Embed Size (px)
Citation preview

/. exp Biol. 158, 275-289 (1991) 275Printed in Great Britain © The Company of Biologists Limited 1991
CIRCULATORY MECHANICS IN THE TOAD BUFO MARINUSI. STRUCTURE AND MECHANICAL DESIGN OF THE AORTA
BY CAROL A. GIBBONS1 AND ROBERT E. SHADWICK2*1 Department of Biology, University of Calgary, Calgary, Alberta, Canada,
T2N1N4 and 2Marine Biology Research Division A-004,Scripps Institution of Oceanography, La Jolla, CA 92093, USA
Accepted 28 March 1991
SummaryThis study describes several important mechanical design features of the aorta
of a typical poikilothermic vertebrate. A strong functional similarity to the aortaof mammals is apparent, but some structural and mechanical differences are seenthat reflect the lower pressure and simpler haemodynamics of the poikilothermiccirculation.
1. The aorta is highly distensible, resilient and non-linearly elastic, giving it therequisite properties to act as an effective storage element in the arterialcirculation.
2. An abrupt transition from high compliance (low elastic modulus) torelatively low compliance (high elastic modulus) takes place at pressures above theresting physiological range of 2-4 kPa. This behaviour reflects the compositenature of the artery wall in which rubbery elastin fibres and relatively rigidcollagen fibres are the predominant elements.
3. The longitudinal tethering of the aorta when inflated is due primarily toanisotropy in elastic properties, rather than to links to the axial skeleton by branchvessels or connective tissue.
4. No significant changes in elastic properties or connective tissue content occuralong the length of the toad arterial tree, in contrast to the situation in mammals.
Introduction
The aorta is an important elastic element in the arterial circulation. The vesselwall expands during systole and recoils during diastole, thereby storing andreleasing elastic strain energy and smoothing the pulsatile output of the heart. Animportant benefit is the reduction of the arterial pressure pulse and, consequently,protection of the small exchange vessels from the high shear forces associated withpulsatile flow, and a reduction of the total energy required to circulate the blood(Taylor, 1964). To perform this function effectively, the aorta must have non-
*To whom reprint requests should be addressed,
rftey words: aorta, elasticity, circulatory system, mechanical design, Bufo marinus.

276 C. A . GIBBONS AND R. E . SHADWICK
linear elastic properties in order to be a compliant reservoir at low pressures but bestiff enough to resist rupture at high pressures.
Most of our information on structure and mechanical properties of arteriescomes from studies on mammalian tissues (Bergel, 1961; Milnor, 1982). The arterywall is a composite structure whose non-linear elastic properties result from thecombination of rubber-like elastic and relatively inextensible collagen fibres. Thetransition from a highly compliant to a relatively stiff vessel takes place at about10-12 kPa, the mean resting blood pressure. In mammals, the distensibility of theaorta decreases as the ratio of collagen to elastin increases along the arterial tree.The elastic properties of the artery wall are important in determining thehaemodynamic behaviour of the arterial system.
Poikilothermic vertebrates generally have much lower blood pressures andoften lower heart rates than do mammals (Shelton and Jones, 1968; Jones et al.191 A; Shelton and Burggren, 1976; Langille and Jones, 1977). Nevertheless, theaortas of reptiles, amphibians and fish appear to have non-linear elastic properties(Goto and Kimoto, 1966; Satchell, 1971; Gibbons and Shadwick, 1989) that arecomparable to those observed in mammals, but presumably appropriate forfunction in a lower-pressure system. However, very little is known about therelationship between structure, connective tissue composition and the elasticproperties of the aorta in any lower vertebrate species. The purpose of thisinvestigation is to make a detailed study of the aortic mechanical properties andtheir structural basis in the toad Bufo marinus, and to compare the design featuresof this arterial system to that of mammals. In a subsequent study (Gibbons andShadwick, 1991), the effects of arterial elasticity on haemodynamic properties inthe toad will be examined.
Materials and methods
Animals
The experiments were performed on the toad, Bufo marinus L. Animalsweighing 200-500 g were maintained in tanks at room temperature with access towater. They were fed mealworms weekly. The animals were killed by injection ofMS-222 into the peritoneal cavity (Sandoz, 1:1000, 0.022 ml g~x body mass). Theheart, aortic arches and dorsal aorta were exposed by a ventral midline incision.Four regions were arbitrarily designated, as shown in Fig. 1, and their in situlengths determined before excision of the whole vessel. Experiments wereperformed on these aortic segments at room temperature within a few hours ofdeath.
