Embed Size (px)
Citation preview

27
Metabolome Analyses for Understanding Abiotic Stress
Responses in Plants to Evolve Management Strategies
Usha Chakraborty, Bhumika Pradhan, and Rohini Lama
Abstract
Abiotic stress responses are of the utmost importance for plants because theycannot survive unless they are able to cope with environmental changes, such ashigh and low temperatures, drought, flooding, salinity, freezing, change in pH,strong light, UV, and heavy metals. Plants respond to various stresses at differentlevels, including molecular and cellular levels, as well as by modifying theirmetabolomes. Hence, studies on plant responses to stresses can be conducted atany of these levels to provide an understanding of the mechanisms involved. Thepresent chapter focuses on the metabolomic approach to understand the responsesof plants to different abiotic stresses, which can then be utilized to evolve strategiesto combat such stress. Osmoprotectant metabolites, such as proline, glycinebetaine, and polyamines, as well as carbohydrates, play important roles in theprotection of plants against osmotic disbalances due to abiotic stresses. In addition,oxidative stresses are also overcome by an array of antioxidants, such as phenols,ascorbate, carotenoids, and a-tocopherol, as well as antioxidative enzymes.Signaling cascades activated during abiotic stresses lead to overexpression ofprotein kinases and stress proteins, and also involve molecules such as jasmonicacid and salicylic acid. Protein kinases and protein phosphatases that are encodedby large gene families often act in tandem to perform the phosphorylationand dephosphorylation leading to their activation and inactivation involved instress signaling in plants. Analysis of microRNAs and transcriptomes hasprovided sufficient understanding of the gene expression levels during periodsof stress. Hence, taken together, all these results can be utilized for identifyinggenes and/or metabolites overexpressed in tolerant species during periods ofstress, and can be utilized to achieve higher tolerance and survivability duringstresses.
727
Climate Change and Plant Abiotic Stress Tolerance, First Edition. Edited by Narendra Tuteja and Sarvajeet S. Gill.� 2014 Wiley-VCH Verlag GmbH & Co. KGaA. Published 2014 by Wiley-VCH Verlag GmbH & Co. KGaA.

27.1
Introduction
Abiotic stress responses are of the utmost importance for plants because theycannot survive unless they are able to cope with environmental changes, such ashigh and low temperatures, drought, flooding, salinity, freezing, change in pH,strong light, UV, and heavy metals. Due to increases in urbanization, deforestation,and other anthropogenic activities, plants face threats for their survival in terms ofavailability of quality water, proper growing space, soil pH, and other favorableenvironmental conditions for growth. It is therefore necessary in the coming yearsto grow plants (which include all forest plants, and economically importantmedicinal and crop plants) that show maximum tolerance to all or most of theabove environmental conditions. The tolerant plants must have the followingimportant features to survive the above challenges: low water-use efficiency, fastgrowth, high productivity, tolerance to changes in the climatic conditions, and so on.Plants can show various strategies, such as stress avoidance and stress tolerance, tocombat the above climatic changes. Stress avoidance includes various protectivemechanisms that delay or prevent the negative impact of a stress factor on a plantthat are stable and inherited, whereas stress tolerance is the ability of a plant toacclimate to a stressful condition, and acclimation is plastic and reversible [1]. Whenthe adverse climatic condition no longer prevails, the induced physiological changesor modifications during acclimation, which are diverse, are usually lost.Water, temperature, and other stresses lead to metabolic toxicity in the plant,
disorganization of the cell membrane system, production of reactive oxygen species(ROS), photosynthetic inhibition, and alteration in the acquisition of nutrients [2].Mild abiotic stresses have been found to be tolerated by plants by alterations inmolecular mechanisms ensuring plant survival and reproduction, as evident by theplants’ ability to evolve and rearrange their genomic expression in response tochanges in their environment, although survival under such stressful conditionsrequires expenditure of energy and metabolism. The adverse effect of abioticstresses is reflected on plant health, habitus, and production, and it is estimated thatabiotic stresses account for 70% reduction in the yield for the major crops [3].Various tolerance mechanisms have been studied and continuous research isongoing at every level to study the different tolerances achieved by plants at thecellular, molecular, metabolomic and genomic level. Mechanisms that operate insignal perception, transduction, and downstream regulatory factors are now beingexamined, and an understanding of cellular pathways involved in abiotic stressresponses provide valuable information on such responses. At the molecular level,it has been suggested that abiotic stress tolerance can be achieved through genetransfer by altering the accumulation of osmoprotectants, production of chaper-ones, superoxide radical-scavenging mechanisms, and exclusion or compartmenta-lization of ions by efficient transporter and symporter systems (see reviews [2,4–9]).In the postgenomic era, comprehensive analyses using three systematic
approaches or “omics” have increased our understanding of the complex molecularregulatory networks associated with stress adaptation and tolerance: (i) “transcrip-
728 27 Metabolome Analyses for Understanding Abiotic Stress Responses in Plants

tomics” for the analysis of coding and non-coding RNAs, and their expressionprofiles, (ii) “metabolomics,” which is a powerful tool to analyze a large number ofmetabolites, and (iii) “proteomics” in which protein and protein modificationprofiles offer an unprecedented understanding of regulatory networks. Proteincomplexes involved in signaling have been analyzed by a proteomics approach[10,11]. The first step in switching on any molecular response is the perception ofstress and then to relay information about it through a signal transduction pathway.To perceive the initial stress signal, a sensor molecule is required that initiates (orsuppresses) a cascade to transmit the signal intracellularly and, in many cases,activate nuclear transcription factors to induce the expression of specific sets ofgenes [12].
27.2
Metabolite Changes During Abiotic Stresses
In this chapter, we focus on the metabolomics of plant responses to various abioticstresses like water stress (drought, flooding), salt (osmotic or ionic) stress, extremetemperatures (high, low, chilling, and freezing), and the involvement of antioxidantmechanisms. Plants show various alterations or changes in their metabolism,which include the production of stress metabolites also termed osmolytes orcompatible solutes (e.g., proline, soluble carbohydrates, starch, glycine betaine,raffinose, and polyamines), that are able to stabilize proteins and cellular structuresand/or to maintain cell turgor by osmotic adjustment, redox metabolism to removeexcess levels of ROS and re-establish the cellular redox balance, as well as stresshormones and kinases during stress perception and tolerance [9,13–15].The advent of various sophisticated technologies such as mass spectroscopy and
bioinformatics has made metabolite profiling along with transcriptome analysispossible, and thus we are now able to pinpoint the exact points of metabolicadjustments during various stresses such as water deficit (dehydration and highsalinity) and extreme temperature (cold and heat), and analyze the final steps ofabiotic stress signal transduction pathways comprehensively [16].
27.2.1
Proline and Glycine Betaine
The two major organic osmolytes glycine betaine and proline accumulate in avariety of plant species during various environmental stresses. The accumulation ofthe amino acid proline in tissues of several plant species is usually regarded as aresponse of the plant tissues to water and other kinds of stresses [17–20]. Theprotective role of proline, especially against damage by drought, high salinity, andheavy metals, as has been reported by several authors [21–24], may be due to itsability to act as an osmolyte, a ROS scavenger, and a molecular chaperonestabilizing the structure of proteins, thereby protecting cells from damage causedby stress [1].
27.2 Metabolite Changes During Abiotic Stresses 729
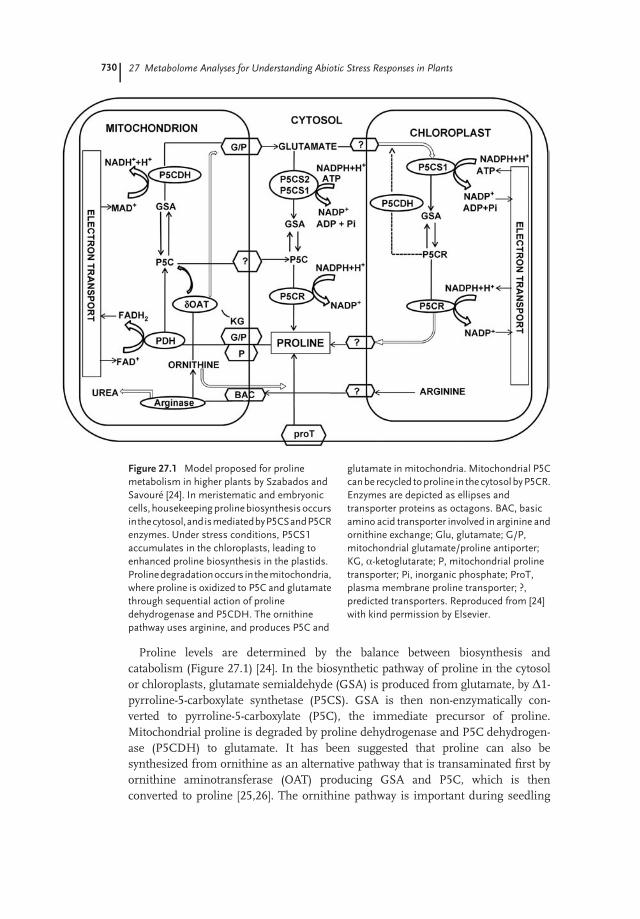
Proline levels are determined by the balance between biosynthesis andcatabolism (Figure 27.1) [24]. In the biosynthetic pathway of proline in the cytosolor chloroplasts, glutamate semialdehyde (GSA) is produced from glutamate, by D1-pyrroline-5-carboxylate synthetase (P5CS). GSA is then non-enzymatically con-verted to pyrroline-5-carboxylate (P5C), the immediate precursor of proline.Mitochondrial proline is degraded by proline dehydrogenase and P5C dehydrogen-ase (P5CDH) to glutamate. It has been suggested that proline can also besynthesized from ornithine as an alternative pathway that is transaminated first byornithine aminotransferase (OAT) producing GSA and P5C, which is thenconverted to proline [25,26]. The ornithine pathway is important during seedling
Figure 27.1 Model proposed for proline
metabolism in higher plants by Szabados and
Savour�e [24]. In meristematic and embryonic
cells, housekeepingprolinebiosynthesisoccurs
inthecytosol,andismediatedbyP5CSandP5CR
enzymes. Under stress conditions, P5CS1
accumulates in the chloroplasts, leading to
enhanced proline biosynthesis in the plastids.
Prolinedegradationoccurs in themitochondria,
where proline is oxidized to P5C and glutamate
through sequential action of proline
dehydrogenase and P5CDH. The ornithine
pathway uses arginine, and produces P5C and
glutamate in mitochondria. Mitochondrial P5C
canbe recycled toproline in thecytosolbyP5CR.
Enzymes are depicted as ellipses and
transporter proteins as octagons. BAC, basic
amino acid transporter involved in arginine and
ornithine exchange; Glu, glutamate; G/P,
mitochondrial glutamate/proline antiporter;
KG, a-ketoglutarate; P, mitochondrial proline
transporter; Pi, inorganic phosphate; ProT,
plasma membrane proline transporter; ?,
predicted transporters. Reproduced from [24]
with kind permission by Elsevier.
730 27 Metabolome Analyses for Understanding Abiotic Stress Responses in Plants

