Embed Size (px)
DESCRIPTION
Climate Change and Range Shifts. Hopkins Marine Station, site of Barry et al. 1995. Example 1: Barry et al. 1995. Hopkins Marine Station is located in Monterey Bay - PowerPoint PPT Presentation
Citation preview

Climate Change and Range Shifts
Hopkins Marine Station, site of Barry et al. 1995

Example 1: Barry et al. 1995
• Hopkins Marine Station is located in Monterey Bay
• In 1931, ecologists sampled a transect (red line) across the intertidal zone. They recorded the identities and densities of every species on the transect.
• In 1993, Barry and colleagues relocated bolts marking the transect and repeated the original survey.
• They then divided the species into three groups: (1) southern species, extending far to the south, but not far north of Monterey (2) northern species, extending far to the north but not south of Monterey, and (3) cosmopolitan species, occurring over a very broad latitudinal range.

Barry et al. 1995: changes in species composition
• They found that southern species increased abundances on the transects (red dots, frame A), northern species deceased abundances (blue dots, frame A, and cosmopolitan species showed no net change (green triangles, frame A). Solid symbols indicate statistically significant changes.
• Plot B is a different representation of the same trend: the means of the ratios of densities (1993/1932) are plotted with respect to species group.
• The tends are consistent with the interpretation that southern species are expanding- and northern species receding to higher latitudes. Such latitudinal rage shifts have been observed for terrestrial species.

Barry et al. 1995: changes in species composition
• southern species abundances ↑
• northern species abundances ↓
• cosmopolitan species: no change
• means of ratios of densities (1993/1932)
Trends are consistent with hypotheses that:
- southern spp. are expanding
- northern spp. receding to higher latitude
similar latitudinal rage shifts have been observed for terrestrial species
solid = P<0.05

Barry et al. 1995: temperature records
• Over the 60 years of the comparison, sea surface temperatures recorded at Hopkins Marine Station increased (plot A).
• Comparing 11-year time spans before the original survey and re-survey revealed higher temperatures preceding the re-survey, with the most extreme differences occurring in summer (plot B).
• They propose that the temperature rise favored southern species and disfavored northern species. However, the specific mechanism of effect is not known.

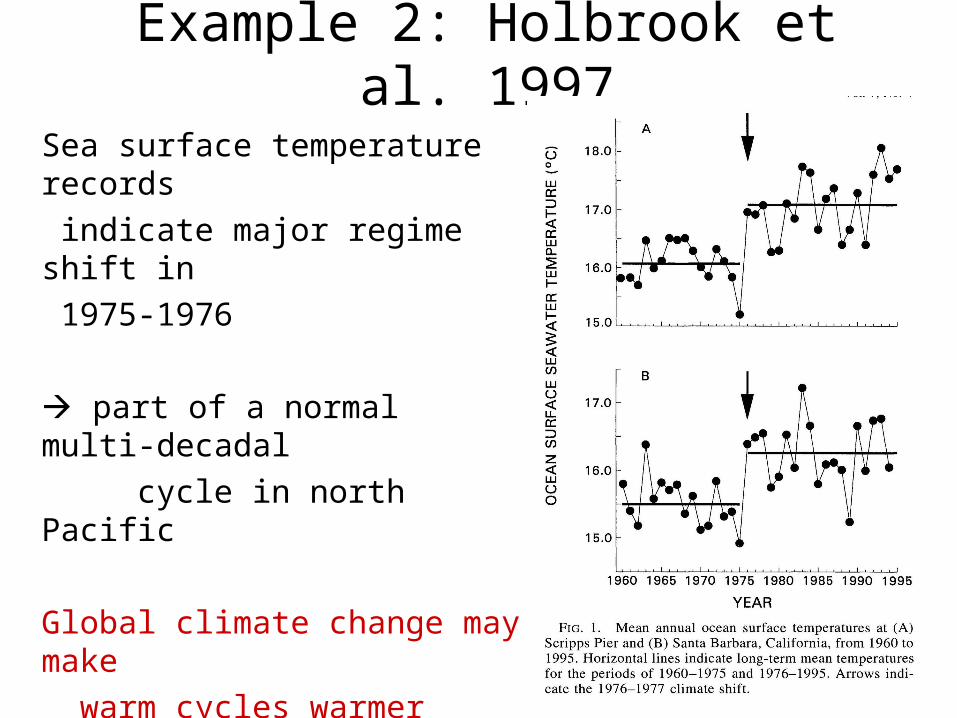
Example 2: Holbrook et al. 1997
•Sea surface temperature records indicate a major shift in temperature regimes occurring in 1975-1976.•Such shifts are part of a normal multi-decadal cycle in the north Pacific ocean, but global climate change appears to be affecting the cycles, making the upward part of the cycles greater than the downward, so that long term temperatures are ratcheting up.
Example 2: Holbrook et al. 1997
Sea surface temperature records
indicate major regime shift in
1975-1976
part of a normal multi-decadal
cycle in north Pacific
Global climate change may make
warm cycles warmer
long term temperatures are
ratcheting up

