Embed Size (px)
Citation preview

CLEARING FACTOR, A HEPARIN-ACTIVATED LIPOPROTEIN LIPASE*
I. ISOLATION AND CHARACTERIZATION OF THE ENZYME FROM NORMAL RAT HEART
BY EDWARD D. KORN
(From the Laboratory of Cellular Physiology, National Heart Institute, National Institutes of Health, United States Public Health Service, Bethesda, Maryland)
(Received for publication, October 15, 1954)
The importance of plasma lipoproteins in normal and abnormal fat metabolism has stimulated interest in the heparin-induced lipemia clearing system first observed by Hahn (2). As a result of the efforts of several independent investigators (3,4), it is now known that,, upon the administra- t’ion of heparin to animals of many different species, an enzyme appears in the blood which is able to “clear” lipemic serum in viva or in vitro.
It has been difficult to evaluate the role of this “clearing factor” in nor- mal fat metabolism since it normally exists in blood in extremely low con- centrations, if at all (5). Certainly the clearing factor content of normal blood is insufficient to account for the rate of disappearance of alimentary lipemia. This is most readily‘evidenced by the fact that lipemic serum does not clear in vitro. Recent investigations of Bragdon and Have1 (6) demonstrated t,hat the administration of prot.amine and other antihep- arinoid agents to fasting rats increases the serum lipide and lipoprotein concentration and, when injected simultaneously with lipoproteins, delays their removal. These results suggest that a heparinoid-requiring system is involved in normal fat metabolism.
The work to be reported in this paper demonstrates the presence in normal rat heart of an enzyme identical with the clearing factor which appears in blood upon heparin administration. In this and the accom- panying paper (7), evidence is presented which supports the view that
, clearing factor is a heparin-activated lipoprotein lipase which may play a major role in lipide metabolism.
Materials and Methods
Sodium heparin, 126 units per mg., was generously supplied by The Up- john Company, protamine sulfate was a product of the Nutritional Bio- chemicals Corporation, and the commercial pancreatic lipase was obtained from the Fisher Scientific Company. The coconut oil emulsion was sup-
* A preliminary report of some of the data presented in this and the accompanying paper has been published (1).
by guest on September 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

2 LIPOPROTEIN LIPASE. I
plied by the Abbott Laboratories. It had the following percentage com- position (weight per volume): coconut oil 15, glucose 5, Tween 60 0.5, phenylmercuric nitrate 0.002, and tocopherol 0.01. It was diluted to a coconut oil concentration of 1.5 per cent before use. The albumin used was bovine serum albumin obtained from the Armour Laboratories; the chylomicrons were obtained from pooled blood from the American Red Cross. The material of low density, which separated on standing in the refrigerator, was washed several times with 0.15 M NaCl by centrifugation and used after dilution with water to approximately 1.5 per cent neutral fat content. Serum was whole normal human serum.
Acetone powders were prepared by homogenizing the tissues in approxi- mately 20 volumes of acetone (- 20’) in a Waring blendor and filtering the mixture rapidly at room temperature. The filter cake was blended again with acetone and the final powder stored in a vacuum desiccator at -20”. Aortas were homogenized briefly in a Potter-Elvehjem glass homogenizer before the acetone treatment.
In all the experiments, unless otherwise stated, the incubation vessel contained 0.2 ml. of 10 per cent albumin (pH KS), 0.02 ml. of 1 M CaC12, 0.1 ml. of the chylomicron suspension, and 0.28 ml. of 0.25 M NH&l-NH3 buffer (pH 8.5) in a total volume of 1 ml. Other additions were as indi- cated. Duplicate 0.05 ml. aliquots of the incubation mixture were removed at 0, 30, 60, and 120 minutes, in all cases, and added directly to 0.1 ml. of cold 1 N HzS04 in a 12 ml. graduated centrifuge tube. Glycerol was then determined by a modified procedure of Lambert and Neish (8). This method is highly reproducible and enables one routinely to determine from 0.005 to 0.1 pmole of glycerol. The contents of the centrifuge: tube were warmed to room temperature and 0.1 ml. of 0.05 M sodium periodate was added. After 5 minutes, 0.1 ml. of 0.5 M sodium arsenite was added to reduce the excess periodate. After an additional 10 minutes, 9 ml. of chromotropic acid reagent (a formaldehyde reagent) were added, and the reaction tubes placed in a covered boiling water bath for 30 minutes. The cooled tubes were adjusted to a volume of 10 ml. with water and the optical density read at 570 rnp in a Coleman junior spectrophotometer. By this procedure 2 moles of formaldehyde are formed from 1 mole of glycerol, and 1 mole of formaldehyde from 1 mole of a-monoglyceride. All the values were calculated in terms of glycerol, although an undetermined percentage of the formaldehyde was actually derived from a-monoglycerides. All the values reported are the averages of two determinations, and in all instances the 30, 60, and 120 minute values were proportional to the incubation times. Where only one value is reported, it is the one obtained after an incubation period of 120 minutes.
