Embed Size (px)
Citation preview

A r t i c l e
CHARACTERIZATION OF FOURESTERASE GENES AND ESTERASEACTIVITY FROM THE GUT OF THETERMITE Reticulitermes flavipes
Marsha M. Wheeler, Matthew R. Tarver, Monique R. Coy,and Michael E. ScharfEntomology and Nematology Department, University of Florida,Gainesville, Florida
Four esterase genes and general esterase activity were investigated in thegut of the termite Reticulitermes flavipes. Two genes (RfEst1 andRfEst2) share significant translated identity with a number of insect JHesterases. The two remaining genes (RfEst3 and RfEst4) apparently codefor much shorter proteins with similarity to fungal phenolic acid esterasesinvolved in hemicellulose solubilization. All four genes showedconsistently high midgut expression. This result was further supported bycolorimetric activity assays and Native polyacrylamide gel electrophoresis,which showed significant esterase activity and a number of isoforms in themidgut. The greatest esterase activity and isoform composition weredetected when a-naphthyl propionate was used as a substrate. Moreover,esterase activity and diverse isoforms were present in gut mitochondrial,microsomal, and cytosolic sub-cellular protein fractions, as well as in thehindgut lumen. These findings reveal an agreement between gut esterasegene expression and activity distributions, and support the idea thatR. flavipes gut esterase activity is host (not symbiont)-derived. Inaddition, these findings support the hypotheses that termite gut esterasesmay play important roles in lignocellulose digestion and castedifferentiation. This study provides important baseline data that willassist ongoing functional-genomic efforts to identify novel genes with
Grant sponsor: USDA-CSREES-NRI; Grant number: 2007-35607-17777; Grant sponsors: University ofFlorida–IFAS; Consortium for Plant Biotechnology Research, Inc., and DOE; Grant number: DE-FG36-02GO12026.Additional Supporting Information may be found in the online version of this article.Marsha M. Wheeler’s present address is Department of Entomology, University of Illinois at Urbana-Champaign, Urbana, IL.Correspondence to: Dr. Michael E. Scharf, Entomology and Nematology Department, PO Box 110620,University of Florida, Gainesville, FL 32611-0620. E-mail: [email protected]
ARCHIVES OF INSECT BIOCHEMISTRY AND PHYSIOLOGY, Vol. 73, No. 1, 30–48 (2010)
Published online in Wiley InterScience (www.interscience.wiley.com).
& 2009 Wiley Periodicals, Inc. DOI: 10.1002/arch.20333

roles in semiochemical, hormone, and lignocellulose processing in thetermite gut. �C 2009 Wiley Periodicals, Inc.
Keywords: hydrolase; general esterase; hemicellulose; hemicellulase;feruloyl esterase; coumaryl esterase; phenolic acid esterase; bioethanol
INTRODUCTION
Esterases are hydrolytic enzymes that cleave ester bonds in a diversity of biomolecules(Oakeshott et al., 2005). Many insect esterases have well-defined biological functions,such as those involved in xenobiotic, lipid, acetylcholine, and JH metabolism. Incontrast, other insect esterases have largely undefined functions. Recently, a number ofmicrobial and fungal esterases have been identified that play roles in hemicellulosedepolymerization by functioning as feruloyl or phenolic acid esterases (Crepin et al.,2004; Benoit et al., 2008). Therefore, esterases with undefined activity in the guts ofphytophagous and xylophagous insects may act as phenolic acid esterases (reviewed byScharf and Tartar, 2008). In addition, because of the highly esterified structure oflignin, it is conceivable that general esterases may also contribute to lignindepolymerization in wood-feeding insects such as termites (Scharf and Tartar, 2008).
Termites are social insects that subsist on sugars and other nutrients obtained fromotherwise nutritionally poor lignocellulose diets (Ohkuma, 2003; Scharf and Tartar,2008). Lignocellulose is a natural complex of the three biopolymers: cellulose,hemicellulose, and lignin. Cellulose is composed of long b-1,4-linked polymers ofglucose that are held together in bundles by hemicellulose (Ljungdahl and Erickson,1985; Lange, 2007). Hemicellulose is composed of shorter b-1,4-linked polymers ofmixed 5- and 6-carbon sugars and uronic acid sugars (Saha, 2003). Lignin is a3-dimensional polymer of phenolic compounds that are linked to each other and tohemicellulose by ester bonds. Another important characteristic of hemicellulose is itsesterification with monomers and dimers of phenolic acid esters, which are identical to themono-lignols that compose lignin (Saha, 2003; Crepin et al., 2004; Benoit et al., 2008).
Termites digest lignocellulose with the assistance of endogenous and symbiont-produced digestive enzymes (Breznak and Brune, 1995; Watanabe et al., 1998;Ohkuma, 2003; Scharf and Tartar, 2008; Tartar et al., 2009). Termite gut symbiontsinclude a diversity of microorganisms such as protozoa, bacteria, spirochetes, fungi,and yeast, among others (Breznak and Brune, 1995; Ohkuma, 2003; Warnecke et al.,2007). The order Isoptera is divided into the higher and lower termites based mostlyon symbiont composition. Lower termites, including Reticulitermes flavipes, hostcellulolytic protozoa in addition to a variety of prokaryotes. Higher termites lackprotozoa altogether, and instead possess cellulolytic prokaryotes (Warnecke et al.,2007). The roles of symbiotic fungi in higher and lower termites are not well defined;however, some higher termites cultivate fungus gardens in their nests and these arethought to assist in lignocellulose digestion by producing cellulases, hemicellulases,and possibly lignases (Okhuma, 2003; Taprab et al., 2005).
A different, but nonetheless important phenomenon linking the termite gut totermite sociality is trophallaxis. Trophallaxis is defined as the transfer of symbionts,food, semiochemicals, or other fluids among members of a social group throughmouth-to-mouth (stomodeal) or anus-to-mouth (proctodeal) feeding (Wheeler, 1918;Wilson, 1971). Trophallaxis enables the flow of socially relevant chemicals such as
Termite Gut Esterases � 31
Archives of Insect Biochemistry and Physiology

