Embed Size (px)
Citation preview

ARTICLE
Chemical ecology of lichens and species composition of cryptogamsamong three boreal habitats in eastern ManitobaS.A. Toni and M.D. Piercey-Normore
Abstract: The Precambrian Shield supports a diversity of cryptogams where environmental conditions predict their distribu-tion. Moisture and light are thought to affect secondary metabolite production in lichens, leading to a hypothesis of chemicalcommunities that may be independent of species assemblages. Hypotheses were that habitats will be characterized by lichensecondary metabolites and that lichen and bryophyte species composition will also distinguish among habitats. Lichens andbryophytes were sampled from six quadrats from each of two sites within each of three habitats (rock faces, rock outcrops, andforest floors). This study showed strong relationships among species assemblages, secondary metabolites, and types of habitat.Species abundance, cover, and species richness also differed among habitats. Individual secondary metabolites could differen-tiate among three habitats, suggesting that certain metabolites such as usnic and squamatic acids would more likely be foundin habitats such as dry exposed rock outcrops than in moist shaded habitats. Individual analyses suggested that some secondarymetabolites may serve multiple functions in a habitat, such as triterpenes, while others may have more specific functions, suchas salazinic acid, where environmental featuresmay be important for secondarymetabolite production. Further investigation ofspecific metabolites is needed to understand their roles in adaptation.
Key words: bryophytes, cyanobacterial lichens, fungal secondary metabolites, mosses.
Résumé : Le bouclier précambrien supporte une diversité de cryptogames où les conditions environnementales permettent deprédire leurs distributions. On croit que l'humidité et la lumière affectent la production de métabolites secondaires chezles lichens conduisant a l'hypothèse de �communautés chimiques' pouvant être indépendantes de l'assemblage des espèces. Leshypothèses sont que les habitats se caractériseraient par lesmétabolites secondaires des lichens et que la composition en espècesde lichens et de bryophytes feraint la différences entre les habitats. Les auteurs ont échantillonné les lichens et lesbryophytes dans six quadrats de chacun de deux sites dans chacun de trois habitats (crêts rocheux, affleurements rocheuxet parterres forestiers). Cette étudemontre qu'il existe une forte relation entre l'assemblage des espèces et leurs métabolitessecondaires, avec les types d'habitat. L'abondance des espèces, la couverture ainsi que la richesse en espèces diffèrentégalement entre les différents habitats. Les métabolites secondaires individuels pourraient faire la différence entre les troishabitats, suggérant que certains métabolites comme les acides usniques et squamatiques se retrouveraient plutôt dansdes habitats comme les crêts rocheux secs et exposés, que dans des habitats humides et ombragés. Les analyses individuellessuggèrent que certains métabolites secondaires pourraient jouer divers rôles dans un habitat, comme les triterpènes, alorsque d'autres auraient des fonctions plus spécifiques comme l'acide salazinique, où les caractéristiques environnementalespourraient être importantes pour la production de métabolites secondaires. De plus, on doit conduire des recherches surles métabolites secondaires pour comprendre leurs rôles dans l'adaptation. [Traduit par la Rédaction]
Mots-clés : bryophytes, lichens cyanobactériens, métabolites fongiques secondaires, mousses.
IntroductionLichens and bryophytes (cryptogams) are known to be effective
indicators of environmental conditions because there is no bar-rier between the ambient environment and the body of the lichenor bryophyte. Water is rapidly gained and lost as external atmo-spheric levels fluctuate resulting in a variation of cryptogam tol-erance to the rate and frequency of desiccation. Becausepollutants found in the atmosphere or substratum are taken upwith water (cf. Bates 2009), lichens and bryophytes have also beenused globally as pollution monitors (Nash 2008). After baselinedata has been established, the species composition of cryptogamcommunities can be useful in monitoring ecosystem health.Whiteshell Provincial Park, located on the Precambrian Shield ineastern Manitoba, has had a survey of its bryophyte communities(Stringer and Stringer 1973), but there are no habitat-specific dataavailable. A habitat-specific survey for lichens has not been under-
taken, so no baseline data are available for boreal forest habitatsin the park. Boreal forests provide diverse habitats where green-algal lichens are superior competitors under high light levels andlow humidity. Bryophytes outcompete green algal lichens underlow light levels and high humidity, and potentially high nutrientlevels in throughfall (Johnson 1981). Cyanobacterial lichens maybe successful in conditions between these two extremes depend-ing on the thallus structure and the number of photobionts. Thal-lus structuremay be homiomerous where cyanobacteria form themajor component of the thallus with a thin outer fungal layer.Alternatively, a heteromerous thallus has distinct layers includ-ing a cortex of fungal hyphae, a layer of photobiont cells, a looselywoven medulla, and sometimes a lower cortex. Cyanobacteriamay form the primary photobiont layer of a heteromerous thal-lus, or theymay form the secondary photobiont restricted to com-partments called cephalodia, while green algae form the primary
Received 12 June 2012. Accepted 22 September 2012.
S.A. Toni and M.D. Piercey-Normore. Department of Biological Sciences, University of Manitoba, Winnipeg, MB R3T 2N2, Canada.
Corresponding author: Michele Piercey-Normore (e-mail: [email protected]).
53
Botany 91: 53–61 (2013) dx.doi.org/10.1139/cjb-2012-0139 Published at www.nrcresearchpress.com/cjb on 31 January 2013.
Bot
any
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Geo
rge
Mas
on U
nive
rsity
on
03/2
1/13
For
pers
onal
use
onl
y.
photobiont layer. The co-existence of different forms of greenalgal and cyanobacterial lichens and bryophytes would be possi-ble if a mosaic of microclimates existed in a habitat.
Habitat conditions may influence cryptogam community com-position (Hauck 2011; Seaward 2008) and the phenotypes of li-chens (Kunkel 1980; Rikkinen 1997) and bryophytes (Hassel et al.2005; Shaw 1985), including the type of secondary metabolite pro-duced by many nonlichenized fungi (Fox and Howlett 2008). Fun-gal secondary metabolites are chemical by-products of primarymetabolism, of which over 1000 secondarymetabolites are knownto be produced by lichen fungi (Huneck 1999). While some studieshave examined the zonation of metabolites produced by siblingspecies and chemical strains (Culberson and Culberson 1967;Culberson et al. 1977), few studies have examined ecologicaltrends among lichen metabolites of co-occurring genera. If sec-ondary metabolites are ecologically adaptive (Lawrey 1986), li-chens should produce similar metabolites or metabolites similarin function to those produced by their neighbours resulting infunctional chemical “communities”, which may not reflect spe-cies assemblages.
While the production of secondarymetabolites is thought to beadaptive, those species that do not produce these metaboliteshave not been eliminated from habitats, and may grow in envi-ronments inwhich secondarymetabolites do not offer substantialselective advantages and (or) where production of the metaboliteis not initiated. The production of squamatic (Stocker-Wörgötter2001) and grayanic acid (Culberson and Armaleo 1992) was onlystimulated when lichen cultures began to dehydrate. Studies havereported roles for lichen metabolites in cation uptake and toler-ance (Hauck et al. 2009), regulation of thallus water content, andprotection from UV radiation (Lawrey 1986; Nybakken et al. 2007).Therefore, the production of particular metabolites or groups ofmetabolites could be expected under those environmental condi-tions in which they could be adaptive. The production of lichenmetabolites may also have been influenced by historic environ-mental conditions and may not be of any present benefit. If someproperties of lichen metabolites are currently ecologically adap-tive, then species or individuals with differing types and quanti-ties of compounds should occupy different niches.
The goal of this study was to elucidate the relationship amongassemblages of species in three habitats with a focus on the sec-ondary metabolites produced by lichen species and to identifyspecificmetabolites or profiles of metabolites that may play a rolein adaptation. Specific hypotheses were primarily that habitatswill be characterized by suites of lichen secondary metabolites,and secondarily that lichen and bryophyte species compositionwill also distinguish among habitats.
