Embed Size (px)
Citation preview

Glycobiology vol. 16 no. 11 pp. 1021–1032, 2006 doi:10.1093/glycob/cwl029 Advance Access publication on July 31, 2006
© The Author 2006. Published by Oxford University Press. All rights reserved. For permissions, please e-mail: [email protected] 1021
Cloning and biochemical characterization of the fucanase FcnA: definition of a novel glycoside hydrolase family specific for sulfated fucans
Sébastien Colin2, Estelle Deniaud2, Murielle Jam2, Valérie Descamps2, Yann Chevolot2, Nelly Kervarec3, Jean-Claude Yvin4, Tristan Barbeyron2, Gurvan Michel1,2,and Bernard Kloareg2
2Equipe Glycobiologie Marine, Centre National de la Recherche Scientifique, Université Pierre et Marie Curie-Paris6, Unité Mixte de Recherche 7139, Station Biologique, F-29682 Roscoff Cedex, Bretagne, France; 3Service Commun de Résonance Magnétique Nucléaire, Université de Bretagne Occidentale, Brest, Bretagne, France; and 4Laboratoires Goëmar, Saint Malo, Bretagne, France
Received on December 5, 2005; revised on June 23, 2006; accepted onJuly 18, 2006
Sulfated fucans are matrix polysaccharides from marinebrown algae, consisting of an a-L-fucose backbone substi-tuted by sulfate-ester groups, masked with ramifications, andcontaining other monosaccharide residues. We here report onthe characterization of a novel glycoside hydrolase (FcnA)specific for the degradation of sulfated fucans. This glycosidehydrolase was purified to electrophoretic homogeneity from aFlavobacteriaceae referred to as SW5. The gene fcnA wascloned and sequenced (3021 nucleotides), and the protein (1007amino acids) was produced in Escherichia coli. FcnA exhib-ited a modular architecture consisting of a 400-residue-longN-terminal domain followed by three repeated domains pre-dicted to adopt an immunoglobulin fold and by an 80-aminoacid-long C-terminal domain. A truncated recombinant proteinencompassing the N-terminal domain and the immunoglobulin-like repeats was shown to retain the enzyme activity. The N-terminal catalytic domain shared ~25% of sequence identitywith two patented fucanase genes, and these three fucanasesdelineate a new family of glycoside hydrolases. As shown bysize-exclusion chromatography (SEC) and 1H-NMR analyses,the fucanase FcnA proceeds according to an endolytic modeof action and cleaves the a-(1®4) glycosidic linkages withinthe blocks of repeating motifs [®4)-a-L-fucopyranosyl-2,3-disulfate-(1®3)-a-L-fucopyranosyl-2-sulfate-(1®]n.
Key words: brown algae/Flavobacteriaceae/fucanase/NMR/sulfated polysaccharide
Introduction
Marine macroalgae synthesize a great diversity of polysac-charides, which are used as cell wall constituents and forenergy storage. Like land plants, these eukaryotic phyla
produce neutral polysaccharides such as cellulose andstarch. However, marine algae are characterized by theirabundance of sulfated polysaccharides that have no equiva-lent in land plants (Kloareg and Quatrano, 1988). The redalgae produce sulfated galactans, agars, and carrageenans,which are commercially used for their gelling properties.The structures of agars and carrageenans have been exten-sively studied to understand and manipulate their physico-chemical properties. The cell walls of brown algae alsocontain sulfated polysaccharides known as sulfated fucans.These anionic polymers encompass a continuous spectrumof highly ramified polysaccharides, ranging from highuronic acid, low-sulfate–containing polymers with signifi-cant proportions of D-xylose, D-galactose, and D-mannoseto highly sulfated homofucan molecules with approxi-mately three substituents per disaccharide (Kloareg andQuatrano, 1988; Mabeau et al., 1990). In the cell wallmatrix, they associate with alginate, a linear polymer of β-D-mannuronic acid and α-L-guluronic acid (Kloareg andQuatrano, 1988). Sulfated fucans exhibit various biologicalactivities (Berteau and Mulloy, 2003), both as cell wallligands and as anticoagulant (Kropf et al., 1988; Goodnerand Quatrano, 1993; Pereira et al., 1999), antiviral, andantiproliferative agents. They were also shown to be recog-nized as fortuitous, defense elicitors in tobacco, where theyinduce systemic resistance against tobacco mosaic virus(Klarzynski et al., 2003). These properties have attracted arenewed interest to their fine chemical structure in the lastdecade. Sulfated fucan oligosaccharides of ∼8–14 fucoseunits, with a regular repeating disaccharide structure builton two residue types, 2-sulfated fucose and 2,3-disulfatedfucose, and with α-(1→3) and α-(1→4) glycosidic linkages,were isolated by chemical methods (Chevolot et al., 2001),indicating a significant degree of repetitiveness in the sul-fated fucan backbone. Yet the general complex structure ofsulfated fucans has remained somewhat elusive.
The algal polysaccharides constitute a crucial carbonsource for many marine bacteria. Those microorganismsdegrade the cell walls of marine algae by secreting specificglycoside hydrolases (Michel et al., 2006). We have so farisolated and structurally characterized several bacterialenzymes that depolymerize the sulfated galactans of redalgae, the family GH16 β-agarases and κ-carrageenases(Michel, Chantalat, Duee, et al., 2001; Allouch et al., 2003;Jam et al., 2005), and the ι-carrageenases, which constitutea distinct family of glycoside hydrolases, the family GH82(Barbeyron et al., 2000; Michel, Chantalat, Fanchon, et al.,2001; Michel et al., 2003). Both the κ- and ι-carrageenaseshave been recently used in combination with liquid chroma-tography and mass spectrometry techniques to analyze thecomplex structure of carrageenans (Antonopoulos,
The nucleotide sequence reported in this article has been submitted to the EMBL Nucleotide Sequence Database with accession number AJ877239.1To whom correspondence should be addressed; e-mail: [email protected]

S. Colin et al.
1022
Favetta, et al., 2005; Antonopoulos, Hardouin, et al.,2005). In this context, the isolation and expression ofhydrolases degrading sulfated fucans would be of specialinterest to further elucidate the fine chemical structure ofthese polysaccharides and to shed light on the structure ofhydrolases that degrade polysaccharides with high lineardensities of sulfate-ester substituents. To date, only twohighly similar sulfated fucan hydrolases, referred to asFda1 and Fda2, were cloned and expressed from the marinebacterium Alteromonas sp. SN-1009. These homologousenzymes have been patented (Takayama et al., 2002), butthey were not subjected to a detailed biochemical analysis.
On the basis of its capacity to degrade sulfated fucansfrom fucoid algae, we recently isolated a flavobacteriaceanstrain, SW5, which secretes sulfated fucan hydrolase activ-ity in its culture medium (Descamps et al., 2006). A crudeenzyme preparation from SW5 culture supernatant wasused to degrade the sulfated fucans from the brown algaPelvetia canaliculata. End products included a tetrasaccha-ride and a hexasaccharide made of the repetition of disac-charidic units consisting of [→4)-α-L-fucopyranosyl-2,3-disulfate-(1→3)-α-L-fucopyranosyl-2-sulfate-(1→]n, withthe 3-linked residues at the nonreducing end (Descampset al., 2006). We here report the purification, the cloning,and the overexpression of the sulfated fucan hydrolasefrom SW5, referred to as the fucanase FcnA. The proteinconsists of an N-terminal catalytic domain followed by threeimmunoglobulin-like repeats, and it harbors a C-terminaldomain conspicuous of proteins from Bacteroidetes bacteria.The enzyme depolymerizes sulfated fucans according to anendolytic mode of action and cleaves the α-(1→4) glyco-sidic linkage between 2-sulfated fucose and 2,3-disulfatedfucose residues. We also show that, together with Fda1and Fda2, FcnA belongs to a new family of glycosidehydrolases (Henrissat and Bairoch, 1996).
