Embed Size (px)
Citation preview

Cloning and characterisation of xylanase genes from phytopathogenic fungi with a special reference to
Helminthosporium turcicum, the cause of northern leaf blight of maize
Yeshitila Degefu
Department of Applied Biology
Plant Pathology
University of Helsinki
Finland
Academic Dissertation
To be presented, with the permission of The Faculty of Agriculture and Forestry,
University of Helsinki, for public criticism in Viikki, Auditorium B2 on September 19,
2003, at 12 o’clock noon
Helsinki 2003

Supervisors Professor Jari Valkonen Department of Applied Biology University Of Helsinki Finland and Dr. Ari Pappinen Department of Applied Biology University of Helsinki Finland Reviewers Professor Annele Hatakka Department of Applied Chemistry and Microbiology University of Helsinki Finland and Dr. Christina Dixelius Department of Plant Biology Swedish University of Agricultural Sciences Uppsala, Sweden Opponent Dr. Anne E. Osbourn Sainsbury Laboratory, John Innes Centre, Norwich United Kingdom
ISSN 1457-8085 ISBN 952-10-1325-7 (paperback) ISBN 952-10-1326-5 (PDF) Electronic version at http://ethesis.helsinki.fi Yliopistopaino, Helsinki 2003

To Nigist, Bethel and Tsion

Authorship statement Publications I, III, IV and V Yeshitila Degefu has been solely responsible for the planning and designing of the experimental work. Alone he has prepared the first draft of the manuscripts and he has been responsible for writing the final versions in collaboration with the other authors. He made all the correspondences concerning the publications. Publication II Yeshitila Degefu has been responsible for the planning of the study and designing of the experiments together with Peter Stephensen Lübeck. He carried out the xylanase production study under different culture conditions and did the xylanase activity assay He set up the induced cultures, collected the induced mycelial samples, isolated total RNA, made the cDNA synthesis and cloned the cDNA fragment for sequencing. He also contributed significantly to the writing of the paper.

TABLE OF CONTENTS
LIST OF ORIGINAL PUBLICATIONS ............................................................................... 7
ABBREVIATIONS USED.......................................................................................................... 8
ABSTRACT...................................................................................................................................... 9
1. GENERAL INTRODUCTION...........................................................................................10
1.1 The plant cell wall as an inert physical barrier...............................................................................12
1.2. The cell wall as a reservoir of signal molecules ............................................................................12
1.3. Components of the plant cell wall.................................................................................................16 1.3.1. Cell wall polysaccharides........................................................................................................17
1.3.1.1. Cellulose ..............................................................................................................................17 1.3.1.2. Hemicelluloses .......................................................................................................................17 1.3.1.3. Pectic polysaccharides ...............................................................................................................17
1.3.2. Cell wall structural proteins....................................................................................................18
1.4. Monocots and dicots differ in their primary wall composition .....................................................19
1.5. Cell wall degradation ....................................................................................................................21
1.6. Cell wall composition, enzymic adaptation and plant-pathogen compatibility........................... 22
1.7. Xylan and xylanases..................................................................................................................... 23 1.7.1. Structure of xylan................................................................................................................... 23 1.7.2. Biodegradation of xylan ........................................................................................................ 25 1.7.3. Occurrence and production of xylanases .............................................................................. 25 1.7.4. Xylanase multiplicity: a mere genetic redundancy or a display of functional diversity? ....... 27 1.7.5. Regulation of xylanase gene expression................................................................................ 28
1.7.5.1. Induction............................................................................................................................. 29 1.7.5.2. Carbon catabolic repression ...................................................................................................... 29
1.7.6. Xylanase inhibitor proteins (XIPs).........................................................................................31 1.7.7. Xylanase in plant pathogenesis ............................................................................................. 33
1.7.7.1. Xylanases as pathogenicity or virulence factors............................................................................... 33 1.7.7.2. Xylanases as elicitors of defence responses in plants ......................................................................... 35
1.8. Molecular evolution of xylanases ................................................................................................. 36
1.9. Helminthosporium turcicum: the cause of northern leaf blight of maize ................................... 37 1.9.1. Taxonomy and etiology ......................................................................................................... 37 1.9.2. Distribution and economic importance ................................................................................ 37 1.9.3. Infection and disease development ....................................................................................... 38 1.9.4. Disease cycle and epidemiology ........................................................................................... 38 1.9.5 Pathogenic races, sources of resistance and control of H. turcicum..................................... 38
1.10. Technologies (new and in use) in plant-pathogen interactions studies. ............ 40 1.10.1. Fungal genomics.................................................................................................................. 40

1.10.2. Fungal transformation and gene disruption ........................................................................ 42
2. AIMS OF THE STUDY ........................................................................................................ 46
3. MATERIALS AND METHODS........................................................................................ 47 3.1. Fungal strains and culture conditions ......................................................................................... 47 3.2. Bacterial strains and plasmids ..................................................................................................... 47 3.3. Production of xylanase ................................................................................................................ 47 3.4. Xylanase activity assay (I, II) ...................................................................................................... 48 3.5. Purification of xylanase (I) .......................................................................................................... 48 3.6. Tryptic cleavage and peptide separation (I)................................................................................ 49 3.7. Protein and peptide sequencing (I) ............................................................................................. 50 3.8. Construction of genomic library (II, III)..................................................................................... 50 3.9. DNA extractions (II, III, IV, V) .................................................................................................. 50 3.10. RNA extraction and first strand cDNA synthesis (II, III, IV) ................................................... 50 3.11. Southern hybridisations (II, III, IV, V) .......................................................................................51 3.12. Northern analysis (II, III, IV) .....................................................................................................51 3.13. Isolation and characterisation of xylanase genes (II, III, IV) .................................................... 52 3.14. Sequence and phylogenetic analyses (II, IV)............................................................................. 52 3.15. Plant infection and reverse transcription–polymerase chain reaction (RT-PCR) (III, IV)........ 53 3.16. Agrobacterium tumefaciens-mediated transformation of H. turcicum (V) ............................... 53
3.16.1. PCR screening of putative transformants (V) ...................................................................... 54 3.16.2. Southern analysis of tranformants (V) ................................................................................. 54
4. RESULTS AND DISCUSSION.............................................................................56 4.1. H. turcicum and A. pisi differ in their ability to produce xylanase activity in culture ................. 56 4.2. Purification properties of xylanase from the H. turcicum (I) ...................................................... 57 4.3. Cloning of the xylanase genes ..................................................................................................... 59 4.4. htxyl1 and htxyl2 exhibit opposite patterns of expression under some test conditions............... 60 4.5. The phylogenetic positions of htxyl1 and htxyl2 ......................................................................... 63 4.6. Agrobacterium tumefaciens-mediated transformation of H. turcicum....................................... 64
5. CONCLUSIONS AND PROSPECTS .............................................................................. 66
6. ACKNOWLEDGEMENTS.................................................................................................. 68
7. REFERENCES........................................................................................................................ 70

- 7 -
LIST OF ORIGINAL PUBLICATIONS
The thesis is based on the following publications, which will be referred in the text in
Roman numerals (I-V). The published papers are reprinted with the permission from the
publishers.
I Degefu, Y, Fagerström, R., and Kalkkinen, N. (1995) Purification and partial
characterisation of xylanase from the fungal maize pathogen Helminthosporium
turcicum Pass. European Journal of Plant Pathology 101: 291-299.
II Lübeck, P.S., Paulin, L., Degefu, Y., Lübeck, M., Alekhina, I., Bullat, S.A., and
Collinge, D.B. (1997) PCR cloning, DNA sequencing and phylogenetic analysis
of a xylanase gene from the phytopathogenic fungus Ascochyta pisi Lib.
Physiological and Molecular Plant Pathology 51: 377-389.
III Degefu, Y., Paulin, L., and Lübeck, P. S (2001) Cloning, sequencing and
expression of a xylanase gene from the maize pathogen Helminthosporium turcicum.
European Journal of Plant Pathology 107: 457-465.
IV Degefu, Y., Lohtander, K.L., and Paulin, L. (2003) Expression patterns and
phylogenetic analysis of two xylanase genes (htxyl1 and htxyl2) from
Helminthosporium turcicum, the cause of northern leaf blight of maize (under
review, Biochimie).
V Degefu, Y., and Hanif, M. (2003) Agrobacterium tumefaciens-mediated
transformation of Helminthosporium turcicum, the maize leaf blight fungus.
Archives of Microbiology (in Press). Published online on 31.7.2003 (DOI
10.1007/s00203-003-0589-5).

- 8 -
ABBREVIATIONS USED AGPs arabinogalactan proteins ATMT Agrobacterium tumefaciens-mediated transformation CWDEs cell wall degrading enzymes cDNA complementary DNA DNA deoxyribonucleic acid DNase deoxyribonuclease DNS dinitrosalicylic acid DP degree of polymerisation EIX ethylene inducing xylanase GAXs glucurunoarabinoxylans GIPs glucanase inhibitor proteins GPD glyceraldehyde phosphate dehydrogenase GRPs glycin rich proteins HHRGPs histidine rich glycoproteins HPLC high pressure liquid chromatography HRGPs hydroxyproline rich glycopoteins ITS internal transcribed spacers KDa kilodaltons mRNA messenger RNA NPR nitrogen pathogenicity regulation PDA potato dextrose agar PDB potato dextrose broth PGA polygalacturonic acid PGIPs polygalacturonase inhibitor proteins PGL polygalacturonic acid lyase PNLIPs pectin lyase inhibitor proteins PCR polymerase chain reaction RNA ribonucleic acid RNase ribonuclease ROS reactive oxygen species RT-PCR reverse transcription PCR SDS-PAGE sodium dodecyl sulphate polyacrylamide gel electrophoresis TAXI Triticum aestivum xylanase inhibitor THRGPs threonine rich glycoproteins XGs xyloglucans XGIPs xyloglucans inhibitor proteins XIPs xylanase inhibitor proteins

- 9 -
ABSTRACT
For the plant cell, the cell wall is a frontier zone, where the cell encounters the challenge
of the invading pathogens. For the plant biologist the cell wall is a research frontier, at
which the old concept of an inert dead structure-less box has given way to a new
concept of a highly complex structure and metabolically active organelle. The cell wall
acts as a reservoir of signal molecules or a "pseudo-gland" which secretes a class of
regulatory molecules known as oligosaccharins. Any organism that attempts to colonise a
plant must contend the cell wall and therefore cell wall degradation is the most
conspicuous effect of microorganisms on plants. Arabinoxylan makes up to 60% of the
primary cell wall of maize. The study in this thesis indicated that Helminthosporium
turcicum, the pathogen of maize, produced high xylanase activity. A comparative study
showed that the pea pathogen, Ascochyta pisi, was less efficient producer of xylanase
activity than H. turcicum under similar test conditions. Xylanase from H. turcicum was
purified and characterised, and the genes encoding xylanases were cloned. Detailed
analysis of the xylanase genes particularly of H. turcicum is presented. According to
sequence alignment and phylogenetic analysis, the cloned xylanase genes encode
xylanases, which belong to family 11 glycosyl hydrolases. Investigation on the
accumulation of the transcript of xylanase genes by northern blotting indicated that the
xylanases are subject to substrate induction and glucose inhibition. Glucose inhibited
transcript accumulation of both htxyl1 and htxyl2 in the presence of the inducing
molecules (xylan or xylose). The two xylanase genes from H. turcicum displayed opposite
patterns of expression with respect to nitrogen sources in vitro. The transcripts of both
xylanase genes were present in infected maize plants both at early and late stages of
infection but differed in transcript abundance. The transcript of htxyl1 was detectable
only by RT-PCR but not in northern blot. Transcript of htxyl2 was readily detectable in
northern blot and accumulated abundantly both at early and late stages of infection
suggesting that htxyl2 encodes a major xylanase induced during infection of maize by H.
turcicum and might be essential for pathogenicity. As an important first step in the
advancement of molecular studies in H. turcicum, an Agrobacterium tumefaciens-mediated
transformation system was developed. Experimentally adequate rate of transformation
was obtained using conidia of H. turcicum. The method is a potential tool for insertional
inactivation and identifying pathogenicity genes in H. turcicum and molecular analysis the
H. turcicum-maize interactions.

- 10 -
1. GENERAL INTRODUCTION
"How pathogens attack plants? And how plants defend themselves? " These are the
central and most relevant questions of plant pathology. During the last one hundred
years of its history (Keen, 2000; Sequeira, 1993), plant disease research has focused on
understanding these interactions and developing more effective means of disease
control. In the beginning of the twentieth century plant pathology embraced a strategy
that encouraged basic research on pathogen and disease defence. These led to the
development of disease resistant crops, and in the process saved billions of dollars and
resulted in reduced use of pesticides. Since recently, plant pathology has rapidly picked
up the powerful technologies of modern molecular biology such as cloning and
sequencing genes and these have helped to make major advances in our basic
understanding of plant diseases. Several fungi have been made amenable to molecular
genetic manipulations and genome wide projects aimed at sequencing fungal genomes
are in progress in several laboratories. A case in point is the Magnaporthe grisea genome
project (http://www-genome.wi.mit.edu/annotation/fungi/magnaporthe/). All these
efforts are geared towards developing genomic resources to obtaining thorough enough
understanding of plant-pathogen interactions and eventually develop durable and
environmentally sound disease control strategy.
Plant diseases have been the potential threat to global food security and continue to be
the focus of extensive research with a wide range of methodologies. Improving methods
(Gold et al., 2001) allow the dissection of fungal pathogenicity determinants in a gene-
by-gene approach. Such significant advances and the knowledge and the development of
efficient technologies for producing transgenic plants create optimism that plant diseases
will be more efficiently controlled in the twenty-first century.
For more than a century, cell wall degrading enzymes (CWDEs) have been implicated to
be involved in pathogenicity but the proof of their actual role had to await the
development of techniques of enzyme purification and the molecular cloning and other
stringent molecular procedures such as targeted gene disruption. The longstanding
questions of cell wall degrading enzymes are now being addressed using a concerted
approach of new molecular and immunocytochemical tools (Gold et al., 2001).

- 11 -
Our views and understanding of the cell wall structure and function has grown
remarkably well due to progress made in carbohydrate chemistry (Carpita, 1996; 2000).
From characteristic repeating unit structures, the sequence and conformation of very
large polymers are deduced (Carpita and Gibeaut, 1993). The major polysaccharides of
the walls of many flowering plants have been defined. The dynamic interactions of
individual components in cell-to-cell communication and molecular dialogue on cell wall
level have been documented. Matching to these advancements in the cell wall
architecture and functions, significant progress has been made in studies involving cell
wall degrading enzymes from various perspectives. Cell wall degrading enzymes have
been purified from a wide range of fungi and bacteria, and genes encoding the enzymes
have been cloned and characterised.
The primary cell wall of maize comprises up to 60% arabinoxylan, and therefore, xylan
degradation may be important for infection by pathogens. In this thesis I present results
of detailed study of production, purification of xylanase and cloning and characterisation
of the genes encoding xylanases from maize pathogen Helminthosporium turcicum and the
pea (Pisum sativum) pathogen Ascochyta pisi. A. pisi was included in the study because it
represents pathogens of dicots; group of plants in which pectin is the predominant
component of the primary cell wall. The fungus causes leaf, stem and pod spot of pea
and it is known to penetrate the plant via the cuticle and the stomata. The fungus has
wide spread distribution in the world. Major emphasis was given to xylanases from H.
turcicum because of the abundance and heterogeneity of xylan in the host plant, maize.
This thesis also provides an Agrobacterium tumefaciens-mediated transformation of H.
turcicum for the first time. The fungus, until this study, was not amenable to any
molecular genetic manipulations.
A comprehensive review of the structure and function of the cell wall, as well as
microbial cell wall degradation with special emphasis on xylan and xylanases is
presented. The current status of the knowledge of fungal xylanases as pathogenicity
factors and elicitors of defence responses in plants are discussed.

- 12 -
1.1 The plant cell wall as an inert physical barrier The pant cell wall is at the forefront of plant pathogen interactions. As the first barrier
encountered by most plant pathogens, it must be degraded in order to allow penetration
and colonisation. The cell wall represents the bulk of plant biomass and it is chemically
and structurally heterogeneous. The various polymers it contains may serve as substrates
to the numerous enzymes secreted by microbial pathogens, providing them nutrients
(Walton, 1994). Owing to its strategic position, it is also involved in plant defence
through an increased deposition of structural polymers, particularly 1,3-β-glucans and
lignin, and through enrichment in several defence proteins. Moreover, studies in
pathogenicity and defence have shown clearly that the cell wall is not only an inert
mechanical support and barrier but it is also a dynamic and metabolically active
organelle, which acts as a media for a molecular dialogue between plants and pathogens.
Albersheim and Darvill (1985) coined the word " pseudo-gland" to describe the cell wall
as a storehouse of precursors of a class of regulatory molecules that on being released
are capable of controlling a number of plant functions.
1.2. The cell wall as a reservoir of signal molecules
Several recent discoveries have changed our view of the plant cell wall from that of inert
and static structure to one in which the wall represents a virtual extension of the
cytoplasm (Robinson, 1991). The cell wall contains components for signalling and
communications by symplastic continuity through plasmodesmata (Roberts and Lucas,
1990; Carpita, 1996). Signals from the cell wall elicited by microbial attack induce the
production of defence molecules against attack by pathogens and insects (Albersheim
and Darvill, 1985; Faure, 2002; Ryan, 1990). Most of our current understanding of cell
wall signalling originates from the studies of a group of biologically active
oligosaccharides commonly known as "oligosaccharins"(Albersheim and Darvill, 1985;
Darvill et al., 1992).
Many of the structural polysaccharides of plant and fungal cell wall undergo enzymic
hydrolysis in vivo, potentially releasing short chain of sugar residues (Degree of
polymerisation (DP) of 10-15 for biologically active oligosaccharides) interconnected by
glycosidic linkages (Fry, 1995). Three mechanisms are known by which oligosaccharins

- 13 -
are released from the cell wall. In some pathogen infections 1) an enzyme from the
infected plant cleaves the fungal cell wall releasing oligosaccharins, and in addition,
oligoglucan, oligochitin and oligochitosan are released by the action of glucanases and
chitinases produced by plants; 2) the fungal or bacterial pathogen supplies an enzyme
that cleaves the infected plant cell wall releasing oligosaccharins, such as
oligogalacturonide, (the well-studied oligosaccharin) which is released by the action of
polygalacturonases and pectate lyase secreted by pathogens, and 3) in some cases the
plant itself supplies an enzyme generating oligosaccharides from its own walls. In all
three cases the oligosaccharins presumably combine with a receptor in the plant to form
an activated signal molecule. The signal causes a number of plant genes to be transcribed
into mRNA, which are further translated into enzymes that regulate different pathways
that control plant growth, organogenesis or activate defence responses against pathogens
(Dravill et al., 1992; Cote and Hahn, 1994; John et al., 1997; Fry et al., 1993; Esquerre-
Tugaye et al., 2000).
Oligosaccharins derived from fungal and plant polysaccharides are one class of well-
characterised elicitors. They are powerful signalling molecules capable of acting at very
low concentration to convey information to the plant under attack (Ryan and Farmer,
1991; Shibuya and Minami, 2001). In response to this information (the elicitor) the plant
defence response is activated often through activation of genes that encode enzymes
responsible for the synthesis of phytoalexins (Peck et al., 2001). Other activated defence
responses include biosynthesis of jasmonates (Nojiri et al., 1996), generation of reactive
oxygen species (ROS, oxidative burst) (Mehdy, 1994), and expression of unique early
responsive genes and typical defence related genes (Ebel and Mithöfer, 1998). Pectic
oligosaccharides are also known to act as a signalling molecule in the early step of cold
acclimation (Balandran-Quintana, et al., 2002). A hypothetical model of oligosaccharide
elicitor signalling which illustrates the events that take place in plant-pathogen
interactions is presented in Figure 1.
Furthermore, the cell walls of plants respond to plant and pathogen signals by
accumulating defence molecules required for cell wall reinforcement. For example, the
deposition of lignin and structural proteins result in resistant cell wall complex harder to
penetrate by pathogens (Farmer and Helgeson, 1987; Schnabelrauch et al., 1996; Brisson
et al., 1994).

