Embed Size (px)
Citation preview

Chemical reaction in cells are often coupled
Glucose + Pi Glucose-6-phosphate
(requires input of energy, endergonic)
ATP ADP + Pi
(releases energy, exergonic)
Reaction 1-
Reaction 2-
Reaction 1 + Reaction 2-Glucose + ATP Glucose-6-phosphate + ADP
(releases energy, exergonic)

Biochemical reactions are generally parts of metabolic pathways
Each step may not be energetically favorable, but the overall pathway must be exergonic
Biochemical reactions are generally parts of metabolic pathways
Each step may not be energetically favorable, but the overall pathway must be exergonic
Most small molecules are synthesized using such pathways

Enzymes are tightly regulated

Cells respond to their environment through the enzymes
Isoleucine starvation
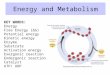
Living organisms are broadly divided into two categories
Eukaryotes(contain a nucleus)
and
Bacteria(no nucleus)
~50 m

Three domains of living organisms

Bacteria lack a nuclear membrane and are relatively simple
Cell envelope: Contains cell walland membrane(s)
Gram positive:one membrane

Eukaryotic cells are much more complex
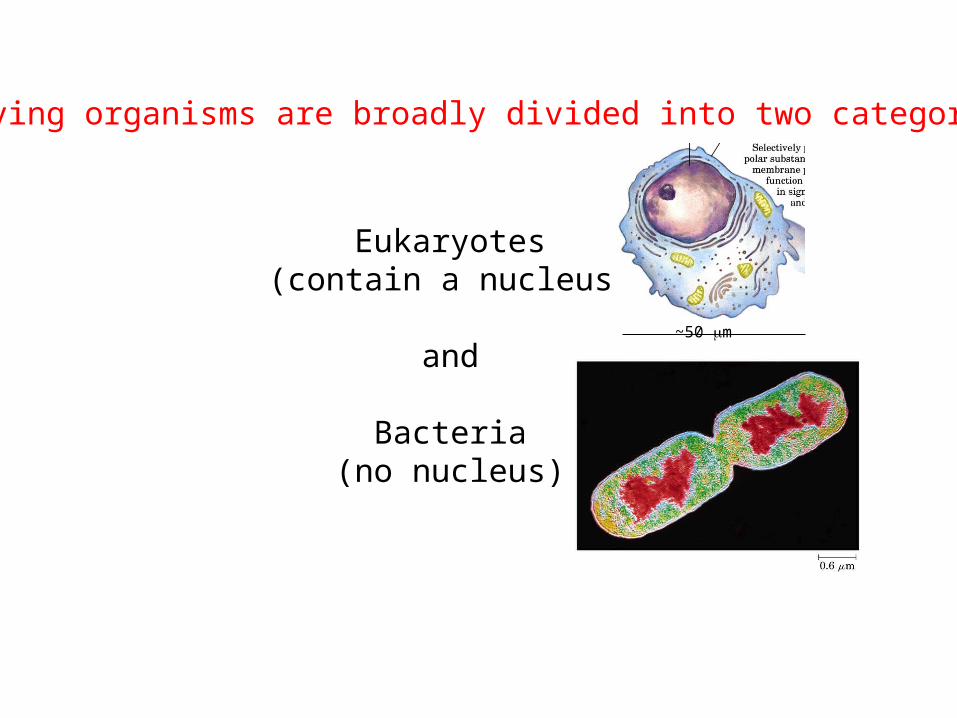
A hydrogen bond is formed through electrical interaction between an atom covalently bonded with a hydrogen and which is more electronegative than hydrogen, and another atom that has a partial negative charge.