Results
Purification of an extracellular sulfated fucan-degrading enzyme from SW5
The purification of the sulfated fucan-degrading activity ofthe SW5 strain was based on a carbohydrate-polyacrylamidegel electrophoresis (C-PAGE) assay of the release ofanionic oligosaccharides from the sulfated fucan fractionreferred to as FS28 (Descamps et al., 2006). From the con-centrated bacterial culture supernatant, purification toelectrophoretic homogeneity was achieved in four steps:protein precipitation with ammonium sulfate at 40–60%saturation, hydrophobic interaction chromatography onPhenyl Sepharose, ion-exchange chromatography ondiethylaminoethyl (DEAE) Sepharose, and gel filtration onSuperdex 200. The active protein fractions corresponded to50, 35, 0.50, and 0.008% of the initial protein content, respec-tively. The latter protein fraction migrated as a single band insodium dodecyl sulfate (SDS)–PAGE analysis, correspond-ing to an apparent molecular weight of 105 kDa (Figure 1).This sulfated fucan-degrading enzyme is referred to as thefucanase FcnA. The SDS–PAGE 105-kDa protein band wasexcised, and three internal peptide sequences were determinedby Edman degradation, ITVDHVAGFTNLGNGAP,
TSGPDWLTIQQTDANS, and TANTTYGINTVASM,referred to as A, B, and C, respectively.
The fucanase gene fcnA
An internal gene probe of 203 nt was generated by poly-merase chain reaction (PCR) from the peptidic microse-quences A and B and used to screen a genomic libraryprepared with SW5 total DNA. Of the 6000 clones in thelibrary, three clones, referred to as pAT153fu11,pAT153fu14, and pAT153fu33 and with respective insertsizes of 4.0, 13.5, and 4.0 kb, hybridized with the probe.Southern blot experiments showed that SW5 strain con-tains only one copy of the fcnA gene. Restriction analysesindicated that the inserts from pAT153fu11 andpAT153fu33 were the same genomic DNA fragment,cloned in opposite directions, and that the pAT153fu14insert overlapped these fragments by ∼1.5 kb. ThepAT153fu11 and pAT153fu14 inserts were sequenced onboth strands. The two sequences perfectly overlapped overa length of 1610 nt, the fucanase open-reading frame (ORF)being interrupted at the 3′ end of pAT153fu11 insert andfinishing on the pAT153fu14 insert. The physical validity ofthis ORF was proven by PCR amplification from thegenomic DNA, leading to the cloning of the entire, 3021nt–long fucanase gene on a contig of 7485 nt. No obviousShine-Dalgarno ribosome-binding site consensus sequence
Fig. 1. Purification to homogeneity of SW5 fucanase, FcnA. The enzyme fraction after Superdex 200 HR was concentrated (Centricon 10 kDa), submitted to electrophoresis on a 12.5% SDS polyacrylamide gel (Phast-system, GE Healthcare, Orsay, France), and stained with Coomassie Blue (lane 1). The molecular weight markers (lane 2) consisted of lysozyme, soybean trypsin, carbonic anhydrase, ovalbumin, serum albumin, phosphorylase B, Escherichia coli β-galactosidase, and myosine (BioRad).

Fucanase, a novel family of glycoside hydrolases
1023
was found upstream of the fcnA gene (Kozak, 1999). Incontrast, 15 nt upstream of the ATG codon, a conservedmotif, AAxTAAAT, was present in the 5′ end of fcnA. Noobvious hairpin was identified at its 3′ end as transcriptionterminator.
The fucanase FcnA displays a modular architecture
The predicted product of the fucanase gene from SW5strain, FcnA, is a preprotein of 1007 amino acids, with atheoretical molecular mass of 110.3 kDa, consistent withthe apparent molecular mass of the secreted protein onSDS–PAGE analysis, 105 kDa (Figure 1). It includes thethree internal peptides determined by microsequencing.Based on the prediction program SignalP V2.0 (Nielsenet al., 1999), a signal peptide is likely to be cleaved betweenA28 and Q29. Using BLASTp and PSI-BLAST searches(Altschul et al., 1997), 60% of the sequence did not shareany significant similarity with the protein sequences available
in GENBANKnr. Only the core and the C-terminal end ofFcnA brought significant matches. However, a BLASTpsearch of the PAT database (NCBI) provided hits with twopatented fucanase sequences from Alteromonas sp. SN-1009 (Takayama et al., 2002), referred to as Fda1(AAO00508) and Fda2 (AAO00509) (E-values: 2E-9 and2E-7, respectively). The three proteins shared a largedomain in their N-terminal ends (∼380 residues long). Fda1and Fda2 are highly similar, displaying 70% identity overtheir entire length, 814 and 881 amino acids, respectively,whereas they displayed 24 and 23% sequence pairwise iden-tities (38 and 37% sequence similarity), respectively, withFcnA over their common N-terminal domain. To furtherassess the significance of these sequence similarities withFcnA, we compared the hydrophobic cluster analysis (HCA)plots of the N-terminal domain of Fda2 and FcnA (Figure 2)(Lemesle-Varloot et al., 1990). HCA plots revealed a similardistribution of the hydrophobic clusters, confirming thatthese domains share similar secondary-structure elements.
Fig. 2. The N-terminal catalytic domain of the fucanase FcnA. (A) HCA of the N-terminal domain of the fucanases FcnA from SW5 strain and Fda2 from Alteromonas sp. SN-1009. The different colors are used to help the visualization of the homologous hydrophobic clusters in both sequences. The conserved hydrophilic residues appear on a red background. The pairwise HCA score was calculated according the following formula: HCA score = 2CR/(RC1 + RC2) × 100, where RC1 and RC2 are the numbers of hydrophobic residues in sequences 1 and 2, respectively, and CR is the number of hydrophobic resi-dues in sequence 1 that are in exact correspondence with those in sequence 2 (Gaboriaud et al., 1987; Lemesle-Varloot et al., 1990). The final HCA score was the average value from all of the hydrophobic clusters delineated by the vertical lines. Amino acids are indicated by the one-letter code and by a star for proline, a black diamond for glycine, a square for threonine, and a dotted square for serine. The HCA plot was manually transformed in a linear form (B) to calculate the identity and similarity scores according the Blosum 62 matrix (sequence identity, 26%; similarity, 44%).

S. Colin et al.
1024
The HCA score based on the correspondences shown inFigure 2 was 74%, a value >60%, which is considered as asignificant threshold in this comparison procedure(Lemesle-Varloot et al., 1990).
In the protein core, RADAR searches detected threerepeated domains of ∼105 residues each (repeats R1, G419-D527; R2, G528-N633; and R3, G634-N737; Figure 3A),which share 31% (R1/R2), 27% (R1/R3), and 48% (R2/R3)pairwise sequence identities (Heger and Holm, 2000). Inthis region, searches on the Pfam database identified threeconsecutive He-PIg domains (PF05345) (Bateman et al.,2004). These domains are predicted to adopt an immuno-globulin fold. A BLASTp search in GENBANKnr usingthe three repeated domains identified mainly bacterialhypothetical conserved proteins, with various annotationssuch as PKD domain protein (NP_953122, 4E-13), putativeimmunoglobulin precursor (ZP_00525205, 2E-9), or RTXtoxin (ZP_00315133, 4E-12). The matching regions are sim-ilar repeated domains with pairwise sequence identitiesranging from 20 to 36% (from 35 to 52% sequence similari-ties). These matching domains were assigned to the He-PIg,cadherin, or PKD Pfam families, which all belong to animmunoglobulin-like-fold superfamily, the E-set Pfam clan(Bateman et al., 2004). It is noteworthy that such domainsare also found associated with catalytic domains belongingto the families GH5 and GH18 of the glycoside hydrolases(trEMBL code: Q48C6, Q693B8).
In the C-terminal region, a stretch of 75 residues dis-played significant matches, ranging from 24 to 40%sequence identity (46–62% sequence similarities), with theC-terminal domains of several proteins, mainly polysaccha-ridases and proteases, exclusively from bacteria of theBacteroidetes phylum, including the genera Porphyromo-nas, Cytophaga, Zobellia, Flavobacterium, Microscilla, andRhodothermus. In particular, the YPNP motif at the beginningof these domain sequences is highly conserved (Figure 3B).
