Embed Size (px)
Citation preview

Chromatin, Gene, and RNA Regulation
mda-7/IL-24 Expression Inhibits Breast Cancer throughUpregulation of Growth Arrest-Specific Gene 3 (gas3) andDisruption of b1 Integrin Function
You-Jun Li7,3, Guodong Liu2, Yanmei Li1,3, Laura M. Vecchiarelli-Federico1,3, Jeff C. Liu4,Eldad Zacksenhaus1,4, Sze W. Shan3, Burton B. Yang3, Qi Li6, Rupesh Dash5,8, Paul B. Fisher5,Michael C. Archer1,2, and Yaacov Ben-David1,3
AbstractMelanoma differentiation-associated gene (MDA)-7)/interleukin (IL)-24, a member of the IL-10 family of
cytokines, inhibits growth of various human cancer cells, yet the underlying mechanism is largely unknown. Here,we report that mda-7/IL-24 efficiently suppresses the development of rat mammary tumors in vivo. Microarrayanalysis for genes differentially expressed in rat mammary tumor cells overexpressingMDA-7/IL-24 compared withthose that do not express this cytokine identified growth arrest-specific gene-3 (gas3) as a target for mda-7/IL-24.Upregulation of gas3 by mda-7/IL-24 was STAT3 dependent. Induction of gas3 inhibited attachment andproliferation of tumor cells in vitro and in vivo by inhibiting the interaction of b1 integrin with fibronectin. AmutatedGAS3,which is unable to bindb1 integrin,was also unable to inhibitfibronectin-mediated attachment andcell growth both in adherent and suspension cultures, suggesting that GAS3 exerts its effects through interactionwith and regulation of b1 integrin. Thus, mda-7/IL-24 inhibits breast cancer growth, at least in part, throughupregulation of GAS3 and disruption of b1 integrin function. Importantly, the expression of the mda-7/IL-24receptor, IL-20R1, is highly correlated with GAS3 expression in human breast cancer (P¼ 1.02� 10�9), and theincidence of metastases is significantly reduced in patients with HER2þ breast cancer expressing high-levels of IL-20R1. Together, our results identify a novel MDA-7/IL-24-GAS3-b1integrin–fibronectin signaling pathway thatsuppresses breast cancer growth and can be targeted for therapy.Mol Cancer Res; 11(6); 593–603.�2013 AACR.
IntroductionBreast cancers are highly heterogeneous and include
luminal-like tumors expressing the estrogen receptor,HER2-like overexpressing HER2 receptor, and basal-likesubtypes that do not express these receptors (triple negative;refs. 1, 2). Although chemotherapy plus hormone therapy
(tamoxifen, aromatase inhibitors) or therapies that targetHER2 (trastuzumab/Herceptin) significantly reduce theincidence of relapse, most women who develop metastaseseventually succumb to the disease. There is, therefore, a greatneed for novel drugs that can inhibit tumor growth byattacking different vulnerabilities of breast cancer cells.Melanoma differentiation-associated gene-7, (mda-7) ini-tially identified by differentiation induction subtractionhybridization from human melanoma cells, is a member ofthe interleukin (IL)-10 family of cytokines, now known asmda-7/IL-24 (3). The activity of MDA-7/IL-24 as a growthsuppressor in melanoma and other cancer cells was firstshown in 1996 (4). Subsequently, ectopic expression of thiscytokine was shown to lead to irreversible growth inhibition,reversal of the malignant phenotype, and terminal differen-tiation in a wide variety of solid tumors including breastcancer (4, 5). Interestingly, these effects are not elicited in thenormal cellular counterparts of these tumors (6). Moreover,the ability of MDA-7/IL-24 to induce bystander cancer-specific cell killing has been confirmed in vivo in a range ofhuman xenograft studies in nude mice and in phase I clinicaltrials involving patients withmelanomas and other advancedsolid cancers (3, 7, 8). Although apoptosis induced byMDA-7/IL-24 involves binding of the cytokine to the IL-20R1–IL-20R2 heterodimeric receptor complex, it can also occurthrough direct intracellular pathways involving binding to
Authors' Affiliations: Departments of 1Medical Biophysics and 2Nutri-tional Sciences, University of Toronto; 3Department of Molecular and CellBiology Research, Sunnybrook Health Sciences Centre; 4Toronto GeneralResearch Institute—University Health Network, Toronto, Ontario, Canada;5Department of Human andMolecular Genetics, VCU Institute ofMolecularMedicine, VCUMasseyCancer Center, Virginia Commonwealth University,School of Medicine, Richmond, Virginia; Departments of 6Pharmacologyand 7Anatomy, Norman Bethune College of Medicine, Jilin University,Changchun, China; and 8Institute of Life Sciences, Bhubaneswar, Orissa,India
Note: Supplementary data for this article are available at Molecular CancerResearch Online (http://mcr.aacrjournals.org/).
Corresponding Authors: Yaacov Ben-David, Sunnybrook HealthSciences Centre, Molecular and Cellular Biology, Rm S216, 2075 BayviewAve, Toronto,OntarioM4N3M5,Canada. Phone: 416-480-6100, ext. 3359;Fax: 416-480-5703; E-mail: [email protected]; and Michael C. Archer,University of Toronto, 150 College, Street, Toronto, Ontario, Canada, M5S3E2. Phone: 416-978-8195; Fax: 416-971-2366; E-mail:[email protected]
doi: 10.1158/1541-7786.MCR-12-0496
�2013 American Association for Cancer Research.
MolecularCancer
Research
www.aacrjournals.org 593
on April 23, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst March 6, 2013; DOI: 10.1158/1541-7786.MCR-12-0496

BiP/GRP78 and translocation to the endoplasmic reticulumresulting in an "unfolded protein stress response" (8–10).The exact mechanism underlying the wide-spectrum, anti-cancer activity of mda-7/IL-24 is not known and may becancer specific, depending on preexisting genetic and epi-genetic alterations (3).Most strains of rats used experimentally such as Wistar-
Furth develop multiple mammary adenocarcinomas follow-ing initiation with a mammary carcinogen, whereas a fewstrains such as the Copenhagen (Cop) are resistant to thedevelopment of mammary tumors (11). To investigate thesedifferences in susceptibility, we previously isolated cell linesfrom tumors induced in resistant Cop � Wistar-Furth F1rats by infusion of a retrovirus harboring the v-Has-ras genedirectly into the main mammary ducts (12). Some of thesecell lines grew in soft agar, whereas others display anchorage-dependent growth. Through microarray analysis, we foundthat the expression of MDA-7/IL-24 and b4 integrin wereinversely correlated with the ability to grow in soft agar (13).Ectopic expression of mda-7/IL-24 in anchorage-indepen-dent cells inhibited growth in monolayer culture and in softagar, their ability to migrate and invade in vitro, and thedevelopment of tumors in nude mice. We showed thatalthough b4 integrin did not play a direct role in regulatingcell growth, it was a downstream target of MDA-7/IL-24,acting in concert with the latter to suppress cell proliferation.Moreover, we showed that growth suppression by MDA-7/IL-24 was associated with upregulation of p27Kip1 viaactivation of STAT3 (13).Here, we show thatmammary tumorigenesis in rats can be
blocked by infection with an adenovirus expressing mda-7/IL-24. We show that growth arrest-specific gene 3 (GAS3/PMP22), is induced by mda-7/IL-24, and that GAS3 inhi-bits the attachment and proliferation of tumor cells, at leastin part, by blocking the interaction of b1 integrin withfibronectin. Moreover, we show that primary human breastcancers exhibit a positive correlation between expression ofthe IL-24 receptor and GAS3 and that receptor expressioncorrelates with metastasis-free survival (MFS). Thus, ourdata identify a novel inhibitory pathway downstream ofmda-7/IL-24, which is important in breast cancer spread andcan be targeted for therapy.
Materials and MethodsTumor inductionTwenty-four 49-day-old Wistar-Furth female rats were
intraperitoneally injected with 75 mg/kg methylnitrosurea(MNU) in 0.05% acetic acid. Thirty days after injection, weinfused via the nipples, the main ducts of thoracic, abdom-inal, or inguinal mammary glands of 12 rats (5–7 glandsinfused/rat) with 15 mL of a solution containing approxi-mately 108 plaque-forming units (pfu) of adenovirus con-taining the human mda-7/IL-24 gene, 8 mg/kg polybrene,and 2 g/mL indigo carmine according to our previouslypublished protocol (12). Glands of 12 control rats weresimilarly infused with approximately 108 pfu of emptyadenovirus. The rats were palpated weekly and tumors wereharvested when they were more than 20 mm in diameter or
when the animals appeared moribund. A portion of eachtumor was fixed in formalin for histologic analysis.
Growth of cells in nude miceFour groups of 6 female athymic nude mice (CD1–Nu/
Nu), 7 weeks of age, were purchased from Charles RiverLaboratories. After a week of acclimatization, 2� 106 FE1.2cells expressing GAS3 or vector alone, in a volume of 50 mL,were injected subcutaneously. Tumor size was measuredtwice weekly with vernier calipers by a single-blinded observ-er. Tumor growth was measured and plotted according tothe nonparametric Kaplan–Meier analysis. Tumors wereharvested 2 weeks after injection.