Hydrogen bonds can also form between other functional groups

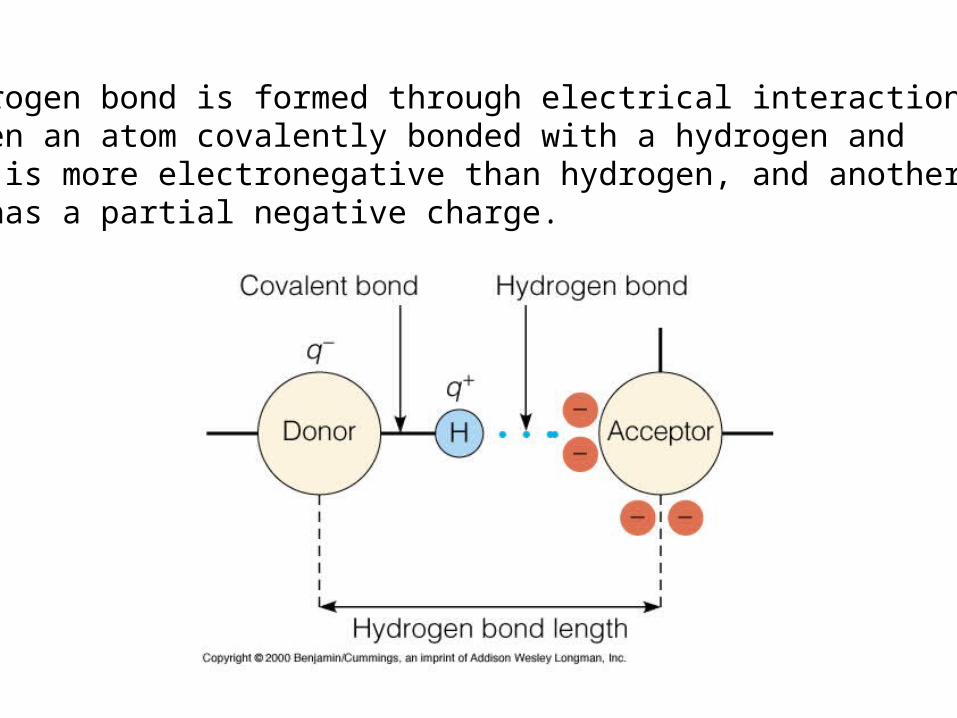
Despite their diversity, all proteins are made from 20 commonmonomeric units called amino acids
Amino acids are carboxylic acids CO
OH
H
2
and contain an amino group

(Uncharged)

(Uncharged)




