Embed Size (px)
Citation preview

JOURNAL OF VIROLOGY, July 2007, p. 7529–7539 Vol. 81, No. 140022-538X/07/$08.00�0 doi:10.1128/JVI.00327-07Copyright © 2007, American Society for Microbiology. All Rights Reserved.
Characterization of Low-Pathogenic H5 Subtype Influenza Virusesfrom Eurasia: Implications for the Origin of
Highly Pathogenic H5N1 Viruses�
L. Duan,1,2 L. Campitelli,3 X. H. Fan,1 Y. H. C. Leung,1 D. Vijaykrishna,1,2 J. X. Zhang,1,2
I. Donatelli,3 M. Delogu,4 K. S. Li,2 E. Foni,5 C. Chiapponi,5 W. L. Wu,1,2 H. Kai,1,2
R. G. Webster,1,6 K. F. Shortridge,1,2 J. S. M. Peiris,1,2 Gavin J. D. Smith,1,2
H. Chen,1,2* and Y. Guan1,2*State Key Laboratory of Emerging Infectious Diseases, Department of Microbiology, The University of Hong Kong, Faculty of
Medicine Building, 21 Sassoon Road, Pokfulam, Hong Kong, SAR, China1; International Institute of Infection and Immunity,Shantou University, Shantou, Guangdong 515031, China2; Department of Infectious, Parasitic and Immune-Mediated Diseases,Istituto Superiore di Sanita, Viale Regina Elena 299, Rome, Italy3; Department of Public Health and Animal Pathology,
Faculty of Veterinary Medicine, University of Bologna, Ozzano Emilia (BO), Italy4; Istituto Zooprofilattico Sperimentale ofLombardia and Emilia, Parma, Italy5; and Virology Division, Department of Infectious Diseases,
St. Jude Children’s Research Hospital, Memphis, Tennessee 380156
Received 14 February 2007/Accepted 3 May 2007
Highly pathogenic avian influenza (HPAI) H5N1 viruses are now endemic in many Asian countries, resultingin repeated outbreaks in poultry and increased cases of human infection. The immediate precursor of theseHPAI viruses is believed to be A/goose/Guangdong/1/96 (Gs/GD)-like H5N1 HPAI viruses first detected inGuangdong, China, in 1996. From 2000 onwards, many novel reassortant H5N1 influenza viruses or genotypeshave emerged in southern China. However, precursors of the Gs/GD-like viruses and their subsequentreassortants have not been fully determined. Here we characterize low-pathogenic avian influenza (LPAI) H5subtype viruses isolated from poultry and migratory birds in southern China and Europe from the 1970s to the2000s. Phylogenetic analyses revealed that Gs/GD-like virus was likely derived from an LPAI H5 virus inmigratory birds. However, its variants arose from multiple reassortments between Gs/GD-like virus andviruses from migratory birds or with those Eurasian viruses isolated in the 1970s. It is of note that unlike HPAIH5N1 viruses, those recent LPAI H5 viruses have not become established in aquatic or terrestrial poultry.Phylogenetic analyses revealed the dynamic nature of the influenza virus gene pool in Eurasia with repeatedtransmissions between the eastern and western extremities of the continent. The data also show reassortmentbetween influenza viruses from domestic and migratory birds in this region that has contributed to theexpanded diversity of the influenza virus gene pool among poultry in Eurasia.
Aquatic birds are considered the natural reservoirs of influ-enza A virus, as all known subtypes (H1 to H16 and N1 to N9)have been isolated from aquatic birds (1, 16, 21, 25, 38, 45, 48).The results of genetic analyses have also suggested that allinfluenza viruses found in other hosts were derived from thoseviruses resident in aquatic birds (45). Influenza viruses fromaquatic birds have occasionally been transmitted to terrestrialpoultry, leading to disease outbreaks; however, only some H5and H7 subtypes of influenza viruses have evolved into highlypathogenic strains that have caused significant mortality inpoultry (2, 4, 13, 14, 23).
The H5N1 “bird flu” incident in Hong Kong, China, in 1997demonstrated that a “pure” avian influenza virus could bedirectly introduced into humans and cause infection (9, 10, 41),suggesting a possible pathway for the generation of a newpandemic influenza (31). Genetic analyses suggested that the
H5N1 virus causing human infection in Hong Kong, repre-sented by A/Hong Kong/156/97 (H5N1/97), might have beengenerated by multiple reassortment between A/goose/Guang-dong/1/96 (Gs/GD)-like and H9N2 or H6N1 viruses found interrestrial poultry (19, 22, 50). Following the 1997 outbreak,highly pathogenic avian influenza (HPAI) H5N1 variants withGs/GD-like hemagglutinin (HA) and neuraminidase (NA)genes became endemic in domestic poultry in southern China(6, 17, 18, 31, 42, 46). Those variants arose through a series ofreassortment events to incorporate different gene segmentswith different origins (17, 18, 31). The endemicity of HPAIH5N1 viruses in poultry in China and its subsequent spreadthroughout Eurasia and to Africa continue to result in out-breaks in poultry and occasional human infections that hasescalated into a new pandemic threat (5, 31, 42). However, thefundamental question of how the Gs/GD-like virus and itsvariants emerged in southern China has not been completelydetermined.
Previous studies have demonstrated that, in the case of H5and H7 subtypes, low-pathogenic avian influenza (LPAI) virusprecursors may evolve into HPAI viruses once transmitted todomestic poultry (2, 23). Since 1997, several LPAI H5 and H7subtype outbreaks have been observed in Europe (2, 4, 13, 26,
* Corresponding author. Mailing address: State Key Laboratory ofEmerging Infectious Disease, Department of Microbiology, The Uni-versity of Hong Kong, Li Ka Shing Faculty of Medicine Building, 21Sassoon Road, Pokfulam, Hong Kong, SAR, China. Phone: (852)28199830. Fax: (852) 28171958. E-mail for H. Chen: [email protected]. E-mail for Y. Guan: [email protected].
� Published ahead of print on 16 May 2007.
7529

35). Influenza surveillance in migratory birds and domesticpoultry in Italy revealed that the precursor virus responsiblefor LPAI outbreaks in domestic poultry originated from mi-gratory birds (4, 11–13). Moreover, a study in northern Europealso suggested that LPAI subtypes of H5 and H7 viruses resi-dent in migratory duck were introduced to poultry and causedHPAI H5N2 and H7N7 outbreaks (35). Since 2003, outbreaksof LPAI H5N2 virus have also occurred in chickens in Japanand Taiwan. However, phylogenetic analyses revealed that thevirus from Taiwan was most closely related to an LPAI H5N2virus that caused multiple outbreaks in Mexico in the mid-1990s and not to Eurasian lineage viruses (8). This virus doesnot match the natural evolutionary pathway of influenza Avirus, thereby implying another mechanism of virus emer-gence.
Systematic influenza surveillance of domestic poultry inHong Kong and southern China was conducted from 1976 to1980 and 2000 onwards, while surveillance of migratory birdpopulations in southern China has been conducted since 2002(6, 15, 31, 39–42). These surveillance studies accumulated com-prehensive data on the influenza virus gene pool in this regionand provide a unique opportunity to explore the possible evo-lutionary pathways of HPAI Gs/GD-like virus and its variants,i.e., different genotypes. In the present study, LPAI H5 subtypeviruses isolated from surveillance of both domestic poultry andmigratory birds in southern China, together with some H5subtype viruses from Italy, were genetically and antigenicallycharacterized. Our findings revealed the dynamic nature of theinfluenza virus gene pool in Eurasia with repeated transmis-sions between the eastern and western extremities of the con-tinent. Furthermore, our findings suggest that current H5N2and H5N3 influenza viruses in domestic birds of southernChina were directly derived from those viruses resident inmigratory birds, particularly from migratory ducks. Epidemio-logical and genetic findings also suggest that those LPAI H5subtype viruses detected in our surveillance have not becomeestablished in poultry in southern China. Accumulated evi-dence suggests that Gs/GD-like virus might be derived fromviruses resident in migratory birds along the western Pacificflyway. Some Gs/GD-like variants from poultry appeared tohave resulted from further reassortment from viruses in mi-gratory birds. However, there are some H5N1 variants that didnot match natural evolutionary pathways, suggesting other or-igins for those viruses. The current study highlights the neces-sity of continued systematic influenza surveillance in both do-mestic and migratory birds in Eurasia to track the emergenceand reemergence of novel influenza viruses with pandemicpotential.
