; Whitham et al., 1994; Grant et al., 1995; Lawrence et al., 1995; Dixon et al., 1996), suggesting a role in protein-pro- tein interactions or protein-peptide ligand binding (Kobe and Deisenhofer, 1994), ora serinehhreonine kinase domain (Martin et al., 1993; Zhou et al., 1995), implicating protein phosphory- lation as a key feature of the pathogen recognition process.

2034 The Plant Cell
These possible roles have been reinforced by sequence analysis of the rice %a21 gene for race-specific resistance to Xanthomonas oryzae (Song et al., 1995). Xa27 possesses predicted extracellular leucine-rich repeats, a membrane- spanning domain, and an intracellular porJion containing a ser- inelthreonine kinase motif. Therefore, this molecule appears to exhibit a combined role in both cell surface recognition of the pathogen and in activation of an intracellular defense signal.
The striking similarities in the predicted structural motifs among the cloned R genes suggest that resistance in differ- ent dicotyledonous and monocotyledonous species to a diverse range of microbial pathogens may operate, at least in part, by similar mechanisms. However, the precise course of bio- chemical events and the number of other components required for a particular R gene-mediated signaling pathway are not yet known.
In recent years, Arabidopsis (mouse-ear cress) has been studied intensively as a model host plant for diseases caused by bacterial, fungal, and vira1 pathogens and provides a ge- netically amenable system to examine the various components of disease resistance (Dangl, 1993; Kunkel, 1996). Two R genes specifying race-specific resistance to fseudomonas syringae strains were isolated using a positional cloning strategy (Bent et al., 1994; Mindrinos et al., 1994; Grant et al., 1995). Also, mutational screens for plants altered in resistance have led to the identification of defective mutant alleles of these respec- tive R genes (Kunkel et al., 1993; Yu et al., 1993; Bisgrove et al., 1994) as well as a mutation in an additional gene, NDR7 (for Conspecific disease resistance), encoding a potential signaling component required for the function of several differ- ent race-specific R genes (Century et a\., 1995). Components of systemically induced resistance responses (Bowling et al., 1994; Cao et al., 1994; Delaney et al., 1995) and other poten- tia1 defense pathways (Dietrich et al., 1994; Glazebrook and Ausubel, 1994; Greenberg et al., 1994; Ausubel et al., 1995) have also been characterized in Arabidopsis.
The interaction between Arabidopsis and the biotrophic oomycete Peronospora parasitica (downy mildew) provides an attractive system for the genetic dissection of pathways deter- mining recognition specificity in race-specific resistance and the proper manifestation of the resistance response (Crute et al., 1993, 1994a, 1994b; Holub and Beynon, 1996). P parasii- ica is a parasite exclusive to members of the Cruciferae (Channon, 1981). Natural F! parasitica isolates have been iden- tified that are able to complete their life cyle on certain ecotypes of Arabidopsis by producing asexual and sexual spores (Koch and Slusarenko, 1990; Crute et al., 1993). Additional genetic studies by Holub et al. (1994) demonstrated a large natural variation among Arabidopsis ecotypes in their interaction phenotypes with particular isolates of the parasite. This is char- acteristic of gene-for-gene relationships (Flor, 1971), and the existence of at least 26 different R f f (for resistance to - /? earasitica) genes (known also as recognition specificities) has been predicted from segregation analyses (Parker et al., 1993; Crute et al., 1994a, 1994b; Holub et al., 1994; Tor et al., 1994; Holub and Beynon, 1996: Reignault et al., 1996).
Two RPP loci have been analyzed in detail with respect to recognition of the Noco2 isolate of P parasitica. RPPS was iden- tified in the ecotype Landsberg erecta (Lera) and was mapped to the upper arm of chromosome 4 (Parker et al., 1993). A sec- ond locus, RPP74, in the ecotype Wassilewskija (Ws-O) maps to a cluster of several other RPP genes on the lower arm of chromosome 3 (Reignault et al., 1996). It has not yet been sepa- rated by recombination from RPP7 or RPP70, associated with recognition of isolates Emoy2 and Cala2, respectively (Holub et al., 1994; Holub and Beynon, 1996). The RPPgensmediated resistance responses are characterized by hypersensitive plant cell necrosis initiated in the mesophyll at sites of attempted mycelium ingress. Funga1 development beyond these lesions is severely restricted. In contrast, ecotype Columbia (Col-0) is susceptible to Noco2, and its mycelium is able to spread systemically through the plant tissue and to develop asexual and sexual spores on inoculated tissue within 4 to 7 days.
A mutational approach was adopted to identify genes that are necessary for resistance mediated by RPP5 and RPP74 and to attempt separation of RPP74 from the other closely linked RPP gene specificities. Therefore, we screened mutagenized populations of Ler-O and Ws-O for mutations that caused a change from Noco2 resistance to susceptibility. Here, we de- scribe a recessive mutation of Ws-O called eds7 (for gnhanced - disease gusceptibility), which abolishes the resistance mediated by RPP74 as well as by other linked and unlinked R f f genes present in the Ws-O background. This mutation also partially suppresses resistance of Ws-O to five Brassica oleracea-infecting isolates of P parasitica to which all Arabidop- sis ecotypes so far tested exhibit resistance, implicating a possible common functional role for the EDSl protein in downy mildew resistance in Arabidopsis and Brassica plants. The potential role of EDSl in resistance mediated by RPP genes and in the context of other disease resistance signaling path- ways is discussed.
R ES U LTS
ldentification of Mutations in Ws-O for Susceptibility to R parasitica lsolate Noco2
The cotyledons of wild-type Ws-O seedlings responded after inoculation with the /? parasitica isolate Noco2 by producing distinct necrotic lesions beginning 3 to 4 days after inocula- tion (DAI), as shown in Figure 1A. The interaction phenotype between Ws-O and Noco2 has been described ata later stage (i' DAI) as "a pitting necrosis with no parasite sporulation" (Holub et al., 1994) and is associated with a single recognition locus, RfP74 (Reignault et al., 1996). This locus cosegregates with RPf7 and R f f 7 0 , which specify resistance in Ws-O to isolates EmoyP and CalaZ, respectively (Holub et al., 1994).
A mutational approach was undertaken to identify genes re- quired for RfP74-mediated resistance and to attempt

Disease Resistance Signaling Mutation 2035
separation of RPf74 from RPP7 and RPPlO in Ws-O. To this end, the isolate NocoP was used to screen for NocoB compatible mutations of Ws-O that were altered solely in re- sponse to the selective isolate. Approximately 30,000 M2 seedlings germinated from ethyl methanesulfonate-mutage- nized Ws-O seed were spray inoculated with Noco2; this included 38 M2 seed populations that were each derived in- dependently from pools of 4 0 M1 plants. Six independently mutated M2 plants were selected, and the first of these (EW80-1) is described here. Characterization of RPP74-specific mutants will be presented elsewhere.
Ten cleaved amplified polymorphic DNA sequence (CAPS; Konieczny and Ausubel, 1993) or microsatellite (Bell and Ecker, 1994) markers representing the five Arabidopsis chromosomes were used to confirm that EW80-1 was in fact a mutation of Ws-O and not a contaminant derived from outcross pollination. The DNA profiles of EWBO-1 were identical to those of wild- type Ws-O with each marker and were distinguished from the two NocoP-susceptible ecotypes COLO and Niederzenz (Nd-O) (results not shown). It was therefore highly unlikely that EW80-1 arose from a contaminant seed. M3 progeny were all suscep- tible to Noco2, and a single M3 plant was selected for subsequent genetic and phenotypic analyses.
Genetic Analyses and Mapping of the eds l Mutation in EW80-1
Crosses were made between EW80-1 and the wild-type par- ent Ws-O (containing RPP74) and with the susceptible ecotype Col-5 (Col-O containing g/7, a recessive phenotypic marker for a glabrous mutation). The segregation of resistance and sus- ceptibility was then analyzed in F1 and F2 progeny generated from the two crosses, and the results are shown in Table 1.
In the cross between EW80-1 and Ws-O, all F1 seedlings were resistant to Noco2, indicating that the mutation is reces- sive. This was confirmed by microscopic examination of the inoculated seedlings after staining with lactophenol trypan blue, which preferentially stains fungal structures as well as dead or dying plant cells (Keogh et al., 1980). The results indicated that the degree of plant cell necrosis and fungal development was similar in F1 and wild-type Ws-O seedlings (data not shown). Analysis of F2 EW80-1 x Ws-O progeny confirmed that EW80-1 contains a single recessive mutation because there was a 3:l segregation of resistance to susceptibility.
The mutation that enhances disease susceptibility in EW80-1 is referred to hereafter as edsl, based on evidence from the phenotypic analyses described below. The Ws-O mutant line EW80-1 is accordingly named Wsedsl. eds has also been used to name several newly identified mutant loci that confer en- hanced susceptibility in COLO to a mildly virulent strain of the bacterial pathogen P syringae pv maculicola (Glazebrook et ai., 1996; see also Discussion).
In the cross between Ws-edsl (EWSO-1) and Col-5, all F1 seedlings were also phenotypically resistant to Noco2 (Table l), indicating that the mutation was complemented by a gene
in Col-5 and was therefore not an allele of RPP74. Segrega- tion analysis of Fp progeny for this cross (Table 1) showed that the ratio of resistant-to-susceptible plants is consistent with the presence of a single recessive mutation in Ws-O at a sec- ond locus required for RPPN function. Linkage of the mutation with RfP14 was inconclusive from these genetic data.