hormones, pheromones, and other signaling molecules through individual termiteguts to other colony members. For example, JH is known to play a role in both termitecaste differentiation and reproductive competence (Park and Raina, 2004, 2005; Maoet al., 2005; Brent et al., 2005; Elliott and Stay, 2007, 2008; Cornette et al., 2008; Korbet al., 2009). Previous studies have shown that JH can act exogenously to inducesoldier caste differentiation in workers via feeding on treated substrates (e.g., Scharfet al., 2005). Other caste-regulatory terpenes that are active via feeding are producedby termite soldiers (Tarver et al., 2009), some of which are derived from the samebiosynthetic pathways as JH (Belles et al., 2005). Finally, sequencing from theendogenous (host) gut transcriptome in R. flavipes has revealed a number of genesencoding members of JH biosynthetic and degradative pathways, including twoputative JH esterases (Tartar et al., 2009; M. R. Tarver, unpublished data). Thus, thetermite gut is of interest not only because of the ability of termites to digestlignocellulose and house symbionts, but also because of the potential role oftrophallaxis in chemical signaling.
This research was conducted to investigate esterase gene and protein expressionin the R. flavipes gut, and to begin addressing questions relating to the physiologicalroles that gut esterases play in termite biology. Our objectives were to (1) determineexpression profiles of several esterase ESTs that were previously sequenced from aR. flavipes gut cDNA library, and (2) functionally characterize esterase activity andisoform composition throughout the gut. From these efforts, we provide evidence thatesterase genes are host-derived, have homology to JH and feruloyl esterases, and arespecifically expressed within the digestive tract, particularly in the symbiont-freemidgut.
MATERIALS AND METHODS
Termites
R. flavipes colonies were collected in Gainesville, FL, and maintained in sealed plasticboxes (30� 24� 10 cm) in complete darkness (L:D 5 0:24), at 221C and 69% RH.Colonies were maintained without soil for more than six months and provisioned withmoist brown paper towels and pine wood shims. The identity of colonies as R. flavipeswas verified by a combination of soldier morphology and 16S-mt-rDNA genesequence. Only worker termites were used because of their lignocellulose digestioncapability. Termites were considered workers if they did not possess any sign of wingbuds or distended abdomens, and had pronotal widths wider than mesonotal widths(Laine and Wright, 2003).
Gene Sequence Identification and Analyses
Four esterase genes were identified from a pool of ESTs obtained previously from asequencing project conducted on a normalized gut cDNA library (Tartar et al., 2009).Initially, three of the four genes were deduced based on multiple ESTs that alignedinto contiguous fragments, while the fourth gene (RfEST-4) was based on a single read.Subsequently, all reported sequences are based on multiple forward and reverse readsof positive clones. The sequences and associated validative details are provided asSupporting Information, which is available online. All sequence alignments were madeunder default settings using MegalignTM in the Lasergene software package (Madison,
32 � Archives of Insect Biochemistry and Physiology, January 2010
Archives of Insect Biochemistry and Physiology
WI). Hydropathy profiles and signal peptides were determined using ProteanTM inthe Lasergene software package. Glycosylation sites in translated peptides werepredicted using the method of Hamby and Hirst (2008) available at http://comp.chem.nottingham.ac.uk/glyco/.
Quantitative PCR
Table 1 shows qPCR primer sequences for the four esterases and the control geneb-actin. qPCR primers were designed with specificity to unique sequence regions of thetarget genes, and to produce products in the 100–300-bp range. qPCR primers weredesigned using Primer3 (Rozen and Skaletsky, 2000). cDNA was synthesized from thetotal RNA of 25 individual gut regions per experimental replicate. Total RNA andcDNA were obtained using the SV total RNA isolation kit (Promega) and the iScriptTM
cDNA Synthesis Kit (Bio-Rad; Hercules, CA), respectively, following manufacturerprotocols. cDNA from the foregut1salivary gland, midgut, and hindgut regions servedas the template for qPCR. Gut dissections were performed as described in thefollowing section, using SV lysis buffer (Promega; Madison, WI) in place of potassiumphosphate. qPCR reactions contained equal template loadings and proceeded for30–35 cycles. PCR products were viewed on 1.5% agarose gels, imaged, and the imagesanalyzed densitometrically using imageJ freeware (http://rsb.info.nih.gov/ij/).
Table 1. Gene Sequence Accession Numbers and qPCR Primer Sequences�
Gene nameAccessionnumbers Primer name
Alternate primername Primer sequence (50–30)
RfEst-1 GQ180944 RfEst1 L Contig515L-set 4 ATGACAGAAGAGGAGCTTGACCFL638851 RfEst1 R Contig515R-set4 CAGGAGTTCAAAGTCACTGTCGFL638692FL637886FL638877
RfEst-2 GQ180945 RfEst2 L Contig493-1R GGTTAGGCGTCATTTGTAGAGGFL638587 RfEst2 R Contig493-1L GGCTCATGGTCATCTAAGAACCFL637361FL637132FL636639
RfEst-3 GQ180946 RfEst3 L Contig275L-set5 AGAGACGGTTGAAGACTTGGTCFL638979 RfEst3 R Contig275R-set5 ACTGATACGTAGTGGCCTGACAFL640151FL636973
RfEst-4 GQ180947 RfEst4 L TG_15_C3 1L GCCAGAATTCAAGCTGCTGTFL638686 RfEst4 R TG_15_C3 1R TGTCCTTGTCTTGCTGTGTCTC
b-actin DQ206832 b-actin L N/A AGAGGGAAATCGTGCGTGACb-actin R N/A CAATAGTGATGACCTGGCCGT
�Accession numbers shown in bold correspond to contiguous sequences; those not shown in bold representindividual ESTs.
Termite Gut Esterases � 33
Archives of Insect Biochemistry and Physiology

Dissections, Protein Preparation, Centrifugation Protocols, and Protein Assays
All manipulations were performed on ice. For tissue localization studies, 25 workertermite guts were removed and dissected into the three regions of foregut1salivarygland, midgut, and hindgut. Each gut region preparation was homogenized using aTenbroeck glass homogenizer in potassium phosphate (0.1 M, pH 7.6), and thencentrifuged for 15 min at 14,000g and 41C. The supernatant was saved for assays andthe pellet discarded.
For isolation of subcellular gut protein fractions, an established protocol was followed(Kupfer and Levin, 1972) with minor modification. Fifty whole worker termite guts werehomogenized in potassium phosphate as above. The homogenate was centrifuged at1,000g and 41C for 5 min and the pellet discarded. The supernatant was transferred to afresh tube and centrifuged at 10,000g for 10 min and 41C. A small aliquot of thesupernatant was saved and the remaining volume transferred to a fresh tube. The resulting10,000g mitochondrial pellet was washed and re-spun 3� in potassium phosphate bufferbefore being re-suspended in potassium phosphate10.1% Triton X-100 (Valles et al., 2001).Next, a volume of calcium chloride was added to the post-mitochondrial supernatant at afinal conc. of 8 mM. The mixture was inverted several times, left on ice for 5 min, and thencentrifuged at 10,000g for 10 min and 41C. The resulting supernatant was transferred to afresh tube and used as the soluble/cytosolic enzyme source. The resulting microsomal pelletwas washed and re-spun 3� in potassium phosphate buffer before being re-suspended inpotassium phosphate 10.1% Triton X-100 (Valles et al., 2001).
Gut luminal contents were isolated from 100 worker termites by holding them bythe abdomen with soft larval forceps and then by gently squeezing to inducedefecation. The termite abdomen was then lowered into potassium phosphate buffer,where the fecal material was collected. Symbiont cells and other debris were pelletedby centrifugation for 10 min at 14,000g and 41C and the remaining supernatant usedas the enzyme source.
Protein content of protein preparations was estimated by a microplate Bradfordassay (Bio-Rad; Hercules, CA) using bovine serum albumin as a standard andcorresponding buffers as blanks.
Esterase Native PAGE
Methods followed an established protocol with slight modification (Scharf et al., 1998).Volumes of supernatant containing 5 mg of total protein were diluted 1:1 with NativePAGE sample buffer (Bio-Rad) and loaded onto native PAGE gels (7.5% resolving gelsand 4% stacking gels). Electrophoresis was conducted in Tris-Glycine running bufferfor 1 hr at 41C. After running, gels were incubated in 100 ml sodium phosphate buffer(0.1 M, pH 7.5) containing 1 ml of 60 mM a-naphthyl ester substrate in acetone (1 mMfinal conc.) for 15 min. Tested substrates included a-naphthyl acetate, propionate,butyrate, valerate, or caprylate (Sigma; St. Louis, MO). To visualize bands, 20 mg ofFast Blue BB (Sigma) in 1 ml water (2.0% final conc.) was added. The Fast Blue BBsolution was filtered through glass wool just before use to remove insoluble particles.Gels were fixed, destained, and stored in 10% acetic acid before photographing.
Colorimetric Esterase Assays
Methods followed an established protocol (Scharf et al., 1998) with adaptation to amicroplate format. The assay mixture consisted of 188 ml of 0.02 M sodium phosphate
34 � Archives of Insect Biochemistry and Physiology, January 2010
Archives of Insect Biochemistry and Physiology