Holbrook et al. 1997: shift in the fish fauna
• They observed that after the shift in temperature regime (arrows, plots A and B) fishes species became less diverse at two survey sites (plot A), southern species increased and northern species declined in relative abundance (Plot B).

Holbrook et al. 1997
• The authors are careful to point out that the negative correlation between temperature and either fish diversity or the abundance of northern species does not reveal the mechanism of change.
• Higher sea surface temperature could directly affect fish physiology, lowering survival or reproduction of northern species. However, increases in surface temperatures could indirectly affect them by diminishing their food supplies. Cold surface water often causes high fertility, high surfaces temperatures often restrict nutrient availability, ocean productivity, and hence food availability.
• Or the correlation could be a shear coincidence. More work is needed.

Correlation vs causationNegative correlation between °C and (a) fish diversity, or (b) abundance of northern species, does not indicate any mechanism or causation for change
Higher sea surface temperature could have:
- direct effects on fish physiology, lowering either survival or reproduction of northern spp.
cold surface water often causes high fertility
- indirect effects – e.g., diminishing food supplies high surfaces temps can restrict nutrient availability, ocean productivity, and hence food availability
... or the correlation could be coincidence; more work needed

Example 3: Harley (2011)
Harley presents evidence to support of the idea that the extinction of mussel beds in some areas of the Salish Sea (Puget Sound) results from temperature stress with climate change.

Harley 2011: The hypothesis
• Harley bases his explanation on the refuge hypothesis (Connell 1970, Paine 1974)
• Under this hypothesis, the lower limits of the mussel zone are set by predation from the sea star Pisaster ochraceus. Upper limits are set by temperature stress.
• Wave exposure (topographic differences in average levels of wave action, including up-shore wave wash) affects temperature stress. On shores with high wave action, wave wash alleviates temperature stress, so that mussels can survive at relatively high shore levels. On shores with low wave action, wave wash does not reach up the shore, and temperature stress extends to lower shores levels, eliminating mussels from higher shore levels.
• The sea stars are unaffected by these differences in wave action, so that the lower boundary of the mussels (lower limit to the refuge) remains at the same shore level regardless of wave exposure.
• Therefore, from wave exposed to sheltered shores the vertical range of the refuge (predator-free space) is compressed, according to the hypothesis.

Diagram of the refuge Hypothesis (Connell 1970, Paine 1974)

A real mussel bed in an apparent spatial refuge.

Harley 2011
• Harley predicted that as temperature stress increases with global warming, invertebrate zones in sheltered areas should compress further, eventually causing extinction.
Wave exposed: broad zones Sheltered: compressed zones

Harley 2011: the sample designAlong a west-east transect, he compared temperature and zonation as it exists today (grey squares) and a change in mussel and barnacle zonation from 1957 to 2010 (gray circles). Lower graph shows low tide temperatures measured at sites indicated by squares.

Along east-west transects, compared: °C & zonation today change in mussel + barnacle zonation from 1957 to 2010 (gray circles)
low tide temperatures
measured at sites
Harley 2011: experimental design

Harley 2011: results from 2009The upper limits of barnacles and mussels measured on 2009 sites (squares in previous slide) are negatively correlated with temperature (plots A-D). Sea star heights measured at low tide show no statistically significant relationship with low tide temperatures (plot E).