Mechanical testing
Artery segments (about 5 cm long) were placed in a chamber containingamphibian saline, and cannulated at one end with a blunt 18-gauge syringe needleand connectors leading to a pressure reservoir and a variable-speed pump. Oncethe air bubbles had been cleared, the distal end and the branch segments we i^

Mechanics of the toad aorta 277
Subclavianartery
Site II
Site III
Site IV
Coeliacomesentericartery
Sciatic artery
Fig. 1. Diagram of the aorta and associated branches in the toad, Bufo marinus,approximately to scale. Four testing sites were used and are labelled as shown. Site I isthe proximal aortic arch, from the branching of the truncus arteriosus to the subclavianartery. Site II is the distal aortic arch, between the subclavian artery and thecoeliacomesenteric artery. Site in is the proximal dorsal aorta, from the coeliaco-mesenteric artery to the urogenital arteries. Site IV is the distal dorsal aorta, betweenthe urogenital arteries and the sciatic arteries. The distance from the beginning of thesystemic arch to the sciatic artery in a 350g animal is approximately 11 cm.
ligated, and the vessel was extended to its in vivo length. The preparations weregenerally leak-free. Slow inflation-deflation cycles (lasting 1-2 min) were per-formed using a variable-speed pump. Conditioning cycles (usually 2-3) were rununtil the pressure-diameter curves were stable. The pressure during one cycle wasraised to approximately lOkPa and lowered back to zero. Pressure was measuredcontinuously using a P23Db Gould pressure transducer, while diameter wasdetermined simultaneously with a video dimension analyzer system or VDA(Instrumentation for Physiology and Medicine, model 303), as described by Fung(1981). Some inflation tests were also performed on untethered vessel segments tomeasure the effect of increasing pressure on longitudinal extension. In this case,lengths were measured for step-wise pressure increments by using a microscopedigital micrometer (Wild-Leitz MMS235).
Pressure and radius data were collected on-line by a PDP11/23 laboratorycomputer (Digital Equipment Corporation) for the inflation-deflation cycles atfour positions along the vessel from the arches to the sciatic arteries (Fig. 1).Values for radius at zero pressure were taken when the artery was stretched and
it was unstretched. Frozen-cut sections were made of each vessel segment

278 C. A. GIBBONS AND R. E. SHADWICK
after the inflation tests. From these, wall thickness and internal radius for theunstretched, unpressurized condition were measured using the digital micrometer.Assuming that the vessel wall is a constant-volume material, the internal radiusand wall thickness could then be calculated at each pressure from the measuredexternal radius and length. With these values, the luminal volume and circumfer-ential stress, strain and elastic modulus were calculated at 0.5 kPa intervals foreach cycle and position, using the laboratory computer. Circumferential wall stresswas defined as:
a = Pr/h, (1)
where r is the inside radius, h is the wall thickness and P is the pressure. Thecircumferential strain was calculated at mid-wall radius as:
e = AR/Ro , (2)
where R=(R+r)/2, R is the outer wall radius and Ro is the unstressed mid-wallradius. The elastic modulus describes the relationship between stress and strainand is a measure of material stiffness. For non-linear materials, such as the arterywall, the modulus varies with the level of strain. We used an incremental formulato calculate the elastic modulus (E) from biaxial stress-strain data obtained atconstant vessel length (Bergel, 1961; Dobrin, 1983):
£ = ( l - ^ ) ( l + e)(Aa/Ae), (3)
where fj. is the Poisson ratio, assumed to be 0.5 (see Dobrin, 1983). This formulauses an incremental strain that is based on the average radius at each pressureincrement, and is therefore equivalent to e/(l + e) (Shadwick and Gosline, 1985).Pressure-strain, stress-strain and modulus-pressure relationships for each aorticposition were calculated as mean curves from data pooled from several animals.Standard errors were calculated, and the mean curves were compared at each0.5 kPa interval using multiple comparison tests to determine whether theydiffered significantly (Zar, 1984).
Uniaxial force-extension tests were made on a tensile testing machine (Mon-santo Tensometer T10) in both the circumferential and longitudinal directions. Forthe circumferential testing, arterial rings were cut with 2 mm widths. They wereplaced around two metal L-shaped hooks. One hook was anchored at the base,while the other was connected to the force transducer attached to the moveablehead. Longitudinal testing was done on vessel segments (2-3 cm long) held in vice-type clamps in the tensometer. Stress was then calculated as o=F/A, where F isthe tensile force applied and A is the tissue cross-sectional area perpendicular tothe force. Length changes (AL) were measured between surface markers by thevideo dimension analyzer. The markers were placed in the central region of thespecimen to avoid any clamp effects. Strain was calculated as e=AL/L0, where Lo
was the initial length.
Connective tissue contentFlistological sections were made from vessel segments fixed in Bouin's solutio^

Mechanics of the toad aorta 279
(Humason, 1979) at constant pressures ranging from 0 to 4kPa. Sections werestained with aldehyde-fuchsin or Verhoeff s stain to identify elastin fibres. Picro-ponceau or Mallory's Trichrome stain were used to identify collagen and muscle(Humason, 1979). The content and arrangement of the collagen and elastin werethen examined in both the transverse and longitudinal directions. Sections werestained from arterial segments at the four arterial sites where inflation testswere made. For comparison, a fresh rat aorta was fixed at OkPa and stained usingthe same methods. To determine the smooth muscle disposition of the aortic wall,sections were stained with haematoxylin and counterstained with eosin.
The elastin content was determined by Lansing's method. Freeze-dried vesselsegments were rehydrated and then treated with 0.1 moll"1 sodium hydroxide for45min at 100 °C, followed by subsequent washings in distilled water. The residuewas then dried and weighed (Lansing et al. 1952). The elastin content was thisfraction's dry mass expressed as a percentage of the total dry mass. Collagencontent was estimated from the hydroxyproline content, which was determinedwith an automated amino acid analyser (LKB Alpha Plus, model 1215) or by amanual colorimetric assay (Berg, 1982). Values for collagen content werecorrected to account for the hydroxyproline present in the elastin according topublished information on the composition of elastin in Amphibia (Sage and Gray,1979). The collagen content was calculated as a percentage of the total dry mass. Acollagen to elastin ratio was also calculated.