development and in some plants for stress-induced proline accumulation.However, quite recently the significance of this pathway and OAT in prolinebiosynthesis has been questioned, because proline levels were not affected inArabidopsis oat knockout mutants [24]. Funck et al. [27] have suggested that OATfacilitates nitrogen recycling from arginine through P5C, which is converted toglutamate by P5CDH.It is not clear whether there exists a direct correlation between the accumulation
of proline and abiotic stress tolerance; however, several examples show there existsa relation. It has been shown that high proline levels can be characteristic of saltand cold-hypersensitive Arabidopsis (Arabidopsis thaliana) mutants [28,29]. Accord-ing to Choudhary et al. [30], proline content is also high in drought-tolerant ricevarieties, but is not correlated with salt tolerance in barley (Hordeum vulgare)[31,32]. Several workers have demonstrated that the metabolism of proline in plantsunder different kinds of stresses is complex, and the accumulation is veryimportant to impart tolerance towards the external environmental conditions withrespect to growth, development, and stress responses of the plant [33–36].Stress conditions stimulate proline synthesis, while proline catabolism is
enhanced during recovery from stress. Overexpression of P5CS in tobacco andpetunia led to increased proline accumulation, and enhanced salt and droughttolerance [33,37], whereas Arabidopsis P5CS1 knockout plants were impaired instress-induced proline synthesis and were hypersensitive to salinity [35]. Consis-tently, proline dehydrogenase antisense Arabidopsis accumulated more proline, andshowed enhanced tolerance to freezing and high salinity [38]. It has also beensuggested that, in an alternative pathway, mitochondrial P5C can be produced byd-OAT from ornithine [36]. Overexpression of Arabidopsis d-OAT has been shownto enhance proline levels, and to increase the stress tolerance of rice, cotton, andtobacco [26,39,40], even though Arabidopsis plants deficient in d-OAT accumulatedproline in response to stress and showed a salt stress tolerance similar to the wild-type [27]. Thus, it is clear that proline has multiple roles in plant metabolism, bothduring the normal state as well as during stress (Figure 27.2).An amino acid derivative by nature, glycine betaine is naturally synthesized in
many plants, with some exceptions in crop species like potato or tomato, which areunable to accumulate glycine betaine [41]. It is a quaternary ammonium compoundthat occurs abundantly in response to dehydration stress [42–45]. In chloroplasts,where glycine betaine is found in abundance, it plays a vital role in the adjustmentand protection of the thylakoid membrane and maintains the efficiency ofphotosynthesis [46,47]. In chloroplasts, serine is converted to ethanolamine, whichgives rise to choline and betaine aldehyde [48,49]. Choline monooxygenase convertscholine to betaine aldehyde, which is then converted to glycine betaine by theaction of betaine aldehyde dehydrogenase and has been identified in plantsshowing glycine betaine accumulation (Figure 27.3). Direct N-methylation ofglycine also produces glycine betaine along with other pathways in the case ofplants [50]. Abou El-Yazied [51] suggested that glycine betaine functions as acompatible solute during salt and drought stress, and is elevated to regulate theintracellular osmotic balance. In addition, positive effects of a foliar spray of glycine
27.2 Metabolite Changes During Abiotic Stresses 731
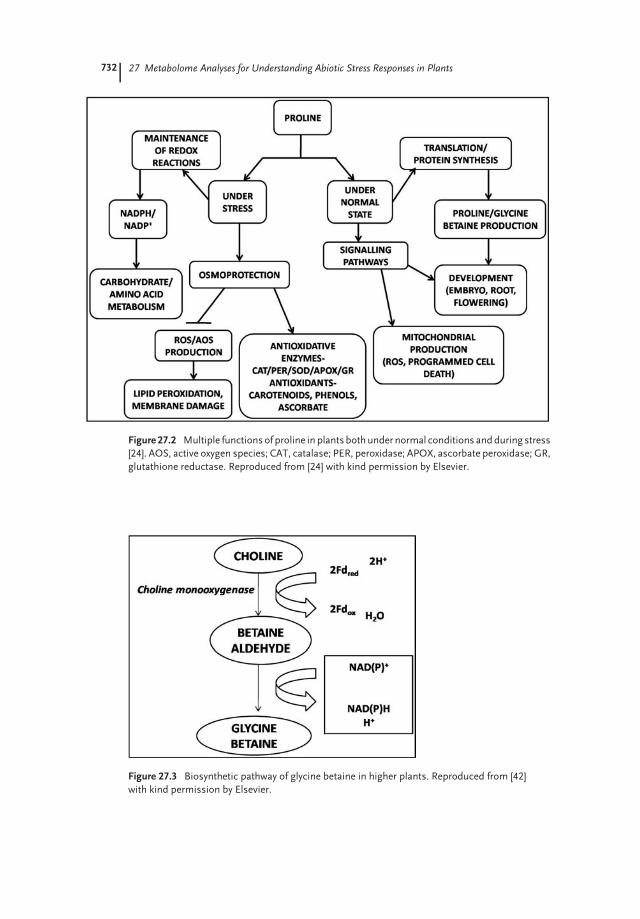
Figure 27.2 Multiple functionsof proline in plants bothunder normal conditions andduring stress
[24]. AOS, active oxygen species; CAT, catalase; PER, peroxidase; APOX, ascorbate peroxidase; GR,
glutathione reductase. Reproduced from [24] with kind permission by Elsevier.
Figure 27.3 Biosynthetic pathway of glycine betaine in higher plants. Reproduced from [42]
with kind permission by Elsevier.
732 27 Metabolome Analyses for Understanding Abiotic Stress Responses in Plants

betaine on yield and yield component in plants grown under water-limitedenvironments have been reported in different crops, such as wheat, common bean,rice, and sunflower [41,51–53]. Exogenous supply of glycine betaine (foliar spray)has also been shown to have an effective role in ameliorating the effects of waterstress on turgor potential and yield of two sunflower lines [53]. On the other hand,it has been suggested that exogenously applied glycine betaine has no effect on theyields of cotton [54].Both proline and glycine betaine are thought to have positive effects on enzyme
and membrane integrity along with adaptive roles in mediating osmoticadjustment in plants grown under stress conditions. Therefore, a better under-standing of the mechanisms of action of exogenously applied glycine betaine andproline is expected to aid their effective utilization in crop production in stressenvironments [42].
27.2.2
Carbohydrates
Under abiotic stress (drought, salinity) plants respond with the activation ofsynthesis or inhibition of catabolism of osmolytes such as sugars – trehalose,raffinose, galactinol, sorbitol, mannitol, fructans, saccharopine, sugar alcohols, andpolyols [55]. The changes in the concentration of carbohydrates, such as glucose,sucrose, sorbitol, galactose, raffinose, stachyose, polysaccharides, starch, and totalsugars, is important because they are directly related to physiological processessuch as photosynthesis, translocation, and respiration, and during abiotic stressthey accumulate as compatible solutes, as they do not interfere with thebiochemical reactions. Soluble sugars are highly sensitive to environmentalstresses, which act on the supply of carbohydrates from source organs to sinkorgans. Sucrose and hexoses both play dual functions in gene regulation asexemplified by the upregulation of growth-related genes and downregulation ofstress-related genes. Although coordinately regulated by sugars, these growth andstress-related genes are up- or downregulated through hexokinase-dependent and/or hexokinase-independent pathways [56]. Drought, salinity, low temperature, andflooding, in general, increased soluble sugar concentrations, whereas high lightirradiance (photosynthetically active radiation, UV-B), heavy metals, nutrientshortage, and ozone decreased sugar concentrations [57–59]. Singh [60] showedthat a greater accumulation of sugar lowers the osmotic potential of cells andreduces loss of turgidity in tolerant genotypes during abiotic stresses. Dkhiland Dendon [61] further suggested that another possible role of sugar may be as areadily available energy source. Glucose has been reported to induce a largenumber of genes involved in various stress responses, thus indicating the role ofsoluble sugars in environmental stress responses [62]. The alterations in thebalance between the source and the sink organs may be the signal for thesechanges that could have induced the change in the metabolism [63]. It was shownthat an artificial increase in leaf carbohydrate content modified gene expression forenzymes of photosynthetic metabolism [64]. Soluble sugars do not only function as
27.2 Metabolite Changes During Abiotic Stresses 733

metabolic resources and structural constituents of cells, they also act as signalsregulating various processes associated with plant growth and development [65,66].Sugar signaling pathways interact with stress pathways in a complex network tomodulate metabolic plant responses [67,68]. Soluble sugars may either act directlyas negative signals or as modulators of plant sensitivity and thus they can also playimportant roles in cell responses to stress-induced remote signals [56]. Never-theless, sugar changes do not follow a static model, and vary with genotype andstress factor [69]. In addition, it has also been reported that not all soluble sugarsplay similar roles in events associated with the metabolism of stressed plants[59,70]. Sucrose and glucose either act as substrates for cellular respiration or asosmolytes to maintain cell homeostasis [63], while fructose seems not to be relatedto osmoprotection, but to secondary metabolite synthesis [56]. Hilal et al. [71]demonstrated that fructose might be related to synthesis of erythrose-4-phoshate,which acts as a substrate in lignin and phenolic compound synthesis. Thus, understress conditions the metabolism of soluble sugars is a dynamic processsimultaneously involving degrading and synthetic reactions [56].Among various sugars, mannitol is a well-characterized sugar alcohol. It is a
significant photosynthetic product in some plant species and has been shown toincrease during abiotic stress [72]. It had been suggested to act as an osmoprotec-tant by reacting with damaging hydroxyl radicals to form mannitol radicals, whichare then converted to mannose in the presence of oxygen [73,74]. Shen et al. [75]transformed tobacco with a construct in which the mt1D enzyme was targeted tochloroplasts. The resulting transgenic plants accumulated mannitol at concentra-tions from 2.5 to 7.0mmol g�1 fresh weight. The presence of mannitol in thechloroplasts resulted in enhanced resistance to oxidative stress induced by thepresence of methyl viologen. Such resistance was due to an increased capacity toscavenge hydroxyl radicals. In the seeds of mannitol-accumulating Arabidopsisplants, the concentration of mannitol reached 10 mmol g�1 dry weight [76].Mannitol-expressing seeds were able to germinate in medium supplemented withup to 400mM NaCl, whereas control seeds ceased to germinate at 100mM NaCl.Fructans are polyfructose molecules that are produced by many plants and
bacteria, and furthermore these may play a role in adaptation to osmotic stress dueto their highly soluble nature [77]. Meer et al. [78] modified non-fructan-storingpotato plants by introducing the microbial fructosyl transferase gene and theregenerated potato plants accumulated fructans. Drought treatment resulted in33% more fresh weight in transformed fructan-accumulating plants than in thecontrol plants [79].Soluble sugars seem to assume a dual role with respect to ROS. Soluble sugars
can be involved in, or related to, ROS-producing metabolic pathways. In reverse,soluble sugars can also feed NADPH-producing metabolic pathways, such as theoxidative pentose phosphate pathway, which can contribute to ROS scavenging[80,81]. The relationship between soluble sugars and ROS production or betweensoluble sugars and ROS responses is not a straightforward positive correlation,since, as discussed above, high sugar levels can correspond to activation of someROS-producing pathways and decrease of other ROS-producing pathways, and
734 27 Metabolome Analyses for Understanding Abiotic Stress Responses in Plants

both high sugar level and low sugar level can result in the enhancement of ROSresponses. Moreover, soluble sugars other than fructose, glucose, and sucrose aredetected at significant levels in plants. Hexoses, such as mannose, and disacchar-ides, such as trehalose, may play important roles in relation to the three mainsoluble sugars. Thus, the positive effects of trehalose on stress responses in plants,including decrease of photooxidative damage, have been ascribed to interactionswith sucrose metabolism, which is significantly modified by trehalose treatment[82]. There is proof to show that abiotic stresses induce sugar signal transductionthat is related to enzyme activities and gene expression (Figure 27.4). Thephosphoinositol cascade is activated by abiotic stress-induced changes in sugarlevels. Activities of enzymes such as sucrose synthase and sucrose phosphatesynthase are upregulated, while sucrose transport is downregulated [62,83–87].
27.2.3
Polyamines
Polyamines can be considered as one of the oldest groups of substances known inbiochemistry [88]. Putrescine, spermidine, and spermine are the most common
Figure 27.4 Abiotic stressors cause the
accumulation of ROS in plant cells. MAPK
cascades respond to the resulting oxidative
burstandmayregulateROSaccumulation.Such
activation of MAPK components may be
mediated by hormone-related pathways. The
kinase modules depicted are supported by
biochemical and in plant analyses inA. thaliana.
Republished from [87] with permission of
Annual Reviews.
27.2 Metabolite Changes During Abiotic Stresses 735

