Embed Size (px)
Citation preview

Environmental Health Perspectives • VOLUME 109 | SUPPLEMENT 1 | March 2001 141
Climate Change
The earth’s climate has always been in a stateof change (1–3). For nearly three centuries ithas been in a warming phase. This was pre-ceded by a cold period, the Little Ice Age,which was itself preceded by a warmer phaseknown as the Medieval Warm Period, or LittleClimatic Optimum. Such changes are entirelynatural, but there is evidence that in recentyears a portion of the current warming may beattributable to human activities, particularlythe burning of fossil fuels (4–6). The potentialimpact of this global warming on humanhealth is a major subject of debate (7–10).
Many of the diseases that currently occurin the tropics are mosquito borne (11). It iscommonly assumed that their distribution isdetermined by climate and that warmer globaltemperatures will increase their incidence andgeographic range (12–14). This reviewexplores the validity of both assumptions byexamining the history of three mosquito-bornediseases—malaria, yellow fever, and dengue—in the context of past climates and of otherfactors that can influence their transmission.
ClimateClimate (Greek κλιµα , an inclination orslope—e.g., of the sun’s rays; a latitude zoneof the Earth) is commonly understood tomean the average weather in a given region orzone. In its older form, clime also included allaspects of the environment, including thezonal distribution of plants and animals. Bothdefinitions are unsatisfactory because theyimply that, unlike the obvious year-to-yearvariations of daily weather, long-term climateis a constant. By contrast, modern climatol-ogy recognizes that change is an inherent andfundamental feature of climate. Therefore,climatic values cannot be quoted withoutspecifying the time span to which they refer.
Natural factors that cause climaticvariability include fluctuations of the sun’sradiant energy, alterations in the trans-parency of the atmosphere (due to sand, vol-canic dust, and other airborne particles), andcyclic changes of the earth’s rotation on itsaxis and its orbit around the sun. In addi-tion, the circulations of the atmosphere andthe oceans, which are major components ofthe climate machine, are subject to internalvariations on time scales ranging from weeksto millennia. It is the complex interaction ofall these variables that generates the continu-ally changing patterns of climate. As a result,just as the yearly averages of climatic ele-ments—such as temperature, humidity,rainfall, wind, and airborne particles—differfrom one another, so too do the averages fordecades, centuries, millennia, and millionsof years (Table 1).
Climate is a major parameter in allecosystems and has always been a fundamen-tal factor in human settlement, economy,and culture. Episodes of second-order cli-mate change, such as the end of the Ice Age,the drying of the Sahara, the waning of theMedieval Warm Period, and the onset of theLittle Ice Age, have had an important impacton human history (1,2,15). However, aware-ness of such change has remained shadowy atbest, probably because the inherent timescales are beyond the span of individualhuman experience.
During most of geologic history, temper-ate conditions extended to the polar circles,and the planet was relatively ice-free (3,16).Indeed, in the perspective of the past 600million years, present global temperaturesare relatively cool. In the context of theselong-term averages, the warming trend thatbegan in the late 1970s (see below) is aminor fluctuation, less than a first-ordervariation.
Weather
Weather, the short-term condition ofclimate, has a much more direct and tangibleimpact on daily life. Since earliest times,weather has been fundamental to the successof human activities, from agriculture to sea-faring, from warfare to leisure. The universalbelief in weather deities, the prominence ofweather events in folklore, and the ubiqui-tous preoccupation with weather signs andportents are evidence that an awareness ofweather, particularly a fear of inclementevents, has been a major feature of thehuman psyche throughout history.
The significance of weather has notdiminished in modern society. Indeed, in thepast few decades, weather awareness, particu-larly in the global context, has reachedunprecedented levels. Weather forecasting hasbecome an important science, fundamental tothe success of agriculture, transportation,trade, tourism, and virtually every otheraspect of human enterprise. Weather data arecollected from every corner of the globe anddisseminated in digested form by governmentand private agencies as an aid to decisionmaking in all walks of life. Continuallyupdated forecasts and other information areavailable to the public via the popular media.Disastrous weather events from around theworld are a major news feature, with detaileddescriptions and graphic illustration. Withthis awareness of weather has come a newrealization of the changeability of climate.
Recent Climate ChangeCurrently, world climate is in a warmingphase that began in the early decades of theeighteenth century. Temperatures, at least inthe Northern Hemisphere, are now broadlysimilar to what they were during the MiddleAges, in the centuries before the Little Ice Age(2). Awareness of this warming has led to anew preoccupation: concern that humanactivities may be affecting the natural climatic
Global atmospheric temperatures are presently in a warming phase that began 250–300 years ago.Speculations on the potential impact of continued warming on human health often focus onmosquito-borne diseases. Elementary models suggest that higher global temperatures will enhancetheir transmission rates and extend their geographic ranges. However, the histories of three suchdiseases—malaria, yellow fever, and dengue—reveal that climate has rarely been the principaldeterminant of their prevalence or range; human activities and their impact on local ecology havegenerally been much more significant. It is therefore inappropriate to use climate-based models topredict future prevalence. Key words: Aedes aegypti, anopheles, climate change, dengue, globalwarming, malaria, mosquito, public health, vector, yellow fever. — Environ Health Perspect109(suppl 1):141–161 (2001).http://ehpnet1.niehs.nih.gov/docs/2001/suppl-1/141-161reiter/abstract.html
Address correspondence to P. Reiter, CDC DengueBranch, 1324 Calle Cañada, San Juan, PR 00920-3860.Telephone: (787) 706 2399. Fax: (787) 706 2496.E-mail: [email protected]
I am grateful to many people who gave helpfulcomments on the manuscript, especially D. Gubler,G. Clark, J. Rigau-Pérez, S. Bernard, K. Ebi, and E.Reiter, and to M. Wolcott for help with the figures.
Received 18 July 2000; accepted 27 September2000.
Climate Change and Mosquito-Borne Disease
Paul Reiter
Dengue Branch, Division of Vector-Borne Infectious Diseases, National Center for Infectious Diseases, Centers for Disease Control andPrevention, U.S. Department of Health and Human Services, San Juan, Puerto Rico
109S1.Part 2 02/21/01 3:32 PM Page 141 (Black plate)

Reiter
142 VOLUME 109 | SUPPLEMENT 1 | March 2001 • Environmental Health Perspectives
regime, just as they are changing many otheraspects of the environment. From the 1940sto the late 1970s, when global temperatureswere in decline, there was concern that partic-ulate industrial pollutants might be exerting aglobal cooling effect (17–19). Since then, asworld climates returned to a warming mode,interest has switched to the greenhouse effect,a natural phenomenon by which atmosphericgases trap solar radiation in the form of heat.
Although the principal greenhouse gas iswater vapor, about 2% by volume, publicattention focuses mainly on carbon dioxide, agas that is essential as the ultimate source ofcarbon for nearly all life on the planet. Fromthe mid–nineteenth century onward, massiveclearance of forests for agriculture, followedby an exponential rise in the combustion offossil fuels (coal, oil, gas), has produced ameasurable increase in atmospheric CO2from around 0.029% in 1890 to 0.037%today. Many climatologists agree that this28% increase in atmospheric CO2 togetherwith an increase in other anthropogenicgreenhouse gases may be contributing to thewarming trend of recent decades (4–6,20).The extent of this contribution remains farfrom clear, but the mere possibility thathuman activities may be involved implies thatthe trend could be reversible, and has gener-ated a spirited scientific (5,7,21–23) and pub-lic (24–26) discussion. Human health—andmosquito-borne disease in particular—is aprominent topic in this debate (12,27–32).
Mosquito-Borne Disease
Mosquitoes As Vectors of Disease
Mosquitoes are found throughout the worldexcept in places that are permanently frozen.There are about 3,500 species, of whichnearly three-quarters are native to the humidtropics and subtropics. The largest popula-tions of individual species occur in the Arctictundra, where colossal numbers emerge in asingle brood each summer from snowmeltpools that overlie the permafrost.
In nearly all mosquito species, the femaleobtains the protein she needs for the devel-opment of her eggs by feeding on vertebrateblood. Some species are highly selective,restricting themselves to one or at most afew closely related host species. Others havea less clearly defined host preference and
may alternate among birds, mammals, andeven reptiles. A complex salivary secretionfacilitates feeding. It is the direct injection ofthis fluid into the capillaries that enablesseveral life forms—viruses, protozoa, andnematode worms—to exploit mosquitoes asa means of transfer between vertebrate hosts.In nearly all cases, there is an obligatoryphase within the insect. This includes a stagein which they multiply prodigiously in thesalivary glands, from which they can beinoculated into a new host during a laterblood meal. Although most such organismsdo not appear to affect either the mosquitoesor their vertebrate hosts, some are pathogensof important human and animal diseases.
Factors That Influence TransmissionClimatic factors. The ecology, development,behavior, and survival of mosquitoes and thetransmission dynamics of the diseases theytransmit are strongly influenced by climaticfactors. Temperature, rainfall, and humidityare especially important, but others, such aswind and the duration of daylight, can also besignificant. The same factors also play a cru-cial role in the survival and transmission rateof mosquito-borne pathogens. In particular,temperature affects their rate of multiplica-tion in the insect. In turn, this affects the rateat which the salivary secretions becomeinfected, and thus the likelihood of successfultransmission to another host. Of course, ifthe development time of the pathogenexceeds the life span of the insect, transmis-sion cannot occur. It is the complex interplayof all these factors that determines the overalleffect of climate on the local prevalence ofmosquito-borne diseases (11,33).
Seasonality is a key component of climate.Summer temperatures in many temperateregions are at least as high as in the warmestseasons of much of the tropics. The crucial dif-ference is that the tropics do not have cold win-ters. Tropical crops such as rice and groundnutscan be cultivated in temperate regions if theyare planted in springtime. Similarly, if tropicalmosquito-borne pathogens are introduced inthe right season, they can be transmitted if suit-able vectors are present; in most cases they areeliminated when winter sets in.
Mosquitoes native to temperate regionshave had to evolve strategies to survive thewinter, as have the pathogens that they
transmit. In the tropics, comparableadaptations are necessary for surviving inunfavorable dry periods, which can last forseveral years. In both cases, such adapta-tions impose a seasonality on transmission.For example, before eradication, the trans-mission season for Plasmodium falciparumin Italy was July–September (34). Thesame 3 months constitute the malaria sea-son in Mali , where the disease is st i l lendemic (35).
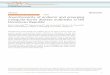
The physical environment is an importantmodifier of local climate (36). In dense forest,daily mean temperatures at ground level canbe as much as 10°C lower than in adjacentopen areas (37). Similarly, natural daytimetemperatures indoors can be several degreeshigher or lower than outdoors, depending ondesign, construction materials, and ventila-tion. Mosquitoes use a variety of strategies toexploit the timing and location of such micro-climates to maximum advantage. For exam-ple, in Anopheles gambiae (38), a physiologic“clock” ensures that whatever the rate of meta-morphosis in the pupal stage, the adultsalways emerge from the water at sundown orin the early hours of the night. In the labora-tory, if An. gambiae pupae are maintained inconstant light, the duration of the pupal stageis a direct function of temperature: It lastsabout 2 days at 22°C but only 1 day at 32°C(Figure 1A). However, in a light regime of 12hr light and 12 hr dark (LD 12:12), the tim-ing is modified (Figure 1B) to minimize emer-gence in daylight (39,40). Similar circadianrhythms ensure that other behaviors such asfeeding, resting, and oviposition are restrictedto optimum times, regardless of ambient tem-perature (41). Thus, An. gambiae can survivein the Sudan (mean monthly temperatures of42°C; actual outdoor temperatures can beover 55°C) by emerging after sunset, hiding inthe thatch of buildings in the daytime, feedingafter midnight, and ovipositing at dawn ordusk (42).
Analogous survival strategies operate intemperate climates. Culex pipiens, a vector ofthe St. Louis encephalitis virus (and therecently imported West Nile virus), is com-mon in North America as far north as NewBrunswick and Nova Scotia, Canada. It over-winters in the adult stage, sequestered at sitesprotected from the cold. Here again, light (inthis case, daylength) is the zeitgeber or cuethat initiates the survival strategy. Studies(43) in Memphis, Tennessee, revealed aremarkably high winter survival rate (p =0.97) in an underground storm sewer.Survival was the same in a cold winter(1981–1982: 42 days of freezing tempera-tures, mean daily minimum temperature[December 1–February 28] –0.74°C; 9 dayswith minimum of less than –10°C) as in amuch warmer “El Niño” winter (1982–1983;
Table 1. Time scale of climatic variation.a
Order Interval (years) Descriptor Example of variation
1 30–100 Instrument record Cooling trend from 1940 to late 1970s2 100–1,000 Historical Medieval Warm Period, Little Ice Age3 1,000–10,000 Glacial Variation within an Ice Age4 10,000–1 million Minor geologic Ice Age5 1–100 million Major geologic Interval between Ice AgesaGlobal events such as the El Niño Southern Oscillation phenomenon occur at a frequency well below that of the first order of variation.
109S1.Part 2 02/21/01 3:32 PM Page 142 (Black plate)

Climate change and mosquito-borne disease
25 days of freezing temperatures; mean dailyminimum temperature [December 1–February 28] +2.9°C; 1 day of less than–10°C, maximum temperatures as high as27.2°C).
Memphis has a typical temperate conti-nental climate. There are normally about 65days per year with maximum temperaturesabove 32°C (temperatures above 38°C arenot unusual) and about 60 days when theminimum falls below 0°C (lows of –15°C arenot unusual) (44). Aedes aegypti, the principalurban vector of dengue and yellow fever, is atropical species for which temperatures below0°C are fatal. Nevertheless, at the time of thestudies mentioned above (and probably for atleast 200 years before), Ae. aegypti was presentthroughout the southern United States, andwas common in Memphis. Presumably it sur-vived during winter in niches protected fromthe cold. Despite the effectiveness of thisstrategy, in the past 15 years the species hasdisappeared from much of the region, includ-ing Memphis, displaced by another non-native species, Ae. albopictus (45). Thisexample and the others given above empha-size that meteorologic variables alone are oflimited value as a guide to the developmenttimes, behavior, and geographic range of vec-tor species and the pathogens they transmit.
Human factors. Human activities are alsocrucial to transmission. Forest clearance elim-inates species that breed in water in tree holes(e.g., the forest Aedes species that transmityellow fever) but provides favorable condi-tions for those that prefer temporary groundpools exposed to full sunlight (e.g., many ofthe Anopheles species that transmit malaria).Drainage of wetlands eliminates the marshypools exploited by many species but can pro-vide the open channels preferred by others(e.g., some important European vectors ofmalaria, and Culex tarsalis, a vector of St.Louis encephalitis). Agricultural fertilizers canpromote the growth of algae and other larvalnutrients, whereas herbicides may eliminatethem altogether. Cisterns, pit latrines,sewage-polluted ditches, storm drains, andblocked gutters can support large populationsof Cx. quinquefasciatus, an important vectorof Bancroftian filariasis. Wells are often a sig-nificant source of malaria vectors. Water-storage jars and drums, cemetery urns,discarded rubber tires, buckets, pots, andother man-made containers can be prolificsources of Ae. aegypti, an important perido-mestic vector of yellow fever and dengue andother species that originally bred in tree holes.
Additional factors arise from behaviorand cultural traits. Daily activity patterns—work, rest, and recreation—the location ofhomes in relation to mosquito breedingsites, the design of buildings, the materialsused to build them, the use of screens and
bed nets, and many other factors can be sig-nificant. Finally, chemotherapy, vaccination,and mosquito control have played majorroles in reducing transmission in many partsof the world.
Predicting the Impact of ClimateChangeTransmission models. Much of the recentspeculation on the possible effects of climatechange on mosquito-borne disease hasfocused on rudimentary concepts of theirtransmission dynamics (30,46–48). An exam-ple is vectorial capacity, a convenient way ofexpressing transmission risk:
where m is the mosquito density per human,a is the average number of bites per day foreach mosquito, p is the probability of a mos-quito surviving through any one day, and n isthe extrinsic incubation period—the timetaken for the pathogen to develop in the mos-quito until the insect becomes infective. Theonly factor directly affected by a climate vari-able is n, which is inversely related to temper-ature. Because p is less than unity, pn willincrease at higher temperatures, although pitself may decrease as a result of other factors.However, the denominator is an exponentialfunction, so p, survival rate, is dominant.
Caged mosquitoes can live for 3–4months, but their median age in the field iscommonly about 4–6 days. Clearly, in naturefew die of senescence; most are killed bypredators, disease, and other hazards longbefore they reach old age (49). Feeding andthe search for oviposition sites are probablythe most hazardous activities. Species thatbecome inactive (e.g., in winter or in the dryseason) are known to survive for as long as 9months.
Vectorial capacity and similar elementarymodels were developed to explore the funda-mental features of the transmission of mos-quito-borne disease (50–53), mainly in thecontext of mosquito control operations.Except for n, the calculation of C depends onquantitative values that can be obtained onlyin the field. However, it is hard to estimatethese values realistically because their meas-urement depends heavily on a complex rangeof assumptions (54–57). Moreover, C is lim-ited to entomologic parameters and the dura-tion of extrinsic incubation; it does notincorporate the parasite rate in humans ormosquitoes, nor any of the many ecologicand behavioral factors outlined in the previ-ous section. Thus, although helpful in ourunderstanding of transmission dynamics andas a practical tool in specific situations, suchmodels have a limited value for assessing theimpact of long-term climate change ondisease transmission (58,59).
Cma p
p
n
e
=−
2
log
Environmental Health Perspectives • VOLUME 109 | SUPPLEMENT 1 | March 2001 143
Figure 1. The influence of light regime on the timing of the emergence of adult An. gambiae from the pupal stage atdifferent temperatures in the laboratory. (A) Duration (mean + SD) of the pupal stage in constant light (LL). (B) Timingof emergence in LL and in a daily regime of 12 hr light:12 hr dark (LD 12:12). In the LL regime (open histograms, down-ward) eclosion occurs as soon as metamorphosis is complete. In the LD 12:12 regime (open histograms, upward)emergence is delayed to give a peak near the time of light-off. P indicates the 6-hr period during which the pupaewere collected in hourly batches. In the laboratory, pupation normally peaks 3–4 hr before the end of the light phase(41). In the field, pupation and pupal/adult eclosion are both delayed until sunset (187) or thereafter. Physiologic“gates” of this type maintain regular rhythms of physiology and behavior regardless of the effects of temperature.Figure reprinted from Reiter and Jones (39), with permission of Blackwell Science Ltd.
●
●
●
●
● ●
50
45
40
35
30
25
20
15
10
5
022 24 26 28 30 32 34 36
Temperature (°C)
Dura
tion
of p
upal
sta
ge (h
r)
A
0 24 48
LD 12:12
B
Adul
ts e
mer
ging
(per
1/2
hr)
P
40
0
40
Time (hr)
34.5°
32.0°
29.5°
27.0°
24.5°
22.0°
LL
109S1.Part 2 02/21/01 3:32 PM Page 143 (Black plate)