Altogether, FcnA consists of a 1007-amino acid-longprotein (110.3 kDa) with a modular architecture. This pro-tein encompasses a 28-amino acid-long signal peptide, a390-amino acid-long N-terminal domain, three successiveHe-PIg domains with ∼105 residues each, followed by aregion of 196 amino acids without any significant sequencesimilarity with known proteins, and finally by a 75-aminoacid-long C-terminal domain.
Overexpression of the fucanase FcnA gene from SW5 strain
The fucanase FcnA was expressed under three differentforms. The first construct (FcnA1) encoded the wholeFcnA sequence without its predicted signal peptide in framewith the N-terminal His-tag of the pDEST17 vector. Thesecond construct (FcnA2) encompassed the N-terminaldomain and the three He-PIg domains of the protein. Thethird construct (FcnA3) only contained the N-terminal
Fig. 3. Sequence comparison of the noncatalytic domains of the fucanase FcnA with representative homologous domains. (A) The immunoglobulin-like repeats. This sequence alignment was created using the sequences of the immunoglobulin-like domains of FcnA (R1, G419-D527; R2, G528-N633; and R3, G634-N737) and of two proteins from Colwellia psycherythraea, a family GH-5 glycoside hydrolase (trEMBL code Q482C6: He-PIg, A358-E456) and a hypothetical conserved protein (trEMBL code Q482B8: He-PIg1, A766-N850, and He-PIg2, A870-S962). (B) The C-terminal domain. This sequence alignment was created using the sequences of the C-terminal domain of FcnA (L940-K1007) and of the following proteins: AgaA_Zg, β-agarase from Zobellia galactanivorans (trEMBL code Q9RGX9: L471-Q539), kappase_Zg, κ-carrageenase from Z. galactanivorans (trEMBL code O84907, I470-E545), antigen_PG102_Pg, immunoreactive 63-kDa antigen PG102 from Porphyromonas gingivalis (trEMBL code Q7MT80, L483-E554), phospholipase_D_Cp, phospholipase D from Chlorobium phaeobacteroides (trEMBL code Q4AHC4, V374-K451), beta1,4xylanase_Ch, xylanase from Cytophaga hutchinsonii (GenBank code ZP_00309327, L1312-R1375), endoglucanase_Ch, endoglucanase from C. hutchinsonii (GenBank code ZP_00308849, L1231-Q1302), and thiol_protease_Pg, thiol-protease from P. gingivalis (trEMBL code Q7MUR6, V774-H843). This figure was prepared using the program ESPript (Gouet et al., 2003).

Fucanase, a novel family of glycoside hydrolases
1025
domain. The expected products were proteins with 1001residues (109.7 kDa), 788 residues (86.5 kDa), and 412 resi-dues (46.6 kDa), respectively. SDS–PAGE analyses indi-cated that soluble products with the expected protein sizeswere obtained from the FcnA1 and FcnA2 constructs,FcnA2 being expressed in a greater amount than FcnA1.Unfortunately, the shortest construct FcnA3 yielded aninsoluble recombinant protein. Contaminant proteins co-eluted with FcnA1 and FcnA2 in the elution peak at ∼150 mMimidazole. Further purification attempts combining anionexchange and size-exclusion chromatography (SEC) didnot significantly improve the purity of FcnA1 and FcnA2(data not shown).
Action of fucanase FcnA on sulfated fucan fractions
The crude sulfated fucan fraction, FS28, was incubated inthe presence of either one or the other partially purifiedrecombinant enzymes, FcnA1 and FcnA2, and the enzy-matic degradation kinetics was monitored by C-PAGEanalysis and by spectrophotometry at 232 nm, the maxi-mum wavelength for the absorbance of 4,5-unsaturatedpyranoses. As illustrated for FcnA2 (Figure 4), both recom-binant enzymes yielded the same profiles in C-PAGEanalysis of the degradation products. The smallest oligosac-charide released by the fucanase approximately migratesat the level of a λ-neocarratetraose, which displayedthree sulfate groups by disaccharide repeating unit (Guibetet al., 2006). No increase of absorbance at 232 nm wasrecorded as function of time, indicating that the degrada-tion products are not unsaturated oligosaccharides.Therefore, FcnA is a glycoside hydrolase and not a polysac-charide lyase.
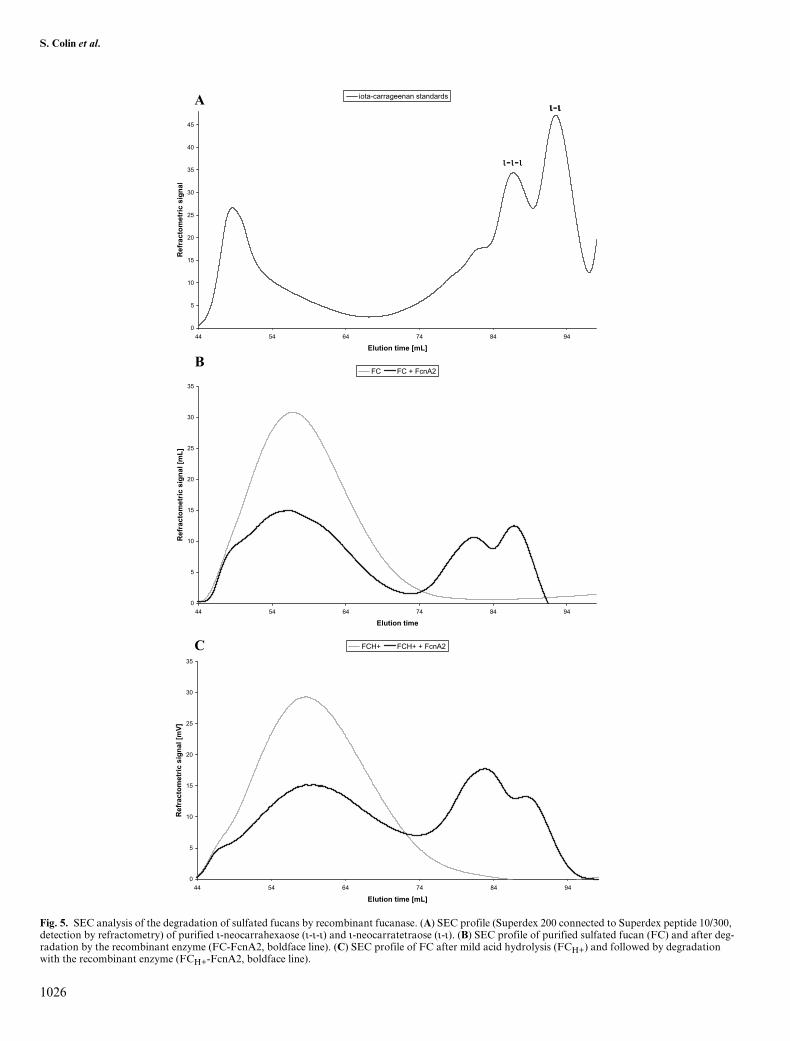
However, as monitored by SEC, FcnA2 did not exten-sively degrade the FS28-sulfated fucan fraction, even afterthe removal of polyphenol contaminants by a chromatog-raphy step on Sephadex LH 20. Low-molecular-weight(LMW) components were also detected in the FS28 frac-tion by SEC analysis. To further purify the native polysac-
charide, we ultrafiltrated the FS28 fraction onto a 50-kDacutoff membrane (Millipore, Molsheim, France). As shownby SEC analyses on a Superdex 200 column connected to aSuperdex peptide 10/300 GL column (GE Healthcare,Orsay, France), the retentate, referred to as FC, eluted atthe exclusion volume and did not contain LMW contami-nants (Figure 5B). 1H-NMR spectrum of the filtrate fractionshowed signals typical of sulfated fucan substrate, espe-cially in the regions of the α-anomeric protons and of themethyl groups. But in comparison with the 1H-NMR spec-trum of FC, these LMW components displayed additional,complex signals in the 3.1–4.9 ppm region, suggesting asubstitution pattern different from that of the polymericFC fraction (data not shown). In contrast to FS28, the sizeprofile of FC was significantly modified on incubation withthe recombinant fucanase FcnA2. The polymeric fractionsignificantly decreased, whereas two peaks eluted in the 74–94 mL region (Figure 5B). In the same elution conditions,purified ι-neocarratetraoses and ι-neocarrahexaoses, whichdisplayed two sulfate groups by disaccharide repeating unit,eluted in the 84–94 mL region (Figure 5A, Guibet M., per-sonal communication). The sulfated fucan fraction FCH+,resulting from the treatment of FC by mild acid hydrolysis,was amenable to a more complete degradation by therecombinant fucanase. In SEC analyses, the size profile ofFCH+ was similar to that of FC, indicating that the mild acidhydrolysis did not result in the breakdown of the polymer(Figure 5C). After incubation in the presence of FcnA2, thesize distribution was markedly shifted toward the lowermolecular weights, with notably an increase of the first peakof oligosaccharides (Figure 5C).