Materials and methodsThe study was conducted in the Whiteshell Provincial Park lo-
cated along the Manitoba–Ontario border. The predominant treecomposition was white spruce (Picea glauca (Moench) Voss), bal-sam fir (Abies balsamea (L.) Mill.), and paper birch (Betula papyriferaMarsh.) on well-drained sites; jack pine (Pinus banksiana Lamb.)stands in drier, sandier areas; balsampoplar (Populus balsamifera L.)and aspen (Populus tremuloidesMichx.) on moist river valley slopes;and black spruce (Picea mariana (Mill.) BSP.) or tamarack (Larixlaricina (DuRoi) K. Koch) bog forests in poorly drained regions(Stringer and Stringer 1973). The underlying geology is primarilyacidic Precambrian gneisses.
Three terrestrial habitats within the southern portion of thepark were selected for their variation in factors such as light andmoisture availability (Table 1). The first habitat consisted of rockfaces in proximity to a persistent water body. The predominantcanopycomposition inthefirst sitewaswhitecedar (ThujaoccidentalisL.),balsam fir, and paper birch with a vertical shaded cliff. The pre-dominant canopy composition for the second site within this hab-itat was paper birch, maple (Acer sp. L.), green ash (Fraxinuspennsylvanica (Vahl) Fern.), and trembling aspen, with beaked ha-zel (Corylus cornuta Marsh.) in the understory with large shadedboulders along a river adjacent to a sloping outcrop. The secondhabitat was exposed granitic outcrops where one site consisted ofrelatively low-lying granitic tables interspersedwith jack pine andblueberry (Vaccinium angustifolium Ait.), and the second site waselevated granitic tables with surrounding vegetation similar tothat of the first site as well as Lycopodium species. The third habitattype consisted of the forest floor of closed coniferous stands not
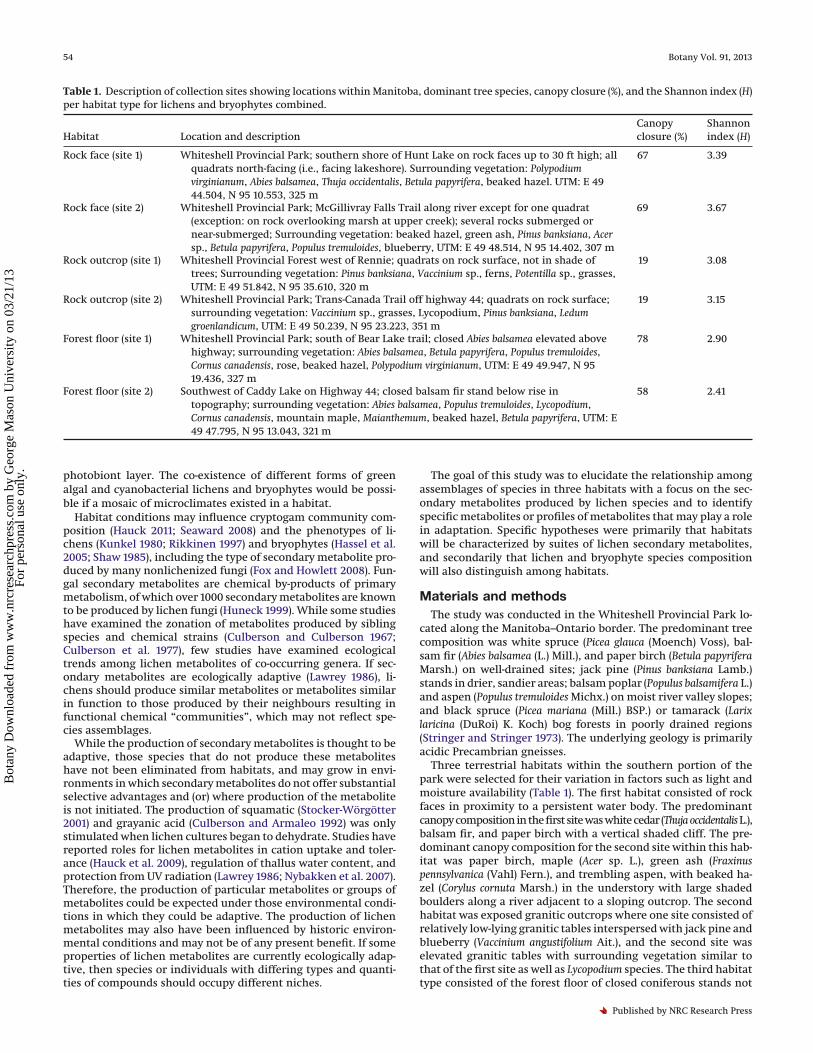
Table 1. Description of collection sites showing locations within Manitoba, dominant tree species, canopy closure (%), and the Shannon index (H)per habitat type for lichens and bryophytes combined.
Habitat Location and descriptionCanopyclosure (%)
Shannonindex (H)
Rock face (site 1) Whiteshell Provincial Park; southern shore of Hunt Lake on rock faces up to 30 ft high; allquadrats north-facing (i.e., facing lakeshore). Surrounding vegetation: Polypodiumvirginianum, Abies balsamea, Thuja occidentalis, Betula papyrifera, beaked hazel. UTM: E 4944.504, N 95 10.553, 325 m
67 3.39
Rock face (site 2) Whiteshell Provincial Park; McGillivray Falls Trail along river except for one quadrat(exception: on rock overlooking marsh at upper creek); several rocks submerged ornear-submerged; Surrounding vegetation: beaked hazel, green ash, Pinus banksiana, Acersp., Betula papyrifera, Populus tremuloides, blueberry, UTM: E 49 48.514, N 95 14.402, 307 m
69 3.67
Rock outcrop (site 1) Whiteshell Provincial Forest west of Rennie; quadrats on rock surface, not in shade oftrees; Surrounding vegetation: Pinus banksiana, Vaccinium sp., ferns, Potentilla sp., grasses,UTM: E 49 51.842, N 95 35.610, 320 m
19 3.08
Rock outcrop (site 2) Whiteshell Provincial Park; Trans-Canada Trail off highway 44; quadrats on rock surface;surrounding vegetation: Vaccinium sp., grasses, Lycopodium, Pinus banksiana, Ledumgroenlandicum, UTM: E 49 50.239, N 95 23.223, 351 m
19 3.15
Forest floor (site 1) Whiteshell Provincial Park; south of Bear Lake trail; closed Abies balsamea elevated abovehighway; surrounding vegetation: Abies balsamea, Betula papyrifera, Populus tremuloides,Cornus canadensis, rose, beaked hazel, Polypodium virginianum, UTM: E 49 49.947, N 9519.436, 327 m
78 2.90
Forest floor (site 2) Southwest of Caddy Lake on Highway 44; closed balsam fir stand below rise intopography; surrounding vegetation: Abies balsamea, Populus tremuloides, Lycopodium,Cornus canadensis, mountain maple, Maianthemum, beaked hazel, Betula papyrifera, UTM: E49 47.795, N 95 13.043, 321 m
58 2.41
54 Botany Vol. 91, 2013
Published by NRC Research Press
Bot
any
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Geo
rge
Mas
on U
nive
rsity
on
03/2
1/13
For
pers
onal
use
onl
y.

directly adjacent to any persistent sources of water. The forestcanopy at the first site was composed of balsam fir, paper birch,and trembling aspen, with an understorey of beaked hazel, Cornuscanadensis L., and Rosa sp. L. The second site was composed ofbalsam fir and trembling aspen, with an understorey of beakedhazel, Acer spicatum Lam., and Cornus canadensis.