Cell cultureThe establishment of the rat breast cancer cell lines was
described previously (12). All cell lines were maintained ina-minimum essential medium (a-MEM), supplementedwith 10% FBS, 10 ng/mL EGF, 1 mg/mL hydrocortisone,and 1 mg/ml 17-b-estradiol (Sigma-Aldrich). All cultureexperiments were repeated 3 times. To examine growthrates, 1 � 104 cells were seeded per well on 24-well platesand counted for 4 consecutive days. To examine the regu-lation of GAS3 by MDA-7/IL-24, FE1.2-IL-24 cells weretreated with 10 or 20 mmol/L of the STAT3 inhibitor AG9,as previously described (13).
Attachment assaysAttachment assays were conducted on tissue culture plates
or on Petri dishes precoated with fibronectin (20 mg/mL) orlaminin (20 mg/mL). Cells (1 � 104/mL) were plated intriplicate and counted and photographed 2 or 24 hours afterculture. In some experiments, the STAT3 inhibitor AG9(Calbiochem) was added to the culture medium of theattached cells and they were photographed 4 hours later.The mean percentage of the attached versus the round cellswas determined for at least 20 fields of view.
Growth of cells in soft agarTo monitor the growth of cells in soft agar, 2 layers of
agarose were used. The bottom layer contained 0.5% aga-rose, the top layer 0.3% agarose, both ina-MEM. Triplicatesamples of 5,000 cells were seeded on 6 cm plates andincubated for 5 to 7 days. At the end of the incubationperiod, colonies with more than 25 cells were counted.
Western blottingWestern blotting was conducted either as we have
described previously (13), or for GAS3, as described in ref.(14). Polyclonal rabbit anti-rat antibodies were obtainedfrom the following sources: rabbit anti-rat GAS3 (Abcam)used at a dilution of 1:500; rabbit anti-rat IL-24 (GenHun-ter) used at a dilution of 1:1,000; b-actin (Sigma) used at adilution of 1/50,000; and pan-cadherin (Cell SignalingTechnology) used at a dilution of 1:1,000. Protein banddensities were determined with the ChemiGenius2 BioImaging System using the Gene Tools Software v4.02(Syngene).
Li et al.
Mol Cancer Res; 11(6) June 2013 Molecular Cancer Research594
on April 23, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst March 6, 2013; DOI: 10.1158/1541-7786.MCR-12-0496

ImmunoprecipitationSemiconfluent FE1.2, FE1.2þGAS3, or FE1.2þGAS3-
GM cells were lysed with NP-40 lysis buffer (1% NonidetP-40, 2 mmol/Lphenylmethlsulfonylfluoride, 10 mg/mLaprotinin, 2 mg/mL pepstatin, 10 mmol/L iodoacetamide,0.1 mmol/L EDTA, 0.1mmol/L EGTA, 10 mmol/LHEPES, and 10 mmol/L KCl). Two micrograms of anti-b1 integrin antibody (Santa Cruz Biotechnology Inc.) wasadded to 0.5 mg of cell lysates in 1 mL and shaken at 4�Covernight. Protein-A beads were then added to the mixtureand shaking was continued for additional 2 hours. The beadswere then washed 3 times with NP-40 buffer, boiled, andproteins separated on 10% SDS-PAGE gel, and Westernblot analyses prepared using GAS3 and b1 antibodies. Toshow loading and input controls, Western blot analyses werealso prepared from the total extracts (20 mg) using b-actinand GAS3 antibodies.
ImmunohistochemistryImmunohistochemistry was conducted on cells grown
on cover slips overnight. Cells were fixed in 4% para-formaldehyde and blocked using 10% normal goat serum(Jackson ImmnoResearch Laboratories). Triplicate coverslips were incubated with 1:500 polyclonal rabbit GAS3antibody (Novus Biologicals) overnight at 4�C. Controlcells were incubated with buffer only. The next day, thecover slips were incubated with 1:1,000 biotinylatedgoat anti-rabbit secondary antibody (Vector Laborato-ries), for 1 hour at room temperature. ABC complex(Vector Laboratories) was then applied to the cover slipsfor 30 minutes at room temperature, followed by 3,30-diaminobenzidine for 2 minutes (Vector Laboratories)and counterstaining with hematoxylin. Cells were dehy-drated before mounting using xylene base mountingmedium.
Infection and transfectionA gas3-producing retrovirus was generated by inserting the
entire 658 bp coding sequence of rat gas3 in either sense orantisense orientations into a unique EcoR1 site in theretroviral expression vector MSCV2.1 (13). Replication-defective viruses were prepared by transfecting the viralplasmid into the helper-free packaging cell line GPþA(B8), as described previously (13). For viral infection, super-natants from the virus-producing cells were used to infectFE1.2 cells, plated at a density of 2 � 106. After 48 hours,G418-resistant cells were pooled and subjected to subclon-ing by limited dilution. Transfection was conducted usingLipofectamine 2000 according to the manufacturer's pro-tocol (Invitrogen).
Fibronectin siRNA transfectionCells were transiently transfected with 2 mg of siRNA
against fibronectin (Gene Pharma) or control oligonucleo-tides for 6 hours according to the manufacturer's instruc-tions, as previously described (15). Fresh medium contain-ing 10%FBSwas then added to the transfected cultures, andthey were maintained overnight, spun down, then seeded
onto 6-well plates, and incubated for 30 minutes. Attachedcells were counted.
Primersmda-7/IL-24 primers: Forward; ACG GCC AGG TCA
TCA CTA TTG, Reverse; AGG TCT TTA CGG ATGTCA ACG, b-actin primers: Forward; GCA ACC CAGTGT GAG TAG CA, Reverse; GAT GGT GTC CCCTGC ATT AC.Gas3 primers and controls were supplied byApplied Biosystems (Rn005668351-AI PMP22, Lot #282134).
Statistical analysisStatistical analyses were conducted using the 2-tailed
Student t test or ANOVA with significance considered atP � 0.05 using Origin 3.5 software (Microcal Software).Values are reported as mean� SE. The correlation betweenIL-20R1 and GAS3 genes was assessed by Pearson Corre-lation method with median-centered, log-2-transformed,and Robust Multi-array Average normalized microarraygene expression values. Kaplan–Meier curves and HRs weregenerated as described (16).
ResultsInhibition of rat mammary carcinogenesis by anadenovirus-harboring mda-7/IL-24We have previously shown that mda-7/IL-24 expression
blocks the proliferation of rat mammary tumor cells in vitro(13). To determine whethermda-7/IL-24 can inhibit tumordevelopment in vivo, 24 female Wistar-Furth rats wereintraperitoneally injected with the mammary carcinogenMNU (75 mg/kg; ref. 12). Thirty days later, when weshowed that the mammary glands of the carcinogen-treatedrats contain a significant number of preneoplastic lesions(17), 12 rats were infused with a recombinant adenovirusexpressing the humanmda-7/IL-24 gene into themain ductsof thoracic, abdominal, or inguinal mammary glands (�108
pfu, 5–7 glands injected/rat). The other 12 control rats wereinfused with empty adenovirus (�108 pfu). Tumors werefirst palpated in the control rats at about 6 weeks followingMNU administration and were harvested over the following20 weeks when tumors reached approximately 2.5 cm indiameter. Six of the control rats developed tumors in thistime period (Supplementary Table S1). Histologic analysisshowed that all tumors were typical adenocarcinomas (12).In contrast, during the same time period, no mammarytumors developed in the glands of rats infused with themda-7/IL-24–containing adenovirus (significantly different fromcontrols, P < 0.005 by c2 analysis). Interestingly, 2 tumorsdeveloped in these rats in glands that had not been infusedwith adenovirusmda-7/IL-24. These results show thatmda-7/IL-24 expression inhibits breast cancer development invivo.