buffer (pH 7.0), 2 ml of 25 mM substrate solution, and 10 ml gut homogenate. Reactionsproceeded at room temperature for 10 min. Reactions were stopped with 50ml of 0.3%Fast Blue BB dissolved in nanopure water containing 3.5% SDS. The assay plates wereincubated at 301C for 15 min before being read at 600 nm with a microplate reader.Formation of the naphthol product was determined from a-naphthol standard curvesstarting at 5 mM, stained with the Fast Blue-SDS stop solution as above. This assay wascarried out for the five naphthyl-based substrates described for native-PAGE in thepreceding section, with three separate homogenates per tissue and replication overtwo termite colonies.
Statistical Analyses
Native PAGE and colorimetric esterase assays were performed on three replicateprotein preparations from two termite colonies. qPCR analyses were performed onfive replicate RNA isolations and cDNA syntheses from two termite colonies. Statisticalanalyses consisted of one-way ANOVA followed by Tukey’s multiple range test forcolorimetric enzyme data and LSD t-tests for qPCR data. With the exception ofhindgut colorimetric data, all variances were equal and ANOVAs were verified as beingsignificant before conducting mean separation testing.
RESULTS
Esterase Gene Identification
A number of putative esterase genes were previously sequenced from a normalized R.flavipes gut cDNA library (Tartar et al., 2009). Here, we focused on four esterase genesthat were deduced from three contigs (515, 493, and 275), as well as a single ESTrepresenting a near full-length cDNA (TG_15_C3) (Table 1). The three contigs wereassembled from 4, 4, and 3 individual EST sequences, respectively. The genes arereferred to, respectively, as Rf (R. flavipes) Est (esterase) genes 1, 2, 3, and 4.Corresponding Genbank accession numbers for each gene are presented in Table 1.Nucleotide sequences, amino acid translations, and protein hydropathy profiles foreach of the four genes are provided in Supplementary Figures S1–S4. The ORFsequences of the four genes are mostly unique in their nucleotide compositions (Supp.Fig. S5).
Esterase Alignments
An alignment of the translated amino acids encoded by the four esterase genes isshown in Figure 1. The deduced proteins encoded by the four esterase genessegregate into two groups that include longer (i.e., RfEst1 and 2) and shorter ORFs(i.e., RfEst3 and 4). The ORFs of RfEst1, 3, and 4 appear to be complete; however,RfEst2 appears to be missing the N-terminal region of its ORF and, thus, will requirefurther sequencing. The alignment reveals several conserved N-terminal esterasemotifs in RfEst1 that are absent in RfEst2, 3, and 4 (see details below), but revealsgreater degrees of conservation in the region homologous to the C-terminal end ofRfEst1. In particular, beginning at consensus residue 422, there are 11 completelyconserved amino acids and a conserved G--H-D- esterase catalytic motif in all fouresterases.
Termite Gut Esterases � 35
Archives of Insect Biochemistry and Physiology

This RfEst1 and 2 group (putative insect JH esterases), is composed of largerproteins with predicted molecular masses of �60 kDa. The predicted RfEst1 proteinhas a molecular mass of 59 kDa, an 11–amino acid signal peptide (MAETVTVTVAQ),30 N-glycosylation sites, and shares significant identity across its entire length with anumber of putative insect JH esterases including representatives from the longicornbeetle Psacothea hilaris (BAE94685), the red flour beetle Tribolium castaneum(XP_967137), the honeybee Apis mellifera (AAU81605), and the saw fly Athalia rosae(BAD91555). The full-length RfEst2 amino acid sequence is not yet available, but theexisting sequence is highly similar to RfEst1. An amino acid alignment of these 6proteins (Fig. 2) reveals a significant proportion of conserved amino acid residues,including a conserved JH esterase catalytic triad (G-SAG, E/D, and G--H-D-), and twosubstrate recognition motifs (RF, DQ).
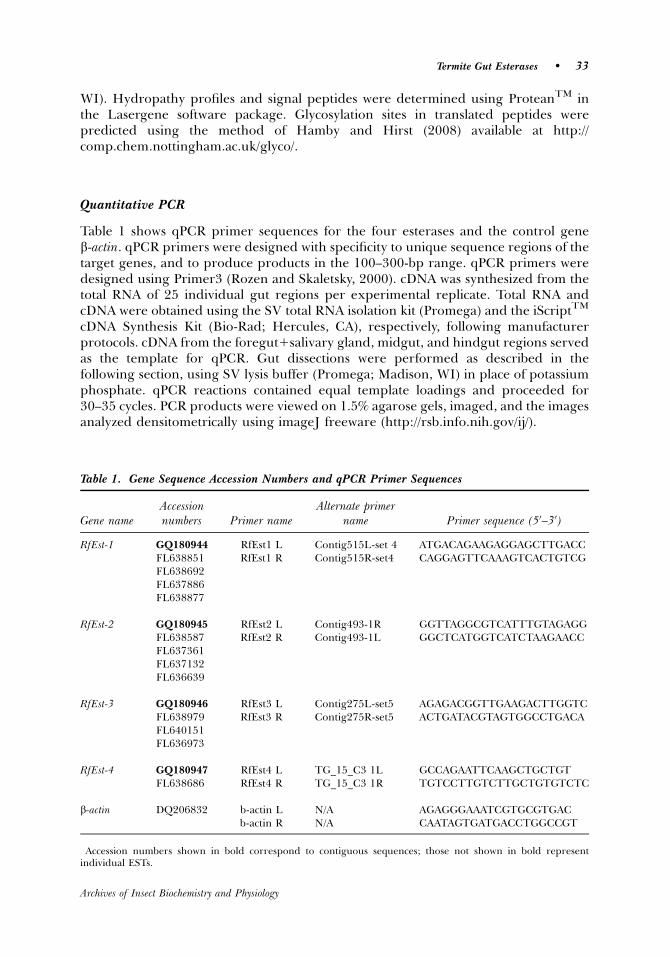
The second esterase group includes RfEst3 and RfEst4. The predicted proteins forthese two genes are smaller (�18–23 kDa) and only share homology with insectcarboxylesterase-like proteins on their C-terminal ends. Other than the carboxyles-terases/JH esterases noted above, there are no other full-length translated databasematches for RfEst3 and 4. Because fungi like Aspergillus niger are lignocellulosedigesters and produce short �30-kDa ferulic acid esterases (fae) that play roles inhemicellulose processing (de Vries et al., 1997), we aligned the translated RfEst3 and 4sequences with the translated faeA sequence of A. niger (Y09330) (Fig. 3). There are 13
Figure 1. Clustal W alignment of translated amino acid sequences for R. flavipes esterases RfEst 1, 2, 3, and 4.Boxes denote conserved esterase catalytic sites and substrate recognition motifs. Triangles (m) indicate aminoacid residues that are 100% conserved across all proteins. Shaded residues are identical to the RfEst1 sequence.See Table 1 for accession numbers.
36 � Archives of Insect Biochemistry and Physiology, January 2010
Archives of Insect Biochemistry and Physiology