Reiter
144 VOLUME 109 | SUPPLEMENT 1 | March 2001 • Environmental Health Perspectives
An alternative approach is to look at thepast. The history of mosquito-borne diseasesat different latitudes and in different climaticeras can help us assess the significance of cli-mate variables in the context of the manyother factors that affect transmission.
History of climates. Climatology is a rela-tively young science. Apart from a handful ofsites, systematic records of climate variablesare available only for the past century, mainlyfrom land-based stations in the NorthernHemisphere. Global data, obtained byweather satellites, are available only for thepast few decades. Thus the time span ofdirect studies of climate is within the firstorder of climatic variation, less than theperiod for valid long-term change (Table 1).
Fortunately, a large amount of alternativedata is available. Documentary informationfrom annals, chronicles, audited accounts, agri-cultural records, tax ledgers, and a wealth ofother archival material provides a rich sourceof indirect information, particularly on thetiming and occurrence of extreme weatherevents such as drought, flood, or unusual cold.Descriptions of wind direction, wind speed,cloud formations, and other weather indicatorsare available from private diaries, ships’ logs,accounts of military campaigns, and similarsources. Substantial lines of evidence are alsoavailable from archaeologic, geologic, fossil, andother sources. They include the study of glacialmoraines, lake and ocean sediments, pollenstrata, deposits of insects, tree rings, coral struc-ture, radiometric analysis of ice cores, andmany other indicators. All this evidence makesit possible to realistically reconstruct past cli-mates over the course of time (2,16).
History of mosquito-borne disease. Thescientific study of mosquito-borne disease isalso limited to the past 100 years. Fortunately,the symptoms of three important diseases—malaria, yellow fever, and dengue—are fairlydistinctive. Thus, as with climatology, we canturn to a variety of sources for evidence oftheir occurrence in past climates. The bulk ofthis review is devoted to such evidence.Particular emphasis is placed on the temperatelatitudes because of the wealth of historicalmaterials that is available and because modelssuggest that the impact of global warmingmay be greatest at these latitudes.
Malaria
The Disease
Malaria (Italian mala aria, bad air) is a proto-zoan (genus Plasmodium) infection transmit-ted by mosquitoes of the genus Anopheles.The four species of Plasmodium that infecthumans appear to have evolved from a com-mon ancestor during the early Tertiaryperiod, some 60 million years ago (60). Thus,some or all of them probably co-evolved with
the genus Homo and have affected humanssince their earliest days.
The clinical course of malaria (33) generallyinvolves distinctive bouts of fever alternatingwith periods of freedom from illness. Eachepisode begins with a short period (15 min–1hr) of violent shivering, with the patient hav-ing cyanotic lips and fingers, a rapid but weakpulse, and a feeling of intense cold (rigor).This cold stage is followed by 2–6 hr of dis-tressing heat, accompanied by flushed skin,intense headaches, nausea, and a full, bound-ing pulse. The hot stage is followed by profusesweating and a rapid drop in temperature,often to levels below normal.
A prominent feature of the febrile boutsis their periodicity: They tend to recur everythird day with Plasmodium. vivax, P. falci-parum, and P. ovale and every fourth daywith P. malariae. From earliest times thisperiodicity gave rise to the descriptors tert-ian and quartan fevers. Another commonterm was aestivo-autumnal or harvest feverbecause in temperate regions epidemic trans-mission tended to occur in late summer andautumn (33).
Malaria in Temperate ClimatesPrehistory. About 60 species of Anophelescan transmit human malaria. Those thatexist in Europe probably began colonizingthe region as the ice caps retreated at the endof the Pleistocene epoch. Human popula-tions, also moving northward, almost cer-tainly brought malaria parasites along withthem. The prevalence of the disease in theNeolithic period and the Bronze Age isuncertain, but it is unlikely to have beenvery high because human groups were gener-ally small and widely scattered. Nevertheless,early Neolithic skeletal remains fromAnatolia and Macedonia exhibit pathologicchanges that have been attributed to chronicanemia caused by P. falciparum infection.The widespread presence of thalassemia(Mediterranean anemia), a genetic conditionthat gives some protection against malaria,may also indicate a long history of contactwith the pathogen.
Ancient Greece and Rome. The introduc-tion of agriculture, around 7000 B.C., led toincreased populations of relatively settled peo-ple and increasingly favorable conditions formalaria transmission (34). The extensivedeforestation that began at this time may alsohave contributed to prevalence by creatingadditional habitat for anopheline mosquitoes.Similar ecologic changes in modern timeshave caused major increases in the prevalenceof the disease.
Contemporary accounts together with fos-sil and other evidence suggest that a gradualwarming and drying occurred in theMediterranean region throughout classical
times until about A.D. 400 (2). Landscapestudies suggest a gradual rise in sea level overthis period. Around 300 B.C., beech trees(genus Fagus) grew in Rome, the Tiber Riverfroze in winter, and snow lay for many days.However, by the first century A.D. theRomans considered the beech a mountaintree, and winters were definitely less severe.Over these centuries, the cultivation of thevine and olive moved gradually northwardalong the Italian peninsula. The Romans wereeven able to introduce wine growing toGermany and Britain, and import data sug-gest that Britain became self-sufficient in wineproduction by around A.D. 300. The warmingtrend is clearly indicated by tree-ring studiesin California, so it may have been a worldwideor at least a hemispheric phenomenon (2).
The first literary mention of an autumnalfever is in Homer’s Iliad (800 or 900 B.C.), asAchilles sets out to fight Hector (61):
And old King Priam was first to see him coming,surging over the plain, blazing like the star that rears at harvest, flaming up in its brilliance—far outshining the countless stars in the night sky, that star they call Orion’s Dog—brightest of all but a fatal sign emblazoned on the heavens, it brings such killing fever down on wretched men.
We cannot be certain that this wasmalaria, but the reference to killing fevers atharvest time is one that recurs many times inother descriptions of malaria. Later texts con-firm that the disease had become a significantfeature in Greek life. Indeed, there is evidencethat a major wave of malaria began with theflowering of Greek civilization and transmis-sion rates continued to increase throughoutthe period of the Roman Empire (2,62).Hippocrates (460–377 B.C.) gave exquisitelydetailed descriptions of the course and relativeseverity of tertian versus quartan infections(63). He also noted their association with wet-lands and even observed that splenomegaly(enlarged spleen, often a symptom of chronicmalaria infection) was particularly prevalent inpeople living in marshy areas.
Praxagoras, Heraclides, and other medicalwriters gave similar descriptions, from whichit appears that much of Greece had becomehighly malarious (62).
There is a wealth of evidence that malariawas common in imperial Rome (34). Horace,Lucretius, Martial, and Tacitus were amongmany Latin authors who mentioned the dis-ease. The Pontine Marshes, close to the city,were notorious as a source of infection.Marcus Terentius Varro (116–127 B.C.)described these loca palustria (fever areas) andattributed the disease to animalia quaedamminuta quae non possunt oculi consequi (ani-mals too small to be seen). He advised thathouses should be built in high, well-ventilated places so that the bestiolae that bred
109S1.Part 2 02/21/01 3:33 PM Page 144 (Black plate)

Climate change and mosquito-borne disease
below would be blown away, and evenmentioned the use of nets to protect againstthe bites of mosquitoes (64). Later, in the sec-ond century A.D., the detailed writings ofGalen and Celsus on the symptoms and treat-ment of “intermittent fevers” give clear evi-dence that three species of parasite—P. falciparum, P. ovale, and P. vivax—werecommonly present (2,65).
The Dark Ages. Relatively l itt le isknown of climate after the Roman era, dur-ing the Dark Ages, but there seems to havebeen a cooling trend from the fifth centuryonward. There were certainly some out-standingly cold winters. In A.D. 763–764there was ice on the sea in the Dardanelles,and in 859–860 the sea ice on the Adriaticwas strong enough to support heavy wag-ons. In 1010–1011 it was even cold enoughfor ice to form on the Nile. Again, tree-ringdata from California indicate that thiscooling was not restricted to Europe.Nevertheless, the armies of Visigoths,Vandals, Ostrogoths, and other barbariansthat swept the continent had to contendwith malaria, often as a major setback totheir campaigns. Several popes and church-men, including St. Augustine, the firstArchbishop of Canterbury, died of malariaduring their journeys to Rome. Around theturn of the millennium, the armies of Ottothe Great, Otto II, and Henry II sufferedseverely from the “Roman Fever” duringtheir sieges of the Holy City.
The Middle Ages. The Medieval WarmPeriod, which reached its peak around 1200,coincided with major advances in technologyand agriculture and a significant increase inpopulation throughout most of Europe. TheVikings were able to establish self-sufficientcolonies, based on oats and barley, in north-ern Scandinavia, Iceland, and Greenland. Inthe British Isles, tillage was extended to muchhigher altitudes than is possible today—sohigh that sheep farmers complained that toolittle land was left for grazing. English vint-ners were able to maintain a flourishing pro-duction of high-quality wine despite effortsby Bordeaux traders to have their exportsrestricted by treaty.
The explosion of economies and culturethat occurred during this warming period hasbeen attributed at least partly to the beneficialimpact of the warming climate. Fromcaliphate Spain to Christian Russia, numer-ous medieval writers mentioned agues, inter-mittent fevers, tertians, quartans, and the like.A classic example appears in The Inferno,where the Florentine poet Dante (1265–1321) wrote (66):
. . . Like those who shake,Feeling the quartan fever coming on—Their nails already blue, so that they shiverAt the mere sight of shade—such was I then;
Later, in The Canterbury Tales, the Englishpoet Chaucer (1342–1400) wrote (67):
Your face is choleric and shows distension;Be careful lest the sun in his ascension Should catch you full of humours, hot and many.And if he does, my dear, I’ll lay a penny It means a bout of fever or a breath Of tertian ague. You may catch your death.
Favorable temperatures and rainfall mayhave enhanced transmission of malaria in ear-lier years, but Chaucer’s lifetime coincidedwith a cooling trend that culminated in aseries of severely cold winters in the firstdecades of the fifteenth century (68). Muchof the earlier agricultural expansion wasreversed. There were many years of famineand a large-scale abandonment of farms.Despite this cooling, malaria persisted, evenin northern regions (34).
The Little Ice Age. The first half of thesixteenth century was warm again.Temperatures were probably quite similar tothose of the period 1900–1950. In the mid-dle of the century, however, a remarkablysharp change occurred. After a decade or so ofparticularly warm years—warm enough foryoung people to bathe in the Rhine River inJanuary—the winter of 1564–1565 was bit-terly cold (2). The next 150–200 years—dubbed the Little Ice Age—were probably thecoldest era of any time since the end of thelast major ice age, some 10,000 years ago(16). Yet despite this spectacular cooling,malaria persisted throughout Europe (34).
William Shakespeare (1564–1616) wasborn in the year of that first fierce winter, yetthere are 12 mentions of ague in his writings.He also made several allusions to the associa-tion between swampy land and disease, andthe name Sir Andrew Aguecheek presumablyreferred to the trembling cheeks of his obesehero. The years 1594–1597 were so cold andwet that wheat harvests were a disaster, yetWilliam Harvey (1578–1657) missed muchof his final year at the University ofCambridge, England, in 1597 because ofmalaria (69). In later years he made carefulobservations of malaria cases in London. Themarshes in the Borough of Westminster,where the Houses of Parliament now stand,were notoriously malarious. In his treatise Onthe Motion of the Heart and Blood in Animals(1628) Harvey described the clinical pathol-ogy of the febrile episodes, including thechanges in the consistency of blood thatoccur in serious cases (70):
In tertian fever, . . . in the first instance . . .the patient [is] short-winded, disposed tosighing, and indisposed to exertion, . . . theblood [is] forced into the lungs and renderedthick. It does not pass through them (as Ihave myself seen in opening the bodies ofthose who had died in the beginning of the
attack), when the pulse is always frequent,small, and occasionally irregular; but the heatincreasing, . . . and the transit made, thewhole body begins to rise in temperature, andthe pulse becomes fuller and stronger. Thefebrile paroxysm is fully formed . . . .
Another notable physician, ThomasSydenham (1624–1689), also lived throughsome of the coldest years of the era, yet madefrequent reference to tertians and quartans(34). He even remarked, “When insects doswarm extraordinarily and when . . . agues(especially quartans) appear early as about mid-summer, then autumn proves very sickly” (34).
Not all the summers of the Little Ice Agewere cool. The overall mean temperature wasprobably at least 1°C cooler than in the twen-tieth century, but there also seems to havebeen an enhanced variability of the climate,with wide differences between clusters of upto 6–8 years. Thus, diarist Samuel Pepys(1633–1703), who suffered from malaria,remarked that the summers of 1661, 1665,and 1666 were remarkably hot (34), andSydenham described an epidemic of “tertianand some quartan” fevers that broke out in1661, which “was doing frightful mischief”by August [quoted in Bruce-Chwatt andde Zulueta (34)].
Warm summers may have contributed tothis and other outbreaks, but transmissionwas not restricted to such years. In1657–1658, snow lay on the ground for 102days—exceptionally cold even with respect tothe climate of the times—yet OliverCromwell (1599–1658) died of malaria inSeptember 1658 just as another severe winterwas setting in. Temperatures were probably attheir lowest from 1670 to 1700, yet this wasthe very period that Robert Talbor (ca.1642–1681) was able to persuade the aristoc-racy of England and Europe to buy the pre-scriptions, based on cinchona bark, that hehad developed in the marshlands of Essex(34,71–74). In the same period, DanielDefoe (1660–1731) described life in theDengie marshes of Essex, England (75):
a strange decay of the [female] sex here. . . . itwas very frequent to meet with men that hadhad from five to six, to fourteen or fifteenwives . . . the reason. . . was this; that they[the men] being bred in the marshes them-selves, and seasoned to the place, did prettywell with it; but that they always went intothe hilly country. . . for a wife: that whenthey took the young lasses out of the whole-some and fresh air, they were healthy, freshand clear, and well; but when they came outof their native aire into the marshes. . . theypresently changed their complexion, got anague or two, and seldom held it above half ayear, or a year at most; and then. . . [the men]would go to the uplands again, and fetchanother; so that marrying of wives was reck-oned a kind of good farm to them (75).
Environmental Health Perspectives • VOLUME 109 | SUPPLEMENT 1 | March 2001 145
109S1.Part 2 02/21/01 3:33 PM Page 145 (Black plate)