mda-7/IL-24 induces GAS3 expressionWe previously showed that proliferation and anchorage-
independent growth of FE1.2 tumor cells were readilysuppressed by MDA-7/IL-24 (12). To begin to address the
mda-7/IL-24 Suppresses Breast Cancer through GAS3
www.aacrjournals.org Mol Cancer Res; 11(6) June 2013 595
on April 23, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst March 6, 2013; DOI: 10.1158/1541-7786.MCR-12-0496
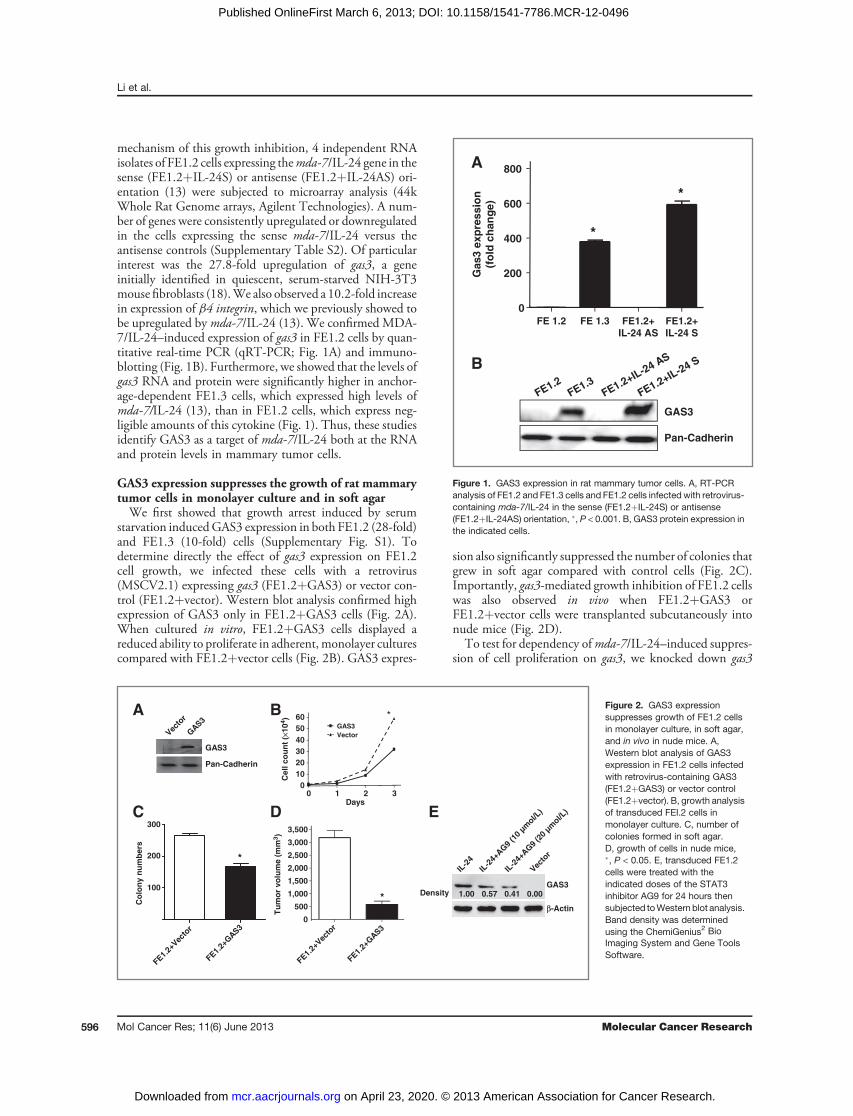
mechanism of this growth inhibition, 4 independent RNAisolates of FE1.2 cells expressing themda-7/IL-24 gene in thesense (FE1.2þIL-24S) or antisense (FE1.2þIL-24AS) ori-entation (13) were subjected to microarray analysis (44kWhole Rat Genome arrays, Agilent Technologies). A num-ber of genes were consistently upregulated or downregulatedin the cells expressing the sense mda-7/IL-24 versus theantisense controls (Supplementary Table S2). Of particularinterest was the 27.8-fold upregulation of gas3, a geneinitially identified in quiescent, serum-starved NIH-3T3mousefibroblasts (18).We also observed a 10.2-fold increasein expression of b4 integrin, which we previously showed tobe upregulated by mda-7/IL-24 (13). We confirmed MDA-7/IL-24–induced expression of gas3 in FE1.2 cells by quan-titative real-time PCR (qRT-PCR; Fig. 1A) and immuno-blotting (Fig. 1B). Furthermore, we showed that the levels ofgas3 RNA and protein were significantly higher in anchor-age-dependent FE1.3 cells, which expressed high levels ofmda-7/IL-24 (13), than in FE1.2 cells, which express neg-ligible amounts of this cytokine (Fig. 1). Thus, these studiesidentify GAS3 as a target of mda-7/IL-24 both at the RNAand protein levels in mammary tumor cells.
GAS3 expression suppresses the growth of rat mammarytumor cells in monolayer culture and in soft agarWe first showed that growth arrest induced by serum
starvation inducedGAS3 expression in both FE1.2 (28-fold)and FE1.3 (10-fold) cells (Supplementary Fig. S1). Todetermine directly the effect of gas3 expression on FE1.2cell growth, we infected these cells with a retrovirus(MSCV2.1) expressing gas3 (FE1.2þGAS3) or vector con-trol (FE1.2þvector). Western blot analysis confirmed highexpression of GAS3 only in FE1.2þGAS3 cells (Fig. 2A).When cultured in vitro, FE1.2þGAS3 cells displayed areduced ability to proliferate in adherent,monolayer culturescompared with FE1.2þvector cells (Fig. 2B). GAS3 expres-
sion also significantly suppressed the number of colonies thatgrew in soft agar compared with control cells (Fig. 2C).Importantly, gas3-mediated growth inhibition of FE1.2 cellswas also observed in vivo when FE1.2þGAS3 orFE1.2þvector cells were transplanted subcutaneously intonude mice (Fig. 2D).To test for dependency ofmda-7/IL-24–induced suppres-
sion of cell proliferation on gas3, we knocked down gas3
0
100
200
300
A
GAS3
Pan-Cadherin
B
*
CDays
Cell c
ou
nt
(×10
4)
D
Co
lon
y n
um
bers
0
500
1,000
1,500
2,000
2,500
3,000
*
3,500
Tu
mo
r vo
lum
e (
mm
3)
32100
10
20
30
40
50
60GAS3
Vector
*
300
200
100Density
GAS3
β-Actin
0.000.410.571.00
E
GAS3
Vecto
r
FE1.2+
Vecto
r
FE1.2+
GAS3
FE1.2+
Vecto
r
FE1.2+
GAS3
Vecto
r
IL-2
4
IL-2
4+AG9
(10
µmol/L
)
IL-2
4+AG9
(20
µmol/L
)
Figure 2. GAS3 expressionsuppresses growth of FE1.2 cellsin monolayer culture, in soft agar,and in vivo in nude mice. A,Western blot analysis of GAS3expression in FE1.2 cells infectedwith retrovirus-containing GAS3(FE1.2þGAS3) or vector control(FE1.2þvector). B, growth analysisof transduced FEI.2 cells inmonolayer culture. C, number ofcolonies formed in soft agar.D, growth of cells in nude mice,�, P < 0.05. E, transduced FE1.2cells were treated with theindicated doses of the STAT3inhibitor AG9 for 24 hours thensubjected toWestern blot analysis.Band density was determinedusing the ChemiGenius2 BioImaging System and Gene ToolsSoftware.
GAS3
Pan-Cadherin
A
B
FE1.2+
IL-24 AS
FE 1.3FE 1.2 FE1.2+
IL-24 S
0
200
400
600
800
*
*
Gas3 e
xp
ressio
n
(fo
ld c
han
ge)
FE1.3
FE1.2+IL-24 AS
FE1.2+IL-24 S
FE1.2
Figure 1. GAS3 expression in rat mammary tumor cells. A, RT-PCRanalysis of FE1.2 and FE1.3 cells and FE1.2 cells infected with retrovirus-containing mda-7/IL-24 in the sense (FE1.2þIL-24S) or antisense(FE1.2þIL-24AS) orientation, �, P < 0.001. B, GAS3 protein expression inthe indicated cells.
Li et al.
Mol Cancer Res; 11(6) June 2013 Molecular Cancer Research596
on April 23, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst March 6, 2013; DOI: 10.1158/1541-7786.MCR-12-0496
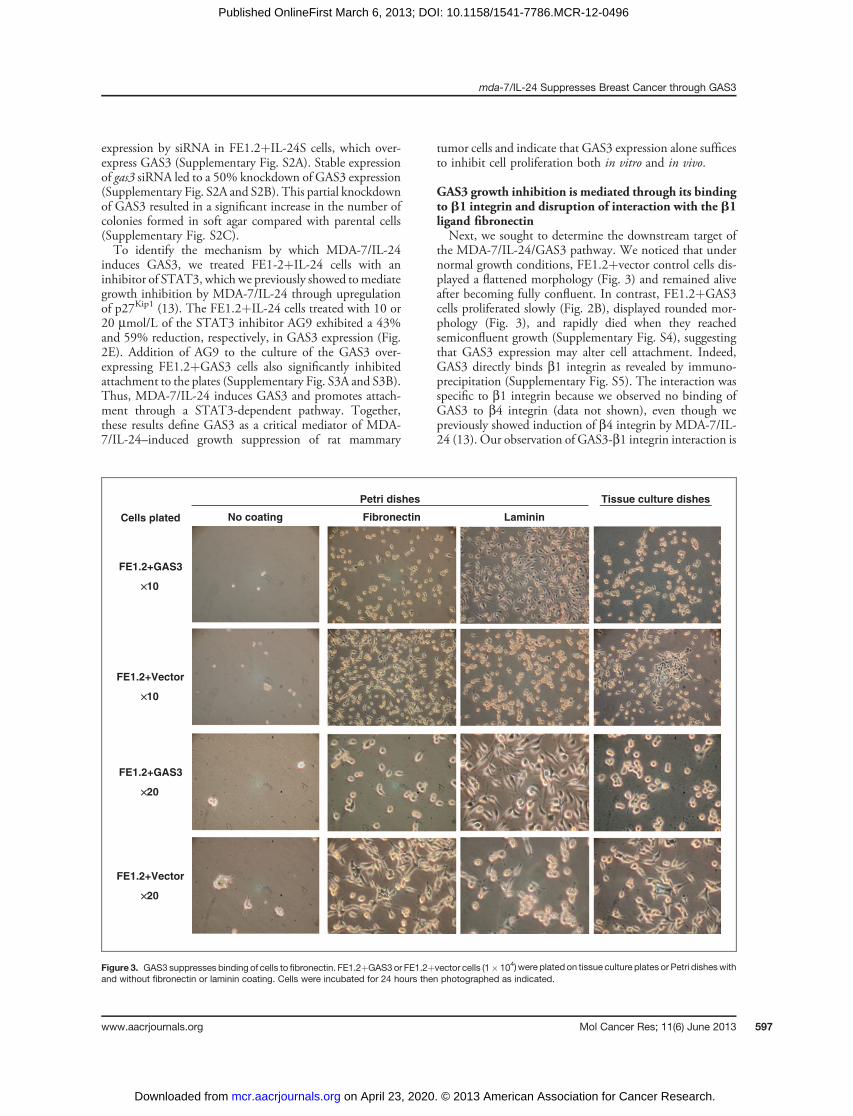
expression by siRNA in FE1.2þIL-24S cells, which over-express GAS3 (Supplementary Fig. S2A). Stable expressionof gas3 siRNA led to a 50% knockdown of GAS3 expression(Supplementary Fig. S2A and S2B). This partial knockdownof GAS3 resulted in a significant increase in the number ofcolonies formed in soft agar compared with parental cells(Supplementary Fig. S2C).To identify the mechanism by which MDA-7/IL-24
induces GAS3, we treated FE1-2þIL-24 cells with aninhibitor of STAT3, which we previously showed tomediategrowth inhibition by MDA-7/IL-24 through upregulationof p27Kip1 (13). The FE1.2þIL-24 cells treated with 10 or20 mmol/L of the STAT3 inhibitor AG9 exhibited a 43%and 59% reduction, respectively, in GAS3 expression (Fig.2E). Addition of AG9 to the culture of the GAS3 over-expressing FE1.2þGAS3 cells also significantly inhibitedattachment to the plates (Supplementary Fig. S3A and S3B).Thus, MDA-7/IL-24 induces GAS3 and promotes attach-ment through a STAT3-dependent pathway. Together,these results define GAS3 as a critical mediator of MDA-7/IL-24–induced growth suppression of rat mammary
tumor cells and indicate that GAS3 expression alone sufficesto inhibit cell proliferation both in vitro and in vivo.