ResultsMechanical properties
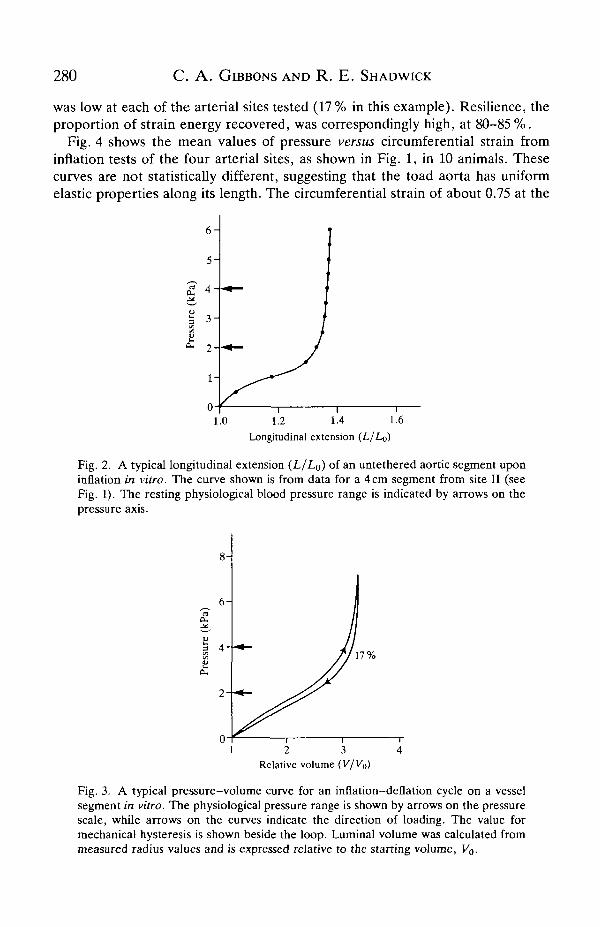
Upon removal from the animal, the aorta shortened by approximately 30% ineach of the designated regions, causing a corresponding increase in diameter.Interestingly, this is comparable to values reported by Bergel (1961) for the dogthoracic and abdominal aorta, which showed shortening of 32-34%. When leftuntethered during an inflation, the length increased with distending pressure untilthe upper physiological pressure was reached (Fig. 2). At this point, the aorta hadregained its in vivo length. Increasing the pressure beyond the physiological rangeresulted in virtually no further lengthening. The vessels were stretched to the invivo length for subsequent inflation tests.
A typical inflation cycle (Fig. 3) illustrates the non-linear and viscoelasticbehaviour of the aorta. Compliance is relatively high up to the physiologicalpressure range (2-4 kPa), but thereafter the slope of the pressure-volume curveincreases sharply as the artery wall becomes very stiff. The difference between theinflation and deflation portions of this curve indicates that some viscous energylosses occur during each cycle. The area under the ascending curve represents thetotal work done on inflation, while the area under the descending curve representsthe strain energy recovered by elastic recoil. Hysteresis, the proportion of energylost through viscous processes during each cycle, is the ratio of the area within the
Dp to the total area beneath the inflation limb. For the toad aorta, the hysteresis
280 C. A . GIBBONS AND R. E . SHADWICK
was low at each of the arterial sites tested (17% in this example). Resilience, theproportion of strain energy recovered, was correspondingly high, at 80-85 %.
Fig. 4 shows the mean values of pressure versus circumferential strain frominflation tests of the four arterial sites, as shown in Fig. 1, in 10 animals. Thesecurves are not statistically different, suggesting that the toad aorta has uniformelastic properties along its length. The circumferential strain of about 0.75 at the
6 -
5 -
3-
2-
1-
1.0 1.2 1.4 1.6Longitudinal extension (L/Lo)
Fig. 2. A typical longitudinal extension (L/Lo) of an untethered aortic segment uponinflation in vitro. The curve shown is from data for a 4cm segment from site II (seeFig. 1). The resting physiological blood pressure range is indicated by arrows on thepressure axis.
6-
17%
1 2 3 4Relative volume (V/K»)
Fig. 3. A typical pressure-volume curve for an inflation-deflation cycle on a vesselsegment in vitro. The physiological pressure range is shown by arrows on the pressurescale, while arrows on the curves indicate the direction of loading. The value formechanical hysteresis is shown beside the loop. Luminal volume was calculated frommeasured radius values and is expressed relative to the starting volume, Vo.

Mechanics of the toad aorta 281
10-
8-
i* 6-a -3
<S 4 -
2 -
o-
I T 1 1
III | IV
I I I I I I 1 I 1 1
0.2 0.4 0 6
Circumferential strain
0.8 1.0
Fig. 4. Pressure-circumferential strain plots from the four sites along the aortaindicated in Fig. 1. Mean values for strain at each pressure increment are plotted fromdata for 10 animals. The standard errors for each of these plots are also shown. Thereare no significant differences between the curves at the four testing sites, as indicatedby the Mest.
mean physiological pressure of 3 kPa is comparable to the level of strain in the ratthoracic aorta at its mean blood pressure of l lkPa (Cox, 1978).