- 14 -
From these observations it is evident that the plant cell walls contribute via their
enzymes to the signal release from pathogens, in addition to being by themselves a
reservoir of signals. It is interesting to note at this point that the hydrolytic enzymes,
which are the vitally important elements of the process, play at least two important roles
in plant tissues: on one hand they disrupt the structure of plant walls allowing fungal
colonisation of plant tissues (virulent or pathogenicty factors) and on the other hand
they are potential avirulent factors (Cervone et al., 1989) since they may activate plant
defence responses by releasing plant cell wall fragments that signal the plant to defend
itself. This raises the question whether this functional dichotomy actually occurs in vivo
or whether plants have a mechanism to balance responses between these two
phenomena, which seem apparently paradoxical.
GIPs PGIPs XGIPs XIPs?
Cell wall degrading enzymes
Fungal pathogen
GIPs Endo-1,3-β-glucanases
Defense-related genes
Defense responses
Plant cell
Receptor ?
Signal cascade
Nucleus
Cell wall
Oligosaccharide elicitors
-Hypersensitive response -Phytoalexin synthesis -Pathogen-related proteins -Ion fluxes -Protein phosphorylation -Active oxygen production
Figure 1. A hypothetical model of oligosaccharide elicitor signalling in plant-pathogen interactions. Adapted and modified from Bergmann et al. http://www.ccrc.uga.edu

- 15 -
Some studies suggest that in the process of molecular cross talk between the pathogen
and the plant, plants seem to have developed a system to regulate the amount and
longevity of the biologically active oligosaccharides. The studies by Garcia-Romera and
Fry (1995, 1997) shade some light on the possible mechanism by which this process is
controlled. According to their findings, endogenous exo - and endo-polygalacturonase
do not operate in the apoplast of healthy plants; instead their co-ordinated function is
assumed to begin during infection. That is, the exo-polygalacturonase is believed to
gradually degrade elicitor-active oligogalacturonides produced by the action of endo-
polygalacturonase of an invading microorganism to smaller elicitor-inactive products.
These co-operate functions (degradation of elicitor-active oligosaccharides) is believed to
be biologically advantageous to the plant. The disappearance of the biologically active
oligogalacturonides would signal to the plant that the potential pathogen has been
defeated, as a result of which the continued synthesis of the defence products would be
an energy cost to the plant and affect its growth and development (Dumville and Fry,
2000).
Furthermore, plant tissues have also systems of sequestering important signalling
molecules such as hormones, so that the message does not persist in vivo after it has been
perceived (Garcia-Romera and Fry, 1995). Another line of evidence also suggests that
the oligosaccharins produced in healthy plant tissues are used to control growth and
development (Darvill et al., 1992).
From the studies reviewed in this thesis and the many other reports on cell wall
signalling, it is noteworthy that the molecular dialogue established at the cell wall level
may direct the outcome of plant-pathogen interactions. Therefore, further studies on
how this dialogue is established and its function in the plant system would greatly add to
our understanding of pathogen-plant interactions.
In view of the well-defined chemical nature and the presence of highly sensitive
perception system in plants, some of these elicitors have provided good model systems
to study how these chemical signals are recognised, transduced and translated into
activities needed for activation of the defence machinery or other plant developmental
processes. These studies include structure-activity relationships of the elicitor molecules,

- 16 -
characterisation of the corresponding receptors and analysis of the signal transduction
cascades and elicitor response genes (Darvill et al., 1992). The details of receptors, signal
transduction pathways and their functions are beyond the scope of this thesis and will
not be discussed further, but the reader is referred to the following very comprehensive
recent reviews (Dumville and Fry, 2000; Esquerre-Tugaye et al., 2000; Shibuya and
Minami, 2001).
1.3. Components of the plant cell wall
The knowledge of cell wall structure is essential to our understanding of its interaction
with microbes. Functionally, the cell wall may be divided into three regions: the middle
lamella, primary wall, and secondary wall. The middle lamella serves as intracellular
cement that binds cells together in tissue system. This layer consists of largely pectic
substances and can be identified as an extremely thin layer between two adjacent cells
(Brett and Waldron, 1996). Primary wall is the first formed and readily visible layer of the
cell wall (Dickison, 2000) and it continues to deposit as long as the cell is growing in
surface area. It is the most dynamic of the wall regions and it functions to support the
protoplast in young growing cells. It also protects the plant against pathogens,
dehydration and other environmental stresses, and serves as a source of signalling
molecules and medium for cell-to-cell interactions (McNiel et al., 1984). Many cells
often limit themselves to two of these regions but certain specialised cells however
proceed to deposit secondary wall after cell elongation is complete. The secondary wall is
mainly for support and is comprised primarily of cellulose and lignin (Brett and
Waldron, 1996).
The plant cell wall may be viewed as a two-phase-system: a diffused phase of cellulose
microfibrils and a continuous matrix (Brett and Waldron, 1996). During the
development, the cell walls are remodelled or undergo conspicuous changes in structure,
form and functions (Carpita and Gibeaut, 1993; Carpita, 1996; Carpita et al., 2000). The
cell walls in young elongating tissues are composed predominantly of polysaccharides
together with lesser amount of structural hydroxyproline-rich glycoproteins and aromatic
compounds such as ferulic acid and lignin, which are thought to play an important role
in the structure and functions of the plant cell wall (Showalter, 1993; Cassab, 1998).

- 17 -
1.3.1. Cell wall polysaccharides
The plant cell wall is composed of approximately 90% polysaccharides (McNeil, 1984)
and can be divided into three groups: cellulose, hemicellulose and pectin. These
polysaccharides are built from monosaccharides or from simple sugars such as glucose
and galatose.
1.3.1.1. Cellulose
Cellulose represents the major constituents of cell wall polysaccharides and comprises
extremely long linear polymer of 1,4-β- linked D-glucose (as many as 25000 glucose
molecules). In plants, cellulose is present in both primary (25-30%) and secondary walls
(with evidence of lower degree of polymerisation in primary walls than in secondary
walls) in highly ordered structure called microfibrils (analogous to the formation of a
thick rope from thin fibbers) (Brett and Waldron, 1996). Microfibrils are highly stable
and their crystalline structure plays a major role in the structural characteristics of cell
wall (Dickison, 2000).
1.3.1.2. Hemicelluloses Hemicelluloses include a heterogeneous group of cellulose-like polysaccharides that are
variable in composition. They also vary greatly in amount and complexity in different
cell types and in different plant species (Huisman et al., 2000). Hemicelluloses include
xylan, mannan, galactan and arabinan. The main heteropolymers and the principal
monomers present in most of the hemicelluloses are D-xylose, D-mannose, D-galactose,
and L-arabinose. Hemicelluloses are linear, flat with a β-1,4 backbone and relatively
short side chains. Xylan, the major hemicellulose polymer in cereals and hard wood,
consists of a β 1,4-linked D-xylose backbone substituted by different side groups such as
L-arabinose, D-galactose, acetyl, feruloyl, p-coumaroyl, and glucuronic acid residues
(Biely, 1985; Bastawade, 1992).
1.3.1.3. Pectic polysaccharides
Pectic polysaccharides are another diverse group of polysaccharides that are particularly
rich in galacturonic acid, rhamnose, arabinose and galactose. They are characteristics of

- 18 -
the middle lamella and primary walls of dicotyledonous plants and to a lesser extent of
monocotyledonous plants (Brett and Waldron, 1996). Polymers of primarily β-1,4
galacturonic acid, also known as homogalacturonans, are particularly common. The
pectic polysaccharides serve a variety of functions including determining wall porosity,
providing wall surface for cell-to-cell adhesion (middle lamella), cell-to-cell recognition
and pathogen recognition (Brett and Waldron, 1996; Dickson, 2000; Dumville and Fry,
2000 and references therein).
1.3.2. Cell wall structural proteins
The plant cell walls contain a variety of proteins, the majority of which are glycosylated
(Showalter, 1993; Cassab, 1998). Among the cell wall structural proteins known thus far,
the most studied and well-documented classes of wall proteins include extensins, the
glycine rich proteins (GRPs), the hydroxyproline rich glycoproteins (HRGPs),
solanaceous lectins and arabinogalactan proteins (AGPs) (Showalter, 1993). These five
classes of proteins are believed to be evolutionarily related to each other (Sommer-
Knudsen, 1998) since all, with the exception of the glycine rich proteins, contain
hydroxyproline. Hydroxyproline is not generally found in protoplast proteins and thus
this particular feature of cell wall structural proteins makes it easier to locate the cell wall
as being the location of such glycoproteins (Brett and Waldron, 1996; Cassab, 1998). To
date much is known about the structure and metabolic regulation of wall structural
proteins, but direct functional evidence and knowledge of their intermolecular
interactions is lacking. The majority of cell wall proteins are cross-linked into the wall
and they are believed to function in strengthening the wall and wound healing for
example in response to mechanical stress such as bending of the plant due to wind and
in response to insect and pathogen attack (Showalter, 1993).
Most of our knowledge about wall proteins is based on extensins, which are the most
studied of all cell wall glycoprtoteins. Primarily extensins are highly glycosylated, a
feature that may reflect their interactions with carbohydrates in the cell wall. Biochemical
evidences supporting the existence of an extensin-pectin cross-link has been reported
(Showalter, 1993). It is assumed that extensins interact ionically with pectins in such a
way that the positively charged lysine and protonated histidine residues of extensins
interact with the negatively charged uronic acids of pectin (Fry, 1986). The cross-linking

- 19 -
of cell wall matrix is an important phenomenon, which results in alteration of
extensibility, digestibility and adherence of the cell wall. It has been found out that the
cell walls of elicitor-treated bean and chickpea cultured cells undergo a rapid extensin
insolubilisation and cross-linking and become tougher than the walls of untreated cells
(as determined by their resistance to digestion by protoplast enzymes) (Brisson et al.,
1994; Otte and Barz, 1996). This rapid insolubilisation and increased mechanical
strength of the wall might contribute to the active defence responses envisaged in
pathogen-plant interactions (Brisson et al., 1994) and resistance against pathogen
penetration. Besides, the HRGPs are also believed to be involved in cell-to-cell
interactions and reproduction. Rubinstein and co-workers (1995 a; b) found that
extensin-like HRGP from maize pollen tube wall is required for location of transmitting
tract of tissue by the pollen tubes. Advancing these findings, Cheung (1995) suggested
that the HRGPs are among the molecules, which participate in compatible pollen-pistil
interactions.
1.4. Monocots and dicots differ in their primary wall composition
In the evolutionary history of plants from their green-algae ancestors, one of the most
noticeable changes that occurred was the development of different kinds of cell walls
(Stebbins, 1992; Cassab, 1993). The plant cell wall determines morphology, thus in
classical sense, the cell wall has been and still is, the focal point for the evolution and
adaptation of plants. Observations from studies ranging from simple survey of
ecological plant anatomy to more sophisticated chemical image sectioning of primary cell
walls (Carpita, 2000) point to the adaptive importance of cell wall differentiation.
Therefore, it is very important to review the current knowledge of plant cell wall
diversity and adaptations of plant pathogens to primary cell walls of their respective host
plants.
The composition of primary walls of cereal grasses is remarkably different from that of
other flowering plants (dicots) and even other monocots (McNeil et al., 1984; Carpita
and Gibeaut, 1993; Carpita, 1996). On the basis of the actual differences existing
between the primary walls of monocots and dicots, structural models for two types of
primary walls has been provided (Carpita and Gibeaut, 1993). A type I primary cell wall,
a generalised cell wall of most flowering plants characterised by a network of cellulose

- 20 -
microfibrils with xyloglucans (XGs) as the principal interlocking polysaccharides
embedded in a pectin gel. In turn a type II primary cell wall represent a special cell walls
of Poaceae (grasses) where, the principal polymer that interlock the cellulose microfibrils
is glucuronoarabinoxylans (GAXs) instead of XGs. Since approximately 20 years ago
when the preliminary data on differences in the wall composition of monocots and
dicots were first documented (York et al., 1985), many studies have focused on
dissecting the structural elements of the primary wall of flowering plants, especially of
grasses (reviewed in Cosgrove, 1997; Carpita; 1996). In general, the wall of enlarging
plant cells is composed of approximately 30% cellulose 30%, hemicellulose, 35% pectin
with apparently 1-5% structural proteins on dry weight basis (Carpita, 1996). Substantial
deviations from these values are found in grasses. For example, the walls of growing
maize coleoptiles consists of approximately 55% hemicellulose, 25% cellulose and only
10% of pectin (Cosgrove, 1997). The two major constituents of flowering plant pectins,
polygalacturonic acid (PGA) and rhamnogalacturonan I (RGI), are found in much
smaller amounts with values as small as 3% in monocots (Darvill et al., 1980; Carpita,
1989; Shibuya and Nakane, 1984). On the other hand, xylan, the predominant
hemicellulose in the cell wall of grasses is generally a minor component (approximately
2%) of the primary walls of dicots (Jarvis et al., 1988).
The other architectural elements of the cell wall where a considerable difference between
grasses and dicots is observed are the kind and composition of structural proteins
(HRGPs). In general, the primary walls of Poales (gramineales) contain substantially less
structural proteins than other species (Carpita, 1996). The most studied HRGP,
extensins, are characteristic to dicot walls. Monocots contain glycoproteins that have
some homology with dicot extensins (Kiellszweski, 1994; Rubinstein et al., 1995 a; b).
However, the glycoproteins in monocots are HRGPs, which are also rich in threonine
(THRGPs), and histidine and alanine (HHRGPs). The biological significance of these
differences is not clearly known but the extensin-like glycoproteins studied from
monocots are reported to have structural and cell-to-cell interaction role similar to those
observed in dicot extensins (Stiefel et al., 1990; Rubinstein et al., 1995 a; b). The primary
wall of grasses has been a subject of intensive research during the recent years (Carpita,
1996; Carpita et al., 2000). Despite the wide differences in cell wall composition, there is
essentially no difference in developmental physiology between monocots and other
flowering plants (Carpita, 1996). Grasses respond similarly to environmental cues and

- 21 -
growth regulators, and undergo similar patterns of wall biogenesis during the
development of specific cell and tissue types (Shedletzky et al., 1992; Carpita, 1996;
Cosgrove, 1997; Carpita et al., 2000). However, in the event of wall disassembly during
microbial invasion, the wall composition would obviously influence the strategy of the
pathogen or enzymatic adaptations to the cell wall matrix.
1.5. Cell wall degradation
The assembly of the cell wall is a very complex process. The models presented, based on
studies on cell wall structure and functions (Carpita and Gibeaut, 1993), have made
significant advances in our understanding of the structure and assembly of the cell wall.
Thus it has provided a useful context from which to project our knowledge on the wall
matrix disassembly and degradation.
Cell wall degradation occurs in a wide variety of situations. Firstly, modification of the
primary wall is required for both cell expansion and for developmental events such as
seed germination, fruit softening and abscission, where the cell size remains static but
wall loosening is an important feature (Brett and Waldron, 1996; Hadfield and Bennet,
1998; Rose and Bennet, 1999). Secondly, it is an important part of the process by which
other organisms degrade plant materials such as, in the process of pathogen attack or
saprophytic invasions. Thirdly, cell wall degradation is a key process in industrial
processes and utilisation of plant biomass.
All the above processes of cell wall degradation involve enzymes that break the
backbone of cell wall polymers resulting in significant changes in the overall integrity of
the cell wall. Exceptions are expansins, which are proteins with no apparent hydrolytic
enzymatic activity but are thought to influence the hydrogen bonding between the
cellulose and hemicellulose (cellulose-xyloglucan network) of the cell wall so that the cell
is compliant enough to expand as the cell grows (Cosgrove, 1997; Rose and Bennet,
1999; Yuan et al., 2001). In general, the degradation of plant cell wall involves a
concerted and/or synergistic action of diverse enzyme families adapted to the different
cell wall polymers. Matching to the complexity of the components that make up the
plant cell wall, fungal plant pathogens are able to produce a broad range of extracellular
enzymes capable of degrading the plant cell wall (Table 1).

- 22 -
Table 1. Examples of cell wall degrading enzymes reported from plant pathogens.
Enzymes EC Reference (s)
-α-Arabinofuranosidase 3.2.1.55 Howell, 1975
-Cellulases
Endo-1,4-β-D-glucanase (carboxymethyl cellulase) 3.2.1.6 Sheppard et al., 1994 cellobiohydrolase 3.2.1.91 Sposato et al., 1995 β-glucosidase 3.2.1.21 Waksman, 1989
-Cutinase 3.1.1.74 Yao and Köller, 1994
-β-galactosidase 3.2.1.46 Cooper et al., 1988; Waksman, 1988
-Endo-1,4-β-D-glucanase 3.2.1.39 Cooper et al., 1988; Wang and Jones, 1995
-Exo-1,3,1,4-β-D-glucanase 3.2.1.58 Schaeffer et al., 1994 Van Hoof et al., 1991
-Laccase 1.10.3.2 Slomezynski et al., 1995
-Pectinases
Polygalacturonic acid lyase 4.2.2.2 Gonzales et al., 1992
Endo-polygalacturonase 3.2.1.15 Walton and Cervone, 1990
Exo-polygalacturonase 3.2.1.82 Garcia et al., 1997
Pectin methylesterase 3.1.1.11 Tans et al., 1998 -Proteases 3.4.22.29 Murphy and Walton, 1996
-Xylanases
Endo -1,4-β- xylanase 3.2.1.8 Southerton et al., 1993; Walton and Holden, 1990 β-xylosidase 3.2.1.37 Ransom and Walton, 1997
1.6. Cell wall composition, enzymic adaptation and plant-pathogen compatibility
The contact between extracellular wall degrading enzymes of pathogens and host cell
walls is often one of the first molecular interactions between the two and the outcome of
which can modify the type and/or balance of the host-pathogen relationship. The
nutritional environment (the amount and accessibility of the wall polymer), in which the
pathogen grows, dictates to a larger degree the type and quantities of cell wall degrading
enzymes produced (Anderson, 1978; Bateman and Basham, 1976). Hence, the disparity

- 23 -
between the wall composition of monocotyledonous and dicotyledonous plants might
determine enzymic adaptation of fungi and presumably contribute to the non-host
resistance. Cell walls from various species have a differential effect on the rate and extent
of enzyme synthesis but show no consistent relationship with compatibility or
incompatibility of the host-pathogen interactions. However, from the observations
presented from the various studies (Cooper, 1983; Cooper et al., 1981; Cooper et al.,
1988) it appears that each host-pathogen relationship should be considered unique and
generalisation may obscure individual effects. The more effective degradation of bean
walls compared to that of rice by an endo-PGL, and conversely of maize cell walls
compared with beans by xylanases is an illustration of enzymic adaptations to wall
structure and composition. Moreover, the adaptations of cereal pathogens to the
production of higher xylanase activities both in vitro and in planta than pathogens of
dicots, is believed to be a reflection of their basic compatibility established with their
monocotyledonous hosts (Cooper et al., 1988).
In view of these facts and the paucity of information on the role of xylanases in plant
pathogenesis, recent studies on cell wall degrading enzymes from cereal pathogens have
focused mainly on xylanases. Since about a decade efforts to provide definitive evidence
on the role of xylanases in plant diseases have been progressing. A detailed review of the
work done on fungal xylanases from only plant disease perspectives will be presented in
the following sections. For wide potential industrial use of xylanases and the remarkable
progress made in that line, the reader is referred to detailed and excellent reviews (Beg et
al., 2001; Sunna and Antranikian, 1997; Viikari et al., 1994).
1.7. Xylan and xylanases
1.7.1. Structure of xylan
Xylan is the predominant hemicelluose in the cell walls of plants. The structure of xylans
found in the cell walls of plants can differ greatly depending on the origin (Huisman et
al., 2000). However, xylans always contain a backbone of 1,4-β linked xylose (a five
carbon sugar) residues, which can be substituted by different side groups such as L-
arabinose, D-galactose, aceytyl, feruloyl, p-cumaroyl and glucuronic acid residues (Biely,
1985; Beg et al., 2001). Most xylans occur as heteropolysaccharides containing different

- 24 -
substituent groups in the backbone chain and in the side chains but some linear
polysaccharides or homoxylans are also known (Beg et al., 2001). Different plants or
different cell types have xylans, which contain these different side chains. The degree of
branching depends on the source. Xylans in the primary walls of monocotyledonous
plants (type II primary walls) contain large quantities of L-arabinose as side chains, and
therefore are often referred to as arabinoxylans (Carpita and Gibeaut, 1993; Ishii, 1984).
The secondary walls of monocotyledonous plants contain an arabinoxylan with more
glucuronic acid. The primary walls of dicotyledonous plants contain small amount of
glucuronoarabinoxylan, containing both glucuronic acid and arabinose side chains.
Arabinose is connected to the backbone of xylan via 1,2-α- or 1,3-α linkage either as
single residue or as short side chains. These side chains may also contain xylose 1,2-β-
linked to arabinose, and galactose, which can be either 1,5-β-linked to arabinose or 1,4-
β-linked to xylose. Acetyl residues are attached to 2-O or 3-O of xylose in the backbone
of xylan, but the degree of acetylation or structural complexity of xylan depends greatly
on the botanic source.
The most potential sources of xylan include many agricultural crops such as straw,
sorghum, sugar cane, corn stalks and cobs, hulls and husks from starch production as
well as forest and pulping waste products (Ebringerová and Henze, 2000). Xylan from
the primary walls of maize and sorghum are much more complex than in other cereals
such as wheat (Huisman, et al., 2000). At least three different components of
arabinoxylans, each with different side branches, are known to occur in maize (Carpita,
1983). This structural complexity makes xylans the most heterogeneous groups of
polysaccharides (Carpita, 1983; Huisman et al., 2000). The arbinoxylans of monocots
primary walls are substituted by ferulic acid (Schooneveld-Bergmans et al., 1998) and
these ferulic acid components are thought to cross-link the arabinoxylans in the wall
(Brett and Waldron, 1996). The occurrence of these different substituents and the
possible cross-linking with other polysaccharides is believed to have influence on the
enzymatic hydrolysis of xylans (Wong et al., 1988). The substituents may limit the
accessibility of the residual xylosyl residues to xylanolytic enzymes thereby preventing its
hydrolysis. On the other hand they may serve a positive role in substrate-enzyme binding
and thus enhance enzyme action. This has been demonstrated by the preferences of
some xylanases for hydrolysing branched xylans (Dekker and Richards, 1975).