MATERIALS AND METHODS
Surveillance and virus studied. Influenza surveillance was conducted from1976 to 1980 in live-poultry markets in Hong Kong, China, and from 2000onwards in six different provinces in southern China, as previously described (6,17, 31, 39–42). Migratory birds were sampled at aggregation sites of PoyangLake, Jiangxi Province, China, and Mai Po Marshes, Hong Kong, China, aspreviously described (6). Further sampling of sentry domestic ducks from farmsat Poyang Lake adjacent to these migratory bird aggregation sites was conductedyear round from October 2002 onwards. In addition, influenza surveillance inwild waterfowl, both migratory and resident, was conducted from 1992 to 2002 atOrbetello wetland in central Italy (4, 11–13). Isolation of H5 influenza virusesfrom domestic poultry in Italy in 1997 to 1998 has been previously reported (14).
Influenza isolates were subtyped by the standard hemagglutination inhibition(HI) test using a panel of the World Health Organization reference antisera(32, 49).
Three different representative groups of LPAI H5 subtype viruses were char-acterized and analyzed (Tables 1 and 2). These included 12 viruses isolated fromdomestic poultry in Hong Kong between 1975 and 1980, 19 viruses isolated insouthern China since 2000, and eight viruses isolated from poultry and mallardsin Italy. To provide further information on the influenza virus gene pool in bothmigratory birds and domestic ducks of this region, 11 viruses from seven differentsubtypes isolated from our surveillance were also characterized and analyzed(Table 2).
Antigenic analysis. Viruses characterized in this study were passaged in 9- to11-day-old chicken embryos as previously described (32). The antigenic charac-terization of H5 influenza viruses were conducted by HI assay performed withpanels of monoclonal antibodies (MAbs) against different hemagglutinin (HA)proteins. MAbs against Ck/PA/1370/83, Ck/PA/8125/83, and VNM/1203/04 wereproduced by the Department of Infectious Diseases at St. Jude Children’s Re-search Hospital, while MAbs against Ck/HK/YU22/02 were produced in ourlaboratory. The HI assay was started at a 1:100 dilution.
To visualize similarity between the antigenic reaction patterns of differentviruses, numerical analysis of HI titers was conducted using PRIMER version5.2.9 (PRIMER-E, Plymouth, United Kingdom). Titers of �100 and �12,800 inthe HI assay were converted to 0 and 25,600 in this analysis, respectively. Thedata were square root transformed, and the Bray-Curtis coefficient (3) was usedto construct a similarity matrix. Hierarchical agglomerative clustering withgroup-average linking (43) was conducted, and a dendrogram was produced.Nonmetric multidimensional scaling (28) was also used to produce two- andthree-dimensional ordinations over 100 iterations. The two-dimensional config-uration with lowest overall stress was presented.
Phylogenetic and molecular analyses. To better understand the origins of theGs/GD lineage of H5 HPAI, 18 LPAI H5N2/N3/N8 viruses obtained from 2002to 2005 and 12 LPAI H5N2/N3 viruses isolated during 1976 to 1980 fromsouthern China, together with eight H5N2/N3 viruses from Italian poultry ormallards, were sequenced for each of the eight gene segments (Tables 1 and 2).To provide the background genetic information from a further 10 differentsubtypes, viruses isolated from migratory ducks in the last several years were alsosequenced and phylogenetically analyzed with all available sequence data inpublic databases (Tables 1 and 2).
RNA extraction, cDNA synthesis and PCR were carried out as previouslydescribed (6). Sequence assembly, editing, alignment, and residue analysiswere performed as previously described (6, 20). Phylogenetic analysis usingMRMODELTEST 2.2 (36), PAUP* 4.0 (44), and MRBAYES 3.1 (24) alsowas carried out as previously described (6).
Nucleotide sequence accession numbers. The nucleotide sequences obtainedin this study are available from GenBank under accession numbers EF597247 toEF597498.
RESULTS
Prevalence of low-pathogenic H5 influenza viruses. BetweenJuly 2000 and December 2005, a total of 167,858 samples fromdomestic ducks, geese, and migratory ducks were collected insouthern China and Hong Kong, China. Seventy-two LPAI H5viruses were isolated on 10 sampling occasions during winter(isolation rate of 0.043%) (Table 1). The main body of thoseisolates were from sentry farm ducks or directly from migratorybirds. Only two isolates, Dk/YN/435/02 and Gs/GY/3799/05were detected from retail market birds. In the surveillance inthe 1970s, among 11,798 market samples, 23 LPAI H5 viruseswere isolated on 10 sampling occasions (isolation rate of 0.2%)(39, 40). Most of the viruses were from domestic ducks, withonly one isolated from a goose (Table 1). It must be noted thatno LPAI H5 viruses were recognized in terrestrial poultry inour surveillance. Thus, even though HPAI H5N1 viruses werepersistently detected in different types of poultry, from 2000onwards, LPAI H5 viruses were isolated predominantly frommigratory ducks or sentry duck and barely detected in marketwaterfowl in southern China.
7530 DUAN ET AL. J. VIROL.
Antigenic analysis. Antigenic analysis with MAbs againstrepresentative H5 influenza viruses demonstrated a diversity ofreaction patterns (Table 3). LPAI H5 viruses isolated fromboth domestic and wild waterfowl in southern China since 2003onwards were antigenically conserved and did not differ signif-icantly (greater than fourfold) from each other in the HI assay.
Those viruses obtained from both poultry and mallards in Italyalso had very similar reactivity patterns. Only Ck/Italy/9097/97had a significantly lower titer to MAb 15A3 against VNM/1203/04. It is interesting to note that most LPAI H5 virusesisolated from Italy and southern China from the 1970s to themid-1990s also showed similar reaction patterns (Table 3). Incontrast, HPAI H5N1 representative strains showed high an-tigenic diversity with different reaction patterns, as has beenpreviously observed (6, 42). It was noted that the reactionpattern of Ck/PA/1370/83 was different from those of all Eur-asian viruses tested (Table 3).
In the numerical analysis of HI titers, all LPAI H5 virusesfrom southern China and Italy isolated from the 1970s to themid-1990s clustered together, with the exception of Dk/JX/7820/03 that had lower titers against most MAbs tested (Fig.1). This highly conserved antigenicity likely reflects the evolu-tionary stasis of those viruses in their natural reservoir. It wasnoted that an early Gs/GD-like virus, Gs/HK/437.4/99, andthree HPAI H5N2 viruses (Ck/Italy/312/97, Ty/Italy/392/97,and Ck/Italy/8/98) isolated from outbreaks in terrestrial poul-try in Italy, also grouped with those LPAI H5 viruses fromEurasia. This is likely due to those viruses having been intro-duced to poultry from this gene pool in the recent past. Thisanalysis also demonstrated the high antigenic diversity ofHPAI H5N1 variants currently prevailing in different regions(Fig. 1).