Harley 2011: results from historical comparison
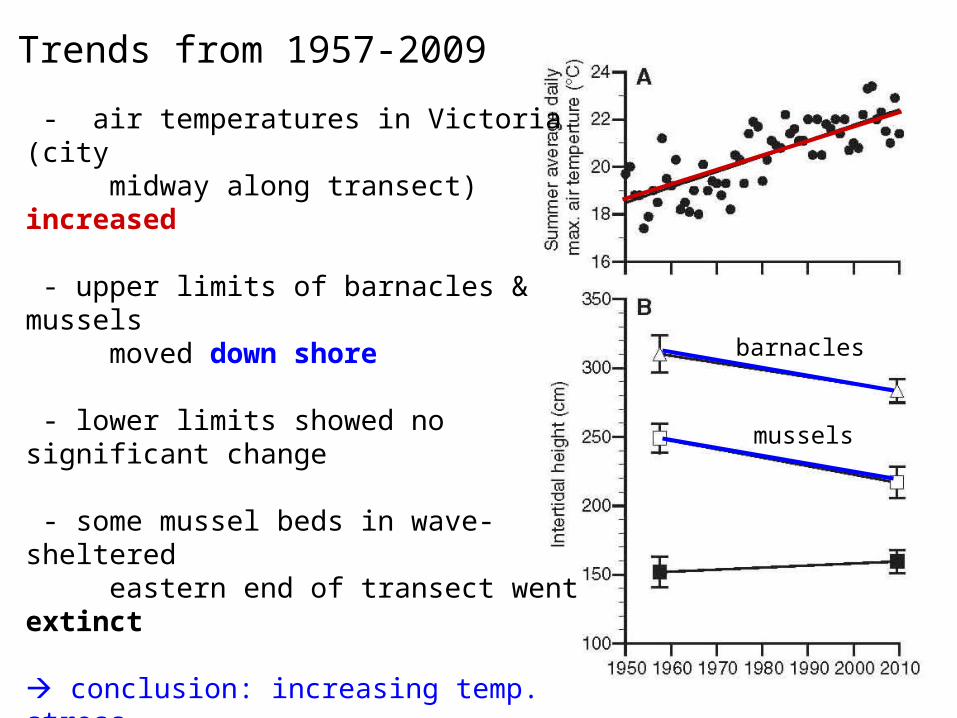
• From 1957 to 2009, air temperatures measured in Victoria, a city midway along the west-east transect, increased steadily (plot A).
• Over the same time span, upper limits of the two species of mussels declined down shore, while lower limits showed no statistically significant trend (plot B).
• Harley also observed that some mussel beds in the wave sheltered eastern extremity of the transect went extinct.
• He concludes that increasing temperature stress with global warming caused the upper limit of the refuge to fall, vertically compressing the mussel zone. The naturally compressed mussel zones in the east eventually vanished.
Trends from 1957-2009
barnacles
mussels
- air temperatures in Victoria (city midway along transect) increased
- upper limits of barnacles & mussels moved down shore
- lower limits showed no significant change
- some mussel beds in wave-sheltered eastern end of transect went extinct
conclusion: increasing temp. stress caused upper limit of refuge to drop, vertically compressing mussel zones - naturally compressed mussel zones in the east just disappeared

Imagining alternative explanations
• In the following slides we learn about… – Pisaster foraging behavior– Temperature effects on Pisaster feeding rates– the coincidence of environmental factors and
mussel bed loss in Southern California

About sea star foraging behavior….
• Most studies of Pisaster predation report densities made at low tide, when the sea stars are resting.
• But Pisaster moves with the tides to forage. On sheltered shores, where the lower limit of prey may occur at high shore levels, the sea stars may forage to high shore levels. On wave exposed sites, they do not forage as high. In plot A, the x-axis measures wave exposure as high tide bottom flow speed; the Y-axis is shore level. Open symbols indicate mean sea star heights at low tide, solid symbols indicate sea star heights at high tide), triangles show highest reach of seas stars, the extreme limit of high tides.
• Panel B shows the corresponding abundance (mean per cover) of mussels over the wave exposure gradient.
• Data are from sites in Barkley Sound, roughly 50 miles from the Salish Sea Transect.

Inviolable refuges?: Sea star “Browse Lines”
Sea stars sometimes forage abovethe lower boundary of mussel beds removing small, preferred prey, including barnacles and small mussels.Their foraging appears to be more flexible than assumed by the refuge hypothesis.