completely conserved residues among faeA, RfEst3, and RfEst4 and many otherregions of similarity.
Expression of Gut Esterase Genes
Expression of the four esterase genes in the R. flavipes foregut1salivary gland, midgut,and hindgut was determined by qPCR. Pooled densitometric analysis results from
Figure 2. Clustal W alignment of the translated RfEst1 and RfEst2 esterase amino acid sequences withother translated insect JHE (juvenile hormone esterase)-like esterases. Boxes denote conserved esterasecatalytic sites and substrate recognition motifs. Shaded residues are identical to the RfEst1 sequence. SeeTable 1 and text for accession numbers.
Termite Gut Esterases � 37
Archives of Insect Biochemistry and Physiology
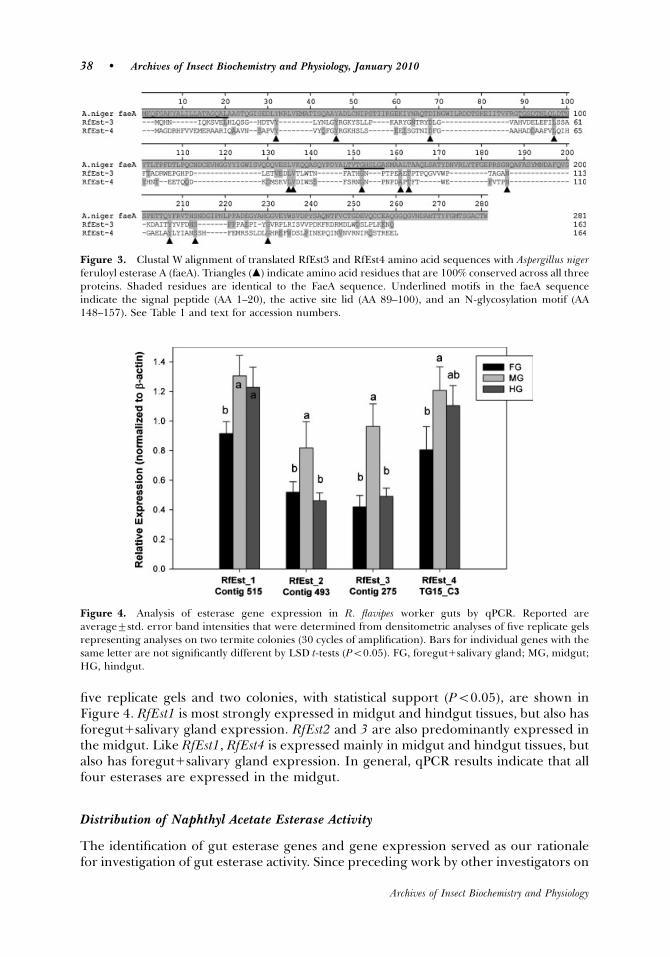
five replicate gels and two colonies, with statistical support (Po0.05), are shown inFigure 4. RfEst1 is most strongly expressed in midgut and hindgut tissues, but also hasforegut1salivary gland expression. RfEst2 and 3 are also predominantly expressed inthe midgut. Like RfEst1, RfEst4 is expressed mainly in midgut and hindgut tissues, butalso has foregut1salivary gland expression. In general, qPCR results indicate that allfour esterases are expressed in the midgut.
Distribution of Naphthyl Acetate Esterase Activity
The identification of gut esterase genes and gene expression served as our rationalefor investigation of gut esterase activity. Since preceding work by other investigators on
Figure 3. Clustal W alignment of translated RfEst3 and RfEst4 amino acid sequences with Aspergillus nigerferuloyl esterase A (faeA). Triangles (m) indicate amino acid residues that are 100% conserved across all threeproteins. Shaded residues are identical to the FaeA sequence. Underlined motifs in the faeA sequenceindicate the signal peptide (AA 1–20), the active site lid (AA 89–100), and an N-glycosylation motif (AA148–157). See Table 1 and text for accession numbers.
Figure 4. Analysis of esterase gene expression in R. flavipes worker guts by qPCR. Reported areaverage7std. error band intensities that were determined from densitometric analyses of five replicate gelsrepresenting analyses on two termite colonies (30 cycles of amplification). Bars for individual genes with thesame letter are not significantly different by LSD t-tests (Po0.05). FG, foregut1salivary gland; MG, midgut;HG, hindgut.
38 � Archives of Insect Biochemistry and Physiology, January 2010
Archives of Insect Biochemistry and Physiology

R. flavipes esterases used whole-body protein preparations, as a first step we initiallycompared gut and carcass esterases. For this purpose, native PAGE was used incombination with hydrolysis of the model esterase substrate a-naphthyl acetate (Fig. 5).Although a significant proportion of total esterase activity is located in the termitecarcass, a diverse number of esterase isoforms are also present in different gut regions.Within gut tissues, naphthyl acetate staining was strongest in the midgut, followed bythe foregut1salivary gland and hindgut regions. Minor differences in bandingpatterns occurred between the two colonies that were investigated; however, resultswere consistent across the two colonies.
Gut Esterase Activity Towards Naphthyl Esters With Varying Carbon Chain Lengths
Activities toward different naphthyl ester substrates were investigated by gut regionusing native PAGE (Fig. 6A). Gut esterases are most active towards naphthyl estersubstrates with side chains of 3 (a-naphthyl propionate) and 4 (a-naphthyl butyrate)carbons, followed by the 2-carbon substrate a-naphthyl acetate. This result wasconsistent for all three termite gut tissues examined. For all three tissues, the lowestesterase staining intensity occurred with a-naphthyl valerate and caprylate (C5 and C6,respectively). PAGE results were confirmed by statistical analysis of absorbance datafrom colorimetric microplate assays (Fig. 6B). Here, all three termite gut regionsshowed the significantly greatest activity toward a-naphthyl propionate (Po0.05).Differences in activity between a-naphthyl butyrate and acetate were more evident incolorimetric assays than native PAGE. Together, colorimetric and native PAGE resultsare in agreement on the points that (1) the majority and greatest diversity of gutesterase activity is located in the midgut, and (2) that activity is greatest towards the 3carbon ester naphthyl propionate. In particular, naphthyl propionate findings arenoteworthy because ferulic acid esterases also show the preferentially highest activitytoward this substrate.
Esterase Activity Among Gut Sub-Cellular Fractions and in Hindgut Excretions
Subcellular esterase assays used only a-naphthyl propionate as a substrate(Fig. 7) based on results presented above showing maximal activity towardthis substrate. All subcellular fractions showed prominent esterase activity in twogroups, hereafter referred to as groups 1 and 2. The group 1 esterases were lessmobile and included a mixture of soluble and membrane-bound forms in both the
Figure 5. Native PAGE analysis of R. flavipes worker gut and carcass esterases using a-naphthyl acetate as asubstrate. Two colonies were examined using 14,000 g supernatants from homogenized tissues of the foregut1salivary gland (FG), midgut (MG), hindgut (HG), and headless carcass that remained after gut removal(carcass).
Termite Gut Esterases � 39
Archives of Insect Biochemistry and Physiology

mitochondrial and microsomal fractions. The group 2 esterases include entirelysoluble forms that only occur in the cytosolic fraction after microsomal precipitation.The three bands observed in the gut microsomal fraction appear very similar to thethree microsomal esterases purified from R. flavipes whole body fractions by Valleset al. (2001). Finally, a strong band of esterase activity that aligned with the group Iesterases on native PAGE gels was present in hindgut luminal supernatant (Fig. 7,right).
Figure 6. Analysis of R. flavipes worker gut esterases by (A) native PAGE and (B) colorimetric microplateassays. Both analyses compared the a-naphthyl ester substrates naphthyl acetate (ace), naphthyl propionate(prop), naphthyl butyrate (but), naphthyl valerate (val), and naphthyl caprylate (cap). 14,000gsupernatants from homogenized tissues of the foregut1salivary gland (FG), midgut (MG), and hindgut(HG) were compared. B: Bars within graphs with the same letter are not significantly different by LSD t-tests(Po0.05).
Figure 7. Native PAGE gel showing the subcellular distribution of esterase activity toward a-naphthylpropionate in the whole R. flavipes worker gut (left) and hindgut lumen (right). See text for isolation protocols.Numbered brackets, respectively, denote (1) isoforms with both soluble and membrane distributions, and (2)isoforms that are exclusively soluble. MITO, solubilized mitochondrial pellet; CYTO1MICRO, post-mitochondrial supernatant containing soluble/cytosolic and membrane/microsomal fractions; CYTO, cytosolicfraction remaining after microsomal precipitation; MICRO, re-suspended microsomal pellet.
40 � Archives of Insect Biochemistry and Physiology, January 2010
Archives of Insect Biochemistry and Physiology

