Embed Size (px)
Citation preview

16
Emerging Strategies to Face Challenges Imposed by Climate
Change and Abiotic Stresses in Wheat
Bharti Garg, Shreelekha Misra, and Narendra Tuteja
Abstract
Abiotic stress cues, including dehydration, salinity, thermotolerance and cold,affect plant growth, posing serious threats for sustainable agriculture. Overcomingthese problems has become an urgent need due to their effects on global foodsecurity. Plant adaptation to stress conditions leads to modulation in the genomic,proteomic, and metabolomic architecture of the plant. Here, we show the series ofphysiological and molecular programs in stress adaptation regarding wheat plants.Basically, we focus on how candidate genes coupled with their networks, proteins,and metabolites change upon exposure to abiotic stress in wheat. Moreover, weraise the role of omics along with a system biological approach to ultimatelyenhance plant fitness in the future under fluctuating climate conditions.
16.1
Introduction
Plants are sessile organisms in nature. To survive they must cope with climatechanges or so-called environmental stress factors, such as drought, heat, salinity,elevated carbon dioxide (CO2), and so on. Climate changes affect all aspects of aplant’s life and pose a serious threat for developing suitable agricultural systems forsignificant growth of the global population [1]. To handle climate changes, plantshave developed a wide spectrum of molecular responses to sense the changes andsurvive accordingly. This also depends on various factors, such as species andgenotype, duration and severity of stresses, tissue type, and developmental stage[2]. Since in nature plants are often exposed to multiple environmental perturba-tions, there is an increasing requirement to breed crops or to produce transgenicplants with enhanced tolerance traits to a variety of environmental stresses.Understanding these adaptation events under constantly changing environmentalconditions has been a matter of research for many decades, although we are still farfrom a complete understanding of the tolerance mechanism of how plants copewith these stresses.
437
Climate Change and Plant Abiotic Stress Tolerance, First Edition. Edited by Narendra Tuteja and Sarvajeet S. Gill.� 2014 Wiley-VCH Verlag GmbH & Co. KGaA. Published 2014 by Wiley-VCH Verlag GmbH & Co. KGaA.

The major cereal crops, such as rice, maize, and wheat, are importantcomponents of the human diet. Among them, wheat is the second most importantcereal on Earth, and provides a large fraction of the dietary protein and total foodsupply. It is considered as a staple food of more than 35% of the world population,whose production is directly related to social stability and sustainability (http://eap.mcgill.ca/Publications/eap_head.htm). However, with progressive global climatechange, wheat production is influenced greatly and reports that correlated theyields of the four largest commodity crops to climate indicate that global wheatproduction has declined by 5.5% due to the effects of climate change in 2012. TheUN Food and Agriculture Organization forecast suggests worldwide wheatproduction in 2013 will be 690 million tonnes; however, with increasing demandfor food supply, this production would be required to be around 1 billion tonnes by2020 and around 1.6 tonnes billion by 2030 (http://www.fao.org/docrep/004/y3557e/y3557e08.htm). Therefore, efforts and new strategies are developing for theimprovement of more abiotic stress-resistant wheat crops [3–5]. Here, we focus onsome of the important physiological and molecular innovations in wheat – a plantknown to retain its evolutionary characteristics and therefore a potential candidateto have a repository of abiotic stress-tolerant genes that might be used for cropimprovement [6]. We mainly focus on the effects of some major limiting factorslike drought, salinity, elevated temperature, and cold tolerance, as well as on therole of functional and comparative genomics in wheat improvement.
16.2
Physiological and Molecular Adaptive Strategies in Wheat
Understanding the physiological, cellular, and molecular mechanism of plantresponses to environmental stress factors that adversely affect plant growth andproductivity is a field of great relevance. Drought, salinity, and extreme temperaturecan induce similar effects on plants. For example, both drought and salinity stressresult in the disruption of ion distribution and homeostasis in the cell [7,8], whileoxidative stress further causes denaturation of structural and functional proteins.As a consequence, these environmental cues often activate more or less similar cellsignaling pathways [9,10] and cellular responses, such as the production of stressproteins, accumulation of compatible solutes [11], and upregulation of antioxidantmachinery. The plant response as a stress tolerance mechanism involves theactivity of a variety of genes and biochemical/molecular pathways as represented inFigure 16.1. These genes can be categorized in three main categories:
i) Genes that are involved in transcriptional and signaling cascades, such as MYC,mitogen-activated protein (MAP), and salt overly sensitive (SOS) kinases [10],phospholipases [12], and transcription factors such as the CRT-binding factor(CBF)/DRE-binding protein (DREB) and ABF/ABAE families (abscisic acid(ABA)-responsive element-binding factor/ABA-responsive promoter element)[13,14].
438 16 Emerging Strategies to Face Challenges Imposed by Climate Change and Abiotic Stresses in Wheat
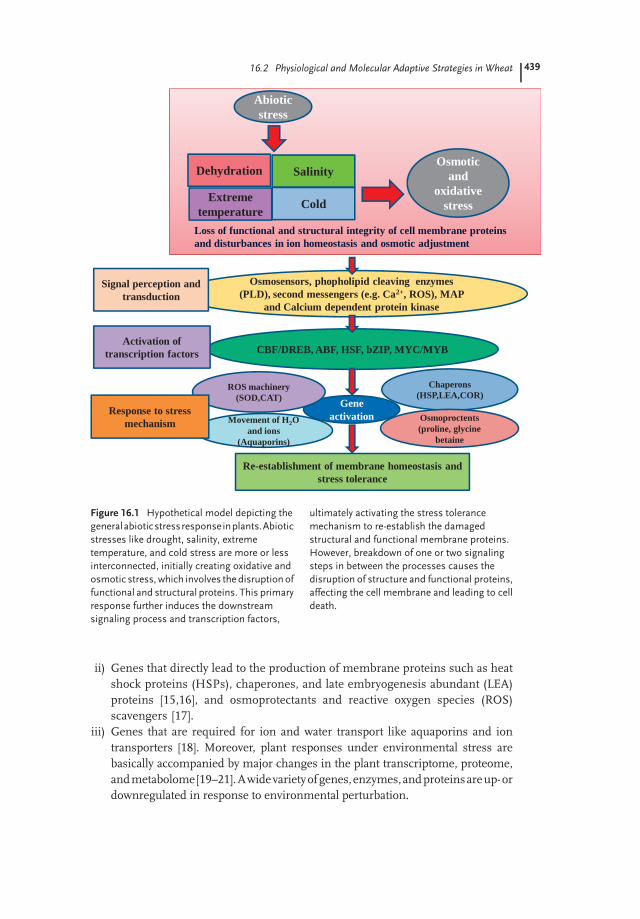
ii) Genes that directly lead to the production of membrane proteins such as heatshock proteins (HSPs), chaperones, and late embryogenesis abundant (LEA)proteins [15,16], and osmoprotectants and reactive oxygen species (ROS)scavengers [17].
iii) Genes that are required for ion and water transport like aquaporins and iontransporters [18]. Moreover, plant responses under environmental stress arebasically accompanied by major changes in the plant transcriptome, proteome,andmetabolome[19–21].Awidevarietyofgenes,enzymes,andproteinsareup-ordownregulated in response to environmental perturbation.
Dehydration Salinity
Extreme temperature
Cold
Abiotic stress
Osmotic and
oxidative stress
Loss of functional and structural integrity of cell membrane proteins and disturbances in ion homeostasis and osmotic adjustment
Osmosensors, phopholipid cleaving enzymes (PLD), second messengers (e.g. Ca2+, ROS), MAP
and Calcium dependent protein kinase
CBF/DREB, ABF, HSF, bZIP, MYC/MYB
Signal perception and transduction
Activation of transcription factors
Gene activation
Chaperons (HSP,LEA,COR)
ROS machinery (SOD,CAT)
Movement of H2O and ions
(Aquaporins)
Osmoproctents (proline, glycine
betaine
Response to stress mechanism
Re-establishment of membrane homeostasis and stress tolerance
Figure 16.1 Hypothetical model depicting the
generalabioticstressresponseinplants.Abiotic
stresses like drought, salinity, extreme
temperature, and cold stress are more or less
interconnected, initially creating oxidative and
osmotic stress, which involves thedisruptionof
functional and structural proteins. This primary
response further induces the downstream
signaling process and transcription factors,
ultimately activating the stress tolerance
mechanism to re-establish the damaged
structural and functional membrane proteins.
However, breakdown of one or two signaling
steps in between the processes causes the
disruption of structure and functional proteins,
affecting the cell membrane and leading to cell
death.
16.2 Physiological and Molecular Adaptive Strategies in Wheat 439
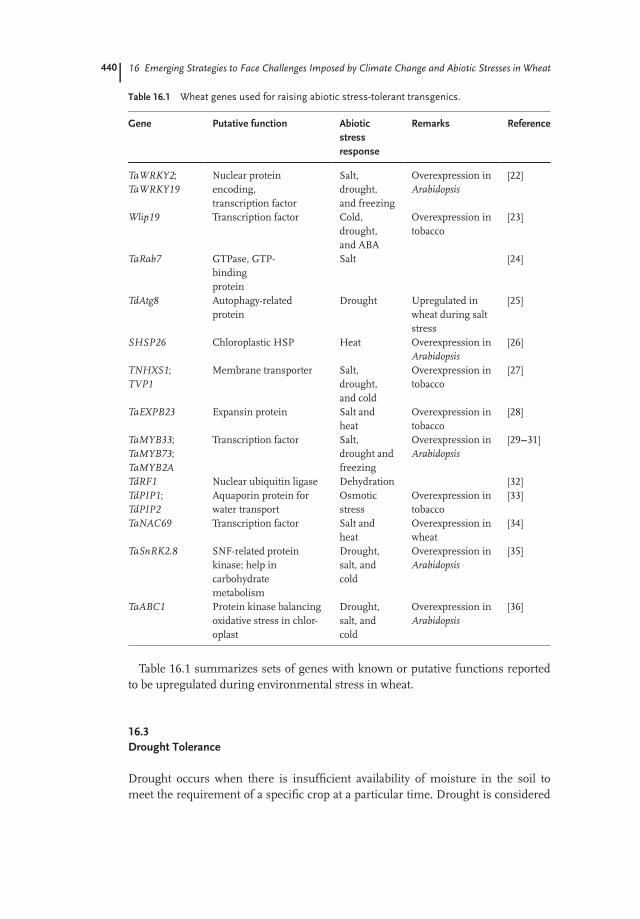
Table 16.1 summarizes sets of genes with known or putative functions reportedto be upregulated during environmental stress in wheat.
16.3
Drought Tolerance
Drought occurs when there is insufficient availability of moisture in the soil tomeet the requirement of a specific crop at a particular time. Drought is considered
Table 16.1 Wheat genes used for raising abiotic stress-tolerant transgenics.
Gene Putative function Abiotic
stress
response
Remarks Reference
TaWRKY2;TaWRKY19
Nuclear proteinencoding,transcription factor
Salt,drought,and freezing
Overexpression inArabidopsis
[22]
Wlip19 Transcription factor Cold,drought,and ABA
Overexpression intobacco
[23]
TaRab7 GTPase, GTP-bindingprotein
Salt [24]
TdAtg8 Autophagy-relatedprotein
Drought Upregulated inwheat during saltstress
[25]
SHSP26 Chloroplastic HSP Heat Overexpression inArabidopsis
[26]
TNHXS1;TVP1
Membrane transporter Salt,drought,and cold
Overexpression intobacco
[27]
TaEXPB23 Expansin protein Salt andheat
Overexpression intobacco
[28]
TaMYB33;TaMYB73;TaMYB2A
Transcription factor Salt,drought andfreezing
Overexpression inArabidopsis
[29---31]
TdRF1 Nuclear ubiquitin ligase Dehydration [32]TdPIP1;TdPIP2
Aquaporin protein forwater transport
Osmoticstress
Overexpression intobacco
[33]
TaNAC69 Transcription factor Salt andheat
Overexpression inwheat
[34]
TaSnRK2.8 SNF-related proteinkinase; help incarbohydratemetabolism
Drought,salt, andcold
Overexpression inArabidopsis
[35]
TaABC1 Protein kinase balancingoxidative stress in chlor-oplast
Drought,salt, andcold
Overexpression inArabidopsis
[36]
440 16 Emerging Strategies to Face Challenges Imposed by Climate Change and Abiotic Stresses in Wheat
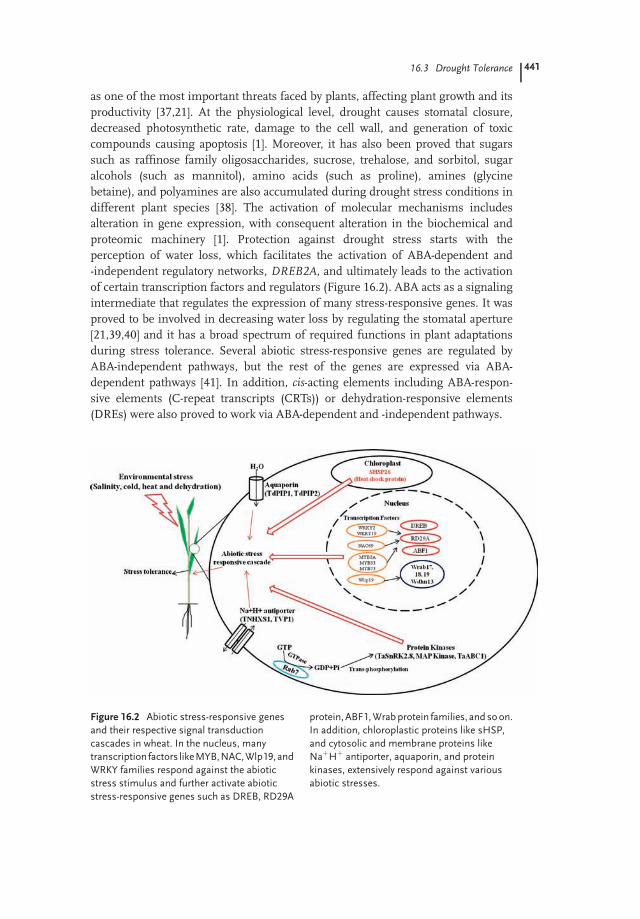
as one of the most important threats faced by plants, affecting plant growth and itsproductivity [37,21]. At the physiological level, drought causes stomatal closure,decreased photosynthetic rate, damage to the cell wall, and generation of toxiccompounds causing apoptosis [1]. Moreover, it has also been proved that sugarssuch as raffinose family oligosaccharides, sucrose, trehalose, and sorbitol, sugaralcohols (such as mannitol), amino acids (such as proline), amines (glycinebetaine), and polyamines are also accumulated during drought stress conditions indifferent plant species [38]. The activation of molecular mechanisms includesalteration in gene expression, with consequent alteration in the biochemical andproteomic machinery [1]. Protection against drought stress starts with theperception of water loss, which facilitates the activation of ABA-dependent and-independent regulatory networks, DREB2A, and ultimately leads to the activationof certain transcription factors and regulators (Figure 16.2). ABA acts as a signalingintermediate that regulates the expression of many stress-responsive genes. It wasproved to be involved in decreasing water loss by regulating the stomatal aperture[21,39,40] and it has a broad spectrum of required functions in plant adaptationsduring stress tolerance. Several abiotic stress-responsive genes are regulated byABA-independent pathways, but the rest of the genes are expressed via ABA-dependent pathways [41]. In addition, cis-acting elements including ABA-respon-sive elements (C-repeat transcripts (CRTs)) or dehydration-responsive elements(DREs) were also proved to work via ABA-dependent and -independent pathways.
Figure 16.2 Abiotic stress-responsive genes
and their respective signal transduction
cascades in wheat. In the nucleus, many
transcription factors likeMYB,NAC,Wlp19,and
WRKY families respond against the abiotic
stress stimulus and further activate abiotic
stress-responsive genes such as DREB, RD29A
protein,ABF1,Wrabprotein families, andsoon.
In addition, chloroplastic proteins like sHSP,
and cytosolic and membrane proteins like
NaþHþ antiporter, aquaporin, and protein
kinases, extensively respond against various
abiotic stresses.
16.3 Drought Tolerance 441