- 25 -
1.7.2. Biodegradation of xylan
The complex chemical structure of xylan has been described as a linear polymer of
repeating xylopyranosyl groups substituted at various carbon positions with different
sugars and/or acidic compounds. Hence, complete and efficient enzymatic hydrolysis of
the complex polymer requires an array of enzymes with diverse specificity and modes of
action. The biodegradation of xylan backbone depends mainly on two classes of
enzymes, endo-1,4-β-xylanases (1,4-β-D-xylanohydrolase; EC 3.2.1.8), which hydrolyse
the 1,4-β-linked xylose backbone, and β-xylosidases (1,4-β-D-xylan xylanohydrolase; EC
3.2.1.37), which hydrolyses xylobiose and other short xylooligosaccharides resulting
from the action of endoxylanases. Debranching enzymes such as α-glucuronidase and α-
L-arabinifuranosidase and esterases such as acetylxylan esterases and ferulyl and p-
coumaroyl esterases, which remove acetyl and phenolic side branches, act synergistically
on the complex polymer (Beg et al., 2001; Thomson, 1993; Uffen, 1997; de Vries et al.,
2000). 1.7.3. Occurrence and production of xylanases
Xylanases are widespread in nature. They occur both in prokaryotes and eukaryotes and
have been reported from marine and terrestrial bacteria, rumen bacteria, fungi, marine
algae, snails, crustaceans, insects and seeds of terrestrial plants (Dekker and Richards,
1975). Microorganisms are primarily responsible for xylan degradation. Most of the
xylanases known to date are from bacterial and fungal sources (Sunna and Antranikian,
1997). Fungi are the most potent producers of xylanases. In culture, high level of
induction of xylanase activity is often achieved using xylan, other hemicellulose-rich
materials and low molecular weight carbohydrates such as xylose (Senior et al., 1989).
Fungi produce xylanases extracellularly (into the medium) thus avoiding the need for cell
lysis. Xylanases have been purified from large number of fungal and bacterial species
(Sunna and Antranikian, 1997 and the references therein) and detailed studies on their
biochemical properties have been carried out (Bastawade, 1992; Kulkarni et al., 1999).
Fungal xylanases have attracted special attention first because of the growing interest on
their potential use in paper and pulp industries (Viikari et al., 1994) and secondly,
because of their potential role in fungal pathogenicity (Walton, 1994). Accordingly,
- 26 -
xylanases have been purified and characterised from an increasing number of
phytopathogenic fungi and the genes encoding xylanases have been cloned and
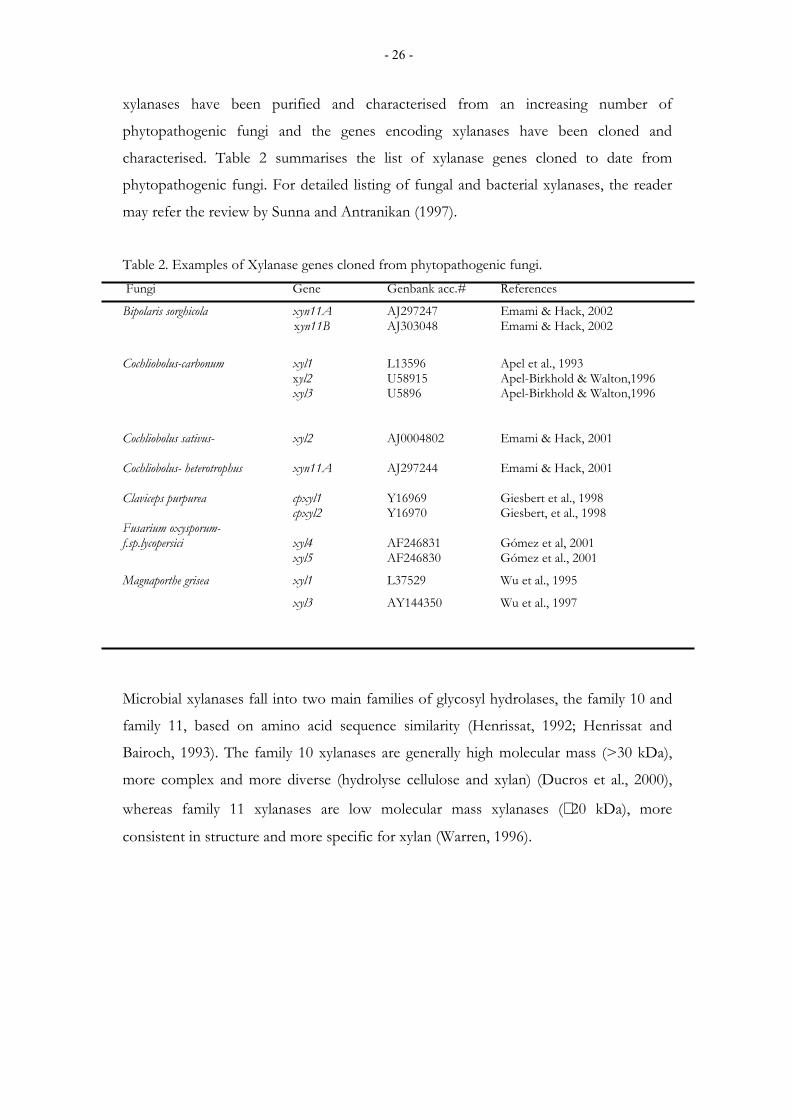
characterised. Table 2 summarises the list of xylanase genes cloned to date from
phytopathogenic fungi. For detailed listing of fungal and bacterial xylanases, the reader
may refer the review by Sunna and Antranikan (1997).
Table 2. Examples of Xylanase genes cloned from phytopathogenic fungi. Fungi Gene Genbank acc.# References
Bipolaris sorghicola xyn11A AJ297247 Emami & Hack, 2002 xyn11B AJ303048 Emami & Hack, 2002
Cochliobolus-carbonum xyl1 L13596 Apel et al., 1993 xyl2 U58915 Apel-Birkhold & Walton,1996 xyl3 U5896 Apel-Birkhold & Walton,1996
Cochliobolus sativus- xyl2 AJ0004802 Emami & Hack, 2001 Cochliobolus- heterotrophus xyn11A AJ297244 Emami & Hack, 2001 Claviceps purpurea cpxyl1 Y16969 Giesbert et al., 1998 cpxyl2 Y16970 Giesbert, et al., 1998 Fusarium oxysporum- f.sp.lycopersici xyl4 AF246831 Gómez et al, 2001 xyl5 AF246830 Gómez et al., 2001
Magnaporthe grisea xyl1 L37529 Wu et al., 1995
xyl3 AY144350 Wu et al., 1997
Microbial xylanases fall into two main families of glycosyl hydrolases, the family 10 and
family 11, based on amino acid sequence similarity (Henrissat, 1992; Henrissat and
Bairoch, 1993). The family 10 xylanases are generally high molecular mass (>30 kDa),
more complex and more diverse (hydrolyse cellulose and xylan) (Ducros et al., 2000),
whereas family 11 xylanases are low molecular mass xylanases (∼ 20 kDa), more
consistent in structure and more specific for xylan (Warren, 1996).

- 27 -
1.7.4. Xylanase multiplicity: a mere genetic redundancy or a display of functional diversity?
Multiplicity is a common phenomenon in microbial xylanases (Wong et al., 1988)
including plant pathogenic fungi (Table 2). Wu et al. (1997) reported as many as five
xylanases from the rice blast fungus Magnaportha grisea, and at least four different
xylanases have been identified from the maize leaf spot fungus, Cochliobolus carbonum
(Apel et al., 1993; Apel-Birkhold and Walton, 1996), each differing in molecular weight
and pI values. The question concerning how many xylanases could occur in a given
fungal species is difficult to answer because practically always the purification and
characterisation of xylanases is conducted from in vitro fungal cultures. The purification
procedures favour the "major" xylanases or those with activity high enough to allow
purification and those activities too little to allow purification or "minor" xylanases
remain obscured. It should be noted, however, that minor xylanases are so called
because of the relatively low amount produced at the given culture condition but this
does not necessarily mean that they have minor role in their biological functions. Minor
xylanases may have biological functions, which are not required in large quantities. It is
also assumed that some xylanases may not be not be detected during the assay because
they might be lost from the culture filtrate due to degradation or adsorption onto
insoluble growth substrate (Wong et al., 1988). Another interesting observation comes
from the xylanase gene knockout studies of the rice blast fungus M. grisea (Wu et al.,
1997) where the removal by gene disruption of one of the major xylanase (xyl2) released
three additional xylanases in the mutant strain that have not been detected in the parent
strain. This indicates the rather complex nature of the phenomenon of xylanase
multiplicity in fungi (Apel et al., 1993; Apel-Birkhold and Walton, 1996). Multiple
xylanases may be allozymes, products of different allels of the same gene, or they could
be distinct gene products produced by a fungus to enhance its utilisation of xylan (Wong
et al., 1988; Uffen, 1997).
The occurrence of multiple xylanases in fungi raises many questions. Firstly, what is the
functional importance of xylanase multiplicity in the fungus? Are the multiple isozymes
redundant enzymes or enzymes with unique functional significance? Are xylanolytic
systems with low degree of xylanase multiplicity inferior to those with high degree of
multiplicity? Does it contribute to plant tissue specificity in plant-pathogen interactions

- 28 -
or to the over all pathogenicity of the fungus? At present it is difficult to find definite
answers to these questions since information on the characterisation of individual
enzymes and xylanolytic systems is very limited. Wong et al. (1986) studied the
functional importance of three xylanases from the saprophytic fungus Trichoderma
harzianum and reported a high degree of complementation of the three xylanases in the
hydrolysis of aspen xylan. The conclusion from their finding was that the three xylanases
are not redundant enzymes since each contributes significantly and uniquely to the
hydrolysis of the xylan. In plant pathogenic fungi, it reported that some of the xylanases
are induced only during infection (Apel-Birkhold and Walton, 1996) suggesting those
different sets of endoxylanases function in saprophytic and pathogenic growth of fungi.
It is also speculated that isozymes of cell wall degrading enzymes (CWDEs) are
produced at different stages during infection of plant tissue (Annis and Goodwin, 1997)
possibly following biochemical changes in the host environment.
The multiple occurrences of xylanases in plant pathogenic fungi have been one of the
hurdles in an attempt to determine their role in pathogenicity by gene disruption. The
generation of mutants lacking xylanase activity has not been possible in C. carbonum and
M. grisea, the two fungal pathogens where xylanase gene disruptions have been carried
out (Apel et al., 1993; Apel-Birkhold and Walton, 1996; Wu et al., 1997). More
information about the extent and nature of the multiplicity, as well as the functional
importance and regulation of this phenomenon in fungal xylanolytic systems would be
useful for better understanding of the system. It would also provide better research
approach to evaluate their role in plant-pathogen interactions.
1.7.5. Regulation of xylanase gene expression
The mechanism of control of xylanolytic enzymes synthesis varies considerably among
different organisms. Individual xylanases from an organism might be under different
control (De Vries and Visser, 2001). In general, xylanase expression in fungi is subject to
substrate induction and glucose or catabolite repression (Apel et al., 1993; De Vries and
Visser, 2001; Gómez-Gómez et al., 2002; Tonukari et al., 2002;). Only a rare case of
constitutive xylanase expression of xylanases has been reported (Srivastava and

- 29 -
Srivastava, 1993). Therefore, induction, catabolic repression, growth rate and other
environmental factors can influence the activity of xylanolytic enzymes.
1.7.5.1. Induction Xylanase induction is a complex phenomenon. High molecular weight xylan cannot
enter the cells and therefore cannot directly induce the synthesis of xylanolytic enzymes.
The low molecular mass fragments of xylan are known to play an important role in
xylanase biosynthesis (Kulkarni et al., 1999). These fragments include xylose, xylobiose
and xylooligosaccharides. In a hypothetical model proposed by Thompson (1993), these
small soluble (signal) fragments are released by the action of low amount of
constitutively produced xylanases which degrade the xylan to xylooligosaccharides and
xylobiose that are further taken up by the cell and induce other xylanase genes. The
inducible xylanases degrade xylan to xylooligosaccharides and xylobiose. The β-
xylosidases, which may be produced constitutively and/or inducibly, convert xylobiose
to xylose and may subsequently transxylanate or transglycosylate it to Xyl1B1-2Xyl and
Glcb1-2Xyl. These compounds are believed to be taken up by the cell and act as
additional inducers of genes encoding xylanases allowing the utilisation of xylan
(Kulkarni et al., 1999; Thompson, 1993). Recent evidence also indicates that xylose is
not always an inducer of xylanase expression in Aspergillus but acts rather as a repressor
at higher concentrations (de Vries et al., 1999) through the well-known process of
carbon catabolic repression. Studies on Aspergillus and Trichoderma spp. at the cellular and
molecular level indicate that xylanase gene expressions are regulated at the
transcriptional level (de Graaff et al., 1994; Margolles-Clark et al., 1997) and that a
transcriptional activator XlnR (positive acting element) is known to regulate induction of
xylanase expression. In general, far less is known about the mechanism of by which
xylanase-encoding genes are induced.
1.7.5.2. Carbon catabolic repression
Carbon catabolic repression in microorganisms is a means to control the synthesis of a
range of enzymes required for the utilisation of less favoured carbon source when more
readily utilisable carbon sources are available in the medium. Microorganisms are
reported to turn off a large number of genes in the presence of glucose as an energy
saving response, as it primarily affects enzymes used to metabolise other carbon sources

- 30 -
which are dispensable in the presence of glucose (Ronne, 1995). Work with yeast and
filamentous fungi has revealed a mechanism for glucose repression in eukaryotes that is
different from that found in bacteria. Zinc finger proteins such as MIG 1 from yeast and
CREA from Aspergillus nidulans, that bind GC-boxes in the promoters, play a key role in
mediating this response (Dowzer and Kelly, 1991; Ronne, 1995). The sequence 5'
SYGGRG 3' is known to be the consensus for CREA (CRE1-homologue) binding in
the promoter region of xylanase genes (de Vries and Visser, 2001; Kulkarni, 1999). The
zink binding domains of both CreA and Mig1 are reported to be very similar since both
genes are involved in glucose repression (Kulumburg et al., 1993; Ronne, 1995).
In yeast the expression of glucose-repressed genes is regulated by a protein kinase
known as SNF1 (Ronne, 1995; Treitel et al., 1998). Cells that lack the SFN1 are
therefore unable to use carbon sources other than glucose. A simple model proposed
(Ronne, 1995) for glucose repression is that glucose generates a signal that inhibits or
counteracts the activity of Snf1. Therefore, mutation in Snf1 gene causes an irreversible
downregulation of the expression of the derepression gene so that glucose repressed
genes remain repressed. The expression of almost all xylanase genes reported from plant
pathogenic fungi is inhibited by glucose (Gómez-Gómez et al., 2002; Tonukari et al.,
2002). An ortholog of the Snf1, ccSnf1, has been cloned for the first time from the plant
pathogenic fungus C. carbonum (Tonukari et al., 2000). It is demonstrated that mutation
of ccSnf1 gene reduced the growth of the fungus on complex carbon sources, down
regulated the expressions of the genes, and eventually reduced the virulence of mutants
on maize. Thus Tonukari and co-workers concluded that the Snf1 gene is required for
the expression of glucose repressed xylanase genes of C. carbonum and for the virulence
on maize. This is the first instance of linking a regulatory pathway to the study of fungal
pathogenesis suggested by Hamer and Holden (1997). So far only one negatively acting
factor CreA (carbon catabolic repression, discussed above) and one positively acting
factor (a transcription activator, XlnR, (van Peij et al., 1998) have been studied in detail.
The actual level of expression of xylanase genes appears to be influenced by the balance
between the induction by XlnR and repression by CreA (de Vries et al., 1999).

- 31 -
1.7.6. Xylanase inhibitor proteins (XIPs)
Plants and their pathogens have evolved several inter-related biochemical processes that
plants use to defend themselves and that pathogens use to colonise their hosts (Rose et
al., 2002). The end result of these interactions determines whether the plant becomes
infected or not. The enzymes from pathogens that degrade plant cell walls and plant-
synthesised proteins that inhibit these cell wall degrading enzymes of pathogens, are one
example of such interacting systems. Polygalacturonase inhibitor proteins (PGIPs) are
present in the cell wall of wide variety of dicots (Albersheim and Anderson, 1971;
Devoto et al., 1998) and a pectin lyase inhibitor protein (PNLIP) has been isolated from
sugar beet (Bugbee, 1993). Moreover, other proteinaceous inhibitors of pectin methyl
esterase (Camardella et al., 2000), α-amylase (Weselake et al., 1983) and invertase
(Greiner et al., 1998) have been identified from plants. The production of inhibitory
proteins appears to be a mechanism of a counterdefence evolved by both plants and
pathogens. The discovery of glucanase inhibitor proteins (GIPs) that inhibit endo-β-
glucanases from the oomycete pathogen Phytophthora sojae (Rose et al., 2002) underlines
the importance of this coevolution of counterdefence in plant-pathogen interactions.
Very recently the occurrence of proteins that inhibit xylanases have been reported from
wheat, barley and rye (Debyster et al., 1999; Gebruers et al., 2001). Currently three
xylanase inhibitor proteins (XIPs) differing in molecular weight, pI and endoxylanase
specificity have been purified and characterised from wheat flour. One of these
inhibitory proteins (XIP1), described by McLauchlan et al. (1999) is monomeric
glycosylated has a molecular mass of 29 kDa, showing no sequence homology with the
two other Wheat (Triticum aestivum) xylanase inhibitor proteins (TAXI I and TAXI II)
characterised by Gebruers et al. (2001). Instead, its N-terminal amino acid sequence has
86% homology with rice chitinase III (McLauchlan, 1999). TAXI I and TAXI II, on the
other hand, have similar N-terminal amino acid sequences, a comparable molecular mass
of approximately 40 kDa but differ in endoxylanase specificity. In contrast to the XIP1,
TAXI I and TAXI II are not glycosylated (Gebruers et al., 2001). These xylanase
inhibitor proteins inhibit both family 10 and family 11 fungal xylanases but show some
degree of specificity depending on the source of the xylanase (Flatman et al., 2002;
Gebruers et al., 2001; McLauchlan, 1999; Tahir et al., 2002). For example a family 11
xylanase from Aspergillus niger is inhibited by TAXI I (Gebruers et al., 2001) and XIP1

- 32 -
(McLauchlan, 1999) but not by TAXI II (Gebruers et al., 2001). Recently, Furniss et al.
(2002) reported from Penicillium funiculosum a xylanase gene (xync) that is strongly
inhibited by all the three known xylanase inhibitors. The mechanism of inhibition is
known to be competitive, that is the inhibitor protein binds at, or close to the active site
of xylanases, thus preventing its access to the substrate (Flatman et al., 2002; Furniss et
al., 2002; Tahir et al., 2002).
Investigations on xylanase inhibitor proteins started only recently. The inhibitor proteins
that are studied in most detail are the polygalacturonase inhibitor proteins (PGIPs).
PGIPs are leucine rich repeat (LRR) proteins present in the cell wall of dicots and non-
graminaceous monocots (Albersheim and Anderson, 1971), and they are proposed to
form part of the plant "immune system" to pathogen attack (De Lorenzo et al., 2001).
PGIPs are capable of inhibiting up to 99% of the activity of fungal polygalacturonases
(Cervone et al., 1989). It is assumed that PGIPs benefit the plant by decreasing the
hydrolytic activity of endo-cleaving fungal PGs. This retards the action of PGs in
opening up the wall to access by other cell wall degrading enzymes and thus increases
the longevity of the elicitor active (DP 10-15) oligogalacturonides (Cook et al., 1999;
Garćia-Romera and Fry, 1995). In the absence of PGIPs, biologically active
oligogalacturonides are rapidly hydrolysed by PGs in vitro, forming smaller, inactive
oligomers and monomers (Cook et al., 1999; Garćia-Romera and Fry, 1995). It has been
found that fungal polygalacturonases exhibit different substrate degradation patterns and
also differ in their susceptibilities to PGIPs (Cook et al., 1999) suggesting the necessity
of several PGIPs with different specificity. This has been confirmed by Desiderio et al.
(1997) in a study where they demonstrated the expression of PGIPs of different
specifities from bean (Phaseolus vulgaris).
Endoxylanase inhibitor proteins may well be involved in plant defence mechanisms.
Therefore, their discovery has profound implications in further unravelling the plant
defence system and opens a new area of research for improvement of plant disease
resistance against fungal pathogens.