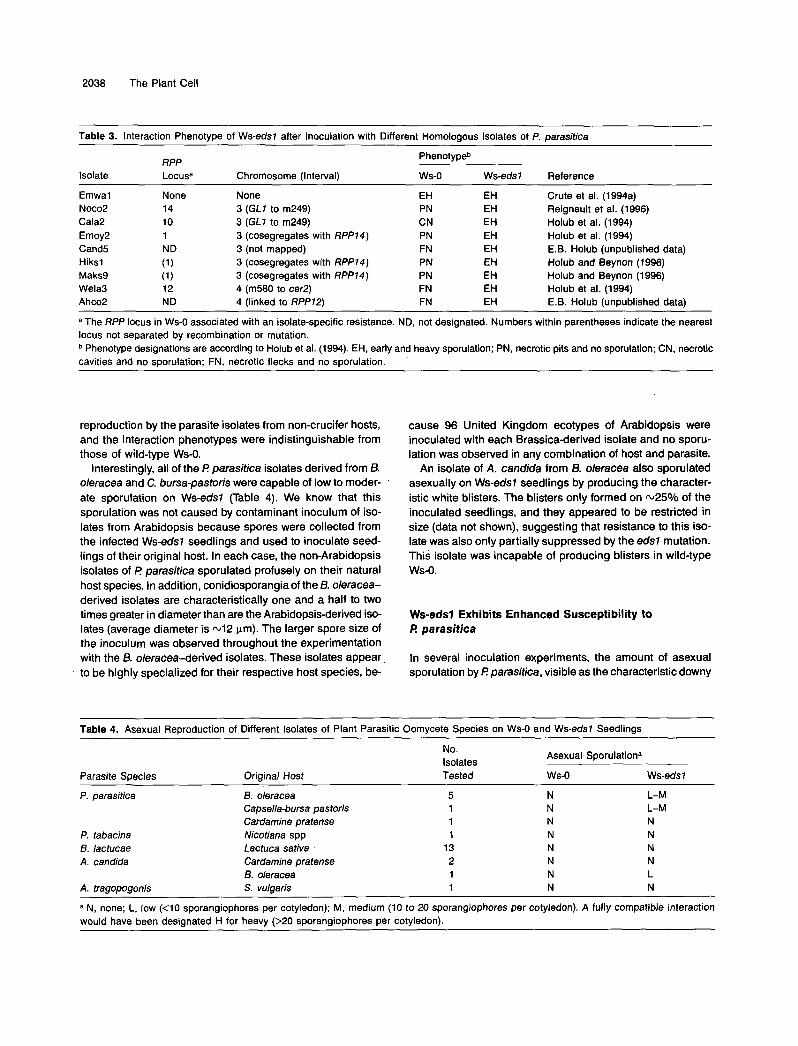
Molecular markers and the glabrous phenotype of g/1 were used to determine a map location for the edsl mutation. Ap- proximately 130 F2 progeny from the Col-5 x EWBO-1 cross, which were segregating for both RPP74 and EDS7, were selected for full Noco2 susceptibility. The selected plants were then further analyzed to determine the presence of the RPPN gene from Ws-O by using closely linked molecular markers. These markers were ve021, lying 1.8 centimorgans (cM) above RPP74, and pAT3-89.1, positioned 1.4 cM below RPP14, as de- scribed by Reignault et al. (1996). F2 plants that were either homozygous or heterozygous at RPP74 were then used for mapping the position of EDS7 relative to CAPS (Konieczny and Ausubel, 1993) and microsatellite (Bell and Ecker, 1994) mark- ers. Linkage to GL7 and other markers on chromosome 3 was identified as shown in Table 2. No linkage was found to mark- ers on chromosomes 1,2,4, and 5 (see Table 2). Examination of the recombination frequencies between EDSl and chromo- some 3 markers and evidence of recombination break points in individual F3 families placed the gene in a 25-cM interval below RPP74, 18 cM above BGL7 and 7 cM below m249.
edsl Suppresses the Function of RPPl4 and Severa1 Other RPP Genes in Ws-O
Detailed phenotypic characterization of the response of Ws- edsl seedlings to Noco2 in comparison to the wild-type resis- tant parent Ws-O and the susceptible ecotype (201-5 was performed with seedlings that had been taken through one parenta1 backcross. In contrast to the necrotic resistant re- sponse of Ws-O cotyledons after inoculation with Nocoe (Figure lA), cotyledons of 9-daydd Wseds7 seedlings supported early (visible 3 DAI) and heavy (by 7 DAI) asexual sporulation of Noco2 in the absence of visible plant cell necrosis (Figure lB). Microscopic analysis of Ws-O cotyledons stained with lac- tophenol trypan blue showed hypersensitive cell death of individual mesophyll cells penetrated by NocoP 24 hr after in- oculation (Figure 1C). This contrasted greatly with Ws-edsl cotyledons in which the Noco2 mycelium had grown unimpeded (Figure 1D). Three days after inoculation, further cell death had occurred in Ws-O cotyledons, and no fungal de- velopment was observed beyond these necrotic areas (Figure lE), whereas Ws-edsl cotyledons were fully infected and the mycelium had grown extensively into the first true leaves (Fig- ure 1F).
Previous genetic studies have identified several other RPP gene specificities in Ws-O. Each recognizes a particular iso- late of /? parasitica, as shown in Table 3. Like RPP74, most of the specificities were mapped to chromosome 3. However, two mapped to chromosome 4. Therefore, it was worthwhile

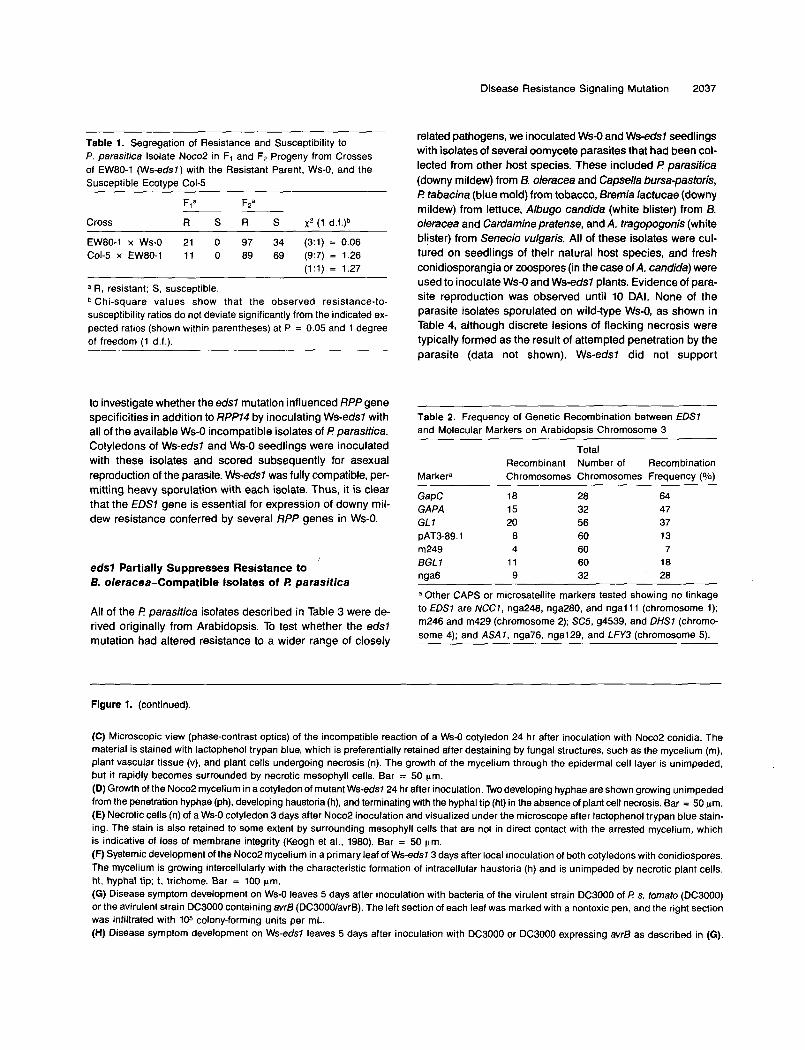
Figure 1. Responses of Ws-0 and Ws-eds7 (EW80-1) to Inoculation with P. parasitica Isolate Noco2 and P syringae pv tomato
(A) Incompatible reaction of a Ws-0 cotyledon 5 days after inoculation with a droplet of conidiospores from P. parasitica isolate Noco2. Necroticlesions start to appear macroscopically after 3 days.(B) Compatible reaction of a cotyledon of mutant Ws-eds7 5 days after inoculation with Noco2 conidiospores. Aerially borne conidiospores developon sporangiophores growing out through the stomata. The first sporangiophores are detectable after 3 days.