ABA induces the activation of transcription factors involved in many develop-mental stages as well as stress responses. It generates a cascade of signals thattriggers stomatal closure and up- or downregulation of genes during waterdeficiency [42]. Transcription factors known to be responsive for drought toleranceand ABA-dependent included the NAC, AREB/ABF, and MYB families. Addition-ally, TaCHP, which is a Cys–His–Pro-rich zinc finger protein with three divergentG-domains and has the capacity to bind with Zn, also showed tolerance todehydration and salinity through ABA-independent pathways [43]. Beside this,TaLTP1 and TaLTP2, encoding lipid transfer proteins in wheat, also showed higherlevels of expression in the tissue layers between the vascular bundle, and theymight play an important role in drought tolerance in wheat [44]. The proteinphosphatase PP2A is a key enzyme that controls potassium channel activity inguard cells and plays a role in signal transduction [45]. Tobacco plants over-expressing PCAPE2/TaPP2AC-1 exhibited stronger drought tolerance whencompared to wild-type plants. This may suggest that TaPP2AC-1 could be a positiveregulator of signal transduction pathways [46,47]. The sucrose non-fermenting(SNF) protein kinase TaSnRK2.8 was proposed to function in drought tolerance viaan ABA-dependent pathway. SNK belongs to the calcium-dependent protein kinase(CDPK)–SnRK superfamily, whose overexpression in Arabidopsis leads to greatertolerance for drought stress [48]. The protein kinase Pkabai was strongly induced inan ABA-dependent manner at the transcriptional level. Analysis of the Pkabaitranscript showed that it could be important in controlling stomatal aperture anddrought tolerance [49]. Wheat CBF homologs such as TaDBEB1 also play a pivotalrole in the signal pathway controlling drought tolerance by mediating the ABA-responsive pathway [50]. Moreover, various MYB-related transcription factors,including TaMYB1, TaMYB2, and TaMYB15, were proposed to function in otherhormone signaling pathways along with ABA [51]. The inactivation of SAL1, anegative regulator of ABA-dependent and -independent stress-responsive path-ways, resulted in higher accumulation of osmoprotectants and higher relative watercontent. Recent research showed that TaNAC4 and TaNAC8 were induced in bothbiotic and abiotic stress responses in wheat [52,53]. Analysis of lines overexpressingTaNAC2 indicated increased tolerance to drought, salt, and freezing stresses inArabidopsis [54]. In addition, previous research showed that TaNAC4 and TaNAC8are also involved in both abiotic and biotic stress responses in wheat [52,53].Interestingly, the plant response under drought conditions was found to be co-
related to the circadian clock. The activity of CAT1 enzyme is regulated by ABA andits induction is under the circadian clock, since it binds to an ABRE promoter [55].The studies above emphasize the regulatory network of transcription factors inplant cells, involving different modes of action via ABA-dependent pathways underdrought stress.Recent evidence showed that ubiquitination also plays an important role in
regulating plant responses to abiotic stress. The TaE3 gene has been recentlylaunched as an E3 ligase that mediates plant tolerance in adverse environmentalconditions [56]. A DREB2A transcription factor modulates water-deficit-inducible
442 16 Emerging Strategies to Face Challenges Imposed by Climate Change and Abiotic Stresses in Wheat