- 33 -
1.7.7. Xylanase in plant pathogenesis
1.7.7.1. Xylanases as pathogenicity or virulence factors
It is widely accepted that pathogens attack plants because during their evolutionary
development they have acquired the ability to live off the substances manufactured by
the host plants (Agrios, 1997). Therefore, as a rule of thumb if the host makes a
substance, the putative parasite will produce an enzyme to metabolise it. But for the
pathogen to invade and obtain nutrients from the plant, it has to first degrade the cell
wall, which is the first line of defence. Although some pathogens may use mechanical
force to penetrate the plant cell wall, the activities of pathogens in plants are largely
biochemical. Therefore, the effects caused by pathogens on plants are almost entirely the
result of biochemical reactions taking place between substances secreted by the
pathogen and those present in, or produced by the plant (Agrios, 1997). Cell wall
degrading enzymes are among the main groups of substances produced by pathogens,
and enzymatic degradation of the cell walls is the most conspicuous effects of
microorganisms on plants (Walton, 1994).
Since over a century, cell wall degrading enzymes have been considered as the primary
agents for tissue maceration and diseases in plants. However, proof of their role in
pathogenesis had to await the development of the technology needed for enzyme
purification and the more powerful molecular techniques such as gene cloning and
targeted gene disruption.
For many years research on cell wall degrading enzymes has focused on pectinases
(endo- and exo-pectin lyase, endo- and exo-polygalacturonases and pectin methyl
esterases) from pathogens of dicotyledonous plants because of the abundance of pectin
in the primary wall of dicots. As a result, significant success has been achieved in
understanding their regulation and their role in pathogenicity in bacterial and fungal
pathogens.
In the last decade, however, fungal xylanases particularly from the pathogens of cereals
have attracted greater attention. Many lines of evidence suggest that xylanases might be a
pathogenicity factor for cereal pathogens. Firstly, the primary wall of cereals comprises
predominately of xylan (Carpita and Gibeaut, 1993) making it a target substrate for

- 34 -
microbial degradation. Secondly, cereal pathogens produce xylanases and little or no
pectinases when grown on isolated plant cell wall in culture as well as during infection in
planta (Cooper et al., 1988; Braun and Rodrigues, 1993). Thirdly, fungal xylanases kill
cultured rice cells (Ishii, 1988). This indirect, but cumulative evidence suggests that
xylanases might be virulence factors in cereal pathogens. Efforts to determine the direct
role of cell wall degrading enzymes in plant diseases has a long history (Bateman, 1976).
Earlier approaches depend on the evaluation of the effect of purified enzymes on plant
cells, and integration of biochemical and immunocytochemical technologies (Bateman,
1976 and references therein). Despite the apparently convincing evidence, this has not
resulted in any definitive conclusions on the importance of cell wall degrading enzymes
in disease process.
Classical genetic studies with fungal mutant strains created by irradiation or mutagenic
chemicals, show reduced cell wall degrading enzyme activities (Durrands and Cooper,
1988). The general drawback of this method is the impression of the mutagenic agent,
where many genes besides the gene of interest are inactivated. This often results in
growth abnormalities and defects in production of other enzymes. During the last
decade a more stringent molecular genetic technique, commonly known as
transformation-mediated gene disruption (discussed in detail under 1.10.2) has been
widely employed to characterise the role of cell wall degrading enzymes in plant diseases.
Accordingly, a large number of fungi have been transformed (Hynes, 1996) and cloned
genes encoding putative pathogenicity factors from several fungi have been disrupted
(Walton, 1994). Xylanase genes have been disrupted and characterised in most detail
from at least two plant pathogenic fungi, namely C. carbonum (Apel et al., 1993; Apel-
Birkhold and Walton, 1996) and M. grisea (Wu et al., 1997). However, in both fungi the
targeted disruption of the individual xylanase genes resulted in mutants of altered
xylanase activity but failed to produce non-pathogenic phenotypes. Genetic redundancy
or enzyme multiplicity in fungi (discussed above) has been a hurdle for scientists carrying
out disruption of genes encoding cell wall degrading enzymes including xylanases (Apel
et al., 1993; Apel-Birkhold and Walton, 1996; Murphy and Walton, 1996; Scott-Craig et
al., 1990; Sposato et al., 1995; Stahl and Schaeffer, 1992; Sweigard et al., 1992; van Kan
et al., 1997; Wu et al., 1997). Surprisingly, disruption of a xylanase xyl2 in M. grisea
revealed three additional xylanases (Wu et al., 1997) distinct from the two other xylanase
genes previously reported (Wu et al., 1995). Therefore, the definitive role of xylanases in

- 35 -
fungal pathogenesis remains to be an open question until mutants lacking xylanase
activity and/or mutants completely deficient in their ability to degrade xylan are
obtained.
1.7.7.2. Xylanases as elicitors of defence responses in plants
The interaction between plants and incompatible pathogens induces a large variety of
defence responses. This inducible reaction results either from transcriptional activation
of cascades of plant defence response genes (Avni et al., 1994; Lotan and Fluhr, 1990) or
from specific enzymatic activation (Bradley et al., 1992). These responses are frequently
associated with localised responses leading to hypersensitive cell death (Atkinson, 1993),
which are often initiated by signals generated by organic compounds known as elicitors.
Elicitors could as well be substances of non-biological origin such as inorganic salts.
Elicitors that trigger defence responses have been isolated from phytopathogenic fungi
and saprophytes (Aktinson, 1994; Blein et al., 1991; Dean and Anderson, 1991; Dean et
al., 1989; De Wit, 1986). A 22 kDa endo-1,4-β-xylanase referred to as ethylene-inducing
xylanase (EIX) was isolated from Trichoderma viride cultures (Dean et al. 1989; Fuchs et
al., 1989). When applied to tobacco (Nicotiana tabacun var xanthi) or tomato (Lycopersicum
esculentum) leaves, EIX induces ethylene biosynthesis, the production of phytoalexins and
pathogenesis-related proteins, and causes necrosis and hypersensitive cell death (Bailey et
al., 1990; Avni et al., 1994 b; Dean et al., 1990; Lotan and Fluhr, 1990). In suspension-
cultured cells of tobacco and tomato EIX induces a rapid medium alkalinization,
oxidative burst, and ethylene biosynthesis (Bailey et al., 1992; Felix et al., 1993; Yano et
al., 1998). The response of tobacco to EIX is cultivar specific (for example the tobacco
cultivar Hicks is insensitive to EIX unlike xanthi) suggesting that there is a requirement
for EIX recognition factors in the plant (Bailey et al., 1993). Like cultivar response in
plant-pathogen interactions, the cultivar specific response of EIX or the sensitivity to
EIX of tobacco var xanthi is controlled by a single dominant gene (Bailey et al., 1993).
Hanania and Avni (1997) also report a high affinity-binding site for EIX elicitor only in
membranes of tobacco and tomato that are genetically capable of responding to the
elicitor.

- 36 -
Proteinaceous elicitors displaying hydrolytic enzyme activity are generally released from
the plant or cell wall fragments of fungus (the specific elicitor compounds) that are
capable of killing plant cells (Bucheli et al., 1990; Walker-Simmons et al., 1984). With
EIX, however, no such active xylan fragments have been identified (Bailey et al., 1993;
Fuchs et al., 1989; Dean et al., 1989). Therefore, the elicitation of plant response by EIX
is believed to be a direct interaction between the peptide and the cell, and it is therefore
not related to the cell wall degradation activity of the enzyme (Enkerli et al., 1999;
Sharon et al., 1993). This conclusion was based on two observations. Firstly, EIX elicits
ethylene biosynthesis in protoplasts (Sharon et al., 1993). Secondly, a mutant xylanase of
reduced activity maintained its potency of elicitor activity by fully inducing medium
alkalinisation and ethylene biosynthesis in suspension cultured tomato cells and
hypersensitive necrosis in leaves of tomato and tobacco plants (Enkerli et al., 1999),
comparable to the wild type elicitor protein.
1.8. Molecular evolution of xylanases
Many microorganisms that hydrolyse xylans also degrade cellulose. In view of the
similarity of the bond cleaved (1,4-β-glucosidic linkages), and cross-specificity sometimes
observed between cellulase and xylanase, the phylogenetic relationships of these
enzymes have been a subject of interest. Xylanases showed no convincing sequence
similarity with cellulases suggesting that the two enzyme species evolved from distinct
ancestral genes. Amino acid sequence alignment and hydrophobic cluster analysis have
been utilised to assign plant cell wall hydrolases into different families (Henrissat and
Bairoch, 1993). Based on these criteria, xylanases are divided into two families; family 10
and family 11, the characteristic of which are discussed elsewhere (section 1.7.3) In
general the two families are not homologous in a phylogenetic sense, and it is therefore
difficult to study their relationship by simple alignments. Few bifunctional xylanases that
contain both family 10 and family 11 xylanases have been identified. These include the
xylanases from the bacterium Ruminococcus flavefaciens (Accession no. P29126 and S61204)
and the fungus Neocallimastix patriciarum (Accession no. P29127). The tight grouping of
fungal and bacterial taxa, which produce bifunctional enzymes, suggests the possibility
that xylanase evolution might proceed through horizontal transfer of the DNA encoding
catalytic domains as suggested by Henrissat et al. (1995). Some phylogenetic studies

- 37 -
suggest that generally less horizontal gene transfer has occurred within the family 11
than the family 10 xylanases (Georis et al., 1999).
1.9. Helminthosporium turcicum: the cause of northern leaf blight of maize 1.9.1. Taxonomy and etiology
Helminthosporium turcicum Pass. (syn. Exserohilum turcicum (Pass.) Leonard and Suggs,
Bipolaris turcica (Pass.) Shoemaker, Drechslera turcica (Pass.) Subram. and Jain.) Perfect state
Trichometasphaeria turcica Luttrell. (Syn. Setosphaeria turcica (Luttrell) Leonard and Suggs.) is
an ascomycete with characteristic conidiophores emerging singly or in groups of 2-6,
straight or flexuous, brown up to 300 µ long. Conidia are straight or slightly curved, pale
to mid straw coloured, 3-8 pseudoseptate and 50-144 µ long (Shurtleff, 1980). T. turcica,
the sexual stage, occurs rarely, if ever in nature but produces black globose pseudothecia
in the laboratory. The asci are cylinderical with a short stipe and contain one to eight,
but usually two to four ascospores. H. turcicum is the causal agent of northern leaf blight
of maize.
1.9.2. Distribution and economic importance
Northern leaf blight is one of the most important diseases of maize (Zea mays L) in warm
and humid part of the world (Ceballos et al., 1991; Shurtleff, 1980). In Africa where
maize is the staple food, northern leaf blight is reported to be widespread and
destructive disease of maize in warm and humid growing regions of Ethiopia, Tanzania
and Uganda (Adipala et al., 1993; Nkonja et al., 1998; Tilahun et al., 2001). In the United
States, northern leaf blight is the major disease of maize (Bowen and Pedersen, 1988;
Perkins and Pedersen, 1987). The magnitude of yield losses caused by northern leaf
blight depends on two factors: the stage of maize growth when the infection occurs and
the severity of the disease. If the disease establishes before silking, yield reductions upto
40% may occur (Raymundo and Hooker, 1981), but if infection is delayed by 6-8 weeks
after silking, yield losses are minimal. H. turcicum infected maize plants are often
predisposed to root and stalk rot infections (Raymundo, 1981).

- 38 -
In general the early maturing varieties of maize are more susceptible to H. turcicum
infection than the late varieties (Ceballos et al., 1991; Degefu, 1990). In Ethiopia,
northern leaf blight is a serious challenge to the national maize-breeding programme for
moisture stress areas, where early maturing maize varieties are the only choice.
1.9.3. Infection and disease development
Conidia are wind-borne over long distances to leaves of maize plants. Conidial
germination occurs 3-6 hours after inoculation (Hilu and Hooker, 1964). Germ tubes
grow at an angle rather than parallel to the veins of the leaf producing an appressoria,
from which a penetration peg develops. Penetration of the maize cell wall, which is very
rich in complex arabinoxylan (up to 60%) (Cosgrove, 1997; Huisman et al., 2000), is
usually direct and occurs very rarely through the stomata (Knox-Davies, 1974). Xylan
degradation may have a key role for penetration. Macroscopic symptoms appear first as
minute light green to whitish flecks. Hyphae grow in the mesophyl cells in these flecks
and in cells beyond the necrotic tissue of the flecks. Flecks enlarge either singly or
coalesce to form long elliptical, greyish-green tan lesions ranging from 2.5 to 15 cm in
length.
1.9.4. Disease cycle and epidemiology
H. turcicum overwinters as mycelia and conidia in infected leaves, husks, and seeds.
Conidia are wind-borne over long distances to leaves of maize plants. Disease
development is favoured by moderate temperatures (18-27 °C) and heavy dew during
the growing season (Shurtleff, 1980). Further spread of the disease within and between
fields occurs by conidia produced abundantly on leaf lesions. Northern leaf blight is
generally known to be sporadic in occurrence, depending on the environmental
conditions and the level of disease resistance in the variety (Degefu, 1990; Perkins and
Pedersen, 1987).
1.9.5 Pathogenic races, sources of resistance and control of H. turcicum
H. turcicum attacks sorghum, Sudangrass and Johnsongrass. Races of H. turcicum on
sorghum and Sudangrass apparently do not infect maize but some maize isolates are

- 39 -
reported to infect Sudangrass (Shurtleff, 1980). The fungus is known to be highly
variable in cultural characteristics and pathogenicity. Knox-Davies and Dickson (1960)
reported sufficient evidence of heterokaryons and their perpetuation through the conidia
and suggested that the high variability in the fungus might be related to heterokaryosis.
At least four races of H. turcicum are reported (Leonard, 1989). Race designation is done
according to the resistance genes in maize that their virulence matches (Table 3). There
are two types of resistance to H. turcicum in maize: monogenic and polygenic resistance.
Monogenic or race specific resistance, controlled by Ht1, Ht2 and Ht3 genes, is
characterised by formation of chlorotic lesions and reduced pathogen sporulation that
delays the onset of epidemics, or as in HtN genes, delays lesion formation in such a way
that plants in the field with HtN gene remain free from lesions until shortly after
pollination (Leonard, 1989). The Ht1 gene is used more extensively in commercial maize
hybrids in the United States, whereas Ht2, Ht3, HtN are limited only to experimental use
(Leonard, 1989). The polygenic resistant (rate reducing partial resistance etc.) genotypes
are characterised by small and few lesions. The occurrence of genes in H. turcicum
matching the four resistance genes in maize is an indication of a potential for many more
pathogenic races to be identified.
Table 3. Races of H. turcicum and matching resistance genes in maize.
Disease reaction
Race designation Ht1 Ht2 Ht3 HtN
0 R R R R 1 S R R R 23 R S S R 23N R S S S
R=resistant, S=Susceptible. Table adapted from Leonard (1989).
The most efficient and economically practical method for the control of northern leaf
blight is using resistant hybrids or varieties. However, in seed production fields
fungicides may be applied, beginning from the time when lesions are first observed
(Shurtleff, 1980).
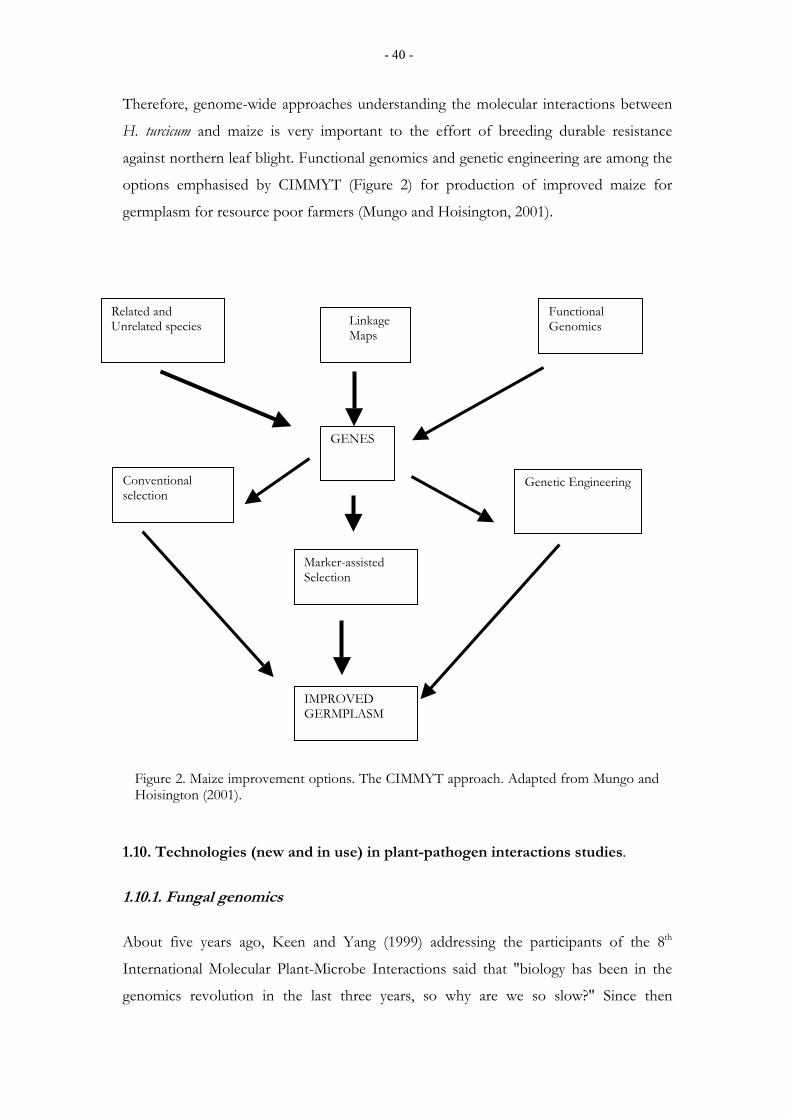
- 40 -
Therefore, genome-wide approaches understanding the molecular interactions between
H. turcicum and maize is very important to the effort of breeding durable resistance
against northern leaf blight. Functional genomics and genetic engineering are among the
options emphasised by CIMMYT (Figure 2) for production of improved maize for
germplasm for resource poor farmers (Mungo and Hoisington, 2001).
1.10. Technologies (new and in use) in plant-pathogen interactions studies.
1.10.1. Fungal genomics About five years ago, Keen and Yang (1999) addressing the participants of the 8th
International Molecular Plant-Microbe Interactions said that "biology has been in the
genomics revolution in the last three years, so why are we so slow?" Since then
Related and Unrelated species Linkage
Maps
Functional Genomics
GENES
Conventional selection
Genetic Engineering
Marker-assisted Selection
IMPROVED GERMPLASM
Figure 2. Maize improvement options. The CIMMYT approach. Adapted from Mungo and Hoisington (2001).