Reiter
146 VOLUME 109 | SUPPLEMENT 1 | March 2001 • Environmental Health Perspectives
Dobson (76–79) has masterfully researchedthe demography, epidemiology, and socialimpact of malaria in England in this period.The disease was especially prevalent in areasof brackish marshland, the preferred habitatof an effective vector, An. atroparvus. Datafrom burial records show that mortality ratesin “marsh parishes” were much higher thanthose in upland areas and were comparable tothose in areas of stable malaria transmissionin sub-Saharan Africa today (80).
After the Little Ice Age. From the earlyeighteenth century until the present, tempera-tures have gradually returned to levels thatprevailed before the mid-sixteenth century.However, the marked variability of the LittleIce Age persisted for at least 150 years. Indeed,in the 1770s, much as is happening today,there was alarm that the climate was becom-ing increasingly erratic, and this prompted anew emphasis on the recording of weathervariables. Some of the cold periods, particu-larly those between 1752 and the 1840s, wereprobably caused by major volcanic eruptions.Whatever their causation, such episodes—accompanied by major advances of the Alpineglaciers from 1820 to 1850—persisted until amore lasting warmth was established in thelate nineteenth century (2,16).
A wealth of records in the eighteenth andnineteenth centuries reveals the northernlimits of malaria transmission. In the BritishIsles, the disease was common in most ofEngland and in many parts of Scotland, withoccasional transmission as far north asInverness. It was endemic throughoutDenmark, coastal areas of southern Norway,and much of southern Sweden (81) andFinland (82). In Russia it was common in theBaltic provinces and eastward at similar lati-tudes throughout Siberia. The current aver-age January temperature in some of theseregions is less than –20°C (–4°F). Clearly, thedistribution of the disease was determined bythe warmth of the summers, not by the cold-ness of the winters. The northern limit oftransmission (83) was roughly defined by thepresent 15°C July isotherm—not the 15°Cwinter isotherm, as stated in several widelycited articles on climate change (27,28).
Decline of malaria. In the second half ofthe nineteenth century malaria began todecline in much of northern Europe.Denmark suffered devastating epidemicsuntil the 1860s, particularly in the country-side around Copenhagen, but thereaftertransmission diminished and had essentiallydisappeared around the turn of the century(84). The picture was similar in Sweden(Figure 2), although isolated cases were stillbeing reported until 1939 (81). In England,there was a gradual decrease in transmissionuntil the 1880s, after which it dropped pre-cipitously and became relatively rare except
in a short period following World War I(34,85). In Germany, transmission alsodiminished rapidly after the 1880s; afterWorld War I it was mainly confined to a fewmarshy localities (34). The last outbreak oflocally transmitted malaria in Paris occurredin 1865 during the construction of theGrands Boulevards, and the disease hadlargely disappeared from the rest of Franceby the turn of the century (86,87). InSwitzerland, most foci had disappeared bythe 1890s (88).
The decline of malaria in all these countriescannot be attributed to climate change becauseit occurred during a warming phase when tem-peratures were already much higher than in theLittle Ice Age. However, a host of other factorscan be identified.
First, improved drainage, reclamation ofswampy land for cultivation, and the adop-tion of new farming methods (there is an oldItalian saying: “malaria flees before theplough”) all helped eliminate mosquito habi-tat. Second, new root crops such as turnipsand mangel-wurzels were adopted as winterfodder. These enabled farmers to maintainlarger numbers of animals throughout theyear, thus diverting mosquitoes from feedingon humans. Third, selective breeding of cattleplus new introductions (e.g., the Chinesedomestic pig) in combination with the newfodder crops enabled farmers to keep largepopulations of stock in farm buildings ratherthan in open fields and woodland. Thesebuildings provided attractive sites for adultmosquitoes to rest and feed, diverting themfrom human habitation.
Changes in demographics and humanliving conditions were also significant. Ruralpopulations declined as manual labor wasreplaced by machinery. This further reducedthe availability of humans (vs. animals) ashosts for the mosquitoes and of humans ashosts for the parasite. New building materialsand improvements in construction methodsmade houses more mosquito-proof, especiallyin winter, another factor that reduced contactwith the vector. Finally, greater access tomedical care and wider use of quininereduced the survival rate of the malariaparasite in its human host.
Much of the decline in malaria camebefore recognition of the role of mosquitoesin its transmission. Thus, for most of theregion, deliberate mosquito control playedlittle or no role in its eventual elimination.
Persistence of malaria in the then-SovietUnion and neighboring countries. In coun-tries where profound changes in crop produc-tion and stock rearing were absent, malariadid not decline.
In Russia, from the Black Sea to Siberia,major epidemics occurred throughout thenineteenth century, and the disease remained
one of the principal public health problemsfor the entire first half of the twentieth cen-tury (34). In 1900, annual incidence in mili-tary garrisons was 6.6 per 1,000 inSt. Petersburg, 31.0 per 1,000 in Moscow,and several hundred per 1,000 in the moresoutherly provinces. Mean annual incidencefrom 1900 to 1904 was 3,285,820, but bythe period from 1933 to 1937, it had risen to7,567,348. Some of this increase can beattributed to more effective reporting, butthere is no doubt that the disease becamemuch more prevalent after World War I andthe 1917 Revolution. In the 1920s, in thewake of massive social and economic disrup-tion, 2 years of severe drought, and a year ofwidespread flooding, a pandemic sweptthrough what was then the entire SovietUnion. Official figures for 1923–1925 listed16.5 million cases, of which not less than600,000 were fatal (89). Tens of thousands ofinfections, many caused by P. falciparum,occurred as far north (Figure 2) as the Arcticseaport of Archangel (61° 30´N).
The Soviet government appeared to makesome headway against the disease in the1930s, mainly by drainage schemes, afforesta-tion, and naturalistic methods such as the useof mosquito-eating fish. World War II inter-rupted these efforts and transmission soared,particularly in the Ukraine, Belorussia, andother occupied areas. Finally, in 1951, a hugemultifaceted antimalaria campaign was initi-ated. It involved widespread use of DDT andother residual insecticides, antimalarial ther-apy, land reclamation, water management,public health education, and many otherapproaches. This mammoth effort finallybrought about a dramatic reduction of trans-mission, so that by the mid-1950s thenational annual incidence was below 1 per10,000 (34).
In neighboring Poland, the disease wasstill present after World War I, and a large-scale epidemic coincided with the 1920s pan-demic in the Soviet Union. In the 1930s,efforts to control the disease were largely suc-cessful, although endemic foci persistedthroughout the country (34). However, anew wave of transmission followed WorldWar II (90). This peaked in 1948, afterwhich an intensive campaign of DDT spray-ing and antimalarial therapy finally elimi-nated the disease (Figure 3). In Finland,malaria was still a problem after it had practi-cally disappeared in neighboring Sweden.Significant transmission occurred during theFinno-Russian war (1939–1940). The diseasefinally died out in the late 1940s (82).
The contrast between the devastationcaused by malaria in the Soviet Union untilthe 1950s, and its quiet withdrawal fromother countries at similar latitudes in the pre-vious century illustrates the importance of
109S1.Part 2 02/21/01 3:33 PM Page 146 (Black plate)

Climate change and mosquito-borne disease
nonclimatic factors in transmission. Until thecollectivization of farmland that began in thewinter of 1929–1930, the Soviet Union hadbeen largely unaffected by the agriculturalrevolution. By 1936, all farming was essen-tially in government hands, but in protestmany peasants had slaughtered their horsesand livestock and destroyed their equipment.These events ran counter to many of thechanges that had reduced transmission inmuch of Europe. In neighboring Poland andFinland farming was also less advanced thanin much of the rest of northern Europe, butthe slow modernization that occurred overthis period probably contributed to the steadydownward trend in malaria cases.
Malaria in southern Europe. Malariaremained highly prevalent in much ofMediterranean Europe, the Balkans, and thecountries bordering the Black Sea until afterWorld War II (89). The presence of severaleffective vector species, an abundance of pro-lific mosquito breeding sites, the warm cli-mate, and the long summer season were allhighly conducive to transmission. In addition,
much of the region was relatively unaffectedby the environmental changes associated withmodern agriculture. Part of this lack of changecan be attributed to the disease itself, forpoverty and lack of progress characterized
many of the highly malarious regions. Innorthern Italy, for example, much ofPiedmont and Lombardy was free of transmis-sion. By contrast, large portions of the rest ofthe country, particularly Sardinia, Calabria,
Environmental Health Perspectives • VOLUME 109 | SUPPLEMENT 1 | March 2001 147
Figure 2. (A). Distribution of malaria in Sweden in the late nineteenth century, showing endemic and epidemic areas. (B) The annual death rate (1860–1940) attributed to the dis-ease. Modified from Ekblom (81) and Renkonen (82). The 15°C July isotherm was the approximate northern limit of transmission. Note the location of Archangel, Russia, site of amajor epidemic of P. falciparum in the 1920s.
A
Arctic Circle
Arctic Circle
9,000
8,000
7,000
6,000
5,000
4,000
3,000
2,000
1,000
01860 1870 1880 1890 1900 1910 1920 1930 1940
YearCa
ses
B
Figure 3. Malaria in Poland, 1946–1955. Malaria persisted in Finland, Poland, and the Soviet Union long after it haddisappeared from other parts of northern Europe. Derived from Dymowska (90).
2,000
1,000
500
100
50
10
5
1
Num
ber o
f inf
ectio
ns
1946 1947 1948 1949 1950 1951 1952 1953 1954 1955
Year
109S1.Part 2 02/28/01 1:32 PM Page 147 (Black plate)
A
Arctic Circle
Arctic Circle

Reiter
148 VOLUME 109 | SUPPLEMENT 1 | March 2001 • Environmental Health Perspectives
and Sicily, remained virtually uncultivateduntil the 1950s, at least partly because of theravages of the disease. The same was true formajor regions in Spain, Greece, Romania, andBulgaria (34).
In World War I, British and French alliedoperations in Macedonia were paralyzed by anepidemic of malaria (34). By 1918, incidenceamong British troops had risen to 459 casesper 1,000. Between the world wars, majormalaria epidemics caused enormous economichardship in the same region. Rail transportwas often interrupted in the summer, and har-vests were badly affected by the shortage oflabor. Serbia, Bosnia-Herzegovina, Kosovo,and Croatia were also seriously affected.P. vivax was the principal parasite species inspring, followed by P. falciparum andP. malariae in late summer and early fall (91).
The elimination of malaria from Europe.Until end of World War II, the only effectiveapproach to mosquito control was to elimi-nate the breeding sites by environmentalmodifications such as drainage and landfilland by the application of insecticidal oils orchemicals. These methods were costly, sothey were applied mainly to urban centersand other areas of high economic importance.The advent of DDT revolutionized malariacontrol (92). It enabled cheap, safe, effectivetreatments to be targeted at the site wheremost infections occur—in the home.
The principal treatment method was toapply 2 g/m2 of DDT to indoor surfaces onceevery 6 months. Mosquitoes were killed bycontact when they alighted on the treatedsurfaces. Initial efforts in Italy, Cyprus, andGreece were so successful that a decision wasmade to eradicate malaria from all of Europe(33,93). The campaign was based on a carefulapplication of scientific principles, meticulousplanning, efficient administration, generousfinancing, and continuous emphasis on evalu-ation. It was orchestrated by several interna-tional agencies, particularly the World HealthOrganization (WHO) and the UnitedNations Children’s Fund, as well as numer-ous national bodies including the U.S. PublicHealth Service. The International HealthDivision of the Rockefeller Foundation alsoprovided generous financial and technicalsupport. By 1961, eradication had alreadybeen achieved in many countries. The entirecontinent was finally declared free of endemicmalaria in 1975 (94). One of the lastcountries affected was The Netherlands.
The complexity of malaria transmission:The Netherlands.
Everything about malaria is so moulded andaltered by local conditions that it becomes athousand different diseases and epidemiologi-cal puzzles. Like chess, it is played with a fewpieces, but is capable of an infinite variety ofsituations (89).
The persistence of malaria in TheNetherlands, a country that has held a centralposition in the economic life of WesternEurope since the Middle Ages, is a good illus-tration of this complexity.
In the nineteenth century, despite greatprogress in drainage and cultivation, theheavily populated Low Countries were themost malarious region of northern Europe.During the Napoleonic Wars the failure ofthe British Walcheren expedition (1809) wasblamed on malaria after 4,000 troops died offever. Severe outbreaks also attributed to thedisease occurred in 1826, 1834, and 1846,with many thousands of deaths (95).
The dominant vector, Anophelesmaculipennis, was present throughout thecountry. However, the disease was particu-larly prevalent in areas that had beenreclaimed from the sea, especially in NoordHolland, a province that includes the cities ofHaarlem and Amsterdam. Moreover, a majorpeak of incidence occurred in the springrather than summer or fall.
The unraveling of this puzzle was one ofthe classic triumphs of medical entomology(89). Investigation revealed that An. maculi-pennis was not one but several sibling species(96): An. atroparvus, which lays its eggs inbrackish water; An. messeae, which prefersfresh water; and An. maculipennis, sensustrictu, which occurred inland in nonmalari-ous areas. In the laboratory, An. messeae andAn. atroparvus were both excellent vectors ofmalaria. In the field the picture was very dif-ferent. Both species preferred to feed ondomestic animals rather than humans, butwhereas An. atroparvus rested in stables,An. messeae preferred uninhabited sheds andother unheated outhouses. In the fall,An. messeae built up a fat body that allowed itto hibernate. By contrast, An. atroparvusremained semiactive, feeding at regular inter-vals throughout the winter. Although most ofits meals were taken on farm animals, it occa-sionally wandered into adjacent humandwellings. If persons infected with the malariapathogen were present, the mosquitoacquired the infection. Thus, as in manytropical countries, transmission occurred atall times of the year despite winter tempera-tures that could dip below –20°C. However,the mosquito’s ovaries did not develop eggsuntil the advent of spring, a condition knownas gonotrophic dissociation. An additionaltwist to the story was that the local strain ofP. vivax had a particularly long incubationperiod, so persons infected in the fall andwinter showed symptoms of illness only inthe spring after the first new brood ofmosquitoes had emerged (34,89).
In 1932 a dyke was built to enclose theZuider Zee, a vast area of brackish water tothe east of Noord Holland. The accumulation
of river water behind this dyke caused agradual decrease in the salinity of the sur-rounding land, greatly reducing the larvalhabitat of An. atroparvus (97). In addition,there were major changes in the living condi-tions of humans. New farmhouses were lessintimately associated with cattle sheds and sta-bles, and their structure and heating technol-ogy made them much less hospitable tooverwintering mosquitoes (34).
Thus, although the disease was finallyeradicated by routine DDT applications andthe administration of antimalarial drugs, theecology, physiology, behavior, and survival ofthe vector, plus the interaction of thepathogen with both vector and host, allcontributed to its demise.
Malaria in North America. Severalefficient malaria vectors are indigenous toNorth America and are common in manyareas. The pathogen probably arrived in thehemisphere in infected European colonistsand African slaves. The history of its declineis similar to that of Europe. In the 1880s, thedisease was widespread in nearly all states eastof the Rocky Mountains (Figure 4), from thesemitropical Gulf Coast states to the northernU.S. border (98) and into Canada (99,100).It was also present west of the RockyMountains, particularly in areas whererainfall is abundant.
As living conditions improved andantimalarial drugs became more widely avail-able, the incidence of the disease declined.During World War II, the U.S. military cre-ated an Office of Malaria Control in WarAreas, which was responsible for militarybases throughout the country. In a sympo-sium published in 1941 (101), the diseasewas described as “of moderate endemic inten-sity in the sixteen southeastern states . . . withscattered epidemics at intervals of approxi-mately seven years.” In 1946 the U.S.Congress established a new agency, theCommunicable Disease Center. This was theforerunner of the U.S. Centers for DiseaseControl and Prevention (CDC), and its prin-cipal mission was to eradicate malaria fromthe entire country. It was based in Atlanta,Georgia, because the southern states were themain region still affected by the disease.When operations began, foci of transmissionwere already hard to find, but the disease wasnot totally eliminated until the late 1950s.Today, as in Europe, there are many parts ofthe country where anopheline vectors areabundant, but the transmission cycles havebeen disrupted and the pathogens are absent.
Malaria in the TropicsEpidemiology. The epidemiology of malariain the tropics is even more complex than thatin temperate climates and varies greatly withlocation (33). In much of equatorial Africa,
109S1.Part 2 02/21/01 3:34 PM Page 148 (Black plate)

Climate change and mosquito-borne disease
parts of northern India, Indonesia, and SouthAmerica transmission is termed stable becauseit is fairly constant from year to year. The dis-ease is endemic, but epidemics are uncom-mon. In other regions, including much ofIndia, Southeast Asia, and Central and SouthAmerica, the disease is also endemic but istermed unstable because transmission canvary greatly from year to year, and the poten-tial for epidemics is high.
Stable endemic malaria. This occurs inregions where the anophelines are anthro-pophilic (they prefer to feed on humans) andhave a high survival rate. Temperature andhumidity are generally high and there is oftenrelatively little seasonal variation. The diseaseis hard to control because transmission isextremely efficient—it can occur with rela-tively low mosquito populations—and trans-mission rates are so high that most peopleexperience many infective bites per year.Severe illness and fatality are generallyrestricted to “new arrivals,” i.e., children andnonimmune immigrants. Older inhabitantsharbor the parasites but have survived multi-ple infections and maintain a high degree ofimmunity by repeated reinfection. Epidemicsin the local population are rare but can some-times occur after a major increase inmosquito populations.
Unstable endemic malaria. This occurs inregions where the anophelines are generallyzoophilic (they tend to bite animals in prefer-ence to humans), where their survival rates arelow, or where both apply. Transmission ratescan vary greatly, with epidemics often sepa-rated by many years of relatively low activity.The factors that precipitate such epidemics are
often difficult to identify. The disease mayappear suddenly for no apparent reason, onlyto disappear again without obvious cause. Ingeneral, transmission is relatively inefficientand therefore requires high mosquito popula-tions. In theory this implies that the disease iseasier to control.
Index of stability. The causes of stabilityversus instability have been the subject ofmuch study and controversy (33). In reality,the terms are merely convenient labels, forthey describe the extremes of a wide range ofsituations. Mathematically the index of stabil-ity is given by the expression
where a is the preference of the principalvector for human versus nonhuman bloodand p is its daily survival probability. Notethat a) the denominator is an exponentialfunction—small changes in survival rate havea major impact on the index; and b) neitherof the two variables is directly affected byclimate variables
The concept of stability has proven valu-able in understanding and solving controlproblems. The same approach is useful inaddressing the question of climate change.
Climatic influences on malaria trans-mission in the tropics. Climatic factors areimportant in the tropics. Their influence iscomplex and varies according to region andthe ecology of the vectors concerned.
Temperature. In theory, high temperaturesshould increase the likelihood of transmissionbecause they reduce the extrinsic incubation
period. However, activities such as biting andegg laying are also likely to be accelerated.These are high-risk activities, so survival rate,and thus transmission rate, may also beaffected.
In equatorial regions, vectors such asAn. gambiae are commonly found as high as3,000 m above sea level, but endemic malariadisappears above 1,800–2,000 m (102,103).A limiting factor is presumably the tempera-ture required for effective extrinsic incuba-tion, analogous to the temperature limitsdefined by isotherms at high latitudes.
Humidity. Survival rate may also bereduced when hot weather is accompanied bylow humidity, but in areas where such condi-tions are normal, local species have adapted tocope with them. For example, in the severedrought and extreme heat of the dry season insemiarid parts of the Sudan, female An. gam-biae survive up to 11 months of the year byresting in dwelling huts and other shelteredplaces. Blood feeding continues, so transmis-sion is not interrupted, but the ovaries do notbegin to develop eggs until the rains return(42,104). This strategy of gonotrophic disso-ciation is remarkably similar to the wintersurvival mechanism of An. atroparvus inThe Netherlands. In both cases, inactivityleads to a high vector survival rate and con-tinued transmission of malaria, even underadverse climatic conditions.
Rainfall. Rainfall can promote transmissionby creating ground pools and other breedingsites, but heavy rains can have a flushing effect,cleansing such sites of their mosquitoes.Drought may eliminate standing water butcause flowing water to stagnate. Thus in aridareas prolonged drought may cause malaria todecline (105), whereas in areas where rainfall isnormally abundant, vast numbers of mosqui-toes can be produced and “drought malaria”may follow. Similar patterns apply to artificialstreams in irrigated regions and storm drainsand sewers in urban areas. Drought may alsostimulate people to store water in cisterns,drums, and other manmade containers thatserve as breeding sites.
Seasonality. Climatic factors often imposeseasonality on transmission. Even in regionsof high stability, transmission may peak at aspecific time of year, although such modula-tions have little impact on the long-termhealth of the population because overallinfection rates are so high. However, indicesof stability are often lower in regions whereseasonality is more pronounced. In somecases, the phenomenon is clearly defined butmay vary according to circumstance.
The complexity of the influence of climateon malaria transmission in the tropics is wellillustrated by the history of epidemics inSri Lanka (106). In the 1930s, the disease wascommon. In a normal year 1.5 million people,
ape− log
Environmental Health Perspectives • VOLUME 109 | SUPPLEMENT 1 | March 2001 149
Figure 4. Malaria in the United States and Canada in 1882.The dark shaded areas are regions where the diseasewas probably endemic. Modified from Faust (98), Stewart (100), and Williams (188).
109S1.Part 2 02/21/01 3:34 PM Page 149 (Black plate)