Field experiments were conducted by choosing three habitats(rock face, rock outcrop, and forest floor), each represented by twosites (for six total sites in three habitats), and six 1 m × 1 m quad-rats were placed within each site and were positioned to includeat least one cyanolichen species within the quadrat boundaries.Within each quadrat, percent cover of all lichen and bryophytespecies was determined by visually estimating the proportion ofthe quadrat occupied by a given species. Canopy closure was esti-mated on an increasing scale of 0–3, where the value corre-sponded to the proportion of the sky directly obscured by thecanopy by looking upward through a 250 cm × 250 cm grid helddirectly above the quadrat (e.g., 0 = open sky, 1 = one-third of theview obstructed by tree canopy, 2 = two-thirds of the view ob-structed by tree canopy, and 3 = completely obscured).
Lichen and bryophyte species were identified to the lowesttaxon possible in the field and vouchers were collected. All lichenvouchers were confirmed in the laboratory using morphology,chemical spot tests, and thin-layer chromatography (TLC) (HindsandHinds 2007; Goward 1999; Brodo et al. 2001). Bryophyte vouch-ers were identified using sporophyte (when available) and ga-metophyte morphology (Crum 1983; Redfearn 1979; Crum andAnderson 1981) to species level. Lichen nomenclature followsEsslinger (2010), and bryophyte nomenclature follows Andersonet al. (1990). All collected vouchers are deposited in the cryptogamdivision of the University of Manitoba Herbarium (WIN).
Lichen secondary compounds were detected by TLC using aprocedure modified from Culberson (1972) and Orange et al.(2001). Compounds were extracted using an acetone rinse andthen run in TLC solvent A (toluene (180 mL); dioxane (45 mL);acetic acid (5 mL)). TLC characteristics were compared with thosecompiled by C. Culberson (Duke University, unpublished) and byOrange et al. (2001) for lichen compounds. Compounds that weredifficult to identify were checked against literature of previousstudies on the chemistry of that particular species. The inability ofTLC to detect the presence of low levels of compounds withinsome samples could further act as an indication of the lesser roleof that compound in ecological adaptation.
Species and habitat data were analysed by one-way ANOVAs andTukey–Kramer HSDs (honestly significant differences) to determinethe significance of differences among habitats in log-transformedlichen,moss, and cyanolichen percent cover. Alpha-diversity (amea-sure of species richness and evenness within a site) was calculatedfor each site and habitat using the Shannon index (H).H values canrange from 0, indicating low species diversity, to over 3.5, indicat-ing high species diversity within a habitat. Beta-diversity (a mea-sure of species replacement in different habitats) was calculatedbetween the two sites in each habitat and among habitats usingthe Sørenson similarity index. The Sørenson similarity indexranges from 0 to 1, where 1 indicates all species were present inboth locations and 0 indicates no species were shared betweenlocations.
Principle coordinates analysis (PCoA) explored the relation-ships between habitat with species assemblages and individualsecondary metabolites. The explanatory variable was the percentcover of species in each quadrat for each habitat type. The Hell-inger index was used to make a similarity matrix and PCoA anal-ysis was done using the program PrCoord to generate thesimilarity matrix among habitats. The matrix was then subjectedto Canoco version 4.5 to perform a principle components analysis(PCA) on the matrix, resulting in a PCoA. Other methods wereexplored for use with species data. PCA assumes linearity of datathat is not appropriate for the data in this study (McCune and
Grace 2002). Correspondence analysis (CA/CCA) assumes uni-modal data but it presents problems associated with ecologicaldata (McCune and Grace 2002) even though it is widely used(Kenkel 2006). PCoA was chosen as the method to analyse data inthis study because it does not make assumptions about species'responses to environmental variables; it can be used on nonlineardata where similarity coefficients are not related to species re-sponse; and it presents the least distortion of the data (McCuneand Grace 2002).
Secondary metabolites were analysed as profiles (combinationofmetabolites produced by each species) and as individual metab-olites (separate metabolites that may be present inmore than onespecies). Secondary metabolite profile data were analysed by one-way ANOVA to test whether the habitats can be distinguishedusing log-transformed percent cover and frequency of occurrenceof each secondary metabolite profile. The profiles were also com-pared to each other by combining habitats to identify profiles thatmay be more abundant than others. Individual secondary metab-olites were analysed by one-way ANOVA to test whether the hab-itats can be distinguished using log-transformed percent coveronly. Pairwise significant differences were tested by Tukey's test.All univariate analyses were performed using JMP (version 8.0,2008; SAS Institute Inc., Cary, North Carolina, USA) and data werelog-transformed.
ResultsSeventy-eight species of lichens and bryophytes (50 lichens,
28 bryophytes) were identified across all three habitats (Fig. 1).Rock faces had the largest number of species (51 species, 34 li-chens, 17 bryophytes), rock outcrops had 35 species (26 lichens,9 bryophytes), and coniferous forest floors had the fewest species(21 species, 8 lichens, 13 bryophytes). The greatest number of spe-cies (30 species) restricted to any one habitat occurred on rockfaces, while 20 were restricted to rock outcrops and 7 species wererestricted to coniferous forest floors. Although 78 species werecollected in this study, a larger number of species are present in theWhiteshell Provincial Park that arenot reported in this study. Themostabundant species in any habitat was Cladonia arbuscula in the rock out-crop habitat andHedwigia ciliata in the rock face habitat.
Prediction of habitat differences by speciesThe Shannon index of species diversity showed that the rock
faces were the most diverse, followed by the rock outcrops, andthen the forest floors (Table 1). Sørenson similarity indices indi-cate the proportion of recurring species within and among habi-tats (Table 2). Rock outcrops and forest floor habitats had thehighest recurrence of species, with more than half the speciesencountered at both sites within each habitat type. Rock faces hadapproximately one-third of their species occur at both sites.Whenhabitat types were compared, rock outcrops and forest floorsshared the fewest species (0.21) (Table 2). Rock faces and forestfloors shared the greatest proportion of species (0.39). Each of thetwo sites of rock faces were slightly more similar to coniferousforest floors and rock outcrops than to each other, owing to rela-tively low numbers of recurring species on rock faces (0.36).
The relationship between lichens and mosses was a negativerelationship. The habitats differed significantly in the percentcover and frequency for each of lichens and bryophytes (Fig. 2).Rock faces had significantly greater lichen cover than forestfloors (P = 0.0003) (Fig. 2A). No difference in moss cover wasobserved among the three habitats. Lichens were significantlymore frequent in the rock outcrops than the forest floors(P = 0.0130) (Fig. 2B). However, there was no difference in thefrequency of occurrence of mosses among the three habitats.The PCoA analysis showed that the quadrats from the samehabitat clustered together (Fig. 3A). The ordination shows onlythe first and second axes, which comprise 17.8% and 11% of thevariation in the data.
Toni and Piercey-Normore 55
Published by NRC Research Press
Bot
any
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Geo
rge
Mas
on U
nive
rsity
on
03/2
1/13
For
pers
onal
use
onl
y.

The percent cover of lichens with different photobionts alsoreflected different habitats. The percent cover of jelly lichenswas significantly higher in the rock face habitat than other
habitats (P = 0.0122). Cephalodiate lichens were present only inthe rock outcrop so results are not shown. The green algallichens were significantly higher in both the rock face and the
Fig. 1. Relative abundance of lichen and bryophyte species in each of three habitats (rock face, rock outcrop, forest floor) showing frequencyof occurrence (%, open bars) and maximum percent cover (solid bars).
56 Botany Vol. 91, 2013
Published by NRC Research Press
Bot
any
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Geo
rge
Mas
on U
nive
rsity
on
03/2
1/13
For
pers
onal
use
onl
y.
outcrop habitats than in the forest floor habitat (P = 0.0002)(Fig. 4).