Stress-strain relationships, calculated from the data in Fig. 4, were all J-shapedand also showed no significant differences among the four arterial sites. The levelof circumferential wall stress attained (approximately lxlf^Pa) at mean bloodpressure was nearly equal to that in the aorta of the rat at its much higher meanblood pressure (Gibbons and Shadwick, 1989).
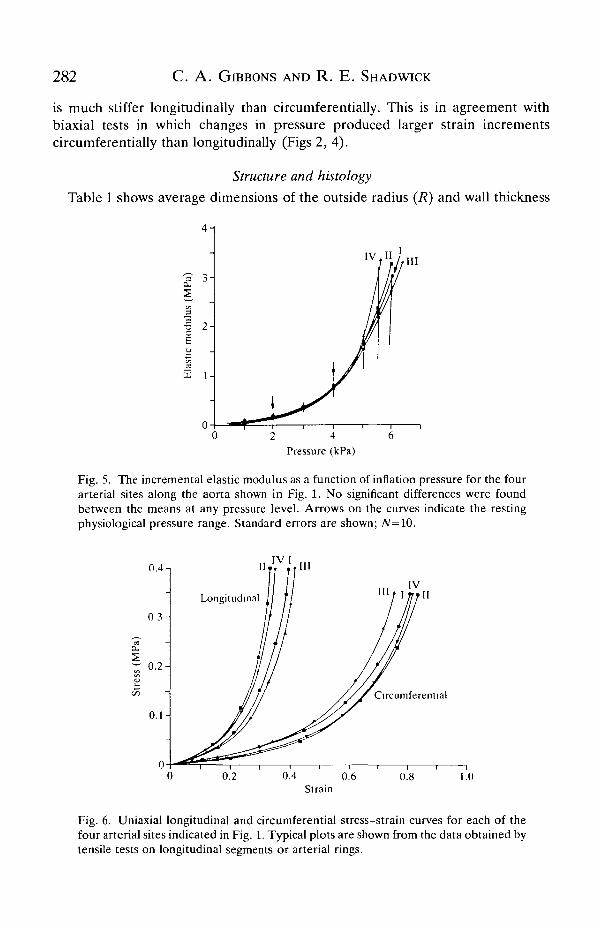
Fig. 5 shows the incremental elastic modulus as a function of distendingpressure. These plots demonstrate that the arterial wall stiffness is highlydependent on pressure and increases considerably over the physiological range,from about 0.2 to 0.7 MPa. At the mean resting blood pressure of 3 kPa, the elasticmodulus is approximately 0.4 MPa. This is similar to values reported for the aortaof other poikilotherms (Gibbons and Shadwick, 1989) as well as for the aorta andupper thoracic aorta of mammals (See Fig. 10) at their respective blood pressures.The similarity of the four plots in Fig. 5 indicates that the toad aorta, unlike that ofmammals, has uniform elastic properties along its entire length.
Potential vascular muscle activators, such as acetylcholine and epinephrine(West and Burrgren, 1984), were applied to the arterial wall and perfused throughthe vessel in an attempt to stimulate muscle activity. These tests were made withina few minutes of death. Although many different concentrations of these drugswere tested, the dorsal aorta showed no measurable mechanical response to thesechemicals.
The stress-strain curves resulting from uniaxial tests (Fig. 6) show that the aorta
282 C. A . GIBBONS AND R. E . SHADWICK
is much stiffer longitudinally than drcumferentially. This is in agreement withbiaxial tests in which changes in pressure produced larger strain incrementscircumferentially than longitudinally (Figs 2, 4).
Structure and histology
Table 1 shows average dimensions of the outside radius (R) and wall thickness
2 4 6Pressure (kPa)
Fig. 5. The incremental elastic modulus as a function of inflation pressure for the fourarterial sites along the aorta shown in Fig. 1. No significant differences were foundbetween the means at any pressure level. Arrows on the curves indicate the restingphysiological pressure range. Standard errors are shown; JV=10.
0.4-!
0 3 -
0.2-
0 . 1 -
IV I
I I I ,IV
0.4 0.6Strain
Fig. 6. Uniaxial longitudinal and circumferential stress-strain curves for each of thefour arterial sites indicated in Fig. 1. Typical plots are shown from the data obtained bytensile tests on longitudinal segments or arterial rings.

Mechanics of the toad aorta 283
Table 1. Average values for radius (R), wall thickness (h) and the R/h ratio at amean pressure of 3 kPa from six different animals at the four arterial sites
Site
I
ninIV
R(mm)
1.341.231.291.20
h(mm)
0.0570.0540.0580.051
R/h
23.522.822.223.5
Elastin(%)
15151414
Collagen(%)
34353535
Elastin and collagen contents are shown as a percentage of total dry mass along the aorta.Each value is the mean of four determinations, each containing arterial segments pooled from
at least 10 animals.