DISCUSSION
JH Esterases and Semiochemical/Hormone Processing
JH esterases are members of the carboxylesterase family (EC 3.1.1.1). They aretypically synthesized in the fat body and secreted into the hemolymph where theycatabolize JH with high affinity (Goodman and Granger, 2005). JH is broadlyimportant in insect physiology, playing roles in juvenile development, adultreproductive competence, polyphenism, and in social insects, caste differentiation,and behavioral polyethism (Gilbert et al., 2000; Hartfelder, 2000; Goodman andGranger, 2005). In termites, JH has established links to worker-to-soldier castedifferentiation (Park and Raina, 2004, 2005; Mao et al., 2005; Elliott and Stay, 2008;Cornette et al., 2008) and vitellogenesis in adult female reproductives (Brent et al.,2005; Elliott and Stay, 2007; Cornette et al., 2008). Within the context of eusociality,the termite gut is not only important to lignocellulose digestion and nutrition; it is alsopotentially important because of the potential flow of hormones, pheromones, andsemiochemicals through the gut to nest-mates via trophallaxis. With respect tohormonal processing and caste regulation, previous sequencing from a R. flavipes gutlibrary revealed a number of endogenous (termite-derived) genes encoding enzymesthat participate in JH biosynthesis and degradation (Tartar et al., 2009; M. R. Tarver,unpublished data). JH biosynthetic genes that have been identified from the R. flavipesdigestive tract encode several enzymes of the mevalonic acid pathway (Belles et al.,2005), including HMG Co-A reductase, two mevalonate kinases, and two farnesoic acidmethyltransferases. Two family-15 cytochrome P450 genes were also identifiedfrom R. flavipes (M. R. Tarver and M. R. Coy, unpublished data) with homology toP450s that play roles in the final step in JH biosynthesis: methyl farnesoateepoxidation (Feyereisen, 2005). Finally, a number of endogenous gut genes wereidentified with homology to insect genes that participate in JH degradation. These JHdegradation genes include P450s, epoxide hydrolases, and the putative JHesterases RfEst1 and 2. As proposed by Henderson (1998), many of these genesincluding JH esterases could play roles in caste regulation/differentiation bymodulating JH, semiochemical, and/or terpene biosynthesis and availability. Theexpression of endocrine-linked genes in insect guts has been reported previously. Forexample, gut expression of mevalonic pathway and P450 genes was previouslyidentified in bark beetles (Keeling et al., 2004, 2006), and a putative JH esterasefrom the beetle P. hilaris was found to have expression in the foregut and midgut(Munyiri and Ishikawa, 2007). Given the importance of chemical communicationand trophallaxis in termite society, it is interesting that genes encoding proteinswith homology to members of JH biosynthetic and degradation pathways areassociated with the termite gut, and that soluble esterases are present in hindgutexcretions (Fig. 7).
The RfEst1 and 2 genes identified in the current study share significant translatedidentity with several putative insect JH esterases, including an RNAi-validated JHesterase from a social insect, the honey bee A. mellifera (Mackert et al., 2008). Commonfeatures shared among the RfEst1 and honey bee JH esterase proteins include a signalpeptide (MAETVTVTVAQ; Figs. 1 and 2) and distinct JH esterase catalytic motifs,such as a catalytic triad (G-SAG, E/D, and G--H-D-) and two putative substraterecognition motifs (RF, DQ) (Campbell et al., 2001; Goodman and Granger, 2005;Munyiri and Ishikawa, 2007; Mackert et al., 2008). Of the five species with significant
Termite Gut Esterases � 41
Archives of Insect Biochemistry and Physiology

homology compared in the JH esterase alignment (Fig. 2), it is also noteworthy thatR. flavipes, P. hilaris, and A. rosae are lignocellulose feeders and are strongly dependenton lignocellulose degradation for nutrient acquisition. Thus, although RfEst-1 and 2have homology to insect JH esterases, it remains unclear if they also play roles inlignocellulose digestion. Gene expression profiling and RNAi studies will help toresolve the physiological roles of RfEst1and 2.
Phenolic Acid Esterases
Phenolic acid esterases, also known as feruloyl, coumaryl, or cinnamoyl esterases, playroles in hemicellulose and pectin depolymerization by cleaving ester bonds thatconnect ferulic and coumaric acids to hemicellulose and pectin sugar monomers (Saha,2003; Crepin et al., 2004; Anderson and Akin, 2008; Benoit et al., 2008). Of these twosubstrate classes, hemicellulose is a significant component of the termite diet.Hemicellulose, also referred to as xylan, is a complex b-linked sugar polymer thatcomposes approximately 25% of lignocellulose; it represents a significant obstacle tocellulose digestion by cellulolytic organisms (Lange, 2007; Scharf and Tartar, 2008).Phenolic acid esterases catalyze a critical step in cellulose and hemicellulosedepolymerization by dissociating both polymers from each other and lignin, makingeach more accessible to hemicellulases and cellulases (Benoit et al., 2008; Scharf andTartar, 2008).
‘‘General’’ esterases are a large group of insect esterases that are usually activetoward naphthyl ester substrates (Oakeshott et al., 2005). By this definition, thenumerous isoforms identified by native PAGE in the current study can be classified asgeneral esterases. Aside from roles in lipid and xenobiotic metabolism, the roles ofinsect general esterases remain mostly undefined (Oakeshott et al., 2005). Onehypothesis is that general esterases from the termite gut may participate inhemicellulose depolymerization by acting as phenolic acid esterases.
Although very little ferulic/phenolic acid esterase activity has been reported fromthe animal kingdom, a large number of such activities have been identified fromvarious microbial sources (reviewed by Crepin et al., 2004; Benoit et al., 2008). Two ofthe first microbial ferulic acid esterases to be identified, named faeA and faeB, comefrom the fungus Aspergillus niger (de Vries et al., 1997, 2002). FaeA is a smaller proteinof approximately 30 kDa, while faeB is a larger protein of approximately 74 kDa. Inamino acid alignments of each A. niger fae with the four translated termite esterases,none were statistically significant; however, faeA does align at key positions with theshort 20–23-kDa peptides encoded by RfEst3 and 4 (Fig. 3). In particular, 13 aminoacids show 100% conservation across the alignment. However, based on their lack ofN-terminal homology with faeA, RfEst3 and 4 may be subunits of larger functionalmultimeric proteins (e.g., Sreerama and Veerabhadrappa, 1991), or their ORFs maybe re-defined after additional sequencing. It is also noteworthy that (1) faeA showspreferential activity toward hemicellulose side-chain esters (de Vries et al., 2002), (2)the termite diet contains significant quantities of hemicellulose, and (3) there is a largecomplement of hemicellulase genes encoded within the R. flavipes gut meta-transcriptome (Scharf and Tartar, 2008; Tartar et al., 2009). Finally, additional supportfor the idea that phenolic acid hydrolysis occurs in the R. flavipes gut is derived fromthe significantly elevated activity observed toward a-naphthyl butyrate, a characteristicdisplayed by some ferulate esterases (Koseki et al., 2006).
42 � Archives of Insect Biochemistry and Physiology, January 2010
Archives of Insect Biochemistry and Physiology