Reiter
150 VOLUME 109 | SUPPLEMENT 1 | March 2001 • Environmental Health Perspectives
about a quarter of the total population, weretreated for malaria in hospitals and dispensaries.However, from 1934 to 1935 a catastrophicepidemic is estimated to have killed 100,000people. Worst hit was the southwestern quad-rant of the country, a region with an averageannual rainfall of more than 250 cm.
The dominant vector in that part of thecountry is An. culicifacies, a species that breedsalong the banks of rivers. In normal years itwas not abundant. Malaria was endemic, butthe stability index was low; in most years thedisease was relatively unimportant.
The monsoons in the preceding 5 yearshad been exceptionally favorable, with abun-dant heavy rainfall leading to excellent ricecrops. Under such conditions, river flow washigh, An. culicifacies was rare, and the popula-tion was exceptionally healthy. However, whentwo successive monsoons failed, the rice cropswere lost and hunger was widespread. Colossalnumbers of An. culicifacies were produced inthe drying rivers and irrigation ditches.
The epidemic that followed was exacer-bated by the weakened condition of the peo-ple. In addition, the herd immunity wasespecially low because the previous 5 yearshad been wet and therefore relatively free ofmalaria. By contrast, in the drier parts of theisland, where An. culicifacies was dominantbut the stability index was higher, immunityprotected the population from the worstravages of the epidemic (107).
Malaria was almost eradicated from all ofSri Lanka in the 1960s, but in recent years ithas returned as a major public health problem,with almost 0.3 million infections reportedannually in its population of 17 million (108).
Nonclimatic influences on malaria trans-mission in the tropics. As in temperateregions, human activities and socioeconomicfactors have a major impact on transmission.
Population. The world’s population was4 billion in 1960, 5 billion in 1987, andmore than 6 billion in the year 2000. Ninety-six percent of this increase has occurred indeveloping regions of Africa, Asia, and LatinAmerica, many of which are malarious (109).For example, in sub-Saharan Africa, there arenow nearly four times as many people(approximately 450 million) as there were in1955. The population of Kenya was 8 millionin 1963, 24 million in 1990, and is expectedto be double this figure within the next 10years. Many of the other factors listed in thissection can be attributed to or associated withthis rapid population increase.
Forest clearance. Many important malariavectors breed in open sunlit pools. Forestclearance provides abundant new habitat forthese species and is a classic cause of theemergence of malaria problems (110).Massive clearance also modifies local micro-climates by reducing shade, altering rainfall
patterns, augmenting air movement, andchanging the humidity regime.
Agriculture. Rice and other irrigated cropsoften create an ideal habitat for the mass pro-duction of mosquitoes. The construction ofdams for irrigation or hydroelectric power canalso lead to high populations of anophelines.Abandoned fish farming projects have hadsimilar results in several countries. Cattlehoofprints can provide a prolific breeding sitefor anthropophilic species such as An. gam-biae. Conversely, as in temperate regions, theproximity of cattle to human habitation candeflect bites from humans in areas where themosquitoes are predominantly zoophilic.
Other ecologic factors. The removal ofreeds from river margins increases water flowand eliminates the shelter preferred bymany—but not all—vector species. Similarly,in still water, floating plants such as duck-weed and water lilies are favored by somespecies but discourage others.
African papyrus produces an oil thatinhibits anopheline development. The elimi-nation of this aquatic plant for rice cultiva-tion provides an environment better suited toAn. gambiae and An. funestus. Conversely, thecultivation of ground depressions in the Sahelcan suppress An. funestus and thereby reducetransmission (105).
In the High Andes of South America,malaria can be transmitted at 2,700 m. At thisaltitude, the vector An. pseudopunctipennisbreeds in volcanic thermal springs (111). Theoutdoor climate is too cold for Plasmodium todevelop, but the adult mosquitoes restindoors, where temperatures are severaldegrees higher. Indoor resting sites also enableAn. gambiae to transmit malaria in the EastAfrican Highlands.
Movement of people. Infected peoplemoving in pursuit of work can introducemalaria to areas where it is absent or relativelyrare. Conversely, nonimmune people are athigh risk if they move to an area of transmis-sion. In recent years, extensive road buildingand modern transportation have greatly exac-erbated this factor. Even in the poorest coun-tries, it is now common for people to travellarge distances by road or rail to work, visitrelatives, or seek medical attention. Tens ofthousands of infected people also arrive by airin nonmalarious temperate countries eachyear, and many people from those countriesbecome infected when they travel to malari-ous regions for work or leisure.
Urbanization. In rapidly expanding urbanareas, extensive water storage and inadequatewater disposal can lead to disastrously highmosquito populations. The absence of cattlecan promote stable transmission by forcingzoophilic species to feed on people.
Many tropical cities are surrounded bylarge satellite settlements that retain rural
characteristics. Their dense populationspromote conditions that are ideal for transmis-sion. Infection rates in these semirural habitatsare often higher than in the cities themselves.
Resistance to insecticides. There is anatural variability in the susceptibility ofinsects to insecticides. Insecticidal controlexerts a selection pressure on the target popu-lation, promoting the survival of the least sus-ceptible individuals. These survivorseventually give rise to strains that are physio-logically resistant even to high dosages.Various biochemical and physiologic processescan be involved. In many cases resistance toone insecticide confers cross-resistance tochemically similar compounds and even tofamilies of compounds that are chemicallyunrelated. For example, knockdown resistance(kdr) to DDT may also confer kdr to thepyrethroids, a family of synthetic insecticidesrelated to natural compounds derived fromplants of the genus Chrysanthemum.
Control operations can also select forbehavioral resistance, i.e., changes in behaviorthat reduce contact with the insecticide. Forexample, species that prefer to feed and restindoors (endophilic) can switch to outdoor(exophilic) activity in response to indoorresidual treatments with DDT. Both types ofresistance have proven a major obstacle tomalaria control (33,112).
Resistance to antimalarial drugs. A rangeof drugs can be used to prevent malaria infec-tion (malaria chemoprophylaxis) or to cureinfections (malaria chemotherapy). The sameselection process that leads to insecticide resis-tance in the vector can foster the emergenceof strains of drug-resistant Plasmodium. Inrecent years, malaria control activities havefrequently placed more reliance on antimalar-ial drugs than on insecticidal strategies.Resistance to several of these drugs is nowwidespread and is a significant factor in theworldwide resurgence of malaria (33,112).
Degradation of the health infrastructure.The public health sector of many countrieshas been eroded by lack of funding, institu-tional difficulties, rapid urbanization, andother problems associated with rapid develop-ment. In much of Africa and parts ofSoutheast Asia the increasing prevalence ofAIDS has overwhelmed the ability of authori-ties to deal with other diseases. In such cir-cumstances, people often rely on ineffectiveself-administration of antimalarial drugs.
War, civil strife, and natural disaster.Mass movements of people, e.g., soldiers andrefugees, often promote malaria transmission.The breakdown of public health services,damage to water distribution and drainagesystems, and the destruction of homes oftenexacerbate the situation. High concentrationsof people in camps for displaced persons canalso be disastrous.
109S1.Part 2 02/21/01 3:34 PM Page 150 (Black plate)

Climate change and mosquito-borne disease
The recent recrudescence of malaria. Ithas been suggested—on the basis of the sim-ple models mentioned earlier—that the inci-dence of malaria in the tropics could increaseby up to 60% as a result of global warming(12,13,113). It has even been claimed thatthe recent recrudescence of the disease may becaused partly by warming that has alreadyoccurred (13,28,114). In the context of suchdiscussion, it is important to review themultiple factors that have shaped the recenthistory of the disease.
The malaria eradication program. Theadvent of DDT after World War II enabledWHO to encourage many newly indepen-dent countries to attempt malaria control.Results were so promising that a worldwidecampaign to eradicate the disease was for-mally endorsed by the Eighth World HealthAssembly in 1955.
Initial results were spectacular: Thirty-sixparticipant countries had been certified as freeof endemic transmission by 1977. Eighty-three percent of the world’s population wasliving in areas from which the disease hadbeen eradicated or where control activitieswere in progress. However, despite thisencouraging success, the momentum of theprogram was already faltering (92), and thedisease began a rapid recrudescence (115).Today malaria has returned to nearly all itsformer territory. Its prevalence continues toincrease, and the goal of eradication has hadto be abandoned. Prevalence is again high inmany parts of Central America, the northernhalf of South America, much of tropical andsubtropical Asia, some Mediterranean coun-tries, and several of the republics that wereonce part of the Soviet Union. Annual casesworldwide are roughly estimated at 250 mil-lion, with 2 million deaths. As before, a largeportion of these deaths are in areas of highstability, particularly in sub-Saharan Africa,where large-scale control was never seriouslyattempted (112).
The failure of the eradication program isoften attributed to the emergence of resistanceto insecticides and antimalarial drugs and theinability of many governments to sustain con-trol operations in the face of competing priori-ties. However, in nearly every country,ecologic changes wrought by rapid develop-ment, population increase, civil strife, and warhave also been highly significant. These arethe same factors that enabled malaria toexpand and flourish in Europe from the timeof Homer until the nineteenth century.
Highland malaria. In equatorial regions,vectors such as An. gambiae are commonlyfound as high as 3,000 m above sea level, butendemic malaria disappears above 1,800–2,000 m. A limiting factor is presumably thetemperature required for effective extrinsicincubation.
In recent decades, outbreaks of malariahave been reported from many such mountainregions. For example:• Serious outbreaks of malaria have recently
been reported in the New GuineaHighlands. Until the 1950s, the diseasehad rarely been reported in the region.
• A devastating malaria epidemic occurredon the central plateau of the MadagascarHighlands from 1986 to 1988.
• Malaria has become a serious public healthproblem in the West Kenya Highlandsover the past 20 years. In previousdecades, transmission was relatively low.
• The Amani hills of northeastern Tanzaniawere once virtually malaria-free, butprevalence has risen to very high levels inthe past three decades.
• In the highlands of southeastern Uganda,records at a well-staffed mission hospitalshow a 70-fold increase in malaria inci-dence from the late 1960s to the mid-1990s, with a significant epidemic in 1994.
• In southern Rwanda, malaria records atanother well-staffed facility show a slowincrease in incidence from 1983 onward,with a sudden jump in 1987 to more thantwice the previous rate.
Several authors have suggested that climatechange may have contributed to these out-breaks (13,28,116,117). It is thereforeinstructive to review them in some detail.
New Guinea Highlands. In the early1930s a human population estimated at 1million, previously unknown to outsiders,was contacted in the mountains of NewGuinea. It appeared that these so-called StoneAge people were malaria free, and this wasattributed to their unique state of isolation(118). By contrast, the lowland coastalregions were highly malarious.
At first, the highlanders became a newsource of labor for the coastal plantations,but after World War II, there was a rapidincrease in the number of small landholdersgrowing Arabica coffee and other crops inthe mountains. In the late 1940s governmentscientists warned that the increasing contactbetween the regions could bring disaster, forepidemic malaria had already appeared inseveral highland areas at around 1,500 m(119). By the mid-1950s several alarmingoutbreaks led to the enforcement of a lawthat required employers of highlanders work-ing on the coast to supply them with anti-malarials and to ensure that the medicationswere actually taken. Highlanders were heldby the government in compulsory quarantinefor 2 weeks and given curative malariatherapy on repatriation (120,121).
These regulations failed to stop theemergence of the disease and its spread tomany isolated valleys. The increase in preva-lence was clearly attributable to a rapid
increase of anopheline populations after forestclearance and to the construction of roads,airstrips, plantations, mines, water impound-ments, and other human artifacts (122).Several vectors were involved, includingAn. farauti and An. punctulatus (122). Thesespecies breed in open sunlit pools and are inmany ways analogous to An. gambiae andAn. funestus in sub-Saharan Africa.
There is some evidence that malaria mayhave been well established in the highlandsbefore the arrival of Europeans, but it didnot become evident until forest clearanceand development were more widespread(119,123). Whatever the chronology, therecent history of the disease is clearly attribut-able to the introduction of the parasite tononimmune populations and the proliferationof its vectors as a result of large-scale ecologicchange.
Madagascar Highlands. Irrigated riceculture was introduced to the plateau regions ofMadagascar around 1800, and the populationexpanded rapidly. Malaria appears to have beenabsent for much of the century, but in 1878 acatastrophic epidemic occurred around the cap-ital, Tananarive (present name: Antananarivo;altitude approximately 1,200 m). A secondmajor epidemic occurred in 1895. Thereafterthe disease became endemic. As in most of sub-Saharan Africa, An. gambiae and An. funestuswere the principal vectors (124). In 1949 theFrench administration initiated a major controlcampaign involving annual treatment of homeswith DDT, and an extensive network ofmalaria clinics and dispensaries. Within 10years the disease had all but disappeared, butthe clinics remained operational to treat spo-radic outbreaks.
The country became independent in 1960and later realigned itself with the SovietUnion. After a military takeover in 1975 andthe enforcement of a new plan of scientificsocialism, the economy went into disastrousdecline. Medication was in short supply and in1980 all but one of the clinics were closed. Theferocity of the transmission that followed wasfueled by the population’s lack of immunity,the absence of treatment centers, and the short-age of medication. In addition, as a result of thebreakdown of law and order, people begankeeping their cattle inside their houses at nightto guard against theft, thereby bringing them-selves in greater contact with the primarilyzoophilic vector, An. arabiensis. During theepidemic, there were press reports of100,000–400,000 deaths, although investiga-tors concluded that the true figure was proba-bly 10,000–25,000 (103,124). The situationwas corrected in 1993–1995 by reinstitutingDDT treatments and reopening the dispen-saries. Despite several claims that transmissionwas the result of unusually warm temperatures,the meteorologic record shows remarkable
Environmental Health Perspectives • VOLUME 109 | SUPPLEMENT 1 | March 2001 151
109S1.Part 2 02/21/01 3:34 PM Page 151 (Black plate)

Reiter
152 VOLUME 109 | SUPPLEMENT 1 | March 2001 • Environmental Health Perspectives
stability of temperatures over the past 30 years(124). All the evidence indicates that thetragedy was brought about by the breakdownof successful control in a highly endemic areaas a result of socioeconomic factors (103).
Kenya Highlands. Malaria was notrecorded in the Kenya Highlands until thesecond decade of the twentieth century(125,126). It began to appear after the clear-ance of forests for the development of teaestates and the importation of infected labor-ers. Initially the disease was seen mainlybelow 1,500 m and was rare in Nairobi(1,680 m), the capital. The first sizable epi-demic (1918–1919) was attributed to thereturn of local soldiers from Tanzania.Thereafter, transmission moved to progres-sively higher altitudes, with major transmis-sion in Nairobi and in the farming districtaround Eldoret (2,040 m). Six epidemicswere recorded between the two World Wars.During World War II, epidemics occurred inthe Londiani district (2,250–2,490 m) andeven at a farm near Mount Timboroa, at2,490–2,550 m (127). Transmission couldoccur at these altitudes because the adultmosquitoes rest indoors, where temperaturesare 3–5°C higher than outside. In the 1940s,British entomologists referred to this as “hutmalaria” and even suggested that the diseasecould be eliminated by persuading thewomen to move their cooking fires outdoorsto make their homes colder!
The fundamental cause of this progressiveupward advance of malaria was widespreaddeforestation and development associatedwith large farming ventures (127). The con-struction of roads and railways generatedinnumerable flooded “borrow pits,” depres-sions left by excavation for materials, and alsocontributed to the dispersal of the mosquito.The introduction of the ox wagon caused aproliferation of rough cart roads; water in thewheel ruts provided a prolific breeding site forvectors. Milldams on rivers interfered withnatural drainage. These and many other fac-tors were components of a drastic ecologicchange, and this change brought transmissionof malaria to the highlands. The disease con-tinued to be a serious public health problemuntil the 1950s, when the colonial govern-ment organized an extensive control program,after which the area was essentially malariafree until the 1970s.
The tea-growing estates (1,780–2,225 m)in the Kericho district have an extensive med-ical service for employees and their depen-dents that was initiated in 1925. Health careat the central hospital of Brookebond KenyaLtd. is extended to some 100,000 inhabitantsof the region. However, there is no attempt atmosquito control, and malaria has reemergedas a serious problem. A recent study showedepidemics in almost every year from 1990 to
1997, with a mean annual attack rates ofapproximately 50% (128). Peak transmissionwas from May to July, after the principalrainy season and before mean monthly tem-peratures drop below 18°C. A questionnairesurvey (June 1997) indicated that only 8% ofpatients had traveled to areas with knownmalaria transmission in the previous 30 days.
It has been suggested that the main factorin this recrudescence may be increased resis-tance to antimalarial drugs, as well as theunsupervised use of ineffective medications,but the picture is not entirely clear (129).Whatever the cause, the history of multipleepidemics in the earlier part of the century,including many at higher altitudes, makes itunnecessary to infer climate change as a con-tributory factor. Moreover, a set of well-maintained meteorologic records shows nosignificant change in temperature over recentdecades (103,128), and a retrospective studyof 33 years of monthly incidence data fromthe Brookebond tea-growing estates did notreveal any relationship between climate vari-ables and the timing or duration of inter-epidemic periods (130). Indeed, in a detailedreport to WHO (131), a group of malariaspecialists based in Nairobi dismissed thosewho claim a global warming link as “scientificNostradamuses.”
Amani Hills, Tanzania. In 1902, theGerman colonial administration built a fieldstation at 1,000 m in the Amani Hills (alti-tude 600–1,000 m) of the eastern UsambaraMountains in northeastern Tanzania. Theheavily forested area had a relatively cool cli-mate and was considered malaria free. Formany years Europeans living on the coastwould spend time there during the hottermonths of the rainy season.
Logging and extensive clearance foragriculture and general settlement began inthe 1960s. Malaria specialists (132) predictedthat this deforestation, along with road con-struction and the proliferation of dams,ponds, ditches, furrows, road ruts, and otherbreeding sites, would be highly favorable toAn. gambiae and An. funestus; both specieshad been scarce or absent because they preferopen sunlit breeding sites. From 1967 to1978, the human population doubled,mainly through an influx of people from thehighly malarious coastal region. At the sametime, malaria became increasingly common.By 1980–1982, 61% of infants less than 1year of age who attended a clinic were posi-tive for malaria; most of the infants (91%)had not left the area in the weeks precedingtheir illness (133).
As in the Kericho tea estates, the Amanistation maintained a good meteorologicrecord. In the first half of the century, meanannual temperatures ranged from 12.8° to15.6°C. In the 1960s there was a sudden
warming directly attributable to theelimination of shade by deforestation. Afterthis warming the recorded temperatures areremarkably consistent for about 10 years,ranging from 20.0° to 20.8°C until refor-estation produced a steady cooling trend ofabout 0.46°C per year. By 1975 the meanannual average was back to 17.4°C, but themalaria incidence remained high, appar-ently because indoor temperatures were sev-eral degrees higher than those recorded atthe field station. In a recent study meanannual prevalence of parasitemia amongchildren 6–71 months of age ranged from33 to 76% (134).
The factors leading to the emergence ofmalaria in Amani were similar to those inNew Guinea and Kenya: A major ecologicdisturbance—deforestation—created a newhabitat for effective vectors, and the elimina-tion of shade produced a marked change inlocal climate. In addition, the prevalence ofthe parasite was augmented by an influx ofinfected people. More recently, substantialresistance to chloroquine has been docu-mented in all three areas, although the drug isstill widely used by the local populations.
Uganda Highlands. Over the past 30years a marked emergence of malaria has beennoted in the Runkungiri and Kabale districtsof southeast Uganda. Incidence of the diseasehas steadily increased among outpatients atKisizi Mission Hospital (altitude 1,650 m),and a significant epidemic occurred in 1994.Peak transmission is generally in July, thedriest, coldest month of the year (103).
Endemic foci of the disease at 1,800–2,000 m were recognized in the area as earlyas 1919, and epidemic transmission has beenknown since 1948 (135). When a malariaeradication program was established in 1961,extensive surveys showed that prevalence wasvery low at altitudes above 1,500 m, except atcertain lakesides up to 1,900 m. As in Kenya,Tanzania, and neighboring countries, trans-mission could occur at these higher altitudesbecause the adult mosquitoes rest indoors,where temperatures were 3–5°C higher thanoutdoors (136). Indoor spraying continuedfrom 1963 to 1966.
In maps drawn before 1961, the humanpopulation was distributed mainly on hill-sides above 1,500 m. The valley bottomswere mostly papyrus marsh. Since then, pop-ulation growth has been rapid (more than 3%per year). Many inhabitants have moved offthe hills to cultivate the marshland with foodcrops and fish ponds. The elimination ofpapyrus has created a habitat for An. gambiaeand An. funestus, leading to increased trans-mission (137). In 1994, rainfall was morethan twice normal, so the vector was probablyespecially abundant. Thus, the increasingprevalence of the disease is attributable to the
109S1.Part 2 02/21/01 3:35 PM Page 152 (Black plate)