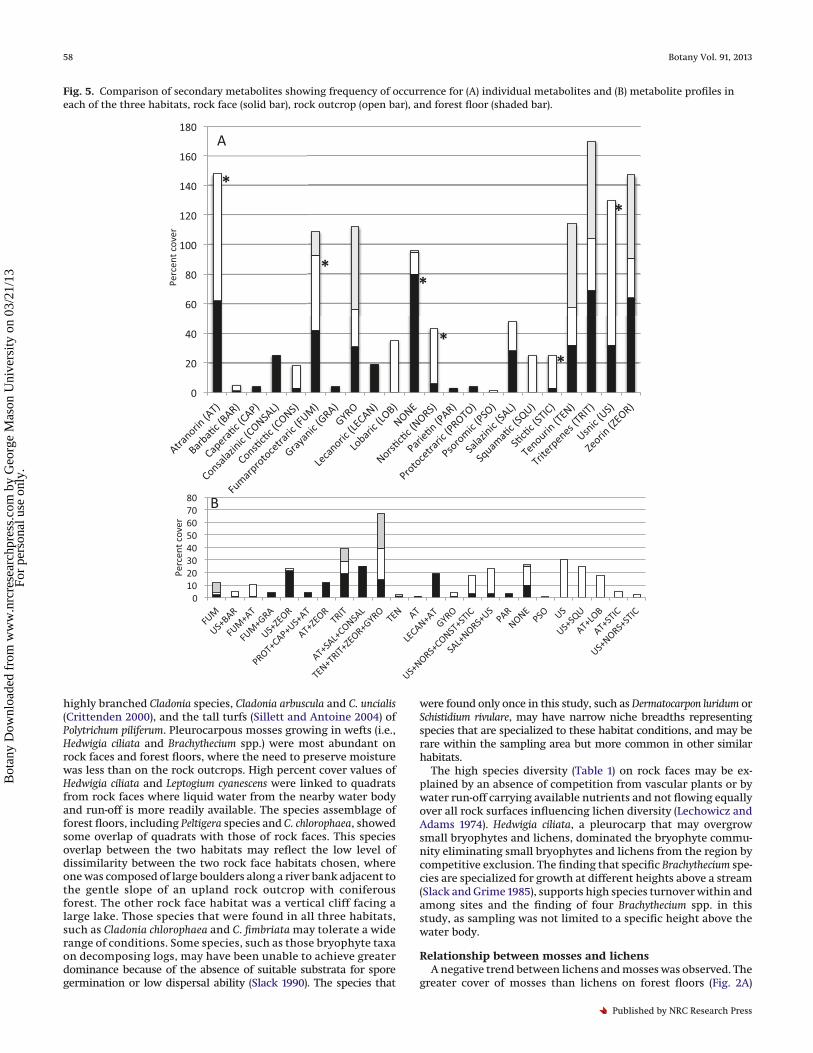
Lichen secondary chemistryTwenty-two different individual secondary metabolites, includ-
ing a category where lichens had no compounds, were identifiedacross the three habitats (Fig. 5A). Twenty-four secondary metab-olite profiles were identified (including a category with “none”)from lichens among the three habitats (Fig. 5B).
To examine whether secondary metabolite profiles can differ-entiate among habitats, a one-way analysis tested the frequencyand percent cover of secondary metabolite profiles among habi-tats. There was significantly lower percent cover of secondarymetabolite profiles in the forest floor habitat than in the othertwo habitats (P = 0.0028), which were not different from eachother. The frequency of occurrence of secondary metabolite pro-files was lowest in the forest floor habitat, but highest in the rockoutcrop (P = 0.0044).
The secondary metabolite profiles were analysed by one-wayANOVA to test for differences in profiles within and among habi-tats to explore potential ecological roles of the profiles (see Fig. 5Afor secondarymetabolite abbreviations). In all habitats combined,the percent cover of TRIT alone and US alone profiles were signif-icantly higher than other profiles (P< 0.0001).When habitats wereexamined separately, the rock face habitat showed that percentcover of TRIT alone, AT + SAL + CONSAL, and the TEN + TRIT +ZEOR + GYRO profiles were significantly higher than other pro-files (P < 0.0001). In the rock outcrop, percent cover of US alone,US + SQU, and TEN + TRIT + ZEOR + GYRO were significantlyhigher than other profiles (P < 0.0001). In the forest floor habitatthe TEN + TRIT + ZEOR + GYRO profile was significantly higherthan other profiles (P < 0.0001).
For the individual secondary metabolites, a PCoA of percentcover showed that quadrats from rock outcrops were clearly sep-arated from the other two habitats (Fig. 3B). Quadrats from rockfaces were overlapping with quadrats from coniferous forestfloors (Fig. 3B). Axis 1 explained 34.3% and axis 2 explained 24.8%of the variation.
A comparison of each individual metabolite among habitats(see Fig. 5A) showed that zeorin was produced in significantlygreatest lichen cover on rock faces. Atranorin, fumarprotocetraricacid, norstictic acid, stictic acid, and usnic acid were produced inthe significantly greatest lichen cover on rock outcrops. The fewcompounds produced by lichens on coniferous forest floors, suchas tenuiorin, triterpenes, and fumarprotocetraric acid, were notproduced by a significantly greater cover of lichens on the forestfloors.
Discussion
Species assemblages as predictors of habitatThis study showed that habitat types could be predicted based
on percent cover of lichens and bryophytes. In the PCoA (Fig. 3A)the species variation in the rock face and the forest floor habitatsmay be explained by the same environmental gradients along axis2. We interpreted this environmental gradient to be a combina-tion of moisture and shade, where species in both habitats werestrongly influenced by moisture and shade but the habitats pre-sented conditions that were sufficient to distinguish among spe-cies assemblages. This pattern contrasted with the speciesassemblage on the rock outcrop habitat where the quadrats arespread out horizontally, suggesting that the variation in the spe-cies of the outcrop may be explained by different environmentalvariables. These environmental variables are interpreted to be dryconditions and high light levels, and the separation is also sup-ported by secondary metabolites (Fig. 3B), suggesting a greaterdifference among rock outcrop and the other two habitats thanbetween rock face and forest floor habitats. The three most abun-dant species on rock outcrops were those whose morphologywould seemingly best conservemoisture and tolerate water stress(Jovan and McCune 2004; Lechowicz and Adams 1974), such as the
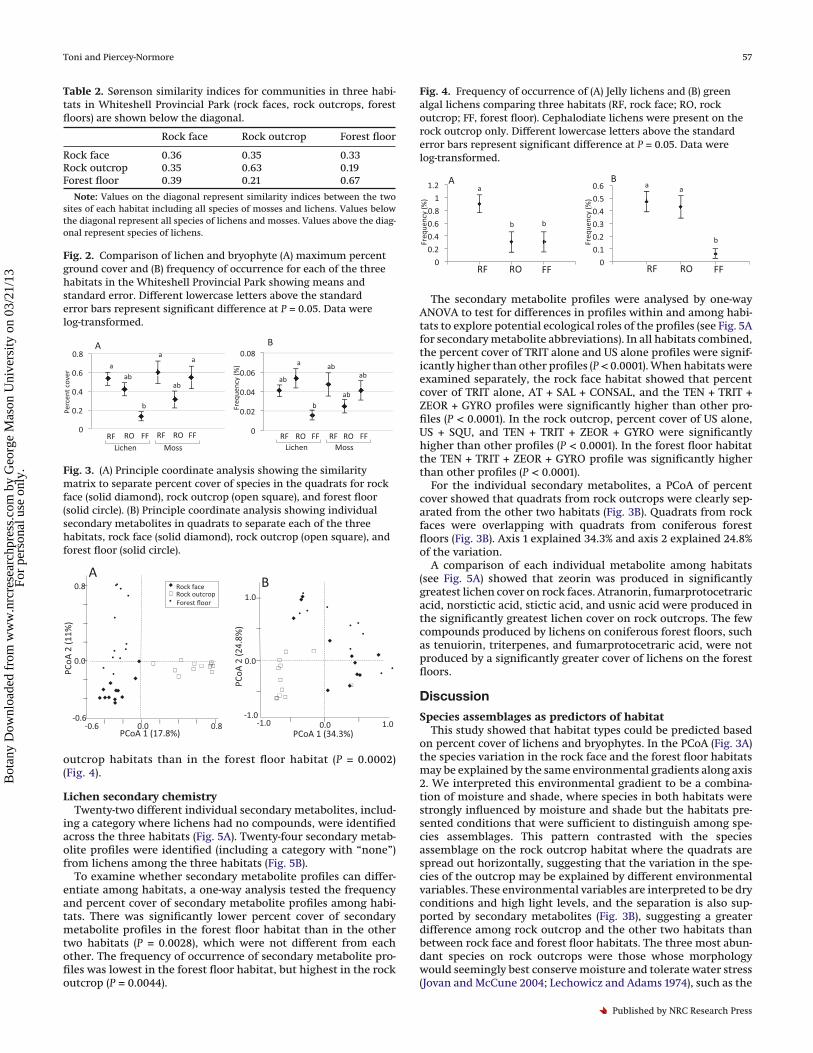
Table 2. Sørenson similarity indices for communities in three habi-tats in Whiteshell Provincial Park (rock faces, rock outcrops, forestfloors) are shown below the diagonal.