gene expression and requires post-translational modification for its activation,although the mechanism is still not clear. DREB2A inter protein 1 and 2 andC3HC4 ring domain-containing protein interact with DREB2A protein, whichfunctions as a E3 ubiquitin ligase and is capable of mediating DREB2Aubiquitination by negatively regulating the drought response by targeting DREB2Ato the 26S proteasome machinery [56]. In some cases, DREB2A overexpression alsoresulted in a significant drought tolerance by positively regulating drought-responsive gene expression [57]. Recent insights proved that two novel proteins,DRIP1 and DRIP2, could act as regulators for the drought response by mediatingDREB2A ubiquitination [1]. The overexpression of PUB22 and PUB23 genes,encoding proteins with U-box domain and E3 ubiquitin activity, leads to droughtintolerance, while pub22 and pub23 mutants are more tolerant to drought stress.In addition, the cytosolic RPN12a is a known target protein for ubiquitinationby PUB22 and PUB23 [58]. These findings emphasize the importance of trans-cription factors like DREB2A and the role of ubiquitination in drought stressresponses. Other studies highlight the importance of ubiquitin E3 ligases in themediation of drought tolerance mechanism. The coordinated role of PUB22 andPUB23 in drought signaling pathways by targeting known RPN12a protein wasalso proven [58].Apart from the role of ABA, DREB2A, and ubiquitination, other systems that also
contribute towards drought tolerance include membrane transport proteins,certain kinases, and ROS signaling pathways [59]. In wheat (Triticum aestivum), thevascular Hþ translocating pyrophosphatase (V-PPase) genes (i.e., TaVP1, TaVP2,and TaVP3) were observed to be differentially regulated in response to differentabiotic stresses, dehydration, and salinity stress [60]. TaRLK-R3 is a novel plasmamembrane-localized receptor-like kinase (RLK) that was shown to play animportant role in combating abiotic stress tolerance in wheat [61]. Recent reportsin rice showed that DSM, a novel nuclear-localized protein kinase, played a criticalrole in drought and oxidative stress tolerance by directly or indirectly modulatingthe POX22.3 and POX8.1 genes, and free radical scavenging [59]. In wheat, theirfunction is yet to be explored. The TdDHN15.2, TaDHN15.1, and TaDHN13(dehydrin) genes were upregulated during drought stress in durum wheat [1].Stress protection machinery include genes that encodes antioxidant enzymes
such as superoxide dismutases (SODs) (FeSOD, MnSOD, and CuZn SOD),catalase (CAT), ascorbate peroxidase (APX), glutathione reductase, andglutathione peroxidase (GPX) [55]. These genes were induced up to 50-foldduring drought stress, while the isoprene synthase (ISPS) transcript and itsprotein concentration decreased in black poplar (Populus nigra) plants [62].Two-dimensional electrophoresis and MALDI-TOF (matrix associated laserabsorption/ionization time of flight) mass spectrometry showed differentiationbetween control and stressed samples by obtaining 36 protein spots thatproved to be involved in gluconeogenesis and glycolysis in wheat [63]. Theseproteins have been considered as important biomarkers at the molecular levelfor improving lines with drought resistance.
16.3 Drought Tolerance 443

16.4
Salinity Tolerance
Salinity is considered one of the severe environmental stresses that impairs a largeportion of agricultural land. High amounts of sodium in the plant cytosol affect cellmetabolism and the photosynthetic rate by inducing osmotic and ionic imbalance[64]. High salinity leads to adverse changes in lipid and protein composition in theplant cell plasma membrane, causes ionic disparity, and eventually affects plantgrowth and development [65]. At the molecular level, the adaptation of wheat plantsto high salinity involves components of the SOS pathway, ABA-induced genes,salinity-induced transcription factors, ROS regulation along with the involvementof peroxisomal and apoplastic enzymes [66–69].The SOS pathway regulates Naþ/Kþ homeostasis during severe salt conditions
and maintains the low concentration of cytoplasmic Naþ by sequestering excessNaþ ions in vacuoles. The overexpression of SOS2 or SOS3, and SOS1 knockdown,showed high tolerance to salinity in wheat. Genetic evidence established that SOS2,SOS3 and SOS1 function via a common pathway [65]. This study suggested thatprotein the kinase complex SOS2/SOS3 can phosphorylate SOS1 and stimulate itsactivity towards salt tolerance. The SOS pathway in Arabidopsis can be operatedcommonly in wheat. The expression of the plasma membrane Naþ/Hþ antiporterSOS1 depends upon large number of kinase complexes like SOS2/SOS3. Autopho-sphorylation of Ser228 in SOS2 is considered it to be of primary importance for itsfunction during salt stress [70]. The DHN5 gene, belonging to the LEA proteinfamily, is also associated with salt stress [71]. Prefoldins, also known as genesinvolved in microtubule biogenesis, form a heterohexameric PFD complex,resulting in tubulin and actin folding. Null mutations in the PFD3 and PFD5genes, encoded in Arabidopsis, led to decreased levels in a- and b-tubulin, causingchanges in microtubule structure [72]. TaMKP1 is a novel, nucleus-localized wheat-responsive MAP kinase phosphatase that showed greater tolerance towards saltstress [73]. In addition, peroxisomes were found to be effective organelles in storingNO in the cytosol during salt stress [74]. NO accumulation during salt stress acts asa prerequisite for the generation of peroxynitrite (ONOO�), thus producing proteintyrosine nitration, which is a biomarker of nitrosative stress. The TaOPR1 (12-oxo-phytodienoic acid reductases) gene mediates salt tolerance by modulating the ABAsignaling pathway.Modification of lipid components in the plasma membrane is also one important
pathway for salinity tolerance in wheat [6]. Recently, it was suggested that thismodification could influence membrane stability due to structural and functionalchanges in plasma membrane proteins such as aquaporins and plasma membrane-localized Hþ-ATPase [75]. Recent reports showed that overexpression of TaAPQ8 intobacco conferred more salinity tolerance by controlling water permeability [76].Furthermore, the activity of TaCHP (Cys-His-Pro-rich protein) was associated withincreased accumulation of DREB2A, AtCBF3, and AtRAB18 under salinity stressconditions [77]. Salt stress greatly affects the structure of apoplastic proteins as well.An apoplastic protein WPM-1, a homolog of the LEA gene OsNAC045, was strongly
444 16 Emerging Strategies to Face Challenges Imposed by Climate Change and Abiotic Stresses in Wheat

induced under salt stress, highlighting the role of apoplastic proteins in salt stresstolerance [78]. Mutation in the AtHKT1 gene suppressed the SOS3 mutantphenotype [79]. The AtHKT1 gene acts by inducing Naþ influx in oocytes and yeast[80]. Ion content analysis revealed that AtHKT1 presented an efficient way fordecreasing Naþ accumulation and salt tolerance in mature shoots [81]. Quantitativeproteomics analysis of the tonoplast in Mesembryanthemum crystalium plantsshowed the interaction of glycolytic enzymes aldolase and enolase along with thetonoplast via direct means of Hþ-ATPase V-ATPase, proving their role in salttolerance [82].The recent findings concerning salt tolerance enhance our knowledge of the
molecular and cellular mechanisms underlying plant adaptation, although moreinformation must be gathered to further explore the signaling pathways for salttolerance in wheat. The apoplastic protein WPM-1 provides significant informationfor the roles that these proteins have in salt tolerance. Further insights will help usgain information on the role of glycolytic enzymes, as multifunctional proteins, indifferent species. Moreover, accumulation of NO in peroxisomes shows thesignificant importance of this molecule in stress tolerance.
16.5
Heat Tolerance
Plants continuously face heat stress throughout their seasonal life cycle [83,84].Increased global temperature affects the ecology and agriculture, and may beconsidered as a major limiting factor for crop productivity. Heat stress influencescellular homeostasis, hence leading to retardation in plant growth and develop-ment. Extensive crop losses are attributed to heat in combination with otherstresses [85]. Two types of thermotolerance have been described: basal and adaptivethermotolerance. The ability of plants to survive under heat stress without priorconditioning is known as basal thermotolerance, while adjustment after exposureto extreme temperature is described as adaptive thermotolerance [86]. Ultimately,plants cope with extreme temperature by both basal and acquired thermotolerance[87]. Among the mechanisms the plants use to maintain continued homeostasis,the accumulation of HSPs under the controlled coordination of heat stresstranscription factors (heat shock factors (HSFs)) is one of utmost importance.HSPs are the major components of thermotolerance reactions and have beenproposed to act as molecular chaperones by inducing protein refolding, andpreventing protein denaturation and aggregation [88].Plant adaptation to thermotolerance not only involves HSP-based mechanisms
[89–91], but also other components like phospholipids or SOR (Pyrococcus fusiosussuperoxide), DREB2A, GSNOR, and RCA genes [1]. CBK3, an importantcomponent of the Ca2þ-regulated heat stress signal transduction pathway, wasfound to be present downstream to calmodulin and is believed to regulate HSPexpression. Several studies showed that HsfA1a is a target of CBK3 and itsphosphorylation by CBK3 targets small HSPs (sHSPs), providing thermotolerance
16.5 Heat Tolerance 445