Disease Resistance Signaling Mutation 2037
Table 1. Segregation of Resistance and Susceptibility to P. parasitica lsolate Noco2 in F, and F2 Progeny from Crosses of EW80-1 (Ws-eds7) with the Resistant Parent, Ws-O, and the Susceptible Ecotype Col-5
Fla Fza
Cross R S R S x2 (1 d.fJb _ _ _ _ _ _
EW80-1 x WS-O 21 O 97 34 (3:l) = 0.06 COI-5 x EW80-1 11 O 89 69 (9:7) = 1.26
(1:l) = 1.27
a R, resistant; S , susceptible. Chi-square values show that the observed resistance-to-
susceptibility ratios do not deviate significantly from the indicated ex- pected ratios (shown within parentheses) at P = 0.05 and 1 degree of freedom (1 d.f.).
to investigate whether the edsl mutation influenced RPPgene specificities in addition to RPP74 by inoculating Ws-edsl with all of the available Ws-O incompatible isolates of F! parasitica. Cotyledons of Ws-edsl and Ws-O seedlings were inoculated with these isolates and scored subsequently for asexual reproduction of the parasite. Ws-edsl was fully compatible, per- mitting heavy sporulation with each isolate. Thus, it is clear that the EDSl gene is essential for expression of downy mil- dew resistance conferred by several RPP genes in Ws-O.
edsl Partially Suppresses Resistance to B. oleracea-Compatible lsolates of I! parasitica
All of the F! parasitica isolates described in Table 3 were de- rived originally from Arabidopsis. To test whether the edsl mutation had altered resistance to a wider range of closely
related pathogens, we inoculated Ws-O and Ws-edsl seedlings with isolates of several oomycete parasites that had been col- lected from other host species. These included F! parasitíca (downy rnildew) from 6. oleracea and Capsella bursa-pastoris, F! tabacina (blue mold) from tobacco, Bremia lactucae (downy mildew) from lettuce, Albugo candida (white blister) from 6. oleracea and Cardamine pratense, and A. tragopogonis (white blister) from Senecio vulgaris, All of these isolates were cul- tured on seedlings of their natural host species, and fresh conidiosporangia or zoospores (in the case of A. candida) were used to inoculate Ws-O and Ws-edsl plants. Evidence of para- site reproduction was observed until 10 DAI. None of the parasite isolates sporulated on wild-type Ws-O, as shown in Table 4, although discrete lesions of flecking necrosis were typically formed as the result of atternpted penetration by the parasite (data not shown). Ws-edsl did not support
Table 2. Frequency of Genetic Recombination between €DS7 and Molecular Markers on Arabidopsis Chromosome 3
Total Recombinant Number of Recombination
Markera Chromosomes Chromosomes Frequency (010)
GapC 18 28 64 GAPA 15 32 47 GL7 20 56 37 pAT3-89.1 8 60 13 m249 4 60 7 BGL 7 11 60 18 nga6 9 32 28
a Other CAPS or microsatellite markers tested showing no linkage to €DSl are NCC7, nga248, nga280, and ngal 1 1 (chromosome 1); m246 and m429 (chromosome 2); SC5, 94539, and DHSl (chromo- some 4); and ASA7, nga76, nga129, and LFY3 (chromosome 5).
Figure 1. (continued).
(C) Microscopic view (phase-contrast optics) of the incompatible reaction of a Ws-O cotyledon 24 hr after inoculation with Noco2 conidia. The material is stained with lactophenol trypan blue, which is preferentially retained after destaining by funga1 structures, such as the mycelium (m), plant vascular tissue (v), and plant cells undergoing necrosis (n). The growth of the mycelium through the epidermal cell layer is unimpeded, but it rapidly becomes surrounded by necrotic mesophyll cells. Bar = 50 pm. (D) Growth of the NocoP mycelium in a cotyledon of mutant Ws-eds724 hr after inoculation. Two developing hyphae are shown growing unimpeded from the penetration hyphae (ph), developing haustoria (h), and terminating with the hyphal tip (ht) in the absence of plant cell necrosis. Bar = 50 wm. (E) Necrotic cells (n) of a Ws-O cotyledon 3 days after NocoP inoculation and visualized under the microscope after lactophenol trypan blue stain- ing. The stain is also retained to some extent by surrounding mesophyll cells that are not in direct contact with the arrested mycelium, which is indicative of loss of membrane integrity (Keogh et al., 1980). Bar = 50 pm. (F) Systemic development of the NocoP mycelium in a primary leaf of Ws-eds73 days after local inoculation of both cotyledons with conidiospores. The mycelium is growing intercellularly with the characteristic formation of intracellular haustoria (h) and is unimpeded by necrotic plant cells. ht, hyphal tip; t, trichome. Bar = 100 ptn.
(G) Disease symptom development on Ws-O leaves 5 days after inoculation with bacteria of the virulent strain DC3000 of I? s. romaro (DC3000) or the avirulent strain DC3000 containing avr6 (DC3000/avrB). The left section of each leaf was marked with a nontoxic pen, and the right section was infiltrated with 105 colony-forming units per mL. (H) Disease symptom development on Ws-eds7 leaves 5 days after inoculation with DC3000 or DC3000 expressing avr6 as described in (G).
2038 The Plant Cell
Table 3. lnteraction Phenotype of Ws-edsl after lnoculation with Different Homologous lsolates of P. parasitica
RPP Phenotypeb
lsolate LOCUSa Chromosome (Interval) ws-o Ws-edsl Reference
Emwal Noco2 Cala2 Emoy2 Cand5 Hiksl Maks9 Wela3 Ahco2
None 14 10 1 ND (1) (1) 12 ND
None 3 (GL7 to m249) 3 (GL7 to 111249) 3 (cosegregates with RPP74) 3 (not mapped) 3 (cosegregates with RPP74) 3 (cosegregates with RPP74) 4 (m580 to cer2) 4 (linked to RPP72)
EH PN CN PN FN PN PN FN FN
EH EH EH EH EH EH EH EH EH
Crute et al. (1994a) Reignault et al. (1996) Holub et al. (1994) Holub et al. (1994) E.B. Holub (unpublished data) Holub and Beynon (1996) Holub and Beynon (1996) Holub et al. (1994) E.B. Holub (unpublished data)
a The RPP locus in Ws-O associated with an isolate-specific resistance. ND, not designated. Numbers within parentheses indicate the nearest locus not separated by recombination or mutation.
Phenotype designations are according to Holub et al. (1994). EH, early and heavy sporulation; PN, necrotic pits and no sporulation; CN, necrotic cavities and no sporulation; FN, necrotic flecks and no sporulation.
reproduction by the parasite isolates from non-crucifer hosts, and the interaction phenotypes were indistinguishable from those of wild-type Ws-O.
Interestingly, all of the P parasitica isolates derived from B. oleracea and C. bursa-pastoris were capable of low to moder- ate sporulation on Ws-edsl (Table 4). We know that this sporulation was not caused by contaminant inoculum of iso- lates from Arabidopsis because spores were collected from the infected Ws-edsl seedlings and used to inoculate seed- lings of their original host. In each case, the non-Arabidopsis isolates of F! parasitica sporulated profusely on their natural host species. In addition, conidiosporangia of the B. oleracea- derived isolates are characteristically one and a half to two times greater in diameter than are the Arabidopsis-derived iso- lates (average diameter is -12 fim). The larger spore size of the inoculum was observed throughout the experimentation with the B. oleracea-derived isolates. These isolates appear to be highly specialized for their respective host species, be-
cause 96 United Kingdom ecotypes of Arabidopsis were inoculated with each Brassica-derived isolate and no sporu- lation was observed in any combination of host and parasite.
An isolate of A. candida from B. oleracea also sporulated asexually on Ws-edsl seedlings by producing the character- istic white blisters. The blisters only formed on -25% of the inoculated seedlings, and they appeared to be restricted in size (data not shown), suggesting that resistance to this iso- late was also only partially suppressed by the edsl mutation. This isolate was incapable of producing blisters in wild-type ws-o.
Ws-edsl Exhibits Enhanced Susceptibility to R parasitica
In severa1 inoculation experiments, the amount of asexual sporulation by F! parasitica, visible as the characteristic downy
Table 4. Asexual Reproduction of Different lsolates of Plant Parasitic Oomycete Species on Ws-O and Ws-eds7 Seedlings
Parasite SDecies Original Host
No. lsolates Tested ws-o Ws-eds 1
Asexual Sporulationa
P. parasitica
P. tabacina B. lactucae A. candida
A. tragopogonis
B. oleracea Capsella-bursa pastoris Cardamine pratense Nicotiana spp Lactuca sativa Cardamine pratense B. oleracea S. vulgaris
5 1 1 1
13 2 1 1
L-M L-M N N N N L N
a N, none; L, low (<I0 sporangiophores per cotyledon); M, medium (10 to 20 sporangiophores per cotyledon). A fully compatible interaction would have been designated H for heavy (>20 sporangiophores per cotyledon).

Disease Resistance Signaling Mutation 2039
Table 5. Conidiosporangia Production of P. parasitica lsolates in Wild-Type Ws-O and Ws-edsl (EW80-1) Seedlings at Different Developmental Stages
Conidiosporangia per Seedlingd
ws-o Ws-edsl Harvest P. parasitica
Experimenta lnoculated Tissue (Age) lsolateC Mean SE Mean SE
1 Cotyledons 7 Emco5 3,333 43 3,567 460 (7 days old) Ahco2 O - 3,933 12
Emoy2 O - 3,467 12 P-O06 O - NQ -
2 Cotyledons and juvenile leaves 3 Emwal 195 138 865 322 (9 days old) Noco2 O - 5,000 812
3 Cotyledons and juvenile leaves 6 Emwal 14,695 3,122 16,938 2,691 (9 days old) Noco2 O - 19,188 2,496
4 Cotyledons and juvenile leaves 10 Emco5 13,900 320 21,967 478 (14 days old) Ahco2 O - 22,200 493
Emoy2 O - 5,300 21 o P-O06 O - NQ -
a Experiments 1 and 4 were conducted simultaneously at Horticultural Research lnternational by using 7- (before emergence of the first juvenile leaves) and 14-day-old seedlings, respectively. Experiments 2 and 3 were conducted simultaneously at the Sainsbury Laboratory by using 9-day-old seedlings for inoculation. At this age, the first juvenile leaves were just beginning to emerge. b Days after inoculation when conidiosporangia were harvested. C lsolates Emco5 and Emwal are compatible with wild-type Ws-O. lsolates Emoy2 and Noco2 are recognized by different RPP genes (RPP1 and RPP74, respectively) on chromosome 3 in Ws-O. Ahco2 is recognized by an unnamed RPP gene on chromosome 4 in Ws-O. All of these isolates were originally collected from Arabidopsis; isolate P-O06 was collected from E. olefacea.