by guest on September 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from
E. D. KORN 3
Results
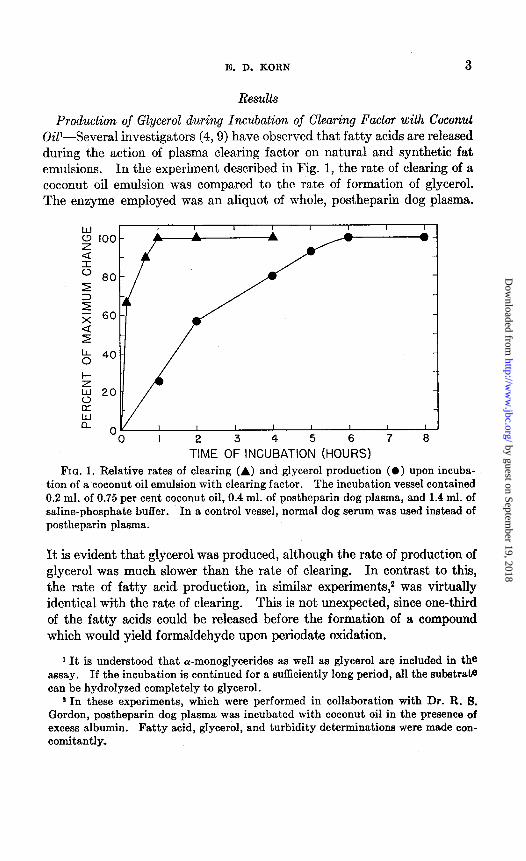
Production of Glycerol during Incubation of Clearing Factor with Coconut OiP-Several investigators (4,9) have observed that fatty acids are released during the action of plasma clearing factor on natural and synthetic fat emulsions. In the experiment described in Fig. 1, the rate of clearing of a coconut oil emulsion was compared to the rate of formation of glycerol. The enzyme employed was an aliquot of whole, postheparin dog plasma.
I 0 I
T?ME OF3 INC”4BATIO: (HO”F?S) 7 a
TIME OF INCUBATION (HOURS) FIG. 1. Relative rates of clearing (A) and glycerol production (0) upon incuba-
tion of a coconut oil emulsion with clearing factor. The incubation vessel contained 0.2 ml. of 0.75 per cent coconut oil, 0.4 ml. of postheparin dog plasma, and 1.4 ml. of saline-phosphate buffer. In a control vessel, normal dog serum was used instead of postheparin plasma.
FIG. 1. Relative rates of clearing (A) and glycerol production (0) upon incuba- tion of a coconut oil emulsion with clearing factor. The incubation vessel contained 0.2 ml. of 0.75 per cent coconut oil, 0.4 ml. of postheparin dog plasma, and 1.4 ml. of saline-phosphate buffer. In a control vessel, normal dog serum was used instead of postheparin plasma.
It is evident that glycerol was produced, although the rate of production of glycerol was much slower than the rate of clearing. In contrast to thii, the rate of fatty acid production, in similar experiments,2 was virtually identical with the rate of clearing. This is not unexpected, since one-third of the fatty acids could be released before the formation of a compound which would yield formaldehyde upon periodate oxidation.
1 It is understood that cu-monoglycerides as well as glycerol are included in the assay. If the incubation is continued for a sufficiently long period, all the substrate can be hydrolyzed completely to glycerol.
2 In these experiments, which were performed in collaboration with Dr. R. 8. Gordon, postheparin dog plasma was incubated with coconut oil in the presence of excess albumin. Fatty acid, glycerol, and turbidity determinations were made con- comitantly.
by guest on September 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

4 LIPOPROTEIN LIPASE. I
The product.ion of compounds (glycerol, a-monoglycerides) which yield formaldehyde upon oxidation with periodate provides an extremely con- venient means for assaying the clearing factor. It is much more accurate than, and not subject to the numerous difficulties of, the turbidity method employed in previous studies of this enzyme.