Climate change and mosquito-borne disease
changes in agriculture and ecology thataccompanied a massive increase in populationin an area of unstable endemic transmission(103), not to any change in climate.
Rwanda Highlands. Rwanda lies close tothe equator, adjacent to Uganda. Most of thecountry is at an elevation above 1,500 m.Population density is the highest in Africa,although more than 90% of the populationis rural. Traditional settlement is not in vil-lages but in homesteads scattered across thehillsides of the mountainous terrain, withseveral hundred persons per square mile. Inthe past few decades, population increase hasbeen extremely high—more than half theinhabitants are less than 16 years old—and isfar above the productive capacity of the envi-ronment. In 1972 this situation was exacer-bated by an influx of 60,000 refugees fromneighboring Burundi.
Data from around the country indicatethat the incidence of malaria has risen steadilyin recent decades. The reasons for this increasehave not been investigated but are probablysimilar to those in neighboring countries. Forexample, outbreaks in the 1980s were attrib-uted to a new road that facilitated travel tomalarious lowland areas (138). Also, as inUganda, population pressures have led toincreased use of valley bottoms, where settle-ments are more concentrated and trans-mission is more likely. Indeed, an associationbetween marsh clearing and an increase inmalaria was noted around Butare as early as1946 (139). In neighboring Burundi, a coun-try with similar topography and culture, anassociation between rice cultivation andmalaria in the Rusizi Valley was noted in the1980s (140).
The Gikonko Health Center in southernRwanda is a well-staffed facility that serves apopulation of 38,000 people who live at amean altitude of 1,500 m. Malaria recordsfor 1983–1990 show a slow increase in inci-dence from 1984 to 1986 and a suddenjump in 1987 that appeared to be especiallyhigh among people living at higher elevations(a 501% increase in incidence in the secondhalf of the year) (114). Temperatures in thatyear were approximately 0.9°C warmer thanthe mean for 1961–1980 and rainfall wasabout double. Loevinsohn (114) suggestedthat this jump was an example of the “sub-stantial epidemiologic effect” of the impactof climatic factors on malaria “near or astridethe altitude and latitude limits” of its trans-mission. He supported this claim with aregression model that showed an autocorrela-tion coefficient (incidence vs. temperatureand rainfall) of 0.89 (p < 0.001). He ruledout the likelihood that other factors—cessation of control measures, chloroquineresistance, population movement, road con-struction, rice cultivation, and the timing of
a famine—had contributed to the suddenchange. His paper is perhaps the most widelyquoted example of a putative link betweenclimate change and transmission.
At first glance, Loevinsohn’s conclusionsappear persuasive. The Gikonko data areconsidered unusually reliable and the coinci-dence between the sudden jump in incidenceand the warm, wet weather of 1987 is strik-ing. However, if the area were truly at thelimit of transmission, we would expect inci-dence to have declined in 1988 and 1989,when temperatures returned to normal andto have increased again in 1990, when it wasnearly 0.5°C warmer. In fact, the raised levelof incidence remained stable from 1987onward. However, a factor that has escapedmention in the climate change debate is thatWHO supervised a complete reorganizationof the national disease surveillance system atthe very time that the malaria “increase”began. The 2.5-year project involved a redef-inition of the list of reportable diseases, anongoing in vivo study system designed toimprove reporting of confirmed malaria,follow-up supervision to check surveillancedata, and a routine of regional feedback let-ters and graphs. It is highly likely, therefore,that the massive jump in incidence in 1987was the result of improved surveillance anddiagnosis rather than a consequence ofclimatic factors (141).
In summary, these examples underscorethe need to avoid simplistic reasoning inexplaining the appearance of “new” malaria athigh altitudes. In every case, transmission waswell below the local maximum altitudinallimit (Figure 5), and there is no evidence thatclimate change played any role in the increaseof incidence. As in South America (142), amajor factor was the cessation of house spray-ing with residual insecticides such as DDT.Most of the other significant variables areultimately attributable to the populationexplosion and an increased mobility of thepopulation.
Yellow Fever
The DiseaseYellow fever is a viral disease native to tropicalAfrica (143). In its original woodland habitatit circulates among wild monkeys. It is trans-mitted by at least 14 species of mosquito—allof the genus Aedes—that breed in tree holesand other natural cavities. Humans can beinfected if bitten by infected mosquitoes.Symptoms include high fever, jaundice, andspontaneous bleeding. The severity of illnessis quite variable, but 20–50% of jaundicecases are fatal.
The virus can also be transmitted betweenhumans by mosquitoes that are more closelyassociated with human habitation. One ofthese is the peridomestic form of Ae. aegypti, aspecies that readily breeds in water jars andother man-made containers. It is widespreadin villages and urban areas throughout thetropics and can also thrive in temperate cli-mates. In the United States its permanentrange extends to North Carolina (144),although breeding populations have beenrecorded as far north as New York, NewYork, and Boston, Massachusetts (145).
History of Yellow FeverAfrica. The endemic zone of yellow fever inAfrica extends from the southern borders ofthe Sahara to Angola. The disease occurs in awide variety of habitats and its epidemiologyis correspondingly complex. The broad pic-ture is of wandering epizootics that moveslowly across vast areas of the continent, gen-erating sporadic human cases in woodlandregions and sparking interhuman transmissionin heavily populated areas where peridomesticAedes species are common.
In the early twentieth century majorepidemics were associated with the rapidgrowth of townships, particularly in WestAfrica. From the 1940s onward, mass immu-nization in territories administered by France(Benin, Burkina Faso, Cameroon, Chad,
Environmental Health Perspectives • VOLUME 109 | SUPPLEMENT 1 | March 2001 153
Figure 5. Occurrence of malaria at various altitudes. Solid circles indicate the maximum recorded altitude of trans-mission from 1880 to 1945. Derived from data in Hackett (111). Open circles indicate recent outbreaks(103,114,122,128,129,133,134,136,137).
●
●●
●
●
●●
●
●
●
●●
●●
●●
●
Madagascar
Papau New GuineaArgentina
Chile
BoliviaKenyaEcuador
PeruUgandaKanyaRwanda
Tanzania
ColombiaEthiopia
Equator
ChinaUnited States
India
3,000
2,500
2,000
1,500
1,000
500
040 30 20 10 0 10 20 30
LatitudeNorth South
Altit
ude
(m)
109S1.Part 2 02/21/01 3:35 PM Page 153 (Black plate)

Reiter
154 VOLUME 109 | SUPPLEMENT 1 | March 2001 • Environmental Health Perspectives
Congo, Gabon, Ivory Coast, Guinea,Senegal, and Togo) greatly reduced the preva-lence of human disease (146), while epi-demics continued to occur in regions withoutimmunization programs. Despite this success,preventive immunization was discontinued inthe mid-1960s. During the past two decadesthe frequency of reported outbreaks hasincreased dramatically (143,147). To date,this increase has occurred mainly in ruralareas, but the risk of urban transmission ishigh. Yellow fever is now classified as animportant reemerging disease, with 34sub-Saharan countries at risk (148).
The Americas. Both Ae. aegypti and yellowfever were brought to the Western Hemispherefrom Africa with the slave trade. Transmissioncould continue at sea because the slave shipscarried breeding colonies of the mosquito intheir freshwater tanks (145). From the lateseventeenth century onward, urban epidemicsplagued the Caribbean and the tropicalAmericas. In addition, a forest cycle was initi-ated among New World primates, transmittedby several indigenous species of mosquito,particularly of the genus Haemagogus. Today,as in Africa, epizootics move across vast areasof the South American rain forest in cycles of5–9 years. These sometimes extend to forestedregions outside the enzootic zone. Thus, dur-ing 1948–1954 the disease moved via Panamaand the Atlantic Coast of Central America toMexico, and during 1964–1966 it spreadfrom southern Brazil into Missiones Province,Argentina (143).
Until the twentieth century, the viruswas frequently introduced to temperateregions. The first major outbreaks in theThirteen Colonies were recorded during theLittle Ice Age. New York was hit in 1668and Philadelphia, Pennsylvania, andCharleston, South Carolina, in 1690.Epidemics became more devastating asurban populations grew, causing widespreadterror and havoc. For example, 17,000 casesoccurred in Philadelphia, a city of 55,000persons in 1793 (149,150). Five thousanddied and a further 12,000 fled the city. Thedisease returned in 1797, 1798, 1799, 1802,1803, and 1805. Many other Atlantic coastcities were affected, including Norfolk,Virginia; Baltimore, Maryland; New York;and Boston (Table 2). Below the equator,major transmission was recorded as far southas Buenos Aires, Argentina.
Epidemics were even more frequent in thesouthern United States. New Orleans,Louisiana, was particularly vulnerable, andexperienced repeated outbreaks from 1793onward. In 1853, there were 40,000 cases with11,000 deaths. Seventy-five thousand peoplewere said to have fled the city. In 1878, thedisease swept northward along the Mississippiand Ohio Rivers, with an estimated 100,000
cases and 20,000 deaths. Worst hit wasMemphis, Tennessee, where at least 9,000died and many inhabitants fled, never toreturn (151). When the disease struck again in1879, there was serious national debate over asuggestion that the federal government issue anexecutive order to destroy Memphis and all itsproperty by fire. This was not carried out, butthe city, which had been the principal com-mercial and cultural center of the South, lostits charter and has never regained its formerimportance.
In 1881 Carlos Finlay proposed that thedisease was transmitted by Ae. aegypti. Hishypothesis was finally confirmed by WalterReed and his team in 1900 (152). Strict mos-quito control measures were applied, first inHavana, Cuba, and later in the United Statesand most of the major tropical ports. The lastoutbreak in the United States occurred inNew Orleans in 1905. The development ofan effective vaccine in the 1930s helped elim-inate the urban disease from the rest of theAmericas (143).
Europe. Ae. aegypti was present insouthern Europe and North Africa untilshortly after World War II. Its disappear-ance is not well documented but was proba-bly associated with the malaria eradicationcampaigns. As in the Americas, yellow feverwas never endemic, but many episodes ofsummertime transmission were initiated byship-borne infections (143,153). Spain,with its extensive overseas colonies, was par-ticularly vulnerable. For example, in 1856,more than 50,000 deaths were recorded inBarcelona, Cadiz, Cartagena, and Jerez. Inthe same year, 18,000 died in Lisbon,Portugal, and the disease also hit Livorno,Italy, and the French ports of Brest,Marseilles, and Saint Nazaire. In someyears, small outbreaks were reported inports as far north as Dublin, Ireland;Southampton, England; and Swansea,Wales (143,153).
Asia. Ae. aegypti is widespread in Asia, butfor reasons that are not clear, yellow fevertransmission has never been recorded there.
1668 New York, NY; Philadelphia, PA; and other settlements
1690 Charleston, SC1691 Boston, MA1693 Charleston, Philadelphia, Boston1694 Philadelphia, New York, Boston1699 Charleston, Philadelphia1702 New York1703 Charleston1728 Charleston1732 Charleston1734 Charleston; Philadelphia; New York;
Albany, NY; Boston1737 Virginia 1739 Charleston1741 Virginia, Philadelphia, New York1743 Virginia, New York1745 Charleston, New York1747 New Haven, CT1748 Charleston1751 Philadelphia, New York1762 Philadelphia1778 Philadelphia1780 Philadelphia1783 Baltimore, MD1791 Philadelphia, New York1792 Charleston1793 Philadelphia1794 Philadelphia1795 Philadelphia1796 Philadelphia1797 Philadelphia1798 Philadelphia1799 Philadelphia1800 Philadelphia1801 Norfolk, VA; New York; Massachusetts 1802 Philadelphia1803 Boston, Philadelphia1804 Philadelphia1805 Philadelphia
1807 Charleston1811 New Orleans, LA; Florida; New Jersey1817 New Orleans, Charleston, Baltimore1819 New Orleans, Charleston, Baltimore,
Philadelphia, New York1820 New Orleans, Philadelphia1821 New Orleans, Mississippi Valley, Alabama,
Charleston, Baltimore, Philadelphia, New York, Boston
1822 New Orleans, New York1823 Key West, FL1824 New Orleans, Charleston1825 Mobile, AL; Natchez, MS; Washington 1827 New Orleans, Mobile1828 New Orleans, Memphis1829 Key West, Mobile, Natchez1837 New Orleans, Mobile, Natchez1839 Galveston, TX; Mobile; Charleston1841 Key West, New Orleans1843 Galveston, Mobile, Mississippi Valley,
Charleston1847 New Orleans, Mobile, Natchez1852 Charleston1853 New Orleans1854 New Orleans, Mobile, Alabama, Charleston 1855 Mississippi Valley, Norfolk1856 New Orleans, Charleston1858 Charleston1867 Key West, Galveston, New Orleans, Mobile,
Philadelphia1870 New York1873 New Orleans, Mississippi Valley, Alabama,
Memphis1876 Charleston1877 Port Royal, SC1878 New Orleans; Memphis; Mississippi
Valley; St. Louis, MO; Chattanooga, TN;many other cities
1879 Memphis1905 New Orleans
Table 2. Notable epidemics of yellow fever in North America, north of Mexico.a,b
Year Location Year Location
aModified from a table published by WHO (186). bSome records refer to states or regions without specifying the cities involved.
109S1.Part 2 02/21/01 3:36 PM Page 154 (Black plate)