in Arabidopsis. Similarly, the Rof1 gene has been showed to bind with HSP90.1 and,under physiological conditions, it localizes in cytoplasm. Exposure to heat stresstriggers this complex relocation to the nucleus in the vicinity of the transcriptionfactor HsfA2, which interacts with HSP90.1 and Rof1 in Arabidopsis [1]. Moreover,in different studies it was observed that HsfA3 is a crucial HSF, which is controlledby DREB2C at the transcriptional level and could lead to thermotolerance byregulating the expression of HSP-coding genes in Arabidopsis [92], while in wheatno such mechanism has yet been reported [93]. However, the overexpression ofArabidopsis HSP101 induced thermotolerance in wheat [94]. The TaSnRK 2.4protein kinase also imparts heat tolerance in wheat [95]. It has been proposed thatwheat chloroplastic sHSPs (sHSP20) were highly upregulated in response to heatstress [95]. Other reports indicated that the cytosolic sHSPs (TaHSP16.9C-1) alsoconfer tolerance to heat stress [96]. Furthermore, the HSP–chaperone complex wasalso found to function in abiotic stress tolerance, even if direct evidence of theprocess is still limited [97]. The HSP/chaperones HSP60, 70, and 90 interact with awide range of cochaperone proteins that facilitate their activity in the folding ofspecific proteins, although their roles were unraveled to a great extent in otherorganisms rather than in plants. Recent reports examined the HSP100/Clpproteins in several plant species along with wheat. These proteins are constitutivelyexpressed in plants, and their expression is regulated by environmental insults andat the developmental level [97].Apart from these genes, other transcription factors are also affected by heat stress
in wheat, although their role in heat tolerance has not been fully explored. Manycandidate genes involved in phytohormone signaling were found to be regulated byheat, such as AREB2 and AREB3, which are also involved in ABA signaling, andshown to be downregulated by heat. ABA signaling is also involved in heat shockresponse in wheat. Additional information about their exact role and downstreampathways in thermotolerance is yet to be elucidated. Interestingly, a link betweenethylene production and heat susceptibility in wheat was reported recently [98]. TheF-box containing auxin receptor (TIR) facilitates 26S proteasome-dependentdegradation of short-lived transcriptional repressors, AUX/IAA, which in turnsallows ARFs to modulate gene expression by binding itself to the promoters ofauxin-related genes and eventually giving positive responses of the auxin level toheat shock [98]. These studies have enhanced the understanding towards exploringthe molecular mechanisms regarding heat shock-mediated responses through heatshock signal transduction pathways, HSPs, and also HSFs such as HsfA1a, HsfA2,and HsfA3. Additional studies had unraveled the role of Ca2þ in heat shock signaltransduction pathways [1]. Moreover, identifying the response of the HSP–chaperone complex is considered as an important concern in providing thermo-tolerance at elevated temperatures. This could be a unique mechanism tounderstand how this complex regulates growth and development in plants underelevated temperatures.In rice and Arabidopsis, it has been reported that with a sudden rise of
temperature, plants accumulate specific phospholipids such as phosphatidylinosi-tol 4,5-bisphosphate and phosphatidic acid to specific locations, mediated by
446 16 Emerging Strategies to Face Challenges Imposed by Climate Change and Abiotic Stresses in Wheat

phosphatidylinositolphosphate kinase and phospholipase D [99]. The activation ofthe phosphatidylinositol 4,5-bisphosphate and phosphatidic acid response requiresmandatory signaling through a G-protein. This adaptation response towards heatwas found to be more or less similar to that observed during the drought response,where phospholipase Da1 facilitates ABA-induced stomatal closure throughinteractions with PP2C and G-protein [100]. Also in this case, several investigationssuggested the role of NO in heat stress tolerance [101]. In wheat, it was evidencedthat NO might act downstream of the ABA-induced proline accumulation. Probablythe mechanism behind this could be that ABA induces the formation of more ABA,which could further increase proline accumulation via the ABA transduction cascadeby complex interaction between ABA and NO [102]. Furthermore, HOT5 (5-hydroxylmethyl glutathione reductase) has been identified as an important factor that couldinteract with GSNOR, required for thermotolerance, suggesting an unexplored roleof NO in thermotolerance and plant growth [102]. GSNOR functions by regulatingnitrosylation levels by maintaining S-nitroglutathione (GSNO), an actual reservoir ofNO in the plant system. This observation shows the urgent need to emphasize themolecular mechanism that controls GSNOR activity in elucidating the overallregulation of stress caused by NO and NO-related signaling in plants [102].
16.6
Cold Tolerance
Plants with the ability to develop freezing tolerance are able to acclimate to coldduring exposure to low, but above freezing, conditions. This phenomenon of coldtolerance involves the accumulation of gene transcripts via cold perception andsignal transduction, eventually leading to promoter activation of the targeted gene.Cold acclimation is cumulative with time and involves the synthesis of specificsubsets of proteins in plants [103,104]. Most of the low-temperature-inducedproteins share common characteristics with other stress-induced proteins in plants[105]. Theses proteins can be variable, but their amino acid sequence repeats arehighly conserved. For instance, glycine is one of them, which is specifically presentat N-terminal postion whereas, lysine is specifically found at C-terminal portion ofprotein. These groups of proteins are induced under stress in order to prepare theplant for dehydration stress, although no specific functions have been assigned tothem. Upon exposure to freezing temperature, water moves from the cellprotoplasm to intercellular ice crystals, thereby causing dehydration in the cells.The stress-related proteins might be induced by one or more stimuli that may actthrough different pathways. Wcs120, a dehydrin-like protein, was found to beinduced by different abiotic stresses in wheat [106]. Moreover, the cold-regulatedtranscription activator CBF3 was also found to be linked to freezing tolerance inwheat [107]. Considerable efforts have reported that genes encoding LEA proteins,containing CRT and DRE, in their promoter regions, generally provide tolerance tocold, desiccation, and salinity. Both ABA-dependent and -independent pathwaysregulate plant exposure to cold conditions [108]. Examples have been quoted in the
16.6 Cold Tolerance 447

literature where ABA was able to activate several genes responsive to cold stress,such as the RD29A (COR78/LT178) gene. This gene contains both CRT/DRE andABA-responsive elements. Activation of RD29A can occur through binding oftranscription factors from the ERF/AP2 family [109,110]. Specifically, the CRT/DRE-binding factor CBF (1–4) recognizes the CRT/DRE and further participates inthe adaptive acclimating responses to either cold (CBF1–3) or desiccation (CBF4)response. In addition, the ecotopic expression of some CBF genes results in bothactivation of target genes, and facilitates freezing, salt, and desiccation tolerance intransgenic plants [109,110]. It has been recently reported that a MYC-typetranscription factor binds to and regulates the activity of CBF3 promoter inresponse to stress [111]. Apart from CBF factors, several studies indicated that othersignaling pathways may also be involved in the stress adaptive responses to coldtolerance [112,113]. For example, the espimo1 mutant is constitutively expressed infreezing tolerance and therefore does require signaling through cold-acclimatedCBF factors [114]. Analysis of hos9-1mutant plants did not reveal the cold inductionof any genes reported to be controlled by CBF, indicating that Hos9 regulates theexpression of genes that are important for cold tolerance, but not a part of the CBFregulon [115]. A family of transcription factors called CRT-binding factors orDREBs is found to be involved in an ABA-independent pathway and regulates CORgene expression through binding to CRT/DRE elements.Recent characterization of cbf2 mutants of Arabidopsis indicated that CBF2/
DREB1C proteins negatively regulate the CBF1/DREB1B and CBF3/DREB1Agenes, thus causing the transient expression of these transcription factors [116].Moreover, another report suggested that the expression of CBF/DREB genes canpartly be activated by ABA [117]. More interestingly, the CBF-mediated coldresponse mechanism in Arabidopsis appears to be conserved in both monocot anddicot plants such as tomato, wheat, and rye [118]. This proves that if there is aconservation of cold tolerance among monocots and dicots, wheat also follows thesame mechanism for cold tolerance. Genes for cold stress tolerance have also beenfound in wheat and its relatives, although the mechanism of their action is stillobscure. A large number of COR and LEA genes have been analyzed in cereals,including wheat [119]. A positive correlation has been observed in the accumulationof Cor/Lea genes with the levels of freezing tolerance in seedlings of commonwheat cultivars [120,121]. The low temperature response of the wheat Cor/Leagenes is also regulated through either ABA-dependent or -independent pathways.At least four of the wheat Cor/Lea genes,, wrab17, wrab18, wrab19, and wco0825,were showed to provide cold tolerance in response to exogenous ABA [121,122].Furthermore, the ABA-independent wheat Cor gene wcor15 encodes for thechloroplast-targeted Cor proteins analogous to the Arabidopsis Cor15a gene [123].The expression of other wheat Cor/Lea genes, including wcor14, wcs19, and wc120,was also found to be upregulated during cold stress in an ABA-independentmanner [120,124]. The conserved CRT/DRE-like sequence motifs were localized inthe promoter region of wcor15 and wcs120 genes, and their promoters proven to becold stress inducible in both monocot and dicot transgenic plants [123]. This studysuggested that the structure and function of the Cor/Lea gene system, including the
448 16 Emerging Strategies to Face Challenges Imposed by Climate Change and Abiotic Stresses in Wheat