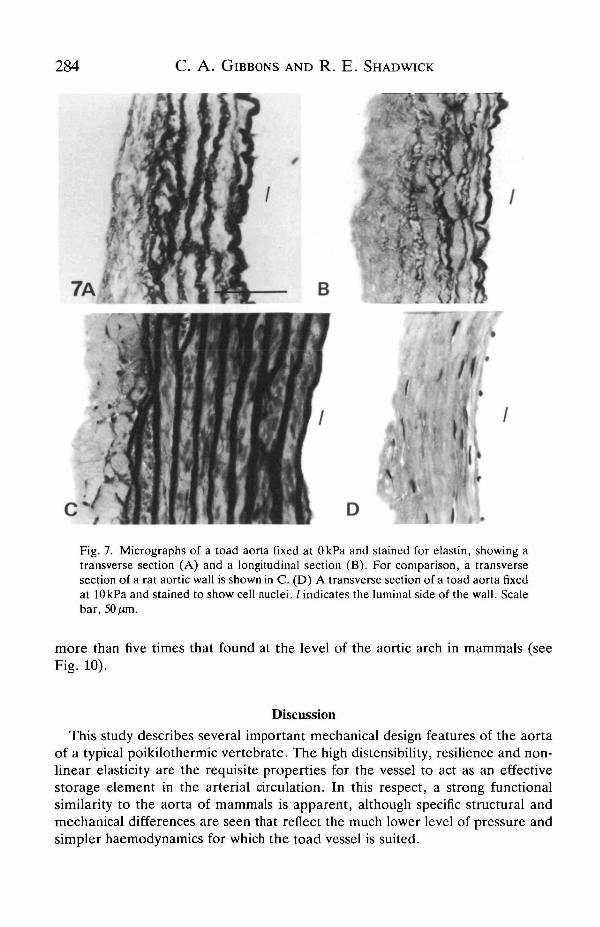
(h) of the toad aorta at a mean pressure of 3 kPa. Both R and h are important indetermining the mechanical properties of the arterial wall. This aorta is surpris-ingly uniform in these dimensions and in the R/h ratio along its length, i.e. there isvirtually no 'geometric taper' as there is in the mammalian aorta (see Fig. 9). Theaverage radius of 1.27 mm and wall thickness of 0.055 mm give an R/h ratio of 23 atmean blood pressure. In contrast, the R/h ratio for a mammalian thoracic aorta atmean blood pressure is only about 10 (McDonald, 1974). The relative thinness ofthe toad aorta is illustrated in Fig. 7; while the diameter of the aorta from the toadand a similar-sized rat are about the same, the wall of the former is only half asthick as that of the latter.
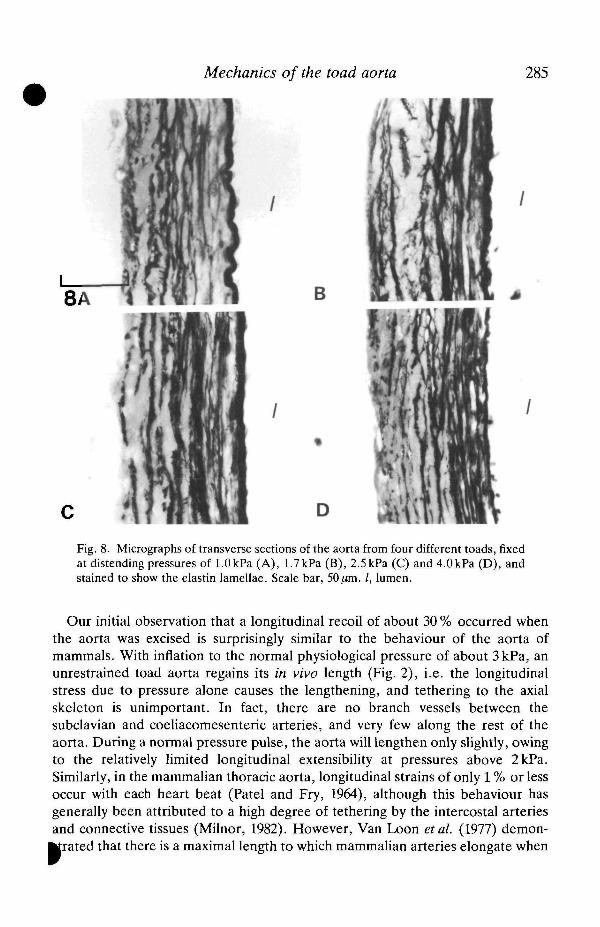
In both the transverse and longitudinal sections of the toad aorta, the elastin(stained black) appears as 4-5 layers within the wall (Fig. 7). These sectionstogether show that the elastin must be arranged as concentric cylindrical sheets. Inuninflated vessels, the luminal elastica is thicker and wavier than the other elastinlayers found throughout the wall. Smooth muscle cells, indicated by the stainednuclei in Fig. 7D, are not very abundant. In both transverse and longitudinalplanes, the elastin layers were relatively thin and sometimes appeared incompleteor fused to adjacent layers. In a rat aorta of similar diameter, the elastin lamellaewere more numerous (9-11), more uniform and well-defined, and thicker than inthe toad aorta (Fig. 7). In the latter there is no distinct outer layer of collagen;rather, it occurs in large amounts among the elastin lamellae. There is relativelyless elastin and more collagen in the arterial wall of the toad than in that of the rat.Fig. 8 shows sections of toad aorta fixed at pressures of 1.0, 1.7, 2.5 and 4.0 kPa.With increasing pressure, the elastin fibres become straighter and more taut asthey are loaded within the physiological pressure range. Concomitantly, theinterlamellar space and lamellar thickness decrease.