Climate change and mosquito-borne disease
Factors that Influence Yellow FeverTransmissionVaccination. Yellow fever vaccine is cheapand highly effective. Nevertheless, as alreadymentioned, enzootic transmission occurs inmany countries that no longer support com-prehensive preventive immunization. In thelast 20 years incidence has increased rapidly,particularly in rural villages in West Africa;some urban epidemics have also occurred(154). The alarming rate of increase of urbandengue throughout the world, also transmit-ted by Ae. aegypti (see below), suggests anincreasingly serious risk of such outbreaks.
Climate. Rainfall. In many enzootic areas,yellow fever is a highly seasonal disease.Rainfall is the principal determinant becausethe tree-hole breeding sites of the mosquitoesare scarce during long dry periods. Verticaltransmission from one generation of mosqui-toes to another via the egg stage is well docu-mented. In areas of interhuman transmission,outbreaks can continue during the dry seasonif water storage containers provide a significantsource of mosquitoes (143).
Temperature. As with malaria, the extrinsicincubation period of the yellow fever virusdecreases with increasing temperature, butthe effect of temperature on mosquito sur-vival and behavior is complex and hard topredict. In many endemic areas, temperaturesin the yellow fever season are cooler than inthe dry season but are much higher than theminimum for effective transmission, so varia-tion among years is probably a minor factorin disease incidence.
In temperate regions where summer trans-mission once occurred, the climate was clearlyadequate for epidemic transmission evenbefore the present warming phase. Coolweather was a limiting factor only when winterapproached. Indeed, in many cities, normalsummer temperatures were well above those oftropical regions where the disease is endemic.
Nonclimatic factors. As with malaria,transmission is influenced by a range of non-climatic factors. Many of these are associatedwith rapid population growth, urbanization,and economic development.
Contact with the forest cycle. In forestedareas, hunting, clearance for agriculture, log-ging, road building, mining, oil prospecting,and other occupations promote contact ofnonimmune humans with the forest trans-mission cycle. These activities eliminate thehabitat that supports the cycle, but their asso-ciated human settlements provide a favorableenvironment for peridomestic species such asAe. aegypti, thus increasing the risk of inter-human transmission.
Urbanization. The rapid growth ofdensely populated towns and cities through-out the tropics provides an increasingly favor-able environment for epidemic transmission.
Even when piped water is available, thesupply is often intermittent, so reserves ofwater are stored in containers that are asource of Ae. aegypti. Other artifacts—buck-ets, flowerpots, bottles, cans, defunct house-hold appliances, discarded tires, and manyother items—can also serve as breeding sites ifthey accumulate rainwater.
Movement of people. Extensive roadbuilding and cheap air travel, even in thepoorest countries, have opened up remoteareas to exploitation and development. InSouth America the resultant movement ofnonimmune people into forested areas hasbeen a major factor in the incidence of yellowfever. Infected people traveling from endemicareas could introduce the virus to urban pop-ulations (155), but despite widespread recog-nition of this increasing risk, many countriesdo not enforce vaccination laws.
Cheap international air travel also hasemerged as a significant factor in disease dis-semination. For example, in 1996 an unvacci-nated man died of yellow fever in Tennesseeafter returning to the United States from afishing trip on the Amazon River (156), andthere have recently been similar cases inEurope.
Urban mosquito control. Little attempthas been made to control yellow fever vectorsin Africa. Residual treatments for malariacontrol were effective against Ae. aegyptibecause the species tends to rest indoors.However, many countries are now abandon-ing the method in favor of bed nets impreg-nated with insecticide. These are not effectivefor yellow fever control because, unlikeanophelines, Ae. aegypti bites mainly in thedaylight hours. During recent epidemics inWest Africa, urban indices of Ae. aegypti weremore than 100 times the minimum forepidemic transmission (157,158).
The Ae. aegypti eradication campaign.With the advent of DDT after WorldWar II, an attempt was made to avert thethreat of urban yellow fever by eradicatingAe. aegypti from the entire WesternHemisphere. The initial years of this cam-paign were remarkably successful. By 1961,22 countries had been declared free of thespecies. However, as with the attempt toeradicate malaria, control was not sustainedand some areas were not affected. The mos-quito has now recolonized all its former ter-ritory, even the relatively remote island ofBermuda. Below the equator it has recentlyextended as far south as Buenos Aires. Untilrecently it remained widespread in theUnited States but has now been displacedfrom many regions by Ae. albopictus, anAsian species that was introduced in ship-ments of used tires (159–161). This dis-placement of one species by anotherillustrates a major flaw in the popular debate
on climate change: Biotic responses toclimate change cannot be predicted on themere basis of climate envelopes. The distrib-ution of a species is determined by its inter-action with other species and by many otherbehavioral and ecologic factors. It is there-fore naïve to suggest that species will moveto higher latitudes and altitudes simply onthe basis of temperature change (162).
Since the eradication attempt, many citieshave grown exponentially. Most countries inthe Americas have a program for Ae. aegypticontrol, ostensibly to prevent dengue trans-mission, but their efforts are largely ineffec-tive and infestation rates are generally high.Thus, in the aftermath of the eradicationcampaign and with the decline in preventiveimmunization, the risks of urban outbreaks ofyellow fever are probably greater than at anytime in the past 100 years (163).
Dengue
The Disease
Dengue fever is a viral disease closely relatedto yellow fever (164). As with yellow fever,forest-dwelling Aedes mosquitoes transmitdengue virus among wild monkeys, andperidomestic species, particularly Ae. aegypti,are responsible for interhuman transmissionin villages and urban areas. Thus, many of thefactors that influence transmission of yellowfever have a similar significance for dengue.
Dengue is caused by any of four closelyrelated viruses. Illness usually begins with asudden onset of fever and can include a vari-ety of other signs and symptoms such asintense headache, severe pain behind the eyes,pains of the muscles and joints, nausea, vom-iting, and a skin rash. Most dengue infectionsare self-limiting, but a small portion developinto a more serious illness—dengue hemor-rhagic fever/dengue shock syndrome(DHF/DSS)—characterized by spontaneoushemorrhage, increased permeability of theblood vessels and a condition known asphysiologic shock. Fatality rates in untreatedDHF/DSS can be as high as 50%. The inci-dence and geographic distribution of denguehave increased dramatically in recent decades:More than half the world’s population nowlives in areas at risk of infection (165). Atpresent, no vaccine is available.
Dissemination of Dengue WorldwideThere are four immunologically distinctdengue viruses. In the forest cycle, all havebeen documented in Asia, whereas only onehas been detected in Africa. This may be evi-dence that the viruses are of Asian origin. InChina, descriptions of a dengue-like illnesswith hemorrhagic manifestations date back toat least the Northern Sung Dynasty, about1,000 years ago (165).
Environmental Health Perspectives • VOLUME 109 | SUPPLEMENT 1 | March 2001 155
109S1.Part 2 02/21/01 3:36 PM Page 155 (Black plate)

Reiter
156 VOLUME 109 | SUPPLEMENT 1 | March 2001 • Environmental Health Perspectives
From the seventeenth century onwardthere are records of major epidemics ofdenguelike illness in Asia, Africa, and theAmericas. As with yellow fever, the diseaseand the vector traveled the world aboardships. For this reason, most early out-breaks—in Panama; Alexandria and Suez,Egypt; Jakarta, Indonesia; Philadelphia;Madras, India; Mombasa, Kenya; Cadiz,;Lima, Peru; and Zanzibar, Tanzania—wereassociated with seaports, and their fre-quency increased as ocean trade grew. Fromthe late eighteenth century onward, pan-demics of human dengue lasting 3–7 yearsswept around the world—much like thewandering epizootics of yellow fever in theforests of Africa and South America—andthe disease became endemic in many tropi-cal cities. In the Americas, the Ae. aegyptieradication campaign eliminated transmis-sion in many countries, but the diseaseinvariably reappeared within a few years ofreinfestation (165).
In recent decades, rapid expansion ofurban populations, the proliferation of mos-quito breeding sites, and the failure of mos-quito control efforts have exacerbated the riskof transmission. In addition, the worldwidemovement of the virus has been greatly facili-tated by international air travel. Todaydengue is by far the most prevalent mosquito-borne virus of humans and is endemic inmost populations of the tropics. Some esti-mates indicate 50–100 million cases per year,with 250,000–500,000 cases of dengue hem-orrhagic fever (165). As with malaria, manyrecent publications have discussed the preva-lence of dengue in the context of climatechange (13,14,27,28,47,48,166,167).
Dengue in the United States. The firstrecorded epidemic of denguelike disease inNorth America occurred in 1780 in the portcity of Philadelphia (168). The virus probablyarrived from the Caribbean, where it wasalready widespread. In succeeding yearsdengue was a regular visitor to the UnitedStates. Eight major pandemics were recordedfrom 1827 to the 1940s. In 1922, an esti-mated 500,000 cases occurred in Texas alone.In that year, the disease spread eastwardthrough Louisiana, Florida, and the adjoiningstates, and eventually moved south into theCaribbean. Thirty thousand cases werereported in Savannah, Georgia, of which28% had hemorrhagic manifestations (169).
Since the 1950s, the disease has beenrelatively rare. Millions of people arrive eachyear from the tropics and there are undoubt-edly many imported cases, yet there is verylittle autochthonous transmission even inareas where Ae. aegypti is common. In a sin-gle year, 1997, U.S. immigration authoritiesreported nearly 70 million personal crossingsfrom three Mexican states—Coahuila,
Nuevo León, and Tamaulipas—into Texas.Nevertheless, in the 20-year period from1980 to 1999, 62,514 suspected cases werereported to the health authorities in thoseMexican states, whereas only 64 locallyacquired cases of dengue were confirmed inTexas (Figure 6). The difference has nothingto do with vector prevalence: In 1999, dur-ing a dengue outbreak in the contiguousborder towns of Nuevo Laredo/Laredo, theBreteau index (a measure of the level of Ae.aegypti infestation; an index of 5 is widelyconsidered sufficient for epidemic transmis-sion) was 38 on the Mexican side but 91 onthe Texas side of the border (170). Thus, aswith malaria, the mere presence of a compe-tent vector is not a guarantee of disease evenif weather conditions are adequate. The con-trast in transmission rates can be attributedto several factors:
Layout of cities. The layout of modernU.S. cities is based on the automobile.Population density is relatively low. Housingareas are often discrete, interspersed withshopping precincts, industrial parks, andother nonresidential land. Much space isdevoted to wide roads, parking lots, and otheropen areas. Plot sizes are large, and the spac-ing between houses is often greater than inmany tropical cities. The number of personsper house is smaller because couples havefewer children, and the extended family is less
prevalent. All these factors limit the numberof humans that are accessible to mosquitoes,thereby reducing the likelihood of epidemictransmission.
Building structure. Ae. aegypti is a day-active species that prefers to bite indoors orin the shade. In the United States, windowsare usually fully glazed to maximize heating/cooling efficiency and are often kept perma-nently closed. The junctions between wallsand roof are well sealed. Insect screens areubiquitous in doorways and windows; manycommunities still have screen-enforcementlaws that date back to the time of malariaeradication. By contrast, most homes,schools, and workplaces in the tropics relyon natural ventilation. Windows and doorsare often kept open, especially during day-light hours. Many buildings do not havescreens, and there is often a gap between thetop of the wall and the underside of theroof. In such circumstances Ae. aegypti hasfar greater access to people than is usual inthe United States.
Air conditioning and human behavior.In the United States, particularly in theSouth, air conditioning is widely used, evenby low-income households. People spendmuch of their daily life sequestered in sealed,air-conditioned buildings. Even if infectedmosquitoes gain entry to these buildings, thelow ambient temperature and artificially dry
Figure 6. Dengue at the Texas/Mexico border from 1980 to 1999 (170). The Texas data refer to laboratory-confirmed,locally acquired infections. The Mexico data refer to clinically diagnosed cases reported to the state health authori-ties of Coahuila, Nuevo León, and Tamaulipas. In 1999, during a dengue outbreak in the contiguous border towns ofLaredo/Nuevo Laredo, indices of Ae. aegypti infestation were considerably higher in Laredo (Texas) than in NuevoLaredo (Tamaulipas, Mexico).
Texas64
Border states62,514
Laredo■
109S1.Part 2 02/21/01 3:37 PM Page 156 (Black plate)

Climate change and mosquito-borne disease
atmosphere will lessen their survival rate andextend the extrinsic incubation period, thusreducing the likelihood of transmission (171).
Air conditioning is much less common inthe tropics. People seek the coolness ofshaded, well-ventilated areas, precisely thesites where Ae. aegypti prefers to feed. Inmany countries the afternoon siesta is takenin such places and coincides with the peakbiting period. The bite of Ae. aegypti is rela-tively painless so bites are often unnoticed.After the mosquito has fed, indoor closets andother dark places provide ideal sites for it torest. All these factors promote contact withthe insect, help ensure its survival, and thusincrease the likelihood of transmission.
Dengue in Europe. A denguelike epidemicwas recorded in Cadiz and Seville in Spainfrom 1784 to 1788. In mid-June 1801, thequeen of Spain had an illness with denguelikesymptoms, including hemorrhage, which shereferred to as dengue (172). In the course ofthe nineteenth century, outbreaks of similardescription occurred in the Canary Islands andin various places around the Mediterranean:Cadiz; Athens and Crete, Greece; Turkey;Beirut, Lebanon; Jerusalem, Israel; Alexandriaand Port Said, Egypt (165). The first dengueepidemic to be confirmed serologically (inthis case, retrospectively) swept throughrefugee camps in Greece in 1927–1928. Itwas one of the most catastrophic outbreakson record anywhere in the world: An esti-mated 1,000,000 people were infected, andmore than 1,000 died of DHF/DSS(173,174). After World War II Ae. aegyptidisappeared from Europe, perhaps as a resultof the widespread use of DDT, and denguetransmission ceased.
In the past 20 years a closely relatedspecies, Ae. albopictus, has infested much ofItaly and large areas of Albania (175). Anative of Asia, it was imported in cargoes ofused tires, a larval habitat that it frequentlyshares with Ae. aegypti. A worldwide com-merce in used tires has been responsible forinfestations of Ae. albopictus in many othercountries, including a large portion of theUnited States (160,176) and much of theAmericas. The Italian infestation has beenattributed to tire imports from Atlanta,Georgia, a city where both Ae. aegypti andAe. albopictus are present (177). TheAlbanian infestation probably came fromChina. Ae. albopictus is also a vector ofdengue, although in most parts of the worldits importance is secondary compared toAe. aegypti . An American species,Ae. atropalpus, has also become establishedin Italy after introduction in used tires(178). It is not known to transmit humandisease, but these examples of recent intro-ductions are a clear warning that Ae. aegypticould also return to the region.
Outlook for the FutureModels of climate change that attempt toproject the potential impact of increased levelsof greenhouse gases indicate that the degree offuture warming is likely to be greatest close tothe poles (4,179). This change of the tempera-ture gradient would alter the world’s wind andocean circulations. In turn, such changeswould affect precipitation, humidity, seasonal-ity, and other climatic factors. The complexi-ties of these systems, along with thecomplexity of all the behavioral and ecologicfactors that influence transmission, make itimpossible to predict their impact on the inci-dence of mosquito-borne diseases. However,some generalizations are permissible.
Temperate ClimatesMalaria. In many countries at higherlatitudes, changes in lifestyles and living con-ditions were the most important factors in theelimination of malaria. Even in countrieswhere these factors were less dominant, eradi-cation of the disease did not require totalelimination of the vector. Residual treatmentswere effective because they reduced the lifespan of the adult insect, reducing the proba-bility of transmission and eventually leadingto the elimination of the parasite. Thus, theanophelines responsible for transmission arestill present in the brackish waters ofThe Netherlands, as they are in Florida. Theystill occur in the rice fields of Arkansas andItaly, the ponds of Pennsylvania and Poland,and the forest pools of Minnesota andFinland. In many of these regions, advancesin agriculture and improvements in livingstandards have limited the mosquito popula-tions and reduced their contact with humans,but this is not always the case. For example,in large areas of Italy, anopheline populationshave returned to levels not seen since beforethe DDT era. In recent surveys the vectorialcapacity of An. labranchiae near rice fields inTuscany and along the west coast of Calabriawas estimated as 7.3–26 for P. falciparum and8.3–32.5 for P. vivax, with peak landing rateson humans of more than 200 mosquitoes perhuman per night (180). In entomologicterms, these infestations are comparable tothose in endemic areas with extremely hightransmission rates; in the Garki Project, amajor study in northern Nigeria, mean vector-ial capacity for P. falciparum during the peaktransmission season (July–October) was 21.74(181). Nevertheless, despite the similarity inthis entomologic parameter, the malariogenicpotential of Italy is considered very low, andreestablishment of the disease is unlikelyunless living standards deteriorate drastically(182). Moreover, if the present warmingtrend continues, human strategies to avoidthese temperatures—particularly indoor livingand air conditioning—are likely to become
more prevalent. In consequence, the stabilityindex, already very low, will continue todecline.
Of course, this does not mean the diseasewill be entirely absent. International traveland population movement will facilitateintroductions from other parts of the world.For example, WHO recorded 12,328 cases ofimported malaria in the European Region in1997. Such cases occasionally lead to sum-mertime transmission (183), recentlyreported as far north as Toronto, Canada,and Berlin, Germany. However, in the all thewealthier countries, such outbreaks are likelyto be small, easily contained, and confined toa limited geographic area.
The same may not be true of less affluentregions. Rapid economic decline combinedwith political instability has already broughtback epidemic typhus, diphtheria, and otherinfectious diseases to several countries of theformer Soviet Union. In the 1990s epidemicmalaria has made a dramatic reappearance inArmenia, Azerbaijan, Tajikistan, andTurkmenistan. Cases have also been reportedfrom Dagestan (Russian Federation),Georgia, Kazakhstan, Kyrgyzstan, andUzbekistan (184). It is quite possible that thedisease could spread northward into Russiaand westward around the Black Sea. The1999 conflict in the Balkans is in the sameregion where hundreds of thousands wereinfected with malaria during World War I.Endemic transmission in such areas could besignificant if the parasite were to be reintro-duced. Climate change might augment thispossibility, particularly at high latitudes (e.g.,in Siberia), although low stability shouldfacilitate control.
Yellow fever and dengue. In the past,except in Asia, yellow fever and dengue haveshared a similar geographic distribution andso can be dealt with together. The existenceof a safe and effective vaccine against yellowfever should prevent the return of the virusto temperate regions. The factors that cur-rently prevent dengue transmission in theUnited States will remain dominant unlessliving standards deteriorate drastically.However, if Ae. aegypti were to reestablishitself in the Mediterranean region, it is morelikely that the disease could return. Summertemperatures are more than adequate fortransmission. Most cities are compact anddensely populated. Air conditioning is rarelyused and windows remain open for most ofthe summer. Many activities, particularlysocial gatherings, occur in outdoor situationssuch as shaded balconies, courtyards, andoutdoor restaurants—all ideal for contactwith the vector. Warmer summers wouldprobably extend the potential range of thedisease. However, as with malaria, thepoorer countries, particularly in North
Environmental Health Perspectives • VOLUME 109 | SUPPLEMENT 1 | March 2001 157
109S1.Part 2 02/27/01 6:12 PM Page 157 (Black plate)

