-
7/27/2019 chenjian-15.pdf
1/10
MIN I-REV IEW
Yin Li . Gongyuan Wei . Jian Chen
Glutathione: a review on biotechnological production
Received: 1 June 2004 / Revised: 16 August 2004 / Accepted: 31
August 2004 / Published online: 12 October 2004# Springer-Verlag
2004
Abstract This Mini-Review summarizes the historicdevelopments
and technological achievements in the
biotechnological production of glutathione in the past 30
years. Glutathione is the most abundant non-protein
thiolcompound present in living organisms. It is used as a
pharmaceutical compound and can be used in foodadditives and the
cosmetic industries. Glutathione can be
produced using enzymatic methods in the presence of ATPand its
three precursor amino acids (L-glutamic acid, L-cysteine, glycine).
Alternatively, glutathione can be
produced by direct fermentative methods using sugar asa starting
material. In the latter method, Saccharomycescerevisiae and Candida
utilis are currently used to produceglutathione on an industrial
scale. At the molecular level,the genes gshA and gshB, which encode
the enzymes -glutamylcysteine synthetase and glutathione
synthetase,
respectively, have been cloned from Escherichia coli
andover-expressed in E. coli, S. cerevisiae, and Lactococcuslactis.
It is anticipated that, with the design and/ordiscovery of novel
producers, the biotechnological
production of glutathione will be further improved toexpand the
application range of this physiologically andmedically important
tripeptide.
Introduction
Glutathione (-glutamyl-L-cysteinylglycine, GSH) is themost
abundant non-protein thiol compound widely
distributed in living organisms and, predominantly, ineukaryotic
cells (Meister and Anderson 1983). While over90% of the glutathione
is normally present in the reduced
form GSH, several additional forms of glutathione arepresent in
(microbial) cells, tissues, and plasmas. Gluta-thione disulfide
GSSG (oxidized glutathione), formedupon oxidation of GSH, can be in
turn be reduced to GSH
by glutathione reductase at the expense of NADPH(Carmel-Harel
and Storz 2000). Besides GSSG, GSHmay occur in other forms of mixed
disulfides, for example,GS-S-CoA, GS-S-Cys (Penninckx 2002), and
GS-S-pro-tein which is formed via glutathionylation.
Although GSH has been found to be involved in manyphysiological
processes and to play various importantroles, the major and general
functions of GSH can besummarized into three major ways, i.e.
serving as
antioxidant, immunity booster, and detoxifier in
highereukaryotic organisms (Pastore et al. 2003). First, thestrong
electron-donating capability of GSH and therelatively high
intracellular concentration (up to millimolarlevels) enable the
maintenance of a reducing cellularenvironment. This makes GSH an
important antioxidantfor protecting DNA, proteins, and other
biomoleculesagainst oxidative damage generated by, for
example,reactive oxygen species. Second, GSH plays an importantrole
in immune function via white blood cell productionand is one of the
most potent anti-viral agents known.Finally, GSH can be conjugated
to exogenous electro-
philes and diverse xenobiotics by glutathione-S-transfer-
ase to accomplish detoxification. GSH is thus consideredto be
one of the most powerful, versatile, and importantself-generated
defense molecules. In humans, GSH defi-ciency has been linked to a
number of disease states: HIVinfection, liver cirrhosis, pulmonary
diseases, gastrointes-tinal and pancreatic inflammations, diabetes,
neurodegen-erative diseases, and aging (Wu et al. 2004). GSH
iswidely used as a pharmaceutical compound and has the
potential to be used in food additives and in the
cosmeticindustries (Sies 1999), given that the price can be
furtherdecreased by improving production methods.
Y. Li (*) . G. Wei . J. ChenThe Key Laboratory of Industrial
Biotechnology, Ministry ofEducation; School of Biotechnology,
Southern YangtzeUniversity,170 Huihe Road,Wuxi, 214036, Peoples
Republic of Chinae-mail: [email protected].: +86-510-5885727Fax:
+86-510-5888301
G. WeiSchool of Life Science, Soochow University,Suzhou, 215007,
Peoples Republic of China
http://www.paper.edu.cn
-
7/27/2019 chenjian-15.pdf
2/10
While the physiological roles of GSH in human andanimal tissues,
in plant cells, and in microbial cells have
been extensively explored and described (Carmel-Hareland Storz
2000; Penninckx and Elskens 1993; Penninckx2000, 2002), reviews
covering the biotechnological
production of this medically important tripeptide arescarce. One
reason might be that much of the outcomeof research on the
production of GSH is patented, due to
the great economic interest. The latest reviews describingthe
production of GSH were published nearly 10 years ago(Murata 1989,
1994; Murata and Kimura 1990), withspecial emphasis on the
production of GSH by geneticallyengineered Escherichia coli or
Saccharomyces cerevisiae.