For experiments 2 and 3, the means were calculated from five samples that each contained conidiosporangia from 20 infected seedlings. For experiments 1 and 4, the means were calculated from six samples that each contained conidiosporangia from 1 O infected seedlings. lsolate P-O06 consistently produced low numbers of sporangiophores on Ws-edsl, but the number of conidiosporangia produced could not be quanti- fied (NO). Dashes indicate that values were not measurable. The method for quantifying conidiosporangia is described in Dangl et al. (1992a).
mildew, appeared to be greater in Wsedsl seedlings compared with sporulation by the same Ws-incompatible isolate, such as Noco2, in the naturally compatible ecotype COLO. An in- creased capacity for asexual sporulation was also observed on Ws-edsl seedlings when compared with sporulation on wild- type Ws-O by naturally Ws-compatible isolates, such as Emwal or Emco5. Ws-incompatible isolates, such as Noco2, Emoy2, Ahco2, and P-006, consistently initiated production of sporan- giophores (trichome-like structures bearing asexual propagules called conidiosporangia) in cotyledons of Ws-edsl by 2 or 3 DAI, which was 24 hr earlier than in a naturally compatible ecotype. Likewise, Ws-compatible isolates (Emwal and Emco5) consistently produced sporangiophores 24 hr earlier in Ws- edsl than in wild-type Ws-O. However, the number of sporan- giophores 7 DAI was always to0 dense to make accurate comparisons.
Quantification of conidiosporangia provided a more tangi- ble means of assessing /? parasifica development in Ws-O and Ws-edsl seedlings, and the results of four experiments, undertaken between the Sainsbury Laboratory and Horticul- tural Research International-East Malling with seedlings of different ages, are shown in Table 5. No differences between Ws-O and Ws-edsl were observed in the numbers of conidi- osporangia produced on 7-day-old seedlings before the
emergence of juvenile leaves (Table 5, experiment 1). How- ever, significant differences were observed when seedlings were inoculated at a stage after the juvenile leaves began to emerge (Table 5, experiments 2 to 4), especially with inocula- tion of 14-day-old seedlings. For example, the Ws-compatible isolate Emco5 produced on average 60% more conidiosporan- gia in Ws-edsl than in wild-type Ws-O. The Ws-incompatible isolate AhcoP produced an amount similar to Emco5 in the mutant. However, Emoy2, a second Ws-incompatible isolate, was not as prolific on Ws-edsl, suggesting that partia1 downy mildew resistance remains within true leaves of Ws-edsl in response to inoculation with this isolate. Altogether, the results confirmed our previous observations of enhanced suscepti- bility in Ws-edsl with respect to the Wscompatible I? parasirica isolates Emwal and Emco5. However, this phenotype was clearly influenced by seedling age.
Resistance to the Bacterial Avirulence Gene avr8 1s Not Affected by edsl
Wild-type Ws-O plants are susceptible to disease caused by strain DC3000 of the bacterial pathogen F! s. pv &"to (Whalen et al., 1991) but are resistant to the same strain harboring the

2040 The Plant Cell
avirulence gene avrB derived from the soybean pathogen P s. pv glycinea (Innes et al., 1993). Therefore, we inoculated leaves of Ws-edsl and wild-type Ws-O plants with the patho- genic strain DC3000 or strain DC3000 expressing avr6 to assess whether the mutant was compromised in race-specific resistance to this bacterial avirulence gene. Leaves of plants were infiltrated with low concentrations of bacteria (105 colony- forming units per mL) and monitored for disease symptom de- velopment and growth of the bacteria in the inoculated leaves. Severe symptoms developed over 5 days in Ws-O and Ws-edsl inoculated with DC3000. Interestingly, the progression of chlo- rosis and subsequent necrosis of the infected tissue was 24 hr more advanced in Ws-edsl than in Ws-O, as shown in Figures 1G and lH, again suggesting an enhanced susceptibility(see above) of Ws-edsl to the virulent strain DC3000. Also, the growth rate of DC3000 in Ws-edsl was marginally greater than in Ws-O, although by day 4 the number of bacteria extracted from infected leaves was not significantly different, as shown in Figure 2.
Ws-O and Ws-edsl plants were equally resistant to DC3000 expressing avr6, as measured by the absence of disease symp- toms (Figures 1G and 1H) and by a 100- to 1000-fold reduction in bacterial number in inoculated leaves over 4 days in both plant lines (Figure 2). Leaves were also inoculated with a con- centration of bacteria (5 x 107 colony-forming units per mL)
-
P.S. tomato DC3000/evr~
1 1 I
O 1 2 3 4 5
Time after infiltration (days)
Figure 2. Growth of Virulent and Avirulent Strains of f? s. tomato in Leaves of Ws-O and Ws-edsl (EWBO-1).
Leaves of Ws-O (O and A) or Ws-edsl (O and A) were infiltrated with 105 colony-forming units (cfu) per mL of the virulent strain DC3000 or the avirulent strain DC3000 expressing avrí3. Concentrations of viable bacteria were measured in the leaves at various time points after inoculation.
that gave rise to a macroscopically visible, local hypersensi- tive plant reaction in the case of the avirulent strain DC3000 expressing avr6 on Ws-O (Innes et al., 1993). This was clearly distinguishable at 16 to 20 hr from the virulent strain DC3000 on Ws-O, which was symptomless at this early time point (results not shown). The timing and appearance of the HR were the same in Ws-O and Ws-edsl (data not shown). As a control, leaves of the natural avr6-susceptible ecotype Blanes (Ela-2; lnnes et al., 1993) were inoculated under the same conditions to test the pathogenic competence of DC3000 expressing avr6, and these produced disease symptoms similar to Ws-O (results not shown). Therefore, we concluded that Ws-edsl retains full resistance to a P syringae avirulence gene avr6, although it appears to allow more rapid disease lesion formation than does the wild-type plant with the virulent strain DC3000 of F? s. fomato.
Ws-edsí Retains Responsiveness to the Chemical lnducer of Systemic Acquired Resistance, 2,6-Dichloroisonicotinic Acid
An inducible plant defense mechanism known as systemic ac- quired resistance (SAR) has been analyzed in detail in tobacco and Arabidopsis plants (Ryals et al., 1994). It is activated by exposure of the plants to necrosis-inducing pathogens and leads to the accumulation of endogenous salicylic acid (SA), a vital component of SAR, increased expression of patho- genesis-related genes, and systemic resistance to a broad spectrum of pathogens (Ward et al., 1991; Gaffney et al., 1993; Uknes et al., 1993; Delaney et al., 1994). SAR is also fully in- duced by the exogenous application of the chemical compound 2,6-dichloroisonicotinic acid (INA; Uknes et al., 1992), which acts as an analog of SA or downstream of SA perception or binding (Vernooij et al., 1995). Therefore, we tested whether Ws-edsl was compromised in its ability to activate SAR after INA application by measuring its response to the genetically compatible I? parasifica isolate Emwal and to the Ws- edsl-compatible isolate Noco2. INA was sprayed onto seed- lings 2 days before spray inoculating with Emwal, Noco2, or water. The results in Table 6 show that INA treatment induced almost complete resistance to Emwal in both wild-type Ws-O and Ws-edsl leaves and to Noco2 in Ws-edsl leaves. These tests indicated that the SAR response pathway, from the point of perception of the inducer INA, is intact in Ws-edsl.
DISCUSSION
By using a mutational approach with Arabidopsis, we have identified a gene, EDSl, that is essential for the expression of severa1 naturally polymorphic RPP genes required for race- specific resistance to the biotrophic parasite P parasitica. The RPP genes influenced by the edsl mutation are located on at least two Arabidopsis chromosomes and specify a range

Disease Resistance Signaling Mutation 2041
Table 6. Development of P. parasitica lsolates Emwal and Noco2 in Ws-O and Ws-edsl Seedlings Pretreated with INA
Asexual Sporulationa
ws-o Ws-edsl
DAI lsolate - lNAb + INA - INA + INA
3 Emwal - - - -
4 Emwal + - + -
7 Emwal + + + (+ ) + + + (+ )
Noco2 - - -
Noco2 - - +
-
-
Noco2 - - + + + (+ )
a The number of sporangiophores was assessed on individual leaves of at least 12 plants per treatment: - , none; (+), rare (1 to 5 on <10% of the leaves); + , low (1 to 10 on < 50% of the leaves); + + , medi- um (5 to 20 on >75% of the leaves); + + + , heavy (>25 on >90% of the leaves).
INA ( + INA) or the control wettable powder formulation ( - INA) at 25 mglL was sprayed to imminent runoff on 21-day-old plants.
of interaction phenotypes in Ws-O with respect to infections by corresponding isolates of F! parasifica. In light of recent ad- vances in understanding the molecular basis of disease resistance, we propose that the EDSl gene product is a neces- sary component in the signal transduction pathways that are specified by several but not necessarily all R f f genes.