Phe
UV absorption

Cysteine can exist in two forms
Oxidation
Reduction


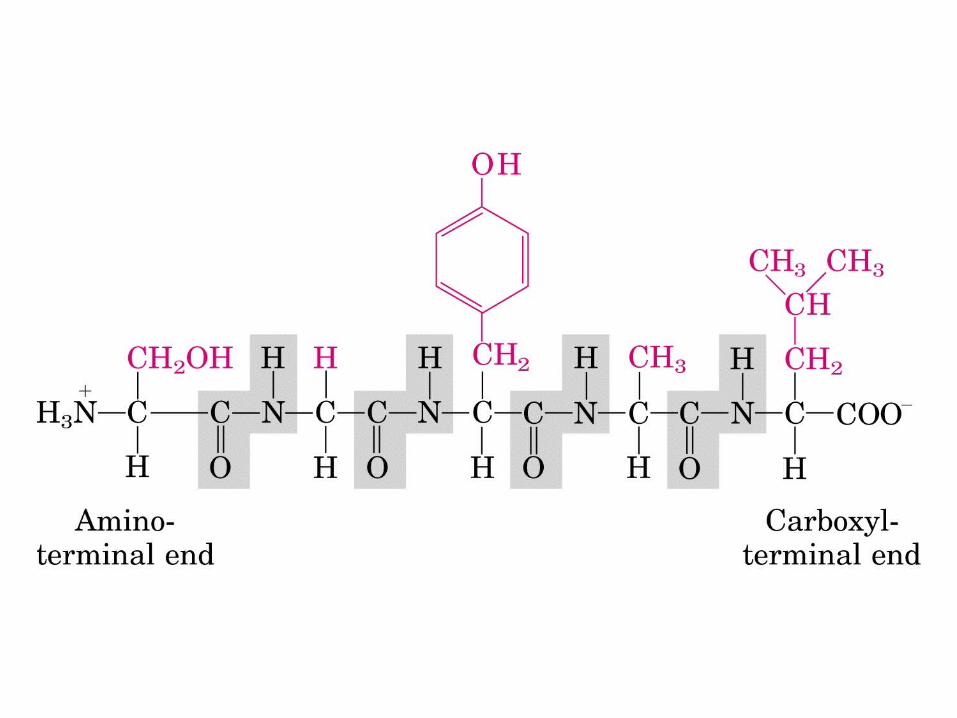

There is delocalization of electrons along the peptide bond
This creates, 1. An electric dipole moment within the peptide unit
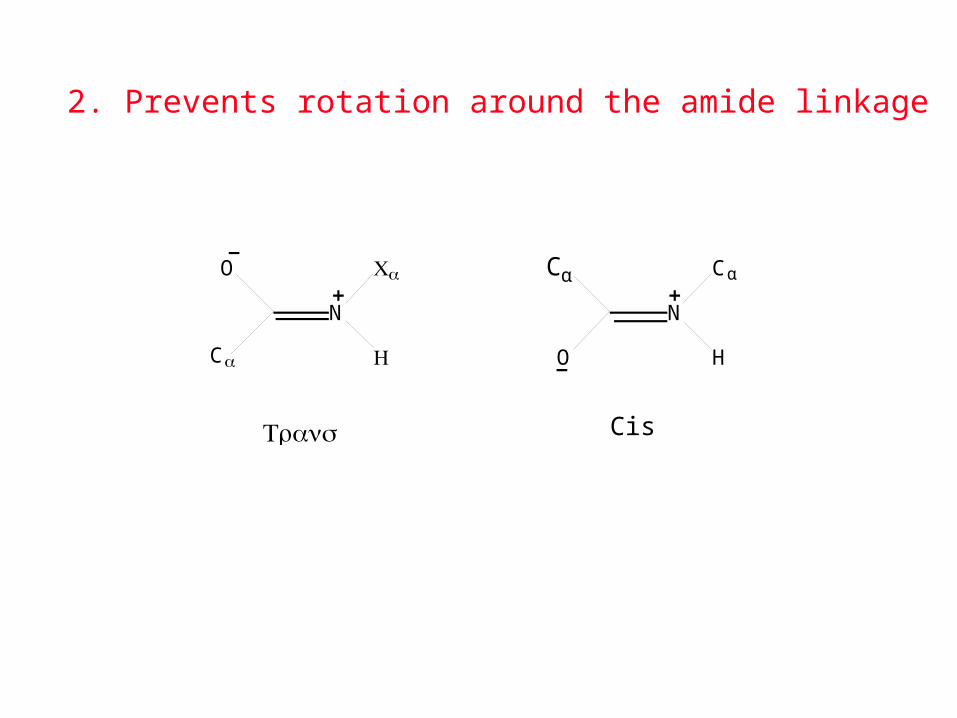
O
N
Cα H
Cα
Trans
Cα
N
O H
Cα
Cis
2. Prevents rotation around the amide linkage