Relation to Previous Studies on Termite Esterase Biochemistry
Esterase biochemistry has previously been studied in several termite species, includingR. flavipes. A series of two reports (Sreerama and Veerabhadrappa, 1991, 1993)provided the first evidence of esterase activity in the termite gut. These studiesinvestigated midgut and fungal symbiont esterases from the higher termiteOdontotermes horni. In the 1991 report, two �78-kDa midgut esterases were purifiedto homogeneity by column chromatography; upon treatment with b-mercaptoethanol,the proteins disassociated into two subunits of �40 kDa. Both termite midgut esterasesshowed optimal catalytic activity at pH 7, as well as preferentially greater activitytoward a-naphthyl acetate than propionate or butyrate. In their 1993 report,Sreerama and Veerabhadrappa purified esterases from the Odontotermes horni fungalsymbiont Xylaria nigripes. Four esterases ranging in size from �85–88 kDa werepurified to homogeneity and found to have considerably different biochemical andcatalytic properties than the endogenous midgut esterases. In addition, starvationstudies revealed little impact on host and symbiont esterase activity, while distributionstudies revealed that 498% of esterase activity was host-derived. These studiessuggested little importance for fungal symbionts in hydrolytic metabolism ofcarboxylester substrates in Odontotermes horni, which agrees with the identification ofesterase genes from only a symbiont-free R. flavipes gut cDNA library (Scharf andTartar, 2008, Tartar et al., 2009), and gene expression and activity from gut regionsthat do not house symbionts (current study).
Prior studies also examined esterases in both hemolymph and whole-bodypreparations of different termite species. Wyss-Huber (1981) investigated hemolymph-soluble esterases in the higher and lower termites Macrotermes subhyalinus andZootermopsis angusticollis and identified caste- and phenotype-specific native PAGEbanding patterns in both species, supporting a role for possible hemolymph JHesterases in caste differentiation and/or caste homeostasis. Another study by Ruvolo-Takasusuki and Collett (2000) investigated esterases from whole-body preparations ofdifferent castes of the higher termite Nasutitermes globiceps. Since whole-bodypreparations were used, it was not possible to determine which N. globiceps esterases,if any, are expressed in the gut. However, as with the work of Wyss-Huber (1981), thefindings of Ruvolo-Takasusuki and Collett (2000) revealed a diversity of esteraseisoforms by native PAGE, which is highly consistent with findings of the current study.
Another study on the related Rhinotermitid lower termite Coptotermes formosanusinvestigated esterase isoform composition in both gut and carcass tissue fractions(Wang and Grace, 2000). Similar to the results of the current research, this studyidentified a significant amount of esterase activity and isoform diversity in gut tissues.Activity in the C. formosanus carcass was also identified, but at much lowers levels thanin the gut. In addition, starvation and forced feeding on low molecular cellulose bothled to reduction of a single esterase band, suggesting that it was from the protistsymbiont Pseudotrichonympha grassii (Yoshimura, 1995; Wang and Grace, 2000). Similar1-wk starvation studies were conducted on R. flavipes (not shown), but revealed noimpact on gut esterase banding patterns, further supporting our conclusion that gutesterases in R. flavipes are host-derived. Thus, in both C. formosanus and R. flavipes gutesterase activity is prominent and apparently produced by host tissues.
Two prior studies focused on esterase biochemistry in R. flavipes, but did notconsider the gut independently. Davis et al. (1995) investigated esterases from workerwhole-body preparations by native PAGE and identified a similar number of isoforms
Termite Gut Esterases � 43
Archives of Insect Biochemistry and Physiology

active toward a-naphthyl acetate as were observed in the present study. They alsoidentified maximal activity at gut physiological pH (7–7.6; Brune et al., 1995) andhighest activity towards the substrate a-naphthyl butyrate. While a-naphthylpropionate hydrolysis was the highest gut activity observed in the present study,Davis et al. (1995) reported higher a-naphthyl butyrate and acetate activity from theirwhole body protein preparations
Valles et al. (2001) purified three trans-permethrin hydrolyzing esterases fromwhole-body microsomal extracts of R. flavipes. In native form, the permethrin esterasesseem to correspond to three of the several isoforms visible at the top of native PAGEgels in the present study (Figs. 5–7). In denatured form, the permethrin esterasesdisplayed molecular masses (�63 kDa) very similar to the predicted RfEst1 mass of59 kDa. It is not known if any of the permethrin esterases are expressed in gutmicrosomes; however, our native PAGE results strongly suggests their presence in thegut. The permethrin esterases were also most active at gut physiological pH, and theywere more active toward a-naphthyl acetate than butyrate (propionate was not tested).Finally, the three permethrin esterases were found to be heavily glycosylated (Valleset al., 2001), while the RfEst1 protein is predicted to have at least 30 glycosylation sitesscattered throughout the mature peptide (results not shown). Thus, despitedifferences in substrate specificity, findings of the current study support the hypothesisthat the RfEst1 gene encodes at least one of the three microsomal permethrin esteraseisoforms reported by Valles et al. (2001), and that these esterases occur in gutmicrosomes.
Host Versus Symbiont Origins for Esterases
While the current study provides evidence suggesting the existence of gut esterases intermite genomes, gut esterases have previously been identified from gut symbioticbacteria and fungi of higher termites. McSweeney et al. (1999) identified esteraseactivity from Clostridium xylanolyticum, a bacterial gut symbiont of the grass-feedinghigher termite Tumilitermes pastinator. Clostridium bacteria have been sampled fromR. flavipes (Fisher et al., 2007); however, there is no existing evidence suggesting thatsymbiotic bacteria in lower termites possess lignocellulolytic abilities (Tokuda andWatanabe, 2007). Pure isolates of C. xylanolyticum were found to produce three esteraseisoforms based on native PAGE and staining with a-naphthyl substrates. TheC. xylanolyticum esterases showed highest activity toward the model substratesa-naphthyl acetate, p-nitrophenyl acetate, and phenyl acetate, but no activity towarda-naphthyl butyrate. Following partial purification, two of the three esterase isoformsshowed significant activity toward the ferulic acid and p-coumaric acid esters FAXX andPAXX (McSweeney et al., 1999). No gene sequences are available for the C. xylanolyticumesterases. Other termite fungal symbiont esterases have been identified from the highertermite Odontotermes horni (Sreerama and Veerabhadrappa, 1993; see above); however,their true substrates are unknown and they represent o5% of gut esterase activity fromtheir host termite. Also, a gut esterase putatively from the C. formosanus protist symbiontP. grassii was previously identified (Wang and Grace, 2000; see above), but again, thisenzyme represents only a small fraction of total gut esterase activity. Our findings in thepresent research do not suggest microbial symbiont origins for R. flavipes esterasesbased on four criteria: (1) the RfEst1-4 genes, as well as 8 others, were identifiedexclusively from a symbiont-free termite gut cDNA library, (2) the genes were notsampled from a hindgut symbiont library, (3) all significant database matches for the
44 � Archives of Insect Biochemistry and Physiology, January 2010
Archives of Insect Biochemistry and Physiology