GAS3 growth inhibition is mediated through its bindingto b1 integrin and disruption of interaction with the b1ligand fibronectinNext, we sought to determine the downstream target of
the MDA-7/IL-24/GAS3 pathway. We noticed that undernormal growth conditions, FE1.2þvector control cells dis-played a flattened morphology (Fig. 3) and remained aliveafter becoming fully confluent. In contrast, FE1.2þGAS3cells proliferated slowly (Fig. 2B), displayed rounded mor-phology (Fig. 3), and rapidly died when they reachedsemiconfluent growth (Supplementary Fig. S4), suggestingthat GAS3 expression may alter cell attachment. Indeed,GAS3 directly binds b1 integrin as revealed by immuno-precipitation (Supplementary Fig. S5). The interaction wasspecific to b1 integrin because we observed no binding ofGAS3 to b4 integrin (data not shown), even though wepreviously showed induction of b4 integrin by MDA-7/IL-24 (13). Our observation of GAS3-b1 integrin interaction is
LamininFibronectinNo coating
Petri dishes Tissue culture dishes
Cells plated
FE1.2+GAS3
××10
FE1.2+Vector
××20
FE1.2+GAS3
××20
FE1.2+Vector
××10
Figure 3. GAS3 suppresses binding of cells to fibronectin. FE1.2þGAS3or FE1.2þvector cells (1� 104)were plated on tissue culture plates or Petri disheswithand without fibronectin or laminin coating. Cells were incubated for 24 hours then photographed as indicated.
mda-7/IL-24 Suppresses Breast Cancer through GAS3
www.aacrjournals.org Mol Cancer Res; 11(6) June 2013 597
on April 23, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst March 6, 2013; DOI: 10.1158/1541-7786.MCR-12-0496
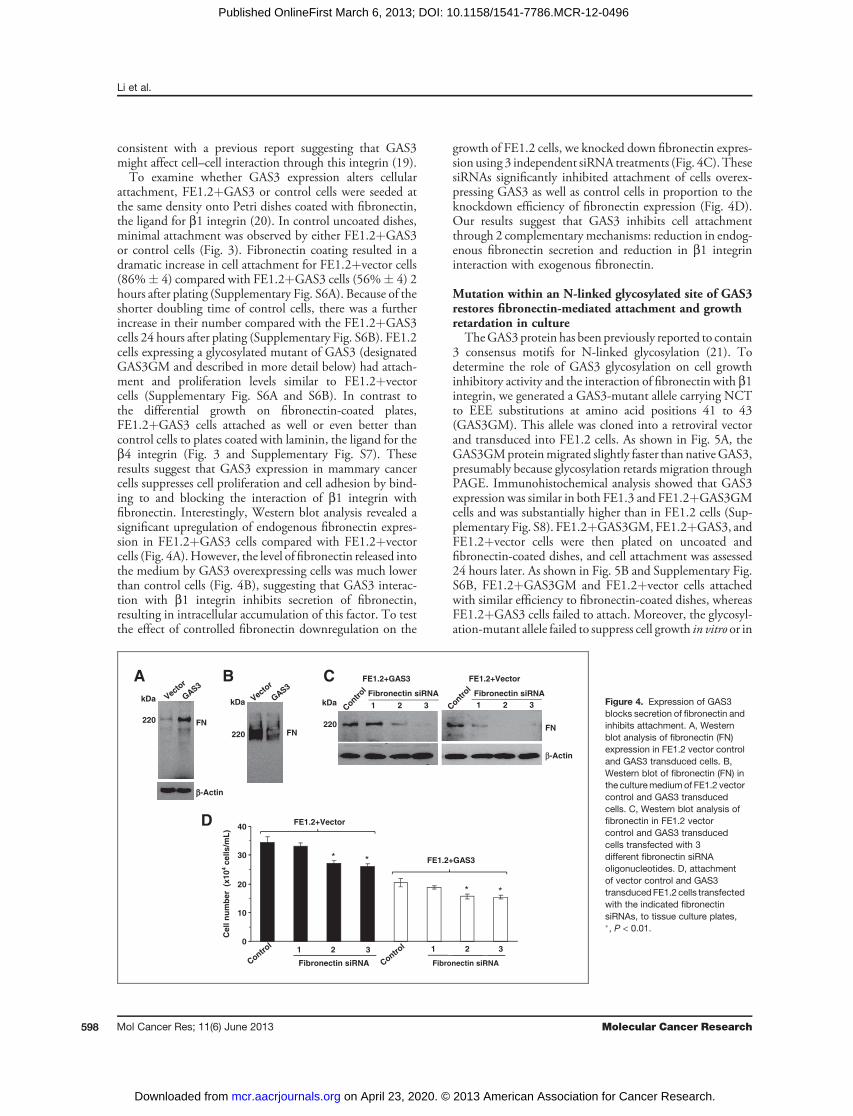
consistent with a previous report suggesting that GAS3might affect cell–cell interaction through this integrin (19).To examine whether GAS3 expression alters cellular
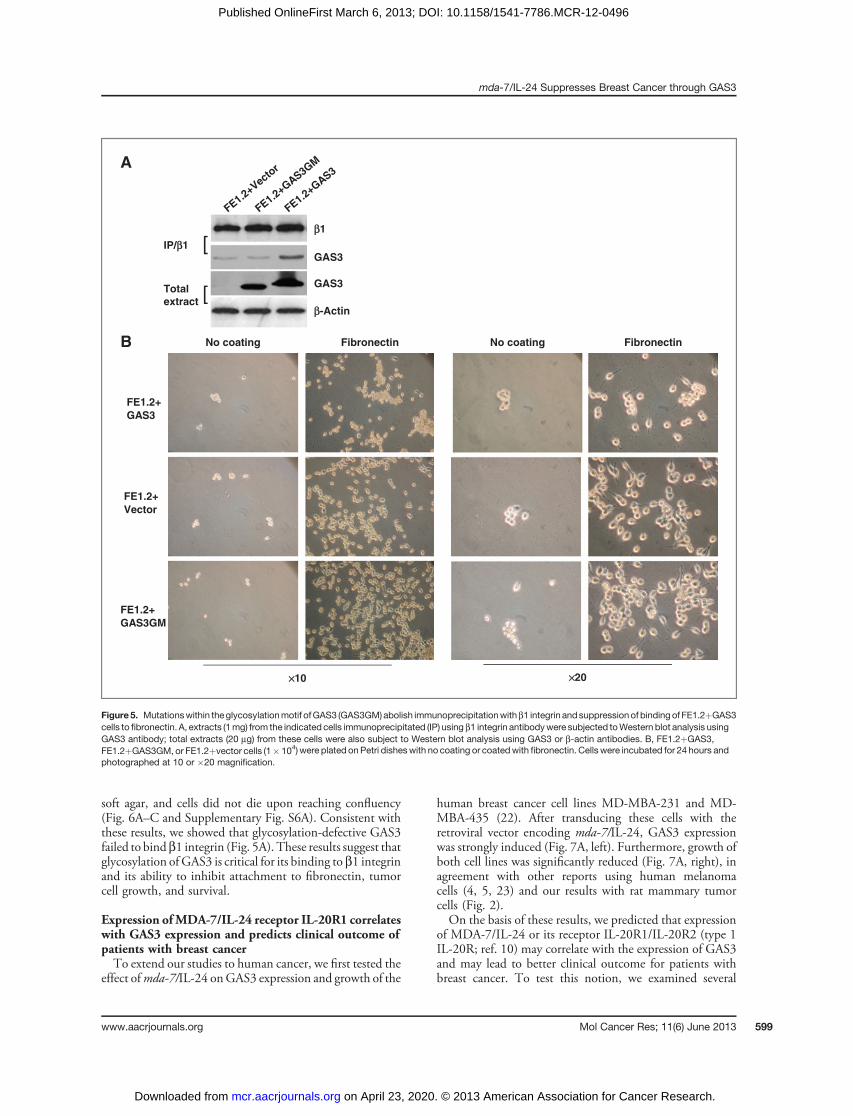
attachment, FE1.2þGAS3 or control cells were seeded atthe same density onto Petri dishes coated with fibronectin,the ligand for b1 integrin (20). In control uncoated dishes,minimal attachment was observed by either FE1.2þGAS3or control cells (Fig. 3). Fibronectin coating resulted in adramatic increase in cell attachment for FE1.2þvector cells(86%� 4) compared with FE1.2þGAS3 cells (56%� 4) 2hours after plating (Supplementary Fig. S6A). Because of theshorter doubling time of control cells, there was a furtherincrease in their number compared with the FE1.2þGAS3cells 24 hours after plating (Supplementary Fig. S6B). FE1.2cells expressing a glycosylated mutant of GAS3 (designatedGAS3GM and described in more detail below) had attach-ment and proliferation levels similar to FE1.2þvectorcells (Supplementary Fig. S6A and S6B). In contrast tothe differential growth on fibronectin-coated plates,FE1.2þGAS3 cells attached as well or even better thancontrol cells to plates coated with laminin, the ligand for theb4 integrin (Fig. 3 and Supplementary Fig. S7). Theseresults suggest that GAS3 expression in mammary cancercells suppresses cell proliferation and cell adhesion by bind-ing to and blocking the interaction of b1 integrin withfibronectin. Interestingly, Western blot analysis revealed asignificant upregulation of endogenous fibronectin expres-sion in FE1.2þGAS3 cells compared with FE1.2þvectorcells (Fig. 4A).However, the level of fibronectin released intothe medium by GAS3 overexpressing cells was much lowerthan control cells (Fig. 4B), suggesting that GAS3 interac-tion with b1 integrin inhibits secretion of fibronectin,resulting in intracellular accumulation of this factor. To testthe effect of controlled fibronectin downregulation on the
growth of FE1.2 cells, we knocked down fibronectin expres-sion using 3 independent siRNA treatments (Fig. 4C). ThesesiRNAs significantly inhibited attachment of cells overex-pressing GAS3 as well as control cells in proportion to theknockdown efficiency of fibronectin expression (Fig. 4D).Our results suggest that GAS3 inhibits cell attachmentthrough 2 complementary mechanisms: reduction in endog-enous fibronectin secretion and reduction in b1 integrininteraction with exogenous fibronectin.