polyamines in higher plants [1]. Putrescine can be produced from either ornithineby the action of ornithine decarboxylase or arginine by arginine decarboxylase.Putrescine, in turn, is converted to spermidine by spermidine synthase and then tospermine by spermine synthase. These are found to be protonated at normalcellular pH for which initially their biological function was associated with thecapability of binding different macromolecules carrying negative charge (i.e., DNA,RNA, chromatin, and proteins), thus confirming them as substances with astructural role [89].Several groups have hence confirmed that polyamines not only stabilize
macromolecular structures, but also act as regulatory molecules in manyfundamental cellular processes, including cell division, differentiation andproliferation, cell death, DNA and protein synthesis, and gene expression [90–93].The importance of polyamines in plants during various physiological processes aswell as abiotic and biotic stress responses has also been reported by several groups[92–94]. Alc�azar et al. [92] suggested that changes in plant polyamine metabolismoccur in response to a variety of abiotic stresses. However, the physiologicalsignificance of increased levels of polyamine in plants during abiotic stressresponses is still not clear [92,93,95]. They have also been implicated in protectingmembranes and alleviating oxidative stress [96–98], but their specific function instress tolerance is not well understood.Early studies on plant polyamine research pointed to their involvement in
responses to different environmental stresses. During the last few years, genetic,transcriptomic, and metabolomic approaches have unraveled key functions ofdifferent polyamines in the regulation of abiotic stress tolerance. Analyses oftransgenic plants and of mutants involved in polyamine metabolism clearly showeda positive role of polyamines in stress tolerance. Plants deficient in ADC1 or ADC2had reduced putrescine levels and were hypersensitive to stress [99,100], whereasconstitutive or stress-induced overexpression of ADC led to higher putrescinelevels and enhanced drought and freezing tolerance [89,101,102]. Nevertheless, theprecise molecular mechanism(s) by which polyamines control plant responses tostress stimuli are largely unknown. Recent studies indicate that polyaminesignaling is involved in direct interactions with different metabolic routes andintricate hormonal cross-talk. Thus, genetic manipulation of crop plants with genesencoding enzymes of polyamine biosynthetic pathways may provide better stresstolerance to crop plants [95].
27.3
Stress Hormones
The five classical phytohormones, auxin, cytokinin, ethylene, gibberellins, abscisicacid (ABA), and the recently identified brassinosteroids, jasmonic acid, and salicylicacid, are chemical messengers present in trace quantities; their synthesis andaccumulation are tightly regulated. Two groups of hormones are recognized: the“stress hormones,” which include ABA, salicylic acid, jasmonic acid, and ethylene,
736 27 Metabolome Analyses for Understanding Abiotic Stress Responses in Plants

and the “positive growth regulators,” which include auxins, cytokinins, gibber-ellins, and brassinosteroids. Depending on the context, they are subject to positiveor negative feedback control and often are affected by cross-talk due toenvironmental inputs. Phytohormones move throughout the plant body via thexylem or phloem transport stream. They move short distances between cells or aremaintained in their site of synthesis to exert their influence on target cells wherethey bind transmembrane receptors located at the plasma membrane orendoplasmic reticulum, or interact with intracellular receptors. The downstreameffects of hormonal signaling include alterations in gene expression patterns and insome cases non-genomic responses. Changes in plant hormone concentrationsand tissue sensitivity to them regulate a whole range of physiological process thathave profound effects on growth and development. The phytohormones affect allphases of the plant life cycle and the responses to environmental stresses, bothbiotic and abiotic. They are essential for the ability of plants to adapt to abioticstresses by mediating a wide range of adaptive responses [103–106] and, accordingto Santner and Estelle [103], they often rapidly alter gene expression by inducing orpreventing the degradation of transcriptional regulators via the ubiquitin–protea-some system. Hormonal signaling is critical for plant defenses against abiotic andbiotic stresses [107–109].
27.3.1
ABA
ABA is a central regulator of many plant responses to environmental stresses,particularly osmotic stresses [110–113], and hence one of the most studied topics inthe response of plants to abiotic stress, especially water stress, is ABA signaling andABA-responsive genes. ABA synthesis is one of the fastest responses of plants toabiotic stress, triggering ABA-inducible gene expression [114] and causing stomatalclosure, thereby reducing water loss via transpiration [115], and eventuallyrestricting cellular growth. Numerous genes associated with ABA de novobiosynthesis and genes encoding ABA receptors and downstream signal relayshave been characterized in A. thaliana (reviewed in [116]). In maize, at least 10viviparous mutants have been identified, most of which (vp2, vp5, vp7, vp9, w3, y3,and y9) were blocked in the biosynthesis of the carotenoid precursors for de novoABA synthesis; in rice (Oryza sativa), four phs mutants, defective in phytoenedesaturase (OsPDS), f-carotene desaturase (OsZDS), carotenoid isomerase (OsCR-TISO), and lycopene b-cyclase (b-OsLCY), were found to affect the biosynthesis ofcarotenoid precursors of ABA [117]. ABA signaling can also be very fast withoutinvolving transcriptional activity; a good example is the control of the stomatalaperture by ABA through the biochemical regulation of ion and water transportprocesses [112]. ABA is a key phytohormone in plant responses to water deficit;therefore, elucidation of the mechanism of ABA signal transduction significantlycontributes to the establishment of a suitable strategy for elevation of planttolerance to abiotic stresses [118]. Agarwal and Jha [119] suggested that ABAs arenot only involved in regulating stomatal opening, growth, and development, but are
27.3 Stress Hormones 737

also involved in coordinating various stress signal transduction pathways in plantsduring abiotic stresses, and that both ABA-dependent and ABA-independent signaltransduction pathways from stress signal perception to gene expression involvedifferent transcription factors such as DREB/CBF (dehydration-responsive elementbinding/C-repeat binding factor), MYC (myelocytomatosis oncogene)/MYB (mye-loblastosis oncogene), AREB/ABF (ABA-responsive element-binding protein/ABA-binding factor), NAC (NAM, ATAF, and CUC), and their corresponding cis-actingelements DRE (dehydration-responsive element), MYCRS/MYBRS, ABRE (ABA-responsive element), and NACRS. ABA also plays an important role during plantadaptations to cold temperatures. Cold stress induces the synthesis of ABA and theexogenous application of ABA improves the cold tolerance of plants [120].
27.3.2
Salicylic Acid
Salicylic acid has been recognized as a regulatory signal mediating plant responsesto abiotic stresses such as drought [121,122], chilling [123,124], heavy metaltolerance [125–127], heat [128,129], and osmotic stress [130]. It functions as anendogenous signal that mediates local and systemic plant defense responsesagainst pathogens, and thus it has been the focus of intensive research. In thissense, salicylic acid appears to be, just like in mammals, an “effective therapeuticagent” for plants [131].Treating plants with exogenous salicylic acid improved thermotolerance and heat
acclimation in several crops [123,132,133]. Using an Arabidopsis transgenic lineexpressing the salicylate hydroxylase gene (NahG) it was demonstrated thatsalicylic acid is required for ozone tolerance by maintaining the cellular redox stateand allowing defense responses [134]. However, by using Cvi-0, an Arabidopsisgenotype that accumulated high levels of salicylic acid, it was shown that salicylicacid activates an oxidative burst and a cell death pathway leading to ozonesensitivity [135].
27.3.3
Jasmonic Acid and Ethylene
Jasmonic acid levels in plants rapidly and transiently increase in response towounding, water deficit, mechanical stimulation, and elicitors, and it also mediatessome of the UV-induced defense responses [136].Abiotic stresses directly trigger jasmonic acid biosynthesis, which functions as a
signal in the cascade of plant reactions to oxidative stress-generating stimuli. Inaddition, jasmonic acid synthesis and signaling are interlinked by a positivefeedback loop whereby jasmonates stimulate their own synthesis [137,138].Jasmonic acid is also involved in the response to wounding. Jasmonic acidbiosynthesis is initiated by wound-mediated release of a-linolenic acid fromchloroplastic membranes, followed by the activity of several chloroplast-locatedenzymes, including 13-lipoxygenase. The combination of jasmonic acid deficiency
738 27 Metabolome Analyses for Understanding Abiotic Stress Responses in Plants

and ethylene insensitivity resulted in a novel growth phenotype characterized bymassive cell expansion around wounds, suggesting that both jasmonic acid andethylene may repress local growth after wounding and/or herbivore attack [139].Recent findings of Mahouachi et al. have suggested an interaction between
jasmonic acid and ABA in plants under drought stress [140]. According to theauthors, the pattern of accumulation of jasmonic acid is compatible with a triggeringsignal upstream ABA. The phytohormone ABA acts in all adaptive responses toenvironmental stresses, while gibberellin and ABA play essential and oftenantagonistic roles in regulating plant growth, development, and stress responses.The transduction of their signal occurs through several types of receptors [141].Ethylene has long been regarded as a stress hormone [142]. However, the roles of
ethylene signaling in abiotic stress responses are still not clear, but it is known thatethylene signaling is important in regulating plant growth and stress responses,and it functions through its receptors. In a study by Cao et al. [143], theytransformed a tobacco type II ethylene receptor homolog gene NTHK1 intoArabidopsis and found that the resulting transgenic plants, with NTHK1 mRNAand protein expression, were salt sensitive as was seen from the severe epinastyphenotype, high electrolyte leakage, and reduced root growth under salt stress.It is apparent from the evidence obtained so far by different workers that ABA,
jasmonic acid, and ethylene signaling pathways interact to regulate diverse stressresponses. The nature of the interaction between these pathways appears to dependon the type of stress experienced by the plant.
27.4
Antioxidants
One of the earliest signals in many abiotic stresses involves ROS and reactivenitrogen species (RNS), which modify enzyme activity and gene regulation[115,144]. The main source of ROS production in plants are chloroplasts andperoxisomes through photorespiration during light [145] and in mitochondriaduring darkness [146]. The chloroplast is one of the major producers of superoxide(O2
�) and hydrogen peroxide (H2O2) in plants. In chloroplast thylakoids, thereaction centers of Photosystems I and II are the major sites of generation of ROS[147].ROS signaling in response to abiotic stresses and its interactions with hormones
has been thoroughly reviewed [148]. ROS and RNS form a coordinated networkthat regulates many plant responses to the environment; there are a large numberof studies on the oxidative effects of ROS on plant responses to abiotic stress, butonly a few studies documenting the nitrosative effects of RNS [149]. ROS are anatural consequence of aerobic metabolism and plants have mechanisms to dealwith them under normal conditions, controlling the formation and removal rates.Under stress conditions, cell homeostasis is disrupted and ROS production canincrease significantly, putting a heavy burden on those antioxidative mechanisms,some of which are activated in order to eliminate the excess ROS [150]. Under
27.4 Antioxidants 739

