Embed Size (px)
Citation preview

Chinese Journal of Oceanology and LimnologyVol. 32 No. 2, P. 377-389, 2014http://dx.doi.org/10.1007/s00343-014-3130-6
Characterization of ribulose-1, 5-bisphosphate carboxylase/oxygenase and transcriptional analysis of its related genes in Saccharina japonica (Laminariales, Phaeophyta)*
SHAO Zhanru (邵展茹) 1, 3 , LIU Fuli (刘福利) 2 , LI Qiuying (李秋莹) 1, 3 , YAO Jianting (姚建亭) 1 , DUAN Delin (段德麟) 1, ** 1 Institute of Oceanology, Chinese Academy of Sciences, Qingdao 266071, China 2 Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China 3 University of Chinese Academy of Sciences, Beijing 100049, China
Received Apr. 3, 2013; accepted in principle Jun. 20, 2013; accepted for publication Jun. 26, 2013 © Chinese Society for Oceanology and Limnology, Science Press, and Springer-Verlag Berlin Heidelberg 2014
Abstract Saccharina japonica is a common macroalga in sublittoral communities of cold seawater environments, and consequently may have highly effi cient ribulose-1, 5-bisphosphate carboxylase/oxygenase (Rubisco) activity for carbon assimilation. In our study, we cloned the full-length Rubisco gene from S . japonica ( SJ - rbc ). It contained an open reading frame for a large subunit gene ( SJ - rbcL ) of 1 467 bp, a small subunit gene ( SJ - rbcS ) of 420 bp, and a SJ - rbcL /S intergenic spacer of 269 bp. The deduced peptides of SJ-rbcL and SJ-rbcS were 488 and 139 amino acids with theoretical molecular weights and isoelectric points of 53.97 kDa, 5.81 and 15.84 kDa, 4.71, respectively. After induction with 1 mmol/L isopropyl- β -D-thiogalactopyranoside for 5 h and purifi cation by Ni 2+ affi nity chromatography, electrophoresis and western blot detection demonstrated successful expression of the 55 kDa SJ-rbcL protein. Real-time quantitative PCR showed that the mRNA levels of SJ - rbcL in gametophytes increased when transferred into normal growth conditions and exhibited diurnal variations: increased expression during the day but suppressed expression at night. This observation implied that Rubisco played a role in normal gametophytic growth and development. In juvenile sporophytes, mRNA levels of SJ - rbcL , carbonic anhydrase, Calvin-Benson-Bassham cycle-related enzyme, and chloroplast light-harvesting protein were remarkably increased under continuous light irradiance. Similarly, expression of these genes was up-regulated under blue light irradiance at 350 μmol/(m 2 ∙s). Our results indicate that long-term white light and short-term blue light irradiance enhances juvenile sporophytic growth by synergistic effects of various photosynthetic elements.
Keyword : Saccharina japonica ; Rubisco; photosynthetic genes; transcription; protein expression; carboxylase activity
1 INTRODUCTION
Nearly 50% of all global CO 2 is assimilated by aquatic primary producers, and algae play a predominant role in global carbon metabolism (Moroney and Ynalvez, 2009). Ribulose-1, 5-bisphosphate carboxylase/oxygenase (Rubisco) is a crucial enzyme in CO 2 assimilation. This bifunctional enzyme catalyzes the initial steps of photosynthetic carbon reduction and photorespiratory carbon oxidation cycles by combining CO 2 and O 2 with ribulose-1, 5-bisphosphate (RuBP) (Berry et al., 1978; Mizohata et al., 2002). RuBP carboxylation determines
the net photosynthetic effi ciency of photoautotrophs (Jensen and Bahr, 1977). Nevertheless, in aquatic environments, the enzymatic effi ciency of Rubisco is limited by the low concentration and diffusion
* Supported by the Agriculture Science & Technology Achievements Transformation Fund (No. 2011GB24910005), the Modern Agricultural-Industry Technology Research Project (No. 200903030), the National High Technology Research and Development Program of China (863 Program) (No. 2012AA10A406), the Shandong Agriculture Breeding Engineering Biological Resources Innovation of Research Project, and the National “Twelfth Five-Year” Plan for Science & Technology Support (No. 2013BAB01B01)SHAO Zhanru and LIU Fuli contributed equally to this work. ** Corresponding author: [email protected]

378 CHIN. J. OCEANOL. LIMNOL., 32(2), 2014 Vol.32
coeffi cients of CO 2 , which is compounded by its low affi nity for CO 2 (Reinfelder, 2011).
Previously, many reports have concentrated on the cloning of full-length sequences of Rubisco genes ( rbc ) from microalgae such as Chlamydomonas reinhardtii (Dron et al., 1982), Euglena gracilis (Ginrich and Hallick, 1985), and Chlorella pyrenoidosa . For brown macroalgae, most studies have focused on taxonomy with partial large subunit ( rbcL ) and/or small subunit ( rbcS ) genes (Phillips et al., 2008; Lee et al., 2011; Win et al., 2011). So far, except for the data available in GenBank (http://www.ncbi.nlm.nih.gov/genbank/), there are few reports on isolation and characterization of macroalgal Rubisco (Kai et al., 2002; Ying et al., 2011). It is therefore necessary to characterize rbc expression in algae, which will facilitate characterization of Rubisco in photosynthesis during algal growth and development.
Saccharina japonica is one of the most important commercial seaweeds worldwide with gametophyte and sporophyte generations in its life cycle (Tseng and Wu, 1962). Because this kelp can grow up to 2–6 m in length and 35–50 cm in width, it shows high plasticity for adaption to low temperatures and weak irradiance (Bartsch et al., 2008). This observation implies that it may possess a complicated carbon metabolism, and the sporophytes may have a high photosynthetic effi ciency. Many elements are believed to be involved in different photosynthetic processes of algae. Expression of light-harvesting fucoxanthin-chlorophyll c antenna protein (Lhcf5), which participates in the light-dependent reaction, is enhanced by light in S . japonica gametophytes (Zou et al., 2009). Carbonic anhydrase (CA) has been reported to mediate carbon-concentrating mechanisms for CO 2 saturation of Rubisco in marine plants (Giordano et al., 2005; Reinfelder, 2011). Calvin-Benson-Bassham cycle-related enzyme (CbbX) has been regarded as a red-like Rubisco activase that activates Rubisco in Chromophyta (Mueller-Cajar et al., 2011). Despite these enzymatic studies, no combined effects of the aforementioned enzymes on photosynthesis have been addressed in higher plants or algae. Because photosynthesis is a light-dependent process, it is important to concentrate on variations of photosynthetic gene expression under different light irradiance (Miller and Connell, 2012). In addition, blue light (BL) is thought to be the most effective light for kelp growth (Dring, 1981; Kirk, 1994; Anderson et al., 1995; Klenell et al., 2002; Walters, 2005; Matsuda et al., 2008). Many studies have shown
that BL stimulates Saccharina / Laminaria gametophytic growth and sporophytic reproduction (Lüning and Dring, 1975; Mizuta et al., 2007; Wang et al., 2010). The effective quantum yield of PSII and relative electron transport rate indicates a linear relationship between BL and stimulation of photosynthesis in algae (MacIntyre et al., 2000; Wang et al., 2010). Nevertheless, the mechanisms underlying BL in photosynthesis are still not clearly understood.
The objective of this study was to characterize Rubisco of S . japonica through gene structure analysis, endogenous enzymatic assays, and prokaryotic expression of S . japonica ( SJ )- rbcL for subsequent purifi cation. Using quantitative PCR detection of the expression of rbc and other photosynthesis-related genes under various light conditions in kelp gametophytes and sporophytes, we explored the optimal environmental conditions for their growth and development. We also investigated the role of Rubisco and the systematic mechanism that is potentially responsible for photosynthesis in S . japonica .