The elastin content of the toad aorta was approximately 15 % of the total drymass, while the collagen content was about 35 % of the dry mass. These valueswere consistent at all four sites (Table 1), indicating that the aorta has a uniformimposition along its length. The collagen to elastin ratio of 2.3 in the toad aorta is
284 C. A. GIBBONS AND R. E. SHAD WICK
B
/ ir»
Fig. 7. Micrographs of a toad aorta fixed at OkPa and stained for elastin, showing atransverse section (A) and a longitudinal section (B). For comparison, a transversesection of a rat aortic wall is shown in C. (D) A transverse section of a toad aorta fixedat lOkPa and stained to show cell nuclei. / indicates the luminal side of the wall. Scalebar, 50^m.
more than five times that found at the level of the aortic arch in mammals (seeFig. 10).
Discussion
This study describes several important mechanical design features of the aortaof a typical poikilothermic vertebrate. The high distensibility, resilience and non-linear elasticity are the requisite properties for the vessel to act as an effectivestorage element in the arterial circulation. In this respect, a strong functionalsimilarity to the aorta of mammals is apparent, although specific structural andmechanical differences are seen that reflect the much lower level of pressure andsimpler haemodynamics for which the toad vessel is suited.
Mechanics of the toad aorta 285
B
Fig. 8. Micrographs of transverse sections of the aorta from four different toads, fixedat distending pressures of 1.0kPa (A), 1.7kPa (B), 2.5 kPa (C) and 4.0kPa (D), andstained to show the elastin lamellae. Scale bar, 50/an. /, lumen.
Our initial observation that a longitudinal recoil of about 30 % occurred whenthe aorta was excised is surprisingly similar to the behaviour of the aorta ofmammals. With inflation to the normal physiological pressure of about 3kPa, anunrestrained toad aorta regains its in vivo length (Fig. 2), i.e. the longitudinalstress due to pressure alone causes the lengthening, and tethering to the axialskeleton is unimportant. In fact, there are no branch vessels between thesubclavian and coeliacomesenteric arteries, and very few along the rest of theaorta. During a normal pressure pulse, the aorta will lengthen only slightly, owingto the relatively limited longitudinal extensibility at pressures above 2kPa.Similarly, in the mammalian thoracic aorta, longitudinal strains of only 1 % or lessoccur with each heart beat (Patel and Fry, 1964), although this behaviour hasgenerally been attributed to a high degree of tethering by the intercostal arteriesand connective tissues (Milnor, 1982). However, Van Loon et al. (1977) demon-
^rated that there is a maximal length to which mammalian arteries elongate when

286 C. A . GIBBONS AND R. E . SHADWICK
pressurized without tethering, and that this is very close to the in vivo length.Thus, to a large extent, the relatively constant aortic length in vivo results from themechanical anisotropy of the wall material, i.e. the stiffness is greater longitudi-nally than it is circumferentially (Fenn, 1957; Patel et al. 1969; Vaishnav et al. 1972;Dobrin, 1983). This type of anisotropy in the toad aorta is evident from theuniaxial tests shown in Fig. 6. Interestingly, similar mechanical behaviour hasbeen observed for the aorta of octopods and squid (Gosline and Shadwick, 1982;Shadwick and Gosline, 1985; Shadwick and Nilsson, 1990), soft-bodied animals inwhich no skeletal tethering of the aorta is possible.
The non-linear and viscoelastic properties of the toad aorta reflect its compositestructure; it consists mainly of highly extensible elastin and relatively inextensiblecollagen. With inflation, the elastic lamellae are loaded and become straightenedas the pressure approaches the physiological range (Fig. 8). Above 3kPa there isan abrupt decrease in the vessel distensibility and an increase in the circumfer-ential elastic modulus (Figs 3, 5), above what could be attributed to elastin alone(Aaron and Gosline, 1981). In the mammahan aorta, an abrupt transition fromhigh compliance to high stiffness also occurs, but at much higher mean bloodpressures of 11-12 kPa (Bergel, 1961; Gibbons and Shadwick, 1989). In both toadand mammal, inflation of the thoracic aortas to their respective physiological meanpressures results in circumferential strains of 0.6-0.8, an elastic modulus of about0.4 MPa and mechanical hysteresis of 15-20% (Bergel, 1961; Cox, 1978; Gibbonsand Shadwick, 1989). Thus, the aorta of the toad appears to be designed to haveessentially the same functional properties as that of mammals, but at much lowerpressures.
A significant finding in this study is that, in contrast to mammals, there is nogeometric or elastic 'tapering' in the arterial tree of the toad. The dimensions andelastic properties are relatively uniform along the length of the aorta, whereas themammahan aorta exhibits a progressive decrease in radius and increase in elasticmodulus peripherally (Figs 9, 10). The distribution of mechanical properties inthese aortas is correlated with their connective tissue composition, i.e. the toadaorta has a relatively constant quantity of elastin and collagen along its length,while in the mammahan aorta an increase in the ratio of collagen to elastin occursdistal to the heart (McDonald, 1974; Fig. 10). In the mammahan system, thiselastic and geometric tapering together cause a continuous increase in pressure-wave velocity and aortic impedance distal to the heart, giving the aorta theproperties of a non-uniform transmission line (McDonald, 1974; Milnor, 1982).The lack of any significant elastic or geometric taper in the toad aorta, by contrast,suggests that this vessel is more suited to function as a simple Windkessel than as acomplex transmission line. These haemodynamic features are considered in moredetail in the following paper.