certain stress conditions (like excess light, cold, heat, drought, heavy metals, etc.)the production of ROS can exceed the capacity of the plant’s defense mechanisms,an imbalance in intracellular ROS content is established, and this results inoxidative stress [95]. The ROS comprising O2
�, H2O2,1O2, HO2
�, OH�, ROOH,ROO�, and RO� are highly reactive and toxic, and cause damage to proteins, lipids,carbohydrates, and DNA, which ultimately results in cell death. Accumulation ofROS as a result of various environmental stresses is a major cause of loss of cropproductivity worldwide [151–156]. ROS may have damaging, protective, orsignaling functions, which are expressed at appropriate times. Stress-induced ROSaccumulation is counteracted by enzymatic antioxidant systems that include avariety of scavengers, such as superoxide dismutase, peroxidases, catalase, andglutathione reductase, as well as non-enzymatic low-molecular-weight metabolites,such as ascorbate, glutathione, a-tocopherol, carotenoids, and flavonoids[22,150,157].
27.5
Stress Proteins and Protein Kinases
Elucidating the function of proteins expressed by genes in stress-tolerant and-susceptible plants would advance our understanding of plant adaptation andtolerance to environmental stresses, but also may provide important informationfor designing new strategies for crop improvement [112]. Protein kinases andprotein phosphatases often act in tandem to perform the phosphorylationand dephosphorylation process, and much work is ongoing to unravel themolecular mechanisms involved in the expression of several of the large genefamilies that encode protein kinases and phosphatases involved in stress signalingin plants. Another large group of stress proteins are the heat shock proteins that areexpressed mainly during temperature stress, but also other stresses.Ca2þ is one of the most important second messengers in response to
extracellular stimuli in plants [158,159]. Signal molecules such as inositol trispho-sphate, diacylglycerol, inositol hexaphosphate, cADP-ribose, or ROS induceincreased levels of intracellular Ca2þ under stress conditions. Two protein kinasesare postulated to be the targets of the Ca2þ signal in plants. One is SnRK3, whoseactivity is dependent on the Ca2þ-binding calcineurin B-like (CBL) proteins [160].Arabidopsis has 25 SnRK3-type kinases, according to genome information, of whichthe best characterized is salt overly sensitive 2 (SOS2)/CIPK24/SnRK3.11, whichwas identified as an essential factor in the salinity stress response [161]. Inconjunction with SOS3/ScaBP8/CBL10 Ca2þ-binding protein, SOS2 activates theplasma membrane Naþ/Hþ antiporter (SOS1) required for salinity tolerance[162,163]. The other protein kinase is calcium-dependent protein kinase (CDPK).CDPKs comprise a large family of Ser/Thr kinases in plants and protozoans, andare calcium-binding, and it has been reported that CDPK transcripts are elevatedafter race-specific defense elicitation and hypo-osmotic stress [164]. Earlier reportshave also indicated CDPK transcript elevation after exposure of Arabidopsis to cold,
740 27 Metabolome Analyses for Understanding Abiotic Stress Responses in Plants

salt, and drought [165,166]. Arabidopsis has more than 30 CDPK genes, and severalof them have been shown to function in abiotic stress and ABA responses [160].CPK3 and CPK6 regulate the ABA response in guard cells [167], and CPK4, CPK11,and CPK32 positively regulate the ABA response [168,169]. In addition, CPK4 andCPK11 phosphorylate AREB/ABF transcription factors in an ABA-dependentmanner [169].Mitogen-activated protein kinase (MAPK) cascades function as major cellular
signaling components in eukaryotes. Therefore, plant genes for MAPK cascadeshave been examined to determine whether they are involved in various stressresponses. MAPK signaling pathways have been found to be involved in abioticstress responses in plants and this cascade links external stimuli with severalcellular responses. The cascade generally consists of MAP4Ks, MAP3Ks, MAP2Ks,and MAPKs. During stress, the stimulated plasma membrane activates MAP3Ks orMAP4Ks, which may act as adaptors linking upstream signaling steps to the coreMAPK cascades. MAP3Ks are also Ser/Thr kinases phosphorylating two aminoacids in the S/T–X–S/T motif of the MAP2K activation loop. MAP2Ks phosphor-ylate MAPKs on threonine and tyrosine residues at a conserved T–X–Ymotif [170].Arabidopsis genome information allowed identification of 20 MAPKs, 10 MAP2Ks,and 60 MAPK Kinase Kinases [171]. These MAPK components appear to functionin several different signaling processes, so they might not constitute simplecascades, but rather networks, making it difficult to identify the function of eachcomponent. Teige et al. [172] reported that an A. thaliana cascade that responds tosalt, drought, and cold may include the components MEKK1, MKK2, MPK4, and/or MPK6. Initially, this work showed that whereas mkk2 mutants had reducedtolerance to salinity, transgenic lines that overexpressed MKK2 exhibited enhancedtolerance to salt and cold. Stressor-specific induction of MAPK genes and increasedMAPK activity have been detected when plants are subjected to touch, cold, salinity,genotoxic agents, UV irradiation, ozone, and oxidative stress [173].MAPK cascades are induced by ROS, but may also regulate ROS levels by
affecting antioxidative enzymes, such as catalase activity. It has been shown inseveral studies that MAPK activity may be mediated through hormones, notably thestress-responsive hormones such as ABA, jasmonic acid, salicylic acid, andethylene [87].
27.6
Stress-Responsive Gene Expression
In the postgenomic era, comprehensive analyses using three systematicapproaches or “omics” have increased our understanding of the complex molecularregulatory networks associated with stress adaptation and tolerance: transcrip-tomics, metabolomics, and proteomics (see Section 27.1). Along with morphologi-cal and physiological studies on the responses of plants to stress conditions, severalmolecular mechanisms from gene transcription to translation as well asmetabolites are currently being investigated. Recent advances in genomic research,
27.6 Stress-Responsive Gene Expression 741

particularly in the field of proteomics, have created an opportunity for dissectingquantitative traits in a more meaningful way. Proteomics is a powerful tool forinvestigating the molecular mechanisms of the responses of plants to stresses andit provides a path toward increasing the efficiency of indirect selection for inheritedtraits [174]. Understanding the function of genes is a major challenge of thepostgenomic era. While many of the functions of individual parts are unknown,their function can sometimes be inferred through association with other knownparts, providing a better understanding of the biological system as a whole. High-throughput “omics” technologies are facilitating the identification of new genesand gene functions. In addition, network reconstructions at the genome scale arekey to quantifying and characterizing the genotype to phenotype relationship [175].There is a need to provide established or new varieties with genotypes havingenhanced or faster induction of expression of genes at the crossroad of permissivegrowth under stress conditions. This group of permissive genes includes aquaporinisoforms able to optimize water fluxes [176]. The stress-responsive genes can beclassified into two classes: early and delayed response genes [177]. The former areinduced quickly and transiently, while the latter are activated more slowly and theirexpression is sustained. The early-response genes encode transcription factors thatactivate downstream delayed-response genes. Regulatory proteins involved indrought stress such as proton antiporters TNHX1 and a proton pyrophosphataseTVP1 have been shown to improve salt and drought stress tolerance in Arabidopsis[178,179]. ERECTA is another gene regulating transpiration efficiency affectingstomatal closure, while the plant is able to maintain biomass production [180].SlERF5 is highly expressed in response to the harpin protein coded in the Hrpgene clusters in many Gram-negative phytopathogens; the overexpression ofSlERF5 is involved in the induction of the dehydration-responsive genes throughthe ABA-mediated abiotic stress response [181]. In rice, GF14 genes contain cis-elements in their promoter regions that are responsive to abiotic stress andpathogen attack. The 14-3-3s family genes are also subject to regulation by certaintranscript factors [182]. In rice, the RO-292 gene is upregulated in roots by salt ordrought stresses and by blast fungus infection [183]. Gene expression profiling hasallowed the identification of hundreds of genes induced when plants are exposed tostress [184–187]. The availability of the complete genome sequence of some modelplants, such as O. sativa and A. thaliana, has allowed the development of whole-genome tiling microarrays. This constitutes a powerful new technology that hasalready made possible the identification of several unannotated transcriptsresponsive to abiotic stress [188,189].
27.7
Role of MicroRNAs in Abiotic Stress
MicroRNAs (miRNAs) are known to play important regulatory roles in plants bytargeting mRNAs for cleavage or translational repression [190]. First discovered in2002, several groups of scientists have found plant miRNAs by cloning small RNAs
742 27 Metabolome Analyses for Understanding Abiotic Stress Responses in Plants

in Arabidopsis [191–193]. According to the miRNA Registry, 731 miRNA genes havebeen identified in various plants, including 117 from Arabidopsis, 178 from rice (O.sativa), 97 from Zea mays, and the rest from Populus trichocarpa, Saccharumoffcinarum, Sorghum bicolor, Medicago truncatula, and Glycine max [194]. While ithas been known that abiotic stresses, such as drought, salinity, and extremetemperatures, regulate the expression of thousands of genes in plants at bothtranscriptional and post-transcriptional levels, currently many miRNAs have beenpredicted and some have been confirmed experimentally to be involved in a varietyof abiotic stress responses.miRNAs generally target miR398, which was discovered in Arabidopsis and
rice (O. sativa), and is encoded by three loci (MIR398a, MIR398b, andMIR398c) in Arabidopsis. miR398 targets two closely related Cu/Zn-superoxidedismutase (SOD) genes, cytosolic CSD1 and chloroplast-localized CSD2[190,195,196], which under oxidative stress are overexpressed as a line of plantdefense. CSD1 and CSD2 mRNA levels were increased in response to highlight, Cu, Fe, and so on, and miR398 was downregulated under oxidative stressconditions. The lack of CSD1 and CSD2 expression in unstressed plantsdepends on miR398-mediated post-transcriptional regulation, and the stressinduction of CSD1 and CSD2 mRNA is mediated by the downregulation ofmiR398. It was also shown by Sunkar et al. [197] that transgenic A. thalianaplants overexpressing a miR398-resistant form of CSD2 accumulate more CSD2mRNA than plants overexpressing a regular CSD2, and are consequently muchmore tolerant to high light, heavy metals, and other oxidative stresses. Sunkarand Zhu [198] reported that the expression of miR393 was strongly upregulatedby ABA treatments, while miR397b and miR402 are slightly upregulated, andmiR389a downregulated, by ABA stress treatment.
27.8
Conclusion
Various abiotic stresses affect plants and are causes of stagnation or reduction incrop productivity. With the rapid increase in population, projected to be 9.4 billionby 2050, worldwide food production needs to be significantly increased to gear upto meet the demands in the coming years. There is evidence that yields of paddyand wheat have shown a declining trend in many parts of South-East Asia due toincreasing water stress [199]. Related to water stress are other abiotic stresses suchas elevated temperature and salinity that also have negative impacts on plants. Inthe Indian subcontinent, it is projected that by 2050 the average temperatures mayrise by 2.5 �C, which is an alarming situation for agriculture. Responses of plants toabiotic stresses are complex and affect several components. Such stresses not onlyaffect cell water potential, induce closure of stomata, and decrease photosynthesis,nitrate assimilation, and various anabolic enzyme reactions, but also induce thegeneration of ROS, such as superoxide radicals, hydrogen peroxide, and hydroxylradicals, which in turn cause lipid peroxidation and consequently membrane
27.8 Conclusion 743

injury, protein degradation, enzyme inactivation, pigment bleaching, and disrup-tion of DNA strands.In this context, metabolomic approaches for detailed analyses of metabolites
before, during, and after stresses can give meaningful inputs to develop crops withenhanced tolerance to different stresses. Although several technological advanceshave been made in the recent past in the field of biotechnology, one of the majorchallenges is the widening gap between the rate of development of new techno-logies and their deployment in applied breeding programs for crop improvement[200]. While many genes for different stresses have been cloned and characterizedin models as well as some crop plant species, and in some cases successfuldevelopment of transgenics has also been reported (e.g., rice for LOS5/ABA3 [201]and AP37 [202]), to date no reports of a released transgenic variety for droughttolerance have been published, even though transgenic crops have been widelyadopted globally [203]. Thus, the success of biotechnology for developing abioticstress-tolerant cultivars has been rather limited, indicating several inherentcomplexities [200]. It would therefore seem imperative to look for othertechnologies that would help plants overcome abiotic stresses and withstand globalclimate changes without affecting their productivity.
References
1 Krasensky, J. and Jonak, C. (2012) Drought,salt, and temperature stress-inducedmetabolic rearrangements and regulatorynetworks. J. Exp. Bot., 63, 1593–1608.
2 Hasegawa, P.M., Bressan, R.A., Zhu, J.K.,and Bohnert, H.J. (2000) Plant cellular andmolecular responses to high salinity. Annu.Rev. Plant Physiol. Plant Mol. Biol., 51,463–499.
3 Acquaah, G. (2007) Principles of PlantGenetics and Breeding, Blackwell, Oxford.
4 Ingram, J. and Bartels, D. (1996) Themolecular basis of dehydration tolerance inplants. Annu. Rev. Plant Physiol. Plant Mol.Biol., 47, 377–403.
5 Apse, M.P. and Blumwald, E. (2002)Engineering salt tolerance in plants. Curr.Opin. Biotechnol., 13, 146–150.
6 Zhu, J.K. (2002) Salt and drought stresssignal transduction in plants. Annu. Rev.Plant Biol., 53, 243–273.
7 Viswanathan, C. and Zhu, J.K. (2004)Molecular perspectives on cross-talk andspecificity in abiotic stress signalling inplants. J. Exp. Bot., 55, 225–236.
8 Sangam, S., Jayasree, D., Reddy, K.J.,Chari, P.V.B., Sreenivasulu, N., and Kavi
Kishor, P.B. (2005) Salt tolerance in plants– transgenic approaches. J. PlantBiotechnol., 7, 1–15.
9 Valliyodan, B. and Nguyen, H.T. (2006)Understanding regulatory networks andengineering for enhanced droughttolerance in plants. Curr. Opin. Plant Biol.,9, 189–195.
10 Lenz, T., Fischer, J.J., and Dreger, M.(2011) Probing small molecule–proteininteractions: a new perspective forfunctional proteomic. J. Proteomics, 75,100–115.
11 Kaufmann, K., Smaczniak, C., de Vries, S.,Angenent, G.C., and Karlova, R. (2011)Proteomics insights into plant signallingand development. Proteomics, 11, 744–755.
12 Chinnusamy, V., Zhu, J., and Zhu, J.K.(2006) Gene regulation during coldacclimation in plants. Physiol. Plant., 126,52–61.
13 Bartels, D. and Sunkar, R. (2005) Droughtand salt tolerance in plants. Crit. Rev. PlantSci., 24, 23–58.
14 Munns, R. and Tester, M. (2008)Mechanisms of salinity tolerance. Annu.Rev. Plant Biol., 59, 651–681.
744 27 Metabolome Analyses for Understanding Abiotic Stress Responses in Plants