2 MATERIAL AND METHOD
2.1 Preculture and treatments of S . japonica
Kelp gametophytes were maintained at 8°C under weak irradiance. About 5 g (wet weight) of gametophytes were homogenized into small pieces (ca 200 μm in length) for preculture. Juvenile sporophytes (about 2–3 cm in width and 15–25 cm in length) were collected at Rongcheng (Shandong Province, China) in December 2011. The collected samples were rinsed three times with sterilized fi ltered seawater and precultured in sterilized seawater supplemented with 11.76 μmol/L NaNO 3 and 7.35 μmol/L KH 2 PO 4 at 15°C under a 12:12 h light/dark photoperiod at a photon fl ux density (PFD) of 45 μmol/(m 2 ∙s) for 48 h.
Gametophytes were exposed to various temperatures and cultivation conditions. The precultured samples were transferred for culture at 5, 10, 15, 20, or 25°C for 2 h, and then about 0.1 g (wet weight) of the samples was collected from each treatment group. In parallel, gametophytes were continuously cultured and sampled (0.1 g) every 24 h for 9 days under a 12 h:12 h light/dark photoperiod. After 9 days of culture, gametophytes were sampled every 3 h for 48 h.
After sampling for 48 h, the juvenile sporophytes were cultured at 15°C under darkness for 12 h. Then, the juvenile sporophytes were exposed to continuous

379No.2 SHAO et al.: Rubisco and its related genes in S . japonica
white light at a PFD of 45 μmol/(m 2 ∙s) for 12 h, followed by darkness for 12 h and 24 h. In addition, sporophytes were cultured in darkness and BL with BL-emitting diodes at about 350 μmol/(m 2 ∙s) for 2 h.
After treatments, all cultures were harvested, rinsed three times with 0.1% DEPC sterile seawater, frozen in liquid nitrogen immediately, and then stored at -80°C for nucleic acid and crude protein extractions.
2.2 Extraction of DNA and RNA, and cDNA synthesis
Extraction of genomic DNA from samples was conducted using a Plant Genomic DNA kit (Tiangen, Beijing, China) according to the manufacturer’s protocol. Qualifi cation and quantifi cation of extracted DNA were performed by 1% agarose gel electrophoresis and a Nanodrop 2000 Spectrophotometer (Thermo Scientifi c, USA).
Total RNA was extracted according to the cetyltrimethylammonium bromide method (Yao et al., 2009). The extracted RNA was dissolved in RNase-free water followed by removal of DNA with a DNase I kit (Fermentas, Burlington, VT, USA). Measurement of total RNA concentration was performed on the Nanodrop 2000 Spectrophotometer and verifi ed by 1% agarose gel electrophoresis.
First-strand cDNA was prepared using a PrimeScript II cDNA synthesis kit (TaKaRa, Tokyo, Japan) according to the manufacturer’s instructions, and then stored at -20°C for subsequent S . japonica ( SJ )- rbc isolation.
2.3 Cloning of SJ - rbc
According to the upstream sequence of rbcL of Ectocarpus siliculosus (Accession: FP102343) and a partial SJ - rbc sequence (Accession: DQ143075), the forward primer SJ-rbcLf and reverse primer SJ-rbcLr were designed, respectively, for PCR amplifi cation of SJ - rbcL from genomic DNA (Table 1). Similarly, the forward primer SJ-rbcSf and reverse primer SJ-rbcSr were designed based on the partial SJ - rbc sequence and downstream sequence of rbcS from E . siliculosus , respectively, to clone SJ - rbcS from genomic DNA (Table 1).
PCR was performed in a 20 μL reaction mixture containing 1 μL genomic DNA (about 50 ng), 10 μL of 2×PCR buffer, 5.15 μL ddH 2 O, 0.25 μL Takara Ex Taq HS (5 U/μL), 1.6 μL dNTP (2.5 mmol/L each), and 1 μL of each primer (10 μmol/L). PCR was carried out at 94°C for 3 min, followed by 35 cycles of 94°C for 30 s, 55°C for 30 s, 72°C for 2 min, and a fi nal extension at 72°C for 10 min. PCR products were subjected to 1% agarose electrophoresis, purifi ed by an agarose gel DNA fragment recovery kit (TaKaRa), subcloned into the pMD-18T vector (TaKaRa), and fi nally sequenced using M13-47 and RV-M primers (Sangon, Shanghai, China).
To obtain the full-length SJ - rbc sequence, eSJ-rbcLf and eSJ-rbcSr primers (Table 1; Fig.1) were designed according to the genomic DNA sequence of
eSJ-rbcSr
eSJ-rbcSfeSJ-rbcLf
-35 element -10 element
TATAAATTTGGTA
qSJ-rbcLf
qSJ-rbcLR ESJ-rbcLR
SJ-rbcS
Fig.1 Gene structure of SJ - rbc The putative promoter elements at –10 and –35 positions and the primers are indicated
Table 1 Primer sequences applied in the study
Primers Sequences
SJ-rbcLf 5'-CGAACTGGGGATAGAGGATTTG-3'
SJ-rbcLr 5'-ACGGTTTGCTGTCGCACCGGATT-3'
SJ-rbcSf 5'-AGAAGGTTTCGGTCTTTATCCA-3'
SJ-rbcSr 5'-CAAAAGGATGGGCTGTTAGTGT-3'
qSJ-rbcLf 5'-AACTGGTGGTCTTGATTTCC-3'
qSJ-rbcLr 5'-AGCCATAGTTTGAATAGCAG-3'
eSJ-rbcSf 5'-ATGAGAGTTACACAAGGATGTT-3'
eSJ-rbcSr 5'-ATAACGGTCTCCAGCAGGT-3'
eSJ-rbcLf 5'-ATGCCTGAGAATGTACAGGAA-3'
eSJ-rbcLr 5'-TTTGCTTTCAGTTGCAACTCC-3'
SSR-F 5'-TCGGACGGTTTTGTGGTGA-3'
SSR-R 5'-GCTGCCTTCCTTGGATGTG-3'
qSJ-CAf 5'-GGTCTTCACTACCCGTATCTAC-3'
qSJ-CA-r 5'-CGATCCACTTCACGCCCTCAG-3'
qSJ-CbbXf 5'-CGACTTCCCCAACTACCTGA-3'
qSJ-CbbXr 5'-CTTGCGTTGGAGAAGAAAGGG-3'
qSJ-Lhcf5f 5'-GCCACTTGACCCAGCAGAACAC-3'
qSJ-Lhcf5r 5'-GGATCTTGGACAGAGCAGCCACA-3'

380 CHIN. J. OCEANOL. LIMNOL., 32(2), 2014 Vol.32
SJ - rbc . PCR was performed according to the procedures described above.
2.4 DNA analysis and deduced amino acid sequences of SJ - rbc
The BLAST program was used to search for SJ - rbc in data available from the National Center for Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.gov/blast). The obtained DNA sequences were translated into the amino acid sequences for analysis of theoretical physical and chemical parameters such as molecular weight and isoelectric point by open reading frame (ORF) Finder and ProtParam (Gasteiger et al., 2005; Sayers et al., 2012). The signal peptide was predicted by the SignalP 4.0 Server. For transmembrane topological structure analysis, we used TMHMM Server version 2.0 (Krogh et al., 2001; Petersen et al., 2011). SWISS-MODEL was used to construct the putative spatial structure of SJ-rbc protein by homology modeling in automated mode (Guex and Peitsch, 1997; Schwede et al., 2003; Arnold et al., 2006).
For phylogenetic analysis, multiple sequence alignments were performed with CLUSTAL X (Thompson et al., 1997), and then analyzed using the neighbor-joining method and MEGA 5.05 software (Tamura et al., 2011). A bootstrap consensus tree was inferred from 1 000 replicates (Felsenstein, 1985).