Extraction of Clearing Factor from Rat Heart Acetone Powder-Anfinsen et al. (5) reported that clearing factor is obtained upon the incubation of heart and lung preparations with heparin and normal plasma. This was interpreted as indicating the existence of a “tissue factor” which converted a “plasma precursor substance” into clearing factor in the presence of heparin. These observations have been reinvestigated.
TABLE I Extraction of Lipoprotein Lipase from Rat Heart Acetone Powder
1 Serum 2 Saline-phosphate 3 NH: 4 Serum (after NH*) 5 “ saline-phosphate, or NH**
Extracting solvent
Glycerol production
30 min. 60 min. 120 min.
~mole pwlc pmole
0.21 0.43 0 0 0 0.19 0.39 0.61 0.03 0.05 0.11 0 0 0
All the vessels contained 0.2 ml. of the solution to be assayed in addition to the other components, as described under “Methods.” All of the incubation vessels contained an equal amount of serum. For other experimental details, see the text.
* In this experiment the three solvents were each assayed separately. No ace- tone powder was present at any time.
50 mg. of rat heart acetone powder were extracted for 30 minutes at 0’ with 1 ml. of normal human serum, or saline-phosphate buffer (1 part of 0.15 M NaCl: 1 part of phosphate buffer (pH 7.4, I’/2 = 0.1))) or 0.025 N ammonia. The insoluble residues were removed by centrifugation (25,000 X g) and the supernatant fluids incubated at 38” for 30 minutes with addition of an equal volume of serum to those samples which did not, already con&n it. In one instance, Experiment 4, the acetone powder was extracted with 0.025 N ammonia, Lhe supernatant fluid discarded, and the residue washed with saline-phosphate and extracted again with serum, which was then assayed for activity.
As shown in Table I, both the serum and ammonia extracts contained an active component, while the saline-phosphate extract was inactive. The serum extract of the acetone powder which had been previously extracted
by guest on September 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

E. D. KORN 5
with ammonia had relatively little lipase activity, and none of the three extracting solvents alone (Experiment 5) had any activity.
The results tabulated in Table II demonstrate that the ammonia extract had lipolytic activity in the complet,e absence of serum and that preincuba- tion of the ammonia extract with serum, even in the presence of heparin, had no stimulatory effect. Thus the rat heart acetone powder contains a lipase which will be shown to be ident.ical to clearing factor, and the serum serves only as a solvent which can be completely replaced by ammonia.
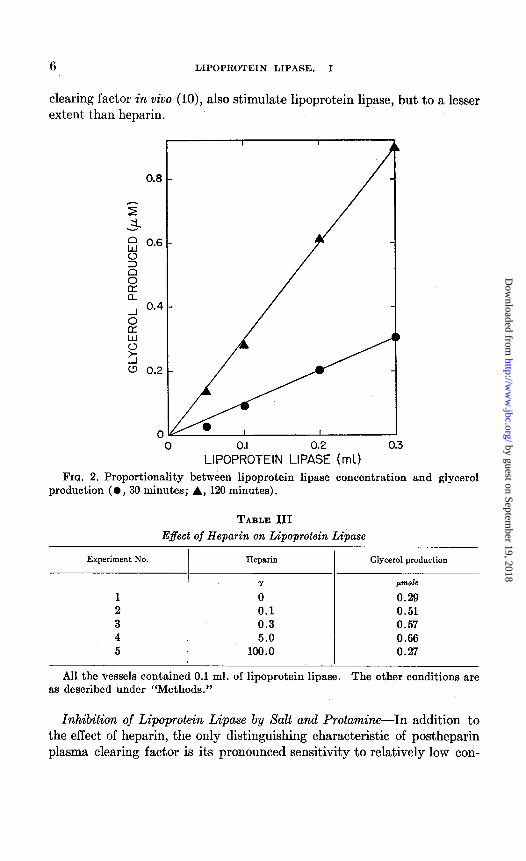
The rate of glycerol production was directly proportional to the amount of enzyme added (Fig. 2) and to the time of incubation (see Table II, for example). The enzyme is non-dialyzable and stable to dialysis for 24 hours against distilled water. It is completely inactivated when heated
TABLE II Non-Essentiality of Serum for Preparation of Lipoprotein Lipase
Glycerol production
Experiment No. Serum added to assay vessel
30 min. 60 min. 120 min.
ml. wok pnwlc *mole
1 0 0.18 0.33 0.65 2 0.2 0.13 0.26 0.49 3 0.2* 0.12 0.24 0.49
All the vessels contained 0.2 ml. of the ammonia extract of rat heart acetone powder and 10 y of heparin in addition to the usual components.