15 Janska, A., Marsik, P., Zelenkova, S.,and Ovesna, J. (2010) Cold stress andacclimation: what is important formetabolic adjustment? Plant Biol., 12,395–405.
16 Urano, K., Kurihara, Y., Seki, M., andShinozaki, K. (2010) ‘Omics’ analyses ofregulatory networks in plant abioticstress responses. Curr. Opin. Plant Biol.,13, 132–138.
17 Chen, D., Kessler, B., and Monselise, S.P.(1964) Studies on water regime andnitrogen metabolism of citrus seedlingsgrown under water stress. Plant Physiol.,39, 379–386.
18 Barnett, N.M. and Naylor, A.W. (1966)Amino acid and protein metabolism inBermuda grass during water stress. PlantPhysiol., 41, 1222–1230.
19 Chu, T.M., Aspinall, D., and Paleg, L.G.(1974) Stress metabolism. VI. Temperaturestress and the accumulation of proline inbarley and radish. Aust. J. Plant Physiol., 1,87–97.
20 Chu, T.M., Aspinall, D., and Paleg, L.G.(1976) Stress metabolism. VII. Salinity andproline accumulation in barley. Aust. J.Plant Physiol., 3, 219–228.
21 Chakraborty, U., Dutta, S., andChakraborty, B.N. (2002) Response oftea plants to water stress. Biol. Plant.,45, 557–562.
22 Chakraborty, U. and Pradhan, B. (2012)Oxidative stress in five wheat varieties(Triticum aestivum L.) exposed to waterstress and study of their antioxidantenzyme defense system, water stressresponsive metabolites and H2O2
accumulation. Braz. J. Plant Physiol., 24,117–130.
23 Verbruggen, N. and Hermans, C. (2008)Proline accumulation in plants: a review.Amino Acids, 35, 753–759.
24 Szabados, L. and Savour�e, A. (2010)Proline: a multifunctional amino acid.Trends Plant Sci., 15, 89–97.
25 Delauney, A.J. and Verma, D.P.S. (1993)Proline biosynthesis and osmoregulationin plants. Plant J., 4, 215–223.
26 Roosens, N.H. et al. (1998) Isolation of theornithine-delta-aminotransferase cDNAand effect of salt stress on its expression inArabidopsis thaliana. Plant Physiol., 117,263–271.
27 Funck, D., Stadelhofer, B., and Koch, W.(2008) Ornithine-delta-aminotransferase isessential for arginine catabolism but notfor proline biosynthesis. BMC Plant Biol.,8, 40.
28 Liu, J. and Zhu, J.K. (1997) Prolineaccumulation and salt-stress induced geneexpression in a salt-hypersensitive mutantof Arabidopsis. Plant Physiol., 114, 591–596.
29 Xin, Z. and Browse, J. (1998) Eskimo1mutants of Arabidopsis are constitutivelyfreezing-tolerant. Proc. Natl. Acad. Sci.USA, 95, 7799–7804.
30 Choudhary, N.L. et al. (2005) Expression ofdelta1-pyrroline-5-carboxylate synthetasegene during drought in rice (Oryzasativa L.). Indian J. Biochem. Biophys., 42,366–370.
31 Chen, Z., Cuin, T.A., Zhou, M., Twomey,A., Naidu, B.P., and Shabala, S. (2007)Compatible solute accumulation and stressmitigating effects in barley genotypescontrasting in their salt tolerance. J. Exp.Bot., 58, 4245–4255.
32 Widodo, Patterson, J.H., Newbigin, E.,Tester, M., Bacic, A. and Roessner, U.(2009) Metabolic responses to salt stress ofbarley (Hordeum vulgare L.) cultivars,Sahara and Clipper, which differ in salinitytolerance. J. Exp. Bot., 60, 4089–4103.
33 Hong, Z., Hong, Z., Lakkineni, K., Zhang,Z., and Verma, D.P.S. (2000) Removal offeedback inhibition of delta (1)-pyrroline-5-carboxylate synthetase results in increasedproline accumulation and protection ofplants from osmotic stress. Plant Physiol.,122, 1129–1136.
34 Mattioli, R., Marchese, D., and D’Angeli, S.(2008) Modulation of intracellular prolinelevels affects flowering time andinflorescence architecture in Arabidopsis.Plant Mol. Biol., 66, 277–288.
35 Szekely, G., Sz�ekely, G., Abrah�am, E.,Cs�eplo, A., Rig�o, G., Zsigmond, L.,Csisz�ar, J., Ayaydin, F., Strizhov, N., J�asik,J., Schmelzer, E., Koncz, C., and Szabados,L. (2008) Duplicated P5CS genes ofArabidopsis play distinct roles in stressregulation and developmental control ofproline biosynthesis. Plant J., 53, 11–28.
36 Miller, G., Honig, A., Stein, H., Suzuki, N.,Mittler, R., and Zilberstein, A. (2009)Unravelling delta1-pyrroline-5-carboxylateproline cycle in plants by
References 745

uncoupled expression of proline oxidationenzymes. J. Biol. Chem., 284, 26482–26492.
37 Yamada, M., Morishita, H., Urano, K.,Shiozaki, N, Yamaguchi-Shinozaki, K.,Shinozaki, K., and Yoshiba, Y. (2005)Effects of free proline accumulation inpetunias under drought stress. J. Exp. Bot.,56, 1975–1981.
38 Nanjo, T., Kobayashi, M., Yoshiba, Y.,Wada, K., Tsukaya, H., Kakaubari, Y.,Yamaguchi-Shinozaki, K., andShinozaki, K. (1999) Biological functionsof proline in morphogenesis andosmotolerance revealed in antisensetransgenic Arabidopsis thaliana. Plant J.,18, 185–193.
39 Parida, A.K., Dagaonkar, V.S., Phalak, M.S., and Aurangabadkar, L.P. (2008)Differential responses of the enzymesinvolved in proline biosynthesis anddegradation in drought tolerant andsensitive cotton genotypes during droughtstress and recovery. Acta Physiol. Plant., 30,619–627.
40 Qu, L.J., Wu, L.Q., Fan, Z.M., Guo, L., Li,Y.Q., and Chen, Z.L. (2005) Over-expression of the bacterial nhaA gene inrice enhances salt and drought tolerance.Plant Sci., 168, 297–302.
41 Aldesuquy, H.S., Mohamed, A.A., Abo-Hamed, S.A., Elhakem, A.H., and Saeed,A.S. (2012) Glycine betaine and salicylicacid induced modification in productivityof two different cultivars of wheat grownunder water stress. J. Stress Physiol.Biochem., 8, 72–89.
42 Ashraf, M. and Foolad, M.R. (2007) Rolesof glycine betaine and proline inimproving plant abiotic stress resistance.Environ. Exp. Bot., 59, 206–216.
43 Mansour, M.M.F. (2000) Nitrogencontaining compounds and adaptation ofplants to salinity stress. Biol. Plant., 43,491–500.
44 Mohanty, A., Kathuria, H., Ferjani, A.,Sakamoto, A., Mohanty, P., Murata, N.,and Tyagi, A.K. (2002) Transgenics of anelite indica rice variety Pusa Basmati 1harbouring the codA gene are highlytolerant to salt stress. Theor. Appl. Genet.,106, 51–57.
45 Yang, W.J., Rich, P.J., Axtell, J.D., Wood,K.V., Bonham, C.C., Ejeta, G., Mickelbart,M.V., and Rhodes, D. (2003) Genotypic
variation for glycine betaine in sorghum.Crop Sci., 43, 162–169.
46 Robinson, S.P. and Jones, G.P. (1986)Accumulation of glycine betaine inchloroplasts provides osmotic adjustmentduring salt stress. Aust. J. Plant Physiol, 13,659–668.
47 Genard, H., Le, S.J., Hillard, J.,Tremolieres, A., and Boucaud, J. (1991)Effect of salinity on lipid composition,glycine betaine content andphotosynthetic activity in chloroplasts ofSuaeda maritime. Plant Physiol. Biochem.,29, 421–427.
48 Hanson, A.D. and Scott, N.A. (1980)Betaine synthesis from radioactiveprecursors in attached, water-stressedbarley leaves. Plant Physiol., 66, 342–348.
49 Rhodes, D. and Hanson, A.D. (1993)Quaternary ammonium and tertiarysulfonium compounds in higher-plants.Annu. Rev. Plant Physiol. Plant Mol. Biol.,44, 357–384.
50 Weretilnyk, E.A., Bednarek, S., McCue,K.F., Rhodes, D., and Hanson, A.D. (1989)Comparative biochemical andimmunological studies of the glycinebetaine synthesis pathway in diversefamilies of dicotyledons. Planta, 178,342–352.
51 Abou El-Yazied, A.A. (2011) Foliarapplication of glycine betaine and chelatedcalcium improves seed production andquality of common bean (Phaseolus vulgarisL.) under water stress conditions. Res. J.Agric Biol. Sci., 7, 357–370.
52 Rahman, M.S., Miyake, H., and Takeoka, Y.(2002) Effects of exogenous glycine-betaineon growth and ultra-structure of salt-stressed rice seedlings (Oryza sativa L.).Plant Prod. Sci., 5, 33–44.
53 Iqbal, N., Ashraf, M., and Ashraf, M.Y.(2008) Glycinebetaine, an osmolyte ofinterest to improve water stress tolerancein sunflower (Helianthus annuus L.):water relations and yield. S. Afr. J. Bot., 74,274–281.
54 Meek, C. Oosterhuis, D. and Gorham, J.(2003) Does foliar-applied glycine betaineaffect endogenous betaine levels and yieldin cotton? Crop Manag., doi: 10.1094/CM-0804-02-RS.
55 Cortina, C. and Culianez-Macia, F.A.(2005) Tomato abiotic stress enhanced
746 27 Metabolome Analyses for Understanding Abiotic Stress Responses in Plants