3. Makes each Cα-CO-NH-Cα flat

Proline does not have a strong preference for the trans configuration
4:1

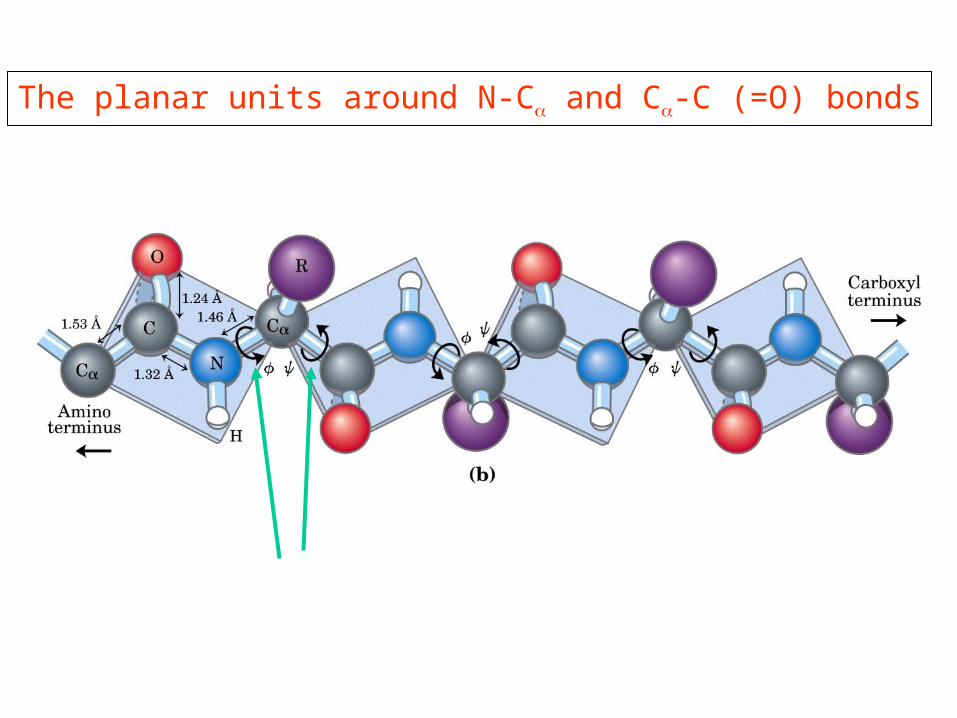
The planar units around N-Cα and Cα-C (=O) bonds

Ramachandran Plot
Only certain combinations of angles are “allowed”

Regular structures in proteins α-helix
=On•••H-Nn+4-

1
2
3 4
5
67
8
9
10

R-groups are on the outside of the helix
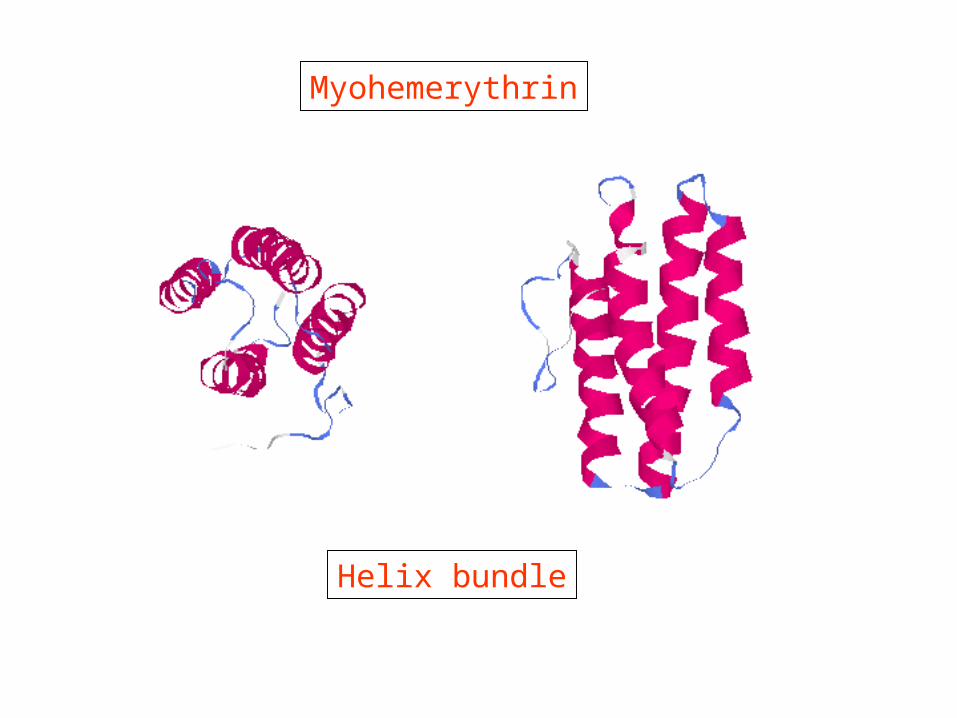
Myohemerythrin
Helix bundle