Rock face Rock outcrop Forest floor
Rock face 0.36 0.35 0.33Rock outcrop 0.35 0.63 0.19Forest floor 0.39 0.21 0.67
Note: Values on the diagonal represent similarity indices between the twosites of each habitat including all species of mosses and lichens. Values belowthe diagonal represent all species of lichens and mosses. Values above the diag-onal represent species of lichens.
Fig. 2. Comparison of lichen and bryophyte (A) maximum percentground cover and (B) frequency of occurrence for each of the threehabitats in the Whiteshell Provincial Park showing means andstandard error. Different lowercase letters above the standarderror bars represent significant difference at P = 0.05. Data werelog-transformed.
Fig. 3. (A) Principle coordinate analysis showing the similaritymatrix to separate percent cover of species in the quadrats for rockface (solid diamond), rock outcrop (open square), and forest floor(solid circle). (B) Principle coordinate analysis showing individualsecondary metabolites in quadrats to separate each of the threehabitats, rock face (solid diamond), rock outcrop (open square), andforest floor (solid circle).
Fig. 4. Frequency of occurrence of (A) Jelly lichens and (B) greenalgal lichens comparing three habitats (RF, rock face; RO, rockoutcrop; FF, forest floor). Cephalodiate lichens were present on therock outcrop only. Different lowercase letters above the standarderror bars represent significant difference at P = 0.05. Data werelog-transformed.
Toni and Piercey-Normore 57
Published by NRC Research Press
Bot
any
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Geo
rge
Mas
on U
nive
rsity
on
03/2
1/13
For
pers
onal
use
onl
y.
highly branched Cladonia species, Cladonia arbuscula and C. uncialis(Crittenden 2000), and the tall turfs (Sillett and Antoine 2004) ofPolytrichum piliferum. Pleurocarpous mosses growing in wefts (i.e.,Hedwigia ciliata and Brachythecium spp.) were most abundant onrock faces and forest floors, where the need to preserve moisturewas less than on the rock outcrops. High percent cover values ofHedwigia ciliata and Leptogium cyanescens were linked to quadratsfrom rock faces where liquid water from the nearby water bodyand run-off is more readily available. The species assemblage offorest floors, including Peltigera species and C. chlorophaea, showedsome overlap of quadrats with those of rock faces. This speciesoverlap between the two habitats may reflect the low level ofdissimilarity between the two rock face habitats chosen, whereonewas composed of large boulders along a river bank adjacent tothe gentle slope of an upland rock outcrop with coniferousforest. The other rock face habitat was a vertical cliff facing alarge lake. Those species that were found in all three habitats,such as Cladonia chlorophaea and C. fimbriata may tolerate a widerange of conditions. Some species, such as those bryophyte taxaon decomposing logs, may have been unable to achieve greaterdominance because of the absence of suitable substrata for sporegermination or low dispersal ability (Slack 1990). The species that
were found only once in this study, such as Dermatocarpon luridum orSchistidium rivulare, may have narrow niche breadths representingspecies that are specialized to these habitat conditions, and may berare within the sampling area but more common in other similarhabitats.
The high species diversity (Table 1) on rock faces may be ex-plained by an absence of competition from vascular plants or bywater run-off carrying available nutrients and not flowing equallyover all rock surfaces influencing lichen diversity (Lechowicz andAdams 1974). Hedwigia ciliata, a pleurocarp that may overgrowsmall bryophytes and lichens, dominated the bryophyte commu-nity eliminating small bryophytes and lichens from the region bycompetitive exclusion. The finding that specific Brachythecium spe-cies are specialized for growth at different heights above a stream(Slack andGrime 1985), supports high species turnoverwithin andamong sites and the finding of four Brachythecium spp. in thisstudy, as sampling was not limited to a specific height above thewater body.
Relationship between mosses and lichensAnegative trend between lichens andmosses was observed. The
greater cover of mosses than lichens on forest floors (Fig. 2A)
Fig. 5. Comparison of secondary metabolites showing frequency of occurrence for (A) individual metabolites and (B) metabolite profiles ineach of the three habitats, rock face (solid bar), rock outcrop (open bar), and forest floor (shaded bar).
58 Botany Vol. 91, 2013
Published by NRC Research Press
Bot
any
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Geo
rge
Mas
on U
nive
rsity
on
03/2
1/13
For
pers
onal
use
onl
y.

where only 8 of 21 cryptogams are lichens, is supported by thefinding that bryophyte growth is favoured by shaded forest floors(Lambert and Maycock 1968). Lambert and Maycock (1968) alsosuggested that suitable substrata, light availability, and the turn-over of litter and organic matter, would influence the speciescomposition in forests. There were 17 lichen species common toboth Lambert and Maycock (1968) and this study, perhaps result-ing from the more easterly geographic location of the study byLambert and Maycock (1968). On the other hand the lichens andmosses on exposed rock outcrops did not show a significant dif-ference even though lichens are considered to be better compet-itors than bryophytes under high light levels and low moistureconditions (Lambert and Maycock 1968). Variation in microhabi-tat of the rock outcrops studied may partially explain this resultbecause foliose lichens, such as Xanthoparmelia spp., are replaced,as soil accumulates, with branched Cladonia species (Ahti andOksanen 1990). Amosaic ofmicrohabitatsmay have been present,and both genera were present in this habitat (Fig. 1).
Lichens with cyanobacterial partners can tolerate low light lev-els andwould be expected to be foundwith byrophytes in habitatssupporting these conditions. Lichens with cyanobacterial part-ners were the most abundant lichens in shady, relatively moisthabitats such as forest floors and rock faces. High moisture levelsand low levels of wind exposure played an important role in thelower rate of desiccation (Sillett and Antoine 2004) in the forestfloor habitat where the cover of green algal lichens was signifi-cantly lower than in other habitats. The six species of Peltigerafound in this study on the forest floor have cyanobacteria as theirprimary photobiont, and represented two-thirds of the encoun-tered lichen species. The jelly lichens, with a homiomerous thal-lus, were significantly higher on the rock faces than on theoutcrops or the forest floor. Thismay reflect the presence of liquidwater requirement for photosynthesis by cyanobacterial lichens(Büdel and Scheidegger 2008), variable substrata, and the lengthof time the habitat has been available for colonization. Highmois-ture is generally linked to less exposed conditions in which evap-oration is reduced. The cyanobacterial lichen species on rock faceswere a more diverse community than that found on forest floorsand included eight Peltigera species, two Leptogium species, oneNephroma species, and one Collema species. Cephalodiate lichenssuch as Stereocaulon spp. were reported only on the rock outcrops,reflecting an adaptation to drier conditions, and early invasion ofrock substratum by S. saxatile, by having green algae as their pri-mary photobiont. The presence of noncephalodiate Peltigera spp.in all three habitats may suggest that the heteromerous thalluscondition can tolerate a wide range of environmental conditions(Büdel and Scheidegger 2008). Some Peltigera species can develop athin cortex with a thick epinecral layer above the cortex whengrowing in high light and low moisture conditions (Büdel andScheidegger 2008). The homiomerous thallus of Leptogium wasonly present in the rock face habitats wheremore liquidmoisturewas available.