2.5 Transcriptional analysis of SJ - rbc
Based on the SJ - rbc sequence, qSJ-rbcLf and qSJ-rbcLr primers (Table 1) were designed to amplify a 262-bp amplicon for real-time quantitative PCR detection. SSR-F and SSR-R primers were used to amplify small subunit rRNA as an internal control. Primers (Table 1) were designed for CA (Accession: AEF33616), chloroplast light-harvesting protein (Lhcf5; Accession: ACE80197), and CbbX (Accession: ADG62366) for quantitative PCR analysis (Table 1).
Quantitative PCR was performed with SYBR Premix Ex Taq ™ II (TaKaRa) on a TP800 Thermal Cycler Dice™ (TaKaRa). Each reaction volume of 25 μL contained 12.5 μL Premix, 1 μL of each primer (10 μmol/L), 2 μL cDNA, and 8.5 μL RNase-free water. PCR conditions were 95°C for 10 s, followed by 40 cycles of 95°C for 5 s and 58°C for 30 s. The specifi city of primers was checked by the relevant dissociation curves, and the baseline was set automatically by the software to maintain consistency. Triplicate repeats were performed independently, and
relative quantitative values were calculated by the 2 -ΔΔ Ct method (Schmittgen et al., 2000). The data were analyzed with the Student’s t -test using SPSS 13.0. P <0.05 was set as the level of statistical signifi cance.
2.6 Determination of crude Rubisco carboxylase activity
Total protein was extracted from S . japonica according to the method described by Rousvoal et al. (2011). Juvenile sporophytes (about 0.1 g wet weight) were ground in liquid nitrogen and then homogenized with 1 mL lysis buffer (25 mmol/L Tris-HCl, pH 8.0) containing 15 mmol/L EGTA, 15 mmol/L MgCl 2 , 2 mmol/L DTT, 0.5% PVP, and protease inhibitors (CWBio, Beijing, China). The sample was then stirred through a tissue grinder and subjected to intermittent sonication (Scieniz, Ningbo, China) for 2 min. After centrifugation at 15 000× g for 20 min, the supernatant was transferred to a new pre-cooled tube. The extracted total protein was quantifi ed using an Easy Protein Quantitative Kit (TransGen, Beijing, China) according to the Bradford method (Bradford, 1976).
To determine Rubisco carboxylase activity, we applied a Plant Rubisco Carboxylation Activity Detection Kit (GenMed Scientifi cs, China) using the methods of Lan and Mott (1991). The enzymatic assay was conducted at 25°C for 5 min in a Microplate Reader (Bio-Tek, Winooski, VT, USA). About 10 μg of crude proteins was added to the 250 μL reaction mixture, and then the reaction was initiated by addition of the substrate, RuBP. Boiled crude algal proteins were used as a negative control.
2.7 Prokaryotic expression of SJ - rbc and purifi cation of SJ-rbcL
After amplifi cation with eSJ-rbcLf/eSJ-rbcLr and eSJ-rbcSf/eSJ-rbcSr primers (Table 1; Fig.1), the desired ORF was subcloned into pEASY -E2 (TransGen Biotech, Beijing, China) to construct recombinant expression vectors pEASY-E2/SJ-rbcL and pEASY-E2/SJ-rbcS, respectively. The E . coli BL21 strain was transformed with the plasmids and then spread onto Lysogeny broth (LB) agar plates containing ampicillin (100 μg/mL) to select positive clones.
After culture in LB liquid medium overnight at 37°C with shaking at 220 r/min on an orbital shaker, 2 mL of the transformed BL21 cells was transferred into 200 mL LB medium for incubation until the OD 600 reached 0.5–0.7 (Biophotometer, Eppendorf, Germany). Then the expression of SJ - rbcL and SJ -

381No.2 SHAO et al.: Rubisco and its related genes in S . japonica
rbcS was induced by addition of isopropyl-β-D-thiogalactopyranoside (IPTG) at various fi nal concentrations (0, 0.5, 1.0, 1.5, and 2.0 mmol/L) to determine the optimal induction dose. Cultures were then collected, centrifuged, and lysed in sodium dodecyl-sulfate (SDS) loading buffer. After boiling for 10 min, the lysates were analyzed by 12% SDS-polyacrylamide gel electrophoresis (PAGE) (ATTO, Tokyo, Japan).
After induction with 1 mmol/L IPTG for 5 h, the cultures (200 mL) were subjected to purifi cation with a His-Purifi cation Kit (Weishibohui, Beijing, China). Gradient elution was applied with 50 and 200 mmol/L imidazole solutions, and the collected samples were analyzed by 12% SDS-PAGE. All purifi cation steps were carried out at 4°C.
2.8 Western blot analysis
Fractions containing recombinant enzyme were pooled, and the mixture was concentrated with Amicon Ultra-15 Centrifugal Filter devices (Millipore, Billerica, MA, USA) to about 1 mg/mL. After SDS-PAGE, the proteins were electrophoretically transferred to a polyvinylidene difl uoride membrane (0.45 μm) at 30 V for 90 min on an AE-6675 HorizBlot system (ATTO, Tokyo, Japan). The membrane was blocked in TBST buffer (150 mmol/L NaCl, 20 mmol/L Tris-HCl, pH 8.0, and 0.05% Tween-20)
containing 5% non-fat milk powder for 2 h at room temperature. A 1:500 dilution of primary rabbit anti-His antibody (Top-Science, Beijing, China) was applied to the membrane, followed by gentle agitation for 1 h at room temperature. After rinsing with TBST buffer, the membrane was exposed to a 1:2 000 dilution of horseradish peroxidase-conjugated goat-anti-rabbit IgG (Tiandz, Beijing, China) for 1 h at room temperature. A DAB-sensitive chromogenic reaction (Tiandz, Beijing, China) was applied to develop the blot.
3 RESULT 3.1 Structural characterization of the SJ - rbc gene
The SJ - rbc gene (2 156 bp) contained a SJ - rbcL ORF (1 467 bp), SJ - rbcL / S intergenic spacer (269 bp), SJ - rbcS ORF (420 bp) (Fig.1), and no introns. The SJ - rbc sequence was deposited in the NCBI database (Accession: HM798587). SJ - rbcL and SJ - rbcS were cotranscribed into the same mRNA, indicating that these genes shared a common operon. In addition, there was a TATAAT box at the -10 position and a TTGCTA box at the -35 position located in the upstream of the promoter (Fig.1).
Deduced sequences of the Rubisco large subunit (LSU) and small subunit (SSU) encoded by SJ - rbcL and SJ - rbcS were 488 and 139 amino acids with a theoretical molecular weights and isoelectric points of 53.97 kDa, 5.81 and 15.84 kDa, 4.71, respectively. Protein prediction indicated that there was no signal peptide, but there was a transmembrane peptide (23 amino acids) in SJ - rbcL . Based on C hlamydomonas reinhardtii and Alcaligenes eutrophuto , we predicted the tertiary structures of SJ-rbcL and SJ-rbcS, respectively (Fig.2). The predicted spatial structure of SJ-rbcL was folded into two separate domains and one α/β-barrel in the C-terminal, which contained critical serial amino acids (Fig.2a). There were six presumed residues (Lys 179, Arg 298, His 330, Lys 337, Gly 406, and Gly 407) for catalysis, two adjacent acidic residues (Asp 207 and Glu 208) for Mg 2+ coordination, and one residue (Lys 205) for CO 2
fi xation (Fig.2b). SJ-rbcS was characterized by an extended C-terminal that folded into a two-stranded anti-parallel β-sheet (Fig.2c).
3.2 Phylogenetic analysis of SJ - rbc
A phylogenetic tree was constructed using the neighbor-joining method based on the deduced full-length amino acid sequences of Rubisco in 28 species
a b
c
a b
Gly407
Gly406Lys179 His330
Arg298Glu208
Asp207Lys205
Lys337
c
Fig.2 Predicted spatial structure of SJ-rbcL (a and b) and SJ-rbcS (c) a. Global tertiary structure of SJ-rbcL. The dotted rectangle indicates the α/β-barrel in the C-terminal; b. Magnifi ed view of the dotted rectangle in (a) containing the barrel and critical serial amino acids Asp 207 and Glu 208 (orange) that were predicted to coordinate Mg 2+ . Lys 205 (green) provided a site for CO 2 fi xation. Amino acids important for catalysis of the carboxylation reaction (Lys 179, Arg 298, His 330, Lys 337, Gly 406, and Gly 407) are indicated in red; c. Global tertiary structure of SJ-rbcS. The dotted rectangle indicates a two-stranded anti-parallel β-sheet in the C-terminal.