Phylogenetic analysis of the surface genes. Phylogeneticanalysis of the H5 HA gene showed that all viruses separatedinto the American and Eurasian lineages (Fig. 2A). Within the
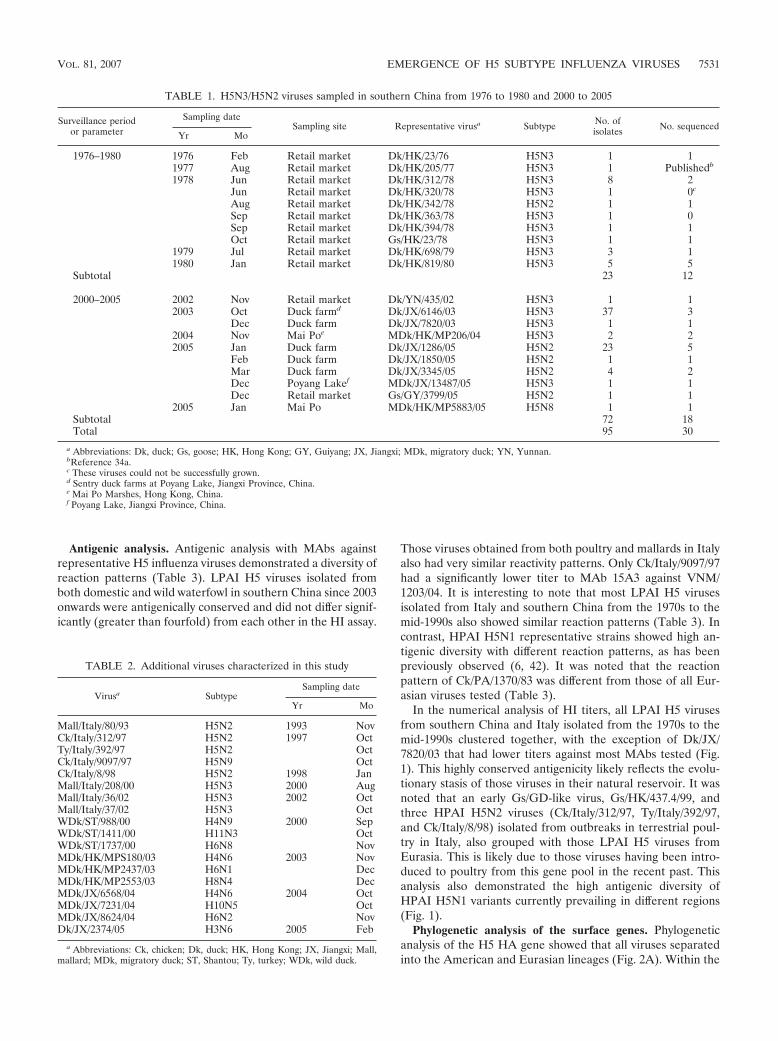
TABLE 1. H5N3/H5N2 viruses sampled in southern China from 1976 to 1980 and 2000 to 2005
Surveillance periodor parameter
Sampling dateSampling site Representative virusa Subtype No. of
isolates No. sequencedYr Mo
1976–1980 1976 Feb Retail market Dk/HK/23/76 H5N3 1 11977 Aug Retail market Dk/HK/205/77 H5N3 1 Publishedb
1978 Jun Retail market Dk/HK/312/78 H5N3 8 2Jun Retail market Dk/HK/320/78 H5N3 1 0c
Aug Retail market Dk/HK/342/78 H5N2 1 1Sep Retail market Dk/HK/363/78 H5N3 1 0Sep Retail market Dk/HK/394/78 H5N3 1 1Oct Retail market Gs/HK/23/78 H5N3 1 1
1979 Jul Retail market Dk/HK/698/79 H5N3 3 11980 Jan Retail market Dk/HK/819/80 H5N3 5 5
Subtotal 23 12
2000–2005 2002 Nov Retail market Dk/YN/435/02 H5N3 1 12003 Oct Duck farmd Dk/JX/6146/03 H5N3 37 3
Dec Duck farm Dk/JX/7820/03 H5N3 1 12004 Nov Mai Poe MDk/HK/MP206/04 H5N3 2 22005 Jan Duck farm Dk/JX/1286/05 H5N2 23 5
Feb Duck farm Dk/JX/1850/05 H5N2 1 1Mar Duck farm Dk/JX/3345/05 H5N2 4 2Dec Poyang Lakef MDk/JX/13487/05 H5N3 1 1Dec Retail market Gs/GY/3799/05 H5N2 1 1
2005 Jan Mai Po MDk/HK/MP5883/05 H5N8 1 1Subtotal 72 18Total 95 30
a Abbreviations: Dk, duck; Gs, goose; HK, Hong Kong; GY, Guiyang; JX, Jiangxi; MDk, migratory duck; YN, Yunnan.bReference 34a.c These viruses could not be successfully grown.d Sentry duck farms at Poyang Lake, Jiangxi Province, China.e Mai Po Marshes, Hong Kong, China.f Poyang Lake, Jiangxi Province, China.
TABLE 2. Additional viruses characterized in this study
Virusa SubtypeSampling date
Yr Mo
Mall/Italy/80/93 H5N2 1993 NovCk/Italy/312/97 H5N2 1997 OctTy/Italy/392/97 H5N2 OctCk/Italy/9097/97 H5N9 OctCk/Italy/8/98 H5N2 1998 JanMall/Italy/208/00 H5N3 2000 AugMall/Italy/36/02 H5N3 2002 OctMall/Italy/37/02 H5N3 OctWDk/ST/988/00 H4N9 2000 SepWDk/ST/1411/00 H11N3 OctWDk/ST/1737/00 H6N8 NovMDk/HK/MPS180/03 H4N6 2003 NovMDk/HK/MP2437/03 H6N1 DecMDk/HK/MP2553/03 H8N4 DecMDk/JX/6568/04 H4N6 2004 OctMDk/JX/7231/04 H10N5 OctMDk/JX/8624/04 H6N2 NovDk/JX/2374/05 H3N6 2005 Feb
a Abbreviations: Ck, chicken; Dk, duck; HK, Hong Kong; JX, Jiangxi; Mall,mallard; MDk, migratory duck; ST, Shantou; Ty, turkey; WDk, wild duck.
VOL. 81, 2007 EMERGENCE OF H5 SUBTYPE INFLUENZA VIRUSES 7531
Eurasian lineage, two major sublineages of “early” and “con-temporary” viruses are apparent. The first sublineage consistspredominantly of older viruses from across the continent, in-cluding those viruses isolated in Hong Kong from 1976 to 1980,along with viruses from Germany, Japan, and the United King-dom. There is only one LPAI H5 virus from Italy (Ck/Italy/9097/97) that falls into this group which was introducedrecently to chicken from this gene pool. Therefore, this sub-lineage of H5 subtype viruses probably represents the earlydomestic Eurasian gene pool of H5 viruses (Fig. 2A).
The second major Eurasian sublineage is composed of threedistinct groups (Fig. 2A). Group 1 contains all but one of therecent viruses from China and Italy characterized in thepresent study, along with other Eurasian isolates, from bothdomestic and migratory birds. However, four Italian virusesisolated during the outbreaks from 1997 to 1998 and one ob-tained from a mallard clustered together within this group,suggesting that after introduction into terrestrial poultry, thoseviruses had undergone significant evolution (Fig. 2A). Thephylogeny of this group, together with that of the “early”sublineage, indicates the continued genetic exchange withinthis influenza virus gene pool between the eastern and westernextremities of the Eurasian continent.
Group 2 consists exclusively of viruses from Western Eu-rope, with the exception of the virus Dk/Chany Lake/9/03 thatwas isolated in Novosibirsk, Russia in Central Asia (Fig. 2A).In contrast, group 3 contained viruses from the western Pacificflyway isolated from domestic ducks and swans, along withHPAI Gs/GD-like H5N1 viruses that have subsequently spread
throughout Eurasia (6, 31, 42). This group therefore seems tomatch one of the major migratory flyways in this region (47).
In the American lineage, viruses from two major outbreaksand those from migratory birds clustered together (Fig. 2A). Ithas been noted that a virus from Japan, Dk/Hokkaido/84/02also joins this lineage, providing an example of gene exchangebetween the American and Eurasian influenza virus genepools. Interestingly, a virus that recently caused outbreaks inpoultry in Japan in 2005 (Ck/Ibaraki/1/2005) also fell in theAmerican lineage and was most closely related to Ck/Hidalgo/28159-232/94 and Ck/Taiwan/1209/03 that caused outbreaks inMexico and Taiwan, respectively (Fig. 2A) (8).