-sheet
R-groupsabove andbelow
-strand

-sheet
R-groupsabove andbelow
-strand
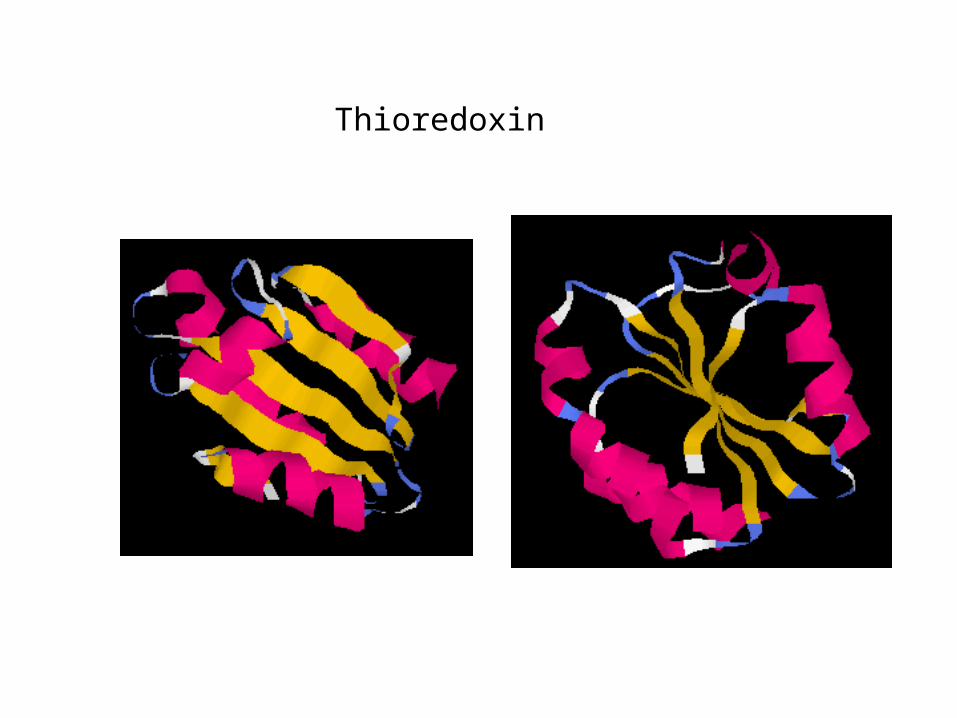
Thioredoxin

Overall folding of the protein

The presence of disulfides reduces the number of alternate structures

Pot
enti
al e
nerg
yP
oten
tial
ene
rgy

Most enzymes are proteins
The exceptions are a handful RNA molecules that can act as enzymes
They are called ribozymes

Consequently, the rate of an enzymatic reaction is highestwhen the enzyme is “saturated”
[E] is constant
Rat
e
Maximumvelocity

Enzymes are often aided by metals
Enzymes that use metals (other than Na+ and K+) are calledmetalloenzymes

…...and by coenzymes

Energetics of enzyme action
S (substrate) P (product)E (enzyme)
Keq =[P][S]
Keq is equillibrium constant
(Molar concentrations)