Feeding rates of Pisaster are temperature dependent
• Sanford (2000) observed feeding rates of Pisaster as sea surface temperatures changed with cycles of coastal upwelling. When water temperatures dropped, sea stars feeding rates decreased, when temperatures increased, feeding rates accelerated.
• Lab studies showed that sea star feeding rates were sensitive to water temperatures when submerged, but not to a normal range of air temperatures during simulated low tides.

Changes in mussel abundance on Bird Rock, Catalina Island, Southern California.
D. R. Blakeway and C. D. Robles, Department of Biological Sciences, California State University at Los Angeles, CA 90032.

Study siteon Bird Rock

Bird Rock east end, winter 1984

Bird Rock east end, March 1987

Bird Rock east end, April 1991

Bird Rock east end, March 1993

Bird Rock east end, October 1999

Bird Rock east end photo-mosaic, March 2001
Using a camera on a pole one can take overhead views, which are easier to analyze

Image analysis software can be used to covert panoramas to an overhead view
0
1
2
3
4
6
Time
5
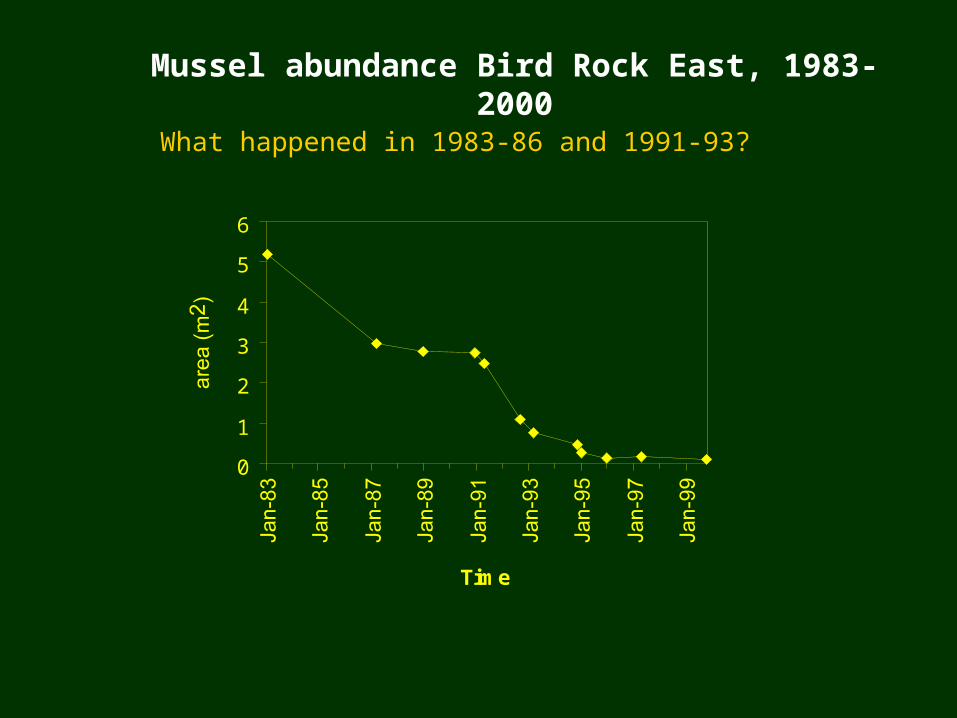
Mussel abundance Bird Rock East, 1983-2000
What happened in 1983-86 and 1991-93?

SST Catalina Ridge/Two Harbors, 1982-2001
Jan-82 Jan-84 Jan-86 Jan-88 Jan-90 Jan-92 Jan-94 Jan-96 Jan-98 Jan-00
25
20
15
5
25
20
15
5
Major el nino (warm water) events in blue

250
53
200
150
100
50
01950 20001990198019701960
years
56
52
51
5455
57 6061
5859
6233
6465
66
67
6869
74
72
75
78
81
85
77
7980 82
83
84
86
87
8889 90
9192
93
9495 96
9798
Zooplankton abundance, SoCal Bight, 1950-2000
Severe El nino years in orange.
Could the recent (since the late 1970s) decline in zooplankton abundance be the result of declining ocean production? If so, what are the implications for mussels?

Assuming that mussel zones are indeed collapsing in the Salish Sea, can you formulate an alternative hypothesis to Harley 2011?





