- 41 -
functional genomics of plant-microbe interactions gained momentum. Just before the
start of the writing this thesis, I attended the tenth New Phytologist Symposium in
Nancy, France, under the theme "Functional Genomics of Plant-Microbe Interactions"
where remarkable progress in a number of genome-wide projects in plant-pathogen and
symbiotic interactions have been presented.
Genomics can be defined as the study of genomes, which includes the discovery of
functions and regulation of gene expressions (functional genomics), the organisation and
structure of genes and chromosomes (structural genomics) and the evolutionary history
of genes and organisms (comparative genomics). However, much more than the study of
individual genes, genomics implies the study of interacting networks of genes that make
up whole organism. The technology facilitates the evaluation of the entire set of genes in
an organism, or the expressions of genes at any one time or particular condition.
By definition, having the complete genome sequence of a pathogen means all genes
required for virulence are in hand. The question is which ones they are and what is their
specific role. To date the identification of new genes is occurring at much faster rate
than the determination of their function. The availability of a complete genome
sequence for a pathogen permits the systematic mutation of all suspect genes and testing
of the resulting mutant strain in growth and pathogenicity assays. The ability to
selectively and reliably create site-specific “gene knockouts” or homologous
recombination of genes of interest is very crucial. Genomic approaches to study fungal
pathogens started very recently compared to that of many other organisms. Therefore,
to date no pathogenic fungus has had its genome completely sequenced (Soanes et al.,
2002) and the contribution of fungal genome data to the general understanding of
pathogenicity and disease control are in their infancy (Mitchell et al., 2003). Most
laboratories focusing on molecular genetic analysis of fungal plant pathogens are still in
"pre genomics era", which is characterised by independent analysis of individual genes.
However, a broader and higher-throughput study for functional analysis pathogenicity
genes is possible and essential. Some examples such efforts are presented in Table 4.

- 42 -
Table 4. Publicly accessible pathogenic fungal and oomycete genome projects
Fungus URL
Aspergillus fumigatus http://www.sanger.ac.uk/projects/A_fumigatus
Blumeria graminis http://cogeme.ex.ac.uk/index.html
Botrytis cinerea http://cogeme.ex.ac.uk/index.html
Candida albicans http://cogeme.ex.ac.uk/index.html
Cladosporium fulvum http://cogeme.ex.ac.uk/index.html
Colletotrichum trifoli http://cogeme.ex.ac.uk/index.html
Cryptococcus neoformans http://www.genome.ou.edu/cneo.html
http://baggage.stanford.edu/group/c.neoformans/menu.html
Fusarium sporotrichioides http://cogeme.ex.ac.uk/index.html
http://www.genome.ou.edu/fsporo.html
Gibberella zeae http://cogeme.ex.ac.uk/index.html
Leptosphaeria maculans http://cogeme.ex.ac.uk/index.html
Magnaporthe grisea http://www.fungalgenomics.ncsu.edu
Mycosphaerella graminicola http://cogeme.ex.ac.uk/index.html
Phytophthora infestans http://cogeme.ex.ac.uk/index.html
Phtophthora sojae http://cogeme.ex.ac.uk/index.html
Verticillium dahliae http://cogeme.ex.ac.uk/index.html
1.10.2. Fungal transformation and gene disruption
An effective way to study the function of genes in an organism is to disrupt the genes
and analyse the gene-disrupted mutants. To date transformation-mediated gene
disruption is a technique widely employed in the molecular genetic analysis of plant-

- 43 -
microbe interactions and study of plant pathogenic fungi (for latest review see Mullins
and Kang, 2001). As a result, fungal mutants of altered activities of cell wall degrading
enzymes have been produced and characterised (Apel-Birkhold and Walton, 1996;
Murphy and Walton, 1997; Rogers et al., 1994; Scot-Craig et al., 1990; Sposato et al.,
1995; Wu et al., 1997).
A common procedure for transforming filamentous fungi involves the preparation of
protoplasts using a cocktail of wall macerating enzymes, introduction of the
transforming DNA, and selection of the emerging putative transformants. Initial systems
were based on complementation of auxotrophic mutations, very often limited to
transformation of fungi with well-developed genetic systems (Yelton et al., 1984). Since
the last decade, more versatile transformation methods have been developed using
vectors containing genes of bacterial origin conferring antibiotic resistance and fungal
promoters to enhance their expression in fungal systems. Upon integration, the vector
enables the resulting transformants to grow in the presence of the specific antibiotic,
which is otherwise toxic to the untransformed wild types. The most commonly used
selectable marker used in fungal transformation are genes conferring resistance to
hygromycin B, geneticin, benomyl and phleomycin (Hynes, 1996). A significant number
of fungi have been transformed using these system (Fincham, 1989).
The ability to transform filamentous fungi has progressed substantially in recent years
due to the development of innovative approaches such as restriction-enzyme-mediated-
integration (REMI) (Kahmann and Basse, 1999) and Agrobacterium tumefaciens-mediated
transformation (de Groot et al., 1998; Gouka et al., 1999; Chen et al., 2000; Rho et al.,
2001). The general mechanisms of ATMT and REMI transformation and the process of
gene disruption are illustrated in Figure 3 and Figure 4. REMI, a method for generating
non-homologous integration of transforming DNA into the chromosomes of eukaryotic
cells, has been used in insertional mutagenesis and isolation of pathogenicity genes in
plant pathogenic fungi (Riggle and Kumamoto, 1998; Thon et al., 2000). In REMI
transformation, a restriction enzyme is added to the transformation mix and by a process
not well understood, the enzyme enters the nucleus of the transformation recipient
fungus and digest the genomic DNA introducing double-stranded breaks at the
recognition sites (Figure 3).

- 44 -
These breaks are compatible with transforming plasmid DNA that has been linearised
with the same restriction enzyme. This often results into what is commonly known as
conservative integration, that is integration into the appropriate restriction site with
conservation of the site. (Riggle and Kumamoto, 1998). By definition, integration of a
plasmid DNA into a gene at a restriction site will cause a mutation, and that mutation
will be tagged by the integrated plasmid. The technique of REMI has been used
effectively in the genetic analysis and production of pathogenicity mutants in several
plant pathogenic fungi (Riggle and Kumamoto, 1998; Thon et al., 2000).
Plasmid
RBLB
T-DNA
Vir. Region
Donor bacterial cellAS
Target DNA
Restriction enzyme
Cell membrane
(A) ATMT (B) REMI
Nuclear membrane
Host cell wall
A1
A2
A3
A4
B1
B2
B3
Circular vector with selectable marker
Figure 3. A diagramatic representation of two forms of transformation , (A) ATMT and (B) REMI. In ATMT the process is initiated by inserting a plasmid with selectable marker gene situated between the left (LB) and the right (RB) border of the transforming DNA sequence into the donor bacterial cell. (A2) In the presence of acetosyringone (AS), it is believed that the bacterial cell adheres to the host cell wall. A single stranded T-DNA complex (T-DNA and Vir proteins) is then produced. The presence of left and right borders are essential for the process. (A3) A channel permits the transport of the complex through the host cell wall and membrane. (A4) The T-DNA crosses the nuclear membrane into the nucleus where it then randomly integrates itself into the host genome. In REMI, the selectable marker is inserted into a circular vector that is then linearised (B1) by a restriction enzyme. (B2) The linear vector is transported across the cell membrane of host protoplast and through the nuclear membrane in the presence of the same restriction enzyme. (B3) The host DNA is digested by a restriction enzyme, and the transforming DNA, compatible with the cut ends of the host genome, inserts itself into these sites. The figure is adapted from Mullins and Kang (2001).
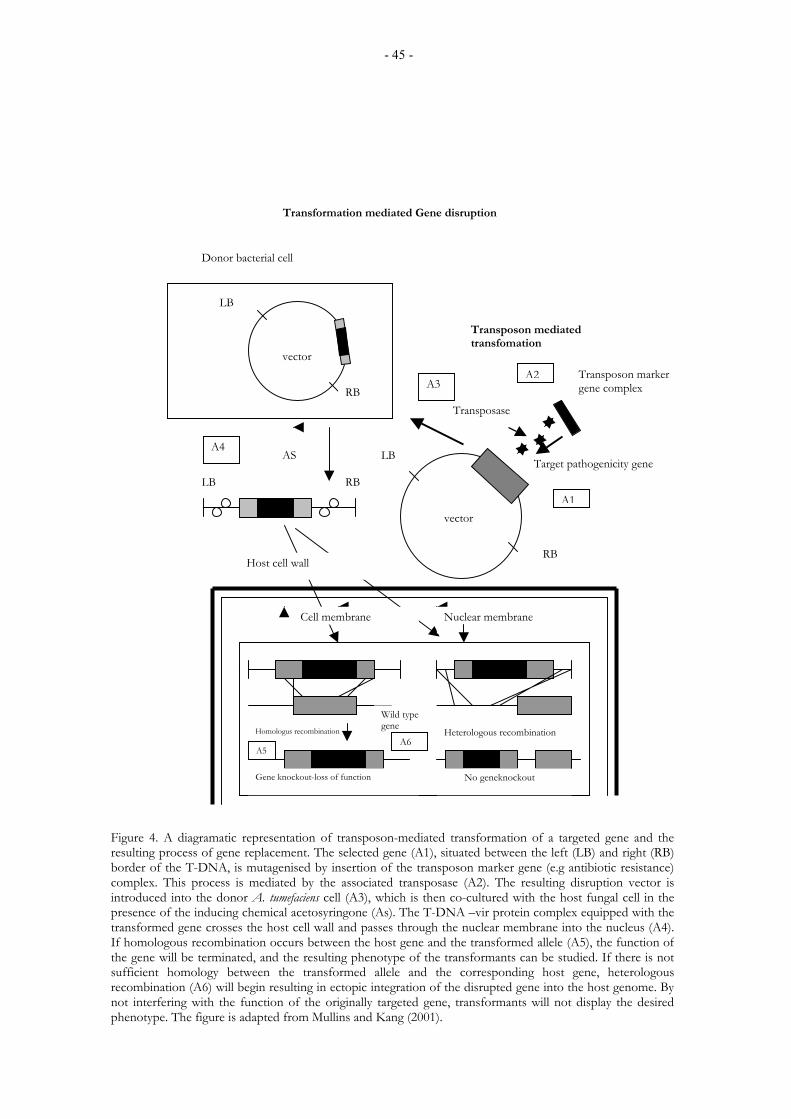
- 45 -
Transposase
A3
RB
LB
Transformation mediated Gene disruption
A1
A2
Target pathogenicity gene
Transposon marker gene complex
Transposon mediated transfomation
A4
RB LB
Wild type gene Homologus recombination Heterologous recombination
Gene knockout-loss of function No geneknockout
A5A6
Host cell wall
Cell membrane Nuclear membrane
Donor bacterial cell
LB
RB
AS
vector
vector
Figure 4. A diagramatic representation of transposon-mediated transformation of a targeted gene and theresulting process of gene replacement. The selected gene (A1), situated between the left (LB) and right (RB)border of the T-DNA, is mutagenised by insertion of the transposon marker gene (e.g antibiotic resistance)complex. This process is mediated by the associated transposase (A2). The resulting disruption vector isintroduced into the donor A. tumefaciens cell (A3), which is then co-cultured with the host fungal cell in thepresence of the inducing chemical acetosyringone (As). The T-DNA –vir protein complex equipped with thetransformed gene crosses the host cell wall and passes through the nuclear membrane into the nucleus (A4).If homologous recombination occurs between the host gene and the transformed allele (A5), the function ofthe gene will be terminated, and the resulting phenotype of the transformants can be studied. If there is notsufficient homology between the transformed allele and the corresponding host gene, heterologousrecombination (A6) will begin resulting in ectopic integration of the disrupted gene into the host genome. Bynot interfering with the function of the originally targeted gene, transformants will not display the desiredphenotype. The figure is adapted from Mullins and Kang (2001).

- 46 -
2. AIMS OF THE STUDY
Helminthosporium turcicum Pass (Exserohilum turcicum Leonard and Suggs) causes leaf blight
maize and inflicts significant yield losses in warm and humid maize growing regions of
the world (Adipala et al., 1993; Tilahun et al., 2001). The molecular genetics of H.
turcicum is poorly studied. The fungus, until this study, was not genetically transformed
and hence was not amenable to any classical or molecular genetic manipulations. There
are no previous efforts to characterise pathogenicity factors from this fungus. Therefore,
the molecular basis of maize-H. turcicum interactions were unknown. The primary wall of
maize contains up to 60% of xylan (Carpita, 1996; Cosgrove, 1997). The fungus
penetrates maize leaves directly through the cuticle and outer epidermal cell walls, and
only rarely penetration occurs through the stomata (Knox-Davies, 1974; Hilu and
Hooker, 1964). So the hypothesise of this study was that xylanase might be an important
pathogenicity or virulence factor for H. turcicum on maize. Thus, the isolation and
characterisation of the gene(s) encoding xylanases and the development of molecular
genetic tools (genetic transformation of the fungus) are necessary prerequisites for the
direct evaluation of the role of these putative pathogenicity factors in plant diseases.
Therefore, for the elucidation of the role of xylanases in H. turcicum-maize interactions,
this study was carried out with the following specific objectives.
1) To study the production and the purification properties of xylanase from the H.
turcicum.
2) To make a comparative study of the production of xylanase between the maize (grass)
pathogen, H. turcicum and the pea (dicot) pathogen, Ascochyta pisi.
3) To clone, characterise and study the expression patterns of xylanase gene(s) from H.
turcicum, both in vitro and during infection of maize by the fungus.
4) To develop an effective transformation procedure for H. turcicum in order to make the
fungus amenable to molecular genetic manipulations and facilitate molecular genetic
studies of its pathogenicity on maize.

- 47 -
3. MATERIALS AND METHODS
The materials used and details of the experimental procedures are described in the
original papers I-V. This section provides only the summary of the materials and
methods used in the study.
3.1. Fungal strains and culture conditions H. turcicum isolate (H-2) was isolated from diseased maize leaves obtained from the
Addis Ababa University, Awassa College of Agriculture, Ethiopia. The fungus was
maintained as conidial suspension in 20% glycerol at -20 ºC and routinely cultured on
potato dextrose agar (PDA) at room temperature. The isolate was periodically revitalised
and checked for pathogenicity in green house infections of maize plants. Ascochyta pisi
Lib. pathotype PF5 (E. Bollerup, Pajbjergfonden, Odder, Denmark) was maintained on
Coon's agar (Ali et al., 1978) at room temperature in alternating cycles of 8 hours under
near-UV light and 16 hours of darkness.
3.2. Bacterial strains and plasmids
Cloning and plasmid amplifications were done using the Escherichia coli strain DH5α
(Life Technologies Inc. Gaithersberg, MD, USA). The E. coli strain LE392 (Stratagene,
La Jolla, CA, USA) was used for the production of genomic library, which was
constructed using Lambda Dash II/BamHI cloning kit (Stratagene), and Lambda phage
infection for plague lifting (II, III, IV). All clonings were carried out in the plasmid
pBluescript II (SK+) (Stratagene). For the Agrobacterium tumefaciens mediated
transformation (ATMT) of H. turcicum, the A. tumefaciens strain AGL-1 carrying the
binary plasmid vector pBGgHg (Peter Romaine, Department of Plant Pathology,
Pennsylvania State University, USA) was used.
3.3. Production of xylanase
The production of xylanase by H. turcicum (I) and A. pisi (II) was studied by growing the
fungi in minimal salt medium (I) supplemented with the carbon sources, oatspelt xylan
(Sigma, MI, USA), birchwood xylan (Sigma), maize cell wall, glucose, and wheat straw (I,

- 48 -
III). The natural or noncommercial sources of xylan, wheat straw and maize cell walls
were prepared as described in (I). The cultures were incubated at room temperature on a
rotary shaker (150 rpm) at room temperature for 14 days. Samples of culture filtrates
were taken at 24 hours interval. Hyphal and other debris were collected by centrifugation
at 10000 rpm at 4 ºC and the resulting clarified culture filtrate was used for xylanase
activity assay either immediately or stored frozen at -70 ºC until analysed.
3.4. Xylanase activity assay (I, II)
Xylanase activity was assayed according to Poutanen and Puls (1988) with slight
modification. Xylan from oatspelt (1%, w/v) in 0.05 M sodium citrate buffer pH 5.3 was
used as a substrate for the assay. The samples (40 µl) from the culture filtrate were
incubated with 360 µl of the substrate solution in a water bath at 50 ºC for exactly 5
minutes. The reaction was terminated by adding 600 µl of dinitrosalicylic acid (DNS)
reagent (Miller, 1959). After heating for 5 min in a boiling water, cooling and
centrifugation, the absorbance of the supernatant was measured at 540 nm using a
Multiscan MCC/340 (Labsystems LTD) microtiterplate spectrophotometer. Xylose
(Merck, IL, USA) was used as a standard and xylanase activities were expressed in
nanokatals.
3.5. Purification of xylanase (I)
Clarified culture medium, stored at -70 ºC, was thawed and centrifuged at 20000 × g
(Sorvall SS-34 rotor). The clear supernatant was passed in 2.5 ml portions through a 1.0 x
8.0 cm Q-Sepharose Fast Flow (Pharmacia, England) column, equilibrated with 20 mM
sodium acetate, pH 4.8, using a flow rate of 1.0 ml min-1 and monitoring at 280 nm. The
column was washed between the runs with 2.0 ml of 1 M sodium chloride and
equilibrated to the buffer above. The xylanase activity containing fractions (about 4 ml)
from four runs were combined and diluted (1:2) with deionised water to reduce the buffer
ionic strength to 10 mM. Cation exchange chromatography was performed on a Mono S
HR5/5 (Pharmacia) column equilibrated with 10 mM sodium acetate, pH 4.8, using a
linear gradient of sodium chloride (0-0.5 M in 30 min) in the equilibration buffer and a
flow rate of 1.0 ml min-1. Xylanase active fractions from cation exchange chromatography
were combined, concentrated to about 1/4 of the original volume in a vacuum centrifuge

- 49 -
and subjected in 250 µl portions to gel filtration in a Superdex S-75 HR10/30 (Pharmacia)
column equilibrated with 20 mM sodium acetate, pH 4.8, 0.1 M sodium chloride.
Chromatography was performed with a flow rate of 0.5 ml min-1 and detection at 280 nm.
Xylanase active fractions were pooled and used for further studies. For the calculation of
specific activities during purification, the protein concentrations of different fractions
were determined using the Bio-Rad protein assay reagent (Bio-Rad, CA, USA) and γ-
globulin (Sigma) as standard. Gel electrophresis under denaturing conditions (12.5% SDS-
PAGE) was carried out according to Laemmli (1970).
3.6. Tryptic cleavage and peptide separation (I)
About 1 nmol of the purified xylanase from gel filtration (I) was concentrated to 100 µl
and passed through a 0.5 x 5 cm Bio-Gel P6 (Bio-Rad) column equilibrated with 10 mM
ammonium bicarbonate. The xylanase active flow through fraction was dried in a vacuum
centrifuge and dissolved in 20 µl of a solution containing 8 M urea and 0.4 M ammonium
bicarbonate. Dithiothreitol (2 µl of 112 mM) was added and the sample incubated at 50
°C for 30 min. To alkylate the cystein residues, 4-vinylpyridine (2 µl, diluted 1:50 with
water) was added and the sample incubated at room temperature for 15 min. Water (200
µl) and 1 µg trypsin (modified trypsin, sequencing grade, V5111 (Promega, Madison, WI,
USA) were added and the solution was incubated in the thermostated (37 °C) autosampler
compartment of an Applied Biosystems 270A-HT Capillary Electrophoresis System.
Digestion was monitored by injecting samples every 30 min followed by peptide
electrophoresis at 25 kV in a 50 µm fused silica (52 cm effective length) capillary in 20
mM sodium citrate buffer, pH 2.5. Electrophoresis was monitored at 200 nm. After
completion of the enzymatic cleavage, the reaction mixture was acidified with 1 µl
trifluoroacetic acid. Peptides were separated by narrow bore reversed phase HPLC
(Applied Biosystems 140B, 785A) on a 2.1 x 150 mm Rexchrom C8 (300Å, 5 µm, Regis
Chemical Company) column using a linear gradient of acetonitrile (3-60% in 60 min) in
0.1% trifluoroacetic acid and detection at 214 nm. Manually collected peptides were
concentrated to about 30 µl in a vacuum centrifuge.