The question of how the non-linear elastic properties occur at much lowerpressures in the toad aorta than in that of the mammal can be addressed byconsidering structural and connective tissue compositional differences. The aorticR/h ratio is nearly 2.5 times higher in the toad, resulting in higher wall stresj|
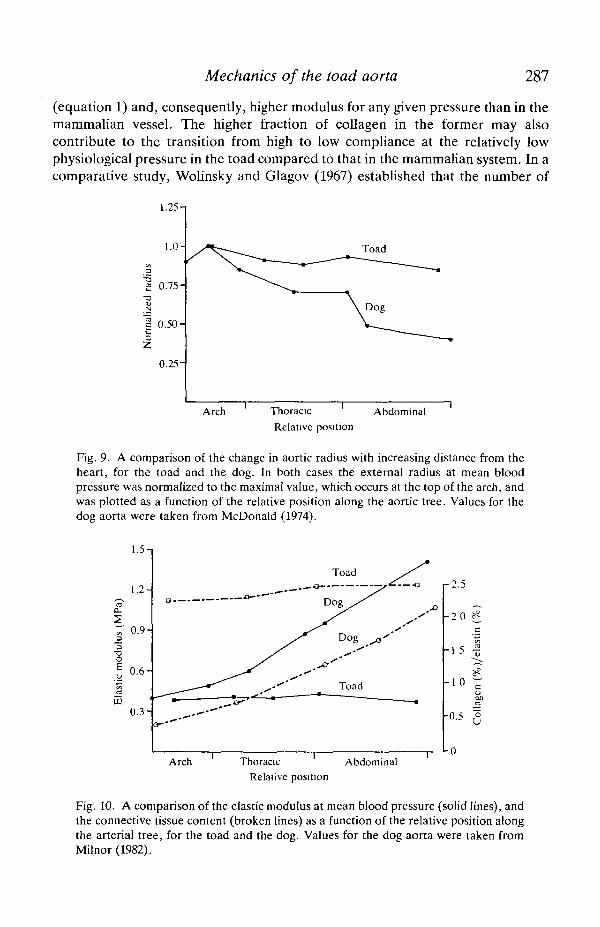
Mechanics of the toad aorta 287
(equation 1) and, consequently, higher modulus for any given pressure than in themammalian vessel. The higher fraction of collagen in the former may alsocontribute to the transition from high to low compliance at the relatively lowphysiological pressure in the toad compared to that in the mammalian system. In acomparative study, Wolinsky and Glagov (1967) established that the number of
1.25-1
1.0-
2 0.75-
1 0.50-
0.25-
Toad
Arch ThoracicRelative position
Abdominal
Fig. 9. A comparison of the change in aortic radius with increasing distance from theheart, for the toad and the dog. In both cases the external radius at mean bloodpressure was normalized to the maximal value, which occurs at the top of the arch, andwas plotted as a function of the relative position along the aortic tree. Values for thedog aorta were taken from McDonald (1974).
1.5-i
1.2-
a.S 0.9-
0.6-
LL)0 .3 -
2.5
20 ?C
15 |
cQJoon3
-0.5 ^
-0Arch Thoracic
Relative position
Abdominal
Fig. 10. A comparison of the elastic modulus at mean blood pressure (solid lines), andthe connective tissue content (broken lines) as a function of the relative position alongthe arterial tree, for the toad and the dog. Values for the dog aorta were taken fromMilnor (1982).

288 C. A. GIBBONS AND R. E . SHADWICK
elastin layers, N, in the mammalian aorta was a function of body size, orspecifically of aortic radius at mean blood pressure. Consequently, the circumfer-ential wall tension per layer (=Pr/N) remains nearly constant at about 2 Pa m in allspecies. In contrast, the quantity and mechanical behaviour of the elastin layers inthe toad aorta do not fit this general pattern, instead becoming straightened at arelatively low pressure (Fig. 8) that generates a tension per layer of only about0.5Pam.
In conclusion, specific differences in structure, connective tissue architectureand mechanical properties are demonstrated in the toad aorta compared to that ofmammals. Overall, however, the mechanical behaviour of the aorta is functionallycomparable in the two taxa, although their range of physiological pressures is verydifferent. Thus, the aorta has the same capability to act as an effective elasticenergy storage element in the circulation of this poikilothermic vertebrate as inthat of mammals. The contribution of these elastic properties to arterialhaemodynamics is explored in the following study.
This work was supported by a Graduate Research Grant from the University ofCalgary to C.A.G. and by an operating grant and Research Fellowship from theNatural Science and Engineering Research Council of Canada to R.E.S.
ReferencesAARON, B. B. AND GOSLINE, J. M. (1981). Elastin as a random network elastomer. Biopolymers
20, 1247-1260.BERG, A. R. (1982). Determination of 3- and 4-hydroxyproline. Meth. Enzymol. 82, 372-398.BERGEL, D. H. (1961). The static elastic properties of the arterial wall. /. Physiol., Lond. 156,
458-469.Cox, R. H. (1978). Comparison of carotid artery mechanics in the rat, rabbit, and dog. Am. J.