Lichen secondary metabolites reflect habitatSecondary metabolite production using individual metabolites
was sufficient to separate rock outcrops from the other two hab-itats (Fig. 6). The significantly greater cover of lichens on rockoutcrops reported to contain the cortical compound usnic acidcan be explained by the correlations between usnic acid and theintensity of UV-B radiation (Bjerke et al. 2002; McEvoy et al. 2006;Rundel 1969) and seasonal changes (Bjerke et al. 2005), or as amechanism to facilitate lichen adaptation to acidic substrata,such as granitic rock (Hauck et al. 2009). The occurrence of an-other cortical compound, atranorin, in both outcrops and rockfaces (Figs. 4 and 5) is consistent with its proposed light regulatoryfunction (Rao and LeBlanc 1965) and the degree of exposure of thehabitat (Solhaug et al. 2010). In a comparison of different habitats,Waring (2008) showed that UV-screening metabolites were more
highly diverse in open pastures than interior forests. Light hasbeen shown to influence production of secondary metabolitesthrough the velvet family of proteins in nonlichenized fungi(Sprote and Brackhage 2007), and these proteins are thought to beconserved among fungi (Dreyer et al. 2007; Kumar et al. 2010).There have been a number of studies on the effect of light onproduction of secondary metabolites, but fewer studies on effectsof other environmental parameters on secondary metabolite pro-duction. In the current study, squamatic acid was produced onlyby lichens growing on exposed rock outcrops, linking its presenceto the driest and most exposed to high light levels of the threehabitats. The larger number of metabolites reported for rock out-crops (Fig. 4) is consistent with the hypothesis that drier condi-tions trigger production of lichen secondary metabolites(Culberson and Armaleo 1992; Stocker-Wörgötter 2001), eventhough these studies were based on culture experiments that in-directly support our findings.
The higher abundance of US alone and TRIT alone over all hab-itats suggests these metabolites may have multiple functions inthe habitat. On the other hand, the low overall levels of profiles inthe forest floor habitat may suggest that they are not neededin that habitat or there is no environmental trigger to induce theirexpression. The higher abundance and cover of TRIT alone andTRIT + TEN + ZEOR +GYROprofiles in the rock face and forest floorhabitats may suggest a link with lower light and higher moisterlevels than on the outcrops. Shaded habitat caused an increase inrhizome production in a plant (Actaea racemosa) that in turn re-sulted in production of higher levels of triterpenes (McCoy et al.2007), but the environmental trigger for production of the triter-penes was not examined. Zeorin is thought to have an antiher-bivory role (Nimis and Skert 2006) but the connection with highmoisture or low light levels is not clear. Since some metabolites,such as usnic acid, may have a myriad of functions serving as alight screen (McEvoy et al. 2006) in herbivore defense (Lawrey1986), regulation of photobiont membrane permeability (Backoret al. 1998), promoting intracellular uptake of copper (Hauck et al.2009), and as an antibiotic or antimicrobial agent (Rundel 1978),the production of usnic acid may be in response to any one of anumber of demands explaining the abundance of usnic acid intwo of the three habitats. The profiles, TRIT alone and TRIT + TEN +ZEOR+GYRO,mayalsoplay similar antifungal or antimicrobial roles(Nimis and Skert 2006).
Varying levels of secondary metabolite production by lichenthalli will also have an effect on the environment in distributionof allelopathic compounds influencing competition among cryp-togams as well as germination and growth of vascular plants(Armstrong andWelch 2007; Lawrey 1995), Collembolans and Dip-terans (Miller et al. 2008), and the growth of intrathalline bacterialcommunities (Bates et al. 2011), which have been shown to varyacross a lichen thallus (Mushegian et al. 2011). The spatial config-uration of noncryptogam communities may be shaped at smalland large spatial scales depending on the levels and types of sec-
Fig. 6. Comparison of secondary metabolite profiles across habitattypes, rock face (RF), rock outcrop (RO), and forest floor (FF) for eachof (A) maximum percent cover and (B) frequency of occurrence ofeach profile. Different lowercase letters above the standard errorbars represent significant difference at P = 0.05. Data were log-transformed.
Toni and Piercey-Normore 59
Published by NRC Research Press
Bot
any
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Geo
rge
Mas
on U
nive
rsity
on
03/2
1/13
For
pers
onal
use
onl
y.

ondary metabolites induced by abiotic conditions. Secondary me-tabolites have even been suggested to have an effect on cariboudigestibility of winter lichen forage (Palo 1983) and their ability todetect lichens beneath snow cover (Skogland 1978; Brown andTheberge 1990).
The finding that multiple lichen genera were involved in theproduction of the same compounds, such as usnic acid producedby some Cladonia spp. and Xanthoparmelia spp., or the triterpenesproduced by several Peltigera spp., or atranorin inmultiple speciesof Physcia, Stereocaulon, Parmelia, and Heterodermia among habitatsmay suggest that secondary metabolite production is plastic (un-der environmental control) over evolutionary time. Just as lichen-ization has been shown to have multiple evolutionary origins(Gargas et al. 1995; Lutzoni et al. 2001), the production of second-ary metabolites may also have multiple evolutionary origins, de-spite chemical profiles having been linked with lichen taxonomy(Hawksworth 1976). Multiple evolutionary origins of secondarymetabolites may be supported by reports of large numbers ofpolyketide synthase gene paralogs and low numbers of polyketideproducts reported for fungi (Deduke et al. 2012 and referencestherein). The presence of large numbers of gene paralogs mightallow certain types or quantities of secondary metabolites to beproduced in one habitat and others to be produced in habitatswith different conditions, while preserving the genetic versatilityor the adaptive nature of secondary metabolites. Studies areslowly accumulating that link the production of a secondary me-tabolite with a particular suite of environmental conditions(Culberson and Armaleo 1992; Lechowicz and Adams 1974; McEvoyet al. 2006; Nybakken et al. 2007; Stocker-Worgotter 2001). Thesestudies arenecessary tounderstand the genetic regulationof second-ary metabolites following the proposed matrix of environmentaland developmental cues described for nonlichenized fungi (Fox andHowlett 2008) and how they might be relevant to lichenized fungi.
In conclusion, this study showed strong relationships amongspecies assemblages and secondarymetabolites with types of hab-itat. We surmise that a complex interaction may exist amongdegree of exposure, moisture levels, and light levels playing asignificant role in species preference and secondary metaboliteproduction. While the relationship between habitat and speciesconfirms results from previous studies, it also provides baselinedata for habitat-specific lists of species and their abundance in theWhiteshell Provincial Park. However, amore significant finding isthat this study has provided one of the first studies on chemicalecology of lichens and the correlation of secondary metaboliteswith habitat types. Secondary compound production differedstrongly among habitats in terms of overall abundance and spe-cific metabolites, potentially owing to the ecological roles ofmany compounds. Exploration of the abundance of individualmetabolites and profiles of metabolites revealed a correlationwith usnic acid, it also provided a correlation with the triterpenesand relatedmetabolites. While functions such as tolerating acidicsubstrata, screening out excess light levels, and herbivore defensemay be adaptive under varying environmental conditions, theenvironment may also constrain metabolite production leadingto different quantities of metabolites produced or the completeabsence of metabolites. Further research on secondary metabo-lites and the regulation of their production in different habitatswill shed light on the interaction between environmental regula-tion and adaptation.