Crosses have been initiated between Ws-edsl and selected ecotypes of Arabidopsis to determine whether other R f P genes, identified in these ecotypes, also require a functional EDSl gene. Recently, a second edsl allele has been identi- fied in the ecotype Ler-O. It exhibits complete suppression of Noco2 recognition conferred by the RfP5 gene located on chro- mosome 4 (J.E. Parker, unpublished data), reinforcing the notion that EDSl encodes a common signaling component re- quired by several different R f P genes. Interestingly, however, this mutant allele does not fully suppress the function of all RPP genes in Ler-O (E.B. Holub and J.E. Parker, unpublished data). These results suggest that there may be RPP gene- mediated resistance pathways that are, at least in part, inde- pendent of EDSl function. A more detailed examination of individual RPP gene-specified phenotypes in Ler-edsl and wild-type Ler-O plants is now in progress. Also, transfer of RPf genes from the Ler4 to the Ws-edsl background using marker- assisted selection will allow us to test more rigorously whether functional differences in specificity can occur between mu- tant alleles of EDSl.
The edsl mutation also causes a reduction in the leve1 of resistance exhibited by Ws-O to isolates of I? parasitica that are pathogens of 8. oleracea. Therefore, we concluded that €OS7 may be required for socalled nonhost resistance to these particular F! parasifica isolates. This suggests that function- ally common defense signaling components may exist between these two crucifer species in response to F! parasifica chal-
lenge. It follows that EDSI-responsive R f f genes are present in the Arabidopsis genome and that they recognize F! parasif- ica pathogens of 6. oleracea. This promises well for intergeneric transfer of potentially useful disease R genes between Arabid- opsis and Brassica species.
The intermediate phenotype of Ws-edsl, after inoculations with B. oleracea-derived isolates, may reflect a deficiency in compatibility factors required for full pathogenesis of a heter- ologous I? parasifica isolate on Arabidopsis. More importantly, it suggests the presence of at least another gene in Ws-0 that is nonresponsive to EDSl and required for resistance to het- erologous isolates. A possible alternative explanation is that this particular edsl mutant allele intrinsically has a less se- vere effect on nonhost F! parasifica-triggered resistance. Several other oomycete parasite species, such as B. lactucae, which causes downy mildew on lettuce (Michelmore et al., 1988), were not able to cause infection in Ws-edsl (Table 4); it may be that these are evolutionarily less related to the Arabidopsis- and 8. oleracea-infecting F! parasifica isolates tested here. The data, however, demonstrate a link between race-specific and nonhost disease resistance, as has been shown in earlier studies with several P syringae avr genes (Kobayashi et al., 1989; Whalen et al., 1991; Dangl et al., 1992b; Fillingham et al., 1992; lnnes et al., 1993; Wood et al., 1994).
Several recently characterized defense response mutations in Arabidopsis are instructive for anticipating the possible role of EDSl in disease resistance. The first mutation, ndrl (Century et al., 1995), suppresses resistance in COLO to four F! syrin- gae avirulence genes, avrRpf2, avrB, avrRpml, and avrPph3, as well as exhibiting variable suppression of resistance to sev- era1 incompatible isolates of I? parasifica. This provides important evidence for the existence of a common step in re- sistance to prokaryotic and eukaryotic pathogens, and supports the hypothesis that convergent signaling pathways exist down- stream of events presumably triggered by different R genes. Our studies clearly show functional differences between EDSl and NDRl. Unlike COLO plants containing the ndrl mutation, Ws-eds7 seedlings retain full resistance to the bacterial aviru- lence gene avrB from F! s. glycinea. This is demonstrated by suppression of avirulent bacterial growth and the absence of disease symptoms at low inoculum concentrations that are comparable to wild-type Ws-O plants. The avirulent bacterial strain was also capable of inciting a hypersensitive (HR) necrotic lesion at high inoculum doses in Ws-edsl with the same timing as in Ws-O. A fuller spectrum of bacterial aviru- lence genes, such as avrRpml, avrRpt2, and avrfph3 (Century et al., 1995) as well as avrRps4 (Hinsch and Staskawicz, 1996), will be tested on both the Ws-edsl and Ler-edsl plants to as- sess the dependence of the corresponding R genes on EDSl function. Experiments have also been initiated to compare the requirements for NDRl and EDSl by the same spectrum of R f P genes in near isogenic backgrounds. This will enable us to assess more critically whether their signaling functions over- lap or are mutually exclusive.
Two other recessive mutations, nprl (for gonexpresser of pathogenesis-related genes; Cao et al., 1994) and niml (for

2042 The Plant Cell
- noninducible hmunity; Delaney et al., 1995), identify gene products that are required for the SAR response in Arabidop- sis and are postulated to be at the site of SA or INA activation of systemic defense responses or at some point downstream. Interestingly, Col-nprl seedlings exhibit strong suppression of several RPP genes (E.B. Holub, unpublished data) as well as suppressing resistance to I? syringae (Cao et al., 1994). Delaney et al. (1995) also provide evidence for a requirement of the wild-type NlMl gene in race-specific resistance. In their study, local, genetically determined resistance to Noco2, in the absence of SAR-inducing agents, is relaxed in niml when compared with Noco2 resistance in the wild-type Ws-O plant.
Our study shows that Ws-edsl plants are fully responsive to INA, becoming systemically resistant to the normally com- patible /? parasitica isolate Emwal and to the Ws-edsl-com- patible isolate Noco2. Therefore, edsl identifies a signaling component that appears to be distinct from NlMl or NPRl. The results are consistent with the SAR pathway being intact in Ws-edsl plants, although the phenotype does not rule out the possibility that edsl leads to a reduction in endogenous SA levels, which would be compensated for by the application of INA (Venooij et al., 1995). However, this is unlikely because it would result in an overall suppression of race-specific resis- tance to avirulent bacterial and fungal isolates, which has been demonstrated in plants engineered to degrade SA (Gaffney et al., 1993; Delaney et al., 1994). We show here that although the enhanced susceptibility phenotype of Ws-edsl to virulent isolates of I? parasitica is reminiscent of the response of SA- depleted Arabidopsis plants (Delaney et al., 1994), resistance is retained to the genetically incompatible I? s. tomato strain expressing avrf3.
In summary, if race-specific resistance can occur in plants as a series of converging pathways that begin with specific recognition of the pathogen, which in some way activate de- fense responses, then it is likely that EDSl controls a common step downstream of several RPP genes but upstream of more broadly convergent events identified by mutations, such as niml and nprl, or possibly ndrl, and acts at a step before SA ac- cumulation and SA-activated responses. It will be instructive to measure both endogenous SA levels and the accumulation of pathogenesis-related protein transcripts in Ws-edsl plants under various inducing and pathogen-challenged conditions to test this model. Currently, this is certainly a far to0 simplis- tic model of biochemical events. Indeed, there is no proof that the R genes themselves are the primary recognition targets for corresponding pathogen Avr genes; therefore, potential sig- naling proteins such as EDSl and NDRl may operate upstream or in concert with R gene products. Additional genetic experi- ments are certainly required to determine the influence of pairwise mutations of genes such as EDS1, NDRl, NlMl, and NPRl on a wide spectrum of R genes in Arabidopsis.
Mutagenesis in several plant-pathogen systems has been a powerful tool in identifying genes that are required for the function of race-specific R genes (Holub and Beynon, 1996; Kunkel, 1996; Schulze-Lefert et al., 1996). In barley, screen-
ing a mutagenized line containing the powdery mildew Rgene Mlarp for susceptible mutants led, in addition to numerous defective Mlalp alleles, to the identification of two loci (Rarl and Rar2) that are necessary components of Mla12-specified resistance (Freialdenhoven et al., 1994). In tomato, screening for mutations susceptible to Pto-mediated resistance to bac- terial speck disease led to the discovery of a second tightly linked gene, Prf, that is required both for f to gene function and for the function of a second linked gene, Fen, that con- trols sensitivity to the herbicide fenthion (Salmeron et al., 1994, 1996). In tomato, an elegant screen for increased fungal bio- mass was used in mutagenized plants to identify two loci, Rcr l and Rcr2, that are necessary for Cf-9-specified resistance to the leaf-spotting fungus Cladosporium fulvum (Hammond- Kosack et al., 1994).
It is likely that different and more refined screens for mu- tants will reveal other genes involved in disease resistance signaling mechanisms, assuming there is not a great degree of redundancy. The activation of signal-response coupled reac- tions triggered by specific pathogen recognition also implicates the participation of negative regulatory circuits to prevent un- controlled stimulation of the plant’s defenses (Schulze-Lefert et al., 1996). The appropriate mutational screens have identi- fied putative negative regulatory genes in Arabidopsis (Bowling et al., 1994; Dietrich et al., 1994; Greenberg et al., 1994) and in barley (Freialdenhoven et al., 1996).
To date, it is unclear to what extent there is cross-signaling between pathways leading to the arrest of development of an avirulent pathogen and mechanisms that limit growth of a viru- lent pathogen. SA-depleted tobacco and Arabidopsis plants (mentioned above) showed increased growth of both avirulent and virulent pathogens (Delaney et al., 1994), implicating SA as a common signal in these two responses as well as in SAR. Also, nprl plants permitted greater ingress of a virulent I? syrin- gae strain than did wild-type COLO (Cao et al., 1994), although it is unclear from that study whether any race-specific bac- terial resistance was affected in the mutant plants. Severa1 RPP genes, however, are influenced by nprl (Holub and Beynon, 1996). Two Arabidopsis pad mutants, deficient in camalexin accumulation, allowed increased growth of virulent F! syrin- gae strains but were resistant to avirulent strains (Glazebrook and Ausubel, 1994), suggesting that certain biochemical processes are unique to each response. In a recent study by Glazebrook et al. (1996), mutagenized Arabidopsis plants were screened for enhanced growth of a pathogenic I? syringae strain, and 10 different loci involved in limiting the spread of the disease lesion were found. Some of these loci were desig- nated eds2 to eds8. Interestingly, defective alleles of Nprl (Cao et al., 1994) and Pad2 (Glazebrook and Ausubel, 1994) were among the isolated mutants, supporting the idea that certain signaling components play a role in multiple defense pathways.