Mutation within an N-linked glycosylated site of GAS3restores fibronectin-mediated attachment and growthretardation in cultureTheGAS3 protein has been previously reported to contain
3 consensus motifs for N-linked glycosylation (21). Todetermine the role of GAS3 glycosylation on cell growthinhibitory activity and the interaction of fibronectin with b1integrin, we generated a GAS3-mutant allele carrying NCTto EEE substitutions at amino acid positions 41 to 43(GAS3GM). This allele was cloned into a retroviral vectorand transduced into FE1.2 cells. As shown in Fig. 5A, theGAS3GMproteinmigrated slightly faster than nativeGAS3,presumably because glycosylation retards migration throughPAGE. Immunohistochemical analysis showed that GAS3expression was similar in both FE1.3 and FE1.2þGAS3GMcells and was substantially higher than in FE1.2 cells (Sup-plementary Fig. S8). FE1.2þGAS3GM, FE1.2þGAS3, andFE1.2þvector cells were then plated on uncoated andfibronectin-coated dishes, and cell attachment was assessed24 hours later. As shown in Fig. 5B and Supplementary Fig.S6B, FE1.2þGAS3GM and FE1.2þvector cells attachedwith similar efficiency to fibronectin-coated dishes, whereasFE1.2þGAS3 cells failed to attach. Moreover, the glycosyl-ation-mutant allele failed to suppress cell growth in vitro or in
ββ-Actin
kDa
220 FN
kDa
220 FN
A
D
* *
0
10
20
30
40
Cell n
um
ber
(x10
4 c
ell
s/m
L)
Fibronectin siRNA Fibronectin siRNA
1 2 31 2 3
B
FN
β-Actin
FE1.2+Vector
Fibronectin siRNA
220
kDaFibronectin siRNA
FE1.2+GAS3
1
C
**
FE1.2+GAS3
FE1.2+Vector
3 2 1 3 2 Vecto
r
GAS3
GAS3
Vector
Control
Control
Contr
ol
Contr
ol
Figure 4. Expression of GAS3blocks secretion of fibronectin andinhibits attachment. A, Westernblot analysis of fibronectin (FN)expression in FE1.2 vector controland GAS3 transduced cells. B,Western blot of fibronectin (FN) inthe culturemediumof FE1.2 vectorcontrol and GAS3 transducedcells. C, Western blot analysis offibronectin in FE1.2 vectorcontrol and GAS3 transducedcells transfected with 3different fibronectin siRNAoligonucleotides. D, attachmentof vector control and GAS3transducedFE1.2 cells transfectedwith the indicated fibronectinsiRNAs, to tissue culture plates,�, P < 0.01.
Li et al.
Mol Cancer Res; 11(6) June 2013 Molecular Cancer Research598
on April 23, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst March 6, 2013; DOI: 10.1158/1541-7786.MCR-12-0496
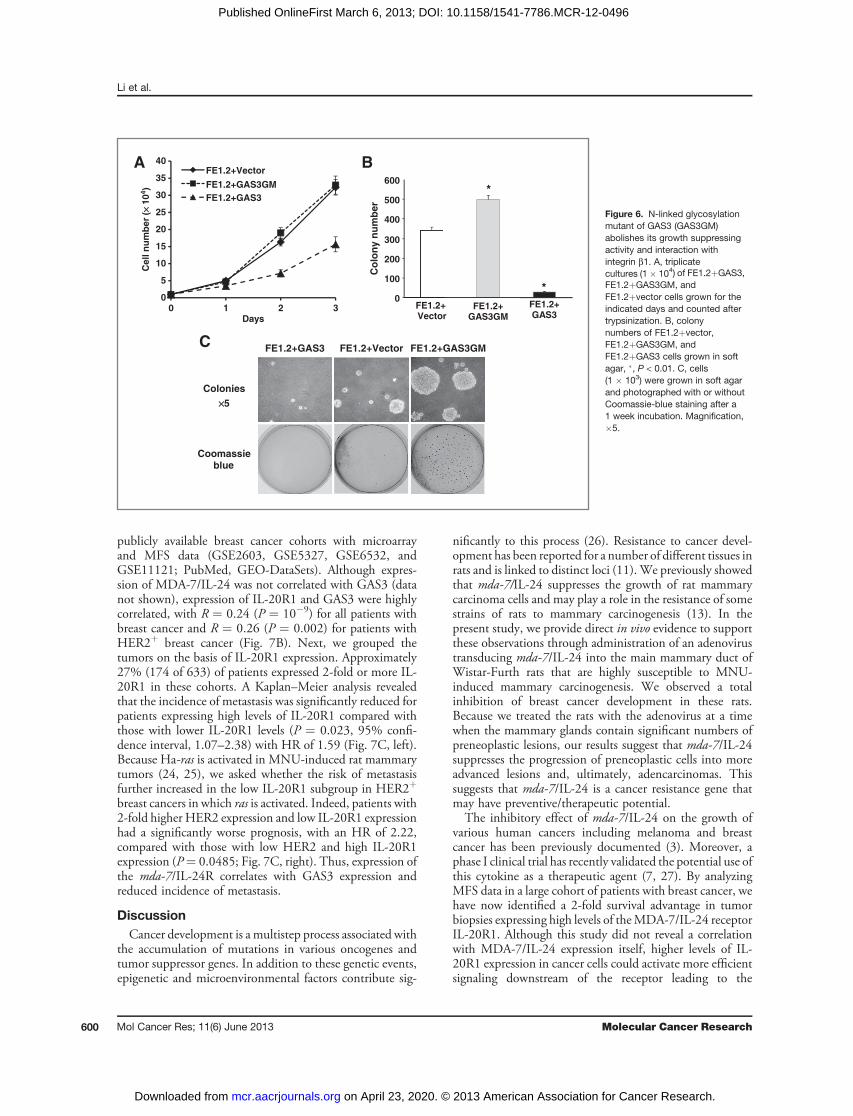
soft agar, and cells did not die upon reaching confluency(Fig. 6A–C and Supplementary Fig. S6A). Consistent withthese results, we showed that glycosylation-defective GAS3failed to bindb1 integrin (Fig. 5A). These results suggest thatglycosylation of GAS3 is critical for its binding to b1 integrinand its ability to inhibit attachment to fibronectin, tumorcell growth, and survival.