- 50 -
3.7. Protein and peptide sequencing (I)
Protein and peptide sequencing was performed on a Gas-Pulsed-Liquid-Phase Sequencer
using on-line PTH-amino acid analysis (Kalkkinen and Tilgmann, 1988).
3.8. Construction of genomic library (II, III)
Genomic libraries of both A. pisi and H. turcicum were constructed in Lambda
DashII /BamHI cloning kit (Stratagene) following the manufacturer's instructions.
Genomic DNA from both fungi was partially digested with Sau3A and size fractionated
by agarose gel electrophoresis. Twelve to twenty kb fragments were isolated from the gel
and cloned into BamHI digested Lambda Dash II . Before use the genomic library was
amplified by making Lambda lysates.
3.9. DNA extractions (II, III, IV, V)
For DNA extraction A. pisi and H. turcicum were grown in potato dextrose broth (PDB)
and Coon’s medium, respectively. Genomic DNA was isolated using a modified protocol
where freshly harvested mycelia were ground in liquid nitrogen, resuspended in an
extraction buffer (150 mM EDTA pH 8.0, 50 mM Tris-HCl pH 8.0, 1% sodium lauryl
sarkosine, 300 µg/ml Proteinase K). The resulting suspension was extracted twice with
equal volume of phenol by centrifugation at 13000 rpm at room temperature. The
supernatant was again extracted once with chloroform:isoamyl alcohol (24:1) and once
with only chloroform. The DNA was double precipitated with half volume of ammonium
acetate (7.5 M) and two volumes of 99% ethanol. The DNA was dissolved in appropriate
amount of TE pH 8 (10 mM Tris-HCL pH 8.0, 1 mM EDTA pH 8.0), treated with
RNases and stored at 4 °C.
3.10. RNA extraction and first strand cDNA synthesis (II, III, IV)
Total RNA was isolated from both fungi and maize plants from approximately 1 g
mycelium or leaf material essentially as described in Chang et al. (1993) with slight
modifications. Fungal mycelia harvested from liquid cultures and/or leaves from maize
seedlings were frozen in liquid nitrogen immediately after harvest. The samples were used

- 51 -
for RNA extraction either immediately or stored at –80 °C until analysed. RNA
concentration and quality were determined spectrophotometrically by measuring A260 and
A280. RNA concentration was calculated according to A260. First strand cDNA synthesis
was carried out using either the Promega (III, IV) or GIBCO BRL (Copenhagen,
Denmark) (II) reverse transcription system where approximately 1 µg of DNA free
(treated with RNases free DNases) total RNA was reverse transcribed using Avian
Myeloblastosis Virus (ALV) reverse transcriptase or Moloney Maurine Leukemia (MLV)
reverse transcriptase for upto one hour at 42 °C.
3.11. Southern hybridisations (II, III, IV, V)
For the Southern hybridisation, approximately 3 µg of genomic DNA was digested with
appropriate restriction enzyme and size fractionated on a 0.7% agarose gel in either Tris
acetate (TAE) or Tris borate (TBE) buffer and capillary blotted to a positively charged
nylon membrane (Roche). Except where stated otherwise, all restriction and modifying
enzymes were from Promega. The membranes were hybridised at 68 °C for at least 18
hours using either radioactive labelled (II) or non-isotopic digoxigenin labelled (Roche)
cDNA or genomic probes (II, IV, V) in 8 × Denhardt solution (II) (0.5% SDS, 4 × SSC,
7% dextran-sulphate, and 1 mM EDTA) or in a modified Church and Gilbert solution
(Church and Gilbert, 1984) containing 0.5 M NaHPO4 and 7% SDS (w/v). After
hybridisation, the membranes were washed once with 5 × SEN (5% SDS, 40 mM
NaHPO4, pH 7.2, 1 mM EDTA pH 8.0) and twice with 1 × SEN (1% SDS, 40 mM
NaHPO4, pH 7.2, 1 mM EDTA, pH 8.0) at 65 °C. After post hybridisation washes and
sufficient blocking of the unspecific sites, the membranes were incubated with a dilution
of antidigoxigenin Fab fragment (Roche) that are conjugated to alkaline phosphatase. The
membranes carrying the hybridised probe and bound antibody conjugate were reacted
with the chemiluminescent substrate CDP star (Roche, Mannheim, Germany) and
exposed to Fuji RX X-ray film to visualise the chemiluminescent signal.
3.12. Northern analysis (II, III, IV)
For northern analysis or analysis of gene expression in vitro and in planta, 10 to 15 µg of
total RNA isolated from fungal mycelia and/or maize leaves was fractionated on

- 52 -
denaturing 1% formaldehyde agarose gels and capillary blotted onto a positively charged
nylon membrane (Roche). Membranes were hybridised with either radioactive labelled
probe (II) or non isotopic digoxigenin labelled probes (II, III, IV). Hybridisation
conditions, post hybridisation washes and detection were as described Southern
hybridisation. Equal loading in each lane was confirmed by ethidium bromide staining.
3.13. Isolation and characterisation of xylanase genes (II, III, IV)
PCR amplification of genomic and cDNA fragments of the xylanase gene fragments was
achieved using oligonucleotides (3475-5′-GGIIGGAA(C/T) CCIGGIAC (A/C/G/T)
GC- (A/C/G/T)(A/C) G-3′) and (3476- 5′-TTI(C/G)(A/T) IGCCCAIGC (A/G)
TC(A/G)AA(A/G)TG-3′) (II, III) designed from amino acid sequence of a xylanase from
H. turcicum (I), (XYL-F-5′-ACAACGGT(A/G)CT(A/T)CTACTC-3′) and (XYL-R-5′-
GGCAA(C/T)AGTCACCGTTC-3′) (IV) designed from multiple alignment of fungal
xylanase genes. The amplified gene products were cloned either into pBluescript II (SK+)
(Stratagene) or pGEMT easy vector system (Promega). Gene products were used as a
probe to screen the genomic library and to isolate the corresponding xylanase genes.
Genomic library screening (II, III, IV) was done by following the standard procedures
(Sambrook et al., 1989). From the genomic library, recombinant phages were plated and
replica membranes were made. Hybridisation, washes and detection were performed as in
Southern hybridisations (see section 3.10). Positive plagues were purified by three or four
rounds of plaque lifting and hybridisations. Lambda DNA from selected plaques were
isolated by the rapid procedure described in Sambrook et al. (1989) or purified on Qiagene
columns (Qiagene, Germany).
3.14. Sequence and phylogenetic analyses (II, IV)
The DNA sequencing was done using the BigDye terminating kit and analysed on an ABI
377XL automated DNA sequencer (Perkin-Elmer) (II, III), or ABI 3100 16 capillary
DNA sequencer (Applied Biosystems) (IV). The sequences obtained were assembled
using X-gap in the Staden Package program on a SUN workstation (Bonfield et al., 1995).
The phylogenetic positions of the cloned xylanase genes (II, IV) were defined using the
Clustal-W program (Thompson et al., 1994). In the case of A. pisi, bacterial and fungal

- 53 -
xylanase genes encoding family 10 and family 11 xylanases (II) and in H. turcicum
conserved regions of xylanase genes encoding family 11 bacterial and fungal xylanases
were used in the phylogenetic analysis. Phylogenetic trees were obtained using the
heuristic search option in PAUP 4.0 (Swofford, 2000) with random addition sequence (10
reps.) and tree–bisection reconnection (TBR) options.
3.15. Plant infection and reverse transcription–polymerase chain reaction (RT-PCR) (III, IV)
To study xylanase expression in infected plants, two-week old maize seedlings grown in
greenhouse were inoculated with conidial suspension of H. turcicum (~106 ml-1 ). Total
RNA was isolated from leaves collected at the different stages of infection and the from
the uninfected control plants. One microgram of total RNA (treated with RNases free
DNases) was reverse transcribed into cDNA using ALV reverse transcriptase (Promega,
Reverse Transcription System) with the reverse primer 3476. The RT reaction product
was used as a template for the PCR amplification with primers 3475 and 3476. The PCR
reaction conditions were as follows: one cycle of denaturation at 95 °C, 2 min of annealing
at 60 °C and 3 min of extension at 72 °C and 34 cycles of 30 sec denaturation at 94° C, 1
min of annealing at 60 °C and 2 min of extension at 72 °C. As a positive control, RT-PCR
was performed on total RNA isolated from mycelium grown for 5 days in a minimal salt
medium containing 1.5% birchwood xylan. Genomic DNA from H. turcicum was used as a
template in the PCR reaction to compare the size of amplified fragment with or without
the intron.
3.16. Agrobacterium tumefaciens-mediated transformation of H. turcicum (V)
The A. tumefaciens strain AGL-1 was grown in minimal medium (MM) (Hanif et al., 2002)
supplemented with kanamycin (100 µg/ml) for two days at 28 °C on a rotary shaker at
100 rpm. The A. tumefaciens cells were then diluted in an induction medium (IM) (Hanif
et al., 2002) in the presence of acetosyringone (200 µM), and were allowed to grow for
additional six hours at 28 °C with shaking at about 100 rpm. The induced cells were
mixed with equal volume of conidial suspension (∼ 105 ml -1). Uninduced cells (A.
tumefaciens culture without acetosyringone) served as a control transformation. The
mixture was incubated at room temperature for 48 hours shaking at about 75 rpm. The

- 54 -
200 µl aliquots of conidial and A. tumefaciens (co-cultivation) mixture were plated on
potato dextrose agar (PDA) in an A. tumefaciens induction medium (IM), which contained
hygromycin B (100 µg/ml) as a selection agent for transformation and cefotaxime (200
µM) or vancomycin (100 µg/ml) to kill A. tumefaciens cells. The plates were incubated at
room temperature and frequently checked for the emergence of putative transformants.
Individual transformants from each plate were counted and transferred to potato
dextrose agar (PDA) amended with hygromycin B, cefotaxime or vancomycin, and
incubated at room temperature until conidiation. Conidia from individual putative
transformants were resuspended in sterile distilled water and plated on potato dextrose
agar containing the appropriate antibiotics. Colonies of different sizes, large and small,
appeared on the plates. Culture from the large and small colonies were maintained as
monoconidial cultures and transferred to small PDA plates (50 mm diameter) containing
hygromycin B and cefotaxime for further selection. Conidia from these monoconidial
cultures of hygromycin B (100 µg/ml) resistant transformants were stored in 30%
glycerol at -80 °C until further analysis.
3.16.1. PCR screening of putative transformants (V)
The screening of putative transformants for the integration of hygromycin resistance
gene (hph) was carried out by PCR using genomic DNA from putative transformants
with the gene specific oligonucleotides hph-F (5′-AAGCCTGAACTCACCGCGACC-3′
and hph-R (5′-CTATTCCTTTGCCCTCGGAC-3′). The PCR reaction program used
was as described in section 3.15. DNA from the wild type strain (H-2) and the plasmid
(pBGgHg) served as negative and positive control, respectively.
3.16.2. Southern analysis of tranformants (V)
For the Southern analysis, approximately 3 µg of DNA from transformant strains and
untransformed wild type was digested with the restriction enzyme SacI (SacI does not cut
within the hph gene but cuts once within the T-DNA) and size fractionated by
electrophoresis on a 0.7% agarose gel in 1×TAE (Tris acetate) buffer. After depurination
(0.25 M HCl), denaturation (1.5 M NaCl and 0.5 M NaOH) and neutralisation (1.5 M
NaCl and 1 M Tris-HCl pH 7.4) steps, the gel was capillary blotted onto a positively

- 55 -
charged nylon membrane (Roche). Pre-hybridisation, hybridisation, post-hybridisation
washes and detection were carried out essentially as described in Southern hybridisations
(section 3.11).

- 56 -
4. RESULTS AND DISCUSSION 4.1. H. turcicum and A. pisi differ in their ability to produce xylanase activity in culture
The production of xylanase activity in time scale of the maize pathogen H. turcicum and
the pea pathogen A. pisi was studied on a minimal salt medium containing 1.5% (w/v)
birchwood xylan as a sole carbon source (I, II). The two fungi differed greatly in their
ability to produce xylanase activity under the test conditions (Figure 5). There was also
marked difference in the growth rate of the two fungi in the induction medium. H.
turcicum grew faster than A. pisi (data not shown) and accumulated also a substantial
amount of mycelial biomass. A number of studies shown that production of xylanase by
fungi differ depending on the species and the xylan growth substrate (Gomes et al.,
1992; Senior et al., 1989). Pure forms of xylan such as oatspelt and birchwood xylans are
less efficient inducers of xylanase activity in fungi compared to that of native sources of
xylan such as isolated plant cell walls and barley or wheat straw. Under all the test
conditions, H. turcicum was superior to A. pisi in production of xylanase activity (I, II).
Supplementing the induction medium with small amounts (< 0.5%) of readily utilisable
carbon source, such as sucrose or glucose, enhanced slightly the production xylanase in
A. pisi (II) but xylanase activity declined as concentration of glucose higher than 0.5%
was present in the medium and completely inhibited in the medium containing 1%
glucose. Holden and Walton (1992) reported similar finding from their xylanase
induction studies of the fungus C. carbonum. The small amount readily utilisable sugars
might support the initial growth of the fungus and accumulation of biomass so that the
fungus starts to hydrolyse the xylan substrate after the sugar is exhausted from the
medium as demonstrated by the delayed induction of xylanase activity in the sugar
supplemented medium. Studies in planta (Dori et al., 1995) also indicate that a fungal
inoculum grown in glucose enriched medium delay infection and symptom development
in wheat.
Pathogens of dicots and monocots appear to have different enzymatic adaptations to the
primary walls of their respective hosts. Cereal pathogens are adapted to production of
xylanase, not pectinases, as a reflection of their basic compatibility established with their

- 57 -
monocotyledonous hosts whose primary wall is composed predominantly of xylan and
very small amount of pectin (Braun and Rodrigues, 1993; Cooper et al., 1988). In
contrast, pectinases appears to predominate in pathogens of dicotyledonous hosts. Thus
the observed difference in the ability of production of xylanase activity between A. pisi, a
pathogen of dicots, and H. turcicum, a pathogen of only grasses, might be a manifestation
of their evolutionary adaptation to the primary cell walls of their respective hosts.
4.2. Purification properties of xylanase from H. turcicum (I)
H. turcicum produced high xylanase activity when it was grown in culture containing
maize cell wall as a sole carbon source (I). The specific activity of the purified enzyme
was comparable to the high-xylanase producing commercial fungi such as Trichoderma
spp. (R. Fagerström, personal communication). The xylanase enzyme was purified from
the culture filtrate by subsequent anion exchange chromatography, cation exchange
chromatography and gel filtration. The elution profile of the culture filtrate applied to a
Q-Sepharose column revealed only one peak in the flow-through and no additional
xylanase activity could be washed from the column with 1 M NaCl. The purified enzyme
gave a single band on sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-
PAGE) corresponding to an apparent molecular weight of 22.5 kDa (Figure 6). It was
determined to have a pI of 7.4; specific activity of 11300 nanokatals mg -1 of protein, pH
Figure 5. Production of xylanase by H. turicum and A. pisi on a minimal salt medium containing 1.5% birchwood xylan as a sole carbon source.
0
50
100
150
200
250
3 5 7 9 11 13 15
Days after inoculation
xyla
nas
e ac
tivi
ty (
nka
t /m
l) H.turcicumA. pisi
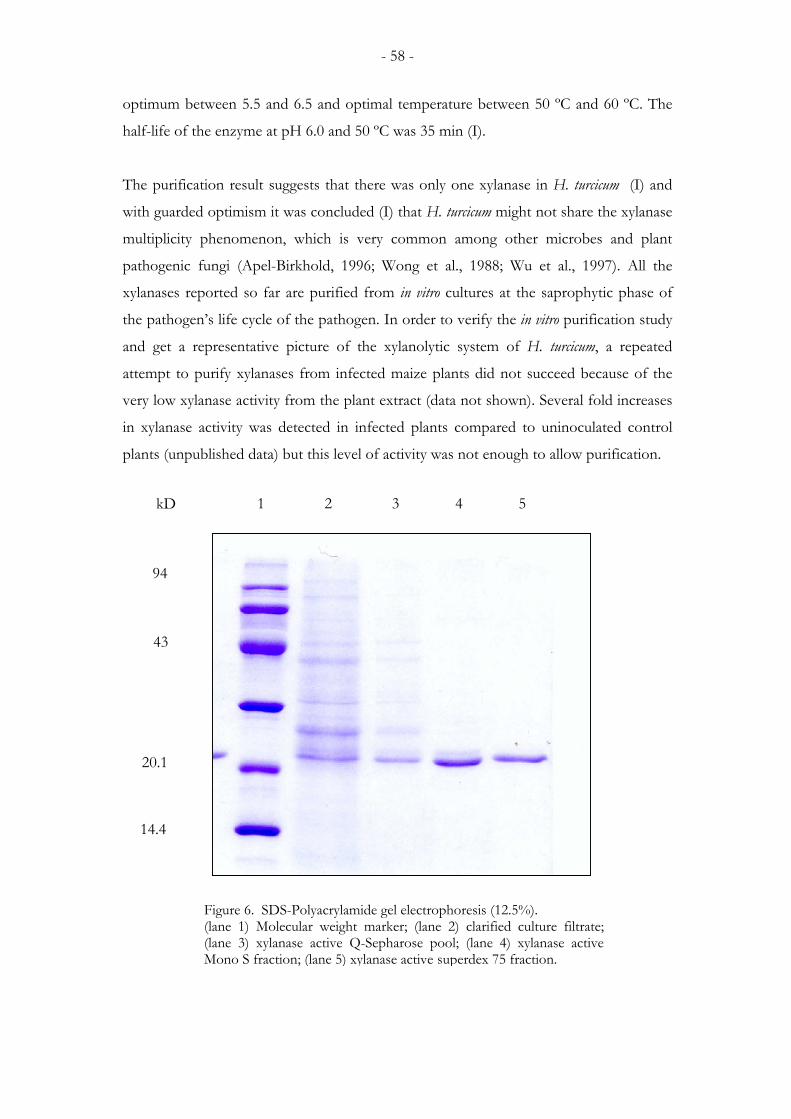
- 58 -
optimum between 5.5 and 6.5 and optimal temperature between 50 ºC and 60 ºC. The
half-life of the enzyme at pH 6.0 and 50 ºC was 35 min (I).
The purification result suggests that there was only one xylanase in H. turcicum (I) and
with guarded optimism it was concluded (I) that H. turcicum might not share the xylanase
multiplicity phenomenon, which is very common among other microbes and plant
pathogenic fungi (Apel-Birkhold, 1996; Wong et al., 1988; Wu et al., 1997). All the
xylanases reported so far are purified from in vitro cultures at the saprophytic phase of
the pathogen’s life cycle of the pathogen. In order to verify the in vitro purification study
and get a representative picture of the xylanolytic system of H. turcicum, a repeated
attempt to purify xylanases from infected maize plants did not succeed because of the
very low xylanase activity from the plant extract (data not shown). Several fold increases
in xylanase activity was detected in infected plants compared to uninoculated control
plants (unpublished data) but this level of activity was not enough to allow purification.
kD
94
43
20.1
14.4
1 2 3 4 5
Figure 6. SDS-Polyacrylamide gel electrophoresis (12.5%). (lane 1) Molecular weight marker; (lane 2) clarified culture filtrate; (lane 3) xylanase active Q-Sepharose pool; (lane 4) xylanase active Mono S fraction; (lane 5) xylanase active superdex 75 fraction.