Our study shows that Ws-edsl allowed more rapid develop- ment of two Ws-O-compatible I? parasitica isolates than did wild-type Ws-O plants (Table 5). However, this trait appears to be dependent on the age of the plants; very young seedlings

Disease Resistance Signaling Mutation 2043
(7 days old at the time of inoculation) did not exhibit enhanced susceptibility. The apparent increased susceptibility during older growth stages of Ws-edsl may be a consequence of greater surface area when inoculated and the larger biomass of host tissue for colonization. Ws-edsl also exhibited more rapid symptom expression of virulent /? s. tomato DC3000 (Fig- ure lH), although this was correlated with only a marginal increase in the rate of bacterial growth (Figure 2). Also, the disease lesion did not extend farther than in wild-type leaves. It is possible that the accelerated symptom development ob- served in Ws-edsl plants compared with wild-type Ws-O under these experimental conditions is disproportionately greater than the limited enhancement of bacterial growth.
Altogether, our results suggest that the wild-type EDSl gene has some influence in restricting development of certain viru- lent pathogens as well as playing a crucial role in RPP gene-mediated resistance, drawing another possible link be- tween pathways limiting pathogen development in both genetically incompatible and compatible interactions. Near- isogenic lines will be invaluable in making further compari- sons and in ruling out possible effects of genetic background in the host and/or parasite. Experiments are in progress to test whether Ws-edsl is allelic with any of the other eds mutations identified by Glazebrook et al. (1996).
EDSl is a suitable target for molecular isolation either by positional cloning or by transposon tagging. Preliminary map- ping of EDSl shows that it is linked to the cluster of RPP specificities residing on chromosome 3 between GL7 and m249 (13% recombination with pAT3-89.1 [Table 21, a marker that was positioned 1.4 cM from RPP74 in a previous study [Reignault et ai., 19961). However, linkage is not tight in con- trast to the relative positions of the tomato Prf and Pto genes (Salmeron et al., 1996). Cloning the EDS7 gene as well as per- forming further genetic analysis of Ws-edsl in combination with different R genes and resistance signaling mutants in Arabidopsis should allow us to establish its biochemical role in plant-pathogen interactions.
METHODS
Maintenance of Bacterial Strains and Funga1 lsolates
Pseudomonas syringae pv tomafo DC3000 containing the avirulence gene avrB in the broad host range vector pVSP6l (Innes et al., 1993) and containing pVSP61 without an insert was kindly provided by 6. Staskawicz (University of California at Berkeley). These were cultured as described previously (Innes et al., 1993). lsolates of Peronospora spp, Albugo spp, and Bremia lactucae were cultured on their corre- sponding compatible host ecotypes, as described previously (Dangl et al., 1992a; Holub et al., 1994, 1995). The other host species were grown in the same manner as Arabidopsis thaliana, and all parasite inocula were prepared and applied as was done for similar isolates from Arabidopsis. B. lacfucae was maintained on lettuce cultivar Cob- ham Green; Brassica-derived /? parasifica was maintained on 6.
oleracea cv Maris Kestral; P fabacina was maintained on Nicotiana rustica; and the other isolates were maintained on wild-type, uncharac- terized ecotypes.
The P parasifica isolates from Arabidopsis were described previ- ously (Parker et al., 1993; Holub et ai., 1994; Holub and Beynon, 1996). All were derived from single oospores (sexual stage), except for "02, which was maintained as a mass conidiosporangia culture derived originally from a single host seedling. Two isolates of I? parasitica from B. oleracea (P-O05 and P-006) were described by Moss et ai. (1994). The isolate of A. candida from B. oleracea (Ac9) was obtained from the Crucifer Genetics Cooperative (University of Wisconsin, Madison). lsolates of B. lactucae were provided from the collection of I.R. Crute (Horticultural Research lnternational) and included 65/92. 87al84, B12l88, 628194, 630194, B32l94, IL4, Si, SFI, SF5, VOll, T, and 87al84. The remaining isolates were uncharacterized mas spore cultures col- lected by E.B. Holub from single leaves of host plants found in the United Kingdom.
Plant Material, Cultivation, and Pathogenicity Tests
Seed of the Arabidopsis ecotype Wassilewskija (Ws-O) were originally obtained from K. Feldman (University of Arizona, Tucson). M2 seed stocks generated from ethyl methanesulfonate-mutagenized Ws-0 seed were a kind gift of F. Chumley (Du Pont, Wilmington, DE). Seed of ecotype Columbia ((201-5, containing the recessive mutation gll ) was obtained from J. Dangl (University of North Carolina, Chapel Hill). Seed of the ecotype Blanes (Bla-2) were obtained from the Nottingham Arabidopsis Stock Centre (Nottingham, UK). The collection of 96 United Kingdom ecotypes used for testing the pathogenicity of Brassica- derived I? parasifica isolates was assembled by E.B. Holub from wild populations throughout Britain.
Ethyl methanesulfonate-mutagenized M2 seedlings were sown at a density of -400 seed per 320 cm'. These seedlings and col-5 x Ws-edsl F3 families (25 to 30 seedlings) were inoculated by spray- ing a suspension of 4 x 104 mL of P parasifica conidia to run off. lnoculated seedlings were incubated at 16OC. Susceptible plants were rescued as previously described (Reignault et al., 1996). 2,6- Dichloroisonicotinic acid (INA), obtained as a 25% active ingredient wettable powder formulation, was a gift from E. Ward (Ciba-Geigy Corp., Research Triangle Park, NC). INA or the control carrier powder formu- lation was sprayed onto seedlings at a concentration of 25 mglL to imminent runoff. Seedlings were then spray inoculated with P parasitica spores 2 days after INA treatment, as described above. Conditions for the growth of plants and bacteria for bacterial inoculations, infiltra- tion of leaves with bacterial suspensions, and in-the-plant bacterial growth assays were as described by May et al. (1996).
,
Light Microscopy
Development of /? parasifica Noco2 was observed in whole infected cotyledons and leaves. Staining was with lactophenol trypan blue fol- lowed by destaining with chloral hydrate, as described previously (Koch and Slusarenko, 1990). The lactophenol trypan blue stain isselectively retained by funga1 tissue as well as by dead plant cells (including xy- lem vessels of the vascular tissue) or cells that have sustained membrane damage (Keogh et ai., 1980). Material was mounted in chloral hydrate and examined using phase-contrast optics on a Zeiss Axioskop microscope (Carl Zeiss Ltd., Welwyn Garden City, UK).

2044 The Plant Cell
lsolation of Plant Genomic DNA, Molecular Markers, and Mapping Analysis
Plant genomic DNA was isolated from flowering plants using the rapid flower head preparation method of Martienssen et al. (1989), as de- scribed by Reignault et al. (1996). DNA prepared in this way was suitable for restriction enzyme digestion and polymerase chain reaction anal- ysis. Polymerase chain reaction analysis of genomic DNA sequences was performed according to Reignault et al. (1996). Oligonucleotide primers were as described by Konieczny and Ausubel(l993) to generate codominant cleaved amplified polymorphic DNA sequence (CAPS) markers. Primers to amplify microsatellite sequences were designed according to Bell and Ecker (1994). Primer sequences to amplify a portion of m249 were kindly provided by I? Bittner-Eddy and J. Beynon (Wye College, Ashford, UK). The amplification products in Ws-O (1.9 kb) and col-5 (2.5 kb) exhibited a simple sequence length polymor- phism. The restriction fragment length polymorphism marker pAT349.1 (Shirley et al., 1992) was obtained from B. Shirley (Virginia Polytech- nic lnstitute and State University, Blacksburg, VA). Restriction digests, DNA gel blotting, phosphorus-32 labeling, and hybridizations were performed according to standard protocols (Ausubel et al., 1987). In the mapping analysis, the percentage of recombinations between mark- ers in selected Col-5 x Ws-eds7 F2 plants or corresponding F3 families was examined to identify linkage groups.
ACKNOWLEDGMENTS
We thank all those who provided seed stocks and molecular probes and acknowledge with thanks Peter Bittner-Eddy and Jim Beynon (Wye College, University of London) for providing the m249 primer sequence. The B. lactucae and t? parasitica isolate collection was kindly assem- bled by Pam Gordon, Debbie Leckie, and Noel Cogan. Many thanks also to Jonathan Jones and Paul Schulze-Lefert at the Sainsbury Lab- oratory for comments on the manuscript and to Andrew Davis for photography. The Sainsbury Laboratory is supported by the Gatsby Charitable Foundation. A.F. is the recipient of a postdoctoral fellow- ship from the Swedish Research Council for Forestry and Agriculture (SJFR). The research at Horticultural Research lnternational was sup- ported by funding from the United Kingdom Biotechnology and Biological Sciences Research Council.
Received August 1, 1996; accepted September 17, 1996.
REFERENCES
Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A., and Struhl, K., eds (1987). Current Protocols in Molecular Biology. (New York: Green Publishing Associates & Wiley-lnterscience).