Expression ofMDA-7/IL-24 receptor IL-20R1 correlateswith GAS3 expression and predicts clinical outcome ofpatients with breast cancerTo extend our studies to human cancer, we first tested the
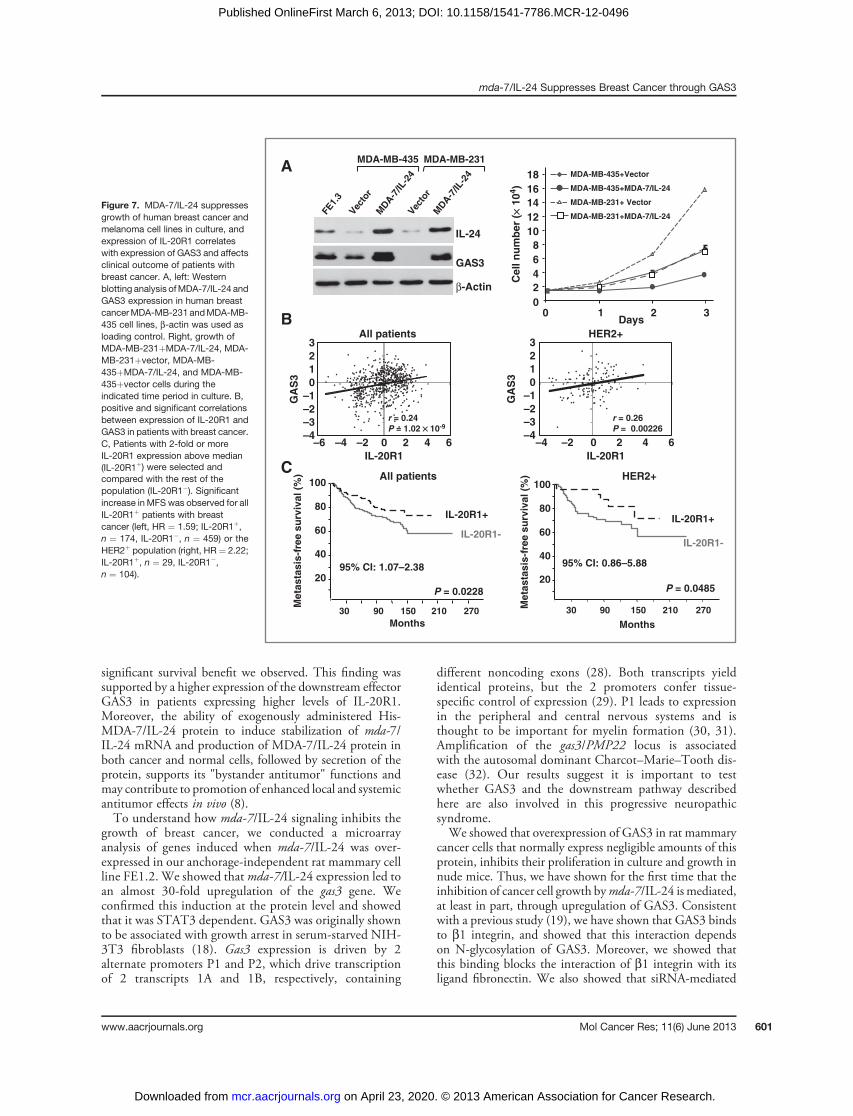
effect ofmda-7/IL-24 onGAS3 expression and growth of the
human breast cancer cell lines MD-MBA-231 and MD-MBA-435 (22). After transducing these cells with theretroviral vector encoding mda-7/IL-24, GAS3 expressionwas strongly induced (Fig. 7A, left). Furthermore, growth ofboth cell lines was significantly reduced (Fig. 7A, right), inagreement with other reports using human melanomacells (4, 5, 23) and our results with rat mammary tumorcells (Fig. 2).On the basis of these results, we predicted that expression
of MDA-7/IL-24 or its receptor IL-20R1/IL-20R2 (type 1IL-20R; ref. 10) may correlate with the expression of GAS3and may lead to better clinical outcome for patients withbreast cancer. To test this notion, we examined several
××10 ××20
No coating
FE1.2+
GAS3
Fibronectin
FE1.2+
GAS3GM
FibronectinNo coating
FE1.2+
Vector
B
A
IP/ββ1
ββ-Actin
GAS3
GAS3Total
extract[
[ββ1
FE1.2+Vecto
r
FE1.2+G
AS3
FE1.2+G
AS3GM
Figure5. Mutationswithin theglycosylationmotif ofGAS3 (GAS3GM)abolish immunoprecipitationwithb1 integrin andsuppressionof bindingof FE1.2þGAS3cells to fibronectin. A, extracts (1mg) from the indicated cells immunoprecipitated (IP) usingb1 integrin antibodywere subjected toWestern blot analysis usingGAS3 antibody; total extracts (20 mg) from these cells were also subject to Western blot analysis using GAS3 or b-actin antibodies. B, FE1.2þGAS3,FE1.2þGAS3GM, or FE1.2þvector cells (1� 104) were plated onPetri disheswith no coating or coatedwith fibronectin. Cells were incubated for 24 hours andphotographed at 10 or �20 magnification.
mda-7/IL-24 Suppresses Breast Cancer through GAS3
www.aacrjournals.org Mol Cancer Res; 11(6) June 2013 599
on April 23, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst March 6, 2013; DOI: 10.1158/1541-7786.MCR-12-0496
publicly available breast cancer cohorts with microarrayand MFS data (GSE2603, GSE5327, GSE6532, andGSE11121; PubMed, GEO-DataSets). Although expres-sion of MDA-7/IL-24 was not correlated with GAS3 (datanot shown), expression of IL-20R1 and GAS3 were highlycorrelated, with R ¼ 0.24 (P ¼ 10�9) for all patients withbreast cancer and R ¼ 0.26 (P ¼ 0.002) for patients withHER2þ breast cancer (Fig. 7B). Next, we grouped thetumors on the basis of IL-20R1 expression. Approximately27% (174 of 633) of patients expressed 2-fold or more IL-20R1 in these cohorts. A Kaplan–Meier analysis revealedthat the incidence of metastasis was significantly reduced forpatients expressing high levels of IL-20R1 compared withthose with lower IL-20R1 levels (P ¼ 0.023, 95% confi-dence interval, 1.07–2.38) with HR of 1.59 (Fig. 7C, left).Because Ha-ras is activated in MNU-induced rat mammarytumors (24, 25), we asked whether the risk of metastasisfurther increased in the low IL-20R1 subgroup in HER2þ
breast cancers in which ras is activated. Indeed, patients with2-fold higher HER2 expression and low IL-20R1 expressionhad a significantly worse prognosis, with an HR of 2.22,compared with those with low HER2 and high IL-20R1expression (P¼ 0.0485; Fig. 7C, right). Thus, expression ofthe mda-7/IL-24R correlates with GAS3 expression andreduced incidence of metastasis.
DiscussionCancer development is a multistep process associated with
the accumulation of mutations in various oncogenes andtumor suppressor genes. In addition to these genetic events,epigenetic and microenvironmental factors contribute sig-
nificantly to this process (26). Resistance to cancer devel-opment has been reported for a number of different tissues inrats and is linked to distinct loci (11). We previously showedthat mda-7/IL-24 suppresses the growth of rat mammarycarcinoma cells and may play a role in the resistance of somestrains of rats to mammary carcinogenesis (13). In thepresent study, we provide direct in vivo evidence to supportthese observations through administration of an adenovirustransducing mda-7/IL-24 into the main mammary duct ofWistar-Furth rats that are highly susceptible to MNU-induced mammary carcinogenesis. We observed a totalinhibition of breast cancer development in these rats.Because we treated the rats with the adenovirus at a timewhen the mammary glands contain significant numbers ofpreneoplastic lesions, our results suggest that mda-7/IL-24suppresses the progression of preneoplastic cells into moreadvanced lesions and, ultimately, adencarcinomas. Thissuggests that mda-7/IL-24 is a cancer resistance gene thatmay have preventive/therapeutic potential.The inhibitory effect of mda-7/IL-24 on the growth of
various human cancers including melanoma and breastcancer has been previously documented (3). Moreover, aphase I clinical trial has recently validated the potential use ofthis cytokine as a therapeutic agent (7, 27). By analyzingMFS data in a large cohort of patients with breast cancer, wehave now identified a 2-fold survival advantage in tumorbiopsies expressing high levels of theMDA-7/IL-24 receptorIL-20R1. Although this study did not reveal a correlationwith MDA-7/IL-24 expression itself, higher levels of IL-20R1 expression in cancer cells could activate more efficientsignaling downstream of the receptor leading to the
A
CFE1.2+GAS3GMFE1.2+GAS3 FE1.2+Vector
××5
Colonies
Coomassieblue
B
FE1.2+GAS3
FE1.2+ Vector
FE1.2+GAS3GM
0
100
200
300
400
500
600
0
5
10
15
20
25
30
35
40
3210
Ce
ll n
um
be
r (×
× 10
4)
Days
*
* FE1.2+GAS3
FE1.2+GAS3GM
FE1.2+Vector
Co
lon
y n
um
ber
Figure 6. N-linked glycosylationmutant of GAS3 (GAS3GM)abolishes its growth suppressingactivity and interaction withintegrin b1. A, triplicatecultures (1� 104) of FE1.2þGAS3,FE1.2þGAS3GM, andFE1.2þvector cells grown for theindicated days and counted aftertrypsinization. B, colonynumbers of FE1.2þvector,FE1.2þGAS3GM, andFE1.2þGAS3 cells grown in softagar, �, P < 0.01. C, cells(1 � 103) were grown in soft agarand photographed with or withoutCoomassie-blue staining after a1 week incubation. Magnification,�5.
Li et al.