- 59 -
It is very likely that the xylanases expressed in planta may not be stable in the buffering
systems of the purification scheme or might be lost due to degradation during the
process as suggested by Wong et al. (1988). Thus, it appears that the purification scheme
favours a xylanase, which is secreted abundantly in culture and may not reflect the
complete xylanolytic system of plant pathogenic fungi. The cloning of a second xylanase
gene (htxyl2) encoding a second xylanase in H. turcicum (IV) presents evidence to this
hypothesis. The xylanase encoded by htxyl2 was highly expressed during the infection of
maize by H. turcicum but was not expressed in culture when the fungus was grown in a
medium containing birchwood xylan as the only carbon source. This differential
expression pattern might have a biological significance related to phytopathogenicity
(Oliver and Osbourn, 1995; Walton, 1994). It is, therefore, important to note that
purification from culture filtrates, even by growing the fungus on medium containing
isolated cell walls with the assumption to induce xylanase with certain biological
functions (Walton, 1994), may not give the whole picture of the xylanolytic system of a
given pathogen.
4.3. Cloning of the xylanase genes
One xylanase gene from the pea pathogen A. pisi (xyl1) (II) and two xylanase genes (htxyl
and htxyl2) from H. turcicum (III, IV), a pathogen of maize were cloned and characterised,
which according to alignment studies with other fungal xylanase sequences (data not
shown) and phylogenetic analysis (Figure 3, in paper II, Figure 7, paper IV) were
identified as endo-1, 4-β-xylanases belonging to the family 11 glycosyl hydrolases
(Henerissat and Bairoch, 1993). The sequence data from the cDNA showed that the
xylanase gene from A. pisi has one intron (55 bp) whereas those of H. turcicum contain
two introns: 52 bp (intron I) and 62 bp (intron II) in htxyl1, and 55bp (intron I) and
52bp (intron II) in htxyl2. All the introns are with the typical intron signal sequence
(Consensus for filamentous fungi: G/GTANGT) (II). The occurrence of two or more
introns is a characteristic feature of the family 10 xylanases and it is less common among
family 11 xylanases. Except xyl4 and xyl 5 of F. oxysporum f.sp. lycopersici (Gómez-Gómez,
et al., 2001) xyl2 of C. carbonum (Apel-Birkhold and Walton, 1996) xyl2 of Cochliobolus
sativus (Emami and Hack, 2001), xyn11B of Bipolaris sorghi (Emami and Hack, 2002), and
htxyl1 and htxyl2 of H. turcicum, all the reported family 11 xylanase genes contain only
one intron. Interestingly, these xylanase sequences (36-51 in table 1) containing two

- 60 -
introns including the two H. turcicum xylanases were found from the same group in the
phylogenetic tree (Figure 7, paper IV).
4.4. htxyl1 and htxyl2 exhibit opposite patterns of expression under some test conditions
The result of enzyme purification (I) and cloning of the encoding gene htxyl1 (III)
suggests that there is only one xylanase in the H. turcicum genome. The purification
scheme resulted in only one peak (Figure 5) and Southern hybridisation, even at less
stringent conditions did not reveal any additional cross-hybridising band other than
htxyl1. However, the cloning of second xylanase gene (htxyl2) (IV) from the fungus,
which has more similarity to other fungal xylanases than to htxyl1, was by itself an
inspiration to look into the patterns of transcript accumulation of the genes under
different media and test conditions. Hence, expression of these two xylanase genes both
in vitro and in infected maize plants was conducted (IV).
The northern analysis of total RNA isolated from the H. turcicum grown in a minimal salt
medium (I) containing 1% birchwood xylan supplemented with additional 1% (w/v)
different carbon sources (glucose, sucrose and xylose) or various concentrations of
nitrogen sources (glutamic acid, ammonium nitrate and ammonium sulphate) were
performed. Among the carbon sources only xylose induced the expression of htxyl2. No
transcript of htxyl2 was detected in the basal inducing medium (minimal salt medium
plus 1% birchwood xylan). Unlike htxyl2 , both xylose and birchwood xylan supported
htxyl1 transcript accumulation. Glucose inhibited the expression of both genes (Figure 4,
paper IV). The lack of htxyl2 transcript in the presence of xylan is very interesting as
xylose is one of the breakdown products of xylan, which therefore and needs further
investigation.
Among the nitrogen sources, ammonium nitrate inhibited completely the expression of
htxyl1 and htxyl2, whereas the two genes differed in their patterns of transcript
accumulation with respect to glutamic acid and ammonium sulphate. The accumulation
of htxyl2 transcript increased with an increasing concentration of glutamic acid in the
medium, while concentrations of glutamic acid higher than 1 mM inhibited the
accumulation of the transcript of htxyl1. A more or less similar trend was also observed

- 61 -
with ammonium sulphate. No transcript of htxyl1 was detected at concentrations of
ammonium sulphate higher than 1 mM, but substantial amount of htxyl2 transcript was
detected at all concentrations with only a slight decrease towards higher molar
concentrations (IV)
Fungal pathogens respond to environmental cues and adjust their intracellular activities
in order to grow and survive in different environmental conditions and complete their
life cycle. Therefore, the occurrence of multiple forms of xylanases that are differentially
expressed could be part of the pathogen's strategy to respond to the molecular
environment at the host-pathogen interface. Plant-pathogen interaction is a complex
process where pathogen induced factors and plant factors interact in co-ordinated
manner. The plant-pathogen interface likely contains substances that influence enzyme
induction or repression (Holz and Knox-Davies, 1986). The substrate derivatives and
the end products of enzyme reactions may often play a key role in the induction of
xylanases, or they may also act as end product inhibitors, possibly at high concentrations.
Xylose, xylobiose, oligosaccharides of xylose and glucose are low molecular mass
fragments of xylan liberated by the action of small amount of constitutively produced
enzymes. These fragments play key role in xylanase biosynthesis (Kulkarni et al., 1999).
The opposite patterns of expression of htxyl1 and htxyl2 might have an important
biological significance in the life cycle of H. turcicum, and might be among the specific
adaptations of the fungus to exploit the host environment (IV).
The inhibition of htxyl1 and htxyl2 transcript accumulation by glucose, in the presence of
the inducing molecules (xylan or xylose), suggests that these two xylanases, like many
other fungal xylanases (Tonukari et al., 2002), are subject to glucose or catabolic
repression a regulatory mechanism well known in microbes (Ronne, 1995). Detailed
promoter analysis and expression studies need to be carried out to verify the result. In
yeast, the release from repression of glucose-repressed genes can be achieved by a
protein kinase, SFN1 (Ronne, 1995; Treitel et al., 1998). An ortholog of the snf1 gene
(ccsnf1) has been cloned recently from the phytopathogenic filamentous fungus, C.
carbonum (Tonukari et al., 2000) which was shown to be essential for the expression of
the glucose repressed xylanase and other cell wall degrading enzymes and for the
pathogenicity of C. carbonum. It is also speculated that mutation in snf1 gene may cause an
irreversible repression of glucose repressed genes, and hence, the approach could be

- 62 -
used for the analysis of the role of cell wall degrading enzymes in pathogenesis
(Tonukari et al., 2000).
Another mechanism that controls gene expression in fungi is the nitrogen metabolite
repression, a global regulatory mechanism that ensures the preferential utilisation of
simple nitrogen sources over complex sources (Marzluf, 1997). In filamentous fungi,
nitrogen utilisation pathways are activated by positive–acting global regulatory DNA
binding proteins of the GATA family encoded by areA and nit-2 in Aspergillus nidulans
and Neurospora crassa, respectively. The genes specify GATA type zinc-finger
transcription factors, which activate nitrogen structural genes when preferred nitrogen
sources are lacking (Marzluf, 1997). Although nitrogen regulatory mechanisms
controlling pathogenic growth of fungi are largely unknown, studies in several other
systems (Gimeno et al., 1992; Prade et al., 2001; Schulte and Bonas, 1992) implicate a
regulatory pathway involving starvation for preferred nitrogen sources. Lau and Hamer
(1996) identified two nitrogen-pathogenicity-regulation genes (NPR1 and NPR2), which
appear to be trans-acting positive effectors of genes in pathogenicity. Mutation in these
genes results in the inability to utilise a wide range of nitrogen sources and causes a
dramatic loss of pathogenicity.
The expression of htxyl1 and htxyl2 was completely inhibited at all concentrations of
ammonium nitrate (IV) but displayed opposite transcription patterns with respect to
ammonium sulphate and glutamic acid. No htxyl1 transcript was detected at
concentration of glutamic acid and ammonium sulphate higher than 1 mM. The htxyl2
transcript accumulation was highest at the highest concentration of glutamic acid
(100mM). Ammonium sulphate supported an accumulation of a substantial amount of
htxyl2 transcript with only slight decrease towards higher concentrations (IV). These
results are consistent with the expression patterns of a xylanase gene (xyl4) from
Fusarium oxysporum (Gómes-Gómes et al., 2002), particularly with respect to ammonium
sulphate in the case of htxyl2 and glutamic acid with htxyl1. The authors detected
increased xyl4 transcript at the concentrations of 25 mM ammonium nitrate.
Ammonium nitrate was inhibitory to both htxyl1 and htxyl2 even at the lowest
concentration (1 mM). Whether these responses to nitrogen supply is a manifestation of
the nitrogen regulatory circuit or not, has not yet been established and therefore needs
further investigation. However, results of this study indicate that htxyl2 expression

- 63 -
responded positively to the increasing concentrations of nitrogen supply That is its
expression was not dependent on nitrogen starvation. This is in contrast to the nitrogen
(nutrition)-linked pathogenicity gene expression model proposed by Lau and Hamer
(1996). However, the view that nitrogen starvation is a cue that controls fungal gene
expression during infection is challenged recently by Solomon et al. (2003) and might
even need to be reassessed.
The knowledge of factors regulating xylanase gene expression in phytopathogenic fungi
is very limited, mainly due to the fact that the composition of nutrients available in
plants and their effect on the xylanase expression is not known. However, fungal
xylanases that are expressed in infected plants (Apel-Birkhold and Walton, 1996;
Giesbert et al., 1998; Gómez-Gómez et al., 2002; and this study III, IV) have been
reported.
The differential accumulation of htxyl1 and htxyl2 transcripts in general, the abundance
of hxyl2 transcript of in infected maize plants in particular are the most interesting
observations in my thesis. The htxyl2 transcript was readily detected by northern
hybridisation both at early and late stages infection stages of maize by H. turcicum (IV). In
turn, htxyl1 transcript, which accumulated abundantly in vitro, was only detectable by the
very sensitive method of RT-PCR (III) but not by northern hybridisation (IV).
Identification of such genes differentially transcribed in a way pertinent to pathogenicity
is believed to be a better approach for a molecular analysis of phytopathogenicity (Oliver
and Osbourn, 1995; Walton, 1994) than those that are expressed only in vitro.
4.5. The phylogenetic positions of htxyl1 and htxyl2
The sequences from A. pisi xylanase and the two xylanase genes from H. turcicum
together with 48 bacterial and fungal xylanases obtained from the Genbank (Table 1,
paper IV) were used in the phylogenetic analysis. Accordingly, the three xylanase genes
encode xylanases which belong to the family 11 glycosyl hydrolases. The two xylanase
genes from H. turcicum grouped into separate clades. Studies with two xylanase genes
(xyn11A and Xyn11B) from several Cochliobolus and Bipolaris species (Emami and Hack,
2002) indicated that the phylogenies of both xylanase groups were consistent with the
phylogeny of the glyceraldehyde phosphate dehydrogenase (GPD) and internal

- 64 -
transcribed spacers (ITS) sequences, indicating that the xylanase genes reflected the
phylogenetic relationships of the studied taxa. Based on these results, the authors Emami
and Hack (2002) suggested that both xylanases were already present in the common
ancestor of all Cochliobolus species. Our results further indicate that both xylanases were
present in the common ancestor of Helminthosporium and Cochliobolus, since the H. turcicum
xylanases formed a sister group with the Cochliobolus sequences in both groups (Figure 7,
paper IV). However, while the xylanase genes may reflect the phylogenetic relationships
of some closely related taxa, they can not be used for studying the relationships of
distantly related organisms due to several gene duplications and possible horizontal gene
transfer events (Syvanen, 1994).
4.6. Agrobacterium tumefaciens-mediated transformation of H. turcicum
An effective way to study the role of putative pathogenicity factors of fungal pathogens
is to disrupt these genes, in both a targeted and random manner. Although, a number of
techniques have been employed for such analysis, those based on transformation are by
far the most commonly used (Gold et al., 2001). In filamentous fungi, the introduction
of DNA by transformation typically results in either the heterologous or homologous
integration of the transformed DNA. Homologous integration permits a targeted gene
disruption by replacing the wild type allele in the genome with a mutant allele.
H. turcicum was transformed (V) using the binary plasmid vector containing the
hygromycin B phosphotrnasferase (hph) gene controlled by gpd promoter from Agaricus
bisporus and CaMV 35 terminator in Agrobacterium tumefaciens strain AGL-1. The A.
tumefaciens-mediated transformation (ATMT) yielded stable transformants capable of
growing on increased concentrations of hygromycin B. The integration of hph gene in
the transformants was first confirmed by PCR (Figure 3, Paper V) and evidence of DNA
integration random sites in the genome was demonstrated by Southern blot analysis
(Figure 4, Paper V)
A. tumefaciens has been found to be capable of transferring its Ti-plasmid known as T-
DNA to other organisms besides its natural host (plants). A wide range of organisms
such as yeast (Bundock et al., 1995), filamentous fungi including phytopathogens (Gouka
et al., 1999; de Groot et al., 1998; Malonek and Meinhardt, 2001; Mullins et al., 2001;

- 65 -
Rho et al., 2001; Covert et al., 2001), mushrooms (Chen et al., 2000; Mikosch et al.,
2001) and even human cells (Kunik et al., 2001) have been transformed via ATMT. The
ATMT allows the use protoplasts, hyphae, spores, conidia, different types of human cells
and even blocks of mushroom mycelial tissue. Due to the versatility of the system,
conidia from the fungus were used for transformation (V). The success in transforming
H. turcicum, a fungus which otherwise proved difficult to transform with other methods,
is an additional confirmation of the efficiency of the ATMT. The transformation
efficiency observed (30-50 putative transformants per ~104 conidia) is comparable to the
efficiency reported from other fungi (Covert et al., 2001). The choice of promoter, the
strain of A. tumefaciens and the species of the recipient fungus influence the efficiency of
ATMT (Chen et al., 2000; Hanif et al., 2002; Mullins et al., 2001). H. turcicum is a readily
conidiating fungus both under natural and laboratory conditions, and furthermore,
conidial transformation is simpler and cheaper than protoplast transformation.
Therefore, with the rate of transformation frequency obtained and the reproducibility of
the system, the ATMT would allow the generation of a large library of H. turcicum
transformants with lower cost than the laborious and/or less efficient protoplast based
methods. ATMT is a promising transformation and gene tagging system (Gold et al.,
2001), which offers increased opportunities for genetic manipulations of diverse fungal
species.

- 66 -
5. CONCLUSIONS AND PROSPECTS
Fungi are the most important plant pathogens and continue to be the focus of extensive
research with a wide range of methodologies. Improving methods allow the dissection of
fungal pathogenicity determinants in a gene-by-gene approach. Identification and
analysis of genes differentially transcribed in a way pertinent to pathogenicity continue to
be a frequent research approach. The development of molecular genetic systems and the
availability of cloned genes provide the only practical way of approaching the problem of
plant-pathogen interactions at molecular level. H. turcicum, the cause of northern leaf
blight of maize, is among the fungi with poorly known genetic system. Before this study,
no transformation system was developed for the pathogen and hence it was not
amenable to any molecular genetic manipulations. No previous attempt was made to
clone and characterise potential pathogenicity genes.
My thesis presents significant advances in the development of the tools for molecular
genetics analysis of particularly H. turcicum, and laid down a useful foundational basis for
the molecular dissection of its pathogenicity. Three different xylanase genes, two from
H. turcicum and one from A. pisi were cloned and further characterised for the first time.
The two xylanase genes from H. turcicum reveal features of fungal xylanolytic systems
interesting for further research. For example, the transcripts of both genes accumulate in
infected maize plants but differed in transcript abundance. The meaning of this
differential transcription is not known and therefore needs further investigations. To my
knowledge, these genes are among the very few xylanase genes found associated with
infection. By definition a prerequisite for a pathogenicity factor is that it is expressed
during penetration and invasion of the host by the pathogen. Hence, htxyl1 and htxyl2
transcript accumulation during infection of maize suggests that xylanase is involved in
the pathogenicity of H. turcicum on maize. The disruption of the cloned genes either
singly or in combination and further analysing the resulting mutants would be very
interesting and would be helpful to further our knowledge about the xylanolytic systems
of fungal pathogens and the role of xylanases in pathogenesis.
Moreover, it is worth noting that within the emerging common phenomenon of xylanase
multiplicity, the clarification of the systems regulating expression of these enzymes is

- 67 -
important and should contribute considerably to devise a meaningful approach towards
evaluation of their role in disease. Further work along this line would be helpful.
The reproducible and conidial based ATMT developed for the fungus opens more
possibilities for the molecular manipulation of the fungus. Although the efficiency of
transformation found in this study is at experimentally useful rate, further optimisation
of the transformation procedure and analysis of larger number of transformants to verify
the plasmid copy number. Further more, experiments concerning the application of the
method, for example, insertional inactivation of a gene and identification of flanking
sequences that demonstrate its potential use in identifying pathogenicity genes are worth
performing.
It has been demonstrated that specific regulatory networks are required for fungal
pathogenesis. An ortholog of the yeast SFN1 gene encoding a protein kinase, which
releases catabolically repressed genes has been found essential for the expression of
xylanases and other glucose repressed cell wall degrading enzymes and virulence of C.
carbonum on maize. The nitrogen pathogenicity regulation genes (NPR1 and NPR 2)
from M. grisea are some examples of a link between metabolic gene regulation and
pathogenicity in fungi. Studies with signal transduction cascades hint that signal
transduction pathways that govern pathogenic growth in fungi are conserved. Linking
these regulatory and signalling pathways to the regulation of expression of cell wall
degrading enzymes and resolving their role in fungal pathogenicity is the real challenge
facing plant pathologists.
The recent discovery of xylanase inhibiting proteins (XIPs) is an indication of a
mechanism involved in plant counter-defence against the action of xylanases. This new
area of research might prove important as a strategy of fungal disease control.
In addition to xylanase genes, I have cloned other genes such as avenacinase and
polygalacturonases, from H. turcicum and these genes will be independently evaluated for
their role in pathogenicity. Taken together, this study is a significant contribution to H.
turcicum-maize pathosystem research and several steps forward in the molecular biology
of H. turcicum. The demonstration of molecular basis of H. turcicum pathogenicity on
maize is on the crossroads and exciting times lie ahead.