* In this experiment the ammonia extract was preincubated with the serum and heparin for 30 minutes at 38” before its lipolytic activity was assayed.
in a boiling water bath for 5 minutes. The ammonia extract of the rat heart acetone powder will be referred to as lipoprotein lipase, since this denotes both its specificity and the type of reaction which it catalyzes, as will be more fully demonstrated in the accompanying paper (7). Unless otherwise indicated, it was always prepared by extracting 50 mg. of rat heart acetone powder with 1 ml. of 0.025 N ammonia for 30 minutes at 0”.
E$ect of Heparin on Lipoprotein Lipase-There has been no evidence that a heparinoid is necessary for the enzymabic action of postheparin plasma clearing factor. Heparin added in vitro to such a clearing factor preparation inhibits its activity (4). The addition of small amount,s of heparin to the ammonia extract of rat, heart acetone powder st.imulated the lipase activity approximately 2-fold (Table III). Relatively large amounts of heparin had an inhibitory effect’. In some instances 3- and I-fold stimulations have been obtained. Mactin-A (a sulfated polysaccharide from clams) and polymetaphosphate, both of which can produce plasma
by guest on September 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from
6 LIPOPROTEIN LIPASE. I
clearing factor in vivo (lo), also stimulate lipoprotein lipase, but to a lesser ext,ent, than heparin.
0.8
0 0.3
FIG. 2. Proportionality between lipoprotein lipase concentration and glycerol production ( l , 30 minutes; A, 120 minutes).
TABLE III Effect of Heparin on Lipoprotein Lipase
Experiment No. Heparin I
Glycerol production
Y pmole
0 0.29 0.1 0.51 0.3 0.57 5.0 0.66
100.0 0.27
All the vessels contained 0.1 ml. of lipoprotein lipase. The other conditions are as described under “Methods.”
Inhibition of Lipoprotein Lipase by Salt and Protamine-In addition to the effect of heparin, the only distinguishing characteristic of postheparin plasma clearing factor is its pronounced sensitivity to relatively low con-
by guest on September 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

E. D. KORN 7
centrations of salt and protamine (4). The inhibitory effects of these compounds on the rat heart preparation are shown in Table IV.
The inhibition by protamine is presumably due to the presence of en- zyme-bound heparin, which is apparently an essential component of the system. Recent experiments, in collaboration with Mr. Thomas W. Quigley, Jr., on lipoprotein lipase extracted from calf heart, have demon- strated that heparin can reverse this inhibition. The inhibition by salt is not reversible by heparin at any concentration nor by the addition of whole serum or a boiled extract of lipoprotein lipase. On the assumption that the active enzyme contains bound heparin, an attempt was made to in-
- Experiment No.
TABLE IV Effect of Salt and Protamine on Lipoprotein Lipase
T
Pretreatment of enzyme
None ‘6
NaCI, 1 M “ 1 “
KCl, 1 Y “ 1 ‘(
Protamine, 5 X 1OF M “ 1 x 10-S “ ‘I 1 x 10-C “
T
Lipase Heparin Glycerol production
ml. -I pfflole
0.1 0.40
0.1 10 0.65 0.2 0 0.2 10 0 0.2 0 0.2 10 0 0.1 0.10 0.1 0.27 0.1 0.33
The other additions are as under “Methods.” Aliquots of lipoprotein lipase were incubated with NaCl, KCl, or protamine in the concentrations indicated for 30 minutes at 0’. All the samples were dialyzed overnight against distilled water and then assayed.
activate it by treatment with Dowex 1 Cl- and activated carbon. Both attempts were unsuccessful.
E$ects of Heparin, Salt, and Protamine on Pancreatic Lipase-The effects of heparin, salt, and protamine on lipoprotein lipase are quite unique. They serve to distinguish it from other lipases which will also hydrolyze (although not as well) chylomicrons. Pancreatic lipase, for example, was neither activated by heparin nor inactivated by salt or protamine under conditions identical with those used to demonstrate the effects of these compounds on lipoprotein lipase.