CBF/DREBs trans-acting factors along with CRT/DRE elements, are highlyconserved in wheat. Recent insights proved that HOS15, a WD40-like protein,functions to control gene expression through histone deacetylation in chromatin.hos15 mutant plants accumulated higher levels of several stress-regulated genes,but proved to be hypersensitive to cold. Additionally, HOS15 was further found tointeract specifically with histone H4 and promotes deacetylation of histone H4,indicating that chromatin remodeling plays a crucial role in gene regulation andhence, tolerance to abiotic stress [124].
16.7
Functional and Comparative Genomics Approaches for Wheat Improvement
Worldwide wheat production is predicted to decline by as much as 8% due to thetremendously increasing climate changes. Therefore, to develop improved wheatvarieties, molecular genetic and functional genomics tools need to be used alongwith existing breeding techniques. Although the wheat genome is quite complexconsidering its large size (16 000 Mb), amplification of transposable elementscoupled with duplication of chromosome segments and polyploidization, differentapproaches have been developed for its improvement. At the proteomic level,functional and structural analysis of proteins is still mandatory. A unique methodfor screening protein folding and function by using a germ cell-free system wasfirst reported by Morita et al. [125]. During the same period, Mojoul et al. [126]observed the effect of high-temperature stress on the wheat endosperm proteomeby using two-dimensional electrophoresis coupled with analysis by MALDI massspectroscopy along with tandem mass spectroscopy for characterizing heat stress-induced proteins. For further enhancing proteome research on wheat, aninteractome of proteins having a role in abiotic stress tolerance was developed byusing the yeast two-hybrid GAL4 system along with specific protein interactiveassays [127]. This report emphasizes the networking of regulatory factors such asphospholipase C and GTP-binding proteins, other proteins related to flowerinitiation, and abiotic stress signaling mediated by the ABA-dependent pathway.Even more significant research has been focused on metabolomics, which includesvarious metabolic reactions, representing the dynamic changes from one conditionto another. Analysis of the amyloplast proteome in developing wheat endospermsidentified 289 proteins involved in carbohydrate metabolism, plastid division,energetics, nitrogen and sulfur metabolism, nucleic acid-related reactions, aminoacids synthesis, and isoprenoid fatty acids and related processes. This studyexplained the broad capability of amyloplasts. Moreover, the best emergingnovel and potential tool for functional genomics in the case of hexaploid wheat isRNA interference (RNAi) silencing. Yan et al. [128] published a report on thedevelopment of the first stable wheat transgenic plants using RNAi transformation.According to their observation, the reduction of VRN2 RNA by RNAi resulted in areduction of flowering time. In addition, Kong et al. [129] cloned a wheat alleleencoding a NAC transcription factor (NAM-B1) and showed that the reduction in
16.7 Functional and Comparative Genomics Approaches for Wheat Improvement 449

RNA levels of TaNAM by RNAi further lead to a delay in senescence and decreasedprotein content along with zinc and iron deficiency in wheat grains. Recentpublished studies showed that any change in the transcriptome during develop-mental processes and abiotic stresses would be useful in dissecting gene functions.For example, Wilson et al. [130] randomly cloned expressed sequence tags (ESTs)from 35 cDNA libraries belonging to different developmental stages and wheatgrains, assigning them functional annotation. Furthermore, high-density microarrayswere produced based on these libraries and, after data analysis, the authors observedthe accumulation of several mRNAs encoding various proteins involved in aminoacid biosynthesis andmetabolism, cell division, cell development, signal transductionpathways, initiation of transcription, and translation. Remarkable knowledge wasgained from the functional genomic studies using the discussed techniques, despitethe fact that the traditional transgenic approaches allow the introduction of candidategenes according to selective modifications. Increasing resistance to biotic and abioticstress, herbicide and pesticide resistance, and improved nutritional quality wouldhelp us to understand the putative function of candidate gene [131]. Using wheat as asystem for transgenic approaches has proved a good tool for the characterization ofparticular genes. Moreover, gene knockout and T-DNA insertion have been found tobe very useful options for exploring gene function in wheat.Since wheat is a heterologous polyploidy plant with highly repetitive sequences,
these attributes limited the progress in wheat genomic studies. Therefore,comparative genomics provided a better platform to study wheat genomics byusing other model plants. Arabidopsis and rice are considered as the best modelspecies for comparative genome analysis. At the DNA and protein levels, thesestudies allow the understanding of structural, functional, and evolutionary relation-ships among various species. Arabidopsis serves as a good model for all floweringplants, while rice acts as a model for cereal species. By using comparative analysis,Mullan et al. [132] identified genes in wheat that control Naþ accumulation such asHKT1 and SOS1 in Arabidopsis. The identified exons in the NHX1 and SOS1 genesof rice and wheat in comparison to Arabidopsis suggested evolutionary relationshipsbetween all three species.Recent insights identified Brachipodium distachyon (L.) as a novel model system
for cereals due to its small genome size, short life cycle, diploid accession, andsimple growth requirements [133]. Moreover, its phylogeny is relatively close towheat and barley, and therefore various genomic studies like construction of cDNAlibraries, bacterial artificial chromosome (BAC) libraries, linkage maps, andelucidation of the genome sequence are under progress [134].
16.8
Conclusion and Future Perspectives
In spite of the fact that several genes have been identified in association withclimate change, the available data is quite descriptive, describing the function ofonly a few proteins. The functional genomics studies related to climate change
450 16 Emerging Strategies to Face Challenges Imposed by Climate Change and Abiotic Stresses in Wheat

present limited information obtained on the modulation of regulatory networksand metabolic pathways associated with plant stress responses. However, appliedscience with regard to crop cultivation is still ambiguous. With the continuousproblem of food security, we ultimately require more specific solutions for cropmanagement. In this regard, the biological-based approaches would proverevolutionary and may finally generate models showing the involvement of manysignaling pathways that define plant “-omic” architectural responses in relation tochanging climate conditions. In order to develop a complete view of plant responseto climate change, so as to enhance plant tolerance to different abiotic stresses, itwould be crucial to merge the omics data with bioinformatic-based strategies andcomputational methods.Some recent breakthroughs represent a promising start [134,135]. Use of
bioinformatics analysis, Arabidopsismicroarray data, and a novel regulatory programhave been proposed [136]. Published reports revealed that this program includestranscriptional and post-transcriptional regulation, triggering the release of aminoacid metabolism in response to various abiotic stresses. Regulation of the stomatalaperture through guard cell signaling would be a real tool for candidate genemanipulations [137]. This could work as the best approach for improving droughttolerance. Weston et al. [138] provided a central approach for the use of a genomicsignature concept on how to characterize the plant stress phenotype and assign a linkto the underlying network pathways, modules, and eventually genes. Basically,modern genomic technology provides a mechanistic understanding of the processesby which genetic changes bring about phenotypic variation and result in theappearance of different physiological traits. By using these approaches, the molecularmechanism and stress-responsive pathways could be explored for the existing genesalong with their phenotypic classification under environmental stresses.At present, the tendency is to move ahead towards the system biological
approaches, which already have various models for certain genes and theirfunctions. Further development of this research is critical due to the ever-changingenvironmental conditions. To acquire a holistic view of plant stress responses, theneed to develop approaches that could use the diverse biological network along withfrequently changing climate cues is of utmost importance. Such models might bevery efficient and could be further used to acquire plant strength under fluctuatingclimates. The use of the wheat genome, coupled with functional and comparativegenomics, has provided significant progress. Since the elucidation of wheat’scomplete genome is not possible just yet, the Brachipodium genome could serve as apotential platform for the identification and characterization of untapped genes inwheat. Ultimately, this strategy will bring our aim of sustainable agriculture in termsof food, fiber, fuel, and oil to every person under climate change catastrophes.
Acknowledgments
Work on signal transduction and plant stress signaling in N.T.’s laboratory ispartially supported by the Department of Science and Technology and Department
16.8 Conclusion and Future Perspectives 451