Mol Cancer Res; 11(6) June 2013 Molecular Cancer Research600
on April 23, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst March 6, 2013; DOI: 10.1158/1541-7786.MCR-12-0496
significant survival benefit we observed. This finding wassupported by a higher expression of the downstream effectorGAS3 in patients expressing higher levels of IL-20R1.Moreover, the ability of exogenously administered His-MDA-7/IL-24 protein to induce stabilization of mda-7/IL-24 mRNA and production of MDA-7/IL-24 protein inboth cancer and normal cells, followed by secretion of theprotein, supports its "bystander antitumor" functions andmay contribute to promotion of enhanced local and systemicantitumor effects in vivo (8).To understand how mda-7/IL-24 signaling inhibits the
growth of breast cancer, we conducted a microarrayanalysis of genes induced when mda-7/IL-24 was over-expressed in our anchorage-independent rat mammary cellline FE1.2. We showed that mda-7/IL-24 expression led toan almost 30-fold upregulation of the gas3 gene. Weconfirmed this induction at the protein level and showedthat it was STAT3 dependent. GAS3 was originally shownto be associated with growth arrest in serum-starved NIH-3T3 fibroblasts (18). Gas3 expression is driven by 2alternate promoters P1 and P2, which drive transcriptionof 2 transcripts 1A and 1B, respectively, containing
different noncoding exons (28). Both transcripts yieldidentical proteins, but the 2 promoters confer tissue-specific control of expression (29). P1 leads to expressionin the peripheral and central nervous systems and isthought to be important for myelin formation (30, 31).Amplification of the gas3/PMP22 locus is associatedwith the autosomal dominant Charcot–Marie–Tooth dis-ease (32). Our results suggest it is important to testwhether GAS3 and the downstream pathway describedhere are also involved in this progressive neuropathicsyndrome.We showed that overexpression of GAS3 in rat mammary
cancer cells that normally express negligible amounts of thisprotein, inhibits their proliferation in culture and growth innude mice. Thus, we have shown for the first time that theinhibition of cancer cell growth bymda-7/IL-24 ismediated,at least in part, through upregulation of GAS3. Consistentwith a previous study (19), we have shown that GAS3 bindsto b1 integrin, and showed that this interaction dependson N-glycosylation of GAS3. Moreover, we showed thatthis binding blocks the interaction of b1 integrin with itsligand fibronectin. We also showed that siRNA-mediated
Figure 7. MDA-7/IL-24 suppressesgrowth of human breast cancer andmelanoma cell lines in culture, andexpression of IL-20R1 correlateswith expression of GAS3 and affectsclinical outcome of patients withbreast cancer. A, left: Westernblotting analysis ofMDA-7/IL-24 andGAS3 expression in human breastcancerMDA-MB-231 andMDA-MB-435 cell lines, b-actin was used asloading control. Right, growth ofMDA-MB-231þMDA-7/IL-24, MDA-MB-231þvector, MDA-MB-435þMDA-7/IL-24, and MDA-MB-435þvector cells during theindicated time period in culture. B,positive and significant correlationsbetween expression of IL-20R1 andGAS3 in patients with breast cancer.C, Patients with 2-fold or moreIL-20R1 expression above median(IL-20R1þ) were selected andcompared with the rest of thepopulation (IL-20R1�). Significantincrease inMFS was observed for allIL-20R1þ patients with breastcancer (left, HR ¼ 1.59; IL-20R1þ,n ¼ 174, IL-20R1�, n ¼ 459) or theHER2þ population (right, HR ¼ 2.22;IL-20R1þ, n ¼ 29, IL-20R1�,n ¼ 104).
Meta
sta
sis
-fre
e s
urv
ival (%
)
IL-20R1-
IL-20R1+
P = 0.0228
30 90 150 210 270
Months
80
60
40
20
100All patients
95% CI: 1.07–2.38
Me
tas
tas
is-f
ree
su
rviv
al
(%)
210
HER2+
IL-20R1-
IL-20R1+
P = 0.0485
30 90 150 270
Months
80
60
40
20
100
95% CI: 0.86–5.88
A
IL-24
β-Actin
GAS3
MDA-MB-435 MDA-MB-231
C
0
2
4
6
8
10
12
14
16
18
0 1 2 3DaysB
–6 –4–4
–3
–2
–1
0
1
2
3
0 2 4 6–2
All patients
IL-20R1
GA
S3
r = 0.24
P = 1.02 ×× 10-9
–4 –2 0 2 4 6
HER2+
–4
–3
–2
–1
0
1
2
3
IL-20R1
GA
S3
r = 0.26
P = 0.00226
Cell n
um
ber
(×× 1
04)
MDA-7
/IL-2
4
MDA-7
/IL-2
4Vec
tor
FE1.
3
Vec
tor
MDA-MB-435+Vector
MDA-MB-435+MDA-7/IL-24
MDA-MB-231+ Vector
MDA-MB-231+MDA-7/IL-24
mda-7/IL-24 Suppresses Breast Cancer through GAS3
www.aacrjournals.org Mol Cancer Res; 11(6) June 2013 601
on April 23, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst March 6, 2013; DOI: 10.1158/1541-7786.MCR-12-0496

downregulation of fibronectin reduced survival of breastcancer cells, suggesting that binding of GAS3 to b1 integrinplays a critical role in its ability to inhibit cancer cell growth.Our results are consistent with previous observations inwhich peptides containing the integrin recognition sequenceRGD (Arg-Ely-Asp) can inhibit metastasis of mouse mela-noma cells by blocking a4–b1 interaction (21) and that thetumor-suppressive activity of mda-7/IL-24 can be signifi-cantly increased when the RGD sequence is fused in framewith the mda-7/IL-24 gene (33).Surprisingly, we showed that mammary cancer cells
expressing a glycosylation-mutant form of GAS3 bind betterto fibronectin-coated plates and exhibit a higher growth ratein soft agar than cells expressing native GAS3. This addi-tional growth potential may stem from the ability of mutantGAS3 to compete out inhibitory effects of the remainingwild-type GAS3. Drugs that inhibit N-linked glycosylationhave been investigated for the treatment of cancers becauseof their ability to block the function of genes that promotetumor growth (34). Our results, however, showed unex-pectedly, an opposite effect on tumor cell growth aftermutating the glycosylation site of GAS3, suggesting thatspecific activation of GAS3 glycosylation may promotemammary tumor cell growth.In summary, we have shown that signaling mediated
through MDA-7/IL-24 and its receptors inhibits mam-mary tumor growth. We provide evidence that growthinhibition by MDA-7/IL-24 is mediated, at least in part,through activation of GAS3. The association of GAS3 andb1 integrin results in the disruption of the binding of b1to fibronectin, thereby inhibiting cell attachment andproliferation. Our results strengthen the notion thatmda-7/IL-24 has cancer preventive/therapeutic potentialand elucidate downstream tumor suppressor signaling
involving GAS3, b1 integrin, and fibronectin. Finally,our observation that expression of the MDA-7/IL-24receptor correlates with reduced metastasis suggests thattherapeutic activation of components of this pathwaymay improve clinical outcome for patients with breastcancer.
Disclosure of Potential Conflicts of interestP. Fisher has ownership interest (including patents) in the Columbia University.
No potential conflicts of interest were disclosed by the other authors.
Authors' ContributionsConception and design: Y.-J. Li, G. Liu, M.C. Archer, Y. Ben-DavidDevelopment of methodology: Y.-J. Li, G. Liu, Y. Li, B.B. Yang, M.C. Archer,Y. Ben-DavidAcquisition of data (provided animals, acquired and managed patients, providedfacilities, etc.):G. Liu, J.C. Liu, S. Shan, R. Dash, P.B. Fisher, M.C. Archer, Y. Ben-DavidAnalysis and interpretation of data (e.g., statistical analysis, biostatistics, compu-tational analysis): Y.-J. Li, G. Liu, L.M. Vecchiarelli-Federico, J.C. Liu, R. Dash,P.B. Fisher, M.C. Archer, Y. Ben-DavidWriting, review, and/or revision of the manuscript: Y.-J. Li, L.M. Vecchiarelli-Federico, E. Zacksenhaus, R. Dash, P.B. Fisher, M.C. Archer, Y. Ben-DavidAdministrative, technical, or material support (i.e., reporting or organizing data,constructing databases): Y.-J. Li, G. Liu, Q. Li, M.C. Archer, Y. Ben-DavidStudy supervision: E. Zacksenhaus, M.C. Archer, Y. Ben-David
AcknowledgmentsThe authors thank Melanie Suttar for excellent secretarial assistance.
Grant SupportThis study was supported by grants from Canadian Breast Cancer Foundation (to
M.C. Archer, and Y. Ben-David), Canadian Breast Cancer Research Alliance (to E.Zacksenhaus) and NIH R01 CA97318 and P01 CA104177 (to P.B. Fisher).
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be herebymarked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
Received August 17, 2012; revised January 31, 2013; accepted February 19, 2013;published OnlineFirst March 6, 2013.
References1. Perou CM, Sorlie T, EisenMB, van de RijnM, Jeffrey SS, Rees CA, et al.
Molecular portraits of human breast tumours. Nature 2000;406:747–52.2. Sørlie T, Perou CM, Tibshirani R, Aas T, Geisler S, Johnsen H, et al.
Gene expression patterns of breast carcinomas distinguish tumorsubclasses with clinical implications. Proc Natl Acad Sci U S A2001;98:10869–74.
3. DashR, Bhutia SK, AzabB, SuZZ, Quinn BA, KegelmenTP, et al. mda-7/IL-24: a unique member of the IL-10 gene family promoting cancer-targeted toxicity. Cytokine Growth Factor Rev 2010;21:381–91.