AcknowledgmentsThe authors thank J. Roth and R. Lastre (University ofManitoba),
R. Belland (University of Alberta) for help with the statistical anal-yses, and C. Deduke (University of Manitoba) for field assistance.Financial support for the study was received by Natural Sciencesand Engineering Research Council of Canada (NSERC).
ReferencesAhti, T., and Oksanen, J. 1990. Epigeic lichen communities of taiga and tundra
regions. Vegetatio, 86: 39–70. doi:10.1007/BF00045134.Anderson, L., Crum, H., and Buck,W.R. 1990. List of themosses of North America
North of Mexico. Bryologist, 93(4): 448–499. doi:10.2307/3243611.Armstrong, R.A., and Welch, A.R. 2007. Competition in lichen communities.
Symbiosis, 43: 1–12.Backor, M., Hudák, J., Repcák, M., Zigerlers, W., and Backorová, M. 1998. The
influence of pH and lichen metabolites (vulpinic acid and (+) usnic acid) onthe growth of the lichen photobiont Trebouxia irregularis. Lichenologist, 30(6):577–582. doi:10.1017/S0024282992000574.
Bates, J.W. 2009. Mineral nutrition and substratum ecology. In Bryophyte biol-ogy. Edited by B. Goffinet and A.J. Shaw. Cambridge University Press, Cam-bridge.
Bates, T.B., Cropsey, G., Caparaso, J., Knight, R., and Fierer, N. 2011. Bacterialcommunities associated with the lichen symbiosis. Appl. Environ. Microbiol.77(4): 1309–1314. doi:10.1128/AEM.02257-10. PMID:21169444.
Bjerke, J.W., Lerfall, K., and Elvebakk, A. 2002. Effects of ultraviolet radiationand PAR on the content of usnic and divaricatic acids in two arctic-alpinelichens. Photochem. Photobiol. Sci. 1: 678–685. doi:10.1039/b203399b. PMID:12665305.
Bjerke, J.W., Elvebakk, A., Dominguez, B., and Dahlback, A. 2005. Seasonaltrends in usnic acid concentrations of Arctic, alpine, and Patagonian popu-lations of the lichen Flavocetraria nivalis. Phytochemistry, 66: 337–344. doi:10.1016/j.phytochem.2004.12.007. PMID:15680990.
Brodo, I., Sharnoff, S.D., and Sharnoff, S. 2001. Lichens of North America. YaleUniversity Press, New Haven and London.
Brown, W.K., and Theberge, J.B. 1990. The effect of extreme snowcover onfeeding-site selection by woodland caribou. J. Wildl. Manage. 54: 161–168.
Büdel, B., and Scheidegger, C. 2008. Thallus morphology and anatomy. In Lichenbiology. 2nd ed. Edited by T.H. Nash, III. Cambridge University Press, Cam-bridge. pp. 40–68.
Crittenden, P.D. 2000. Aspects of the ecology of mat-forming lichens. Rangifer,20(2–3): 127–139.
Crum, H. 1983. Mosses of the Great Lakes Forest. University Herbarium, Univer-sity of Michigan, Ann Arbor.
Crum, H.A., and Anderson, L.E. 1981. Mosses of eastern North America. Vols. Iand II. Columbia University Press, New York.
Culberson, C.F. 1972. Improved conditions and new data for the identification oflichen products by a standardized thin-layer chromatographic method. J.Chromatogr. 72: 113–125. doi:10.1016/0021-9673(72)80013-X. PMID:5072880.
Culberson, C.F., and Armaleo, D. 1992. Induction of a complete secondary-product pathway in a cultured lichen fungus. Exp. Mycol. 16(1): 52–63.
Culberson, C.F., Culberson, W.L., and Arwood, D.A. 1977. Physiography and fu-marprotocetraric acid production in the Cladonia chlorophaea group in NorthCarolina. Bryologist, 80: 71–75. doi:10.2307/3242511.
Culberson, W.L., and Culberson, C.F. 1967. Habitat selection by chemically dif-ferentiated races of lichens. Science, 158(3805): 1195–1197. doi:10.1126/science.158.3805.1195. PMID:6057292.
Deduke, C., Timsina, B., and Piercey-Normore, M.D. 2012. Effect of environmen-tal change on secondary metabolite production in lichen-forming fungi. InInternational perspectives on global environmental change. Edited byStephen Young. Available at http://www.intechopen.com/books/international-perspectives-on-global-environmental-change/effect-of-environmental-change-on-secondary-metabolite-production-in-lichen-forming-fungi. [ISBN: 978-953-307-815-1.]
Dreyer, J., Eichhorn, H., Friedlin, E., Kurnsteiner, H., and Kuck, U. 2007. A ho-mologue of the Aspergillus velvet gene regulates both cephalosporin C biosyn-thesis and hyphal fragmentation in Acremonium chrysogenum. Appl. Environ.Microbiol. 73: 3412–3422.
Esslinger, T.L. 2010. A cumulative checklist for the lichen-forming, lichenicolousand allied fungi of the continental United States and Canada. North DakotaState University, Fargo. Available at http://www.ndsu.nodak.edu/instruct/~esslinge/chcklst/chcklst7-8.htm [accessed 25 April 2011].
Fox, E.M., and Howlett, B.J. 2008. Secondary metabolism: regulation and role infungal biology. Curr. Opin. Microbiol. 11: 481–487. doi:10.1016/j.mib.2008.10.007. PMID:18973828.
Gargas, A., DePriest, P.T., Grube, M., and Tehler, A. 1995. Multiple origins oflichen symbioses in fungi suggested by SSU rDNA phylogeny. Science,268(5216): 1492–1495. doi:10.1126/science.7770775. PMID:7770775.
Goward, T. 1999. Lichens of British Columbia. Part II: Fruticose species. Ministryof Forests Research Program, British Columbia.
Hassel, K., Pedersen, B., and Söderström, L. 2005. Changes in life-history traits inan expanding moss species: phenotypic plasticity or genetic differentiation?A reciprocal transplantation experiment with Pogonatum dentatum. Ecogra-phy, 28: 71–80.
Hauck, M. 2011. Eutrophication threatens the biochemical diversity in lichens.Lichenologist, 43: 147–154. doi:10.1017/S0024282910000654.
Hauck, M., Jürgens, S.-R., Willenbruch, K., Huneck, S., and Leuschner, C. 2009.Dissociation and metal-binding characteristics of yellow lichen substancessuggest a relationship with site preferences of lichens. Ann. Bot. 103: 13–22.PMID:18977765.
Hawksworth, D.L. 1976. Lichen chemotaxonomy. In The Systematics Association
60 Botany Vol. 91, 2013
Published by NRC Research Press
Bot
any
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Geo
rge
Mas
on U
nive
rsity
on
03/2
1/13
For
pers
onal
use
onl
y.

Special Vol. 8. Edited by D.H. Brown, D.L. Hawksworth, and R.H. Bailey. Aca-demic Press, London and New York. pp. 139–184.
Hinds, J.W., and Hinds, P.L. 2007. The macrolichens of New England. The NewYork Botanical Garden Press, Bronx.
Huneck, J. 1999. The significance of lichens and their metabolites. Naturwissen-schaften, 86: 559–570. doi:10.1007/s001140050676. PMID:10643590.