of Biotechnology, Government of India. We would like to acknowledge Dr AncaMacovei for critical reviewing and Mr Dipesh K. Trivedi for his help withFigure 16.2.
References
1 Ahuja, I. et al. (2010) Plant molecularstress responses face climate change.Trends Plant Sci., 15, 664–674.
2 Ergen, N.Z. et al. (2009) Transcriptomepathways unique to dehydration tolerantrelatives of modern wheat. Funct. Integr.Genomics, 9, 377–396.
3 Guy, C., Porat, R., and Hurry, V. (2006)Plant cold and abiotic stress gets hot.Physiol. Plant., 126, 1–4.
4 Vasil, I.K. (2007) Molecular and geneticimprovement of cereals, transgenic wheat(Triticum aestivum L.). Plant Cell Rep.,26, 1133–1154.
5 Collins, N.C., Tardieu, F., and Tuberosa, R.(2008) Quantitative trait loci and cropperformance under abiotic stress,where do we stand? Plant Physiol.,147, 469–486.
6 Garg, B. et al. (2013) Transcript profilingidentifies novel transcripts with unknownfunctions as primary responsecomponents to osmotic stress in wheat(Triticum aestivum L.). Plant Cell TissueOrgan Cult., 113, 91–101.
7 Serrano, R. et al. (1999) A glimpse of themechanisms of ion homeostasis duringsalt stress in plants. J. Exp. Bot., 50,1023–1036.
8 Zhu, J.K. (2001) Plant salt tolerance. TrendsPlant Sci., 6, 66–71.
9 Knight, H. and Knight, M.R. (2001) Abioticstress signalling pathways, specificity andcross-talk. Trends Plant Sci., 6, 262–267.
10 Zhu, J.K. (2001b) Cell signaling under salt,water and cold stresses. Curr. Opin. Plant.Biol., 4, 401–406.
11 Zhu, J.K. (2002) Salt and drought stresssignal transduction in plants. Annu. Rev.Plant Biol., 53, 247–273.
12 Vierling, E. and Kimpel, J.A. (1992) Plantresponses to environmental stress. Curr.Opin. Biotechnol., 3, 164–170.
13 Frank, W. et al. (2000) Water deficit triggersphospholipase D activity in the
resurrection plant Craterostigmaplantagineum. Plant Cell, 12, 111–124.
14 Choi, H.I. et al. (2000) ABFs, a family ofABA-responsive element binding factors.J. Biol. Chem., 275, 1723–1730.
15 Shinozaki, K. and Yamaguchi-Shinozaki,K. (2000) Molecular responses todehydration and low temperature,differences and cross-talk between twostress signalling pathways. Curr. Opin.Plant Biol., 3, 217–223.
16 Ingram, J. and Bartels, D. (1996) Themolecular basis of dehydration tolerance inplants. Annu. Rev Plant Biol., 47, 377–403.
17 Bray, E.A., et al. (2000) Responses to abioticstresses, in Biochemistry and MolecularBiology of Plants (eds W. Gruissem, B.Buchannan, and R. Jones), AmericanSociety of Plant Physiologists, Rockville,MD, pp. 1158–1249.
18 Bohnert, H.J. and Sheveleva, E. (1998)Plant stress adaptations makingmetabolism move. Curr. Opin. Plant Biol.,1, 267–274.
19 Blumwald, E. (2000) Sodium transport andsalt tolerance in plants. Curr. Opin. CellBiol., 12, 431–434.
20 Chae, L. et al. (2009) Diversetranscriptional programs associated withenvironmental stress and hormones in theArabidopsis receptor like kinase genefamily.Mol. Plant, 2, 84–107.
21 Shulaev, V. et al. (2008) Metabolomics forplant stress response. Physiol. Plant.,132, 199–208.
22 Niu, C.F. et al. (2012) Wheat WRKYgenesTaWRKY2 and TaWRKY19 regulate abioticstress tolerance in transgenic Arabidopsisplants. Plant Cell Environ., 35, 1156–1170.
23 Kobayashi, F. et al. (2008) Development ofabiotic stress tolerance via bZIP-typetranscription factor LIP19 in commonwheat. J. Exp. Bot., 59, 891–905.
24 Liu, F. et al. (2012) Wheat TaRab7 GTPaseis part of the signaling pathway in
452 16 Emerging Strategies to Face Challenges Imposed by Climate Change and Abiotic Stresses in Wheat

responses to stripe rust and abiotic stimuli.PLoS ONE, 7, e37146.
25 Kuzuoglu, O.D. et al. (2012) Autophagy-related gene, TdAtg8, in wild emmer wheatplays a role in drought and osmotic stressresponse. Planta, 236, 1081–1092.
26 Chauhan, H. et al. (2012) The wheatchloroplastic small heat shock protein(sHSP26) is involved in seed maturationand germination and impartstolerance to heat stress. Plant Cell Environ.,35, 1912–1931.
27 Gouiaa, S. et al. (2012) Expression of wheatNaþ/Hþ antiporter TNHXS1 and Hþ-pyrophosphatase TVP1 genes in tobaccofrom a bicistronic transcriptional unitimproves salt tolerance. Plant Mol. Biol.,79, 137–155.
28 Han, Y. et al. (2012) Characterization of awheat (Triticum aestivum L.) expansin gene,TaEXPB23, involved in the abiotic stressresponse and phytohormone regulation.Plant Physiol. Biochem., 54, 49–58.
29 Qin, Y. et al. (2012) Over-expression ofTaMYB33 encoding a novel wheat MYBtranscription factor increases salt anddrought tolerance in Arabidopsis.Mol. Biol.Rep., 39, 7183–7192.
30 He, Y. et al. (2012) Evidence supporting arole of jasmonic acid in Arabidopsis leafsenescence. Plant Physiol., 128, 876–884.
31 Mao, X. et al. (2011) Transgenic expressionof TaMYB2A confers enhanced tolerance tomultiple abiotic stresses in Arabidopsis.Funct. Integr. Genomics, 11, 445–465.
32 Guerra, D. et al. (2012) Identification of aprotein network interacting with tdrf1, awheat ring ubiquitin ligase with aprotective role against cellular dehydration.Plant Physiol., 158, 777–789.
33 Ayadi, M. et al. (2011) Identification andcharacterization of two plasma membraneaquaporins in durum wheat (Triticumturgidum L. subsp. durum) and their rolein abiotic stress tolerance. Plant Physiol.Biochem., 49, 1029–1039.
34 Xuea, G.P. et al. (2011) Overexpression ofTaNAC69 leads to enhanced transcriptlevels of stress up-regulated genes anddehydration tolerance in bread wheat.Mol. Plant, 4, 697–712.
35 Zhang, H. et al. (2012) Overexpression of acommon wheat gene TaSnRK2.8 enhances
tolerance to drought, salt and lowtemperature in Arabidopsis. PLoS ONE,12, e16041.
36 Wang, X. et al. (2013) TaAbc1, a member ofAbc1-like family involved in hypersensitiveresponse against the stripe rust fungalpathogen in wheat. PLoS ONE, 8, e58969.
37 Seo, P.J. et al. (2009) TheMYB96transcription factor mediates abscisic acidsignaling during drought stress responsein Arabidopsis. Plant Physiol., 151, 275–289.
38 Seki, M. et al. (2003) Molecular responsesto drought, salinity and frost, common anddifferent paths for plant protection. Curr.Opin. Plant Biol., 14, 194–199.
39 Hong, Y. et al. (2008) Dual functions ofphospholipase Da1 in plant response todrought.Mol. Plant, 1, 262–269.
40 Wilson, P.B. et al. (2009) The nucleotidase/phosphatase SAL1 is a negative regulatorof drought tolerance in Arabidopsis. PlantJ., 58, 299–317.
41 Yamaguchi-Shinozaki, K. and Shinozaki,K. (2005) Organization of cis-actingregulatory elements in osmotic- and cold-stress-responsive promoters. Trends PlantSci., 10, 88–94.
42 Chaves, M.M., Maroco, J.P., and Pereira,J.S. (2003) Understanding plant responsesto drought from genes to whole plant.Funct. Plant Biol., 30, 239–264.
43 Cuiling, L. et al. (2010) TaCHP, a wheatzinc finger protein gene down-regulated byabscisic acid and salinity stress plays apositive role in stress tolerance. PlantPhysiol., 154, 211–221.
44 Jang, C.S. et al. (2002) Isolation andcharacterization of lipid transfer protein(LTP) genes from a wheat–ryetranslocation line. Plant Cell Rep.,20, 961–966.
45 Millward, T.A., Zolnierowicz, S., andHemmings, B.A. (1999) Regulation ofprotein kinase cascades by proteinphosphatase 2A. Trends Biochem. Sci.,24, 186–191.
46 Chongyi, X. et al. (2006) A wheat(Triticum aestivum) protein phosphatase 2Acatalytic subunit gene provides enhanceddrought tolerance in tobacco. Ann. Bot.,99, 439–450.
47 Pernas, M. et al. (2007) A proteinphosphatase 2A catalytic subunit is a
References 453