Physiol. 234, H280-H288.DOBRIN, J. B. (1983). Vascular mechanics. In Handbook of Physiology, section 2, The
Cardiovascular System, vol. II (eds. D. F. Bohr, A. P. Somylo and H. V. Sparks, Jr), pp.65-102. Bethseda, MD: American Physiological Society.
FENN, W. O. (1957). Changes in length of blood vessel on inflation. In Tissue Elasticity (ed.J. Remington), pp. 154-167. Washington: American Physiological Society.
FUNG, Y. C. (1981). Biomechanics. Mechanical Properties of Living Tissues. New York:Springer-Verlag.
GIBBONS, C. A. AND SHADWICK, R. E. (1989). Mechanical design in the arteries of lowervertebrates. Experientia 45, 1083-1089.
GIBBONS, C. A. AND SHADWICK, R. E. (1991). Circulatory mechanics in the toad Bufo marinus.II. Haemodynamics of the arterial Windkessel. /. exp. Biol. 158, 291-306.
GOSLINE, J. M. AND SHADWICK, R. E. (1982). The biomechanics of the arteries of Nautilus,Notodarus, and Sepia. Pacific Sci. 36, 283-2%.
GOTO, M. AND KIMOTO, Y. (1966). Hysteresis and stress-relaxation of the blood vessels studiedby a universal tensile testing instrument. Jap. J. Physiol. 76, 169-184.
HUMASON, G. L. (1979). Animal Tissue Techniques. San Francisco: W. H. Freeman andCompany.
JONES, D. R., LANGILLE, B. L, RANDALL, D. J. AND SHELTON, G. (1974). Blood flow in dorsaland ventral aortas of the cod, Gadus morhua. Am. J. Physiol., Lond. 226, 90-95.
LANGILLE, B. L. AND JONES, D. R. (1977). Dynamics of blood flow through the hearts andarterial systems of anuran amphibia. J. exp. Biol. 68, 1-17.
LANSING, A. I., ROSENTHAL, T. B., ALEX, M. AND DEMPSEY, E. W. (1952). The structure and|

Mechanics of the toad aorta 289
chemical characterization of elastic fibres as revealed by elastase and electron microscopy.Anat. Rec. 114, 555-575.
MCDONALD, D. A. (1974). Blood Flow in Arteries. London: Edward Arnold.MILNOR, W. R. (1982). Hemodynamics. Baltimore: Williams and Wilkens.PATEL, D. J. AND FRY, D. L. (1964). In situ pressure-radius-length measurements in the
ascending aorta of anaesthetized dogs. /. appl. Physiol. 19, 413-416.PATEL, D. J., JANICKI, J. S. AND CAREW, T. E. (1969). Static anisotropic elastic properties of the
aorta in living dogs. Circulation Res. 25, 765-769.SAGE, E. H. AND GRAY, W. R. (1979). Studies on the evolution of elastin. I. Phylogenetic
distribution. Comp. Biochem. Physiol. 66B, 313-327.SATCHELL, G. H. (1971). Circulation in Fishes. Cambridge: Cambridge University Press.SHADWICK, R. E. AND GOSLLNE, J. M. (1985). Mechanical properties of the octopus aorta. /. exp.
Biol. 114, 259-284.SHADWICK, R. E. AND NILSSON, E. K. (1990). The importance of vascular elasticity in the
circulatory system of the cephalopod Octopus vulgaris. J. exp. Biol. 152, 471-484.SHELTON, G. AND BURGGREN, W. W. (1976). Cardiovascular dynamics of the chelonia during
apnoea and lung ventilation. J. exp. Biol. 64, 323-343.SHELTON, G. AND JONES, D. R. (1968). A comparative study of central blood pressures in five
amphibians. /. exp. Biol. 49, 631-643.TAYLOR, M. G. (1964). Wave travel in arteries and the design of the cardiovascular system. In
Pulsatile Blood Flow (ed. E. O. Attinger), pp. 343-372. New York: McGraw-Hill.VAISHNAV, R. N., YOUNG, J. T., JANICKI, J. S. AND PATEL, D. J. (1972). Non-linear anisotropic
elastic properties of the canine aorta. Biophys. J. Yl, 1008-1027.VAN LOON, P., KLIP, W. AND BRADLEY, E. L. (1977). Length-force and volume-pressure
relationships of arteries. Biorheology 14, 181-201.WEST, N. H. AND BURGGREN, W. W. (1984). Factors influencing pulmonary and cutaneous
arterial blood flow in the toad, Bufo marinus. Am. J. Physiol., Lond. 247, R884-R894.WOLINSKY, H. AND GLAGOV, S. (1964). Structural basis for the static mechanical properties of
the aortic media. Circulation Res. 14, 400-413.WOLINSKY, H. AND GLAGOV, S. (1967). A lamellar unit of aortic medial structure and function in
mammals. Circulation Res. 20, 99-111.ZAR, J. H. (1984). Biostatistical Analysis. New Jersey: Prentice-Hall.






