In this Mini-Review, we aim to provide a summary ofthe historic
developments and technological achievementsin the biotechnological
production of GSH in the past 30years. Patents relevant to
enzymatic and fermentative
production of GSH are cited, to show the diversity
ofglutathione-producing approaches. Furthermore, future
perspectives on the biotechnological production of gluta-thione
are given.
Production of glutathione: a brief historic overview
GSH was discovered in ethanol-extract of bakers yeast in1888 and
was referred to as philothion. It wassubsequently renamed
glutathione follow the establish-ment of its molecular structure in
1921 (Penninckx andElskens 1993). With the discovery of the
presence of GSHin many living organisms, solvent extraction of GSH
fromanimal or plant tissues was exploited as a preparativeapproach.
However, the limited raw materials availableand the relatively low
intracellular content of GSH made
the end-product expensive, thus hampering its
practicalapplication.
Nearly 70 years ago, it was demonstrated that glutathi-one could
be synthesized by a chemical method(Harington and Mead 1935). The
introduction andremoval of sulfhydryl protective agents on a
cysteineresidue were crucial steps in the chemical synthesis ofGSH
(Douglas 1989). Although this process was com-
mercialized in the 1950s, chemically synthesized GSHwas an
optically inactive (racemic) mixture of the D- and L-isomers. As
only the L-form is physiologically active, anoptical resolution is
required to separate the L-form fromits D-isomer.
Following the observation of the biosynthesis of GSH inan
isolated liver and the characterization of the biosyn-thetic
pathway for GSH (Bloch 1949), explorations on theenzymatic and
fermentative production of GSH were
pursued. GSH is synthesized in two consecutive ATP-dependent
reactions (Meister and Anderson 1983). Thedipeptide
-glutamylcysteine (-GC) is first synthesizedfrom L-glutamic acid
and L-cysteine by -glutamylcysteine
synthetase (also known as glutamate-cysteine ligase, EC6.3.2.2,
GSHI). In the second step, catalyzed by glutathi-one synthetase (EC
6.3.2.3, GSHII), glycine is added tothe C-terminal site of-GC to
form GSH. Generally, theactivity of GSHI is feedback-inhibited by
GSH (but notGSSG) to avoid over-accumulation of GSH, which is
of
physiological significance (Richman and Meister 1975).Meanwhile,
cellular GSH can be degraded by -
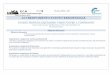
Fig. 1 Overview of GSH-related metabolism pathways and
thegeneral physiological roles played by GSH in microorganisms.
Cyscysteine, Gly glycine, Glu glutamate, G6PDH
glucose-6-phosphatedehydrogenase, GPx glutathione peroxidase, GR
glutathione reduc-tase, GRX glutaredoxin, GSHI -glutamylcysteine
synthetase,GSHII glutathione synthetase, GTP -glutamyltransferase,
ROOHperoxides, ROH reduced peroxides, SS disulfide bond
Table 1 Development of thebiotechnological production ofGSH
Research interest in GSH production Period
Breeding of yeast (predominantly S. cerevisiae and Candida
utilis) for glutathione
production, using either fermentative or enzymatic methods
19761985,
1998present
Purification of glutathione from biological systems
19761987,19931998
Selection of bacteria to produce glutathione using enzymatic
methods 19781991
Construction of ATP regeneration systems 19781982
Molecular biology of GSH biosynthesis in E. coli and
construction of recombinant
E. coli to produce GSH
19821990
Construction of recombinant S. cerevisiae to produce GSH
19861990,
19961998
Optim ization and control of glutathione production process
19911994,
1997present
Production of glutathione by lactic acid bacteria
2001present
234 http://www.paper.edu.cn
-
7/27/2019 chenjian-15.pdf
3/10
Table2
Anoverviewoftheliteratureontheenzymaticproductionofglutathionebymicroorganisms.Enzymaticproductionofglutathioneusesareactionmix
turecontainingL-glutamic
acid,L-cysteine,andglycine
GSHbiosynthesis
ATPregeneration
ATPsupply
T (C)
pH
t (h)
GSH
(mgl1)
Yieldon
Cys(%)
Reference
C.
kruseiIFO0011
Notnecessary
37
6.0
16
350
1.4
Yokozekietal.