Reiter
158 VOLUME 109 | SUPPLEMENT 1 | March 2001 • Environmental Health Perspectives
Africa, the Levant, and the Balkans, wouldbe at greatest risk. In Macedonia, for exam-ple, current refugee camps are less than 200km from the location of similar camps inGreece that were the site of the devastatingdengue/dengue hemorrhagic fever epidemicof 1927–1928.
Tropics and SubtropicsMalaria. In regions of high endemicity andstability, particularly in sub-Saharan Africa,childhood infection rates have remainedremarkably static over the past half-century.Transmission rates are so high that even thecolossal increases in anopheline populationsthat result from the introduction of rice culti-vation do not alter the indices of infection(185). In such circumstances it is unlikelythat a warmer climate would have mucheffect on incidence. Moreover, climate mod-els suggest that the low latitudes are leastlikely to be affected by any global trend.
Farther from the equator, in zones ofunstable malaria, the picture is less clear.Transmission is more sensitive to climatic fac-tors, so the impact of climate change could besignificant. A long-term increase in averagerainfall might promote high mosquito popu-lations in areas that are currently arid butreduce them where drought malaria is nowthe norm. Lower rainfall could suppresstransmission in arid areas, as appears to havehappened in recent years on the southernedge of the Sahara (105), but promotesdrought malaria in wetter ones. Increasedtemperatures could facilitate transmission inhumid areas but reduce it if associated withlow humidity. Such changes could alter therelative abundance of local mosquito species,and natural selection could modify the speciesthemselves. Climate change would induceother ecologic changes, which could lead toagricultural and economic change that mightincrease or decrease transmission potential.The plethora of factors involved is complex,and the overall impact on endemicity andstability is even harder to predict.
Higher temperatures would probably raisethe maximum altitude for transmission andcould bring the disease back to major citiessuch as Quito, Ecuador, and Mexico City,Mexico, where it has not been seen for manydecades. In most such situations, stabilitywould remain low, so control measuresshould be relatively effective. However, atlower altitudes, which are at the present limitsof transmission, increased stability and preva-lence might be more of a problem.
Other factors could have a crucialimpact. The recent history of Ae. albopictusdemonstrates that modern transportationhas greatly increased the mobility of manyspecies. The introduction of an anthro-pophilic mosquito to a new location could
drastically alter transmission potential byaltering stability. This has happened in thepast—the worldwide distribution ofAe. aegypti is perhaps the best example.Climate change might affect the likelihoodthat such introductions would lead todisease transmission.
Yellow fever and dengue. Rainfall is amajor determinant of transmission in the for-est cycle of yellow fever. Climate models donot suggest major changes in equatorialregions of Africa but indicate that rainfallmight decrease toward the Sahara. This couldreduce the overall potential for enzootic activ-ity, but the resultant reduction of immunityin the monkey population might exacerbatethe rate of epizootic transmission during rainyepisodes and enhance the likelihood of trans-fer of virus to the urban cycle. Transmissionwould also be modified if climate changeaffected the monkey populations.
Transmission by peridomestic mosquitoesis often highly seasonal. The causes of thisseasonality vary in different regions and inmany cases are complex and poorly under-stood. Thus, in areas where mosquitoes breedin stored water, their populations may beextremely high all year, yet peak transmissionoccurs in the rainy season. In many instancesthis probably occurs because it is also thehottest season and/or because the survival rateis greater at higher humidity. In other situa-tions transmission may be enhanced during
drought, when water storage becomes moreimportant. However, the mere fact thattransmission is seasonal does not imply that,for example, warmer seasons will producehigher rates of transmission. This is welli l lustrated by the record of dengue inPuerto Rico: There is no reason to supposethat climatic factors affect transmission ratesof the four serotypes of this virus in differ-ent ways, but it is clear from Figure 7 thatthe patterns of their incidence are not iden-tical. Immunologic history, expressed as theherd immunity, is presumably the domi-nant factor. Moreover, a retrospectiveanalysis of 33 years of monthly data onDHF incidence in Bangkok, Thailand, didnot reveal any relationship between climatevariables and the timing of epidemics (130).
Final Comment
The natural history of mosquito-bornediseases is complex, and the interplay ofclimate, ecology, vector biology, and manyother factors defies simplistic analysis. Therecent resurgence of many of these diseasesis a major cause for concern, but it is facileto attribute this resurgence to climatechange. The principal determinants are pol-itics, economics, and human activities. Acreative and organized application ofresources is urgently required to controlthese diseases regardless of future climatechange.
Figure 7. Monthly isolates of dengue virus, by serotype, in Puerto Rico from 1986 to 1999 (189). The viruses areessentially identical, yet the patterns of their incidence are not identical, indicating that factors other than climateare dominant in their rates of transmission.
Dengue 1
Dengue 2
Dengue 3
Dengue 4
100
80
60
40
20
0
80
60
40
20
0
80
60
40
20
0
100
80
60
40
20
0
1986 1987 1988 1989 1990 1991 1992 1993 1994 1995 1996 1997 1998 1999
Conf
irmed
cas
es
109S1.Part 2 02/27/01 6:12 PM Page 158 (Black plate)

Climate change and mosquito-borne disease
REFERENCES AND NOTES
1. Wigley TML, Ingram MJ, Farmer G, eds. Climate and History.Cambridge:Cambridge University Press, 1981.
2. Lamb HH. Climate, History and the Modern World. London:Routledge, 1995.
3. Chorley RJ, Barry RG. Atmosphere, Weather and Climate.London:Routledge, 1998.
4. Houghton JT, Meira Filho LG, Callander BA, Harris N,Kattenberg A, Maskell K, eds. The Science of Climate Change.Contribution of Working Group I to the Second Assessment ofthe Intergovernmental Panel on Climate Change (IPCC).Cambridge:Cambridge University Press, 1996.
5. Tett SFB, Stott PA, Allen MR, Ingram WJ, Mitchell JFB. Causesof twentieth-century temperature change near the earth's sur-face. Nature 399:569–572 (1999).
6. Wigley TML, Schimel D, eds. The Carbon Cycle. Cambridge:Cambridge University Press, 2000.
7. Kerr RA. Greenhouse forecasting sti l l cloudy. Science276:1040–1042 (1997).
8. Taubes G. Apocalypse not. Science 278:1004–1006 (1997).9. Longstreth J. Public health consequences of global climate
change in the United States—some regions may suffer dispro-portionately. Environ Health Perspect 107:169–179 (1999).
10. Gubler DJ. Climate change: implications for human health. HlthEnv Digest 12:54–55 (1998).
11. Cook G, ed. Manson's Tropical Diseases. London:W.B. SaundersCo., 1996.
12. McMichael AJ, Haines A, Slooff R, Kovats S. Climate Changeand Human Health. Geneva:World Health Organization (WHO),1996.
13. Watson RT, Zinyowera MC, Moss RH, eds. Impacts,Adaptations and Mitigation of Climate Change: Scientific-Technical Analyses. Contribution of Working Group II to theSecond Assessment of the Intergovernmental Panel on ClimateChange (IPCC). Cambridge:Cambridge University Press, 1996.
14. Watson RT, Zinyowera MC, Moss RH, eds. The RegionalImpacts of Climate Change: An Assessment of Vulnerability.Special Report of the Intergovernmental Panel on ClimateChange (IPCC) Working Group II. Cambridge:CambridgeUniversity Press, 1998.
15. Rampino M. Climate—History, Periodicity and Predictability.New York:Van Nostrand Reinhold, 1987.
16. Grove JM. Little Ice Age. London:Routledge and Kegan Paul, 1988.17. Calder N. The Weather Machine. New York:Viking, 1974.18. Ponte L. The Cooling. Englewood Cliffs, NJ:Prentice-Hall, 1976.19. Halacy DS. Ice or Fire? Can We Survive Climate Change? New
York:Harper and Row, 1978.20. USGCRP. Climate Change Impacts on the United States: The
Potential Consequences of Climate Variability and Change. U.S.Global Change Research Program, Washington, DC, 2001,[Available: http://www.grio.org/nationalassessment/ ].
21. Karl TR, Knight RW, Easterling DR, Quayle RG. Indices of cli-mate change in the United States. Bull Am Meteorol Soc77:279–292 (1996).
22. Michaels PJ, Knappenberger PC. Human effect on globalclimate? Nature 384:522–523 (1996).
23. Lindzen RS. Can increasing carbon dioxide cause climatechange? Proc Natl Acad Sci USA 94:8335–8342 (1997).
24. Ross A. Strange Weather. Culture, Science, and Technology inthe Age of Limits. London:Verso, 1991.
25. Gelbspan R. The Heat is On: The High Stakes Battle OverEarth's Threatened Climate. Reading, MA:Addison-Wesley,1997.
26. Michaels PJ, Balling RC. The Satanic Gases. Washington,DC:Cato Institute, 2000.
27. Patz JA, Epstein PR, Burke TA, Balbus JM. Global climatechange and emerging infectious diseases. JAMA 275:217–223(1996).
28. Epstein PR, Diaz HF, Elias S, Grabherr G, Graham NE, MartensWJM. Biological and physical signs of climate change: focus onmosquito-borne diseases. Bull Am Meteorol Soc 79:409–417(1998).
29. McMichael AJ, Patz J, Kovats RS. Impacts of global environ-mental change on future health and health care in tropicalcountries. Br Med Bull 54:475–488 (1998).
30. Martens P. Health and Climate Change: Modelling the Impactsof Global Warming and Ozone Depletion. London:EarthscanPublications, 1998.
31. Epstein PR. Climate and health. Science 285:347–348 (1999).32. Kovats RS, Haines A, Stanwell-Smith R, Martens P, Menne B,
Bertollini R. Climate change and human health in Europe. BrMed J 318:1682–1685 (1999).
33. Gil les HM, Warrell DA, eds. Bruce-Chwatt’s EssentialMalariology. London:Edward Arnold, 1993.
34. Bruce-Chwatt LJ, de Zulueta J. The Rise and Fall of Malaria inEurope, a Historico-Epidemiological study. Oxford:OxfordUniversity Press, 1980.
35. Craig MH, Snow RW, le Sueur D. A climate-based distributionmodel of malaria transmission in sub-Saharan Africa. ParasitolToday 15:105–111 (1999).
36. Geiger R. The Climate Near the Ground. Cambridge,MA:Harvard University Press, 1980.
37. Ricklefs RE, Miller GL. Ecology. New York:W.H. Freeman & Co.,1999.
38. Note: In this text, the generic name Anopheles gambiae refersto a group of “sibling” species that are identical in appearancebut can be distinguished by certain behavioral traits or by chro-mosomal mapping and other special techniques.
39. Reiter P, Jones MDR. An eclosion timing mechanism in themosquito Anopheles gambiae. J Ent (A) 50:161–168 (1975).
40. Reiter P. A study of mosquito biology in relation to contol byinsoluble surfactants [DPhil thesis]. Brighton: University ofSussex, 1978.
41. Jones MDR, Reiter P. Entrainment of the pupation and adultactivity rhythms during development in the mosquito Anophelesgambiae. Nature 254:242–244 (1975).
42. Omer SM, Cloudsley-Thompson JL. Survival of femaleAnopheles gambiae Giles through a 9-month dry season inSudan. Bull WHO:319–330 (1970).
43. Reiter P. Vector-borne Diseases Division, Annual Reports. FortCollins, CO:Centers for Disease Control (CDC), 1982, 1983.
44. National Oceanographic and Atmospheric Administration(NOAA). Online climate data:Environmental data and informationservice, National Climatic Center, Asheville, NC 28801-5001,USA. Available: http://www.ncdc.noaa.gov/ol/climate/climatedata.html [cited 16 January 2001].
45. O'Meara GF, Evans LF Jr, Gettman AD, Cuda JP. Spread ofAedes albopictus and decline of Ae. aegypti (Diptera: Culicidae)in Florida. J Med Entomol 32:554–562 (1995).
46. Lindsay SW, Birley MH. Climate change and malaria transmis-sion. Ann Trop Med Parasitol 90:573–588 (1996).
47. Jetten TH, Focks DA. Potential changes in the distribution ofdengue transmission under climate warming. Am J Trop MedHyg 57:285–297 (1997).
48. Patz JA, Martens WJM, Focks DA, Jetten TH. Dengue feverepidemic potential as projected by general circulation modelsof global climate change. Environ Health Perspect 106:147–153(1998).
49. Krebs CJ. Ecology. The Experimental Analysis of Distributionand Abundance. New York:Harper and Row, 1972.
50. Macdonald G. The Epidemiology and Control of Malaria.London:Oxford University Press, 1957.
51. Bailey NTJ. The Mathematical Theory of Infectious Diseasesand its Applications. London:Charles Griffin and Company Ltd.,1975.
52. Molineaux L. The pros and cons of modelling malaria transmis-sion. Trans R Soc Trop Med Hyg 79:743–747 (1985).
53. Molineaux L, Dietz K. Review of intra-host models of malaria.Parassitologia 41:221–231 (1999).
54. Garrett-Jones C. Problems of epidemiological entomology asapplied to malariology. Entomol Soc Am 7:168–180 (1970).
55. Najera JA. A critical review of the field application of a mathe-matical model of malaria eradication. Bull WHO 50:449–457(1974).
56. Clements AN, Paterson GD. The analysis of mortality and sur-vival rates in wild populations of mosquitoes. J Appl Ecol18:373–399 (1981).
57. Dye C. The analysis of parasite transmission by bloodsuckinginsects. Annu Rev Entomol 37:1–19 (1992).
58. Rogers DJ, Randolph SE. The global spread of malaria in afuture, warmer world. Science 289:1763–1766 (2000).
59. Dye C, Reiter P. Temperatures without fevers? Science289:1697–1698 (2000).
60. Garnham PCC. Distribution of simian malaria parasites in vari-ous hosts. J Parasitol 49:905–911 (1963).
61. Homer. The Iliad. Robert Fagles, translator. Book 22, lines31–37. New York:Viking Penguin, 1990.
62. Jones WHS. Malaria and Greek History. Manchester:Manchester University Press, 1909.
63. Langholf V. Medical theories in Hippocrates: early texts and the'Epidemics' (Untersuchungen zur Antiken Literature undGeschichte, Band 34). Berlin:Walter de Gruyter, 1990.
64. Cheeseman DF. Varro and the small beasts. Nature203:911–912 (1964).
Environmental Health Perspectives • VOLUME 109 | SUPPLEMENT 1 | March 2001 159
Dengue. A disease caused by any of four closely related viruses that are transmittedby the bite of infected Aedes mosquitoes, principally Ae. aegypti. The illness usuallybegins with a sudden onset of fever and can include various other signs and symptomssuch as intense headache, severe pain behind the eyes, pains of the muscles and joints,nausea, vomiting, and a skin rash. Dengue hemorrhagic fever/dengue shock syndrome(DHF/DSS) is a serious manifestation of dengue infection characterized by spontaneoushemorrhage, increased permeability of the blood vessels, and a condition known as physi-ologic shock. Fatality rates in untreated DHF/DSS can be as high as 50%.
Endemic transmission. Continuous transmission of a human disease pathogenwithin a given population group or geographic region. Enzootic transmission describesthe same concept among animals.
Epidemic transmission. The occurrence of numbers of cases of human disease ina community or region that are clearly in excess of the normal expectancy for that loca-tion and time. The term is thus a relative one, based on average long-term frequency in aspecified population at a specific time of year. Epizootic transmission describes the sameconcept among animals.
Malaria. A mosquito-borne infectious disease caused by protozoan parasites of thegenus Plasmodium. Human malaria is transmitted by mosquitoes of the genusAnopheles, and is characterized by cycles of chills, fevers, and sweating.
Mosquito. Insects of the family Culicidae and order Diptera (two-winged flies).There are about 3,500 species, all of which lay their eggs on or near water and areentirely aquatic until the adult stage. The mouthparts of the adults are in the form of anelongated proboscis about half as long as the body. The females of most species can usethis proboscis to pierce skin and imbibe blood from peripheral blood capillaries. A sali-vary secretion that is injected into the tissues during blood feeding can serve as a vehiclefor the transmission of pathogens.
Quartan malaria. Malaria in which the febrile episodes occur every third day (i.e.,days 1, 4, 7, etc.). This periodicity is characteristic of P. malariae infections.
Tertian malaria. Malaria in which the febrile episodes occur every alternate day(i.e., days 1, 3, 5, etc.). This periodicity is characteristic of P. vivax, P. falciparum, andP. ovale infections.
Vector species (of mosquito). A species of mosquito capable of transferring aninfectious agent from an infected animal or human to a noninfected one. Transfer nor-mally includes an obligatory incubation period that involves multiplication of thepathogen in various tissues of the insect, including the salivary glands.
Yellow fever. A viral disease of primates transmitted by the bite of infectedmosquitoes. Symptoms in humans include high fever, jaundice, and spontaneous bleed-ing. The severity of illness is quite variable, but 20–50% of jaundice cases are fatal.
Appendix. Glossary
109S1.Part 2 02/27/01 6:13 PM Page 159 (Black plate)