4. Jiang H, Su ZZ, Lin JJ, Goldstein NI, Young CS, Fisher PB. Themelanoma differentiation associated gene mda-7 suppresses cancercell growth. Proc Natl Acad Sci U S A 1996;93:9160–5.
5. Jiang H, Lin JJ, Su ZZ, Goldstein NI, Fisher PB. Subtractionhybridization identifies a novel melanoma differentiation associatedgene, mda-7, modulated during human melanoma differentiation,growth and progression. Oncogene 1995;11:2477–86.
6. Su ZZ, Lebedeva IV, Sarkar D, Gopalkrishnan RV, Sauane M, SigmonC, et al. Melanoma differentiation associated gene-7, mda-7/IL-24,selectively induces growth suppression, apoptosis and radiosensiti-zation in malignant gliomas in a p53-independent manner. Oncogene2003;22:1164–80.
7. CunninghamCC,ChadaS,Merritt JA, TongA,SenzerN, ZhangY, et al.Clinical and local biological effects of an intratumoral injection of mda-7 (IL24; INGN 241) in patients with advanced carcinoma: a phase Istudy. Mol Ther 2005;11:149–59.
8. Sauane M, Su ZZ, Gupta P, Lebedeva IV, Dent P, Sarkar D, et al.Autocrine regulation of mda-7/IL-24 mediates cancer-specific apo-ptosis. Proc Natl Acad Sci U S A 2008;105:9763–8.
9. Gupta P, Walter MR, Su ZZ, Lebedeva IV, Emdad L, Randolph A, et al.BiP/GRP78 is an intracellular target for MDA-7/IL-24 induction ofcancer-specific apoptosis. Cancer Res 2006;66:8182–91.
10. Zheng M, Bocangel D, Doneske B, Mhashilkar A, Ramesh R, Hunt KK,et al. Human interleukin 24 (MDA-7/IL-24) protein kills breast cancercells via the IL-20 receptor and is antagonized by IL-10. CancerImmunol Immunother 2007;56:205–15.
11. WoodGA,Korkola JE, ArcherMC.Tissue-specific resistance to cancerdevelopment in the rat: phenotypes of tumor-modifier genes. Carci-nogenesis 2002;23:1–9.
12. Li YJ, Song R, Korkola JE, Archer MC, Ben-David Y. Cyclin D1 isnecessary but not sufficient for anchorage-independent growth ofrat mammary tumor cells and is associated with resistance of theCopenhagen rat to mammary carcinogenesis. Oncogene 2003;22:3452–62.
13. Xuan W, Li YJ, Liu G, Ben-David Y, Archer MC. Interleukin-24 inducesexpression of beta4 integrin but suppresses anchorage-independentgrowth of rat mammary tumor cells by a mechanism that is indepen-dent of beta4. Mol Cancer Res 2009;7:433–42.
14. Pareek S, Notterpek L, Snipes GJ, Naef R, Sossin W, Lalibert�e J, et al.Neurons promote the translocation of peripheral myelin protein 22 intomyelin. J Neurosci 1997;17:7754–62.
Li et al.
Mol Cancer Res; 11(6) June 2013 Molecular Cancer Research602
on April 23, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst March 6, 2013; DOI: 10.1158/1541-7786.MCR-12-0496

15. Shan SW, Lee DY, Deng Z, Shatseva T, Jeyapalan Z, Du WW, et al.MicroRNA MiR-17 retards tissue growth and represses fibronectinexpression. Nat Cell Biol 2009;11:1031–8.
16. Liu JC, Voisin V, Bader GD, Deng T, Pusztai L, Symmans WF, et al.Seventeen-gene signature from enriched Her2/Neu mammarytumor-initiating cells predicts clinical outcome for human HER2þ:ERalpha- breast cancer. Proc Natl Acad Sci U S A 2012;109:5832–7.
17. Korkola JE, Archer MC. Resistance to mammary tumorigenesis inCopenhagen rats is associated with the loss of preneoplastic lesions.Carcinogenesis 1999;20:221–7.
18. Karlsson C, Afrakhte M, Westermark B, Paulsson Y. Elevated level ofgas3 gene expression is correlated with G0 growth arrest in humanfibroblasts. Cell Biol Int 1999;23:351–8.
19. Wadehra M, Iyer R, Goodglick L, Braun J. The tetraspan proteinepithelial membrane protein-2 interacts with beta1 integrins and reg-ulates adhesion. J Biol Chem 2002;277:41094–100.
20. Hynes RO. Integrins: bidirectional, allosteric signaling machines. Cell2002;110:673–87.
21. Danen EH, Marcinkiewicz C, Cornelissen IM, van Kraats AA, PachterJA, Ruiter DJ, et al. The disintegrin eristostatin interferes with integrinalpha 4 beta 1 function and with experimental metastasis of humanmelanoma cells. Exp Cell Res 1998;238:188–96.
22. Chambers AF. MDA-MB-435 and M14 cell lines: identical but not M14melanoma? Cancer Res 2009;69:5292–3.
23. Lebedeva IV, Su ZZ, Chang Y, Kitada S, Reed JC, Fisher PB.The cancer growth suppressing gene mda-7 induces apoptosisselectively in human melanoma cells. Oncogene 2002;21:708–18.
24. Lu SJ, Archer MC. Ha-ras oncogene activation in mammary glandsof N-methyl-N-nitrosourea-treated rats genetically resistant tomammary adenocarcinogenesis. Proc Natl Acad Sci U S A 1992;89:1001–5.
25. Sukumar S, Notario V, Martin-Zanca D, Barbacid M. Induction ofmammary carcinomas in rats by nitroso-methylurea involves malig-nant activation of H-ras-1 locus by single point mutations. Nature1983;306:658–61.
26. Ponder BA. Cancer genetics. Nature 2001;411:336–41.27. TongAW,Nemunaitis J, SuD,ZhangY,CunninghamC,SenzerN, et al.
Intratumoral injection of INGN 241, a nonreplicating adenovectorexpressing the melanoma-differentiation associated gene-7 (mda-7/IL24): biologic outcome in advanced cancer patients. Mol Ther2005;11:160–72.
28. Suter U, Snipes GJ, Schoener-Scott R, Welcher AA, Pareek S, LupskiJR, et al. Regulation of tissue-specific expression of alternative periph-eral myelin protein-22 (PMP22) gene transcripts by two promoters.J Biol Chem 1994;269:25795–808.
29. Hai M, Bidichandani SI, Patel PI. Identification of a positive regulatoryelement in the myelin-specific promoter of the PMP22 gene. J Neu-rosci Res 2001;65:508–19.
30. Garbay B, Boiron-Sargueil F, Cassagne C. Expression of the exon 1A-containing PMP22 transcript is altered in the tremblermouse. NeurosciLett 1995;198:157–60.
31. Ohsawa Y, Murakami T, Miyazaki Y, Shirabe T, Sunada Y. Peripheralmyelin protein 22 is expressed in human central nervous system.J Neurol Sci 2006;247:11–5.
32. ValentijnLJ,BolhuisPA,ZornI,HoogendijkJE,vandenBoschN,HenselsGW, et al. The peripheral myelin gene PMP-22/GAS-3 is duplicated inCharcot-Marie-Tooth disease type 1A. Nat Genet 1992;1:166–70.
33. Zhang BF, Liu JJ, Pei DS, Yang ZX, Di JH, Chen FF, et al. Potentantitumor effect elicited by RGD-mda-7, an mda-7/IL-24 mutant, viatargeting the integrin receptor of tumor cells. Cancer Biother Radio-pharm 2011;26:647–55.
34. Contessa JN, Bhojani MS, Freeze HH, Rehemtulla A, Lawrence TS.Inhibition of N-linked glycosylation disrupts receptor tyrosine kinasesignaling in tumor cells. Cancer Res 2008;68:3803–9.
mda-7/IL-24 Suppresses Breast Cancer through GAS3
www.aacrjournals.org Mol Cancer Res; 11(6) June 2013 603
on April 23, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst March 6, 2013; DOI: 10.1158/1541-7786.MCR-12-0496

2013;11:593-603. Published OnlineFirst March 6, 2013.Mol Cancer Res You-Jun Li, Guodong Liu, Yanmei Li, et al.
1 Integrin FunctionβDisruption of ) andgas3Upregulation of Growth Arrest-Specific Gene 3 (
7/IL-24 Expression Inhibits Breast Cancer throughmda-
Updated version
10.1158/1541-7786.MCR-12-0496doi:
Access the most recent version of this article at:
Material
Supplementary
http://mcr.aacrjournals.org/content/suppl/2013/03/06/1541-7786.MCR-12-0496.DC1
Access the most recent supplemental material at:
Cited articles
http://mcr.aacrjournals.org/content/11/6/593.full#ref-list-1
This article cites 34 articles, 12 of which you can access for free at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://mcr.aacrjournals.org/content/11/6/593To request permission to re-use all or part of this article, use this link
on April 23, 2020. © 2013 American Association for Cancer Research. mcr.aacrjournals.org Downloaded from
Published OnlineFirst March 6, 2013; DOI: 10.1158/1541-7786.MCR-12-0496









![[IV] The Role of Chromatin Structure in Control of Gene Expression Overview of levels of control of gene expression Chromatin structure in active or potentially](https://img.pdfslide.net/doc/110x75/56649e9e5503460f94ba080b/iv-the-role-of-chromatin-structure-in-control-of-gene-expression-overview.jpg)


