382 CHIN. J. OCEANOL. LIMNOL., 32(2), 2014 Vol.32
(Table 2). As a result, 21 species from Stramenopiles and Rhodophyta were clustered into one clade (Red-like Form I Rubisco), while the other seven species in Cyanobacteria, Chlorophyta, and higher plants were
grouped in another clade (Green-like Form I Rubisco) (Fig.3). Moreover, brown algae were phylogenetically closer to Xanthophyta, Cryptophyta, and Haptophyta than to diatoms.
Xanthophyta
Cryptophyta
Rhodophyta
Bacillariophyta
Cyanobacteria
Higher plants
Chlorophyta
Phaeophyta
Haptophyta
Red-likeForm IRubisco
Green-likeForm IRubisco
Xanthophyta
Cryptophyta
Rhodophyta
Bacillariophyta
Cyanobacteria
Higher plants
Chlorophyta
Phaeophyta
Saccharina japonicaNereocystis luetk eanaChorda filumEctocarpus siliculosusPetrospongium rugosumScytosiphon dotyiFucus vesiculosusVaucheria litoreaTribonema virideBumilleria sp. SAG 2160Guillardia theta
Rhodomonas salinaCryptomonas parameciumEmiliania huxleyiPorphyridium aerugineumPorphyra purpureaPalmaria palmata
Phaeodactylum tricomutumCylindrotheca sp. Thalassiosira nordenskioeldiiOdontella sinensisProchlorothrix hollandicaSynechococcus sp.
Ulva proliferaChlamydomonas reinhardtiiOryza sativa Japonica GroupSpinacia oleraceaZea mays
Haptophyta
8397
80
997576
86
98988296
98
76
78
89
100100
66
0.232
6051
100638890
Fig.3 Phylogenetic tree based on alignment of Rubisco LSU amino acid sequences Putative Rubisco sequences were aligned using CLUSTAL X and analyzed with the MEGA 5.05 program using the neighbor-joining method. Bootstrap values calculated from 1 000 replicates are shown, and the branch lengths are proportional to the estimated evolutionary change. The scale bar corresponds to an estimation of 0.05 amino acid substitutions per site
Table 2 Sequences from all selected species for phylogenetic analysis
Taxonomy Accession number Taxonomy Accession number
Bumilleria sp. SAG 2160 ABS20374 Phaeodactylum tricornutum AF195952
Chlamydomonas reinhardtii J01399 Porphyra purpurea NC000925
Chorda fi lum AY372983 Porphyridium aerugineum X17597
Cryptomonas paramecium ACT46756 Rhodomonas salina ABO70740
Cylindrotheca sp. M59080 Prochlorothrix hollandica X57359
Ectocarpus siliculosus X52503 Saccharina japonica HM798587
Emiliania huxleyi AEI29479 Scytosiphon dotyi ABB72077
Fucus vesiculosus FM957154 Spinacia oleracea V00168
Guillardia theta AAC35639 Synechococcus sp. D13539
Nereocystis luetkeana DQ372565 Thalassiosira nordenskioeldii AB018007
Odontella sinensis NC001713 Tribonema viride ABS20372
Oryza sativa Japonica Group X15901 Ulva prolifera AFK24486
Palmaria palmata U28421 Vaucheria litorea AF207527
Petrospongium rugosum AAY53539 Zea m ays X86563

383No.2 SHAO et al.: Rubisco and its related genes in S . japonica
3.3 Transcriptional profi les of SJ - rbcL
At temperatures from 5 to 15°C, the mRNA expression of SJ - rbcL in kelp gametophytes was up-regulated continuously until the maximum at 15°C, but decreased at temperatures above 25°C (Fig.4). The trends of mRNA expression of SJ - Lhcf5 , SJ - CA ,
and SJ - CbbX were remarkably consistent, and reached peak levels at 15°C (Fig.4).
After incubation of gametophytes for 9 days (15°C, 45 μmol/(m 2 ∙s) PFD), the transcripts exhibited lower levels at the storage stage, which increased dramatically on day 1 (4.3-fold), and then increased gradually with the incubation time. It was up-regulated by 11.8-fold on day 7 compared with that at the beginning (Fig.5a), and fi nally remained stable during days 7–9.
The expression levels of SJ - rbcL in gametophytes were increased by photoperiods and exhibited a biological rhythm (Fig.5b). Transcript levels of rbcL were increased during the light period (simulated daytime) and reached a maximum after illumination for 12 h. Then, rbcL transcript levels declined sharply by 5.6-fold after 3 h of darkness (15 h point) and continuously decreased to the lowest level after 12 h of darkness (simulated night). From 24–48 h, rbcL transcript levels exhibited tendencies similar to those in the fi rst 24 h (Fig.5b).
00.050.1
0.150.2
0.250.3
0.350.4
SJ-CA SJ-CbbX SJ-Lhc SJ-RbcLGenes
Rel
ativ
e tra
nscr
iptio
n le
vel
5°C10°C15°C20°C25°C
Fig.4 mRNA expression of SJ - CA , SJ - CbbX , SJ - Lhc , and SJ - RbcL at various temperatures for 2 h Data are the means of three replicates, and the error bars indicate one standard deviation.
10
9
8
7
6
6
6 12 15 18 21 24 27 30 33 36 39 42 45 48
7 8 9
9
5
5
4
4Time (days)
Time (h)
Rel
ativ
e tra
nscr
iptio
n le
vel
Rel
ativ
e tra
nscr
iptio
n le
vel
b
a
3
3
3
2
2
1
10
0
0
7
6
5
4
3
2
1
0
Fig.5 mRNA expression of SJ - rbcL in gametophytes a. Expression of SJ - rbc during gametophyte incubation for 0–9 days; b. Diurnal variation of SJ - rbcL expression during 48 h. Data are the means of three replicates, and error bars indicate one standard deviation.