negative regulator of abscisic acidsignalling. Plant J., 51, 763–768.
48 Mao, X. et al. (2010) TaSnRK2.4, an SNF1-type serine/threonine protein kinase ofwheat (Triticum aestivum L.), confersenhanced multistress tolerance inArabidopsis. J. Exp. Bot., 61, 683–693.
49 Anderberg, R.J. and Walker-Simmons, M.K. (1992) Isolation of a wheat cDNA clonefor an abscisic acid-inducible transcriptwith homology to protein kinases. Proc.Natl. Acad. Sci. USA, 89, 10183–10187.
50 Kurashashi, Y., Terashima, A., and Takumi,S. (2009) Variation in dehydrationtolerance, ABA sensitivity and related geneexpression patterns in D-genomeprogenitor and synthetic hexaploid wheatlines. Int. J. Mol. Sci., 10, 2733–2751.
51 Chen, R. et al. (2005) Isolation andcharacterization of genes encoding mybtranscription factors in wheat (Triticumaestivum L.). China Sci. J., 169, 1146–1154.
52 Xia, N. et al. (2010) Characterization of anovel wheat NAC transcription factor geneinvolved in defense response against striperust pathogen infection and abioticstresses.Mol. Biol., 37, 3703–3712.
53 Xia, N. et al. (2010) TaNAC8, a novel NACtranscription factor gene in wheat,responds to stripe rust pathogen infectionand abiotic stresses. Physiol. Mol. PlantPathol., 74, 394–402.
54 Mao, X. et al. (2012) TaNAC2, a NAC-typewheat transcription factor conferringenhanced multiple abiotic stress tolerancesin Arabidopsis. J. Exp. Bot., 63, 2933–2946.
55 Luna, C.M. et al. (2005) Drought controlson H2O2 accumulation, catalase (CAT) andCATgene expression in wheat. J. Exp. Bot.,56, 417–423.
56 Zhang, Y. et al. (2011) Cloning andprokaryotic expression of TaE3 from wheatand preparation of antiserum. Front. Agric.China, 5, 437–442.
57 Kobayashi, F. et al. (2008) Development ofabiotic stress tolerance via bZIP-typetranscription factor LIP19 in commonwheat. J. Exp. Bot., 59, 891–905.
58 Cho, S.K. et al. (2008) Arabidopsis PUB22and PUB23 are homologous U-box E3ubiquitin ligases that play combinatoryroles in response to drought stress. PlantCell, 20, 1899–1914.
59 Ning, J. et al. (2010) A Raf like MAPKKKgene DSM1 mediates drought resistancethrough reactive oxygen speciesscavenging in rice. Plant Physiol., 152,876–890.
60 Wang, X. et al. (2009) Expression andresponses to dehydration and salinitystresses of V-PPase gene membersin wheat. J. Genet. Genomics, 36,711–720.
61 Zhou, H. et al. (2007) Molecular analysis ofthree new receptor like kinases genes fromhexaploid wheat and evidence forparticipation in wheat hypersensitiveresponses to strip rust fungus. Plant J.,52, 420–434.
62 Fortunati, A. et al. (2008) Isopreneemission is not temperature dependentduring and after severe drought-stress, aphysiological and biochemical analysis.Plant J., 55, 687–697.
63 Caruso, G. et al. (2009) Analysis of droughtresponsive proteins in wheat (Triticumdurum) by 2D-PAGE and MALDI-TOFmass spectrometry. Plant Sci.,177, 570–576.
64 Bini, C. et al. (2000) Fertility gain andheavy metal accumulation in plants andsoil, studied by means of compostamended cultivation of Taraxacumofficinale. Toxicol. Environm. Chem.,77, 131–142.
65 Mahajan, S. and Tuteja, N. (2005)Cold, salinity and drought stress, anoverview. Arch. Biochem. Biophys.,444, 139–158.
66 Rodriguez-Milla, M.A. and Salinas, J.(2009) Prefoldins 3 and 5 play an essentialrole in Arabidopsis tolerance to salt stress.Mol. Plant, 2, 526–534.
67 Sakamoto, H. et al. (2008) ITN1, a novelgene encoding an ankyrin repeat proteinthat affects the ABA-mediated productionof reactive oxygen species and is involvedin salt-stress tolerance in Arabidopsisthaliana. Plant J., 56, 411–422.
68 Corpas, F.J. et al. (2009) Peroxisomes arerequired for in vivo nitric oxideaccumulation in the cytosol followingsalinity stress of Arabidopsis plants. PlantPhysiol., 15, 2083–2094.
69 Møller, I.S. et al. (2009) Shoot Naþ
exclusion and increased salinity tolerance
454 16 Emerging Strategies to Face Challenges Imposed by Climate Change and Abiotic Stresses in Wheat

engineered by cell type-specific alterationof Naþ transport in Arabidopsis. Plant Cell,21, 2163–2178.
70 Fujii, H. and Zhu, J.K. (2009) Anautophosphorylation site of the proteinkinase SOS2 is important for salttolerance in Arabidopsis.Mol. Plant,2, 183–190.
71 Kosova, K., Prasil, I.T., and Vitamvas, P.(2013) Protein contribution to plantsalinity response and tolerance acquisition.Int. J. Mol. Sci., 14, 6757–6789.
72 Rodriguez, A.A. et al. (2009) Polyamineoxidase activity contributes to sustainmaize leaf elongation under saline stress.J. Exp. Bot., 60, 4249–4262.
73 Zaidi, I. et al. (2010) TMKP1, is a novelwheat stress responsive MAP kinasephosphatase localized in the nucleus. PlantMol. Biol., 73, 325–338.
74 LoPez-Perez, L., Martnez-Ballesta, M.C.,and Maurel, C. (2009) Changes in plasmamembrane lipids, aquaporins and protonpump of broccoli roots, as an adaptationmechanism to salinity. Phytochemistry,70, 492–500.
75 Hu, W. et al. (2010) Overexpression ofwheat aquaporins to TaAQP8 enhances salttolerance in transgenic tobacco. Plant CellPhysiol., 53, 2127–2141.
76 Swami, A.K. et al. (2011) Differentialproteomic analysis of salt stress responsein sorghum bicolor leaves. Environ. Exp.Bot., 71, 321–328.
77 Rus, A. et al. (2001) AtHKT is a salttolerance determinant that controls Naþ
entry in to plant roots. Proc. Natl. Acad. Sci.USA, 98, 14150–14155.
78 Uozumi, N. et al. (2000) The ArabidopsisHKT, gene homologue mediates inwardNaþ currents in Xenopus laevis oocytesand Naþ uptake in Saccharomycescerevisiae. Plant Physiol., 122,1249–1260.
79 Møller, I.S. et al. (2009) Shoot Naþ
exclusion and increased salinity toleranceengineered by cell type-specific alterationof Naþ transport in Arabidopsis. Plant Cell,21, 2163–2178.
80 Barkla, B.J. et al. (2009) Quantitativeproteomics of the tonoplast reveals a rolefor glycolytic enzymes in salt tolerance.Plant Cell., 21, 4044–4058.
81 Sarkar, N. et al. (2009) Rice sHsp genes,genomic organization and expressionprofiling under stress and development.BMC Genomics., 10, 393.
82 Perez, D.E. et al. (2009) BOBBER1 is anoncanonical Arabidopsis small heatshock protein required for bothdevelopment and thermotolerance. PlantPhysiol., 151, 241–252.
83 Mittler, R. (2006) Abiotic stress, the fieldenvironment and stress combination.Trends Plant Sci., 11, 15–19.
84 Chen, J. et al. (2006) FtsH11 protease playsa critical role in Arabidopsisthermotolerance. Plant J., 48, 73–84.
85 Qin, D. et al. (2008) Heat stress-responsivetranscriptome analysis in heat susceptibleand tolerant wheat (Triticum aestivum L.) byusing Wheat Genome Array. BMCGenomics, 9, 432.
86 Kotak, S. et al. (2007) Complexity of heatstress response in plants. Curr. Opin. PlantBiol., 10, 310–316.
87 Meiri, D. and Breiman, A. (2009)Arabidopsis ROF1 (FKBP62) modulatesthermotolerance by interacting withHSP90.1 and affecting the accumulationof HsfA2-regulated sHSPs. Plant J.,59, 387–399.
88 Ginzberg, I. et al. (2009) Transcriptomicprofiling of heat-stress response inpotato periderm. J. Exp. Bot., 60,4411–4421.
89 Lim, C.J. et al. (2007) Overexpression ofthe Arabidopsis DRE/CRT bindingtranscription factor DREB2C enhancesthermotolerance. Biochem. Biophys. Res.Commun., 362, 431–436.
90 Jackson, S. and Chen, Z.J. (2010)Genome and expression plasticity ofpolyploidy. Curr. Opin. Plant Biol.,13, 153–159.
91 Campbell, J.L. et al. (2001) Cloning of newmembers of heat shock proteins HSP101gene family in wheat (Triticum aestivum (L.)Moench) inducible by heat, dehydrationand ABA. Biochim. Biophys. Acta,1517, 270–277.
92 Chauhan, H. et al. (2012) The wheatchloroplastic small heat shock protein(sHSP26) is involved in seed germinationand imparts tolerance to heat stress. PlantCell Environ., 35, 1912–1931.
References 455