Johnson, E.A. 1981. Vegetation organization and dynamics of lichen woodlandcommunities in the Northwest Territories. Ecology, 62(1): 200–215. doi:10.2307/1936682.
Jovan, S., and McCune, B. 2004. Regional variation in epiphytic macrolichencommunities in northern and central California forests. Bryologist, 107: 328–339. doi:10.1639/0007-2745(2004)107[0328:RVIEMC]2.0.CO;2.
Kenkel, N.C. 2006. On selecting an appropriate multivariate analysis. Can. J.Plant Sci. 86(3): 663–676. doi:10.4141/P05-164.
Kumar, L., Breakspear, A., Kistler, C., Ma, L., and Xie, X. 2010. Systematic discov-ery of regulatory motifs in Fusarium graminiarum by comparing four Fusariumgenomes. BMC Genomics, 11: 208. doi:10.1186/1471-2164-11-208. PMID:20346147.
Kunkel, G. 1980. Microhabitat and structural variation in the Aspicilia desertorumgroup (lichenized Ascomycetes). Am. J. Bot. 67: 1137–1144. doi:10.2307/2442357.
Lambert, J.D.H., andMaycock, P.F. 1968. The ecology of terricolous lichens of thenorthern conifer–hardwood forests of central Eastern Canada. Can. J. Bot.46(8): 1043–1078. doi:10.1139/b68-138.
Lawrey, J.D. 1986. Biological role of lichen substances. Bryologist, 89(2): 111–122.doi:10.2307/3242751.
Lawrey, J.D. 1995. Lichen allelopathy: a review. In Allelopathy: organisms, pro-cesses, and applications. Edited by M.M. Inderjit, K. Dakshini, andF.A. Einhellig. ACS Symposium Series, Vol. 582. American Chemical Society.pp. 26–38.
Lechowicz, M.J., and Adams,M.S. 1974. Ecology of Cladonia lichens. I. Preliminaryassessment of the ecology of terricolous lichen–moss communities in On-tario and Wisconsin. Can. J. Bot. 52(1): 55–64.
Lutzoni, F., Pagel, M., and Reeb, V. 2001. Major fungal lineages are derived fromlichen symbiotic ancestors. Nature, 411: 937–940. doi:10.1038/35082053.PMID:11418855.
McCoy, J.A., Davis, J., Camper, N., Kahn, I., and Bharathi, A. 2007. Influence ofrhizome propagule size on yields and triterpene glycoside concentrations onBlack Cohosh [Actaea racemosa L. syn. Cimicifuga racemosa (L.) Nuttal]. Hort-Science, 42(1): 61–64.
McCune, B., and Grace, J.B. 2002. Analysis of ecological communities. MjM Soft-ware Design, Geleneden Beach, Oregon, U.S.A.
McEvoy, M., Nybakken, L., Solhaug, K.A., and Gauslaa, Y. 2006. UV triggers thesynthesis of the widely distributed secondary lichen compound usnic acid.Mycological Progress. 5: 221–229. doi:10.1007/s11557-006-0514-9.
Miller, K., Wagner, R., andWoods, S. 2008. Arboreal arthropod associations withepiphytes following gap harvesting in the Acadian forest of Maine. Bryolo-gist, 111(3): 424–434. doi:10.1639/0007-2745(2008)111[424:AAAWEF]2.0.CO;2.
Mushegian, A., Peterson, C., Baker, C., and Pringle, A. 2011. Bacterial diversityacross individual lichens. Appl. Environ. Microbiol. 77(12): 4249–4252. doi:10.1128/AEM.02850-10. PMID:21531831.
Nash, T.H., III (Editor). 2008. Lichen sensitivity to air pollution. In Lichen biology.2nd ed. Cambridge University Press, Cambridge. pp. 299–314.
Nimis, P.L., and Skert, N. 2006. Lichen chemistry and selective grazing by thecoleopteran Lasioderma serricorne. Environ. Exp. Bot. 55: 175–182. doi:10.1016/j.envexpbot.2004.10.011.
Nybakken, L., Asplund, J., Solhaug, K., and Gauslaa, Y. 2007. Forest successionalstage affects the cortical secondary chemistry of three old forest lichens.J. Chem. Ecol. 33: 1607–1618. doi:10.1007/s10886-007-9339-5. PMID:17636451.
Orange, A., James, P.W., and White, F.J. 2001. Microchemical methods for theidentification of lichens. British Lichen Society.
Palo, R.T. 1983. Usnic acid, a secondary metabolite of lichens and its effect on invitro digestibility in reindeer. Rangifer, 13(1): 39–43.
Rao, D.N., and LeBlanc, F. 1965. A possible role of atranorin in the lichen thallus.Bryologist, 68(3): 284–289. doi:10.2307/3240598.
Redfearn, P.L., Jr. 1979. How to know themosses and liverworts. Revised ed.WCBMcGraw-Hill, USA.
Rikkinen, J. 1997. Habitat shifts and morphological variation of Pseudeverniafurfuracea along a topographical gradient. Symb. Bot. UPS. 32: 223–245.
Rundel, P.W. 1978. The ecological role of secondary lichen substances. Biochem.Syst. Ecol. 6: 157–170. doi:10.1016/0305-1978(78)90002-9.
Seaward, M.R.D. 2008. Environmental role of lichens. In Lichen biology. 2nd ed.Edited by T.H. Nash III. Cambridge University Press, Cambridge. pp. 274–298.
Shaw, J. 1985. The relevance of ecology to species concepts in bryophytes. Bry-ologist, 88: 199–206. doi:10.2307/3243029.
Sillett, S.C., and Antoine, M.E. 2004. Lichens and bryophytes in forest canopies.In Forest canopies. Edited byM.D. Lowman andH.B. Rinker. Elsevier AcademicPress.
Skogland, T. 1978. Characteristics of the snow cover and its relationship to wildmountain reindeer (Rangifer tarandus tarandus L.) feeding strategies. Arct. Alp.Res. 10: 569–580.
Slack, N.G. 1990. Bryophytes and ecological niche theory. Bot. J. Linn. Soc. 104:187–213. doi:10.1111/j.1095-8339.1990.tb02218.x.
Slack, N.G., and Grime, J.M. 1985. Niche relationships of mountain stream bryo-phytes. Bryologist, 88: 7–18. doi:10.2307/3242643.
Solhaug, K.A., Larsson, P., and Gauslaa, Y. 2010. Light screening in lichen corticescan be quantified by chlorophyll fluorescence techniques for both reflectingand absorbing pigments. Planta, 231: 1003–1011. doi:10.1007/s00425-010-1103-3. PMID:20135325.
Sprote, P., and Brackhage, A.A. 2007. The light dependent regulator velvet A ofAspergillus nidulans acts as a repressor of the penicillin biosynthesis. Arch.Microbiol. 188: 69–79.
Stocker-Wörgötter, E. 2001. Experimental lichenology and microbiology of li-chens: culture experiments, secondary chemistry of cultured mycobionts,resynthesis, and thallus morphogenesis. Bryologist, 104(4): 576–581. doi:10.1639/0007-2745(2001)104[0576:ELAMOL]2.0.CO;2.
Stringer, M.H.L., and Stringer, P.W. 1973. Studies on the bryophytes of southernManitoba. V. Collections from Whiteshell Provincial Park. Can. Field. Nat.87(4): 419–425.
Waring, B. 2008. Light exposure affects secondary compound diversity in lichencommunities in Monteverde, Costa Rica. PennScience, 6(2): 11–13.
Toni and Piercey-Normore 61
Published by NRC Research Press
Bot
any
Dow
nloa
ded
from
ww
w.n
rcre
sear
chpr
ess.
com
by
Geo
rge
Mas
on U
nive
rsity
on
03/2
1/13
For
pers
onal
use
onl
y.





