1H-NMR analyses of the enzymatic degradation of FCH+
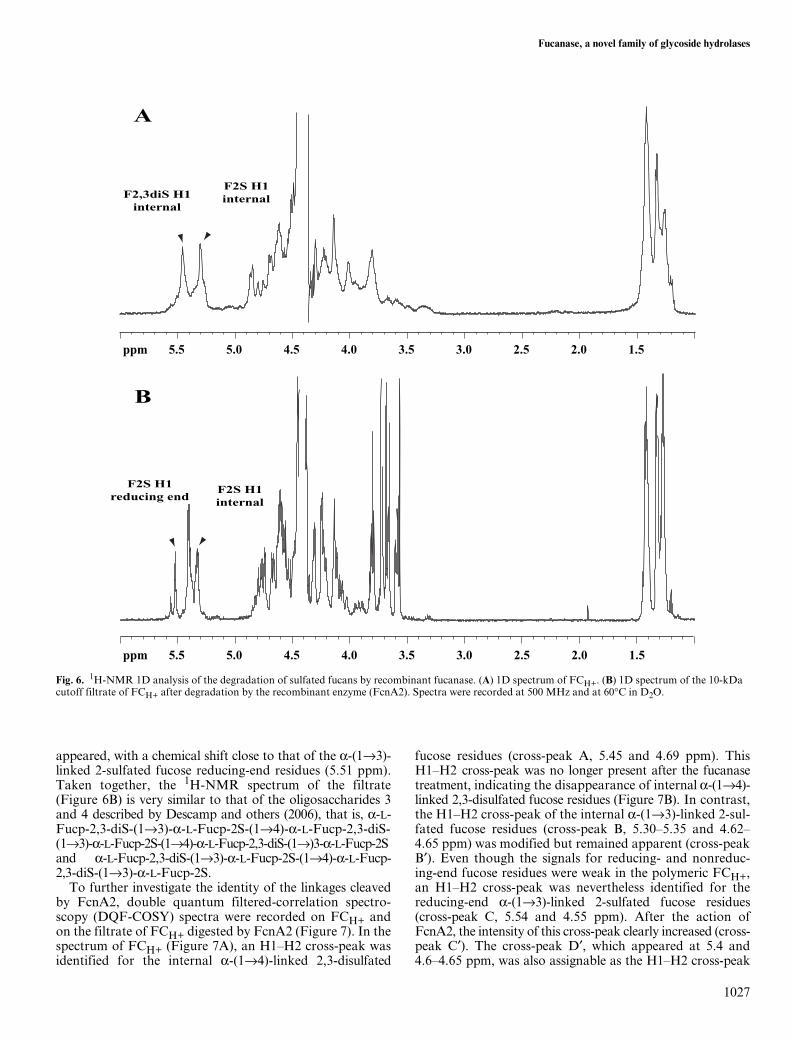
As shown in Figure 6A, the 1H-NMR spectrum of sulfatedfucan that had been subjected to mild acid hydrolysis,FCH+, was comparable with that of polymeric sulfatedfucan. In the region of the anomeric protons (5.6–5.2 ppm),signals were found with chemical shifts similar to thosedescribed by Chevolot and others (2001) as assignable toalternating α-(1→4)-linked 2,3-disulfated fucose andα-(1→3)-linked 2-sulfated fucose residues (F2,3S-F2S).This pattern, which was proposed to be the main structurein the sulfated fucans from Ascophyllum nodosum and Fucusvesiculosus fucoid algae (Chevolot et al., 2001), is alsopresent in the sulfated fucan of the fucalean alga P. canalic-ulata (Descamps et al., 2006). After incubating FCH+ withFcnA2, the reaction mixture was ultrafiltrated onto a 10-kDa cutoff membrane (Amicon) to separate the releasedoligosaccharides (filtrate) from the resistant fraction (reten-tate). In comparison with FCH+, the 1H-NMR spectrum ofthe filtrate was markedly modified (Figure 6B), particularlyin the regions of the α-anomeric protons (5.6–5.2 ppm) and ofthe methyl groups (1.4–1.0 ppm). In particular, the α-anomericsignals corresponding to the repeating motif F2,3S-F2Swere affected by the enzyme action. The signal assignableto the H1 of the internal α-(1→3)-linked 2-sulfated fucoseresidues (5.30 ppm) was refined, but its intensity did notdecrease on hydrolysis. In contrast, the signal of H1 ofthe internal α-(1→4)-linked 2,3-disulfated fucose residues(5.45 ppm) was significantly diminished. A doublet also
Fig. 4. C-PAGE analysis of the degradation products of the recombinant fucanase FcnA2. (A) Comparison between purified λ-carrageenan oligosaccharides (λ-λ, tetraose; λ-λ-λ, hexose) and the sulfated fucan oligosaccharides released by the recombinant fucanase FcnA2 after 42 h incubation with FC and FCH+, respectively. (B) Hydrolysis kinetics of crude, native sulfated fucan (FS28) by FcnA2, showing the endolytic action of the fucanase and the end products of the enzyme.
S. Colin et al.
1026
Fig. 5. SEC analysis of the degradation of sulfated fucans by recombinant fucanase. (A) SEC profile (Superdex 200 connected to Superdex peptide 10/300, detection by refractometry) of purified ι-neocarrahexaose (ι-ι-ι) and ι-neocarratetraose (ι-ι). (B) SEC profile of purified sulfated fucan (FC) and after deg-radation by the recombinant enzyme (FC-FcnA2, boldface line). (C) SEC profile of FC after mild acid hydrolysis (FCH+) and followed by degradation with the recombinant enzyme (FCH+-FcnA2, boldface line).
0
5
10
15
20
25
30
35
40
45
44 54 64 74 84 94
Elution time [mL]
langis cirtemotcarfe
R
iota-carrageenan standards
ι-ι-ι
ιι-ιι
0
5
10
15
20
25
30
35
44 54 64 74 84 94
Elution time
]Lm[ langis cirte
motcarfeR
FC FC + FcnA2
0
5
10
15
20
25
30
35
44 54 64 74 84 94
Elution time [mL]
]Vm[ langis cirte
motcar feR
FCH+ FCH+ + FcnA2
A
B
C
Fucanase, a novel family of glycoside hydrolases
1027
appeared, with a chemical shift close to that of the α-(1→3)-linked 2-sulfated fucose reducing-end residues (5.51 ppm).Taken together, the 1H-NMR spectrum of the filtrate(Figure 6B) is very similar to that of the oligosaccharides 3and 4 described by Descamp and others (2006), that is, α-L-Fucp-2,3-diS-(1→3)-α-L-Fucp-2S-(1→4)-α-L-Fucp-2,3-diS-(1→3)-α-L-Fucp-2S-(1→4)-α-L-Fucp-2,3-diS-(1→)3-α-L-Fucp-2Sand α-L-Fucp-2,3-diS-(1→3)-α-L-Fucp-2S-(1→4)-α-L-Fucp-2,3-diS-(1→3)-α-L-Fucp-2S.