Phylogenetic analysis showed that three N2 genes from re-cent southern China H5N2 isolates (Gs/GY/3799/05, Dk/JX/1286/05, and Dk/JX/3345/05) were closely related to the N2genes of viruses obtained from mallards in Italy and ducks inHokkaido, Japan (data not shown). Analysis also showed thatthe N3 genes from the recent H5N3 viruses isolated in south-ern China are closely related to H7N3 subtype viruses fromdomestic and migratory birds in Italy from 2001 to 2003 (datanot shown). Therefore, both the HA and NA genes of thoseH5N2 and H5N3 viruses recently isolated from domestic ducksor migratory birds in southern China were from the Eurasianlineage and appear to be derived from viruses resident inmigratory birds. Phylogenetic analysis of N1 NA also revealedthat the NA gene of Gs/GD-like virus was most closely relatedto Dk/Hokkaido/55/95 (H1N1) (data not shown). AnotherH5N1 variant (Ck/Hebei/718/01) had an N1 that was almostidentical to African starling/England-Q/983/79 (H7N1) (un-
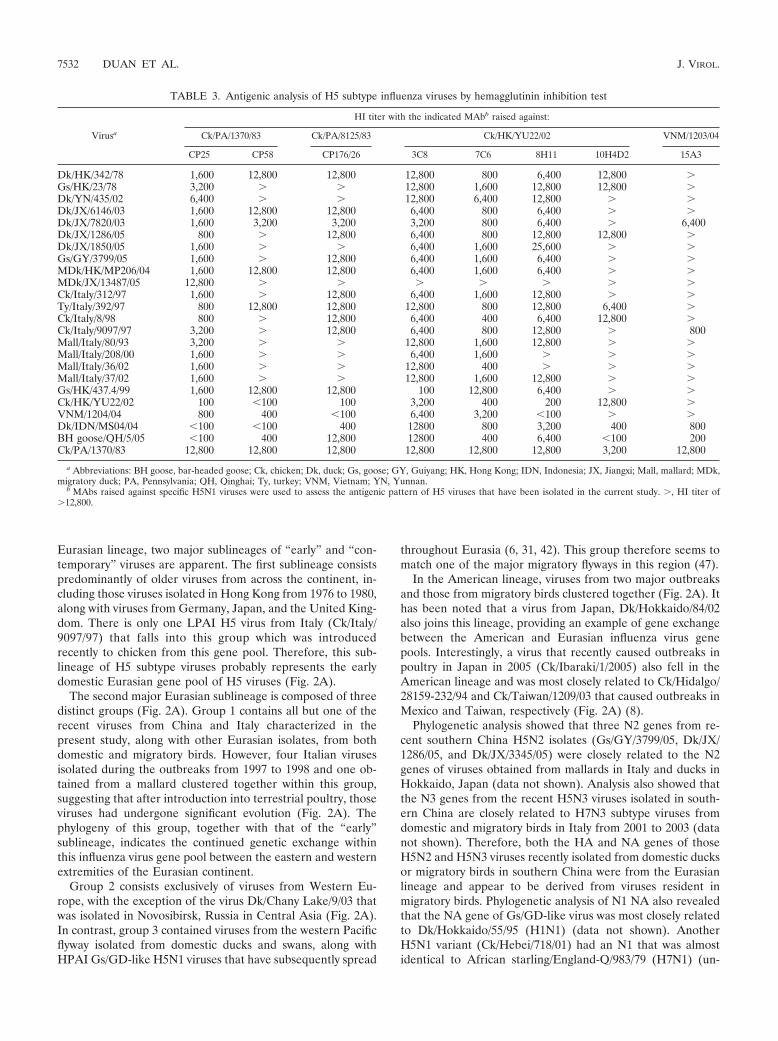
TABLE 3. Antigenic analysis of H5 subtype influenza viruses by hemagglutinin inhibition test
Virusa
HI titer with the indicated MAbb raised against:
Ck/PA/1370/83 Ck/PA/8125/83 Ck/HK/YU22/02 VNM/1203/04
CP25 CP58 CP176/26 3C8 7C6 8H11 10H4D2 15A3
Dk/HK/342/78 1,600 12,800 12,800 12,800 800 6,400 12,800 �Gs/HK/23/78 3,200 � � 12,800 1,600 12,800 12,800 �Dk/YN/435/02 6,400 � � 12,800 6,400 12,800 � �Dk/JX/6146/03 1,600 12,800 12,800 6,400 800 6,400 � �Dk/JX/7820/03 1,600 3,200 3,200 3,200 800 6,400 � 6,400Dk/JX/1286/05 800 � 12,800 6,400 800 12,800 12,800 �Dk/JX/1850/05 1,600 � � 6,400 1,600 25,600 � �Gs/GY/3799/05 1,600 � 12,800 6,400 1,600 6,400 � �MDk/HK/MP206/04 1,600 12,800 12,800 6,400 1,600 6,400 � �MDk/JX/13487/05 12,800 � � � � � � �Ck/Italy/312/97 1,600 � 12,800 6,400 1,600 12,800 � �Ty/Italy/392/97 800 12,800 12,800 12,800 800 12,800 6,400 �Ck/Italy/8/98 800 � 12,800 6,400 400 6,400 12,800 �Ck/Italy/9097/97 3,200 � 12,800 6,400 800 12,800 � 800Mall/Italy/80/93 3,200 � � 12,800 1,600 12,800 � �Mall/Italy/208/00 1,600 � � 6,400 1,600 � � �Mall/Italy/36/02 1,600 � � 12,800 400 � � �Mall/Italy/37/02 1,600 � � 12,800 1,600 12,800 � �Gs/HK/437.4/99 1,600 12,800 12,800 100 12,800 6,400 � �Ck/HK/YU22/02 100 �100 100 3,200 400 200 12,800 �VNM/1204/04 800 400 �100 6,400 3,200 �100 � �Dk/IDN/MS04/04 �100 �100 400 12800 800 3,200 400 800BH goose/QH/5/05 �100 400 12,800 12800 400 6,400 �100 200Ck/PA/1370/83 12,800 12,800 12,800 12,800 12,800 12,800 3,200 12,800
a Abbreviations: BH goose, bar-headed goose; Ck, chicken; Dk, duck; Gs, goose; GY, Guiyang; HK, Hong Kong; IDN, Indonesia; JX, Jiangxi; Mall, mallard; MDk,migratory duck; PA, Pennsylvania; QH, Qinghai; Ty, turkey; VNM, Vietnam; YN, Yunnan.
b MAbs raised against specific H5N1 viruses were used to assess the antigenic pattern of H5 viruses that have been isolated in the current study. �, HI titer of�12,800.
7532 DUAN ET AL. J. VIROL.

published data), which was investigated as a vaccine candidatein China (7, 33, 37).
Phylogenetic analysis of the internal genes. In general, allinternal gene trees were divided into the American and Eur-asian lineages, and the “early” and “contemporary” subline-ages were observed within the Eurasian lineage (Fig. 2B and 3and data not shown). The internal gene phylogenies furtherhighlight the dynamic nature of the influenza virus gene pool inEurasia with viruses from the east and west frequently cluster-
ing together. For the LPAI H5N2 virus Ck/Ibaraki/1/05, allavailable gene segments clustered with the Mexico-like HPAIfrom 1994. It is noteworthy that several Gs/GD-like variantsisolated from northern China from 1997 to 2004 consistentlyclustered within the “early” Eurasian sublineage (Fig. 2B and3 and data not shown). Most of the LPAI H5 viruses fromsouthern China clustered or grouped together with some vi-ruses from European viruses, including those Italian HPAI andLPAI viruses (Fig. 2B and 3 and data not shown).
FIG. 1. Numerical analysis of HI titers (see Table 2) by nonmetric multidimensional ordination in two dimensions (A) and using hierarchicalagglomerative clustering (B). Colors indicate viruses isolated from Italy (blue), southern China (green), and A/goose/Guangdong/1/96-like (red).Abbreviations: IDN, Indonesia; Mall, mallard; MDk, migratory duck; BHG, bar-hooded goose; QH, Qinghai.
VOL. 81, 2007 EMERGENCE OF H5 SUBTYPE INFLUENZA VIRUSES 7533

In the M-gene tree, the Eurasian lineage separated into fourdifferent sublineages, except for those H9N2 influenza virusesfrom terrestrial poultry in China that formed an outgroupsublineage to the rest (Fig. 2B). All recent H5 viruses fromboth southern China and Italy, along with those viruses re-cently isolated from migratory birds, fell into two distinguish-able groups (groups 1 and 2) that appeared to represent anatural evolutionary pathway. It is noteworthy that the Gs/GD-like viruses and its variants, which formed an independentsublineage, were most closely related to Dk/Hokkaido/120/01(H6N2), a virus that consistently has a close phylogenetic re-
lationship to those viruses from migratory ducks, suggesting itmay have originated in migratory birds (Fig. 2B and 3 and datanot shown). Furthermore, while almost all 1970s H5 virusesfrom Hong Kong clustered together with the “early” subline-age, there was a single virus (Dk/HK/23/76) that fell at the baseof those group 2 viruses (Fig. 2B).
In the NP gene tree, all recent LPAI H5 viruses from sentryducks and retail markets in southern China clustered togetherwith viruses isolated directly from migratory ducks at Mai PoMarshes in Hong Kong or Poyang Lake in China (Fig. 3B). All1970s H5 viruses joined the “early” sublineage, while the Ital-
FIG. 2. Phylogenetic relationships of the HA (A) and M (B) genes of representative influenza A viruses isolated in Asia, America, and Europe.Numbers above and below the branch nodes indicate neighbor-joining bootstrap values of �50% and Bayesian posterior probabilities of �95,respectively. Analyses were based on nucleotides 22 to 1032 of the HA gene and 26 to 947 of the M gene. The HA and M gene trees were rootedto A/Japan/305/57 and A/equine/Prague/1/56, respectively. Colors indicate viruses isolated from Italy (blue), southern China (green), andA/goose/Guangdong/1/96 (red). Red branches indicate A/goose/Guangdong/1/96-like highly pathogenic influenza virus and its variants. Bar, 0.1nucleotide substitution per site. Abbreviations: Am, America; Mall, mallard; MDk, migratory duck; Gf, guinea fowl; SCk, silky chicken; BH goose,bar-hooded goose; QH, Qinghai; Qa, quail; WDk, wild duck.