tolerance by trehalose biosynthesis. PlantSci., 169, 75–82.
56 Rosa, M., Prado, C., Podazza, G.,Interdonato, R., Gonz�alez, J.A., Hilal, M.,and Prado, F.E. (2009) Soluble sugars:metabolism, sensing and abiotic stress: Acomplex network in the life of plants. PlantSignal. Behav., 4, 388–393.
57 Strand, A�., Hurry, V., Henkes, S., Huner,
N., Gustafsson, P., Gardestr€om, P., andStitt, M. (1999) Acclimation of Arabidopsisleaves developing at low temperatures.Increasing cytoplasmic volumeaccompanies increased activities ofenzymes in the Calvin cycle and in thesucrose-biosynthesis pathway. PlantPhysiol., 119, 1387–1397.
58 Dubey, R.S. and Singh, A.K. (1999) Salinityinduces accumulation of soluble sugarsand alters the activity of sugarmetabolising enzymes in rice plants. Biol.Plant., 42, 233–239.
59 Gill, P.K., Sharma, A.D., Singh, P., andBhullar, S.S. (2001) Effect of various abioticstresses on the growth, soluble sugars andwater relations of sorghum seedlingsgrown in light and darkness. Bulg. J. PlantPhysiol., 27, 72–84.
60 Singh, A.K. (2004) The physiology of salttolerance in four genotypes of chickpeaduring germination. J. Agric. Sci. Technol.,6, 87–93.
61 Dkhil, B.B. and Denden, M. (2010) Saltstress induced changes in germination,sugars, starch and enzyme of carbohydratemetabolism in Abelmoschus esculentus L.(Moench.) seeds. Afr. J. Agric. Res., 5,1412–1418.
62 Price, J., Laxmi, A., Martin, S.K., and Jang,J.C. (2004) Global transcription profilingreveals multiple sugar signal transductionmechanisms in Arabidopsis. Plant Cell, 16,2128–2150.
63 Gupta, A.K. and Kaur, N. (2005) Sugarsignalling and gene expression inrelation to carbohydrate metabolismunder abiotic stresses in plants. J.Biosci., 30, 761–776.
64 Krapp, A. and Stitt, M. (1995) Anevaluation of direct and indirectmechanisms for the sink-regulation ofphotosynthesis in spinach: changes in gasexchange, carbohydrates, metabolites,enzyme activities and steady state
transcript levels after cold-girdling sourceleaves. Planta, 195, 313–323.
65 Jang, J.C. and Sheen, J. (1997) Sugarsensing in plants. Trends Plant Sci., 2, 208–214.
66 Pego, J.V., Kortstee, A.J., Huijser, C., andSmeekens, S.C.M. (2000) Photosynthesis,sugars and the regulation of geneexpression. J. Exp. Bot., 51, 407–416.
67 Ho, S.L., Chao, Y.C., Tong, W.F., and Yu,S.M. (2001) Sugar co-ordinately anddifferentially regulates growth and stress-related gene expression via a complexsignal transduction network and multiplecontrol mechanisms. Plant Physiol., 125,877–890.
68 Tran, L.S., Nakashima, K., Sakuma, Y.,Osakabe, Y., Qin, F., Simpson, S.D.,Maruyama, K., Fujita, Y., Shinozaki, K.,and Yamaguchi-Shinozaki, K. (2007) Co-expression of the stress-inducible zincfinger homeodomain ZFHD1 and NACtranscription factors enhances expressionof the ERD1 gene in Arabidopsis. Plant J.,49, 46–63.
69 Morsy, M.R., Jouve, L., Hausman, J.F.,Hoffmann, L., and Stewart, J.M. (2007)Alteration of oxidative and carbohydratemetabolism under abiotic stress in two rice(Oryza sativa L.) genotypes contrasting inchilling tolerance. J. Plant Physiol., 164,157–167.
70 Almodares, A., Hadi, M.R., and Dosti, B.(2008) The effects of salt stress on growthparameters and carbohydrates contents insweet sorghum. Res. J. Environ. Sci., 2,298–304.
71 Hilal, M., Parrado, M.F., Rosa, M.,Gallardo, M., Orce, L., Massa, E.M.,Gonzalez, J.A., and Prado, F.E. (2004)Epidermal lignin deposition in quinoacotyledons in response to UV-B radiation.Photochem. Photobiol., 79, 205–210.
72 Zamski, E., Guo, W.W., Yamamoto, Y.T.,Pharr, D.M., and Williamson, J.D. (2001)Analysis of celery (Apium graveolens)mannitol dehydrogenase (Mtd) promoterregulation in Arabidopsis suggests roles forMTD in key environmental and metabolicresponses. Plant Mol. Biol., 47, 621–631.
73 Franzini, E., Sellak, H., Hakim, J., andPasquier, C. (1994) Comparative sugardegradation by (OH) produced by the iron-driven Fenton reaction and gamma
References 747

radiolysis. Arch. Biochem. Biophys., 309,261–265.
74 Shen, B., Jensen, R.G., and Bohnert, H.J.(1997) Mannitol protects against oxidationby hydroxyl radicals. Plant Physiol., 115,527–532.
75 Shen, B., Jensen, R.G., and Bohnert, H.J.(1997) Increased resistance to oxidativestress in transgenic plants by targetingmannitol biosynthesis to the chloroplast.Plant Physiol., 113, 1177–1183.
76 Thomas, J.C., Sepahi, M., Arendall, B., andBohnert, H.J. (1995) Enhancement of seedgermination in high salinity byengineering mannitol expression inArabidopsis thaliana. Plant Cell Environ., 18,801–806.
77 Gupta, A.K. and Kaur, N. (2000) Fructanmetabolism in Jerusalem artichoke andchicory, in Carbohydrate Reserves in Plants.Synthesis and Regulation (eds A.K. Guptaand N. Kaur), Elsevier, Amsterdam,pp. 223–248.
78 Meer, I.M., Ebskamp, M.J.M., Visser, R.G.F., Weisbeek, P.J., and Smeekens, S.C.M.(1994) Fructan as a new carbohydrate sinkin transgenic potato plants. Plant Cell, 6,561–570.
79 Pilon-Smits, E.A.H., Ebskamp, M.J.M.,Paul, M.J., Jeuken, M.J.W., Weisbeek, P.J.,and Smeekens, S.C.M. (1995) Improvedperformance of transgenic fructan-accumulating tobacco under droughtstress. Plant Physiol., 107, 125–130.
80 Russell, D.G., Mwandumba, H.C., andRhoades, E.E. (2002)Mycobacterium andthe coat of many lipids. J. Cell Biol., 158,421–426.
81 Barros, M.H., Bandy, B., Tahara, E.B., andKowaltowksi, A.J. (2004) Higherrespiratory activity decreasesmitochondrial reactive oxygen release andincreases life span in Saccharomycescerevisiae. J. Biol. Chem., 279, 49883–49888.
82 Bae, H.H., Herman, E., Bailey, B., Bae, H.J., and Sicher, R. (2005) Exogenoustrehalose alters Arabidopsis transcriptsinvolved in cell wall modification, abioticstress, nitrogen metabolism, and plantdefense. Physiol. Plant., 125, 114–126.
83 Kaur, S., Gupta, A.K., and Kaur, N. (1998)Gibberellin A3 reverses the effect of saltstress in chickpea (Cicer arietinum L.)seedlings by enhancing the amylase
activity and mobilization of starch incotyledons. Plant Growth Regul., 26, 85–90.
84 Umemura, T., Perata, P., Futsuhara, Y.,and Yamaguchi, J. (1998) Sugar sensingand a-amylase gene expression in riceembryos. Planta, 204, 420–428.
85 Lalonde, S., Boles, E., Hellmann, H.,Barker, L., Patrick, J.W., Frommer, W.B.,and Wood, J.M. (1999) The dual action ofsugar carriers: transport and sugarsensing. Plant Cell, 11, 707–726.
86 Seki, M., Narusaki, M., Ishida, J., Nanjo,T., Fujita, M., Oone, Y., Kamiya, A.,Nakajima, M., Enju, A., Sakurai, T., Satou,M., Akiyama, K., Taji, T., Shinozaki, K.Y.,Carninci, P., Kawai, J., Hayashizaki, K.Y.,and Shinozaki, K. (2002) Monitoring theexpression profiles of 7000 Arabidopsisgenes under drought, cold and highsalinity stresses using a full length cDNAmicro array. Plant J., 31, 279–292.
87 Rodriguez, M.C.S., Petersen, M., andMundy, J. (2010) Mitogen-activated proteinkinase signalling in plants. Annu. Rev.Plant. Biol., 61, 621–649.
88 Galston, A.W. (1991) On the trail of a newregulatory system in plants. New Biol., 3,450–453.
89 Alc�azar, R., Altabella, T., Marco, F.,Bortolotti, C., Reymond, M., Koncz, C.,Carrasco, P., and Tiburcio, A.F. (2010)Polyamines: molecules with regulatoryfunctions in plant abiotic stress tolerance.Planta, 231, 1237–1249.
90 Igarashi, K. and Kashiwagi, K. (2000)Polyamines: mysterious modulators ofcellular functions. Biochem. Biophys. Res.Commun., 271, 559–564.
91 Seiler, N. and Raul, F. (2005) Polyaminesand apoptosis. J. Cell Mol. Med., 9, 623–642.
92 Alc�azar, R., Marco, F., Cuevas, J.C., Patr�on,M., Ferrando, A., Carrasco, P., Tiburcio, A.F., and Altabella, T. (2006) Involvement ofpolyamines in plant response to abioticstress. Biotechnol. Lett., 28, 1867–1876.
93 Kusano, T., Berberich, T., Tateda, C., andTakahashi, Y. (2008) Polyamines: essentialfactors for growth and survival. Planta,228, 367–381.
94 Bagni, N. and Tassoni, A. (2001)Biosynthesis, oxidation and conjugation ofaliphatic polyamines in higher plants.Amino Acids, 20, 301–317.
748 27 Metabolome Analyses for Understanding Abiotic Stress Responses in Plants

95 Gill, S. and Tuteja, N. (2010) Polyaminesand abiotic stress tolerance in plants. PlantSignal. Behav., 5, 26–33.
96 Groppa, M.D. and Benavides, M.P. (2008)Polyamines and abiotic stress: recentadvances. Amino Acids, 34, 35–45.
97 Alc�azar, R., Cuevas, J.C., Planas, J., Zarza,X., Bortolotti, C., Carrasco, P., Salinas, J.,Tiburcio, A.F., and Altabella, T. (2011)Integration of polyamines in the coldacclimation response. Plant Sci., 180,31–38.
98 Hussain, S.S., Ali, M., Ahmad, M., andSiddique, K.H. (2011) Polyamines: naturaland engineered abiotic and biotic stresstolerance in plants. Biotechnol. Adv., 29,300–311.
99 Urano, K., Yoshiba, Y., Nanjo, T., Ito,T., Yamaguchi-Shinozaki, K., andShinozaki, K. (2004) Arabidopsis stress-inducible gene for argininedecarboxylase AtADC2 is required foraccumulation of putrescine in salttolerance. Biochem. Biophys. Res.Commun., 313, 369–375.
100 Cuevas, J.C., L�opez-Cobollo, R., Alc�azar,R., Zarza, X., Koncz, C., Altabella, T.,Salinas, J., Tiburcio, A.F., and Ferrando, A.(2008) Putrescine is involved in Arabidopsisfreezing tolerance and cold acclimation byregulating abscisic acid levels in responseto low temperature. Plant Physiol., 148,1094–1105.
101 Capell, T., Bassie, L., and Christou, P.(2004) Modulation of the polyaminebiosynthetic pathway in transgenic riceconfers tolerance to drought stress. Proc.Natl. Acad. Sci. USA, 101, 9909–9914.
102 Alet, A.I., S�anchez, D.H., Ferrando, A.,Tiburcio, A.F., Alcazar, R., Cuevas, J.C.,Altabella, T., Pico, F.M., Carrasco-Sorli, P.,Men�endez, A.B., and Ruiz, O.A. (2011)Homeostatic control of polyamine levelsunder long-term salt stress in Arabidopsis:changes in putrescine content do notalleviate ionic toxicity. Plant Signal. Behav.,6, 237–242.
103 Santner, A. and Estelle, M. (2010) Theubiquitin–proteasome system regulatesplant hormone signalling. Plant J., 61,1029–1040.
104 Argueso, C.T., Ferreira, F.J., and Kieber, J.J. (2009) Environmental perceptionavenues: the interaction of cytokinin and
environmental response pathways. PlantCell Environ., 32, 1147–1160.
105 Messing, S.A.J., Gabelli, S.B., Echeverria,I., Vogel, J.T., Guan, J.C., Tan, B.C., Klee,H.J., McCarty, D.R., and Amzel, L.M.(2010) Structural insights into maizeViviparous14, a key enzyme in thebiosynthesis of the phytohormone abscisicacid. Plant Cell, 22, 2970–2980.
106 Wang, L., Wang, Z., Xu, Y., Joo, S.H., Kim,S.K., Xue, Z., Xu, Z., Wang, Z., andChong, K. (2009) OsGSR1 is involved incrosstalk between gibberellins andbrassinosteroids in rice. Plant J., 57,498–510.
107 Crozier, A., Kamiya, Y., Bishop, G., andYokota, T. (2000) Biosynthesis of hormonesand Elicitor Molecules, in Biochemistry andMolecular Biology of Plants (eds B.B.BuchananW., Gruissem and R.L. Jones),American Society of Plant Biology,Rockville, MD, pp. 850–929.
108 Taiz, T.L. and Zeiger, E. (2010) PlantPhysiology, 5th edn, Sinauer Associates,Boston, MA.
109 Williams, M.E. (2010) Introduction tophytohormones. Plant Cell, 22, 1–9.
110 Cramer, G.R. (2010) Abiotic stress andplant responses from the whole vine to thegenes. Aust. J. Grape Wine Res., 16, 86–93.
111 Hubbard, K.E., Nishimura, N., Hitomi, K.,Getzoff, E.D., and Schroeder, J.I. (2010)Early signal transduction mechanisms:newly discovered components andnewly emerging questions. Genes Dev., 24,1695–1708.
112 Chae, L., Pandey, G.K., Luan, S., Cheong,Y.H., and Kim K.-N. (2010) Protein kinasesand phosphatases for stress signaltransduction in plants, in Abiotic StressAdaptation in Plants: Physiological,Molecular and Genomic Foundation (eds A.Pareek, S.K. Sopory, H.J. Bohnert andGovindjee), Springer, Dordrecht,pp. 123–163.
113 Chinnusamy, V., Gong, Z., and Zhu, J.K.(2008) ABA-mediated epigenetic processesin plant development and stress responses.J. Integr. Plant Biol., 50, 1187–1195.
114 Yamaguchi-Shinozaki, K. and Shinozaki,K. (2006) Transcriptional regulatorynetworks in cellular responses andtolerance to dehydration and cold stresses.Annu. Rev. Plant Biol., 57, 781–803.
References 749