To further investigate the identity of the linkages cleavedby FcnA2, double quantum filtered-correlation spectro-scopy (DQF-COSY) spectra were recorded on FCH+ andon the filtrate of FCH+ digested by FcnA2 (Figure 7). In thespectrum of FCH+ (Figure 7A), an H1–H2 cross-peak wasidentified for the internal α-(1→4)-linked 2,3-disulfated
fucose residues (cross-peak A, 5.45 and 4.69 ppm). ThisH1–H2 cross-peak was no longer present after the fucanasetreatment, indicating the disappearance of internal α-(1→4)-linked 2,3-disulfated fucose residues (Figure 7B). In contrast,the H1–H2 cross-peak of the internal α-(1→3)-linked 2-sul-fated fucose residues (cross-peak B, 5.30–5.35 and 4.62–4.65 ppm) was modified but remained apparent (cross-peakB′). Even though the signals for reducing- and nonreduc-ing-end fucose residues were weak in the polymeric FCH+,an H1–H2 cross-peak was nevertheless identified for thereducing-end α-(1→3)-linked 2-sulfated fucose residues(cross-peak C, 5.54 and 4.55 ppm). After the action ofFcnA2, the intensity of this cross-peak clearly increased (cross-peak C′). The cross-peak D′, which appeared at 5.4 and4.6–4.65 ppm, was also assignable as the H1–H2 cross-peak
Fig. 6. 1H-NMR 1D analysis of the degradation of sulfated fucans by recombinant fucanase. (A) 1D spectrum of FCH+. (B) 1D spectrum of the 10-kDa cutoff filtrate of FCH+ after degradation by the recombinant enzyme (FcnA2). Spectra were recorded at 500 MHz and at 60°C in D2O.
5.5 5.0 4.5 4.0 3.5 3.0 2.5 2.0 1.5ppm
5.5 5.0 4.5 4.0 3.5 3.0 2.5 2.0 1.5ppm
F2S H1internalF2,3diS H1
internal
F2S H1internal
F2S H1reducing end
A
B

S. Colin et al.
1028
of nonreducing-end α-(1→4)-linked 2,3-disulfated fucose resi-dues (Descamps et al., 2006). Altogether, the above resultsindicate that the filtrate fraction included oligosaccharidesbelonging to the series of the sulfated fucan oligosaccharidesdescribed by Descamp and others (2006). Therefore, FcnAcleaves the α-(1→4) linkages within the repeating motifs [→4)-α-L-fucopyranose-2,3-disulfate-(1→3)- α-L-fucopyranose-2-sulfate-(1→]n of the sulfated fucan polysaccharidic backbone.
Discussion
FcnA is a sulfated fucan a-1,4-endohydrolase
We here report the purification of the fucanolytic activitysecreted by a marine Flavobacteriaceae referred to as SW5(Descamps et al., 2006). Using a four-step procedure, theprotein content was decreased 12,500-fold from the culturemedium down to the enzyme electrophoretic homogeneity.Because the bioassay used to monitor protein purification,observation by C-PAGE of the release of oligosaccharidesfrom a complex fucan substrate, was not amenable to quan-tification, the ratio of enzyme recovery was not assessed.Based on internal peptides sequenced from the purified pro-tein, the enzyme was cloned and successfully produced inEscherichia coli, as both full-length and truncated proteinsand tagged with an N-terminal polyhistidine tail. Severallines of evidence indicate that the full-length recombinantprotein is indeed a fucanase: (1) its sequence featured thethree internal peptide sequences found in the wild-type pro-tein (I620-P636, T677-S692, and T743-M756); (2) based onC-PAGE analysis, it released oligosaccharides from the sul-fated fucan fraction FS28 (Figure 4); (3) as seen by SEC(Figure 5) and NMR (Figures 6 and 7) analyses, the recom-binant protein significantly depolymerized the purified sul-fated fucan fractions FC and FCH+; and (4) the N-terminaldomain of FcnA exhibited low yet significant sequence simi-larities with two patented fucanases (Takayama et al., 2002).
Based on the marked size decrease of the initial polysac-charide substrate (Figure 5) leading to the release of interme-diary-sized fucan oligosaccharides (Figure 4), the fucanaseproceeds according to an endo mode of action. However,FcnA did not extensively degrade the native sulfated fucanFS28, whereas it significantly degraded the purified fractionsFC and FCH+. The difference of degradation susceptibilitybetween FS28 and FC suggests that the LMW sulfated fucancomponents contained in FS28 may inhibit the fucanase.Moreover, the sulfated fucans of fucalean algae consist of apolysaccharidic backbone of alternating α-(1→4)-linkedfucose and α-(1→3)-linked fucose residues, masked by vari-ous substitutions, mainly ester-sulfate substituents and lat-eral branches (Pereira et al., 1999; Chevolot et al., 2001;Bilan et al., 2004). The increased efficiency of FcnA onFCH+ indicates that the mild acid hydrolysis has likelyremoved some specific substituents, unmasking the sulfatedfucan core structure and increasing the accessibility of fuca-nase to its substrate. For instance, mild acid hydrolysis hasbeen reported to remove 2-sulfate esters at specific sites in thesulfated fucans from sea urchins (Pomin et al., 2005).
The 1H-NMR analyses of the oligosaccharides released bythe enzymatic hydrolysis of FCH+ (Figures 6 and 7) indicatethat they belong to the homologous series of the sulfated fucanoligosaccharides described by Descamp and others (2006).Therefore, FcnA cleaves α(1→4) glycosidic linkages within theblocks of the repeating disaccharidic motif [→4)-α-L-fucopyra-nosyl-2,3-disulfate-(1→3)-α-L-fucopyranosyl-2-sulfate-(1→]n.
Sulfated fucan endohydrolases constitute a new family of glycoside hydrolases
FcnA is a modular enzyme featuring five domains, theN-terminal region, three consecutive He-PIg domains, and a
Fig. 7. 1H-NMR 2D analysis of the degradation of sulfated fucans by recombinant fucanase. (A) COSY DQF spectrum of FCH+. (B) COSY DQF spectrum of the 10-kDa cutoff filtrate of FCH+ after degradation by FcnA2. Spectra were recorded at 500 MHz and at 60°C in D2O.
ppm
4.55.05.5 ppm
4.5
5.0
5.5
ppm
5.5 5.0 4.5 ppm
5.5
5.0
4.5
A B
B
C
C
A
B
D

Fucanase, a novel family of glycoside hydrolases
1029
C-terminal conserved domain. The C-terminal region ofFcnA is conserved at the C-terminus of several proteinsexpressed by bacteria from the Bacteroidetes phylum, mainlyincluding degrading enzymes such as polysaccharidases orproteases (Figure 3B). Nothing is known yet on the functionof this specific peptidic sequence in these bacteria. Given theretention of fucan-degrading activity by the truncated formof the enzyme, FcnA2, this C-terminal domain does not par-ticipate in the catalytic machinery of FcnA. The three centralrepeats are predicted to adopt an immunoglobulin fold, buttheir exact functions remain elusive. Considering their smallsize and their association with other catalytic domains suchas glycoside hydrolases of the families GH5 and GH18(trEMBL code: Q48C6, Q693B8; Figure 3A), it is unlikelythat these repeated domains possess a catalytic function.
The N-terminal region of FcnA, which was alignable over alength of 384 residues with the N-terminal domains of the∼800 amino acid-long sulfated fucan hydrolases of Alteromo-nas sp. SN-1009, Fda1 and Fda2 (Figure 2), likely is the cata-lytic domain of the fucanase. This observation is furthersubstantiated by the fact that the truncated protein FcnA2,which comprises the N-terminal domain and the three He-PIgrepeats only, retained the enzyme activity. Our attempt to sep-arately express the N-terminal domain resulted in an insolublerecombinant protein, suggesting an incorrect delineation ofthe domain limits. Another possibility is that the N-terminaldomain interacts with the first He-PIg module and that itcannot be separated from this latter domain. Such an inter-action has been observed between the catalytic domain andthe carbohydrate binding module (CBM) module of theendo/exocellulase E4 (Sakon et al., 1997).