Keq =[P][S]
Keq is equillibrium constant
ΔG' 0 =−RT lnKeq'
G is Gibbs free energy, G’0 is change in Gibbs free energyunder the following conditions: Temp 37°C (298 °K), pH 7.0,1M reactantsR is Gas constant (8.3 J/mol or 2 Cal/mol)
All but the last condition is always satisfiedWe may drop the superscript “’” and “0”, but it is always assumedto be present
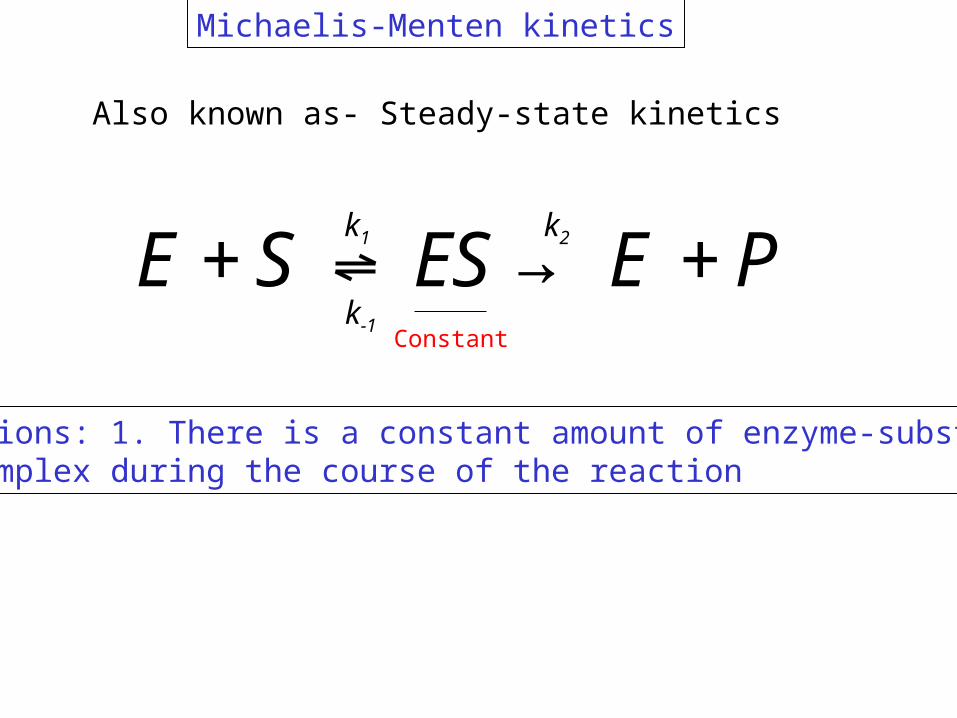
E+S↔ ES→ E +Pk1
k-1
k2
Michaelis-Menten kinetics
Assumptions: 1. There is a constant amount of enzyme-substrate complex during the course of the reaction
Also known as- Steady-state kinetics
Constant
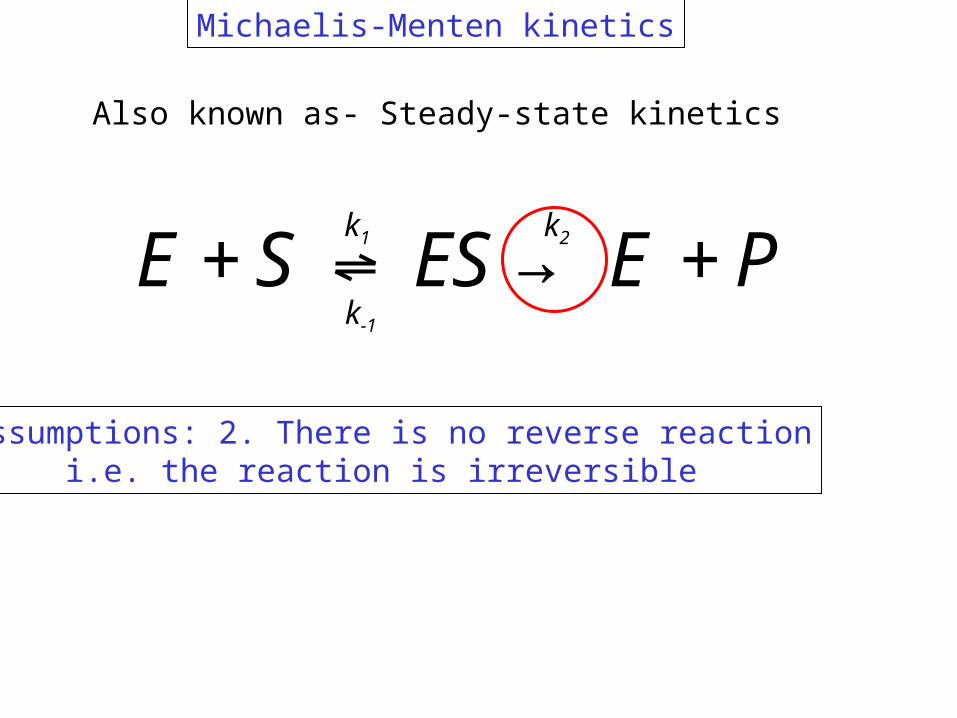
E+S↔ ES→ E +Pk1
k-1
k2
Michaelis-Menten kinetics
Assumptions: 2. There is no reverse reactioni.e. the reaction is irreversible
Also known as- Steady-state kinetics