(1985)
S.cerevisiaeIFO2044(immobilize
d)
Glycolysis
(glucose)
30
7.5
5
240
7.8
Chibataetal.
(1979b)
S.cerevisiaeIFO0304(treatedbysodiumlaurylsulfate)
Glycolysis
(glucose)
30
7.5
18
11.1
24.1
Takesueetal.
(1979)
S.cerevisiaeIFO0021(treatedbysodiumdodecylsulfateand
-1,3-glucanase)
Glycolysis
(glucose)
37
7.3
15
9,060
98.4
Miwa(1978)
S.glyoxalphilusCDA-5(treatedby
sodiumdodecylsulfate,
immobilized)
Glycolysis
(glucose)
37
7.3
Continuous8,989
97.6
MiwaandTajima
(1978a)
S.cerevisiae500(immobilizedcrudeenzymesofGSHI,
GSHII)
Immobilizedcarbamyl-
phosphokinase
fromStreptococcus
faecalisR600
ATP
35
7.3
5
325
58.6
Miwa(1976)
S.cerevisiae(immobilizedGSHI,G
SHII)
ATP
30
7.5
2,517
82
MiyamotoandMiwa
(1977)
AchromobacterlacticumFERM-P7401
Notnecessary
37
6.0
16
560
2.2
Yokozekietal.
(1985)
CorynebacteriumglutamicumATCC21171(frozencells
treatedwithsurfactant,xylene)
Brevibacterium
ammoniagenes
ATCC21170
(frozen
cellstreated
withsurfactan
t,
xylene)
Glycolysis
(glucose)
37
7.4
6
2,456
49.0
KyowaHakkoKogyo
Co.(1985b)
E.
coli(treatedwithsurfactant,xylene)
ATP
37
7.4
6
2,714
58.9
KyowaHakkoKogyo
Co.(1985a)
E.
coliATCC11303
Glycolysis
(glucose)
37
7.4
2,700
35.2
Fujioetal.
(1985)
E.
coliBATCC23226(crudeenzy
mesofGSHI,GSHII)
Acetatekinase
preparedfrom
E.
coliBATC
C23226
Dextran-N-[(6-ami-
nohexyl)-
carbamoylmethyl]
ATP
37
7.0
3
46.3
3.0
Chibataetal.
(1979a)
Proteusmirabilis(cell-freeextract)
Driedyeast
ATP,glucose
37
8.5
2
1,320
61.4
AjinomotoCo.(1984)
P.mirabilisIFO3849
ATP
37
9.1512
280
1.3
AjinomotoCo.(1982a)
P.vulgaris(crudeGSHI,GSHIIenzymesmodifiedbyN-
ethylmaleimide
ATP
37
9.1512
480
AjinomotoCo.(1982b)
Phormidiumlapideum(2,5
00lx)
Lightusedasan
externalenerg
ysource
ATP
47
7.5
12
1.5mgg
1
wetcell
Sawaetal.
(1986)
235
http://www.paper.edu.cn
-
7/27/2019 chenjian-15.pdf
4/10
glutamyltranspeptidase (-GTP) to transfer the -glutamylmoiety
(Meister and Anderson 1983), which is importantin amino acid
transport. To overproduce GSH using eitherenzyme/intact cell
biocatalysis or fermentation, the releaseof GSHI from feedback
inhibition by GSH and/or theinactivation or deficiency of-GTP are
necessary (Murata1994). Figure 1 gives a biochemical overview,
showingthe biosynthetic and metabolic pathways and illustrating
the general physiological roles of GSH.Table 1 summarizes the
development of the biotechno-
logical production of GSH. Research on fermentative andenzymatic
production of GSH was very active between1976 and 1985 in Japan;
and fermentative production ofGSH by yeast was commercialized in
the early 1980s.Since then, the number of patents on the production
ofGSH declined dramatically. To date, the enzymatic
production of GSH has not been commercialized becauseof the
relatively high production cost.
Enzymatic production of glutathione
One of the well studied approaches to glutathioneproduction is
the enzymatic method. In principle, theessential elements to
constitute an enzymatic synthesissystem include: GSHI, GSHII,
precursor amino acids (L-glutamic acid, L-cysteine, glycine), ATP,
necessarycofactors (Mg2+) to maintain the activities of GSHI
andGSHII, and a suitable pH (usually pH 7.5). Table 2 givesan
overview of the enzymatic production of GSH usingeither intact
cells or crude enzymes. It can be generallyconcluded from Table 2
that the addition of ATP is notnecessary if using S. cerevisiae,
but is required if using
prokaryotes or crude enzymes. This is due to the fact that,
in S. cerevisiae, the strong ATP regeneration capability
ofglycolysis compensates the consumption of ATP. Also,increasing
the cell wall permeability (induced by freezingthawing and/or
treatment with surfactants or enzymes)enhances enzymatic production
significantly when intactcells are used.