It follows that the N-terminal portions of Fda1 and Fda2,which share 24 and 23% sequence identity with that of FcnA,respectively, also are the catalytic domains of these fucanases.These three sequences do no match significantly with any pro-teins in the public databases, and no other fucanase sequencesare known so far. We thus propose that they constitute anovel family of glycoside hydrolases (CAZy server: http://afmb.cnrs-mrs.fr/CAZY/) (Henrissat and Bairoch, 1996).
In conclusion, the fucanase from SW5, referred to asFcnA, should help in cracking the structure of the sulfatedfucans of brown algae, a task which so far has proven dif-ficult because of the lack of appropriate, specific fucanhydrolases. This fucan α-(1→4) endohydrolase depolymer-izes the sulfated fucan backbone, made of stretches ofalternating α-(1→4)-linked 2,3-disulfated fucose and α-(1→3)-linked 2-sulfated fucose residues. FcnA thus providesanother case study for understanding the structure–functionrelationships in the degradation of polysaccharides with highdensities of sulfate-ester substituents. We indeed show herethat, together with two distant homologous bacterial fuca-nases, FcnA delineates a novel structural family, thus open-ing a new opportunity to further decipher the structuralbases of the degradation of sulfated polysaccharides.
Materials and methods
Preparation and analysis of sulfated fucan fractions
The preparation of the crude sulfated fucan fraction fromthe fucoid brown alga P. canaliculata, referred to as FS28,
was described previously (Descamps et al., 2006). Briefly,thalli were macerated overnight in 1 L of ethanol/formalde-hyde/water (80:5:15; v/v) and exhausted by 2 L of ethanol/formaldehyde/water followed by 2 L of acetone. The result-ing pellet was dried at 60°C and extracted twice for 3 h at70°C by a 0.01 N HCl solution supplemented with 4% (w/v)calcium chloride. The extract was filtered and neutralizedwith ammonium carbonate, and fucans were precipitatedwith 2.5 volumes of ethanol, re-suspended in water, andfreeze-dried. The resulting FS28 fraction containing 36%fucose (Descamps et al., 2006) was hygroscopic and dis-played a brownish color, indicating the presence of polyphe-nols. To eliminate polyphenols, we further fractionated FS28as follows. Aliquots (∼500 mg) were re-suspended in water(1%; w/v) and applied onto a Sephadex LH 20 column(GE Healthcare; total volume, 100 mL) equilibrated withwater. The water-eluted fractions (total volume, 250 mL) werepooled, extensively dialyzed (cutoff 3500 Da) against distilledwater, and freeze-dried. This white-colored sulfated fucanfraction contained 41% fucose. To eliminate LMW compo-nents, we ultrafiltrated this latter fraction against a 50-kDacutoff membrane (Amicon). The retentate is a polymeric sul-fated fucan fraction referred to as FC. To eliminate possiblebranching osidic residues (Chevolot et al., 1999) or some spe-cific sulfate-ester substituents (Pomin et al., 2005), we dis-solved FC in 0.5 M sulphuric acid (0.5%; w/v) and incubatedat 40°C for 30 min. After neutralization with 4 M sodiumhydroxide, the sample was dialyzed (cutoff 500 and 3500 Da)against water and freeze-dried. This sulfated fucan fraction isreferred as FCH+.
Assays for fucanase activity
Attempts to quantitatively assay for fucanolytic activity usingthe release of sulfated fucan oligosaccharides by a conven-tional reducing sugar assay were unsuccessful (Nelson, 1944;Kidby and Davidson, 1973; Descamps et al., 2006). Sulfatedfucan-degrading activity was thus monitored by a C-PAGEassay of the release of anionic oligosaccharides (Zablackis andPerez, 1990). Briefly, 100–250 μL of 0.1% (w/v) FS28 sulfatedfucan in 20 mM Tris–HCl buffer (pH 7.5) was incubated with1–5 μL of enzyme fraction for 1 h at room temperature. Theproducts (5 μL) were electrophoresed through a 6% (m/v)stacking and a 27% running, 1-mm-thick polyacrylamide gelin 50 mM Tris–HCl, 2 mM EDTA buffer (pH 8.7), andstained with Alcian Blue followed by silver nitrate (Min andCowman, 1986). Fucanolytic activity was detected by theoccurrence of anionic oligosaccharide bands in the bottompart of the running gel. Product formation was also moni-tored as an increase of absorbance at 232 nm as a function oftime. At room temperature, 5 μL of enzyme was added to a 1-mL cuvette containing 0.1% (w/v) FS28-sulfated fucan, and20 mM Tris–HCl buffer (pH 7.5). The absorbance at 232 nmof the reaction mixture was followed for 30 min against ablank cuvette containing 0.1% (w/v) FS28-sulfated fucan, 20mM Tris–HCl buffer (pH 7.5), and 5 μL of enzyme inacti-vated by heating at 100°C for 10 min.
Production and purification of fucanase FcnA
A 2-day-old culture in ZoBell-Fucan (ZF) medium of SW5strain was inoculated (10 mL) in a 1-L Erlenmeyer flask

S. Colin et al.
1030
containing 250 mL of ZF medium (Descamps et al., 2006)and cultured at 22°C, with a shaking speed of 200 rpm, for1 day. Aliquots (40 mL) were then grown for 5 days in 5-LErlenmeyer flasks in 6 × 1 L of the same medium under thesame conditions, and bacterial growth was monitored bythe absorbance at 600 nm. The culture supernatant was col-lected by centrifugation at 12,000 g for 20 min, filtered usinga Pellicon system with 0.45-μm membrane (Millipore), andconcentrated by ultrafiltration on a 30-kDa cutoff mem-brane (Millipore). The retentate (∼620 mL) was brought to40% saturation with (NH4)2SO4, the suspension was centri-fuged at 12,000 g for 15 min, and the resulting supernatantwas brought to 60% (w/v) saturation with (NH4)2SO4. Theprecipitate was then collected by centrifugation at 12,000 gfor 15 min and dissolved in 60 mL of 20 mM Tris–HCl buffer(pH 7.5), 5 mM MgCl2, 5 mM CaCl2, and 50 mM NaCl.This fraction was diluted 20-fold in a 20 mM Tris–HClbuffer (pH 7.5), resaturated at 40% with (NH4)2SO4, anddeposited on a Phenyl Sepharose CL4B column (GE Health-care) equilibrated with 100 mL of 20 mM Tris–HCl buffer(pH 7.5) and 40% (NH4)2SO4. The enzyme was eluted withthe same buffer followed by a linear gradient of (NH4)2SO4from 1.8 to 0 M (180 mL) in the same buffer. The activefractions (determined by their ability to release oligosaccha-rides from the FS28-sulfated fucan fraction as assayed byC-PAGE) were pooled and diluted 5-fold in a 20-mM Tris–HCl buffer (pH 7.5) containing 5 mM NaCl. The solutionwas applied onto a DEAE Sepharose CL6B column (GEHealthcare) equilibrated with 100 mL of 20 mM Tris–HClbuffer (pH 7.5, 5 mM NaCl). The column was washed withthe same buffer (140 mL) and eluted with a linear gradient of 5mM–1 M NaCl (210 mL). The active fractions were pooled(∼27 mL) and concentrated down to ∼1 mL by ultrafiltra-tion on a 10-kDa cutoff membrane (Centricon, Millipore).The concentrate (1 mL) was applied onto a Superdex 200 (GEHealthcare) equilibrated with 50 mM sodium phosphatebuffer (pH 7.0), 150 mM NaCl (100 mL), and the enzyme waseluted with the same buffer (30 mL). All purification stepswere performed at 4°C. The apparent molecular weight of thepurified enzyme was estimated by SDS–PAGE analysis, usinga 12.5% polyacrylamide gel and a Phast-system (GE Health-care), with standard proteins ranging from 14.5 to 200 kDa(broad range, Biorad, Marnes-la-Coquette, France). Proteincontents were measured according to the Bradford method(Bradford, 1976) using a Biorad protein assay reagent withbovine serum albumin (Sigma, Lyon, France) as standard.Internal peptide microsequences were determined by Edmandegradation at the Pasteur Institute (Paris, France).