Inhibition of Lipoprotein Lipase by Pyrophosphate-The data tabulated in Table V demonstrate that sodium pyrophosphate completely inhibited lipoprotein lipase when present in the assay vessel at a concentration of 10 pmoles per ml. At levels of 5 pmoles per ml. the pyrophosphate inhibi-
by guest on September 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

8 LIPOPROTEIN LIPASE. I
tion was net always complete but was still considerable, and could then be reversed by 5 y of heparin. This inhibition could also be overcome by polymetaphosphate.
TABLE V
Inhibition of Lipoprotein Lipase by Pyrophosphate
Experiment No. Sodium pyrophosphate Heparin
pm&r
1 2 3 4 5 6 7 8 9
10
1 1 5 5 5 5
10 10
-
-- Glycerol production
pmde
0.46 0.68 0.49 0.63 0.14 0.57 0 0.51 0 0
All the vessels contained 0.1 ml. of lipoprotein lipsse.
TABLE VI
Inhibition of Lipoprotein Lipase by Phosphate-Containing Compounds
Experiment No. ’ Inhibitor* Heparin :lycerol production -
WWlCS /&+7tOk
0.34 0.55
P 50 0.14 “ 50 0.42 ATP 20 0
“ 10 0.10 “ 10 0.40
UTP 10 0.07 “ 10 0.44
ITP 20 0.05 AMP 20 0.05
‘I 10 0.24 “ 10 0.44
- All the vessels contained 0.1 ml. of lipoprotein lipase. * P, orthophosphate; ATP, adenosine triphosphate; UTP, uridine triphosphate;
ITP, inosine triphosphate; AMP, adenylic acid.
1 2 3 4 5 6 7 8 9
10 11 12 13
Other phosphate derivatives were also assayed for their ability to inhibit lipoprotein lipase (Table VI), Inorganic phosphate, adenosine triphos- phate, uridine triphosphate, inosine triphosphate, and adenylic acid all
-
by guest on September 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

E. D. KORN 9
inhibited, although at somewhat higher concentrations than was necessary with the pyrophosphate. Heparin could reactivate the enzyme in all cases.
This inhibition is due to a direct effect of the phosphate compound on lipoprotein lipase itself. Pyrophosphate had no inhibitory effect when present in the assay vessel at a concentration of 0.5 pmole per ml. How- ever, preincubation of the enzyme with pyrophosphate at a concentration of 5 pmoles per ml. caused definite inhibition even though the inhibitor was diluted to the non-effective concentration in the assay vessel (Table VII).
TABLE VII Effect of Preincubation of Lipoprotein Lipase with Pyrophosphate
Experiment No. Sodium pyrophosphate Heparin Glycerol production
0.5 0.5 5.0 5.0 0.5* 0.5*
pmole
0.49 0.57 0.50 0.58 0.19 0.50 0.27 0.43
All the vessels contained 0.1 ml. of lipoprotein lipase. * In these two experiments, 0.5 pmole of pyrophosphate was preincubated with
0.1 ml. of lipoprotein lipase for 30 minutes at room temperature. In the absence of pyrophosphate, there was no decrease in enzymatic activity during a preincubation period at room temperature.
These results are most readily explained by the assumption that lipo- protein lipase, as isolated from normal rat hearts, contains enzyme-bound heparin (or heparinoid) which may be competitively replaced by polyvalent anions which possess no cofactor activity.
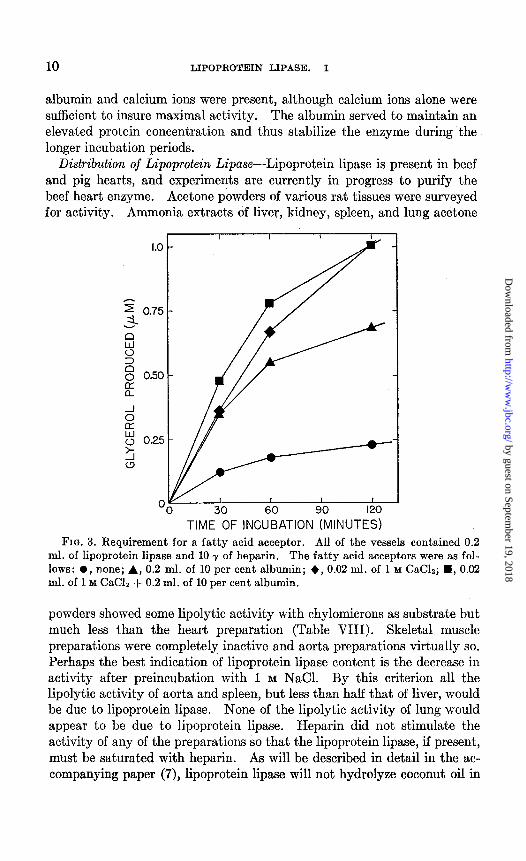
Requirement for Fatty Acid Acceptor-Recent experiments by Gordon et al. (11) have demonstrated that serum albumin stimulates the clearing of coconut oil emulsions by postheparin plasma clearing factor by virtue of its ability to remove from solution the fatty acids produced during the reaction. Other investigators (12) have postulated a special, and more obscure, r6le for albumin. It can be shown, however, that there is no specific requirement for albumin for the hydrolysis of chylomicrons by lipoprotein lipase (Fig. 3). The chylomicrons used as substrate in this experiment were washed completely free of albumin. Calcium ions were at least as good a fatty acid acceptor as albumin.