The requirement for ATP in the enzymatic production
ofglutathione makes this process difficult to scale-up, since itis
impractical from an economic point of view to add ATPdirectly on an
industrial scale. Accordingly, it would be ofindustrial interest to
construct a highly efficient ATPregeneration system, which can be
briefly defined as asystem in which ATP-requiring reactions are
coupled with
ATP-producing reactions. ATP regeneration systems canbe
categorized into self-coupling systems which workwithin one
organism and co-coupling systems which areconstructed between two
or more organisms. When a co-coupling ATP regeneration system is
used, the efficiencyof ATP/ADP transport across the respective cell
mem-
branes should be carefully taken into account.Self-coupling ATP
regeneration systems have not been
intensively investigated, since it is difficult to
simulta-neously improve the activities of both GSH biosynthesisand
ATP regeneration in one organism (Shimosaka et al.1982). The
co-coupling ATP regeneration system is the
only feasible approach to date. The reaction catalyzed byacetate
kinase in E. coli cells was explored as an ATPgeneration system to
be coupled with GSH biosynthesisreactions, resulting in the
successful production of GSH(Langer et al. 1976). However, the
acetyl phosphatesubstrate required for this approach was unstable
andexpensive. Murata and co-workers considered the glyco-lytic
pathway of S. cerevisiae to be the simplest and the
most capable system for regenerating sufficient ATP forGSH
biosynthesis (Murata et al. 1981b). When theeconomic supply of ATP
was no longer a problem, thelow activities of GSHI and GSHII became
the limitingfactors in GSH biosynthesis. The requirement to
enhancethe activities of GSHI and GSHII accelerated theapplication
of molecular cloning and genetic engineeringapproaches in GSH
biosynthesis.
Fermentative production of glutathione
GSH producer screening
The advantage of the enzymatic production of GSH is thata high
concentration (up to 9 g l1; Table 2) can beachieved. However,
since the usage of three precursoramino acids increases the
production cost, the fermentative
production of glutathione using sugar materials assubstrates has
been extensively studied and is the majorcommercial method
currently used. S. cerevisiae andCandida utilis are the most
commonly used microorgan-isms on an industrial scale; and the GSH
contents of thewild-type strains are usually high (0.11.0% dry
cellweight). Therefore, these two microorganisms werechosen as
targets for mutagenesis. The physical or
chemical mutagenesis methods used included UV, X-radiation,
-radiation, and N-methyl-N-nitro-N-nitroso-guanidine (NTG)
treatment, while resistance to com-
pounds such as glutathione analogues (methionine, ethi-onine),
1,2,4-triazole, sodium cyanide, and sulfite wasused to screen GSH
over-producers. The mechanism forthe selection, in most cases, was
to disrupt or release thefeedback inhibition of GSH on GSHI.
Several examplesillustrate how powerful the screening strategy was:
(1) C.utilis n74-8 sensitive to DL-ethionine was cultured to give
aGSH content of 3.0% compared with the parent content of0.665%
(3.5-fold increase; Ikeno et al. 1977), (2) C. utilisresistant to
methionine was treated with UV, X-radiation,
and NTG to select a mutant resistant to both sulfite
andethionine and the intracellular GSH content increased from0.52%
to 4.0% (6.7-fold increase; Kono et al. 1977), (3) S.cerevisiae
TRZ-6 with resistance to 1,2,4-triazole and
NaN3 had a GSH content of 2.5% (4.0-fold increase;Hamada et al.
1983). The production of GSH byfermentative methods is summarized
in Table 3. Generally,a GSH content of 35% can be obtained using
thesemutants. A particularly high level of GSH content, 9.5%,was
also reported in S. cerevisiae (Ishii and Miyajima1989).
236
http://www.paper.edu.cn
-
7/27/2019 chenjian-15.pdf
5/10
Table3
Anoverviewofliterature
onthefermentativeproductionofglutath
ionebymicroorganisms.TTemperature,
tculturetime,superscriptrresistant,su
perscriptssensitive
Strain
Phenotype
Carbon
source
NitrogensourceSpecialnutrient
T (C)
t (h)
GSHa
GSH
(mgl1)b
Extraction
Recovery
rate
ScaleReference
C.
tropicalisFERM-P
3368
Wildtype
3.0%Glucose(NH4)2SO4,
(NH4)2HPO4
N
icotinamide
30
24
550
FlaskTannoetal.