7534 DUAN ET AL. J. VIROL.
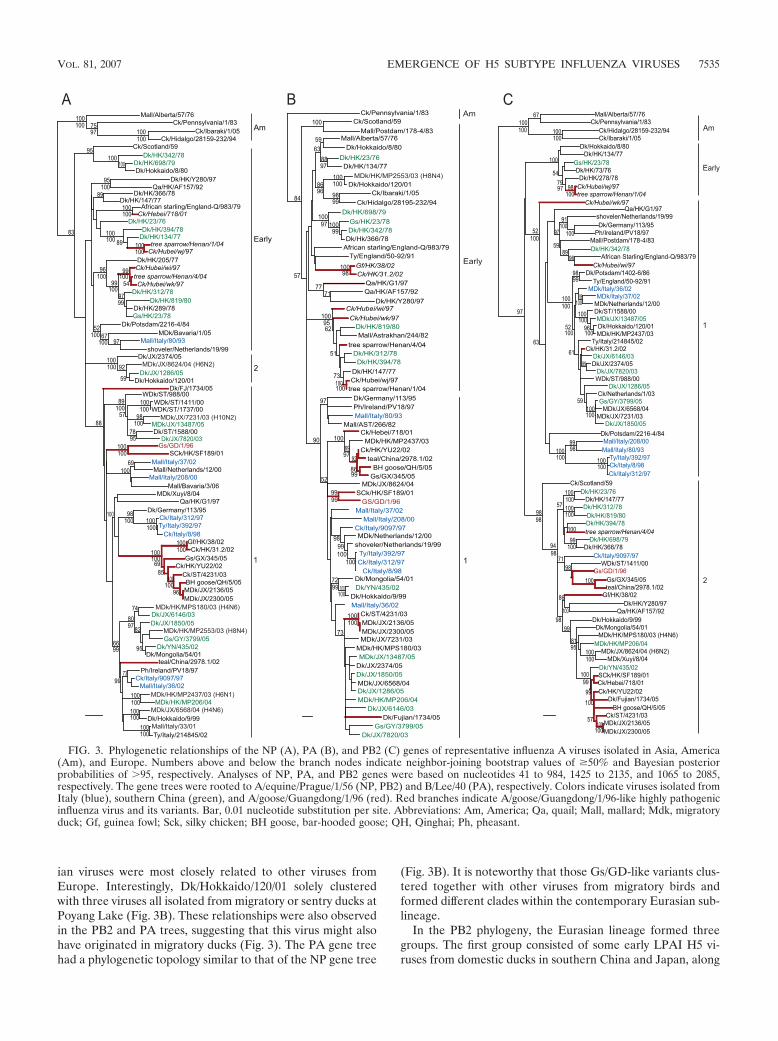
ian viruses were most closely related to other viruses fromEurope. Interestingly, Dk/Hokkaido/120/01 solely clusteredwith three viruses all isolated from migratory or sentry ducks atPoyang Lake (Fig. 3B). These relationships were also observedin the PB2 and PA trees, suggesting that this virus might alsohave originated in migratory ducks (Fig. 3). The PA gene treehad a phylogenetic topology similar to that of the NP gene tree
(Fig. 3B). It is noteworthy that those Gs/GD-like variants clus-tered together with other viruses from migratory birds andformed different clades within the contemporary Eurasian sub-lineage.
In the PB2 phylogeny, the Eurasian lineage formed threegroups. The first group consisted of some early LPAI H5 vi-ruses from domestic ducks in southern China and Japan, along
FIG. 3. Phylogenetic relationships of the NP (A), PA (B), and PB2 (C) genes of representative influenza A viruses isolated in Asia, America(Am), and Europe. Numbers above and below the branch nodes indicate neighbor-joining bootstrap values of �50% and Bayesian posteriorprobabilities of �95, respectively. Analyses of NP, PA, and PB2 genes were based on nucleotides 41 to 984, 1425 to 2135, and 1065 to 2085,respectively. The gene trees were rooted to A/equine/Prague/1/56 (NP, PB2) and B/Lee/40 (PA), respectively. Colors indicate viruses isolated fromItaly (blue), southern China (green), and A/goose/Guangdong/1/96 (red). Red branches indicate A/goose/Guangdong/1/96-like highly pathogenicinfluenza virus and its variants. Bar, 0.01 nucleotide substitution per site. Abbreviations: Am, America; Qa, quail; Mall, mallard; Mdk, migratoryduck; Gf, guinea fowl; Sck, silky chicken; BH goose, bar-hooded goose; QH, Qinghai; Ph, pheasant.
VOL. 81, 2007 EMERGENCE OF H5 SUBTYPE INFLUENZA VIRUSES 7535
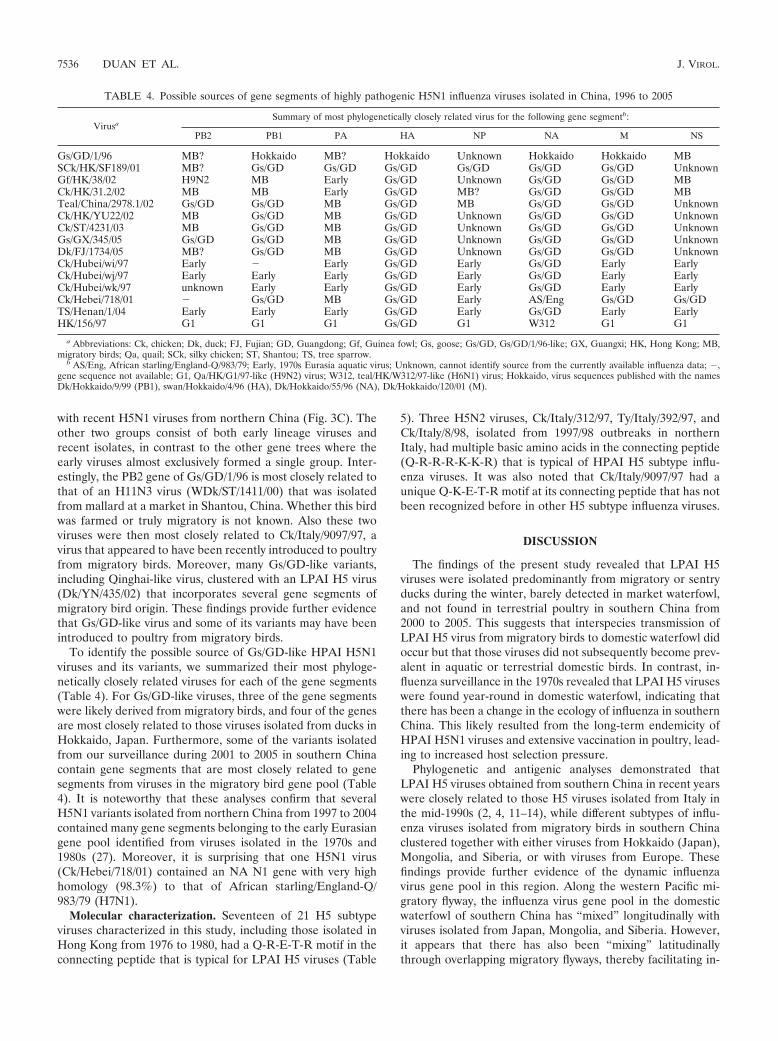
with recent H5N1 viruses from northern China (Fig. 3C). Theother two groups consist of both early lineage viruses andrecent isolates, in contrast to the other gene trees where theearly viruses almost exclusively formed a single group. Inter-estingly, the PB2 gene of Gs/GD/1/96 is most closely related tothat of an H11N3 virus (WDk/ST/1411/00) that was isolatedfrom mallard at a market in Shantou, China. Whether this birdwas farmed or truly migratory is not known. Also these twoviruses were then most closely related to Ck/Italy/9097/97, avirus that appeared to have been recently introduced to poultryfrom migratory birds. Moreover, many Gs/GD-like variants,including Qinghai-like virus, clustered with an LPAI H5 virus(Dk/YN/435/02) that incorporates several gene segments ofmigratory bird origin. These findings provide further evidencethat Gs/GD-like virus and some of its variants may have beenintroduced to poultry from migratory birds.
To identify the possible source of Gs/GD-like HPAI H5N1viruses and its variants, we summarized their most phyloge-netically closely related viruses for each of the gene segments(Table 4). For Gs/GD-like viruses, three of the gene segmentswere likely derived from migratory birds, and four of the genesare most closely related to those viruses isolated from ducks inHokkaido, Japan. Furthermore, some of the variants isolatedfrom our surveillance during 2001 to 2005 in southern Chinacontain gene segments that are most closely related to genesegments from viruses in the migratory bird gene pool (Table4). It is noteworthy that these analyses confirm that severalH5N1 variants isolated from northern China from 1997 to 2004contained many gene segments belonging to the early Eurasiangene pool identified from viruses isolated in the 1970s and1980s (27). Moreover, it is surprising that one H5N1 virus(Ck/Hebei/718/01) contained an NA N1 gene with very highhomology (98.3%) to that of African starling/England-Q/983/79 (H7N1).
Molecular characterization. Seventeen of 21 H5 subtypeviruses characterized in this study, including those isolated inHong Kong from 1976 to 1980, had a Q-R-E-T-R motif in theconnecting peptide that is typical for LPAI H5 viruses (Table
5). Three H5N2 viruses, Ck/Italy/312/97, Ty/Italy/392/97, andCk/Italy/8/98, isolated from 1997/98 outbreaks in northernItaly, had multiple basic amino acids in the connecting peptide(Q-R-R-R-K-K-R) that is typical of HPAI H5 subtype influ-enza viruses. It was also noted that Ck/Italy/9097/97 had aunique Q-K-E-T-R motif at its connecting peptide that has notbeen recognized before in other H5 subtype influenza viruses.