Isolation and analysis of fucanase clones
Total DNA from SW5 strain was prepared as previouslydescribed (Barbeyron et al., 1984), cut by the restrictionendonuclease Sau3AI, and fractionated on a sucrose gradi-ent. DNA fragments of ∼4–10 kb were cloned at the BamHIsite of plasmid vector pAT153 which was used to transformE. coli strain DH5α. The genomic library contained ∼6000clones. Using the microsequences obtained from internalpeptides of the purified fucanase, we designed degeneratedDNA primers and we obtained an internal DNA probe of203 nt by PCR. The probe was labeled (kit Megaprime DNA
labeling systems, GE Healthcare) and used to screen thelibrary according to Sambrook and Russel (2001); then theclones of interest were mapped by restriction endonucleases.The probe was used for a Southern blot experiment carriedout on genomic DNA, using various endonuclease combina-tions (HinDIII/BamHI, HinDIII, HinDIII/XcmI, EcoRI/NdeI, PstI/NdeI, EcoRI/PstI/NdeI, ClaI/NdeI, XcmI/NdeI,XcmI/PvuII, and EcoRI/SalI) (Sambrook and Russel, 2001).Sequencing was carried out by gene walking on both strandsusing a 3100 Genetic Analyser (Applied Biosystems, Courta-boeuf, France) with BigDye Terminator V3.0 chemistry(DNA sequencing kit, Applied Biosystems) and synthetic oli-gonucleotides as primers. Sequences were verified at leastfive times and most of them >10 times.
Sequence analysis
Nucleotide sequences were searched for ORFs, as well forputative ribosomal binding sequences, putative promoters,and putative transcription terminators hairpins via themfold v.3.1 prediction server (Zuker et al., 1999). The signalpeptide was predicted with SignalP V2.0 (Nielsen et al.,1999). The searches of homology were run with GEN-BANKnr and PAT via the NCBI web server usingBLASTp and PSI-BLAST (Altschul et al., 1997). Evidencefor specific motifs and domains was queried on the Pfamdatabase (Bateman et al., 2004). The fucanase sequence waschecked for repeats using the program RADAR (Hegerand Holm, 2000).
Production of recombinant fucanase
The overexpression vector was constructed using GATE-WAY cloning technology (Invitrogen, Cergy Pontoise,France), according to the supplier’s instructions. The cod-ing region of SW5 fucanase, either as a whole or truncatedafter the He-PIg repeats or the N-terminal domain, wasamplified by PCR from genomic DNA with high fidelityPLATINUM Pfx DNA polymerase (Invitrogen). PCRproducts were inserted into pDONR201 vector, thesequence was checked, and a valid clone was inserted intothe pDEST17 vector with a N-terminal 6xHistidine tag.Plasmid vectors were used to transform BL21(DE3) pLysSE. coli strain (Novagen, Fontenay-Sous-Bois, France), andoverexpression was carried out in the M9 medium supple-mented with 2% casaminoacids, 100 μg mL–1 ampicillin,and 35 μg mL–1 chloramphenicol. Recombinant bacteriawere grown in a Bioflow 3000 fermentor at 37°C (pH 7.2)and under-regulated oxygen concentration (70% of air).When cultures reached 0.8 OD600, expression was inducedat 20°C with 1 mM isopropyl-1-thio-β-D-galactopyranoside(IPTG). Partially purified recombinant fucanases were pro-duced as follows. After an overnight induction, bacterialcultures (500 mL) were centrifuged for 15 min at 10,000 g,and pellets were either frozen (–20°C) or resuspended in50 mL of 20 mM Tris buffer (pH 7.5) containing 250 mMNaCl, 80 mM imidazol, and one tablet of Complete pro-tease inhibitors (Roche Diagnostics, Meylan, France) andthen disrupted by a French Press (4°C). The lysate wasultracentrifuged at 40,000 g for 1 h, and the supernatantwas loaded on a Fast Flow Chelating Sepharose (GEHealthcare) column saturated with 100 mM NiSO4. The

Fucanase, a novel family of glycoside hydrolases
1031
affinity chromatography column (V = 10 mL) was washedwith lysis buffer (5 V) and eluted with an 80–300 mM imi-dazol gradient (12 V) and then by 1 M imidazol (2 V). Pro-tein contents were estimated by SDS–PAGE, and aliquotsof the recombinant enzyme were assayed for sulfated fucan-degrading activity by C-PAGE, as above.
SEC and 1H-NMR monitoring of the enzymatic degradation of sulfated fucan fractions
The enzymatic degradation of sulfated fucan was moni-tored by SEC as follows. Sulfated fucan aliquots (20–50 mg,0.5–1% in water) were extensively digested with native orrecombinant fucanase for 42 h at room temperature, thenboiled for 10 min, and freeze-dried. Aliquots (10 mg) of thedigestion products were resuspended (0.5–1%; w/v) in waterand chromatographied with 50 mM ammonium carbonateusing a Superdex 200 column whose outlet is connected to aSuperdex peptide 10/300 GL column (GE Healthcare),equipped with refractometric detection. Liquid 1H-NMRspectra were recorded at 60°C on a Bruker DRX 500 Avance(Bruker, Wissembourg, France) at 500 MHz. Samples(∼10 mg) were previously exchanged twice in D2O (100%)before solubilization in 100% D2O.
Acknowledgments
We are grateful to Drs Mirjam Czjzek, William Helbert,and Bernard Henrissat for helpful discussions. We thankalso Miss Marion Guibet for her gift of purified ι- andλ-carrageenan oligosaccharides. S.C. and V.D. were therecipients of PhD fellowships co-funded by the ANRT,whose help is gratefully acknowledged.
Conflict of interest statement
None declared.
Abbreviations
C-PAGE, carbohydrate-polyacrylamide gel electrophore-sis; HCA, hydrophobic cluster analysis; LMW, low molec-ular weight; ORF, open-reading frame; SDS, sodiumdodecyl sulfate; SEC, size-exclusion chromatography.
References
Allouch, J., Jam, M., Helbert, W., Barbeyron, T., Kloareg, B., Henrissat, B.,and Czjzek, M. (2003) The three-dimensional structures of two beta-agarases. J. Biol. Chem., 278, 47171–47180.
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z., Miller, W.,and Lipman, D.J. (1997) Gapped BLAST and PSI-BLAST: a newgeneration of protein database search programs. Nucleic Acids Res.,25, 3389–3402.
Antonopoulos, A., Favetta, P., Helbert, W., and Lafosse, M. (2005)On-line liquid chromatography electrospray ionization mass spec-trometry for the characterization of kappa- and iota-carrageenans.Application to the hybrid iota-/nu-carrageenans. Anal. Chem., 77,4125–4136.
Antonopoulos, A., Hardouin, J., Favetta, P., Helbert, W., Delmas, A.F.,and Lafosse, M. (2005) Matrix-assisted laser desorption/ionisation
mass spectrometry for the direct analysis of enzymatically digestedkappa- iota- and hybrid iota/nu-carrageenans. Rapid Commun. MassSpectrom., 19, 2217–2226.
Barbeyron, T., Kean, K., and Forterre, P. (1984) DNA adenine methyla-tion of GATC sequences appeared recently in the Escherichia colilineage. J. Bacteriol., 160, 586–590.
Barbeyron, T., Michel, G., Potin, P., Henrissat, B., and Kloareg, B. (2000)iota-Carrageenases constitute a novel family of glycoside hydrolases, unre-lated to that of kappa-carrageenases. J. Biol. Chem., 275, 35499–35505.
Bateman, A., Coin, L., Durbin, R., Finn, R.D., Hollich, V., Griffiths-Jones, S., Khanna, A., Marshall, M., Moxon, S., Sonnhammer, E.L.,and others (2004) The Pfam protein families database. Nucleic AcidsRes., 32, 138–141.