In the experiments reported in this and the accompanying paper, both
by guest on September 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from
10 LIPOPROTEIN LIPASE. I
albumin and calcium ions were present, although calcium ions alone were sufficient to insure maximal activity. The albumin served to maintain an elevated protein concentration and thus stabilize the enzyme during the longer incubation periods.
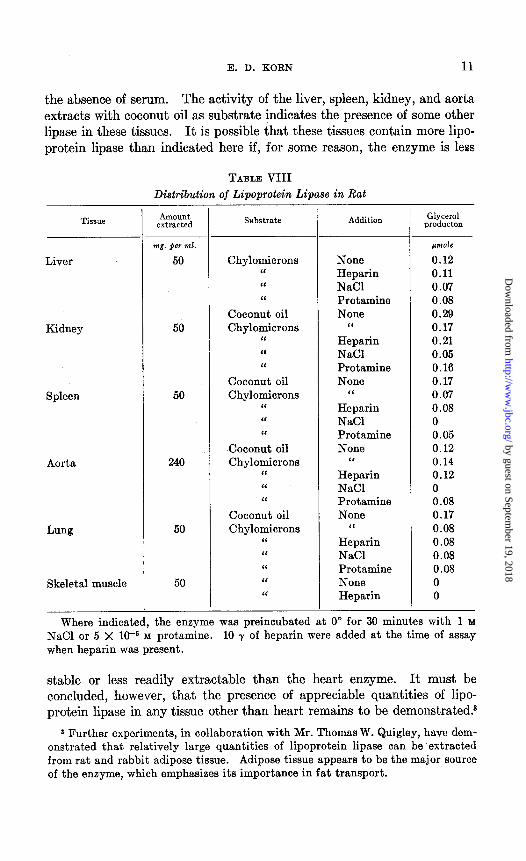
Distribution of Lipoprotein fipase-Lipoprotein lipase is present in beef and pig hearts, and experiments are currently in progress to purify the beef heart enzyme. Acetone powders of various rat tissues were surveyed for activity. Ammonia extracts of liver, kidney, spleen, and lung acetone
TIME OF INCUBATION (MINUTES) FIG. 3. Requirement for a fatty acid acceptor. FIG. 3. Requirement for a fatty acid acceptor. All of the vessels contained 0.2 All of the vessels contained 0.2
ml. of lipoprotein lipase and 10 y of heparin. ml. of lipoprotein lipase and 10 y of heparin. The fatty acid acceptors were as fol- The fatty acid acceptors were as fol- lows: 0, none; A, 0.2 ml. of 10 per cent albumin; +, 0.02 ml. of 1 M CaC12; H, 0.02 lows: 0, none; A, 0.2 ml. of 10 per cent albumin; +, 0.02 ml. of 1 M CaC12; H, 0.02 ml. of 1 M CaClz + 0.2 ml. of 10 per cent albumin. ml. of 1 M CaClz + 0.2 ml. of 10 per cent albumin.
powders showed some lipolytic activity with chylomicrons as substrate but much less than the heart preparation (Table VIII). Skeletal muscle preparations were completely inactive and aorta preparations virtually so. Perhaps the best indication of lipoprotein lipase content is the decrease in activity after preincubation with 1 M NaCl. By this criterion all the lipolytic activity of aorta and spleen, but less than half that of liver, would be due to lipoprotein lipase. None of the lipolytic activity of lung would appear to be due to lipoprotein lipase. Heparin did not stimulate the activity of any of the preparations so that the lipoprotein lipase, if present, must be saturated with heparin. As will be described in detail in the ac- companying paper (7), lipoprotein lipase will not hydrolyze coconut oil in
by guest on September 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from
E. D. KORN 11
the absence of serum. The activity of the liver, spleen, kidney, and aorta extracts with coconut oil as substrate indicates the presence of some other lipase in these tissues. It is possible that these tissues contain more lipo- protein lipase than indicated here if, for some reason, the enzyme is less
TABLE VIII Distribution of Lipoprotein Lipase in Rat -
I Liver
Kidney 50
Spleen
Aorta 240
Lung
Skeletal muscle
Amount extracted
mg. fier ml.