DISCUSSION
The findings of the present study revealed that LPAI H5viruses were isolated predominantly from migratory or sentryducks during the winter, barely detected in market waterfowl,and not found in terrestrial poultry in southern China from2000 to 2005. This suggests that interspecies transmission ofLPAI H5 virus from migratory birds to domestic waterfowl didoccur but that those viruses did not subsequently become prev-alent in aquatic or terrestrial domestic birds. In contrast, in-fluenza surveillance in the 1970s revealed that LPAI H5 viruseswere found year-round in domestic waterfowl, indicating thatthere has been a change in the ecology of influenza in southernChina. This likely resulted from the long-term endemicity ofHPAI H5N1 viruses and extensive vaccination in poultry, lead-ing to increased host selection pressure.
Phylogenetic and antigenic analyses demonstrated thatLPAI H5 viruses obtained from southern China in recent yearswere closely related to those H5 viruses isolated from Italy inthe mid-1990s (2, 4, 11–14), while different subtypes of influ-enza viruses isolated from migratory birds in southern Chinaclustered together with either viruses from Hokkaido (Japan),Mongolia, and Siberia, or with viruses from Europe. Thesefindings provide further evidence of the dynamic influenzavirus gene pool in this region. Along the western Pacific mi-gratory flyway, the influenza virus gene pool in the domesticwaterfowl of southern China has “mixed” longitudinally withviruses isolated from Japan, Mongolia, and Siberia. However,it appears that there has also been “mixing” latitudinallythrough overlapping migratory flyways, thereby facilitating in-
TABLE 4. Possible sources of gene segments of highly pathogenic H5N1 influenza viruses isolated in China, 1996 to 2005
VirusaSummary of most phylogenetically closely related virus for the following gene segmentb:
PB2 PB1 PA HA NP NA M NS
Gs/GD/1/96 MB? Hokkaido MB? Hokkaido Unknown Hokkaido Hokkaido MBSCk/HK/SF189/01 MB? Gs/GD Gs/GD Gs/GD Gs/GD Gs/GD Gs/GD UnknownGf/HK/38/02 H9N2 MB Early Gs/GD Unknown Gs/GD Gs/GD MBCk/HK/31.2/02 MB MB Early Gs/GD MB? Gs/GD Gs/GD MBTeal/China/2978.1/02 Gs/GD Gs/GD MB Gs/GD MB Gs/GD Gs/GD UnknownCk/HK/YU22/02 MB Gs/GD MB Gs/GD Unknown Gs/GD Gs/GD UnknownCk/ST/4231/03 MB Gs/GD MB Gs/GD Unknown Gs/GD Gs/GD UnknownGs/GX/345/05 Gs/GD Gs/GD MB Gs/GD Unknown Gs/GD Gs/GD UnknownDk/FJ/1734/05 MB? Gs/GD MB Gs/GD Unknown Gs/GD Gs/GD UnknownCk/Hubei/wi/97 Early � Early Gs/GD Early Gs/GD Early EarlyCk/Hubei/wj/97 Early Early Early Gs/GD Early Gs/GD Early EarlyCk/Hubei/wk/97 unknown Early Early Gs/GD Early Gs/GD Early EarlyCk/Hebei/718/01 � Gs/GD MB Gs/GD Early AS/Eng Gs/GD Gs/GDTS/Henan/1/04 Early Early Early Gs/GD Early Gs/GD Early EarlyHK/156/97 G1 G1 G1 Gs/GD G1 W312 G1 G1
a Abbreviations: Ck, chicken; Dk, duck; FJ, Fujian; GD, Guangdong; Gf, Guinea fowl; Gs, goose; Gs/GD, Gs/GD/1/96-like; GX, Guangxi; HK, Hong Kong; MB,migratory birds; Qa, quail; SCk, silky chicken; ST, Shantou; TS, tree sparrow.
b AS/Eng, African starling/England-Q/983/79; Early, 1970s Eurasia aquatic virus; Unknown, cannot identify source from the currently available influenza data; �,gene sequence not available; G1, Qa/HK/G1/97-like (H9N2) virus; W312, teal/HK/W312/97-like (H6N1) virus; Hokkaido, virus sequences published with the namesDk/Hokkaido/9/99 (PB1), swan/Hokkaido/4/96 (HA), Dk/Hokkaido/55/96 (NA), Dk/Hokkaido/120/01 (M).
7536 DUAN ET AL. J. VIROL.

teraction between the influenza virus gene pool in domesticwaterfowl in the eastern and western extremities of the Eur-asian continent. This helps to explain the latitudinal spread ofthe Qinghai-like (clade 2.2) H5N1 virus in the last 2 years,while H5N1 outbreaks in Korea and Japan may represent thelongitudinally transmitting pathway (5).
In the present study, the phylogenies of Gs/GD-like H5N1virus genes either clustered with viruses from migratory birdsor with viruses isolated from ducks in Hokkaido, Japan (Table4). As no systematic surveillance was conducted from 1980 to1997 in southern China, we failed to identify the direct pre-cursor of the Gs/GD-like virus which emerged in 1996 (50).Even though it is not clear whether those Hokkaido viruseswere from domestic or migratory ducks, many of them clus-tered directly with the viruses isolated from migratory ducks atPoyang Lake, Jiangxi Province, China, and Mai Po Marshes,Hong Kong, since 2002 (Fig. 2 and 3 and data not shown). Wetherefore speculate that the duck viruses from Hokkaido mayalso be of migratory bird derivation. Thus, the available find-ings suggest that Gs/GD-like H5N1 virus may be derived froman LPAI H5N1 virus in migratory waterfowl along the westernPacific migratory flyway. A precedent for such an introductionand subsequent change from LPAI to HPAI has also beenobserved in Europe (2, 4, 35).
Phylogenetic analysis also showed that those H5N1 variantsisolated in southern China from 2001 to 2005 were mainly
reassortants between Gs/GD-like virus and virus from migra-tory birds. Even though there are still some gene segments forwhich we could not identify the source, current results suggestthat these reassortment events very likely occurred in the field.However, those H5N1 variants from northern China in 1997and afterwards contained almost all internal genes that wereobviously from early gene pool viruses (Table 5). The remain-ing question is to how those old gene segments had beenincorporated into Gs/GD-like virus to generate those variants.
Two possibilities could explain this situation, either thesedata were a result of laboratory contamination or the viruseswere isolated in the field through the use of incompletelyinactivated poultry vaccines that utilized older strains of LPAIH5 virus. For instance, one virus (Ck/Hebei/718/01) containedan N1 gene that was almost identical to that from Africanstarling/England-Q/983/79 (H7N1). This H7N1 virus, alongwith Ty/Wisconsin/1/66 (H9N2) and Ty/England/N28/73 (H5N2),were investigated as vaccine candidates in China (33, 37). AnH9N2 virus (Ck/Heilongjiang/35/00), isolated in a field innorthern China, had almost identical HA and NP genes withTy/Wisconsin/1/66 (30, 34). Furthermore, genetic analyses ofviruses recently isolated from LPAI H5N2 outbreaks in Taiwan(8) and Japan confirmed that those H5N2 viruses were derivedfrom a Mexico-like LPAI H5N2 virus, suggesting that thoseviruses may have been directly derived from incompletely in-activated vaccine (29). Therefore, it seems likely that thoseH5N1 variants containing early gene segments were not gen-erated under natural conditions.
Currently, HPAI H5N1 viruses are endemic in poultry inmany Eurasian countries (6, 31, 42), and as this article wasbeing prepared, a new outbreak wave was occurring acrosseastern and southeastern Asia. Genetic analysis of the recentoutbreak suggested that the outbreaks in Korea and Japanwere caused by BH goose/QH/65/05-like virus. If this is thecase, it is not impossible that those Gs/GD-like variants mayhave successfully become established in the natural reservoirsof influenza viruses, i.e., migratory waterfowls. If so, control ofHPAI H5N1 in the short term has become an impossible mis-sion.
This study demonstrates the dynamic nature of the influenzavirus gene pool in Eurasia with continuing gene exchangesbetween the eastern and western parts of the continent. Ourfindings suggest that the HPAI Gs/GD-like virus were likelyderived directly from viruses resident in migratory birds. Thefailure to identify the source for all gene segments of Gs/GD-like viruses and its variant highlights the need for continuedand extensive surveillance in both migratory and domestic pop-ulations in larger regions. Such surveillance is crucial for ef-fective pandemic influenza preparedness.
ACKNOWLEDGMENTS
This study was supported by the Research Fund for Control ofInfectious Diseases and Research Grants Council (HKU 7512/06 Mand HKU1/05C) of the Hong Kong SAR Government, the Li Ka-ShingFoundation, and the National Institutes of Health (NIAID contractHHSN266200700005C).