Ausubel, F.M., Katagiri, F., Mindrinos, M., and Glazebrook, J. (1995). Use of Arabidopsis thaliana defense-related mutants to dissect the plant response to pathogens. Proc. Natl. Acad. Sci. USA 92,
Bell, C.J., and Ecker, J.R. (1994). Assignment of 30 microsatellite loci 4189-4196.
to the linkage map of Arabidopsis. Genomics 19, 137-144.
Bent, A.F., Kunkel, B.N., Dahlbeck, D., Brown, K.L., Schmidt, R., Giraudat, J., Leung, J., and Staskawicz, B.J. (1994). RPSP of Arabidopsis thaliana: A leucine-rich repeat class of plant disease resistance genes. Science 265, 1856-1860.
Bisgrove, S.R., Simonich, M.T., Smith, N.M., Sattler, A., and Innes, R.W. (1994). A disease resistance gene in Arabidopsis with speci- ficity for two different pathogen avirulence genes. Plant Cell 6,
Bowling, S.A., Guo, A., Cao, H., Gordon, S., Klessig, D.F., and Dong, X. (1994). A mutation in Arabidopsis that leads to constitu- tive expression of systemic acquired resistance. Plant Cell 6,
Cao, H., Bowling, S.A., Gordon, A.S., and Dong, X. (1994). Char- acterization of an Arabidopsis mutant that is nonresponsive to inducers of systemic acquired resistance. Plant Cell 6, 1583-1592.
Century, K., Holub, E.B., and Staskawicz, B.J. (1995). NDR7, a lo- cus of Arabidopsis thaliana that is required for disease resistance to both a bacterial and a fungal pathogen. Proc. Natl. Acad. Sci.
Channon, A.G. (1981). Downy mildew of Brassicas. In The Downy Mil- dews, D.M. Spencer, ed (London: Academic Press), pp. 321-339.
Crute, I.R. (1985). The genetic basis of relationships between para- sites and their hosts. In Mechanisms of Resistance to Plant Disease, R.S.S. Fraser, ed (Dordrecht, The Netherlands: Kluwer Academic Publishers), pp. 80-143.
Crute, I.R., Holub, E.B., Tor, M., Brose, E., and Beynon, J.L. (1993). The identification and mapping of loci in Arabidopsis thaliana for recognition of the fungal pathogens I? parasitica (downy mildew) and Albugo candida (white blister). In Advances in Molecular Genetics of Plant-Microbe Interactions, Vol. 2, E.W. Nester and D.P.S. Verma, eds (Dordrecht, The Netherlands: Kluwer Academic Publishers),
Crute, I.R., Beynon, J., Dangl, J., Holub, E., Mauch-Mani, B., Slusarenko, A., Staskawicz, B., and Ausubel, F. (1994a). Microbial pathogenesis of Arabidopsis. In Arabidopsis (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory), pp. 705-747.
Crute, I.R., Holub, E.B., and Beynon, J.L. (1994b). Phenotypic vari- ation and non-allelic interaction in the gene-for-gene relationship between Arabidopsis thaliana and I? parasitica (downy mildew). In Advances in Molecular Genetics of Plant-Microbe Interactions, Vol. 3, M.J. Daniels, J.A. Downie, and A.E. Osbourn, eds (Dordrecht, The Netherlands: Kluwer Academic Publishers), pp. 267-272.
Dangl, J.L. (1993). Applications of Arabadopsis thaliana to outstand- ing issues in plant-pathogen interactions. Int. Rev. Cytol. 114,5343.
Dangl, J.L., Holub, E.B., Debener, T., Lehnackers, H., Ritter, C., and Crute, I.R. (1992a). Genetic definition of loci involved in Arabidop sis-pathogen interactions. In Methods in Arabidopsis Research, C. Koncz, N.-H. Chua, and J. Schell, eds (Singapore: World Scientific Publishing Co.), pp. 393-418.
Dangl, J.L., Ritter, C., Gibbon, M.J., Mur, L.A.J., Wood, J.R., Goss, S., Mansfield, J., Taylor, J.D., and Vivian, A. (1992b). Functional homologs of the RPM7 disease resistance gene in bean and pea. Plant Cell 4, 1359-1369.
Delaney, T.P., Uknes, S., Vernooij, B., Friedrich, L., Weymann, K., Negrotto, D., Gaffney, T., Gut-Rella, M., Kessman, H., Ward, E., and Ryals, J. (1994). A central role of salicylic acid in plant disease resistance. Science 266, 1247-1250.
Delaney, T.P., Friedrich, L., and Ryals, J.A. (1995). Arabidopsis sig- na1 transduction mutant defective in chemically and biologically
927-933.
1845-1857.
USA 92, 6597-6601.
pp. 437-444.

Disease Resistance Signaling Mutation 2045
induced disease resistance. Proc. Natl. Acad. Sci. USA 92,
Dietrich, R.A., Delaney, TR, Uknes, S. J., Ward, E.R., Ryals, J.A., and Dangl, J.L. (1994). Arabidopsis mutants simulating disease re- sistance response. Cell 77, 565-577.
Dixon, M.S., Jones, D.A., Keddie, J.S., Thomas, C.M., Harrison, K., and Jones, J.D.G. (1996). The tomato 0-2 disease resistance locus comprises two functional genes encoding leucine-rich repeat proteins. Cell 84, 451-459.
Fillingham, A.J., Wood, J.R., Crute, I.R., Mansfield, J.W., Taylor, J.D., and Vivian, A. (1992). Avirulence genes from Pseudomonas syringae pathovars phaseolicola and pisi confer specificity towards both host and non-host species. Physiol. MOI. Plant Pathol. 40, 1-15.
Flor, H.H. (1971). Current status of the gene-for-gene concept. Annu. Rev. Phytopathol. 9, 275-296.
Freialdenhoven, A., Scherag, B., Hollricher, K., Collinge, D.B., Thordal-Christensen, H., and Schulze-Lefert, P. (1994). Nar-1 and Nar-2, two loci required for Mfa,,specified race-specific resistance to powdery mildew in barley. Plant Cell 6, 983-994.
Freialdenhoven, A., Peterhansel, C., Kurth, J., Kreuzaler, F., and Schulze-Lefert, P. (1996). ldentification of genes required for the function of non-race-specific mlo resistance to powdery mildew in barley. Plant Cell 8, 5-14.
Gaffney, T., Friedrich, L., Vernooij, B., Negrotto, D., Nye, G., Uknes, S., Ward, E., Kessman, H., and Ryals, J. (1993). Requirement of salicylic acid for the induction of systemic acquired resistance. Science 261, 754-756.
Glazebrook, J., and Ausubel, F.M. (1994). lsolation of phytoalexin- deficient mutants of Arabidopsis thaliana and characterization of their interactions with bacterial pathogens. Proc. Natl. Acad. Sci. USA 91, 8955-8959.
Glazebrook, J., Rogers, E.E., and Ausubel, F.M. (1996). lsolation of Arabidopsis mutants with enhanced disease susceptibility by di- rect screening. Genetics 143, 973-982.
Grant, M.R., Godiard, L., Straube, E., Ashfield, T., Lewald, J., Sattler, A., Innes, R.W., and Dangl, J.L. (1995). Structure of the Arabidopsis RPMl gene enabling dual specificity disease resistance. Science 269, 843-846.
Greenberg, J.T., Guo, A., Klessig, D.F., and Ausubel, F.M. (1994). Programmed cell death in plants: A pathogen-triggered response activated coordinately with multiple defense functions. Cell 77,
Hammond-Kosack, K.E., Jones, D.A., and Jones, J.D.G. (1994). Iden- tification of two genes required in tomato for full Cf-9-dependent resistance to Cladosporium fulvum. Plant Cell 6, 361-374.
Hinsch, M., and Staskawicz, B.J. (1996). ldentification of a new Arabidopsis disease resistance locus, RPS4, and cloning of the cor- responding avirulence gene, avrRps4, from Pseudomonas syringae pv. pisi. MOI. Plant-Microbe Interact. 9, 55-61.
Holub, E.B., and Beynon, J.L. (1996). Symbiology of mouse-ear cress (Arabidopsis thaliana) and oomycetes. Adv. Bot. Res., in press.
Holub, E.B., Beynon, J.L., and Crute, I.R. (1994). Phenotypic and genotypic characterization of interactions between isolates of i? parasitica and ecotypes of Arabidopsis thaliana. MOI. Plant-Microbe Interact. 7, 223-239.
Holub, E.B., Brose, E., Tor, M., Clay, C., Crute, I.R., and Beynon, J.L. (1995). Phenotypic and genotypic variation in the interaction between Arabidopsis thaliana and Albugo candida. MOI. Plant- Microbe Interact. 8, 916-928.
6602-6606.
551-563.
Innes, R.W., Bisgrove, S.R., Smlth, N.M., Bent, A.F., Staskawlw, B.J., and Liu, Y.-C. (1993). ldentification of a disease resistance lo- cus in Arabidopsis that is functionally homologous to the RPGl locus of soybean. Plant J. 4, 813-820.
Jones, D.A., Thomas, C.M., Hammond-Kosack, K.E., Balint-Kurti, P.J., and Jones, J.D.G. (1994). lsolation of the tomato Cf-9 gene for resistance to Cladospotfum fulvum by transposon tagging. Science
Keen, N.T. (1990). Gene-for-gene complementarity in plant-pathogen interactions. Annu. Rev. Genet. 24, 447-463.