(1979)
C.
tropicalisFERM-P
4173
Wildtype
4.2%molas-
ses
Urea,
NH4H2PO4
30
14
0.73
Water(90C)
FlaskSuzukiandMat-
subayashi
(1980)
C.
tropicalisPK233
Wildtype
5%ethanol,
0.5%glucose
(NH4)2SO4
C
asaminoacid,
biotin30
96
60(extracel-
lular)
FlaskNipponZeon
Co.(1983)
C.
tropicalis239D5
Ethioniner
,
sulfiter
3.0%glucose(NH4)2SO4,
(NH4)2HPO4
N
icotinamide
30
24
5
FlaskHamazawaetal.
(1998)
C.
utilisIFO1086
Wildtype
3.0%glucose(NH4)2HPO4
Y
eastextract,corn
s
teepliquor,L-cys-
t
eine
30
20
2.2
250
Water(95C),
Cu2+-H2S
70
20lTannoetal.
(1976)
C.
utilisn74-8
Ethionines
3.0%glucose(NH4)2HPO4
Y
eastextract
30
15
3.0
450
H2SO4,
Cu
2+-
H2S
74.6
70lIkenoetal.
(1977)
C.
utilisFERM-P6907Ethioniner
,
sulfiter
3.0%glucose(NH4)2SO4
30
24
4.5
680
H2SO4,
Cu2O-
H2S
70
200lKohjinCo.
(1984)
C.
utilisFERM-P7396Ethioniner
,
sulfiter
3.0%glucose(NH4)2SO4
302630
5.0
735
200lHinoetal.
(1985)
C.
utilisWSH02-06
Wildtype
2.8%glucose(NH4)2SO4
24
28
2.5
385
7l
Weietal.
(2003a)
S.cerevisiaeIAM4207
3%methanol,
2%molasses
(NH4)2SO4
30
48
1.85
80
H2SO4,
Cu2O-
H2S
4050
25lMiyamotoetal.
(1977)
S.cerevisiaeTRZ-6
1,2,4-triazoler,
NaN3
r
6%molasses
(NH4)2SO4,
NH4H2PO4
C
ornsteepliquor
30
24
3.5
860
H2SO4,
Cu2O-
H2S
70
2l
Hamadaetal.
(1983)
S.cerevisiaeR-3
Amider,
indophenolr
6%molasses
(NH4)2SO4,
NH4H2PO4
C
ornsteepliquor
30
24
680
H2SO4,
Cu2O-
H2S
70
2l
Kawamuraetal.
(1985)
S.cerevisiaeK-2
Deficientinga-
lactoseforma-
tion
5%sucrose
Urea,(NH4)
2SO4,
(NH4)
2HPO4
G
alactose
33
24
2,700
FlaskNomuraetal.
(1985)
S.cerevisiae
Glucose
Urea
B
iotin
30
40
9.5
4,320
125lIshiiandMiyaji-
ma(1989)
S.cerevisiaeKY6186
Molasses
(NH4)2SO4,
NH4H2PO4
30
60
3.7
120
m3
SakatoandTa-
naka(1992)
S.cerevisiae
Glucose
(NH4)2SO4
30
7.6
300
3l
Alfafaraetal.
(1993)
S.cystinovolensFERM-
P3831
Wildtype
2%cornsteep
liquor
(NH4)2SO4
Y
eastextract,DL-
m
ethionine,L-cys-
t
eine
30
32
116
H2SO4
(95C)
FlaskMiwaandTaji-
ma(1978b)
237
http://www.paper.edu.cn
-
7/27/2019 chenjian-15.pdf
6/10
There are few reports on the selection of prokaryoticGSH
producers. Immobilized methylglyoxal (MG)-resis-tant E. coli
produced GSH to levels 1.6-fold higher thanthat of the control in
the presence of MG. Presumably thestress induced by the toxic MG
increased the production ofGSH, which functioned as a detoxifier
(Murata et al.1981c). Selenite-resistant mutants of E. coli
producedhigher amounts of GSH (ranging from 9% to 173%,
compared with the wild type; Schmidt and Konetzka1986).