Berteau, O. and Mulloy, B. (2003) Sulfated fucans, fresh perspectives:structures, functions, and biological properties of sulfated fucans andan overview of enzymes active toward this class of polysaccharide. Gly-cobiology, 13, 29–40.
Bilan, M.I., Grachev, A.A., Ustuzhanina, N.E., Shashkov, A.S., Nifantiev,N.E., and Usov, A.I. (2004) A highly regular fraction of a fucoidan fromthe brown seaweed Fucus distichus L. Carbohydr. Res., 339, 511–517.
Bradford, M.M. (1976) A rapid and sensitive method for the quantitationof microgram quantities of protein utilizing the principle of protein–dyebinding. Anal. Biochem., 72, 248–254.
Chevolot, L., Foucault, A., Chaubet, F., Kervarec, N., Sinquin, C.,Fisher, A.M., and Boisson-Vidal, C. (1999) Further data on the struc-ture of brown seaweed fucans: relationships with anticoagulant activ-ity. Carbohydr. Res., 319, 154–165.
Chevolot, L., Mulloy, B., Ratiskol, J., Foucault, A., and Colliec-Jouault, S.(2001) A disaccharide repeat unit is the major structure in fucoidansfrom two species of brown algae. Carbohydr. Res., 330, 529–535.
Descamps, V., Colin, S., Lahaye, M., Jam, M., Richard, C., Potin, P.,Barbeyron, T., Yvin, J.C., and Kloareg, B. (2006) Isolation and cultureof a marine bacterium degrading the sulfated fucans from marinebrown algae. Mar. Biotechnol., 8, 27–39.
Gaboriaud, C., Bissery, V., Benchetrit, T., and Mornon, J.P. (1987)Hydrophobic cluster analysis: an efficient new way to compare andanalyse amino acid sequences. FEBS Lett., 224, 149–155.
Goodner, B. and Quatrano, R.S. (1993) Fucus embryogenesis: a model tostudy the establishment of polarity. Plant Cell, 5, 1471–1481.
Gouet, P., Robert, X., and Courcelle, E. (2003) ESPript/ENDscript:extracting and rendering sequence and 3D information from atomicstructures of proteins. Nucleic Acids Res., 31, 3320–3323.
Guibet, M., Kervarec, N., Genicot, S., Chevolot, Y., and Helbert, W.(2006) Complete assignment of (1)H and (13)C NMR spectra of Gigartinaskottsbergii lambda-carrageenan using carrabiose oligosaccharidesprepared by enzymatic hydrolysis. Carbohydr. Res., 341, 1859–1869.
Heger, A. and Holm, L. (2000) Rapid automatic detection and alignmentof repeats in protein sequences. Proteins, 41, 224–237.
Henrissat, B. and Bairoch, A. (1996) Updating the sequence-based classifi-cation of glycosyl hydrolases. Biochem. J., 316, 695–696.
Jam, M., Flament, D., Allouch, J., Potin, P., Thion, L., Kloareg, B.,Czjzek, M., Helbert, W., Michel, G., and Barbeyron, T. (2005) Theendo-beta-agarases AgaA and AgaB from the marine bacterium Zobelliagalactanivorans: two paralogue enzymes with different molecularorganizations and catalytic behaviours. Biochem. J., 385, 703–713.
Kidby, D.K. and Davidson, D.J. (1973) A convenient ferricyanide estima-tion of reducing sugars in the nanomole range. Anal. Biochem., 55,321–325.
Klarzynski, O., Descamps, V., Plesse, B., Yvin, J.C., Kloareg, B., andFritig, B. (2003) Sulfated fucan oligosaccharides elicit defenseresponses in tobacco and local and systemic resistance against tobaccomosaic virus. Mol. Plant Microbe Interact., 16, 115–122.
Kloareg, B. and Quatrano, R. (1988) Structure of the cell walls of marinealgae and ecophysiological functions of the matrix polysaccharides.Oceanogr. Mar. Biol. Ann. Rev., 26, 259–315.
Kozak, M. (1999) Initiation of translation in prokaryotes and eukaryotes.Gene, 234, 187–208.
Kropf, D.L., Kloareg, B., and Quatrano, R.S. (1988) Cell wall isrequired for fixation of the embryonic axis in Fucus zygotes. Science,239, 187–190.
Lemesle-Varloot, L., Henrissat, B., Gaboriaud, C., Bissery, V., Morgat, A.,and Mornon, J.P. (1990) Hydrophobic cluster analysis: procedures to

S. Colin et al.
1032
derive structural and functional information from 2-D-representation ofprotein sequences. Biochimie, 72, 555–574.
Mabeau, S., Kloareg, B., and Joseleau, J.-P. (1990) Fractionation andanalysis of fucans from brown algae. Phytochemistry, 29, 2441–2445.
Michel, G., Chantalat, L., Duee, E., Barbeyron, T., Henrissat, B.,Kloareg, B., and Dideberg, O. (2001) The kappa-carrageenase of P.carrageenovora features a tunnel-shaped active site: a novel insight inthe evolution of Clan-B glycoside hydrolases. Structure, 9, 513–525.
Michel, G., Chantalat, L., Fanchon, E., Henrissat, B., Kloareg, B., andDideberg, O. (2001) The iota-carrageenase of Alteromonas fortis. Abeta-helix fold-containing enzyme for the degradation of a highly poly-anionic polysaccharide. J. Biol. Chem., 276, 40202–40209.
Michel, G., Helbert, W., Kahn, R., Dideberg, O., and Kloareg, B. (2003)The structural bases of the processive degradation of iota-carrageenan,a main cell wall polysaccharide of red algae. J. Mol. Biol., 334,421–433.
Michel, G., Nyval-Collen, P., Barbeyron, T., Czjzek, M., and Helbert, W.(2006) Bioconversion of red seaweed galactans: a focus on bacterialagarases and carrageenases. Appl. Microbiol. Biotechnol., 71, 23–33.
Min, H. and Cowman, M.K. (1986) Combined alcian blue and silver stain-ing of glycosaminoglycans in polyacrylamide gels: application to elec-trophoretic analysis of molecular weight distribution. Anal. Biochem.,155, 275–285.
Nelson, N. (1944) A photometric adaptation of the Somogyi method forthe determination of glucose. J. Biol. Chem., 153, 375–380.
Nielsen, H., Brunak, S., and von Heijne, G. (1999) Machine learningapproaches for the prediction of signal peptides and other protein sort-ing signals. Protein Eng., 12, 3–9.
Pereira, M.S., Mulloy, B., and Mourao, P.A. (1999) Structure and anticoagu-lant activity of sulfated fucans. Comparison between the regular, repeti-tive, and linear fucans from echinoderms with the more heterogeneousand branched polymers from brown algae. J. Biol. Chem., 274, 7656–7667.
Pomin, V.H., Valente, A.P., Pereira, M.S., and Mourão, P.A. (2005) Mildacid hydrolysis of sulfated fucans: a selective 2-desulfation reactionand an alternative approach for preparing tailored sulfated oligosac-charides. Glycobiology, 15, 1376–1385.
Sakon, J., Irwin, D., Wilson, D.B., and Karplus, P.A. (1997) Structure andmechanism of endo/exocellulase E4 from Thermomonospora fusca.Nat. Struct. Biol., 4, 810–818.
Sambrook, J. and Russel, D. (2001) Molecular Cloning: A LaboratoryManual. Cold Spring Harbor Press, Cold Spring Harbor, NY.
Takayama, M., Koyama, N., Sakai, T., and Kato, I. (2002) Enzymescapable of degrading a sulfated-fucose-containing polysaccharide andtheir encoding genes. Patent US6489155 B1.
Zablackis, E. and Perez, J. (1990) A partially pyruvated carrageenan fromHawaiian Grateloupia filicina (Cryptonemiales, Rhodophyta). Bot.Mar., 33, 273–276.
Zuker, M., Mathews, D.H., and Turner, D.H. (1999) Algorithms and Ther-modynamics for RNA Secondary Structure Prediction: A PracticalGuide. Kluwer Academic Publishers, Dordrecht.





