384 CHIN. J. OCEANOL. LIMNOL., 32(2), 2014 Vol.32
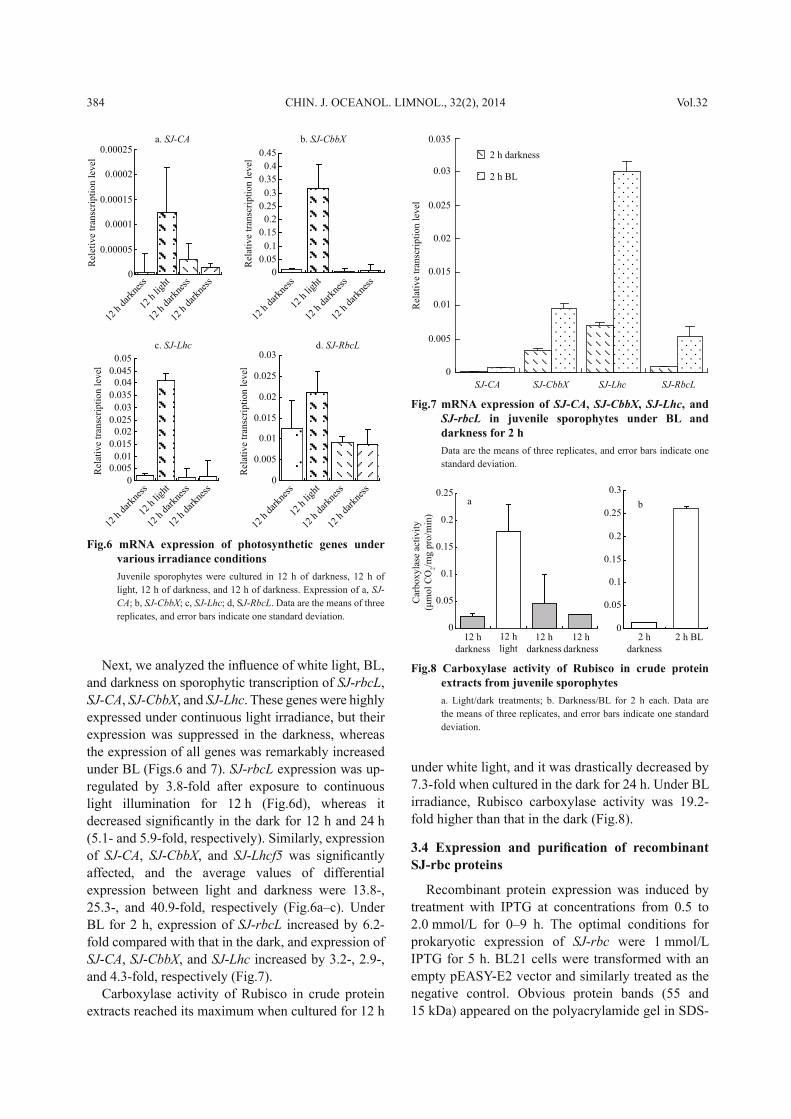
Next, we analyzed the infl uence of white light, BL, and darkness on sporophytic transcription of SJ - rbcL , SJ - CA , SJ - CbbX , and SJ - Lhc . These genes were highly expressed under continuous light irradiance, but their expression was suppressed in the darkness, whereas the expression of all genes was remarkably increased under BL (Figs.6 and 7). SJ - rbcL expression was up-regulated by 3.8-fold after exposure to continuous light illumination for 12 h (Fig.6d), whereas it decreased signifi cantly in the dark for 12 h and 24 h (5.1- and 5.9-fold, respectively). Similarly, expression of SJ - CA , SJ - CbbX , and SJ - Lhcf5 was signifi cantly affected, and the average values of differential expression between light and darkness were 13.8-, 25.3-, and 40.9-fold, respectively (Fig.6a–c). Under BL for 2 h, expression of SJ - rbcL increased by 6.2-fold compared with that in the dark, and expression of SJ - CA , SJ - CbbX , and SJ - Lhc increased by 3.2-, 2.9-, and 4.3-fold, respectively (Fig.7).
Carboxylase activity of Rubisco in crude protein extracts reached its maximum when cultured for 12 h
under white light, and it was drastically decreased by 7.3-fold when cultured in the dark for 24 h. Under BL irradiance, Rubisco carboxylase activity was 19.2-fold higher than that in the dark (Fig.8).
3.4 Expression and purifi cation of recombinant SJ-rbc proteins
Recombinant protein expression was induced by treatment with IPTG at concentrations from 0.5 to 2.0 mmol/L for 0–9 h. The optimal conditions for prokaryotic expression of SJ - rbc were 1 mmol/L IPTG for 5 h. BL21 cells were transformed with an empty pEASY-E2 vector and similarly treated as the negative control. Obvious protein bands (55 and 15 kDa) appeared on the polyacrylamide gel in SDS-
a. SJ-CA0.00025
0
0.00005
0.0001
0.00015
0.0002
12 h
darkn
ess
12 h
light
12 h
darkn
ess
12 h
darkn
ess
Rel
etiv
e tra
nscr
iptio
n le
vel
b. SJ-CbbX
d. SJ-RbcL
0
0.005
0.01
0.015
0.02
0.025
0.03
12 h
darkn
ess
12 h
light
12 h
darkn
ess
12 h
darkn
ess
Rel
ativ
e tra
nscr
iptio
n le
vel
00.05
0.10.15
0.20.25
0.30.35
0.40.45
12 h
darkn
ess
12 h
light
12 h
darkn
ess
12 h
darkn
ess
Rel
ativ
e tra
nscr
iptio
n le
vel
c. SJ-Lhc
00.005
0.010.015
0.020.025
0.030.035
0.040.045
0.05
12 h
darkn
ess
12 h
light
12 h
darkn
ess
12 h
darkn
ess
Rel
ativ
e tra
nscr
iptio
n le
vel
Fig.6 mRNA expression of photosynthetic genes under various irradiance conditions Juvenile sporophytes were cultured in 12 h of darkness, 12 h of light, 12 h of darkness, and 12 h of darkness. Expression of a, SJ - CA ; b, SJ - CbbX ; c, SJ - Lhc ; d, S J - RbcL . Data are the means of three replicates, and error bars indicate one standard deviation.
0
0.005
0.01
0.015
0.02
0.025
0.03
0.035
SJ-CA SJ-CbbX SJ-Lhc SJ-RbcL
Rel
ativ
e tra
nscr
iptio
n le
vel
2 h darkness
2 h BL
Fig.7 mRNA expression of SJ - CA , SJ - CbbX , SJ - Lhc , and SJ - rbcL in juvenile sporophytes under BL and darkness for 2 h Data are the means of three replicates, and error bars indicate one standard deviation.
0
0.05
0.1
0.15
0.2
0.25
12 hdarkness
2 hdarkness
12 hdarkness
12 hdarkness
12 hlight
Car
boxy
lase
act
ivity
(μm
ol C
O2/m
g pr
o/m
in)
0
0.05
0.1
0.15
0.2
0.25
0.3
2 h BL
a b
Fig.8 Carboxylase activity of Rubisco in crude protein extracts from juvenile sporophytes a. Light/dark treatments; b. Darkness/BL for 2 h each. Data are the means of three replicates, and error bars indicate one standard deviation.

385No.2 SHAO et al.: Rubisco and its related genes in S . japonica
PAGE (Fig.9a), which exhibited the presumed protein molecular weights. After elution with 200 mmol/L imidazole, the purifi ed SJ-rbcL protein was subjected to SDS-PAGE and western blot analysis. A 55 kDa protein band was positive for the anti-His antibody with high specifi city (Fig.9b).
4 DISCUSSION
4.1 Structural and phylogenetic characterization of SJ - rbc
Similar to Pylaiella littoralis , S accharina angustata , and S . latissima in Phaeophyta (Assali et al., 1991; Lane et al., 2006), both SJ-rbcL and SJ-rbcS are encoded by one sequence in the chloroplast genome with an A/T rich (83%) rubisco intergenic spacer (269 bp) in S . japonica . There are similar structures in species of Rhodophyta and Cryptophyta (Reith and Cattolico, 1986; Douglas and Durnford, 1989). Therefore, we assumed that rbc genes in Stramenopiles and Rhodophyta might have similar evolutional origins. Typical variations in algal rbc sequences exist in the Rubisco spacer, which can be applied to construct a phylogenetic tree for algal evolutional analysis.
Although rbc genes are intensively applied for evolutional analysis, our study is the fi rst to construct a phylogenetic tree using full-length sequences of rbcL . As a result, the close relationships between Stramenopiles and Rhodophyta were consistent with previous fi ndings, because Stramenopiles likely arose from secondary endosymbiosis with red algae based
on plastid-encoded genes and some nucleus-encoded proteins involved with the plastid (Yoon et al., 2002; Reyes-Prieto et al., 2007; Gould et al., 2008; Baurain et al., 2010). Nevertheless, it showed that the divergence of Bacillariophyta appeared earlier than other lineages in the Stramenopiles, which is different from other studies on the phylogeny of eukaryotes (Falkowski and Raven, 2007; Baldauf, 2008). We therefore assumed that diatoms might be the pioneer algal groups in Stramenopiles, while the Haptophytes and Cryptophytes are more closely related to brown algae. However, this difference may be partially because of the methods and gene sequences adopted, and needs to be verifi ed based on more sequence data (Balakirev et al., 2012).