Reiter
160 VOLUME 109 | SUPPLEMENT 1 | March 2001 • Environmental Health Perspectives
65. Jones WHS. Malaria: a neglected factor in the history ofGreece and Rome. Cambridge:Macmillan and Bowes, 1907.
66. Dante A. The Inferno. Robert Pinsky, translator. Canto XVII,lines 75–78. New York:Farrar, Straus and Giroux, 1997.
67. Chaucer G. The Canterbury Tales. Nevill Coghill, translator. TheNun's Priest's Tale, lines 134–140. London:Penguin, 1977.
68. Campbell BMS, ed. Before the Black Death: Studies in the“Crisis” of the Early Fourteenth Century. Manchester:Manchester University Press, 1991.
69. Harrison WC. Dr. William Harvey and the Discovery ofCirculation. New York:MacMillan, 1967.
70. Harvey W. On the Motion of the Heart and Blood in Animals.Robert Willis, translator. Amherst, New York:PrometheusBooks, 1993.
71. Dock G. Robert Talbor, Madame de Sévigné, and the introduc-tion of cinchona. An episode illustrating the influence of womenin medicine. Annals of Medical History 4:241–247 (1927).
72. Siegel RE, Poynter FNL. Robert Talbor, Charles II and Cinchona.A contemporary document. Med Hist 6:82–85 (1962).
73. Dobson MJ. Bitter-sweet solutions for malaria: exploring nat-ural remedies from the past. Parassitologia 40:69–81 (1998).
74. Poser CM, Bruyn GW. An Illustrated History of Malaria. NewYork:Parthenon, 1999.
75. Defoe D. A Tour Through the Whole Island of Great Britain.London:Penguin, [1724–26]1986.
76. Dobson MJ. “Marsh Fever”—the geography of malaria inEngland. J Hist Geog 6:357–389 (1980).
77. Dobson MJ. History of malaria in England. J Roy Soc Med82:3–7 (1989).
78. Dobson MJ. Malaria in England: a geographical and historicalperspective. Parassitologia 36:35–60 (1994).
79. Dobson MJ. Contours of death and disease in early modernEngland. Cambridge:Cambridge University Press, 1997.
80. Reiter P. From Shakespeare to Defoe: Malaria in England in theLittle Ice Age. Emerg Infect Dis 6:1–11 (2000).
81. Ekblom T. Les races Suédoises de l’Anopheles maculipennis etleur role épidémiologique. Bull Soc Pathol Exot 31:647–655(1938).
82. Renkonen KO. Über das Vorkommen von Malaria in Finnland.Acta Med Scand 119:261–275 (1944).
83. Russell PF. World-wide malaria distribution, prevalence, andcontrol. Am J Trop Med Hyg 5:937–965 (1956).
84. Wesenberg-Lund C. Sur les causes du changement intervenudans le mode de nourriture de l'Anophèle maculipennis. C rSéanc Soc Biol 85:383–386 (1921).
85. James SP. The disappearance of malaria from England. Proc RSoc Med 23:71–85 (1920).
86. Crosnier F. De quelques considérations sur le paludisme métro-politain. Revue Path hum comp 53:1299–1388 (1953).
87. Laigret M. Remarques d'ordre général concernant la régressiondu paludisme dans nos pays. Revue Path hum comp53:1308–1312 (1953).
88. Galli-Valerio B. La distribution géographique des Anophèlinesen Suisse au point du vue du danger de formation de foyers demalaria. Bull schweiz Gesundheit Amt 40:440–454 (1917).
89. Hackett LW. Malaria in Europe, an ecological study.London:Oxford University Press, 1937.
90. Dymowska Z. Zimnica (malaria). In: Choroby zakazne w Polsce iich zwalczanie w latach 1919–1962 [Infectious diseases inPoland and their control, 1919–1962] (Kostrzewski J, ed).Warsaw:Panstwowy Zaklad Wydawnictw Lekarskich, 1964.
91. Simic C. Le paludisme en Yougoslavie. Bull World Health Organ15:753–766 (1956).
92. Brown AWA, Haworth J, Zahar AR. Malaria eradication andcontrol from a global standpoint. J Med Entomol 13:1–25(1976).
93. Bruce-Chwatt LJ. Malaria and its control: present situation andfuture prospects. Annu Rev Public Health 8:75–110 (1987).
94. WHO. The malaria situation in 1976. WHO Chron. 32:9–17(1978).
95. Lloyd C, Coulter JLS. Medicine and the Navy, Vol 1.Edinburgh:Livingstone, 1961.
96. Hackett LW, Missiroli A. The varieties of Anopheles maculipen-nis and their relation to the distribution of malaria in Europe.Riv Malar 14:45–109 (1935).
97. de Jong JCM. The influence of changes in chlorine content ofinland waters on malaria in Friesland. Documenta Med GeogrTrop 4:206–209 (1952).
98. Faust EC. The distribution of malaria in North America, Mexico,Central America and the West Indies. In: A Symposium onHuman Malaria, with Special Reference to North America andthe Caribbean Region. Washington, DC:American Associationfor the Advancement of Science, 1941;8–18.
99. Fisk GH. Malaria and the Anopheles mosquito in Canada. Can
Med Assoc J 25:679–683 (1931).100. Stewart DA. Malaria in Canada. Can Med Assoc J 26:239–241
(1932).101. Moulton FRE. In: A Symposium on Human Malaria, with Special
Reference to North America and the Caribbean Region.Washington, DC:American Association for the Advancement ofScience, 1941;398.
102. Reiter P. Global warming and vector-borne disease in temper-ate regions and at high altitude. Lancet 351:839–840 (1998).
103. Mouchet J, Manguin S, Sircoulon J, Laventure S, Faye O,Onapa AW, Carnevale P, Julvez J, Fontenille D. Evolution ofmalaria in Africa for the past 40 years: impact of climatic andhuman factors. J Am Mosq Control Assoc 14:121–130 (1998).
104. Omer SM. Dry season biology of Anopheles gambiae Giles inthe Sudan. Nature 217:879–880 (1968).
105. Mouchet J, Faye O, Juivez J, Manguin S. Drought and malariaretreat in the Sahel, West Africa [letter]. Lancet 348:1735–1736(1996).
106. Macdonald G, ed. Dynamics of Tropical Disease: A Selection ofPapers with Bibliographic Introduction and Bibliography.London:Oxford University Press, 1973.
107. Wigglesworth VB. Insects and the Life of Man: CollectedEssays on Pure Science and Applied Biology. London:Chapmanand Hall, 1976.
108. Gunawardena DM, Wickremasinghe AR, Muthuwatta L,Weerasingha S, Rajakaruna J, Senanayaka T, Kotta PK,Attanayake N, Carter R, Mendis KN. Malaria risk factors in anendemic region of Sri Lanka, and the impact and cost implica-tions of risk factor-based interventions. Am J Trop Med Hyg58:533–542(1998).
109. McDevitt TM. World Population Profile: U.S. Bureau of theCensus, Report WP/98. Washington, DC:U.S. GovernmentPrinting Office, 1999.
110. Walsh JF, Molyneux DH, Birley MH. Deforestation: effects onvector-borne disease. Parasitology 106:S55–S75 (1993).
111. Hackett LW. The malaria of the Andean region of SouthAmerica. Rev Inst Salubr Enferm Trop 6:239–252 (1945).
112. WHO Technical Report Series 892. WHO Expert Committee onMalaria: Twentieth Report. Geneva:World Health Organization,2000.
113. Martens P, Kovats RS, Nijhof S, de Vries P, Livermore MTJ,Bradley DJ, Cox J, McMichael AJ. Climate change and futurepopulations at r isk of malaria. Global Environ Chang9(suppl):S89–S107 (1999).
114. Loevinsohn ME. Climatic warming and increased malaria inci-dence in Rwanda. Lancet 343:714–718 (1994).
115. Harrison G. Mosquitoes, Malaria and Man: A History of theHostilities Since 1880. London:John Murray, 1978.
116. Epstein PR. Global warming and vector-borne disease [letter,with reply by Reiter]. Lancet 351:1737–1738 (1998).
117. Haines A. Global warming and vector-borne disease [letter,with reply by Reiter]. Lancet 351:1738 (1998).
118. Nelson HE. Disease, demography, and the evolution of socialstructure in highland New Guinea. Journal of the PolynesianSociety 80:204–216 (1971).
119. Christian SH. Series of typescript reports to the Director,Department of Public Health, Territory of Papua and NewGuinea. Unpublished data (manuscripts on file in Port Moresby),1947–1949.
120. Spencer TET, Spencer M, Jemesen MT, Tommerup JWJ.Special patrol report. Malaria in the Mount Hagen area. P N GMed J 1:110–113 (1956).
121. Peters W, Christian SH, Jameson JL. Malaria in the highlandsof Papua and New Guinea. Med J Aust 2:409–416 (1958).
122. Peters W, Christian SH. Studies on the epidemiology of malariain New Guinea. Trans R Soc Trop Med Hyg 54:529–536 (1960).
123. Spencer TET, Spencer DM. Malaria in the Middle Wahgi Valley(Report of presentation). Medical Journal of Australia 42(vol 2,issue 17):688 (1955).
124. Mouchet J, Laventure S, Blanchy S, Fioramonti R,Rakotonjanabelo A, Rabarison P, Sircoulon J, Roux J. La recon-quête des Hautes Terres de Madagascar par le paludisme. [Thereconquest of the Madagascar highlands by malaria]. Bull SocPathol Exot 90:162–168 (1997).
125. Garnham PCC. The incidence of malaria at high altitudes.Journal of the National Malaria Society 7:275–284 (1948).
126. Matson AT. The history of malaria in Nandi. East Afr Med J34:431–441 (1957).
127. Garnham PCC. Malaria epidemics at exceptionally high alti-tudes in Kenya. Br Med Bull 2:456–457 (1945).
128. Malakooti MA, Biomndo K, Shanks GD. Reemergence of epi-demic malaria in the highlands of Western Kenya. Emerg InfectDis 4:671–676 (1998).
129. Shanks GD, Biomndo K, Hay SI, Snow RW. Changing patterns of
clinical malaria since 1965 among a tea estate populationlocated in the Kenyan highlands. Trans R Soc Trop Med Hyg94:253–255 (2000).
130. Hay SI, Myers MF, Burke DS, Vaughn DW, Endy T, Ananda N,Shanks GD, Snow RW, Rogers DJ. Etiology of interepidemicperiods of mosquito-borne disease. Proc Natl Acad Sci USA97:9335–9339 (2000).
131. Snow RW, Ikoku A, Omumbo J, Ouma J. The epidemiology, pol-itics and control of malaria epidemics in Kenya: 1900–1998.Report prepared for Roll Back Malaria, Resource Network onEpidemics. Geneva:World Health Organization, 1999.
132. Clyde DF. Malaria in Tanzania. London:Oxford University Press,1967.
133. Matola YG, White GB, Magayuka SA. The changed pattern ofmalaria endemicity and transmission at Amani in the easternUsambara mountains, north-eastern Tanzania. J Trop Med Hyg90:127–134 (1987).
134. Ellman R, Maxwell C, Finch R, Shayo D. Malaria and anaemia atdifferent altitudes in the Muheza district of Tanzania: childhoodmorbidity in relation to level of exposure to infection. Ann TropMed Parasitol 92:741–753 (1998).
135. Garnham DB, Wilson ME. Malaria in Kigezi, Uganda. J TropMed Hyg 51:156–159 (1948).
136. Meyus H, Lips M, Caubergh H. L'état actuel du problème dupaludisme d'altitude au Rwanda, Burundi. Ann Soc Belg MedTrop 42:771–782 (1962).
137. Lindblade KA. Land use change alters malaria transmissionparameters by modifying temperature in a highland area ofUganda. Trop Med Int Health 5:263–274 (2000).
138. Gascon J, Pluymaekaers J, Bada JL. Changing patterns ofmalaria in Nyarutovu (Rwanda). Trans R Soc Trop Med Hyg78:421–422 (1984).
139. Vincke I, Jadin JB. Contribution à l'étude de l'anophélisme enpays d'altitude. Ann Soc Belg Med Trop 26:483–500 (1946).
140. Coosemans M, Wery M, Storme B, Hendrix L, Mfisi P.Epidémiologie du paludisme dans la plaine de la Ruzizi,Burundi. Ann Soc Belg Med Trop 64:135–158 (1984).
141. Niell M, Sexton J, Steketee R. Personal communications.142. Roberts DR, Laughlin LL, Hsheih P, Legters LJ. DDT, global
strategies, and a malaria control crisis in South America [seecomments]. Emerg Infect Dis 3:295–302 (1997).
143. Monath TP. Yellow fever. In: The Arboviruses: Epidemiologyand Ecology, Vol V (Monath TP, ed). Boca Raton, FL:CRC,1988;139–231.
144. Darsie RF, Ward RA. Identification and geographical distributionof the mosquitoes of North America, north of Mexico. MosqSyst Suppl 1:1–313 (1981).
145. Christophers SR. Aëdes aegypti (L.), the Yellow Fever Mosquito.Its Life History, Bionomics and Structure. Cambridge:CambridgeUniversity Press, 1960.
146. Durieux C. Mass yellow fever vaccination in French Africa southof Sahara. In: Yellow Fever Vaccination, Monograph Series, Vol30 (Smithburn KC, Durieux C, Koerber R, Penna HA, Dick GWA,Courtois G, de Sousa Manso C, Stuart G, Bonnel PH, eds).Geneva:World Health Organization (WHO), 1956;115–121.
147. Monath TP. Yellow fever: Victor, Victoria? Conqueror, con-quest? Epidemics and research in the last forty years andprospects for the future. Am J Trop Med Hyg 45:1–43 (1991).
148. Robertson SE, Hull BP, Tomori O, Bele O, LeDuc JW, Esteves K.Yellow fever: a decade of reemergence. JAMA 276:1157–1162(1996).
149. Estes JW, Smith BG, eds. A Melancholy Scene of Devastation.The Public Response to the 1793 Philadelphia Yellow FeverEpidemic. Canton, MA:Science History Publications/USA, 1997.
150. Foster KR, Jenkins MF, Toogood AC. The Philadelphia yellowfever epidemic of 1793. Sci Am 279(2):88–93 (1998).
151. Bloom KJ. The Mississippi Valley's great yellow fever epidemicof 1878. Baton Rouge:Louisiana State University Press, 1993.
152. Delaporte F. The History of Yellow Fever. An Essay on the Birthof Tropical Medicine. Cambridge, MA:MIT Press, 1991.
153. Eager JM. Yellow Fever in France, Italy, Great Britain andAustria, and bibliography of yellow fever in Europe. YellowFever Institute, Bulletin No. 8:25–35 (1902).
154. Monath TP, Nasidi A. Should yellow fever vaccine be includedin the expanded program of immunization in Africa? A cost-effectiveness analysis for Nigeria. Am J Trop Med Hyg48:274–299 (1993).
155. Reiter P, Cordellier R, Ouma JO, Cropp CB, Savage HM, SandersEJ, Marfin AA, Tukei PM, Agata NN, Gitau LG, et al. Firstrecorded outbreak of yellow fever in Kenya, 1992–1993. II:Entomologic investigations. Am J Trop Med Hyg 59:650–656(1998).
156. McFarland JM, Baddour LM, Nelson JE, Elkins SK, Craven RB,Cropp BC, Chang G-J, Grindstaff AD, Craig AS, Smith RJ.
109S1.Part 2 02/27/01 6:14 PM Page 160 (Black plate)

Climate change and mosquito-borne disease
Imported yellow fever in a United States citizen. Clin Infect Dis25:1143–1147 (1997).
157. Nasidi A, Monath TP, DeCock K, Tomori O, Cordellier R, OlaleyeOD, Harry TO, Adeniyi JA, Sorungbe AO, Ajose-Coker AO, et al.Urban yellow fever epidemic in western Nigeria, 1987. Trans RSoc Trop Med Hyg 83:401–406 (1989).
158. Miller BR, Monath TP, Tabachnick WJ, Ezike VI. Epidemic yel-low fever caused by an incompetent mosquito vector. Trop MedParasitol 40:396–399 (1989).
159. Reiter P, Sprenger D. The used tire trade: a mechanism for theworldwide dispersal of container breeding mosquitoes. J AmMosq Control Assoc 3:494–501 (1987).
159. Reiter P, Sprenger D. The used tire trade: a mechanism for theworldwide dispersal of container breeding mosquitoes. J AmMosq Control Assoc 3:494–501 (1987).
160. Hawley WA, Reiter P, Copeland RS, Pumpuni CB, Craig GB Jr.Aedes albopictus in North America: probable introduction inused tires from northern Asia. Science 236:1114–1116 (1987).
161. Reiter P. Aedes albopictus and the world trade in used tires,1988–1995: the shape of things to come? J Am Mosq ControlAssoc 14:83–94 (1998).
162. Davis AJ, Jenkinson LS, Lawton JH, Shorrocks B, Wood S.Making mistakes when predicting shifts in species range inresponse to global warming. Nature 391:783–786 (1998).
163. Monath TP. Facing up to re-emergence of urban yellow fever[comment]. Lancet 353:1541 (1999).
164. Gubler DJ, Kuno G, eds. Dengue and Dengue HemorrhagicFever. Wallingford, UK:CAB International, 1997.
165. Gubler DJ. Dengue and dengue hemorrhagic fever: its historyand resurgence as a global public health problem. In: Dengueand Dengue Hemorrhagic Fever (Gubler DJ, Kuno G, eds).Wallingford, UK:CAB, 1997;1–22.
166. Martens WJM, Jetten TH, Focks DA. Sensitivity of malaria,schistosomiasis and dengue to global warming. Clim Change35:145–156 (1997).
167. Martens WJ. Health impacts of climate change and ozonedepletion: an ecoepidemiologic modeling approach. EnvironHealth Perspect 106(suppl 1):241–251 (1998).
168. Rush AB. An Account of the Bilious Remitting Fever as ItAppeared in Philadelphia in the Summer and Autumn of theYear 1780. Philadelphia, PA:Prichard and Hall, 1789.
169. Ehrenkranz NJ, Ventura AK, Cuadrado RR, Pond WL, Porter JE.Pandemic dengue in Caribbean countries and the southernUnited States—past, present and potential problems. N Engl JMed 285:1460–1469 (1971).
170. National Institute of Health, Mexico; Texas Department ofHealth; U.S. Public Health Service. Unpublished data.
171. Gahlinger PM, Reeves WC, Milby MM. Air conditioning andtelevision as protective factors in arboviral encephalitis risk.Am J Trop Med Hyg 35:601–610 (1986).
172. Rigau-Perez JG. The early use of break-bone fever (Quebrantahuesos, 1771) and dengue (1801) in Spanish. Am J Trop MedHyg 59:272–274 (1998).
173. Cardamatis JP. La dengue en Grèce. Bull Soc Pathol Exot22:272–292 (1929).
174. Halstead SB, Papaevangelou G. Transmission of dengue 1 and 2viruses in Greece in 1928. Am J Trop Med Hyg 29:635–637(1980).
175. Adhami J, Reiter P. Introduction and establishment of Aedes(Stegomyia) albopictus skuse (Diptera: Culicidae) in Albania.J Am Mosq Control Assoc 14:340–343 (1998).
176. Moore CG, Mitchell CJ. Aedes albopictus in the United States:ten-year presence and public health implications. Emerg InfectDis 3:329–334 (1997).
177. Dalla Pozza GL, Romi R, Severini C. Source and spread of Aedesalbopictus in the Veneto region of Italy. J Am Mosq ControlAssoc 10:589–592 (1994).
178. Romi R, Sabatinell i G, Savell i LG, Raris M, Zago M,Malatesta R. Identification of a North American mosquitospecies, Aedes atropalpus (Diptera: Culicidae), in Italy. J Am
Mosq Control Assoc 13:245–246 (1997).179. Houghton JT. Global Warming: the Complete Briefing.
Cambridge:Cambridge University Press, 1997.180. Romi R, Pierdominici G, Severini C, Tamburro A, Cocchi M,
Menichetti D, Pili E, Marchi A. Status of malaria vectors in Italy.J Med Entomol 34:263–271 (1997).
181. Molineaux L, Gramiccia G. The Garki Project. Research on theEpidemiology and Control of Malaria in the Sudan Savanna ofWest Africa. Geneva:World Health Organization (WHO), 1980.
182. Sabatinelli G. WHO, Regional Office for Europe, Copenhagen,Denmark. Personal communication, October 2000.
183. Zucker JR. Changing patterns of autochthonous malaria trans-mission in the United States: a review of recent outbreaks.Emerg Infect Dis 2:37–43 (1996).
184. Sabatinelli G. Malaria situation and implementation of theglobal malaria control strategy in the WHO European region.Geneva:WHO Expert Committee on Malaria MAL/EC20/98.9(1998).
185. Robert V, Gazin P, Boudin C, Molez JF, Ouedraogo V, CarnevaleP. La transmission du paludisme en zone de savane arborée eten zone rizicole des environs de Bobo-Dioulasso, Burkina-Fasso.Ann Soc Belg Med Trop 65:201–214 (1985).
186. Vaino J, Cutts F. Yellow fever. Geneva:WHO/EPI/GEN/98.11:78(1998).
187. Reiter P. The action of lecithin monolayers on mosquitoes. III:studies in irrigated rice-fields in Kenya. Ann Trop Med Parasitol74:541–557 (1980).
188. Williams LL. The anti-malaria program in North America. In: ASymposium on Human Malaria, With Special Reference toNorth America and the Caribbean Region, Vol. 15 (Moulton FR,ed). Washington, DC:American Association for theAdvancement of Science, 1941;365–370.
189. CDC Dengue Branch and Puerto Rico Health Department.Unpublished data, 2000.
Environmental Health Perspectives • VOLUME 109 | SUPPLEMENT 1 | March 2001 161
109S1.Part 2 02/27/01 6:14 PM Page 161 (Black plate)