93 Basha, E. et al. (2004) Chaperon activity ofcytosolic small heat shock proteins fromwheat. Eur. J. Biochem., 271, 1426–1430.
94 Wang, W. et al. (2004) Role of plant heatshock proteins and molecular chaperons inabiotic stress response. Trends Plant Sci.,9, 244–252.
95 Zhang, H. et al. (2010) Overexpression of acommon wheat gene TaSnRK2.8 enhancestolerance to drought, salt and lowtemperature in Arabidopsis. PLoS ONE,5, e16041.
96 Qin, F. et al. (2008) Arabidopsis DREB2A-interacting proteins function as RING E3ligases and negatively regulate plantdrought stress-responsive gene expression.Plant Cell., 20, 1693–1707.
97 Mishkind, M. et al. (2009) Heat stressactivates phospholipase D and triggers PIP2accumulation at the plasma membrane andnucleus. Plant J., 60, 10–21.
98 Mishra, G. et al. (2006) A bifurcatingpathway directs abscisic acid effects onstomatal closure and opening inArabidopsis. Science, 312, 264–266.
99 Siddiqui, M.H., Al-Whaibi, M.H., andBasalah, M.O. (2011) Role of nitric oxide intolerance of plants to abiotic stress.Protoplasma, 242, 447–455.
100 Wang, P. and Song, C.P. (2008) Guardcell signaling for hydrogen peroxideand abscisic acid. New Phytol.,178, 703–718.
101 Lee, U. et al. (2008) Modulation ofnitrosative stress by S-nitrosoglutathionereductase is critical for thermotoleranceand plant growth in Arabidopsis. Plant Cell.20, 786–802.
102 Guy, C.L. (1990) Cold acclimation andfreezing stress tolerance: role of proteinmetabolism. Annu. Rev. Plant Physiol. PlantMol. Biol., 41, 187–223.
103 Houde, M. et al. (1992) Cloning,characterization and expression of a cDNAencoding a 50-kilodalton proteinspecifically induced by cold acclimation inwheat. Plant Physiol., 99, 1381–1387.
104 Nordin, R., Vahela, T., and Palva, E.T.(1993) Differential expression of tworelated, low-temperature-induced genes inArabidopsis thaliana (L.) Heynh. Plant Mol.Biol., 26, 641–653.
105 Limin, A.E. et al. (1997) Chromosomemapping of low temperature induced
wcs120 family genes and regulation of coldtolerance experience in wheat.Mol. Gen.Genet., 253, 720–727.
106 Vagujfalvi, A. et al. (2003) The cold-regulated transcriptional activator cbf3 islinked to the frost-tolerance locus fr-A2 onwheat chromosome 5A.Mol. Genet.Genomics., 209, 60–67.
107 Shinozaki, K., Yamaguchi-Shinozaki, K.,and Seki, M. (2003) Regulatory network ofgene expression in drought and coldstress response. Curr. Opin. Plant Biol.,6, 410–417.
108 Jaglo-Ottosen, K.R. et al. (1998) ArabidopsisCBF1 overexpression induces COR genesand enhances freezing tolerance. Science,280, 104–106.
109 Kasuga, M. et al. (1999) Improving plantdrought, salt, and freezing tolerance bygene transfer of a single stress inducibletranscription factor. Nat. Biotechnol.,17, 287–291.
110 Chinnusamy, V., Schumaker, K., and Zhu,J.K. (2004) Molecular genetic prospectiveon crosstalk and specificity in abioticstress signaling in plants. J. Exp. Bot.,155, 225–236.
111 Fowler, S. and Thomashow, M.F.(2002) Arabidopsis transcriptome profilingindicates that multiple regulatorypathways are activated during coldacclimation in addition to the CBFcold response pathway. Plant Cell,14, 1675–1690.
112 Kreps, J.A. et al. (2002) Transcriptomechanges for Arabidopsis in response to salt,osmotic and cold stress. Plant Physiol.,130, 2129–2141.
113 Xin, Z. and Browse, J. (1998) Eskimomutants of Arabidopsis are constitutivelyfreezing tolerant. Proc. Natl. Acad. Sci.USA, 95, 7799–7804.
114 Zhu, J.K., Hasegawa, P.M., and Bressan,R.A. (2004) An Arabidopsis homeodomaintranscription factor gene,HOS9, mediatescold tolerance through a CBF-independentpathway. Proc. Natl. Acad. Sci. USA,101, 9873–9878.
115 Novillo, F. et al. (2004) CBF2/DREB1C is anegative regulator of CBF1/DREB1B andCBF3/DREB1A expression and plays acentral role in stress tolerance inArabidopsis. Proc. Natl. Acad. Sci. USA,101, 3985–3990.
456 16 Emerging Strategies to Face Challenges Imposed by Climate Change and Abiotic Stresses in Wheat

116 Knight, H. et al. (2004) Abscisic acidinduces CBF gene transcription andsubsequent induction of cold-regulatedgenes via the CRT promoter element. PlantPhysiol., 135, 1710–1717.
117 Jaglo, K.R. et al. (2001) Components of theArabidopsis C-repeat/dehydrationresponsive element binding factor cold-responsive pathway are conserved inBrassica napus and other plant species.Plant Physiol., 127, 910–917.
118 Cattivelli, L. and Bartels, D. (1990)Molecular cloning and characterization ofcold-regulated genes in barley. PlantPhysiol., 93, 1504–1510.
119 O hno, R. , Tak umi, S . , a n d N aka mura, C.(2001) Ex pression of a c old-responsive Lt - C o rgene and development of fr eezing toleranceduring cold acclim ation in wheat (Triticumaestivum L. ). J. Exp. Bot., 52, 2367–2374.
120 Kobayashi, F., Takumi, S., Kume, S.,Ishibashi, M., Ohno, R., Expressionpro files of wheat CBF and Cor/Lea genes197 Murai, K. and Nakamura, C. (2005)Regulation by Vrn-1/Fr - 1 chromosomalintervals of CBF-mediated Cor/Lea geneexpression and freezing tolerance incommon wheat. J. Exp. Bot., 56, 887–895.
121 Tsuda, K. et al. (2000) New members of acold-responsive group-3. Lea/Rab-relatedCor gene family from common wheat( Triticum aestivum L.). Gene. Genet. Syst.,75 , 179–188.
122 Takumi, S et al. (2003) Cold-speci fic andlight-stimulated expression of a wheat( Triticum aestivum L.) Cor gene Wcor15encoding a chloroplast-targeted protein.J. Exp. Bot., 54, 2265–2274.
123 Tsvetanov, S. et al. (2000) A cold-responsivewheat (Triticum aestivum L.) gene wcor14identified in a winter-hardy cultivar‘Mironovska 808’. Genes Genet. Syst.,75 , 49–57.
124 Zhu, J. et al. (2008) Involvement ofArabidopsis HOS15 in histonedeacetylation and cold tolerance. Proc. Natl.Acad. Sci. USA, 15, 4945–4950.
125 Morita, E.H. et al. (2003) A wheat germcell-free system is a novel way to screenprotein folding and function. Protein Sci.,12 , 1216–1221.
126 Majoul, T. et al. (2003) Proteomic analysisof the effect of heat stress on hexaploidwheat grain. Characterization of heat-
responsive proteins from total endosperm.Proteomics, 3 , 175–183.
127 Tardif, G. et al. (2007) Interaction networkof proteins associated abiotic stress anddevelopment in wheat. Plant Mol. Biol.,63 , 703–718.
128 Yan, L. et al. (2004) Regulation of VRN-1vernalization genes in normal andtransgenic polyploid wheat. Science,303, 1640–1644.
129 Kong, F.J., Oyanagi, A., and Komatsu, S.(2010) Cell wall proteome of wheat rootsunder flooding stress using gel-based andLC MS/MS-based proteomics approaches.Biochim. Biophys. Acta, 1804, 124–136.
130 Wilson, I.D. et al. (2005) Alteration of theembryo transcriptome of hexaploid winterwheat (Triticum aestivum cv. Mercia) duringmaturation and germination. Funct. Integr.Genomics, 5, 144–154.
131 Dunca, R.R. and Carrow, R.N. (1999)Turfgrass molecular genetic improvementfor abiotic/edaphic stress resistance. Adv.Agron., 67, 233–305.
132 Mullan, D.J. et al. (2007) Arabidopsis–rice –wheat gene orthologues for Naþ
transport and transcript analysis in wheat–L. elongatum aneuploids under salt stress.Mol. Genet. Genomics, 277, 199–212.
133 Ozdemir, B.S. et al. (2008) Brachypodiumgenomics. Int. J. Plant Genomics,2008, 536104.
134 Vogel, J. and Hill, T. (2008) High-efficiencyagrobacterium-mediated transformationof Brachypodium distachyon inbred lineBd21-3. Plant Cell Rep., 27, 471–478.
135 Shulaev, V. et al. (2008) Metabolomics forplant stress response. Physiol. Plant.,132, 199–208.
136 Carnegie, I. (2010) Gene functiondiscovery: new computation modelpredicts gene function. Science Daily, 1February, http://www.sciencedaily.com/releases/2010/01/100131142436.htm.
137 Less, H. and Galili, G. (2008) Principaltranscriptional programs regulatingplant amino acid metabolism inresponse to abiotic stresses. Plant Physiol.,147, 316–330.
138 Weston, D.J. et al. (2008) Connectinggenes, co-expression modules, andmolecular signature to environmentalstress phenotypes in plants. BMC Syst.Biol., 2 (16). doi: 10.1186/1752-0509-216.
References 457





