If you multiply both sides of the equation with k2 then...
k2[ES]=k2[ET ][S]Km +[S]
k2[ES] = v (velocity of the reaction)k2[ET] = k2[ETS] = vmax (the maximum velocity of the reaction)
∴ v=vmax[S]Km +[S]
Michaelis-Menten Equation

Turnover number (kcat) is defined as
kcat = vmax/[ET]
This defines the rate of catalysis per enzyme molecule
Turnover number (kcat) is defined as
kcat = vmax/[ET]
This defines the rate of catalysis per enzyme molecule
Specificity constant is defined as
kcat/Km

Types of enzyme inhibition
1. Competitive inhibition

Types of enzyme inhibition2. Non-competitive inhibition

Types of enzyme inhibition3. Irreversible inhibition

Other ways of regulating enzymes
1. Allosteric control
An enzyme is said to be regulated allosterically when a molecule binds to a site other than the active site and modulates enzyme activity
2. Covalent modifications
Enzymes are often (reversibly) covalently modified to changethere activity
3. Proteolytic cleavage
Some enzymes are activated by proteolytic cleavage

Schematic view of allosteric activation
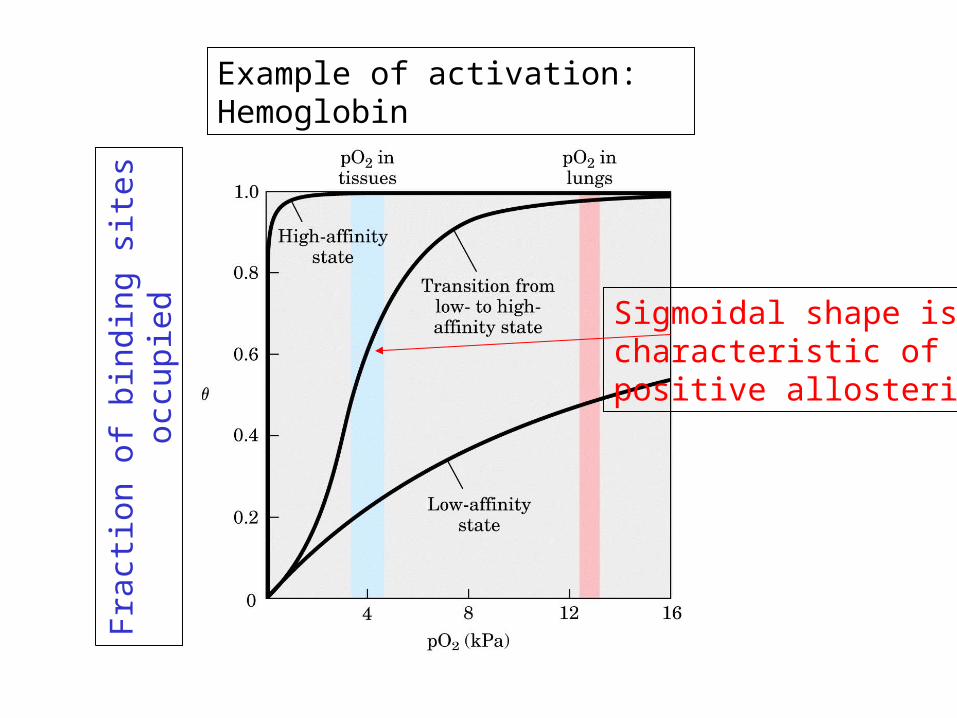
Example of activation: Hemoglobin
Fra
ctio
n of
bin
ding
sit
es
occ
upie
d
Sigmoidal shape ischaracteristic ofpositive allosterism

Negative regulation of biochemical pathwaysis often allosteric
Threonine dehydratase is negatively regulated by L-isoleucine
Feedback Inhibition