50
50
50
50
Substrate Addition
Chylomicrons “ “ ‘L
Coconut oil Chylomicrons
“ ‘C “
Coconut oil Chylomicrons
“ ‘< I‘
Coconut oil Chylomicrons
“ “ “
Coconut oil Chylomicrons
“ “ “ “ ‘<
None Heparin NaCl Protamine None
I‘ Heparin NaCl Protamine None
“
Heparin NaCl Protamine None
I‘ Heparin NaCl Protamine None
‘I Heparin NaCl Protamine None Heparin
Glycerol producton
pm&
0.12 0.11 0.07 0.08 0.29 0.17 0.21 0.05 0.16 0.17 0.07 0.08 0 0.05 0.12 0.14 0.12 0 0.08 0.17 0.08 0.08 0.08 0.08 0 0
Where indicated, the enzyme was preincubated at 0” for 30 minutes with 1 M NaCl or 5 X 1V M protamine. 10 y of heparin were added at the time of assay when heparin was present.
stable or less readily extractable than the heart enzyme. It must be concluded, however, that the presence of appreciable quantities of lipo- protein lipase in any tissue other than heart remains to be demonstrated.3
8 Further experiments, in collaboration with Mr. Thomas W. Quigley, have dem- onstrated that relatively large quantities of lipoprotein lipase can be extracted from rat and rabbit adipose tissue. Adipose tissue appears to be the major source of the enzyme, which emphasizes its importance in fat transport.
by guest on September 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from
12 LIPOPROTEIN LIPASE. I
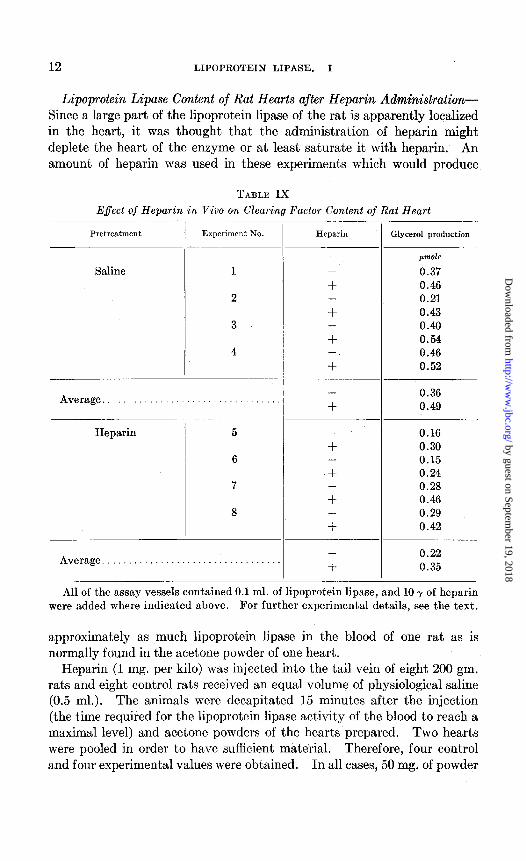
Lipoprotein Lipase Content of Rat Hearts after Heparin Administration- Since a large part of the lipoprotein lipase of the rat is apparently localized in the heart, it was thought that the administration of heparin might deplete the heart of the enzyme or at least saturate it with heparin. An amount of heparin was used in these experiments which would produce
TABLE IX
Effect of Heparin in Viva on Clearing Factor Content of Rat Heart
Pretreatment
Saline
Experiment No. Experiment No.
Average..................................
Heparin
6
7
Heparin Glycerol production
-
+ -
+ -
+ -
+
-
+
-
+ -
+ -
+ -
+
-
+
jmole
0.37 0.46 0.21 0.43 0.40 0.54 0.46 0.52
0.36 0.49
0.16 0.30 0.15 0.24 0.28 0.46 0.29 0.42
0.22 0.35
All of the assay vessels contained 0.1 ml. of lipoprotein lipase, and 10 y of heparin were added where indicated above. For further experimental details, see the text.
approximately as much lipoprotein lipase in the blood of one rat as is normally found in the acetone powder of one heart.