RfEst1-4 genes are from insects, and (4) esterase activity is clearly abundant in symbiont-free termite gut regions (Scharf and Tartar, 2008; Tartar et al., 2009; Figs. 5 and 6).
Summary and Conclusions
This study investigated esterase gene expression and activity in the gut of the termiteR. flavipes. Our goals were to attempt to correlate esterase gene and protein expressionacross the R. flavipes digestive tract, and to begin to develop an understanding of thepotential physiological roles played by esterases in gut tissues and the gut lumen. Fouresterase genes and deduced protein sequences from the gut were first investigated.These genes were determined to encode two JH esterase-like proteins and two muchsmaller proteins with similarity to fungal phenolic acid esterases. Each of the fourgenes has midgut expression, with variable expression among other gut regions,depending on the particular gene. Additionally, an unexpected diversity of esteraseprotein isoforms was identified in different gut regions, particularly in the midgut.Activity towards a number of model substrates was also highest in the midgut region,and esterase protein isoforms were ubiquitously expressed among the mitochondrial,cytosolic, and microsomal sub-cellular fractions. Soluble esterase isoforms are alsopresent in hindgut excretions, suggesting that they and other soluble enzymes can betransferred among termites via proctodeal trophallaxis.
Based on the activity and gene expression observed in gut regions that do nothouse microbial symbionts (foregut and midgut), as well as other observations, ourfindings strongly suggest that R. flavipes gut esterase activity is not symbiont-derived.Thus, the four esterase genes are likely encoded in the termite genome, transcribedand translated by termite gut cells, and potentially secreted into the gut lumen. Thesefindings suggest several interesting possibilities not considered previously in termitedigestive research, namely that ‘‘general’’ esterases in the termite gut may be playingroles in hormone/semiochemical biosynthesis and degradation, as well as lignocellulosedepolymerization, and that they may be distributed among nestmates via trophallaxis.The research presented here (1) helps to validate our gut meta-transcriptome databaseas a functional-genomic resource (Scharf and Tartar, 2008; Tartar et al., 2009), and (2)provides important background information to assist in the testing of hypothesesrelating to the termite gut as a physiological foundation that underlies termite sociality.
ACKNOWLEDGMENTS
We thank Aurelien Tartar for esterase contig assembly and preliminary sequenceanalyses and Drion Boucias for helpful advice and manuscript critiques. Support of theresearch by DOE and the Consortium for Plant Biotechnology Research, Inc., does notconstitute an endorsement by DOE or by the Consortium for Plant BiotechnologyResearch, Inc., of the views expressed in this publication.
LITERATURE CITED
Anderson WF, Akin DE. 2008. Structural and chemical properties of grass lignocelluloses relatedto conversion for biofuels. J Ind Microbiol Biotechnol 35:355–366.
Belles X, Martın D, Piulachs MD. 2005. The mevalonate pathway and the synthesis of juvenilehormone in insects. Annu Rev Entomol 50:181–199.
Termite Gut Esterases � 45
Archives of Insect Biochemistry and Physiology

Benoit I, Danchin EGJ, Bleichrodt RJ, de Vries RP. 2008. Biotechnological applications andpotential of fungal feruloyl esterases based on prevalence, classification and biochemicaldiversity. Biotechnol Lett 30:387–396.
Brent CS, Schal C, Vargo EL. 2005. Endocrine changes in maturing primary queens ofZootermopsis angusticollis. J Insect Physiol 51:1200–1209.
Breznak JA, Brune A. 1995. Role of microorganisms in the digestion of lignocellulose bytermites. Annu Rev Entomol 39:453–487.
Brune A, Emerson D, Breznak JA. 1995. Termite gut microflora as an oxygen sink,microelectrode determinations of oxygen and pH gradients in guts of lower and highertermites. Appl Env Microbiol 61:2681–2687.
Campbell PM, Harcourt RL, Crone EJ, Claudianos C, Hammock BD, Russell RJ, Oakeshott JG.2001. Identification of a juvenile hormone esterase gene by matching its peptide massfingerprint with a sequence from the Drosophila genome project. Insect Biochem Mol Bio31:513–520.
Cornette R, Gotoh H, Koshikawa S, Miura T. 2008. Juvenile hormone titers and castedifferentiation in the damp-wood termite Hodotermopsis sjostedti (Isoptera, Termopsidae).J Insect Physiol 54:922–930.
Crepin VF, Faulds CB, Connerton IF. 2004. Functional classification of the microbial feruloylesterases. Appl Microbiol Biotechnol 63:647–652.
Davis RW, Kamble ST, Prabhakaran SK. 1995. Characterization of general esterases in workersof the Eastern subterranean termite. J Econ Entomol 88:574–578.
de Vries RP, Michelsen B, Poulsen CH, Kroon PA, van den Heuvel RH, Faulds CB,Williamson G, van den Hombergh JP, Visser J. 1997. The faeA genes from Aspergillusniger and Aspergillus tubingensis encode ferulic acid esterases involved in degradation ofcomplex cell wall polysaccharides. Appl Environ Microbiol 63:4638–4644.
de Vries RP, vanKuyk PA, Kester HC, Visser J. 2002. The Aspergillus niger faeB gene encodes asecond feruloyl esterase involved in pectin and xylan degradation and is specifically inducedin the presence of aromatic compounds. Biochem J 363:377–386.
Elliott KL, Stay B. 2007. Juvenile hormone synthesis as related to egg development in neotenicreproductives of the termite Reticulitermes flavipes, with observations on urates in the fatbody. Gen Comp Endocrinol 152:102–110.
Elliott KL, Stay B. 2008. Changes in juvenile hormone synthesis in the termite Reticulitermesflavipes during development of soldiers and neotenic reproductives from groups of isolatedworkers. J Insect Physiol 54:492–500.
Feyereisen R. 2005. Insect cytochrome P450. In: Gilbert LI, Iatrou K, Gill SS, editors.Comprehensive molecular insect science, Vol. 4. Oxford: Elsevier-Pergamon. p 1–77.
Fisher M, Miller D, Brewster C, Husseneder C, Dickerman A. 2007. Diversity of gut bacteria ofReticulitermes flavipes as examined by 16S rRNA gene sequencing and amplified rDNArestriction analysis. Curr Microbiol 55:254–259.
Gilbert LI, Granger NA, Roe RM. 2000. The juvenile hormones, historical facts and speculationson future research directions. Insect Biochem Mol Bio 30:617–644.
Goodman WG, Granger NA. 2005. The juvenile hormones. In: Gilbert LI, Iatrou K, Gill SS, editors.Comprehensive molecular insect science, Vol. 3. Oxford: Elsevier-Pergamon. p 319–408.
Hamby SE, Hirst JD. 2008. Prediction of glycosylation sites using random forests. BMCBioinformatics 9:500.
Hartfelder K. 2000. Insect juvenile hormone, from ‘‘status quo’’ to high society. Braz J Med BiolRes 33:157–177.
Henderson G. 1998. Primer pheromones and possible soldier caste influence on the evolution ofsociality in lower termites. In: Vander Meer RK, Breed MD, Winston ML, Espelie KE,editors. Pheromone communication in social insects. Boulder: Westview Press. p 314–330.
46 � Archives of Insect Biochemistry and Physiology, January 2010
Archives of Insect Biochemistry and Physiology