4.2 Transcriptional analysis of SJ - rbcL in gametophytes
Presently, cryopreservation of gametophytes is a common method for kelp seedstock preservation (Liu et al., 2006; Yang et al., 2007), and the methods for recovery and ensuring normal growth after long-term preservation are still in their infancy (Li et al., 1999). Wang et al. (2011) reported that the expression level of SJ - rbcL is notably higher in gametophytes than that in sporophytes, inferring the presence of a C4-like pathway in sporophytes. However, no studies were performed on rbc expression during recovery of gametophytic growth. Our results show a gradual increase of SJ - rbcL transcripts during the algal de-differentiation and differentiation stages. The regulatory mechanism of gene expression changes still needs to be investigated further.
In higher plants, the photoperiod regulates expression of rbc genes (Yoon et al., 2001; Nishimura et al., 2008). However, in brown algae, the photoperiod also has effects on growth and development (Hsiao and Druehl, 1973; Henry, 1988; Wellnitz and Rinne, 1999; Gao et al., 2005). We found that SJ - rbcL expression was infl uenced by diurnal variations: increased expression during irradiance and suppressed in the darkness. This observation implied that SJ - rbcL is a photoperiod-regulated gene in S . japonica , even though the light-regulated network is currently unknown.
4.3 Transcriptional profi les of photosynthesis-related genes
In our study, mRNA levels of SJ - Lhcf5 , SJ - CA , SJ - CbbX , and SJ - rbcL increased to a maximum at 15°C, which is consistent with studies on optimal
kDa
95
7062
51
42
M 1M M1 2 3 6
97.266.4
14.3
44.3
29.0
20.1
kDa
a b
Fig.9 SDS-PAGE and the western blot analysis of recombinant rbc proteins M: protein marker; lanes 1 and 4 in a, negative controls with induction of empty expression vectors; lanes 2 and 5 in a, negative control for rbcS/rbcL recombinant vectors without IPTG induction; lane 3 and 6 in a, expression of rbcS/rbcL induced by 1 mmol/L IPTG for 5 h; lane 1 in b, western blot for rbcL obtained after 5 h of induction. Arrows indicate the proteins of interest

386 CHIN. J. OCEANOL. LIMNOL., 32(2), 2014 Vol.32
gametophytic growth conditions (Lüning, 1980; Li et al., 2004). Therefore, 15°C may be the optimal temperature to support highly effi cient photosynthesis for kelp.
Expression of SJ - Lhcf5 was up-regulated in juvenile sporophytes, which we believe may be more effective under light to capture more energy for PSII and indirectly for carbon fi xation during growth. These data are consistent with Lhcf5 expression in Cyclotella cryptica (Eppard et al., 2000) and diatoms (Papagiannakis et al., 2005). In addition, SJ - CbbX expression was elevated synchronously by white light, implying that CbbX might serve as a Rubisco activase in S . japonica . Surprisingly, after irradiance for 12 h, the expression of SJ - CA was nearly 2 500-fold less than that of SJ - CbbX and 300-fold less than that of SJ - Lhc . We suspect that the CCM process is less dependent on light for carboxylation of Rubisco in kelp.
Based on transcriptome data, Deng et al. (2012) found that many pathways are closely related to BL induction of gene expression. Here, we investigated genes potentially involved in BL-mediated metabolism. Based on the analysis of SJ - rbcL , SJ - CA , SJ - CbbX , and SJ - Lhcf5 expression, we found that BL might broadly up-regulate photosynthesis processes including light-harvesting, light-dependent, and dark reactions to produce primary substances more effectively and persistently for kelp growth. Interestingly, a recent study by Huysman et al. (2013) indicated that BL activates a combination of BL sensors and cyclin regulatory elements, leading to the onset of the cell cycle in diatoms. Our results implied an additional physiological process induced by BL and provide the foundation for further systematic studies of its underlying mechanism. In addition, under white light for 12 h and BL for 2 h, the carboxylase activity of Rubisco was increased by 7.3- and 19.2-fold in crude extracts, while SJ - rbcL expression was increased by 3.8- and 6.2-fold, respectively. Therefore, Rubisco activity is highly regulated by both light quantity and quality at both transcriptional and post-transcriptional levels.
As the dominant brown algae in sublittoral areas, S . japonica may have developed effi cient biological and biochemical mechanisms to overcome weak irradiance in seawater. One of the mechanisms is related to the light-induced expression of genes described in this study. Considering the above results, we propose that there might be an integrated pathway activated by light, mainly by blue light, which
underlies the photosynthesis of S . japonica . BL might be captured by light-harvesting proteins on the thylakoid membrane, which increases the level of chemical energy and ATP. ATP is then used by CbbX in the plastid, which has a typical ATPase domain and conserved ATP-binding motif (Shi et al., 2010), to activate Rubisco catalysis downstream in carbon fi xation.
4.4 Prokaryotic expression and purifi cation of Rubisco proteins
For prokaryotic expression, the Rubisco proteins reached maximal expression by induction at 1 mmol/L IPTG for 5 h. Furthermore, recombinant rbcL was completely eluted with 200 mmol/L imidazole. However, the purifi ed proteins may not have the correct folding and assembled structure. In fact, no attempts to reconstitute the active homologous enzyme have succeeded for form I Rubisco to our knowledge (Whitney et al., 2011), because of the indispensability of chaperonin RbcX, GroEL, GroES, Mg-ATP, and Proteinase K (Tarnawski et al., 2008; Liu et al., 2010). Thus, optimization and investigation of additional expression systems are needed for functional protein analysis.
5 CONCLUSION
In conclusion, our study provides an insight into the basic characters and physiological features of Rubisco in S . japonica . Various elements including Lhc, CA, CbbX, and Rubisco might be involved in the enhancement of photosynthetic effi ciency of gametophytes and juvenile sporophytes. Therefore, these proteins might play crucial roles in maintaining the robust status of kelp in habitats with weak irradiance and low temperatures.
6 ACKNOWLEDGEMENT
The authors are grateful to all the laboratory members for their technical advice and helpful suggestions. We acknowledge anonymous reviewers for the critical comments and suggestions for the manuscript.
References
Anderson J M, Chow W S, Park Y I. 1995. The grand design of photosynthesis: acclimation of the photosynthetic apparatus to environmental cues. Photosynth esis Res earch , 46 : 129-139.
Arnold K, Bordoli L, Kopp J, Schwede T. 2006. The SWISS-MODEL workspace: a web-based environment for protein

387No.2 SHAO et al.: Rubisco and its related genes in S . japonica
structure homology modelling. Bioinformatics , 22 : 195-201.
Assali N E, William F M, Charles C S, Susan L G. 1991. Evolution of the Rubisco operon from prokaryotes to algae: Structure and analysis of the rbcS gene of the brown alga Pylaiella littoralis . Plant Molecular Biolog y , 17 (4): 853-863.
Balakirev S E, Krupnova N T, Ayala J F. 2012. DNA variation in the phenotypically-diverse brown alga Saccharina japonica . BMC Plant Biology , 12 : 108.
Baldauf S L. 2008. An overview of the phylogeny and diversity of eukaryotes. Journal of Systematics and Evolution , 46 (3): 263-273.
Bartsch I, Wiencke C, Bischof K et al. 2008. The genus Laminaria sensulato : recent insights and developments. European Journal of Phycology , 43 (1): 1-86.
Baurain D, Brinkmann H, Petersen J, Rodríguez-Ezpeleta N, Stechmann A, Demoulin V, Roger A J, Burger G, Lang B F, Philippe H. 2010. Phylogenomic evidence for separate acquisition of plastids in Cryptophytes, Haptophytes, and Stramenopiles. Mol ecular Biol ogy and Evol ution , 27 (7): 1 698-1 709.
Berry J A, Osmond C B, Lorimer G. 1978. Fixation of CO 2 during photorespiration. Plant Physiol ogy , 62 : 954-967.