115 Wilkinson, S. and Davies, W.J. (2010)Drought, ozone, ABA and ethylene: newinsights from cell to plant to community.Plant Cell Environ., 33, 510–525.
116 Cutler, S.R., Rodriguez, P.L., Finkelstein,R.R., and Abrams, S.R. (2010) Abscisicacid: emergence of a core signallingnetwork. Annu. Rev. Plant Biol., 61,651–679.
117 Fang, J., Chai, C., Qian, Q., Li, C., Tang,J., Sun, L., Huang, Z., Guo, X., Sun, C.,and Liu, M. (2008) Mutations of genes insynthesis of the carotenoid precursors ofABA lead to pre-harvest sprouting andphoto oxidation in rice. Plant J., 54,177–189.
118 Vankova, R. (2012) Abscisic acidsignalling in plants, in Abiotic StressResponses in Plants – Metabolism,Productivity and Sustainability (eds P.Ahmad and M.N.V. Prasad), Springer,New York, pp. 359–368.
119 Agarwal, P.K. and Jha, B. (2010)Transcription factors in plants and ABAdependent and independent abiotic stresssignalling. Biol. Plant., 54, 201–212.
120 Xue-Xuan, X., Hong-Bo, S., Yuan-Yuan,M., Gang, X., Jun-Na, S., Dong-Gang, G.,and Cheng-Jiang, R. (2010)Biotechnological implications fromabscisic acid (ABA) roles in cold stress andleaf senescence as an important signal forimproving plant sustainable survival underabiotic-stressed conditions. Crit. Rev.Biotechnol., 30, 222–230.
121 Munn�e-Bosch, S. and Pe~nuelas, J. (2003)Photo and antioxidative protection, and arole for salicylic acid during drought andrecovery in field grown Phillyreaangustifolia plants. Planta, 217, 758–766.
122 Chini, A., Grant, J.J., Seki, M., Shinozaki,K., and Loake, G.J. (2004) Droughttolerance established by enhancedexpression of the CCI-NBS-LRR gene,ADR1, requires salicylic acid, EDS1 andABI1. Plant J., 38, 810–822.
123 Janda, T., Szalai, G., Tari, I., and P�aldi, E.(1999) Hydroponic treatment with salicylicacid decreases the effects of chilling injuryin maize (Zea mays L.) plants. Planta, 208,175–180.
124 Kang, H.M. and Saltveit, M.E. (2002)Chilling tolerance of maize, cucumber andrice seedling leaves and roots are
differentially affected by salicylic acid.Physiol. Plant., 115, 571–576.
125 Metwally, A., Finkemeier, I., Georgi, M.,and Dietz, K.J. (2003) Salicylic acidalleviates the cadmium toxicity in barleyseedlings. Plant Physiol., 132, 272–281.
126 Yang, Z.M., Wang, J., Wang, S.H., and Xu,L.L. (2003) Salicylic acid-inducedaluminium tolerance by modulation ofcitrate efflux from roots of Cassia tora L.Planta, 217, 168–174.
127 Freeman, J.L., Garcia, D., Kim, D., Hopf,A.M, and Salt, D.E. (2005) Constitutivelyelevated salicylic acid signals glutathione-mediated nickel tolerance in Thlaspi nickelhyper-accumulators. Plant Physiol., 137,1082–1091.
128 Larkindale, J. and Knight, M.R. (2002)Protection against heat stress inducedoxidative damage in Arabidopsis involvescalcium, abscisic acid, ethylene, andsalicylic acid. Plant Physiol., 128, 682–695.
129 Larkindale, J., Hall, J.D., Knight, M.R., andVierling, E. (2005) Heat stress phenotypesof Arabidopsismutants implicate multiplesignalling pathways in the acquisitionof thermotolerance. Plant Physiol., 138,882–897.
130 Borsani, O., Valpuesta, V., and Botella,A. (2001) Evidence for a role of salicylicacid in the oxidative damage generatedby NaCl and osmotic stress inArabidopsis seedlings. Plant Physiol.,126, 1024–1030.
131 Rivas-San Vicente, M. and Plasencia, J.(2011) Salicylic acid beyond defense: itsrole in plant growth and development.J. Exp. Bot., 62, 3321–3338.
132 Dat, J.F., Lopez-Delgado, H., Foyer, C.H.,and Scott, I.M. (1998) Parallel changes inH2O2 and catalase during thermotoleranceinduced by salicylic acid or heatacclimation in mustard seedlings. PlantPhysiol., 116, 1351–1357.
133 Chakraborty, U. and Tongden, C. (2005)Evaluation of heat acclimation and SAtreatments as potent inducers ofthermotolerance in Cicer arietinum L. Curr.Sci., 89, 384–389.
134 Sharma, Y.K., Leon, J., Raskin, I., andDavis, K.R. (1996) Ozone-inducedresponses in Arabidopsis thaliana: the roleof salicylic acid in the accumulation ofdefense-related transcripts and induced
750 27 Metabolome Analyses for Understanding Abiotic Stress Responses in Plants

resistance. Proc. Natl. Acad. Sci. USA, 93,5099–5104.
135 Rao, M.V. and Davis, R.D. (1999) Ozone-induced cell death occurs via two distinctmechanisms in Arabidopsis: the role ofsalicylic acid. Plant J., 1, 603–614.
136 Xiang, C. and Oliver, D.J. (1998)Glutathione metabolic genes co-ordinately respond to heavy metals andjasmonic acid in Arabidopsis. Plant Cell,10, 1539–1550.
137 Sasaki, T., Kajino, T., Bo, L., Sugiyama, H.,and Takahashi, H. (2001) New pulpbiobleaching system involving manganeseperoxidase immobilized in a silica supportwith controlled pore sizes. Appl. Environ.Microbiol., 67, 2208–2212.
138 Acosta, I.F. and Farmer, E.E. (2010)Jasmonates. Arabidopsis Book, 8, e0129.
139 Onkokesung, I., Dahl, C.C.V., Matsuoka,K., Saluz, Hans-Peter, and Baldwin, I.T.(2010) Jasmonic acid and ethylenemodulate local responses to wounding andsimulated herbivory in Nicotiana attenuataLeaves1 [W] [OA] Nawaporn. Plant Physiol.,153, 785–798.
140 Mahouachi, J., Arbona, V., and Gomez-Cadenas, A. (2007) Hormonal changes inpapaya seedlings subjected to progressivewater stress and re-watering. Plant GrowthRegul., 53, 43–51.
141 Razem, F.A., El-Kereamy, A., Abrams, S.R., and Hill, R.D. (2006) The RNA-bindingprotein FCA is an abscisic acid receptor.Nature, 439, 290–294.
142 Morgan, P.G. and Drew, M.C. (1997)Ethylene and plant responses to stress.Physiol. Plant., 100, 620–630.
143 Cao, W.H., Liu, J., He, X.J., Mu, R.L.,Zhou, H.L., and Chen, S.Y. (2007)Modulation of ethylene responses affectsplant salt-stress responses. Plant Physiol.,143, 707–719.
144 Mittler, R. (2006) Abiotic stress, the fieldenvironment and stress combination.Trends Plant Sci., 11, 15–19.
145 Foyer, C.H. and Noctor, G. (2003) Redoxsensing and signalling associated withreactive oxygen in chloroplasts,peroxisomes and mitochondria. Physiol.Plant., 119, 355–364.
146 Moller, I.M. (2001) Plant mitochondria andoxidative stress: electron transport,NADPH turnover and metabolism of
reactive oxygen species. Annu. Rev. PlantPhysiol. Plant Mol. Biol., 52, 561–591.
147 Asada, K. (2006) Production andscavenging of reactive oxygen species inchloroplasts and their functions. PlantPhysiol., 141, 391–396.
148 Mittler, R., Vanderauwera, S., Suzuki, N.,Miller, G., Tognetti, V.B., Vandepoele, K.,Gollery, M., Shulaev, V., and VanBreusegem, F. (2011) ROS signalling: thenew wave? Trends Plant Sci.,16, 300–309.
149 Molassiotis, A. and Fotopoulos, V. (2011)Oxidative and nitrosative signalling inplants: two branches in the same tree?Plant Signal. Behav., 6, 210–214.
150 Mittler, R, Vanderauwera, S, Gollery, M.,and Van Breusegem, F. (2004) Reactiveoxygen gene network of plants. TrendsPlant Sci., 9, 490–498.
151 Mittler, R. (2002). Oxidative stress,antioxidants and stress tolerance. TrendsPlant Sci., 7, 405–410.
152 Apel, K. and Hirt, H. (2004) Reactiveoxygen species: metabolism, oxidativestress, and signal transduction. Annu. Rev.Plant Biol., 55, 373–399.
153 Mahajan, S. and Tuteja, N. (2005) Cold,salinity and drought stresses: anoverview. Arch. Biochem. Biophys., 444,139–158.
154 Tuteja, N. (2007) Mechanisms of highsalinity tolerance in plants.MethodsEnzymol., 428, 419–438.
155 Tuteja, N. and Hirt, H. (2010) Cold, saltand drought stress, in Plant Stress Biology:From Genomics Towards System Biology(ed. H. Hirt), Wiley-Blackwell, Weinheim,pp. 137–159.
156 Khan, N.A. and Singh, S. (2008) AbioticStress and Plant Responses, IK International,New Delhi.
157 Grat~ao, P.L., Polle, A., Lea, P.J., andAzevedo, R.A. (2005) Making the life ofheavy metal-stress plants a little easier.Funct. Plant Biol., 32, 481–494.
158 Harper, J.F., Breton, G., and Harmon, A.(2004) Decoding Ca2þ signals throughplant protein kinases. Annu. Rev. PlantBiol., 55, 263–288.
159 Ludwig, A.A., Romeis, T., and Jones, J.D.G. (2004) CDPK-mediated signalingpathways: specificity and cross-talk. J. Exp.Bot., 55, 181–188.
References 751