Keeling CI, Blomquist GJ, Tittiger C. 2004. Coordinated gene expression for pheromonebiosynthesis in the pine engraver beetle, Ips pini (Coleoptera, Scolytidae). Naturwissenschaften91:324–328.
Keeling CI, Bearfield JC, Young S, Blomquist GJ, Tittiger C. 2006. Effect of juvenile hormoneon gene expression in the pheromone-producing midgut of the pine engraver beetle, Ipspini. Insect Mol Biol 15:207–216.
Korb J, Hoffman K, Hartfelder K. 2009. Endocrine signatures underlying plasticity inpostembryonic development of a lower termite, Cryptotermes secundus. Evol Dev 11:269–277.
Koseki T, Takahashi K, Handa T, Yamane Y, Fushinobu S, Hashizume K. 2006. N-linkedoligosaccharides of Aspergillus awamori feruloyl esterase are important for thermostabilityand catalysis. Biosci Biotechnol Biochem 70:2476–2480.
Kupfer D, Levine E. 1972. Monooxygenase drug metabolism activity in CaCl2-aggregatedhepatic microsomes from rat liver. Biochem Biophys Res Commun 47:611–618.
Laine LV, Wright DJ. 2003. The life cycle of Reticulitermes spp (Isoptera: Rhinotermitidae): whatdo we know? Bull Entomol Res 93:267–378.
Lange JP. 2007. Lignocellulose conversion, an introduction to chemistry, process and economics.Biofuels Bioprod Bioref 1:39–48.
Ljungdahl LG, Erickson KE. 1985. Ecology of microbial cellulase degradation. Adv Micro Ecol8:237–299.
Mackert A, de Nascimento AM, Bitondi MM, Hartfelder K, Simoes ZL. 2008. Identification of ajuvenile hormone esterase-like gene in the honey bee, Apis mellifera L.-expression analysisand functional assays. Comp Biochem Physiol B Biochem Mol Bio 150:33–44.
Mao L, Henderson G, Liu Y, Laine RA. 2005. Formosan subterranean termite (Isoptera,Rhinotermitidae) soldiers regulate juvenile hormone levels and caste differentiation inworkers. Ann Entomol Soc Am 98:340–345.
McSweeney CS, Dulieu A, Webb RI, Del Dot T, Blackwell LL. 1999. Isolation andcharacterization of a Clostridium sp. with cinnamoyl esterase activity and unusual cellenvelope ultrastructure. Arch Microbiol 172:139–149.
Munyiri FN, Ishikawa Y. 2007. Molecular cloning and developmental expression of the geneencoding juvenile hormone esterase in the yellow-spotted longicorn beetle, Psacothea hilaris.Insect Biochem Mol Bio 37:497–505.
Oakeshott JG, Claudianos C, Campbell PM, Newcombe RD, Russell RJ. 2005. Biochemicalgenetics and genomics of insect esterases. In: Gilbert LI, Iatrou K, Gill SS, editors.Comprehensive molecular insect science, Vol. 5. Oxford: Elsevier-Pergamon. p 309–382.
Ohkuma M. 2003. Termite symbiotic systems: efficient bio-recycling of lignocellulose. ApplMicrobiol Biotechnol 61:1–9.
Park YI, Raina AK. 2004. Juvenile hormone III titers and regulation of soldier caste inCoptotermes formosanus (Isoptera, Rhinotermitidae). J Insect Physiol 50:561–566.
Park YI, Raina AK. 2005. Regulation of juvenile hormone titers by soldiers in the Formosansubterranean termite, Coptotermes formosanus. J Insect Physiol 51:385–391.
Rozen S, Skaletsky HJ. 2000. Primer3 on the WWW for general users and for biologistprogrammers. In: Krawetz S, Misener S, editors. Bioinformatics methods and protocols,methods in molecular biology. Totowa, NJ: Humana Press. p 365–386.
Ruvolo-Takasusuki MCR, Collet T. 2000. Characterization of Nasutitermes globiceps esterases.Biochem Genet 38:367–375.
Saha BC. 2003. Hemicellulose conversion. J Indust Microbiol Biotechnol 30:279–291.
Scharf ME, Tartar A. 2008. Termite digestomes as sources for novel lignocellulases. BiofuelsBioprod Biorefin 2:540–552.
Termite Gut Esterases � 47
Archives of Insect Biochemistry and Physiology

Scharf ME, Neal JJ, Bennett GW. 1998. Changes in insecticide resistance levels and detoxicationenzymes following insecticide selection in the German cockroach, Blattella germanica. PesticBiochem Physiol 59:67–79.
Scharf ME, Ratliff CR, Wu-Scharf D, Zhou X, Pittendrigh BR, Bennett GW. 2005. Effects ofjuvenile hormone III on Reticulitermes flavipes: changes in hemolymph protein compositionand gene expression. Insect Biochem Mol Biol 35:207–215.
Sreerama L, Veerabhadrappa PS. 1991. Purification and properties of carboxylesterases fromthe mid-gut of the termite Odontotermes horni. Insect Biochem 21:833–844.
Sreerama L, Veerabhadrappa PS. 1993. Isolation and properties of carboxylesterases of thetermite gut-associated fungus, Xylaria nigripes, and their identity from the host termite,Odontotermes horni. Int J Biochem 25:1637–1651.
Taprab Y, Johjima T, Maeda Y, Moriya S, Trakulnaleamsai S, Noparatnaraporn N, Okhuma M,Kudo T. 2005. Symbiotic fungi produce laccases potentially involved in phenol degradationin fungus combs of fungus-growing termites in Thailand. Appl Env Microbiol 71:7696–7704.
Tartar A, Wheeler MM, Zhou X, Coy MR, Boucias DG, Scharf ME. 2009. Meta-transcriptomeanalyses of host and symbiont gene expression in the gut of the termite Reticulitermes flavipes.Biotechnol Biofuels (in press).
Tarver MR, Schmelz EA, Rocca JR, Scharf ME. 2009. Effects of soldier-derived terpenes onsoldier caste differentiation in the termite Reticulitermes flavipes. J Chem Ecol 35:256–264.
Tokuda G, Watanabe H. 2007. Hidden cellulases in termites, revision of an old hypothesis. BiolLett 3:336–339.
Valles SM, Oi FM, Strong CA. 2001. Purification and characterization of trans-permethrinmetabolizing microsomal esterases from workers of the eastern subterranean termite,Reticulitermes flavipes. Insect Biochem Mol Biol 31:715–725.
Wang J, Grace JK. 2000. Allozyme patterns of protozoan symbionts of Coptotermes formosanus.Sociobiology 35:467–474.
Warnecke F, Luginbuhl P, Ivanova N, Ghassemian M, Richardson TH, Stege JT, Cayouette M,McHardy AC, Djordjevic G, Aboushadi N, Sorek R, Tringe SG, Podar M, Martin HG,Kunin V, Dalevi D, Madejska J, Kirton E, Platt D, Szeto E, Salamov A, Barry K,Mikhailova N, Kyrpides NC, Matson EG, Ottesen EA, Zhang X, Hernandez M, Murillo C,Acosta LG, Rigoutsos I, Tamayo G, Green BD, Chang C, Rubin EM, Mathur EJ,Robertson DE, Hugenholtz P, Leadbetter JR. 2007. Metagenomic and functional analysis ofhindgut microbiota of a wood-feeding higher termite. Nature 450:560–565.
Watanabe H, Noda H, Lo N. 1998. A cellulase gene of termite origin. Nature 394:330–331.
Wheeler WM. 1918. A study of some ant larvae with a consideration of the origin and meaningof social habits among insects. Proc Am Phil Soc 57:293–343.
Wilson EO. 1971. The insect societies. Cambridge: Harvard University Press. 560p.
Wyss-Huber M. 1981. Caste differences in haemolymph proteins in two species of termites.Insect Soc 28:71–86.
Yoshimura T. 1995. Contribution of protozoan fauna to nutritional physiology of the lowertermite, Coptotermes formosanus. Ph.D. Dissertation. University of Kyoto, Japan.
48 � Archives of Insect Biochemistry and Physiology, January 2010
Archives of Insect Biochemistry and Physiology





