Heparin (1 mg. per kilo) was injected into the tail vein of eight 200 gm. rats and eight control rats received an equal volume of physiological saline (0.5 ml.). The animals were decapitated 15 minutes after the injection (the time required for the lipoprotein lipase activity of the blood to reach a maximal level) and acetone powders of the hearts prepared. Two hearts were pooled in order to have sufficient material. Therefore, four control and four experimental values were obtained. In all cases, 50 mg. of powder
by guest on September 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

E. D. KORN 13
were extracted with 1 ml. of 0.025 N ammonia for 30 minutes at O’, the mixture was centrifuged, and the clear supernatant fluid assayed.
Although the series is not sufficiently great to permit a definite conclusion, there was an apparent tendency for the hearts of the heparin-treated ani- mals to have a lower lipoprotein lipase content than the controls (Table IX). A major portion of the original activity still remained, however, and the loss does not appear to be sufficient to account for the enzyme level in the blood. This might be explained by the contribution to the blood from other sources, or by an increased synthesis of enzyme protein in response to the elevated heparin concentration. It is obvious that the lipoprotein lipase had not been saturated with heparin, since its activity was stimulated by heparin added in vitro.
DISCUSSION
The experiments described in this paper definitely establish the presence in normal rat heart of an enzyme with all the known properties of post- heparin plasma clearing factor. This removes the major obstacle in the attempt to relate the phenomena which occur in postheparin plasma to normal lipide metabolism. These phenomena do occur in the absence of injected heparin but at the cellular level. The administration of heparin, therefore, appears to cause an overflow of the lipase into the blood, where it does not exist normally in significant quantities.
SUMMARY
Clearing factor, the enzyme present in postheparin plasma, can be ex- tracted from acetone powders of normal rat hearts. This enzyme catalyzes the hydrolysis of the neutral fat of chylomicrons. It is activated by heparin and inhibited by salt and protamine. A fatty acid acceptor is also required. The distribution of the enzyme in other tissues and the effect of heparin administration on the concentration of the enzyme in heart have been investigated.
The author wishes to thank Dr. Christian B. Anfinsen for his continued interest in this problem.
BIBLIOGRAPHY
1. Korn, E. D., Science, 120, 399 (1954). 2. Hahn, P. F., Science, 98, 19 (1943). 3. Anderson, N. G., and Fawcett, B., Proc. Sot. Exp. Biol. and Med., ‘74, 768 (1950). 4. Brown, R. K., Boyle, E., and Anfinsen, C. B., J. Biol. Chem., 204, 423 (1953). 5. Anfinsen, C. B., Boyle, E., and Brown, R. K., Science, 116, 583 (1952). 6. Bragdon, J. H., and Havel, R. J., Am. J. Physiol., 177, 128 (1954). 7. Kern, E. D., J. Biol. Chem., 216, 15 (1955).
by guest on September 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

14 LIPOPROTEIN LIPASE. I
8. Lambert, M., and Neish, A. C., C&ad. J. Res., !I&83 (1950). 9. Shore, B., Nichols, A. V., and Freedman, N. K., Proc. Sot. Exp. Biol. and Med.,
83, 216 (1953). 10. Havel, R. J., and Bragdon, J. H., Circulation, 10, 591 (1954). 11. Gordon, R. S., Boyle, E., Brown, R. K., Cherkes, A., and Anfinsen, C. B., Proc.
Sot. Exp. Biol. and Med., 84, 168 (19531. 12. Nikkilii, E. A., and Haahti, E., Acta them. Stand., 8, 363 (1954).
by guest on September 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from

Edward D. KornENZYME FROM NORMAL RAT HEART
CHARACTERIZATION OF THE LIPASE: I. ISOLATION AND
HEPARIN-ACTIVATED LIPOPROTEIN CLEARING FACTOR, A
1955, 215:1-14.J. Biol. Chem.
http://www.jbc.org/content/215/1/1.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
#ref-list-1
http://www.jbc.org/content/215/1/1.citation.full.htmlaccessed free atThis article cites 0 references, 0 of which can be
by guest on September 19, 2018
http://ww
w.jbc.org/
Dow
nloaded from





