160 Hrabak, E.M., Chan, C.W., and Gribskov,M. (2003) The Arabidopsis CDPK–SnRKsuper family of protein kinases. PlantPhysiol., 132, 666–680.
161 Hirayama, T. and Shinozaki, K. (2010)Research on plant abiotic stress responsesin the post-genome era: past, present andfuture. Plant J., 61, 1041–1052.
162 Mahajan, S., Pandey, G.K., and Tuteja, N.(2008) Calcium and salt stress signaling inplants: shedding light on SOS pathway.Arch. Biochem. Biophys., 471, 146–158.
163 Luan, S. (2009) The CBL–CIPK network inplant calcium signaling. Trends Plant Sci.,14, 37–42.
164 Romeis, T., Ludwig, A.A., Martin, R., andJones, J.D.G. (2001) Calcium-dependentprotein kinases play an essential role in aplant defense response. EMBO J., 20,5556–5567.
165 Urao, T., Katagiri, T., Mizoguchi, T.,Yamaguchi-Shinozaki, K., Hayashida, N.,and Shinozaki, K. (1994) Two genes thatencode Ca2þ-dependent protein kinasesare induced by drought and high-saltstresses in Arabidopsis thaliana.Mol. Gen.Genet., 244, 331–340.
166 Taehtiharju, S., Sangwan, V., Monroy, A.F.,Dhindsa, R.S., and Borg, M. (1997) Theinduction of kin genes in cold-acclimatingArabidopsis thaliana. Evidence of a role forcalcium. Planta, 203, 442–447.
167 Mori, I.C., Murata, Y., Yang, Y., Munemasa,S., Wang, Y.F., Andreoli, S., Tiriac, H.,Alonso, J.M., Harper, J.F., Ecker, J.R.,Kwak, J.M., and Schroeder, J.I. (2006)CDPKs CPK6 and CPK3 function in ABAregulation of guard cell S-type anion- andCa2þ permeable channels and stomatalclosure. PLoS Biol., 4, e327.
168 Choi, H., Park, H.J., Park, J.H., Kim, S.,Im, M.Y., Seo, H.H., Kim, Y.W., Hwang, I.,and Kim, S.Y. (2005) Arabidopsis calcium-dependent protein kinase CPK32 interactswith ABF4, a transcriptional regulator ofabscisic acid-responsive gene expression,and modulates its activity. Plant Physiol.,139, 1750–1761.
169 Zhu, S.Y., Yu, X.C., Wang, X.J., Zhao,R., Li, Y., Fan, R.C., Shang, Y., Du, S.Y.,Wang, X.F., Wu, Y.H., Zhang, X.Y., andZhang, D.P. (2007) Two calcium-dependent protein kinases, CPK4 andCPK11, regulate abscisic acid signal
transduction in Arabidopsis. Plant Cell,19, 3019–3036.
170 Sinha, A.K., Jaggi, M., Raghuram, B., andTuteja, N. (2011) Mitogen-activated proteinkinase signalling in plants under abioticstress. Plant Signal. Behav., 6, 196–203.
171 Ichimura, K., Shinozaki, K., and Tena, G.(2002) Mitogen-activated protein kinasecascades in plants: a new nomenclature.Trends Plant Sci., 7, 301–308.
172 Mizoguchi, T., Irie, K., Hirayama, T.,Hayashida, N., Yamaguchi-Shinozaki, K.,Matsumoto, K., and Shinozaki, K. (1996) Agene encoding a mitogen-activated proteinkinase is induced simultaneously withgenes for a mitogen-activated proteinkinase and S6 ribosomal protein kinase bytouch, cold, and water stress in Arabidopsisthaliana. Proc. Natl. Acad. Sci. USA, 93,765–769.
173 Teige, M., Scheikl, E., Eulgem, T., Doczi,R., Ichimura, K., Shinozaki, K, Dangl, J.L.,and Hirt, H. (2004) The MKK2 pathwaymediates cold and salt stress signalling inArabidopsis.Mol. Cell, 15, 141–152.
174 Komatsu, S. and Ahsan, N. (2009) Soybeanproteomics and its application tofunctional analysis. J. Proteomics, 72,325–336.
175 Feist, A.M. and Palsson, B.O. (2008) Thegrowing scope of applications of genome-scale metabolic reconstructions usingEscherichia coli. Nat. Biotechnol., 26,659–667.
176 Javot, H., Lauvergeat, V., Santoni, V.,Martin-Laurent, F., G€uxcl€u, J., Vinh, J.,Heyes, J., Franck, K.I., Sch€affner, A.R.,Bouchez, D. et al. (2003) Role of a singleaquaporin isoforms in root water uptake.Plant Cell, 15, 509–522.
177 Sairam, R.K. and Tyagi, A. (2004)Physiology and molecular biology ofsalinity stress tolerance in plants. Curr.Sci., 86, 407–421.
178 Brini, F., Gaxiola, R.A., Berkowitz, G.A.,and Masmoudi, K. (2005) Cloning andcharacterization of a wheat vacuolarcation/proton antiporter andpyrophosphatase proton pump. PlantPhysiol. Biochem., 43, 347–354.
179 Brini, F., Hanin, M., Lumbreras, V., Irar,S., Pag�es, M., and Masmoudi, K. (2007)Functional characterization of DHN-5, adehydrin showing a differential
752 27 Metabolome Analyses for Understanding Abiotic Stress Responses in Plants

phosphorylation pattern in two Tunisiandurum wheat (Triticum durum Desf.)varieties with marked differencesin salt and drought tolerance. Plant Sci.,172, 20–28.
180 Masle, J., Gilmore, S.R., and Farquhar, G.D. (2005) The ERECTA gene regulatesplant transpiration efficiency inArabidopsis. Nature, 436, 866–870.
181 Chuang, H., Harnrak, A., Chen, Y., andHsu, C. (2010) A harpin-induced ethylene-responsive factor regulates plant growthand responses to biotic and abioticstresses. Biophys. Res. Commun.,402, 410–420.
182 Chen, X., Shang, J., Chen, D., Lei, C., Zou,Y., Zhai, W., Liu, G., Xu, J., Ling, Z., Cao,G., Ma, B., Wang, Y., Zhao, X., Li, S., andZhu, L. (2006) A B-lectin receptor kinasegene conferring rice blast resistance. PlantJ., 46, 794–804.
183 Hashimoto, M., Kisseleva, L., Sawa, S.,Furukawa, T., Komatsu, S., and Koshiba, T.(2004) A novel rice PR10 protein,RSOsPR10, specifically induced in roots bybiotic and abiotic stresses, possibly via thejasmonic acid-signalling pathway. PlantCell Physiol., 45, 550–559.
184 Kreps, J.A., Wu, Y, Chang, H.-S., Zhu, T.,Wang, X., and Harper, J.F. (2002)Transcriptome changes for Arabidopsis inresponse to salt, osmotic, and cold stress.Plant Physiol., 130, 2129–2141.
185 Oono, Y., Seki, M., Satou, M., Iida, K.,Akiyama, K., Sakurai, T., Fujita, M.,Yamaguchi-Shinozaki, K., and Shinozaki,K. (2006) Monitoring expression profiles ofArabidopsis genes during cold acclimationand deacclimation using DNAmicroarrays. Funct. Integr. Genomics, 6,212–234.
186 Jianping, P.W. and Suleiman, S.B. (2007)Monitoring of gene expression profiles andidentification of candidate genes involvedin drought responses in Festuca mairei.Mol. Genet. Genomics, 277, 571–587.
187 Mantri, N., Ford, R., Coram, T., and Pang,E. (2007) Transcriptional profiling ofchickpea genes differentially regulated inresponse to high-salinity, cold and drought.BMC Genomics, 8, 303.
188 Gregory, B., O’Malley, R., Lister, R., Urich,M., and Tonti-Filippini, J. (2008) A linkbetween RNA metabolism and silencing
affecting Arabidopsis development. Dev.Cell, 14, 854–866.
189 Matsui, A., Ishida, J., Morosawa, T.,Mochizuki, Y., Kaminuma, E., Endo, T.A.,Okamoto, M., Nambara, E., Nakajima, M.,Kawashima, M., Masakazu Satou, M., Kim,J.M., Kobayashi, N., Toyoda, T., Shinozaki,K., and Seki, M. (2008) Arabidopsistranscriptome analysis under drought,cold, high-salinity and ABA treatmentconditions using a tiling array. Plant CellPhysiol., 49, 1135–1149.
190 Bartel, D.P. (2004) MicroRNAs: genomics,biogenesis, mechanism, and function. Cell,116, 281–297.
191 Reinhart, B.J., Weinstein, E.G., Rhoades,M.W., Bartel, B., and Bartel, D.P. (2002)MicroRNAs in plants. Genes Dev., 16,1616–1626.
192 Llave, C., Kasschau, K.D., Rector, M.A.,and Carrington, J.C. (2002) Endogenousand silencing-associated small RNAs inplants. Plant Cell, 14, 1605–1619.
193 Park, W., Li, J., Song, R., Messing, J., andChen, X. (2002) CARPEL FACTORY, aDicer homolog, and HEN1, a novelprotein, act in microRNA metabolismin Arabidopsis thaliana. Curr. Biol., 12,1484–1495.
194 Lu, X.Y. and Huang, X.L. (2008) PlantmiRNAs and abiotic stress responses.Biochem. Biophys. Res. Commun., 368,458–462.
195 Bonnet, E., Wuyts, J., Rouze, P., and Van,Y.P. (2004) Evidence that microRNAprecursors, unlike other non-coding RNAs,have lower folding free energies thanrandom sequences. Bioinformatics, 20,2911–2917.
196 Jones-Rhoades, M.W. and Bartel, D.P.(2004) Computational identification ofplant microRNAs and their targets,including a stress-induced miRNA.Mol.Cell, 14, 787–799.
197 Sunkar, R., Kapoor, A., and Zhu, J.K.(2006) Post-transcriptional induction oftwo Cu/Zn superoxide dismutase genes inArabidopsis is mediated by down regulationof miR398 and important for oxidativestress tolerance. Plant Cell, 18, 2051–2065.
198 Sunkar, R. and Zhu, J.K. (2004) Novel andstress-regulated microRNAs and othersmall RNAs from Arabidopsis. Plant Cell,16, 2001–2019.
References 753

199 Grover, S., Talwar, P., Baghel, R., Kaur, H.,Gupta, M., Devi, M.G., Bala, K., Sharma,S., and Kukreti, R. (2010). Geneticvariability in estrogen disposition:potential clinical implications forneuropsychiatric disorders. Am. J. Med.Genet. B., 153, 1391–1410.
200 Varshney, R.K., Bansal, K.C., Aggarwal, P.K., Datta, S.K., and Craufurd, P.Q. (2011)Agricultural biotechnology for cropimprovement in a variable climate: hope orhype? Trends Plant Sci., 16, 363–371.
201 Xiao, B.Z., Chen, X., Xiang, Cheng-Bin,Tang, N., Zhang, Qi-Fa, and Xiong,
Li-Zhong (2009) Evaluation of sevenfunction-known candidate genes for theireffects on improving drought resistance oftransgenic rice under field conditions.Mol.Plant., 2, 73–83.
202 Oh, S.J., Oh, S.J., Kim, Y.S., Kwon, C.W.,Park, H.K., Jeong, J.S., and Kim, J.K.(2009) Overexpression of the transcriptionfactor AP37 in rice improves grain yieldunder drought conditions. Plant Physiol.,150, 1368–1379.
203 James, C. (2009) Global Status ofCommercialized Biotech/GM Crops: ISAAABrief 41, ISAAA, Ithaca, NY.
754 27 Metabolome Analyses for Understanding Abiotic Stress Responses in Plants





