We gratefully acknowledge the excellent technical assistance of Li-Juan Zhang and Justin Bahl and the continual and invaluable collab-oration of Marco Carsughi, Antonio Canu, and Luigi Calchetti (WWFItalian Delegation).
TABLE 5. Connecting peptide of H5 subtype viruses characterizedin this study
VirusAmino acid at HA1 positiona
�7 �6 �5 �4 �3 �2 �1
Dk/HK/23/76 Q R E – – T RDk/HK/205/77 Q R E – – T RDk/HK/312/78 Q R E – – T RDk/HK/320/78 Q R E – – T RDk/HK/342/78 Q R E – – T RDk/HK/363/78 Q R E – – T RDk/HK/394/78 Q R E – – T RGs/HK/23/78 Q R E – – T RDk/HK/698/79 Q R E – – T RDk/HK/819/80 Q R E – – T RDk/YN/435/02 Q R E – – T RDk/JX/6146/03 Q R E – – T RDk/JX/7820/03 Q R E – – T RMDk/HK/MP206/04 Q R E – – T RDk/JX/1286/05 Q R E – – T RDk/JX/1850/05 Q R E – – T RDk/JX/3345/05 Q R E – – T RMDk/JX/13487/05 Q R E – – T RGs/GY/3799/05 Q R E – – T RMall/Italy/80/93 Q R E – – T RCk/Italy/312/97 Q R R R K K RTy/Italy/392/97 Q R R R K K RCk/Italy/9097/97 Q K E – – T RCk/Italy/8/98 Q R R R K K RMall/Italy/208/00 Q R E – – T RMall/Italy/36/02 Q R E – – T RMall/Italy/37/02 Q R E – – T RCk/Hidalgo/28159-232/94 Q R E – – T RCk/Ibaraki/1/05 Q R E – – T RCk/Taiwan/1209/03 Q R E – – K R
a Dashes indicate gaps in the connecting peptide multiple sequence alignment.Bolded residues indicate changes from the consensus alignment.
VOL. 81, 2007 EMERGENCE OF H5 SUBTYPE INFLUENZA VIRUSES 7537

REFERENCES
1. Alexander, D. J. 2000. A review of avian influenza in different bird species.Vet. Microbiol. 74:3–13.
2. Banks, J., E. S. Speidel, E. Moore, L. Plowright, A. Piccirillo, I. Capua, P.Cordioli, A. Fioretti, and D. J. Alexander. 2001. Changes in the haemagglu-tinin and the neuraminidase genes prior to the emergence of highly patho-genic H7N1 avian influenza viruses in Italy. Arch. Virol. 146:963–973.
3. Bray, R. J., and J. T. Curtis. 1957. An ordination of the upland forestcommunities of southern Wisconsin. Ecol. Monogr. 27:325–349.
4. Campitelli, L., E. Mogavero, M. A. De Marco, M. Delogu, S. Puzelli, F.Frezza, M. Facchini, C. Chiapponi, E. Foni, P. Cordioli, R. Webby, G.Barigazzi, R. G. Webster, and I. Donatelli. 2004. Interspecies transmission ofan H7N3 influenza virus from wild birds to intensively reared domesticpoultry in Italy. Virology 323:24–36.
5. Chen, H., G. J. D. Smith, S. Y. Zhang, K. Qin, J. Wang, K. S. Li, R. G.Webster, J. S. M. Peiris, and Y. Guan. 2005. H5N1 virus outbreak in migra-tory waterfowl. Nature 436:191–192.
6. Chen, H., G. J. D. Smith, K. S. Li, J. Wang, X. H. Fan, J. M. Rayner, D.Vijaykrishna, J. X. Zhang, L. J. Zhang, C. T. Guo, C. L. Cheung, K. M. Xu,L. Duan, K. Huang, K. Qin, Y. H. Leung, W. L. Wu, H. R. Lu, Y. Chen, N. S.Xia, T. S. Naipospos, K. Y. Yuen, S. S. Hassan, S. Bahri, T. D. Nguyen, R. G.Webster, J. S. Peiris, and Y. Guan. 2006. Establishment of multiple sub-lineages of H5N1 influenza virus in Asia: implications for pandemic control.Proc. Natl. Acad. Sci. USA 103:2845–2850.
7. Chen, J. M., H. C. Ma, J. W. Chen, Y. X. Sun, J. M. Li, and Z. L. Wang. 2007.A preliminary panorama of the diversity of N1 subtype influenza viruses.Virus Genes 35:33–40.
8. Cheng, M. C., M. S. Lee, and C. H. Wang. 2005. Molecular characterizationof H5N2 low pathogenic avian influenza viruses from Taiwan poultry in 2004,p. 30. In Proceedings of the APEC Conference on Avian Influenza. NationalChung Hsing University, Taichung, Taiwan.
9. Claas, E. C., A. D. Osterhaus, R. Vanbeek, J. C. de Jong, G. F. Rimmel-zwaan, D. A. Senne, S. Krauss, K. F. Shortridge, and R. G. Webster. 1998.Human influenza A H5N1 virus related to a highly pathogenic avian influ-enza virus. Lancet 351:472–477.
10. de Jong, J. C., E. C. J. Claas, A. D. M. E. Osterhaus, R. G. Webster, andW. L. Lim. 1997. A pandemic warning? Nature 389:554.
11. de Marco, M. A., L. Campitelli, E. Foni, E. Raffini, G. Barigazzi, M. Delogu,V. Guberti, L. di Trani, M. Tollis, and I. Donatelli. 2004. Influenza surveil-lance in birds in Italian wetlands (1992–1998): is there a host restrictedcirculation of influenza viruses in sympatric ducks and coots? Vet. Microbiol.98:197–208.
12. de Marco, M. A., G. E. Foni, L. Campitelli, E. Raffini, L. di Trani, M. Delogu,V. Guberti, G. Barigazzi, and I. Donatelli. 2003. Circulation of influenzaviruses in wild waterfowl wintering in Italy during the 1993–99 period: evi-dence of virus shedding and seroconversion in wild ducks. Avian Dis.47(Suppl. 3):861–866.
13. de Marco, M. A., E. Foni, L. Campitelli, M. Delogu, E. Raffini, C. Chiapponi,G. Barigazzi, P. Cordioli, L. di Trani, and I. Donatelli. 2005. Influenza viruscirculation in wild aquatic birds in Italy during H5N2 and H7N1 poultryepidemic periods (1998 to 2000) Avian Pathol. 34:480–485.
14. Donatelli, I., L. Campitelli, L. di Trani, S. Puzelli, L. Selli, A. Fioretti, D. J.Alexander, M. Tollis, S. Krauss, and R. G. Webster. 2001. Characterizationof H5N2 influenza viruses from Italian poultry. J. Gen. Virol. 82:623–630.
15. Ellis, T. M., R. B. Bousfield, L. A. Bissett, K. C. Dyrting, G. S. M. Luk, S. T.Tsim, K. Strum-Ramirez, R. G. Webster, Y. Guan, and J. S. M. Peiris. 2004.Investigation of outbreaks of highly pathogenic H5N1 avian influenza inwaterfowl and wild birds in Hong Kong in late 2002. Avian Pathol. 33:492–505.
16. Fouchier, R. A. M., V. Munster, A. Wallensten, T. M. Bestebroer, S. Herfst,D. Smith, G. F. Rimmelzwaan, B. Olsen, and A. D. M. E. Osterhaus. 2005.Characterization of a novel influenza A virus hemagglutinin subtype (H16)obtained from black-headed gulls. J. Virol. 79:2814–2822.
17. Guan, Y., M. Peiris, K. F. Kong, K. C. Dyrting, T. M. Ellis, T. Sit, L. J.Zhang, and K. F. Shortridge. 2002. H5N1 influenza viruses isolated fromgeese in southeastern China: evidence for genetic reassortment and inter-species transmission to ducks. Virology 292:16–23.
18. Guan, Y., J. S. M. Peiris, A. S. Lipatov, T. M. Ellis, K. C. Dyrting, S. Krauss,L. J. Zhang, R. G. Webster, and K. F. Shortridge. 2002. Emergence ofmultiple genotypes of H5N1 avian influenza viruses in Hong Kong SAR.Proc. Natl. Acad. Sci. USA 99:8950–8955.
19. Guan, Y., K. F. Shortridge, S. Krauss, and R. G. Webster. 1999. Molecularcharacterization of H9N2 influenza viruses: were they the donors of the“internal” genes of H5N1 viruses in Hong Kong? Proc. Natl. Acad. Sci. USA16:9363–9367.
20. Hall, T. A. 1999. BioEdit: a user-friendly biological sequence alignmenteditor and analysis program for Windows 95/98/NT. Nucleic Acids Symp.Ser. 41:95–98.