Keogh, R.C., Dwerall, B.J., and Mcleod, S. (1980). Comparison of histological and physiological responses to Phakopsora pachyrhizi in resistant and susceptible soybean. Trans. Br. Mycol. SOC. 74,
Kobayashi, D.Y., Tamaki, S.J., and Keen, N.T. (1989). Cloned aviru- lence genes from the tomato pathogen Pseudomonas syringae pv. tomato confer cultivar specificity on soybean. Proc. Natl. Acad. Sci. USA 86, 157-161.
Kobe, B., and Deisenhoter, J. (1994). The leucine-rich repeat: A ver- satile binding rnotif. Trends Biochem. Sci. 19, 415-421.
Koch, E., and Slusarenko, A. (1990). Arabidopsis is susceptible to infection by a downy mildew fungus. Plant Cell 2, 437-445.
Konieczny, A., and Ausubel, F.M. (1993). A procedure for mapping Arabidopsis mutations using co-dominant ecotype-specific PCR- based markers. Plant J. 4, 403-410.
Kunkel, B.N. (1996). A useful weed put to work: Genetic analysis of disease resistance in Arabidopsis thaliana. Trends Genet. 12,63-69.
Kunkel, B.N., Bent, A.F., Dahlbeck, D., Innes, R.W., and Staskawicz, B.J. (1993). RPS2, an Arabidopsis disease resistance locus specifying recognition of Pseudomonas syringae strains expressing the aviru- lence gene avrRpt2. Plant Cell 5, 865-875.
Lamb, C.J. (1994). Plant disease resistance genes in signal percep- tion and transduction. Cell 76, 419-422.
Lawrence, G.J., Finnegan, E.J., Ayliffe, M.A., and Ellis, J.G. (1995). The L6 gene for flax rust resistance is related to the Arabidopsis bacterial resistance gene RPS2 and the tobacco vira) resistance gene N. Plant Cell 7, 1195-1206.
Martienssen, R.A., Barkan, A., Freeling, M., and Taylor, W.C. (1989). Molecular cloning of a maize gene involved in photosynthetic mem- brane organisation that is regulated by Robertson's mutator. EM60
Martin, G.B., Brommonschenkel, S.H., Chunwongse, J., Frary, A., Ganal, M.W., spivey, R., Wu, T., Earle, E.D., and Tanksley, S.D. (1993). Map-based cloning of a protein kinase gene conferring dis- ease resistance in tomato. Science 262, 1432-1436.
May, M.J., Parker, J.E., Daniels, M.J., Leaver, C.J., and Cobbett, C.S. (1996). An Arabidopsis mutant depleted in glutathione shows unaltered responses to funga1 and bacterial pathogens. MOI. Plant- Microbe Interact. 9, 349-356.
Michelmore, R.W., llott, T., Hulbert, S.H., and Farrara, B. (1988). The downy mildews. Adv. Plant Pathol. 6, 53-79.
Mindrinos, M., Katagiri, F., Yu, G.-L., and Ausubel, F.M. (1994). The A. thaliana disease resistance gene RPS2 encodes a protein con- taining a nucleotide-binding site and leucine-rich repeats. Cell 78,
Moss, N.A., Crute, I.R., and Lucao, J.A. (1994). Laboratory produc- tion of oospores of i? parasitica (crucifer downy mildew) and the
266, 789-793.
329-333.
J. 8, 1633-1639.
1089-1099.

2046 The Plant Cell
recovery and characterization of sexual progeny from crosses be- tween isolates with different host specificity. Plant Pathol. 43,713-725.
Parker, J.E., Szab6, V., Staskawia, B. J., Lister, C., Dean, C., Daniels, M.J., and Jones, J.D.G. (1993). Phenotypic characterization and molecular mapping of the Arabidopsis thaliana locus, RPP-5, deter- mining resistance to i? parasitica. Plant J. 4, 821-831.
Pryor, T., and Ellis, J. (1993). The genetic complexity of funga1 resis- tance genes in plants. Adv. Plant Pathol. 10, 281-305.
Reignault, P., Frost, L.N., Richardson, H., Daniels, M.J., Jones, J.D.G., and Parker, J.E. (1996). Four Arabidopsis RPf loci control- ling resistance to the Noc02 isolate of i? parasitica map to regions known to contain other RPf recognition specificities. MOI. Plant- Microbe Interact. 9, 464-473.
Ryals, J., Uknes, S., and Ward, E. (1994). Systemic acquired resis- tance. Plant Physiol. 104, 1109-1112.
Salmeron, J.M., Barker, S.J., Carland, F.M., Mehta, A.Y., and Staskawiu, B.J. (1994). Tomato mutants altered in bacterial dis- ease resistance provide evidence for a new locus controlling pathogen recognition. Plant Cell 6, 511-520.
Salmeron, J.M., Oldroyd, G.E.D., Rommens, C.M.T., Scofield, S.R., Kim, H.-S., Lavelle, D.T., Dahlbeck, D., and Staskawicz, B.J. (1996). Tomato Prf is a member of the leucine-rich repeat class of plant dis- ease resistance genes and lies embedded within the Pto kinase gene cluster. Cell 86, 123-133.
Schulze-Lefert, P., Peterhaensel, C., and Freialdenhoven, A. (1996). Mutation analysis for the dissection of resistance. In The Gene-for- Gene Relationship in Host-Parasite Interactions, I. Crute, J. Bur- don, and E. Holub, eds Wallingford, UK: CAB International), in press.
Shirley, B.W., Hanley, S., and Goodman, H.M. (1992). Effects of ioniz- ing radiation on a plant genome: Analysis of two Arabidopsis transparent testa mutations. Plant Cell 4, 333-347.
Song, W.X., Wang, O.-L., Chen, L.-L., Kim, H.-S., Pi, L.Y., Holsten, T., Gardner, J., Wang, B., Zhai, W.-X., Fauquet, C., and Ronald, I? (1995). A receptor kinase-like protein encoded by the rice dis- ease resistance gene, Xa27. Science 270, 1804-1806.
Staskawicz, B.J., Ausubel, F.M., Baker, B.J., Ellis, J.G., and Jones, J.D.G. (1995). Molecular genetics of plant disease resistance. Science 268, 661-667.
Tor, M., Holub, E.H., Bmse, E., Musker, R., Gunn, N., Can, C, Crute, IR., and Beynon, J.L. (1994). Map positions of three loci in Ambidop sis iBa/iana associated with isolate-specific recognition of i? pemskica (downy mildew). MOI. Plant-Microbe Interact. 7, 214-222.
Uknes, S., Mauch-Mani, B., Moyer, M., Potter, S., Wllliams, S., Dincher, S., Chandler, D., Slusarenko, A., Ward, E., and Ryals, J. (1992). Acquired resistance in Arabidopsis. Plant Cell4,645-656.
Uknes, S., Winter, A.M., Delaney, T., Vernooij, B., Morse, A., Friedrich, L., Nye, G., Potter, S., Ward, E., and Ryals, J. (1993). Biological induction of systemic acquired resistance in Arabidop- sis. MOI. Plant-Microbe Interact. 6, 692-698.
Vernooij, B., Friedrich, L., Ahl Goy, P., Staub, T., Kessmann, H., and Ryals, J. (1995). 2,6-Dichloroisonicotinic acid-induced resis- tance to pathogens without the accumulation of salicylic acid. MOI. Plant-Microbe Interact. 8, 228-234.
Ward, E.R., Uknes, S.J., Wllliams, S.C., Dincher, S.S., Wiederhold, D.L., Alexander, D.C., Ahl-Goy, P., Mbtraux, J.-P., and Ryals, J.A. (1991). Coordinate gene activity in response to agents that induce systemic acquired resistance. Piant Cell 3, 1085-1094.
Whalen, M.C., Innes, R.W., Bent, A.F., andstaskawicz, B.J. (1991). ldentification of Pseudomonas syringae pathogens of Arabidopsis and a bacterial locus determining avirulence on both Arabidopsis and soybean. Plant Cell 3, 49-59.
Whitham, S., Dinesh-Kumar, S.P., Choi, D., Hehl, R., Corr, C., and Baker, 8. (1994). The product of the tobacco mosaic virus resistance gene N: Similarity to To11 and the interleukin-1 receptor. Cell 78,
Wood, J.A., Vivian, A., Jenner, C., Mansfield, J.W., and Taylor, J.D. (1994). Oetection of a gene in pea controlling nonhost resistance to Pseudomonas syringae pv. phaseolicola. MOI. Plant-Microbe In- teract. 7, 534-537.
Yu, G., Katagiri, F., and Ausubel, F.M. (1993). Arabidopsis mutations at the RfS2 locus result in loss of resistance to Pseudomonas syrin- gae strains expressing the avirulence gene avrRpt2. MOI. Plant-Microbe Interact. 6, 434-443.
Zhou, J., Loh, Y.-T., Bressan, R.A., and Martin, G.B. (1995). The tomato gene Ptil encodes a serinehhreonine kinase that is phos- phorylated by Pto and is involved in the hypersensitive response. Cell 83, 925-935.
101 1-1 115.

DOI 10.1105/tpc.8.11.2033 1996;8;2033-2046Plant Cell
J E Parker, E B Holub, L N Frost, A Falk, N D Gunn and M J Danielsparasitica specified by several different RPP genes.
Characterization of eds1, a mutation in Arabidopsis suppressing resistance to Peronospora
This information is current as of March 14, 2019
Permissions https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298X
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists





