Following the observation that GSH was present ina blue-green
bacterium (Fahey et al. 1978), someresearchers were interested in
finding GSH producersamong green algae or cyanobacteria. These
bacteriadisplayed GSH-related different characteristics,
comparedwith yeast or E. coli. For instance, the cyanobacterium
Phormidium lapideum produced GSH in the presence ofprecursor
amino acids and ATP; and the ATP wasefficiently regenerated from
ADP using light as anexternal energy source. GSH was also produced
underdark conditions without ATP, when ATP was
presumablyregenerated by a respiratory system active in the
dark
(Sawa et al. 1986). The production of GSH by the greenalga
Dunaliella sp. was also of industrial interest. A GSHcontent of
2.38% was achieved in a medium containingonly inorganic nutrients
(Yamaoka and Takimura 1990).Although the relatively low total
biomass concentrationlimited the final GSH concentration, this
study providedan alternative way to produce GSH, or
high-GSH-contentfood additives.
Process optimization
The ultimate goal of the biotechnological production of
GSH is to achieve a high total GSH concentration
throughincreasing the intracellular GSH content and cell
density.While the intracellular GSH content was
improvedsignificantly by mutagenesis, the enhancement of
cellconcentration could be achieved by process optimizationand
control. The intracellular GSH content might also bemaximized under
the control of a suitable cultivationstrategy.
Selection of nutrients in the culture medium is usuallyimportant
for the fermentative production of GSH and theconcentration of
nutrients should also be optimized.Central composition experimental
design has been appliedto examine the effects of certain nutrients
(glucose,
peptone, KH2PO4, biotin, cysteine) on the cell growthand
intracellular GSH content of S. cerevisiae S-8H. Themean of total
GSH concentration obtained at the optimalconditions was 160.1 mg
l1, which was nearly 2-fold thatof the control (Udeh and
Achremowicz 1997). BoxBehnken design and response surface
methodology havealso been used to optimize the concentration of
glucose,
peptone, and MgSO4 in the production of GSH by S.cerevisiae CCRC
21727. The derived neural networkmodels predicted cell growth and
GSH production more
precisely than the second-order response surface models(Liu et
al. 1999).S
train
Phenotype
Carbon
source
NitrogensourceSpecialnutrient
T (C)
t (h)
GSHa
GSH
(mgl1)b
Extraction
Recovery
rate
ScaleReference
Methylomonasmetha-
nolvorensFERM-P
3330
Wildtype
Methanol
(NH4)2SO4
30
48
1.82
50
Water(90C),
chromotog-
raphy
80
25lMiyamotoetal.
(1978)
ProteusmirabilisIFO
3849
Wildtype
1.0%glucosePeptone,
NaNH4HPO4
Serine,H3BO3
31
36
180(extracel-
lular)
Chromotography61
2l
AjinomotoCo.
(1983)
Dunaliellasp.(
irradia-
tion1200lx)
Wildtype
CO2
NaNO3
24
144
2.38
FlaskYamaokaand
Takimura
(1990)
Phormidiumlapideum
(irradiation2500lx)
Wildtype
CO3
2,
HCO3
48
24
0.15
FlaskOchiai(1987)
aTheintracellularGSHcontent,sho
wnasthemasspercentageofGSHinth
etotaldrycellweight(%)
bThetotalGSHconcentrationinthefermentationmedium,whichequalsthe
intracellularGSHcontentmultipliedby
thedrycellconcentration
Table3
(continued)
238 http://www.paper.edu.cn
-
7/27/2019 chenjian-15.pdf
7/10
Fed-batch culture is one of the most efficient methods toachieve
a high cell density of yeast culture. However, thespecific growth
rate () during fed-batch culture should becarefully controlled, to
avoid a decrease in intracellularGSH content. Based on the mass
balance, together withthe relationship between and the specific
GSH
production rate, a simple mathematic model was devel-oped, to
determine the optimal profile of in a yeast fed-
batch culture (Shimizu et al. 1991). An ideal profile offor
maximizing the production of GSH was realized bymanipulating the
glucose-feeding rate with the extendedKalman filter and a
programmed-controller/feedback-compensator system. As a
consequence, the maximum
production of GSH was 41% higher than that of thecontrol
(Shimizu et al. 1991). Furthermore, a fuzzy logiccontroller was
developed to control the ethanol concen-tration in fed-batch
cultures of S. cerevisiae, to maximizeGSH production. Moreover,
when of cells in the GSH
production phase was adjusted to maintain a specific
GSHproduction rate of 6.2 mg g1 h1, the total GSHconcentration
achieved was 56% higher than that of the
control, where was kept at a constant level throughoutthe
process (Alfafara et al. 1993). Alternatively, using afeed-forward
and a feed-back control system based on theon-line data of oxygen
and ethanol concentrations in theexhaust gas, an average of 40%
improvement in GSH
production was obtained, compared with the
conventionalprogrammed control of exponential fed-batch
operationwith S. cerevisiae (Sakato and Tanaka 1992).