21. Hinshaw, V. S., G. M. Air, V. J. Gibbs, L. Graves, B. Prescott, and D.Karunakaran. 1982. Antigenic and genetic characterization of a novel hemag-glutinin subtype of influenza A viruses from gulls. J. Virol. 42:865–872.
22. Hoffmann, E., J. Stech, I. Leneva, S. Krauss, C. Scholtissek, P. S. Chin, M.Peiris, K. F. Shortridge, and R. G. Webster. 2000. Characterization of theinfluenza A virus gene pool in avian species in southern China: was H6N1 aderivative or a precursor of H5N1? J. Virol. 74:6309–6315.
23. Horimoto, T., E. Rivera, J. Pearson, D. Senne, S. Krauss, Y. Kawaoka, andR. G. Webster. 1995. Origin and molecular changes associated with emer-gence of a highly pathogenic H5N2 influenza virus in Mexico. Virology213:223–230.
24. Huelsenbeck, J. P., and F. R. Ronquist. 2001. MRBAYES: Bayesian infer-ence of phylogenetic trees. Bioinformatics 17:754–755.
25. Kawaoka, Y., S. Yamnikova, T. M. Chambers, D. K. Lvoy, and R. G. Web-ster. 1990. Molecular characterization of a new hemagglutinin, subtype H14,of influenza A virus. Virology 179:759–767.
26. Koopmans, M., B. Wilbrink, M. Conyn, G. Natro, H. van der Nat, H.Vennema, A. Meijer, J. van Steenbergen, R. Fouchier, A. Osterhaus, and A.Bosman. 2004. Transmission of H7N7 avian influenza A virus to humanbeings during a large outbreak in commercial poultry farms in the Nether-lands. Lancet 363:587–593.
27. Kou, Z., F. M. Lie, J. Yu, Z. J. Fan, Z. H. Yin, C. X. Jia, K. J. Xiong, Y. H.Sun, X. W. Zhang, X. M. Wu, X. B. Gao, and T. X. Li. 2005. New genotypeof avian influenza H5N1 viruses isolated from tree sparrows in China. J. Vi-rol. 79:15460–15466.
28. Kruskal, J. B. 1964. Nonmetric multidimensional scaling: a numericalmethod. Psychometrika 29:115–129.
29. Lee, C. W., D. A. Senne, and D. L. Suarez. 2004. Effect of vaccine use in theevolution of Mexican lineage H5N2 avian influenza virus. J. Virol. 78:8372–8381.
30. Li, C., K. Yu, G. Tian, D. Yu, L. Liu, B. Jing, J. Ping, and H. Chen. 2005.Evolution of H9N2 influenza viruses from domestic poultry in MainlandChina. Virology 340:70–83.
31. Li, K. S., Y. Guan, J. Wang, G. J. D. Smith, K. M. Xu, L. Duan, A. P.Rahardjo, P. Puthavathana, C. Buranathai, T. D. Nguyen, A. T. S. Esto-epangestie, A. Chaisingh, P. Auewarakul, H. T. Long, N. T. H. Hanh, R. J.Webby, L. L. M. Poon, H. Chen, K. F. Shortridge, K. Y. Yuen, R. G. Webster,and J. S. M. Pieris. 2004. Genesis of highly pathogenic and potentiallypandemic H5N1 influenza virus in eastern Asia. Nature 430:209–213.
32. Li, K. S., K. M. Xu, J. S. M. Peiris, L. L. M. Poon, K. Z. Yu, K. Y. Yuen, K. F.Shortridge, R. G. Webster, and Y. Guan. 2003. Characterization of H9subtype influenza viruses from the ducks of southern China: a candidate forthe next influenza pandemic in humans? J. Virol. 77:6988–6994.
33. Liu, L. L., X. R. Wang, H. L. Chen, G. B. Tian, K. Sun, and P. Wei. 2006. Thegrowth characteristics of different subtypes of avian influenza virus in theMDCK cell line. Chin. J. Vet. Med. 42:18–20. (In Chinese.)
34. Lu, J. H., X. F. Liu, W. X. Shao, Y. L. Liu, D. P. Wei, and H. Q. Liu. 2005.Phylogenetic analysis of eight genes of H9N2 subtype influenza virus: amainland China strain possessing early isolates’ genes that have been circu-lating. Virus Genes 31:163–169.
34a.Mastrosovich, M., N. Zhou, Y. Kawaoka, and R. Webster. 1999. Thesurface glycoproteins of H5 influenza viruses isolated from humans,chickens, and wild aquatic birds have distinguishable properties. J. Virol.73:1146–1155.
35. Munster, V. J., A. Wallensten, C. Baas, G. F. Rimmelzwaan, M. Schutten, B.Olsen, A. D. M. E. Osterhaus, and R. A. M. Fouchier. 2005. Mallards and highlypathogenic avian influenza ancestral viruses, northern Europe. Emerg. Infect.Dis. 11:1545–1551.
36. Nylander, J. A. A. 2004. MRMODELTEST 2. Evolutionary Biology Centre,Uppsala University, Uppsala, Sweden.
37. Qiao, C., G. Tian, Y. Jiang, Y. Li, J. Shi, K. Yu, and H. Chen. 2006. Vaccinesdeveloped for H5 highly pathogenic avian influenza in China. Ann. N. Y.Acad. Sci. 1081:182–192.
38. Rohm, C., N. N. Zhou, J. Suss, J. Mackenzie, and R. G. Webster. 1996.Characterization of a novel influenza hemagglutinin, H15: criteria for deter-mination of influenza A subtypes. Virology 217:508–516.
39. Shortridge, K. F. 1982. Avian influenza A viruses of southern China andHong Kong: ecological aspects and implications for man. Bull. W. H. O.60:129–135.
40. Shortridge, K. F. 1992. Pandemic influenza—a zoonosis? Semin. Respir.Infect. 7:11–25.
41. Shortridge, K. F., N. N. Zhou, Y. Guan, P. Gao, T. Ito, Y. Kawaoka, S.Kodihallli, S. Krauss, D. Markwell, K. Gopal Murti, M. Norwood, D.Senne, L. Sims, A. Takada, and R. G. Webster. 1998. Characterization ofavian H5N1 influenza viruses from poultry in Hong Kong. Virology 252:331–342.
42. Smith, G. J. D., X. H. Fan, J. Wang, K. S. Li, K. Qin, J. X. Zhang, D.Vijaykrishna, C. L. Cheung, K. Huang, J. M. Rayner, J. S. M. Peiris, H.Chen, R. G. Webster, and Y. Guan. 2006. Emergence and predominance ofan H5N1 influenza variant in China. Proc. Natl. Acad. Sci. USA 103:16936–16941.
43. Sokal, R. R., and C. D. Michener. 1958. A statistical method for evaluatingsystematic relationships. Univ. Kans. Sci. Bull. 38:1409–1438.
44. Swofford, D. L. 2001. PAUP*: phylogenetic analysis using parsimony (andother methods) 4.0 Beta. Sinauer Associates, Sunderland, MA.
7538 DUAN ET AL. J. VIROL.

45. Webster, R. G., W. J. Bean, O. T. Gorman, T. M. Chambers, and Y.Kawaoka. 1992. Evolution and ecology of influenza A viruses. Microbiol.Rev. 56:152–179.
46. Webster, R. G., Y. Guan, M. Peiris, D. Walker, S. Krauss, N. N. Zhou, E. A.Govorkova, T. M. Ellis, K. C. Dyrting, T. Sit, D. R. Perez, and K. F.Shortridge. 2002. Characterization of H5N1 influenza viruses that continueto circulate in geese in southeastern China. J. Virol. 76:118–126.
47. Webster, R. G., J. S. M. Peiris, H. Chen, and Y. Guan. 2006. H5N1 outbreaksand enzootic influenza. Emerg. Infect. Dis. 12:3–8.
48. Webster, R. G., M. Yakhno, V. S. Hinshaw, W. J. Bean, and K. G. Murti.
1978. Intestinal influenza: replication and characterization of influenza vi-ruses in ducks. Virology 84:268–278.
49. Xu, K. M., K. S. Li, G. J. D. Smith, J. W. Li, H. Tai, J. X. Zhang, R. G.Webster, J. S. M. Peiris, H. Chen, and Y. Guan. 2007. Evolution and mo-lecular epidemiology of H9N2 influenza A viruses from quail in southernChina, 2000 to 2005. J. Virol. 81:2635–2645.
50. Xu, X., K. Subbarao, N. J. Cox, and Y. Guo. 1999. Genetic characterizationof the pathogenic influenza A/goose/Guangdong/1/96 (H5N1) virus: similar-ity of its hemagglutinin gene to those of H5N1 viruses from the 1997 out-breaks in Hong Kong. Virology 261:15–19.
VOL. 81, 2007 EMERGENCE OF H5 SUBTYPE INFLUENZA VIRUSES 7539





