Bradford M M. 1976. Rapid and sensitive method for quantitation of microgram quantities of protein utilizing principle of protein-dye binding. Anal ytical Biochem istry , 72 : 248-254.
Deng Y, Yao J, Wang X, Guo H, Duan D. 2012. Transcriptome sequencing and comparative analysis of Saccharina japonica (Laminariales, Phaeophyceae) under blue light induction. PLoS ONE , 7 (6): e39704.
Douglas S E, Durnford D G. 1989. The small subunit of ribulose-l, 5-bisphosphate carboxylase is plastid-encoded in the chlorophyll c-containing alga Cryptomonas Ф. Plant Molecular Biology , 13 : 13-20.
Dring M J. 1981. Chromatic adaptation of photosynthesis in benthic marine algae: an examination of its ecological signifi cance using a theoretical model. Limnol ogy and Oceanogr aphy , 26 : 271-284.
Dron M, Rahire M, Rochaix J D. 1982. Sequence of the chloroplastDNA region of Chlamydomonas reinhardii containing the gene of the large subunit of ribulose bisphosphate carboxylase and parts of its fl anking genes. Journal of Molecular Biology , 162 : 775-793.
Eppard M, Krumbein W E, von Haeseler A, Rhiel E. 2000. Characterization of fcp4 and fcp12, two additional genes encoding light harvesting proteins of Cyclotella cryptica (Bacillariophyceae) and phylogenetic analysis of this complex gene family. Plant Biol ogy , 2 : 283-289.
Falkowski P G, Raven J A. 2007. Aquatic Photosynthesis. 2 nd edn. Princeton University Press, New Jersey, United States. ISBN 0-632-06139-1.
Felsenstein J. 1985. Confi dence limits on phylogenies: an approach using the bootstrap. Evolution , 39 : 783-791.
Gao J, Zhang Y, Zhang W, Wu S, Qin S, Zhang W, Yu X. 2005. Optimal light regime for the cultivation of transgenic
Laminaria japonica gametophytes in a bubble-column bioreactor. Biotechnol ogy Lett ers , 27 (18): 1 417-1 419.
Gasteiger E, Hoogland C, Gattiker A, Duvaud S, Wilkins M R, Appel R D, Bairoch A. 2005. Protein Identifi cation and Analysis Tools on the ExPASy Server. In : J M Walker ed. The Proteomics Protocols Handbook. Humana Press, New York, United States. p.571-607.
Ginrich J C, Hallick R B. 1985. The Euglena gracilis chloroplast ribulose-1, 5-bisphosphate carboxylase gene. II. The spliced mRNA and its product. J ournal of Biol ogical Chem istry , 260 : 16 162-16 168.
Giordano M, Beardall J, Raven J A. 2005. CO 2 concentrating mechanisms in algae: mechanisms, environmental modulation, and evolution. Annu al Rev iew of Plant Biol ogy , 56 : 99-131.
Gould S B, Waller R F, McFadden G I. 2008. Plastid evolution. Annu al Rev iew of Plant Biol ogy , 59 : 491-517.
Guex N, Peitsch M C. 1997. SWISS-MODEL and the Swiss-PdbViewer: an environment for comparative protein modelling. Electrophoresis , 18 : 2 714-2 723.
Henry E C. 1988. Regulation of reproduction in brown algae by light and temperature. Botanica Marina , 31 : 353-357.
Hsiao S I C, Druehl L D. 1973. Environmental control of gametogenesis in Laminaria Saccharina IV in situ development of gametophytes and young sporophytes. Journal of Phycology , 9 : 160-165.
Huysman M J J, Fortunato A E, Matthijs M, Costa B S, Vanderhaeghen R, Daele H Van den, Sachse M, Inzé D, Bowler C, Kroth P G, Wilhelm C, Falciatore A, Vyverman W, Veylder L D. 2013. AUREOCHROME1a-mediated induction of the diatom-specifi c cyclin dsCYC2 controls the onset of cell division in diatoms ( Phaeodactylum tricornutum ). The Plant Cell , http://dx.doi.org/10.1105/tpc.112.106377.
Jensen G R, Bahr T J. 1977. Ribulose 1.5-bisphosphate carboxylase-oxygenase. Ann ual Rev iew of Plant Physiol ogy , 28 : 379-400.
Kai B, Gudrun K, Christian W, Dieter H. 2002. Solar ultraviolet radiation affects the activity of ribulose-1,5-bisphosphate carboxylase-oxygenase and the composition of photosynthetic and xanthophyll cycle pigments in the intertidal green alga Ulva lactuca L. Planta , 215 : 502-509.
Kirk J T O. 1994. Light and Photosynthesis in Aquatic Ecosystems. Cambridge University Press, Cambridge, England.
Klenell M, Snoeijs P, Pedersén M. 2002. The involvement of a plasma membrance H + -ATPase in the blue-light enhancement of photosynthesis in Laminaria digitata (Phaeophyta). J ournal of Phycology , 38 : 1 143-1 149.
Krogh A, Larsson B, von Heijne B, Sonnhammer E L L. 2001. Predicting transmembrane protein topology with a hidden Markov model: application to complete genomes. J ournal of Mol ecular Biol ogy , 305 (3): 567-580.
Lan Y, Mott K A. 1991. Determination of apparent KM values for ribulose 1,5-bisphosphate carboxylase oxygenase (Rubisco) activase using the spectrophotometric assay of

388 CHIN. J. OCEANOL. LIMNOL., 32(2), 2014 Vol.32
Rubisco activity. Plant Physiol ogy , 95 : 604-609. Lane C E, Mayes C M, Druehl L G, Saunders G W. 2006. A
multi-gene molecular investigation of the kelps (Laminariales, Phaeophyceae) resolves competing phylogenetic hypotheses and supports substantial taxonomic re-organization. J ournal of Phycol ogy , 42 : 493-512.
Lee S R, Oak J H, Keum YS, Lee J A, Chung I K. 2011. Utility of rbc S gene as a novel target DNA region for brown algal molecular systematics. Phycological Research , 59 : 34-41.
Li D P, Zhou Z G, Liu H H, Wu C Y. 1999. A new method of Laminaria japonica strain selection and sporeling raising by the use of gametophyte clones. Hydrobiologia , 398/399 : 473-476.
Li X J, Wang G W, Zhang Q S, Zhang Z Z, Luo S J. 2004. The effects of temperature on growth of gametophytes of Laminaria japonica 901. Fisheries Science & Technology Information , 31 (4): 66-168. (in Chinese with English abstract)
Liu C M, Young A L, Starling-Windhof A, Bracher A, Saschenbrecker S, Rao B V, Rao K V, Berninghausen O, Mielke T, Hartl F U, Beckmann R, Hayer-Hartl M. 2010. Coupled chaperone action in folding and assembly of hexadecameric Rubisco. Nature , 463 : 197-202.
Liu T, Zhang J, Meng X L, Wang X Y, Cui J J. 2006. Studies of ultralow cryopreservating female gametophytes clone of Laminaria japonica . Acta Oceanologica Sinica , 28 (2): 175-177. (in Chinese with English abstract)
Lüning K, Dring M J. 1975. Reproduction, growth and photosynthesis of gametophytes of Laminaria saccharina growth in blue and red light. Mar ine Biol ogy , 29 : 195-200.
Lüning K. 1980. Critical levels of light and temperature regulating the gametogenesis of the three Laminaria species (Phaeophyceae). J ournal of Phycol ogy , 16 : 1-15.
MacIntyre H L, Kana T M, Geider R J. 2000. The effect of water motion on short-term rates of photosynthesis by marine phytoplankton. Trends Plant Sci ence , 5 : 12-17.
Matsuda R, Ohashi-Kaneko K, Fujiwara K, Kurata K. 2008. Effects of blue light defi ciency on acclimation of light energy partitioning in PSII and CO 2 assimilation capacity to high irradiance in Spinach leaves. Plant Cell Physiol ogy , 49 (4): 664-670.