Although sugar material was the major substrate in
thefermentative production of GSH, the addition of precursoramino
acids required for GSH was an easy approach intrials to increase
GSH production. L-Cysteine wasconfirmed to be a key amino acid for
increasing the
specific GSH production rate, but it showed some
growthinhibition in the second growth phase of S. cerevisiaewhen
glucose was used as the sole carbon source (Alfafaraet al. 1992a).
Therefore, a suitable L-cysteine additionstrategy should be
developed to increase GSH productionwithout causing growth
inhibition. Alfafara et al. (1992b)found that single-shot addition
of L-cysteine was betterthan continuous addition, where the
concentration of L-cysteine was maintained at a constant level
(Alfafara et al.1992b). Using this knowledge, they developed a
mass
balance model-based feeding strategy in which L-cysteinewas
added in a single-shot manner to a culture entering theGSH
production phase. As a consequence, the specific
GSH production rate increased approximately 2-foldcompared with
that of the control (Alfafara et al.1992b). A similar stimulatory
effect of L-cysteine onGSH production was observed in recombinant
E. coli (Liet al. 1998), where the total GSH concentration and
theintracellular GSH content increased by 40% and
100%,respectively, when 9 mM L-cysteine was added to theculture at
12 h. Besides L-cysteine, several other materialswere found to have
a stimulatory effect on GSH
production: amino acid supplements (yeast extract, pep-tone) on
S. cerevisiae (3.4-fold increase; Watanabe et al.1986), ethanol (26
g l1) on S. cerevisiae (1.4-fold
increase; Kyowa Hakko Kogyo Co. 1984), p-aminoben-zoic acid (200
mg l1) on Hansenula capsuleita (94%increase; Kinoshita et al.
1986), and sodium lactate (10 gl1) on S. cerevisiae (82% increase;
Hirakawa et al. 1985).
Genetic/metabolic engineering
Early studies on the properties of GSHI and GSHIIshowed that
these enzymes were feedback-inhibited byGSH and GSSG, respectively
(Murata and Kimura 1990),indicating that GSHI was the rate-limiting
step in GSH
biosynthesis. The genes gshA and gshB encoding GSHI(Murata et
al. 1981a) and GSHII (Murata et al. 1983) werecloned and sequenced.
Specifically, an E. coli B strainwith GSHI desensitized to feedback
inhibition of GSHwas screened and the desensitized GSHI-coding
gene
gshA* was cloned (Murata and Kimura 1982). Theintroduction of
the recombinant plasmid pGS500 harbor-ing the gshA* and gshB genes
into E. coli RC912 resultedin a simultaneous increase in the
activities of GSHI (10.0-
fold) and GSHII (14.5-fold; Gushima et al. 1983).Although the
intracellular GSH concentration in E. coliRC912 (pGS500) cells only
increased 1.3-fold comparedwith the wild type, the intact
recombinant E. coli cellswere used as an excellent GSH biosynthesis
system, bywhich 5 g l1 GSH was produced in the presence of
three
precursor amino acids and ATP. Similar work was done inS.
cerevisiae, where the expression of GSHI and GSHIIincreased
1,039-fold and 33-fold, respectively, and theintracellular GSH
content increased 2-fold (Ohtake et al.1988, 1989).
The overexpression of gshA and gshB did not lead to asignificant
increase in GSH content in either E. coli or S.
cerevisiae. This might be due to: (1) feedback inhibitionon GSHI
caused by GSH, or (2) the presence of -GTPdegrading the GSH
synthesized intracellularly. Recently,an extremely high
intracellular concentration of GSH, upto 140 mM upon addition of 5
mM L-cysteine, wasachieved in Lactococccus lactis NZ9000 expressing
the
gshA and gshB from E. coli under the control of a nisin-induced
control expression system. This is the highestintracellular GSH
content achieved for a bacterium to date(Li et al. 2004). L. lactis
is not capable of GSH
biosynthesis and therefore there is no -GTP
activity.Furthermore, the feedback inhibition of GSH on GSHIseems
not to occur in L. lactis (where the intracellular
GSH concentration of 140 mM achieved was obviouslyhigher than
the concentration normally required forfeedback inhibition),
although the reason for that remainsunclear. The development of
genetic engineering ap-
proaches for GSH production is summarized in Table 4.