Covalent Modifications of Proteins

Perhaps the most important of these is phosphorylation
O P O
O-
O-
OH + ADP+ ATP
Kinase

ATP ADP
Kinase
PhosphataseActive Inactive

Proteolytic cleavage of proteins

Purification of proteins
1. Size Exclusion chromatography (aka Gel Filtration)Separation of proteins by size
Larger (bigger) proteins are “excluded” and go through faster
Smaller proteins are retained by the beads and go through slower

Ion-exchange chromatography
In a cation exchanger, thepositively charged proteins will tend to stick to the column and move slowly through the column.In contrast, negatively charged proteins will move faster through the column
Opposite is true for the anion exchangers.

2. Analysis of the subunit composition of the protein
A. Determination of MW by Gel filtration Chromatography
RelativeElutionVolume
+
++
++
++
++
+
Log MW
Proteins with known MW
Larger proteins elute first (in smaller elution volume)

B. SDS-Polyacrylamide Gel Electrophoresis (SDS-PAGE)
S
O
O
-O O (CH2)11 CH3Na+
Sodium dodecyl sulfate
Polyacrylamidematrix
Breaks disulfide bridges
HOH2C CH2SH
2-mercaptoethanol

1 2 3 4 5 6

Larger proteins elute later (migrate less distance on the gel)

3. Amino acid composition of the protein
Heat (>105°C)24 Hr
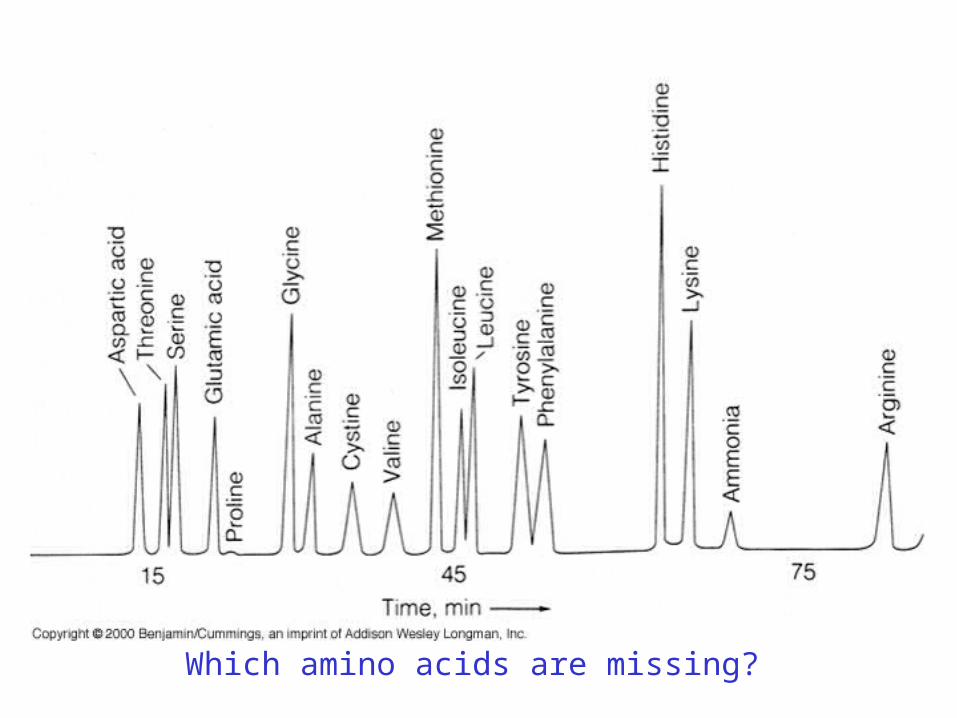
Which amino acids are missing?

Tryptophan (May be accounted for by UV absorbance)
Gln Glu
Asn Asp
O
NH2
Acid HydrolysisO
O-
Hence,Amt. of Glu (by acid hydrolysis) = Amt. of Glu + Amt. of GlnAmt. of Asp (by acid hydrolysis) = Amt. of Asp + Amt. of Asn

4. Amino acid sequence of protein
Edman degradation