Miller A D, Connell M U. 2012. Photoperiodic mediation of differential gene expression in Scytosiphon lomentaria (Phaeophyceae). Phycological Research , 60 : 105-113.
Mizohata E, Matsumura H, Okano Y, Kumei M, Inoue T, Yokota A, Kai Y. 2002. Crystal structure of activated ribulose-1,5-bisphosphate carboxylase/oxygenase from green alga Chlamydomonas reinhardtii complexed with 2-carboxyarabinitol-1,5-bisphosphate. J ournal of Mol ecular Biol ogy , 316 : 679-691.
Mizuta H, Kai T, Tabuchi K, Yasui H. 2007. Effects of light quality on the reproduction and morphology of sporophytes of Laminaria japonica (Phaeophyceae). Aquac ulture Res earch , 38 : 1 323-1 329.
Moroney J V, Ynalvez R A. 2009. Algal Photosynthesis. In:
eLS. John Wiley & Sons Ltd., Chichester. http://www.els.net. http://dx.doi.org/10.1002/9780470015902.a0000322.pub2.
Mueller-Cajar O, Stotz M, Wendler P, Hartl U, Bracher A, Hayer-Hartl M. 2011. Structure and function of the AAA + protein CbbX, a red-type Rubisco activase. Nature , 479 : 194-199.
Nishimura K, Ogawa T, Ashida H, Yokota A. 2008. Molecular mechanisms of RuBisCO biosynthesis in higher plants. Plant Biotechnology , 25 : 285-290.
Papagiannakis E, van Stokkum I H M, Fey H, Büchel C, van Grondelle R. 2005. Spectroscopic characterization of the excitation energy transfer in the fucoxanthin-chlorophyll protein of diatoms. Photosynth esis Res earch , 86 : 241-250.
Petersen T N, Brunak S, von Heijne G, Nielsen H. 2011. SignalP 4.0: discriminating signal peptides from transmembrane regions. Nat ure Methods , 8 : 785-786.
Phillips N, Burrowes R, Rousseau F, De Reviers B, Saunders G W. 2008. Resolving evolutionary relationships among the brown algae using Chloroplast and Nuclear genes. Journal of Phycology , 44 : 394-405.
Reinfelder J R. 2011. Carbon concentrating mechanisms in Eukaryotic marine phytoplankton. Annu al Rev iew of Mar ine Sci ence , 3 : 291-315.
Reith M, Cattolico R A. 1986. Inverted repeat of Olisthodiscus luteus chloroplast DNA contains genes for both subunits of ribulose-1, 5-bisphosphate carboxylase and the 32,000-dalton QB protein: phylogenetic implications. Proc eedings of Nat ional Acad emy of Sci ences , 83 : 8 599-8 603.
Reyes-Prieto A, Weber A P, Bhattacharya D. 2007. The origin and establishment of the plastid in algae and plants. Annu al Review of Genetics , 41 : 147-168.
Rousvoal S, Groisillier A, Dittami S M, Michel G, Boyen C, Tonon T. 2011. Mannitol-1-phosphate dehydrogenase activity in Ectocarpus siliculosus , a key role for mannitol synthesis in brown algae. Planta , 233 : 261-273.
Sayers E W, Barrett T, Benson D A, Bolton E, Bryant S H et al. 2012. Database resources of the National Center for Biotechnology Information. Nucleic Acids Res earch , 40 : D13-D25.
Schmittgen T D, Zakrajsek B A, Mills A G, Gorn V, Singer M J, Reed M W. 2000. Quantitative reverse transcription-polymerase chain reaction to study mRNA decay: comparison of endpoint and real-time methods. Anal ytical Biochem istry , 285 : 194-204.
Schwede T, Kopp J, Guex N, and Peitsch M C. 2003. SWISS-MODEL: an automated protein homology-modeling server. Nucleic Acids Research , 31 : 3 381-3 385.
Shi W W, Wang L L, Chen J, Ou-Yang L L, Zhou Z Z. 2010. Characterization and differential expression of CbbX gene between female and male gametophytes of Laminaria japonica . Journal of Fisheries of China , 34 (1): 80-88. (in Chinese with English abstract)
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. 2011. MEGA5: molecular evolutionary genetics

389No.2 SHAO et al.: Rubisco and its related genes in S . japonica
analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol ecular Biol ogy and Evol ution , 28 : 2 731-2 739.
Tarnawski M, Gubernator B, Kolesinski P, Szczepaniak A. 2008. Heterologous expression and initial characterization of recombinant RbcX protein from Thermosynechococcus elongatus BP-1 and the role of RbcX in Rubisco assembly. Acta Biochim ica Pol onica , 55 : 777-785.
Thompson J D, Gibson T J, Plewniak F, Jeanmougin F, Higgins D G. 1997. The ClustalX windows interface: fl exible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research , 24 : 4 876-4 882.
Tseng C K, Wu C Y. 1962. Cultivation of Laminaria japonica . 1 st edn. Science Press, Beijing, China. p.14-15. (in Chinese)
Walters R G. 2005. Towards an understanding of photosynthetic acclimation. J ournal of Exp erimental Bot any , 411 : 435-447.
Wang C, Fan X, Wang G, Niu J, Zhou B. 2011. Differential expression of rubisco in sporophytes and gametophytes of some marine macroalgae. PLoS ONE , 6 (1): e16351.
Wang W J, Sun X T, Wang F J. 2010. Effect of blue light on early sporophyte development of Saccharina japonica (Phaeophyta). Mar ine Biol ogy , 157 : 1 811-1 817.
Wellnitz T, Rinne B. 1999. Photosynthetic response of stream periphyton to fl uctuating light regimes. J ournal of Phycol ogy , 35 : 667-672.
Whitney S M, Houtz R L, Alonso H. 2011. Advancing our understanding and capacity to engineer nature’s CO 2 -sequestering enzyme, Rubisco. Plant Physiology , 155 : 27-35.
Win N N, Hanyuda T, Arai S, Uchimura M, Prathep A, Draisma S G A, Phang S M, Abbott I A, Millar A J K, Kawai H. 2011. A taxonomic study of the genus Padina (Dictyotales, Phaeophyceae) including the descriptions of four new species from Japan, Hawaii, and the Andaman Sea. Journal of Phycology , 47 : 1 193-1 209.
Yang G P, Li X J, Cong Y Z, Qu S C, Li Z L, Zhang Z Z, Luo S J. 2007. Trends in the cloning methodology of Laminaria gametophytes: research and application. Periodical of Ocean University of China , 37 (4): 569-572. (in Chinese with English abstract)
Yao J T, Fu W D, Wang X L, Duan D L. 2009. Improved RNA isolation from Laminaria japonica Aresch (Laminariaceae, Phaeophyta). J ournal of Appl ied Phycol ogy , http://dx.doi.org/10.1007/s10811-008-9354-0.
Ying C Q, Yin S J, Shen Y, Lin S J, He P M. 2011. Cloning and analysis of the full-length Rubisco large subunit (rbcL) cDNA from Ulva linza (Chlorophyceae, Chlorophyta). Botanica Marina , 54 : 303-312.
Yoon H S, Hackett J D, Pinto G, Bhattacharya D. 2002. The single, ancient origin of chromist plastids. Proc eedings of Nat ional Acad emy of Sci ences , 99 : 15 507-15 512.
Yoon M, Putterill J J, Ross G S, Laing W A. 2001. Determination of the relative expression levels of Rubisco small subunit genes in Arabidopsis by rapid amplifi cation of cDNA ends. Analytical Biochemistry , 291 : 237-244.
Zou D Y, Bi Y H, Zhou Z G. 2009. Effects of light on the relative transcription of lhcf5 and lhcf6 genes from the gametophytes of Laminaria japonica . Journal of Fishery Sciences of China, 16 (6): 850-858. (in Chinese with English abstract)





