Conclusions and perspectives
One topic that is not discussed in this Mini-Review is
thesecretion of GSH, a process which might increase the totalGSH
concentration in the culture and facilitate the
239
http://www.paper.edu.cn
-
7/27/2019 chenjian-15.pdf
8/10
recovery process. C. tropicalis PK233 (Nippon Zeon Co.1983) and
Proteus mirabilis IFO 3849 (Ajinomoto Co.1983) are capable of
producing a certain amount of GSHextracellularly. We have shown
that the addition of acritical concentration of surfactants
enhanced the extra-cellular accumulation of GSH by S. cerevisiae
(Wei et al.2003b). We recently showed that C. utilis 02-08
secretedGSH into the medium when the pH of the culture
decreased to pH 1.5 (Nie W et al., unpublished data).Based on
this observation, a pH shift strategy wasdeveloped and the total
concentration of GSH increased(up to 2.0-fold). If GSH can be
produced extracellularly byfood-grade microorganisms, i.e., lactic
acid bacteria, theGSH present in the food matrix will no doubt
increase thenutritional value, as GSH is considered as a
nutraceutical.
Related to another aspect for future studies, previousefforts on
GSH production mainly focused on improvingthe activity of the GSH
biosynthesis system itself.However, as cysteine availability is
usually the limitingstep for GSH biosynthesis, we propose
developing afurther metabolic engineering strategy to improve
the
cysteine biosynthesis capability, instead of focusing onlyon the
GSH biosynthesis pathway. In addition, it is widelyrecognized that
GSH plays important roles in oxidativestress resistance. However,
this property was not related tothe production of GSH until Kimura
et al. (1996) reportedthe cloning and expression of oxidative
stress-resistancegenes from a peroxide-resistant S. cerevisiae.
Theintroduction of these genes into another S. cerevisiaestrain
enhanced GSH production (up to 2.5-fold; Kimuraet al. 1996). This
investigation gave a clue how to design anovel metabolic
engineering strategy for GSH production.
In conclusion, GSH is successfully produced on anindustrial
scale using fermentative methods and serves as
an important pharmaceutical. The range of applicationswhere GSH
is employed is expanding in line with thediscovery of new
functional roles for this bioactivetripeptide. With the design
and/or discovery of novel
producers, or the deregulation of biosynthesis in theknown
producers, the biotechnological approach offerssignificant
opportunities to further decrease the productioncost of GSH, which
will further the application andutilization of GSH.
Acknowledgements The authors thank Dr. Paul W. OToole
forcritically reading this manuscript. This study was supported by
theNational Science Foundation of China (contract no.
30300009).
References
Ajinomoto Co. (1982a) Production of glutathione. JP
patent57,002,698
Ajinomoto Co. (1982b) Production of glutathione. JP
patent57,005,699
Ajinomoto Co. (1983) Microbial production of glutathione.
JPpatent 58,016,694
Ajinomoto Co. (1984) Production of glutathione. JP
patent59,156,298
Table4
Productionofglutathione
byrecombinantmicroorganisms.Formo
reinformationontheproductionofglutathionebyengineeredE.
coliorS.cerev
isiae,refertoMurataand
Kimura(1990)
Hoststrain
Cloningstrategy
GSHinmu
tant
GSHinwildtypeIncrement
(x-fold)
Reference
S.cerevisiae
Astrongpromoter,P8fromS.cerevisiaeYNN27,wasused
toreplacethepromoterof
GSHI,resultingin
plasmidpGRS2518-x
24mgg
1
3mgg
1
7
Tezukaetal.
(1987)
E.
coli
HindIIIdigestedchromosomalDNAofE.
coliwasinsertedintopBR322
andtransformedintoGSH-deficientE.
colitoselectpositiveclones
7.9molg1
(2.4
2mgg
1)
1.3molg
1
(0.5
5mgg
1)
3.4
KimuraandMur-
ata(1983)
E.
coli
DNAfragmentcontainingthestructuralandpromoterregion
ofgshB(encodingGSHII)
wasclonedinsingle,
double,andtriplecopiesintopBR325
Littleimproved
Watanabeetal.
(1986)
E.
colia
ArecombinantbacteriophagecontainingtwocopiesofgshA
andonecopyofgshBofE.
coliwasconstructedtotransfectE.
coli
7.9molm
g1
proteinh
1