- 68 -
6. ACKNOWLEDGEMENTS
The Finnish International Development Agency (FINNIDA) and the Department of
Applied Biology are acknowledged for financial support.
The journey to this point has been a marathon, not a sprint. First of all I thank God for
His amazing grace, love and encouragement. I am also very grateful to many people,
who in many different ways, have supported and encouraged me through all these years.
I sincerely thank Professor (emer.) Eeva Tapio for giving me good background
knowledge in Plant Pathology and for providing me the opportunity for an advanced
learning and research in the field. I thank you for the concern and kindness you have
shown to my family and myself. I am deeply grateful to Professor (emer.) Katri Bremer
for her contribution to my education and for the good and fruitful discussion. I thank
Professor (emer.) Peter Tigerstedt for organising funding for part of my study.
I sincerely thank Professor Kim von Weissenberg for the great interest he has shown in
my studies and for creating a room for me to work in his research group. I thank you for
your support and encouragement through all the years. I greatly appreciate Professor
Annele Hatakka and Dr. Christina Dixelius for the excellent and expert reviewing work.
I thank them for their comments and suggestions to improve my thesis.
I deeply thank my supervisors; Professor Jari Valkonen for his valuable advice and
encouragement especially during the finalising of this thesis and Dr. Ari Pappinen for
creating a good working atmosphere, arranging funding, and valuable advice. I thank you
for your good will, positive attitude and interest in my work.
I wish to express my deepest gratitude to Dr. Nisse Kalkkinen for his important
contribution to the success of my work. Your help with purification of the first xylanase
and protein sequencing has been a great input indeed. I thank you for the kindness and
good will you have shown me. I also thank you for introducing me to Lars Paulin.
Words are not enough to thank Lars Paulin. My heart felt thanks to you Lars. Without
your help and determination, it would have been difficult to come to this stage. We faced
enormous challenges along the way but you never gave up. You always saw things
beyond the problems and in all cases we achieved the goal. I thank you for the excellent

- 69 -
sequencing work and for the valuable ideas and discussion. I thank all the co-authors of
the articles for their credible scientific contributions for the thesis. I thank Dr. Jorma
Vahala for carefully and critically reading the thesis manuscript and for his valuable
comments and suggestions including the language. I am very grateful to Dr. Peter
Stephensen Lübeck and Dr. Mette Lübeck for the fruitful collaboration we have had, for
teaching me the techniques of molecular biology and for their help in construction of
fungal genomic library. You are friends indeed. I also wish to thank Dr. Dan Funck
Jensen for inviting me to work for two months, as a visiting scientist, in his laboratory at
the Department of Plant Biology Royal Veterinary and Agricultural University,
Copenhagen, Denmark. I am very grateful to Mrs. Tuula Laine for well-organised and
excellent technical assistance and for her consistent friendly expressions. Thank you
Tuula for teaching me Finnish language. I deeply thank Mrs. Tuula Öhman for her help
in all administrative matters and accurate record keeping.
My heartfelt thanks to all my former and present colleagues and friends at the
Department of Applied Biology for their friendship and excellent working atmosphere.
My special thanks to Dr. Jorma Vahala, Dr. Ari Hietala, Kari Kammiovirta, Dr. Nicola
Laporta, Dr. Hanna Pasonen, Sanna-Kaisa Seppänen, Anna-Maija Niskanen, Jouko
Närhi, Hilkka Koponen, Minna Terho, Dr. Marja Poteri, Leena Syrjälä, Terttu Parkkari,
Marjo Kilpinen, Jinrong Lu, Gunnar Salingre, Dr. Tuula Mäki-Valkama, Dr. Anna-Liisa
Fabritius, Dr. Elina Roine, Dr. Katileena Lohtander, Lahja Pesonen, Sanna Koutaniemi,
Ursula Malm, Leena Tulisalo and Javier Brumos. I feel very privileged in having you as
my friends and colleagues.
Finally, my heartfelt and most genuine thanks go to my wife Nigist and my daughters
Bethel and Tsion to whom I dedicate this work. My wife Nigist has given me
tremendous support, love and understanding throughout the years. Bethel and Tsion,
every time you visited the lab you asked me what I am doing and why I am doing this
and that. That always made me think a lot to find a simple and clear way to explain.
Thank you for your help. My sincere gratitude to my mother, brothers and sisters back
in Ethiopia.
Helsinki, August 2003

- 70 -
7. REFERENCES
Adipala, E., Lipps, P.E., and Madden, L.V. (1993) Reaction of maize cultivars from Uganda to Exserohilum turcicum. Phytopathology 83:217-223
Agrios, G.N. (1997) Plant Pathology. Fourth edition. Academic Press, New York, London.
Albersheim, P., and Anderson, A. (1971) Proteins from plant cell walls inhibit polygalacturonases secreted by plant pathogens. Proceedings of the National Academy of Sciences, USA 68: 1815-1819
Albersheim, P., and Dravill, A.G. (1985) Oligosaccharins. Scientific American 253: 44-50 Ali, S.M., Nitscke, L.F., Dube, A.J., Krause, M.R., and Cameron, B. (1978) Selection for pea lines for resistance to pathotypes of Ascochyta pisi and Phoma medicaginis var pinodella. Australian Journal of Agricultural Research 29: 841-849
Anderson, A.J. (1978) Extracellular enzymes produced by Colletotrichum lindemuthianum and Helminthosporium maydis during growth on isolated bean and corn cell walls. Phytopathology 68: 1585-1589
Annis, S.L., and Goodwin, P.H. (1997) Recent advances in the molecular genetics of plant cell wall–degrading enzymes produced by plant pathogenic fungi. European Journal of Plant Pathology 103: 1-14
Apel-Birkhold, P.C., and Walton, J.D. (1996) Cloning, disruption and expression of endo-β, 1,4-xylanase genes, XYL2 and XYL3 from the maize pathogen Cochliobolus carbonum. Applied and Environmental Microbiology 62: 4129-4135
Apel, P.C., Pannacione, D.G., Holden, F.R., and Walton, J. D. (1993) Cloning andtargeted disruption of XYL1, a β-1,4-xylanase gene from the maize pathogen Cochliobolus carbonum. Molecular Plant-Microbe Interactions 6: 467-473 Atkinson, M.M. (1993) Molecular mechanisms of a pathogen recognition by plants. In J.H Andrews and J.C Tommercup (ed). Advances in Plant Pathology Vol. 10. Academic Press. London, pp 35-64 Avni, A., Bailey, B.A., Mattoo, A.K., and Anderson, J.D. (1994) Induction of ethylene biosynthesis in Nicotiana tabacum by Trichoderma viride xylanase is correlated to the accumulation of 1-Aminocyclopropane-1-carboxylic acid (ACC) synthase and ACC oxidase transcripts. Plant Physiology 106: 1049-1055 Bailey, B.A., Dean, J.F.D., and Anderson J.D. (1990) An ethylene biosynthesis-inducing endoxylanase elicits electrolyte leakage and necrosis in Nicotiana tabacum cv xanthi leaves. Plant Physiology 94: 1849-1854

- 71 -
Bailey, B.A., Korcak, R.F., and Anderson, J.D. (1992) Alteration in Nicotiana tabacumL. cv xanthi cell membrane function following treatment with an ethylene biosynthesis-inducing endoxylanase. Plant Physiology 100: 749-755 Bastawade, K.B (1992) Xylan structure, microbial xylanases, and their mode of action. World Journal of Microbiology and Biotechnology 8: 353-368 Balandran-Quintana, R.R., Wilson-Mendoza, A, M., Manilla-Alvarez,G.,Bateman, D.F., and Basham, H.G. (1976) Degradation of plant cell walls and membranes by microbial enzymes In: Heitefuss, R., and Williams, P.H (ed.), Encyclopaedia of Plant Physiology. Springer-Verlag, New York, pp 316-355 Beg, Q.K., Kapoor, M., Mahajan, L., and Hoondal, G.S. (2001) Microbial xylanases and their industrial applications. Applied Microbiology and Biotechnology 56:326-338 Bergmann, C.W., Vargas, A., and Martinez-Tellez, M.A. (2002) Effect of pectic oligomers on physiological responses of chilling injury in discs excised from Zucchini (Cuccurbita pepo L). Biochemical and Biophysical Research Communications 299: 577-584 Biely, P. (1985) Microbial xylanolytic systems. Trends in Biotechnology 3: 286-290 Blein, J.P., Milat, M-L., and Ricci, P. (1991) Response of cultured tobacco cells tocryptogenin, a proteinaceous elicitor from Phytophthora cryptogea: possible plasmalemma involvement. Plant Physiology 95: 486-491 Bonfield, J.K, Smith; K.F., and Staden, R. (1995) A new DNA sequence assembly program. Nucleic Acid Research 23: 4992-4999 Bowen, K.L., and Pedersen, W.L. (1988) Effects of northern leaf blight and detasseling on yields and yield components of corn inbreds. Plant Diseases 72: 952-956 Bradley, D. J., Kjellborn, P., and Lamb, C.J. (1992) Elicitor and wound-induced oxidative cross-linking of a proline-rich plant cell wall protein: a novel defence response. Cell 70: 21-30 Braun, E.J., and Rodrigues, C.A. (1993) Purification properties of an endoxylanase from a corn stalk rot of Erwinia chrysanthemi. Phytopathology 83: 332-338 Brett, C., and Waldron, K. (1996) Physiology and biochemistry of plant cell walls, second edition. Champman and Hall, New York. Brisson, I.F., Tenhaken, R., and Lamb, C. (1994) Function of oxidative cross-linking of cell wall structural proteins in plant disease resistance. Plant Cell 6: 1703-1712 Bucheli, P., Doares; S.H., Albersheim, P., and Darvill, A. (1990) Host-pathogen Interactions XXXVI. Partial characterisation of heat-labile molecules secreted by the rice blast pathogen that solubilise plant cell wall fragments that kill plant cells. Physiological and Molecular Plant Pathology 36: 159-173

- 72 -
Bugbee, W.M. (1993) Pectin lyase inhibitor protein from cells of sugar beet Phytopathology 83: 63-68 Bundock, P., Den Dulk-Ras, A., Beijersbergen, A., and Hooykas, P.J.J. (1995) Trans kingdom T-DNA transfer from Agrobacterium tumefaciens to Saccharomyces cerevisiae. EMBO Journal 14: 3206-3214 Camardella , L., Carratore, V., Ciardiello, M.A., Servillo, Luigui, Balstrieri, C., and Giovane, A. (2000) Kiwi protein inhibitor of pectin methylesterase: amino acid sequence structure and importance of two disulphide bridges. European Journal of Biochemistry 267: 4561-4565 Carpita, N.C. (1983) Hemicellulose polymers of the cell walls of Zea coleoptiles. Plant Physiology 75: 515-521 Carpita, N.C. (1989) Pectic polysaccharides of maize coleoptiles and porsso millet cells in liquid culture. Phytochemistry 28: 121-125 Carpita, N.C., and Gibeaut, D. M. (1993) Structural models of primary cell walls in flowering plants; consistency of molecular structure with the physical properties of the walls during growth. The Plant Journal 3: 1-30 Carpita, N.C. (1996) The structure and biogenesis of the cell walls of grasses. Annual Review of Plant Physiology and Plant Molecular Biology 47: 445-476 Cassab, G. I. (1998) Plant cell wall proteins. Annual Review of Plant Physiology and Plant Molecular Biology 49: 281-309 Ceballos, H., Deutsch, J.A., and Gutiérrez, H. (1991) Recurrent selection for resistance to Exserohilum turcicum in eight subtropical maize populations. Crop Science 31: 964-971 Cervone, F., Hahn, M.G., De Lorenzo, G., Darvil, A., and Albersheim P. (1989) Host-pathogen Interactions XXXIII. A plant protein converts a fungal pathogenicity factor into an elicitor of plant defence responses. Plant Physiology 90: 542-548 Chang, S., Puryear, J., and Cairney, J. (1993) A simple and efficient method for isolating RNA from pine trees. Plant Molecular Biology Reporter 11: 113-116 Chen, X., Stone, M., Schalgnhaufer, C., and Romaine, C.P. (2000). A fruiting body tissue method for efficient transformation of Agaricus bisporus. Applied and Environmental Microbiology 66: 4510-4513 Cheung, A.Y. (1995). Pollen-pistil interactions in compatible pollination. Proceedings of the National Academy of Sciences, USA 92: 3077-3080 Church, G.M., and Gilbert, W. (1984) Genomic sequencing. Proceedings of the National Academy of Sciences, USA 81: 1991-1995 Cook, B.J., Clay, R.P., Bergmann, C.W., Albersheim, P., and Darvill, A.G. (1999) Fungal polygalacturonases exhibit different substrate degradation patterns and differ in

- 73 -
their susceptibilities to polygalacturonase inhibiting proteins. Molecular Plant-Microbe Interactions 12: 703-711 Cooper, R.M., Longman, D., Campell, A., Henry, M., and Lee, P.E. (1988) Enzymic adaptation of cereal pathogens to the monocotyledonous primary walls. Physiological and Molecular Plant Pathology 32: 33-47 Cosgrove, D.J. (1997) Assembly and enlargement of the primary cell wall in plants. Annual Review of Cellular and Developmental Biology 13.171-201 Côte,F., and Hahn, M.G. (1994) Oligosaccharins: structure and signal transduction. Plant Molecular Biology 26: 1397-1411 Covert, S.F., Kapoor, P., Lee, M-H., Briley, A., and Nairn, C.J. (2001) Agrobacteriumtumefaciens-mediated transformation of Fusarium circinatum. Mycological Research 105: 259-264 Darvill, A.G., Augur, C., Bergmann, C., Carlson, R.W., Cheong, J.J., Eberhard, S.,Hahn, M.G., L.O. V.M., Marfa, V., Meyer, B., Mohnen, D., Ó Neill, M.A., Spiro, M.D., van Helbeek, H., York, W.S., and Albersheim, P. (1992) Oligosaccharins-oligosaccharides that regulate growth, development and defence responses in plants. Glycobiology 2: 181-198 Dean, J.F.D., and Anderson, J.D. (1991) Ethylene biosynthesis inducing xylanase. II. purification and physical characterisation of the enzyme produced by Trichoderma viride. Plant Physiology 95: 316-323 Dean, J.F.D., Gamble, H.R., and Anderson, J.D. (1989) The ethylene biosynthesis-inducing xylanase: its induction in Trichoderma viride and certain plant pathogens. Phytopathology 79: 1071-1078 Debyster, W., Peumans, W.J., Van Dammel, E.J.M., and Delcour, J.A. (1999) Triticum aestivum xylanase inhibitor (TAXI), a new class of enzyme inhibitor affecting breadmaking performance. Journal of Cereal Science 30: 39-43 Degefu, Y. (1990) Evaluation of Ethiopian maize germplasms for resistance against leaf blight caused by Helminthosporium turcicum. Research Progress Report, Awassa College of Agriculture, Addis Ababa University De Graaff, L.H., van den Broek, C., van Ooijien, A.J.J., and Visser, J. (1994) Regulation of the xylanase–encoding xylnA gene of Aspergillus tubingensis. Molecular Microbiology 12: 479-490 De Groot, M.J.A., Bundock, P., Hooykaas P.J.J., and Beijersbergen, A.G.M. (1998) Agrobacterium tumefaciens-mediated transformation of filamentous fungi. Nature Biotechnology 16: 839-842 Dekker, R.F.H., and Richards, G.N. (1975) Purification properties and mode of action of hemicellulase II produced by Ceratocystis paradoxa. Carbohydrate Research 42: 102-123

- 74 -
De Lorenzo, G., D’Ovidio, R., and Cervone, F. (2001) The role of polygalacturonase-inhibiting proteins (PGIPs) in defence against pathogenic fungi. Annual Review of Phytopathology 39: 313-335 Desiderio, A., Aracri, B., Leckie, F., Mattel, B., Salvi, G., Tigelaar, H., vanRoekel, J.S.C., Baulcombe, D,C., Melchers, L.S., De Lorenzo, G., and Cervone, F. (1997) Polygalacturonase-inhibiting proteins (PGIPs) with different specificities are expressed in Phaseolus vulgaris. Molecular Plant-Microbe Interactions 10: 852-860 De Vries, R.P., Visser, J., and de Graaf, L.H. (1999) CreA modulates the XlnR-induced expression on xylose of Aspergillus niger genes involved in xylan degradation. Research in Microbiology 150: 281-285 De Vries, R.P., Kester, H.C.M., Poulsen, C.H., Benen, G.A.E., and Visser, J. (2000) Synergy between enzymes from Aspergillus involved in the degradation of plant cell wall polysaccharides. Carbohydrate Research 327: 401-410 De Vries, R.P., and Visser, J. (2001) Aspergillus enzymes involved in degradation of plant cell wall polysaccharides. Microbiology and Molecular Biology Reviews 65: 497-522 DeWit, P.J.G.M., Hofman, A.E., Velthuis, G.C.M., and Kuc, J.A. (1986) Isolation and characterisation of an elicitor of necrosis isolated from intercellular fluids of compatible interactions of Cladosporium fulvum (syn Fulvia fulva) and tomato. Plant Physiology 77: 642-647 Dickison, W.C. (2000) Integrative plant anatomy. Academic press. San Diego, San Francisco, New York, Boston, London, Toronto, Sydney Tokyo Dori, S., Solel, Z., and Barash, I. (1995) Cell wall-degrading enzymes produced by Gaeumannomyces graminis var. tritici in vitro and in vivo. Physiological and Molecular Plant Pathology 46: 189-198 Dowzer, C.E.A., and Kelly, J.M. (1991) Analysis of the CreA gene, a regulator of carbon catabolite repression in Aspergillus nidulans. Molecular and Cellular Biology 11: 5701-5709 Ducros,V., Charnock, S.J., Derewenda, U., Derevwenda, Z.S., Dauter, Z., Dupont, C., Shareck, F., Morosoli, R., Kluepfel, D., and Davies, G.J. (2000)Substrate specificity in glycoside hydrolase family 10: Structure and kinetic analysis of the Streptomyces lividans xylanase 10A. The Journal of Biological Chemistry 275: 23020-23026 Dumville, J.C., and Fry, S.C. (2000) Uronic acid-containing oligosaccharins: their biosynthesis, degradation and signalling roles in non-diseased plant tissues. Plant Physiology and Biochemistry 38: 125-140 Durrands, P.K., and Cooper, R.M. (1988) The role of pectinases in vascular wiltdisease as determined by defined mutants of Verticillium albo-atrum. Physiological and Molecular Plant Pathology 32: 363-371

- 75 -
Ebel, J., and Mithöfer, A. (1998) Early events in elicitation of plant defence. Planta 206: 335-348 Ebringerová, A., and Heinze, T. (2000) Xylan and xylan derivatives-biopolymers withdesirable properties, 1 Naturally occurring xylans structures, isolation procedures and properties. Macromolecular Rapid Communications 21: 542-556 Emami, K., and Hack, E. (2002) Conservation of XYN11A and XYN11B xylanase genes in Bipolaris sorghicola, Cochliobolus sativus, Cochliobolus heterotrophus, and Cochliobolus spicifer. Current Microbiology 45: 303-306 Enkerli, J., Felix, G., and Boller, T. (1999) The enzyme activity of fungal xylanase is not necessary for its elicitor activity. Plant Physiology 121: 391-397 Esquerre-Tugaye, M.T., Bourdat, G., and Dumas, B. (2000) Cell wall degrading enzymes, inhibitory proteins and oligosaccharides participate in molecular dialogue between plants and pathogens. Plant Physiology and Biochemistry 38: 157-163 Farmer, E.E., and Helgeson, J.P. (1987) An extracellular protein from Phytophthora parasitica var nicotianae is associated with stress metabolite accumulation in tobacco callus. Plant Physiology 85: 733-740 Faure, D. (2002) The family-3 glycoside hydrolases: from housekeeping functions to host-microbe interactions. Applied and Environmental Microbiology 68: 1485-2002 Felix, G., Regenass, M., and Boller, T. (1993) Specific perception of sub nanomolar concentration of chitin fragments by tomato cells: induction of extracellular alkalinization, changes in protein phosphorylation, and establishment of a refractory state. The Plant Journal 4: 307-316 Fincham, J.R.S. (1989). Transformation in Fungi. Microbiological Reviews 53: 148-170 Flatman, R., MacLauchlan,W.R., Juge, N., Furniss, C., Berrin, J-G., Kughe, R.K., Manzanares, P., Ladbury, J.E., O’briens, R., and Williamson, G. (2002)Interactions defining the specificity between fungal xylanases and xylanase-inhibiting protein XIP-I from wheat. Biochemical Journal 365: 773-781 Fry, S.C (1986). Cross-linking of matrix polymers in the growing cell walls of angiosperms. Annual Review of Plant Physiology 37: 165-186 Fry, S.C., Aldington, S., Hetherington, P.R., and Aitken, J. (1993). Oligosaccharidesas signals and substrates in the plant cell wall. Plant Physiology 103: 1-5 Fuchs, Y., Saxena, A., Gamble, H.R., and Anderson, J.D. (1989) Ethylenebiosynthesis-inducing protein from cellulysin is an endoxylanase. Plant Physiology 89: 38-143 Furniss, C.S.M., Belshaw, N.J., Alcocer, M.J.C., Williamson, G., Elliot, G.O., Gebrueres, K., Haigh, N.P., Fish, N.M., and Kroon, P.A. (2002) A family 11 xylanase from the Penicillium funiculosum is strongly inhibited by three wheat xylanase inhibitors. Biochimica et Biophysica Acta 1598: 24-29

- 76 -
Garcia-Maceira, F.I., Di Pietro, A., and Roncero, M.I.G. (1997) Purification andcharacterisation of a novel exopolygalacturonase from Fusarium oxysporum f.sp. lycopersici. FEMS Microbiology Letters 154: 37-43 Garcia-Romera, I., and Fry, S.C. (1995) The longevity of biologically-active oligogalacturonides in rose cell cultures: degradation by exo-polygalacturonase. Journal of Experimental Botany 46: 1853-1857 Garcia-Romera, I., and Fry, S.C. (1997) Upper limits for endogenous oligogalacturonides and free galacturonic acid in rose cell-suspension cultures: implications for the action of exo and endo-polygalacturonases. Journal of Plant Physiology 150: 241-246 Gebruers, K., Debyser, W., Goesaret, H., Proost, P., Van Damme, J., and Delcour, J.A. (2001). Triticum aestivum l. endoxylanase inhibitor (TAXI) consists of two inhibitors, TAXI I and TAXI II, with different specificities. Biochemical Journal 353: 239-244 Ghosh, V.K., and Deb, J.K. (1988) Production and characterisation of xylanase from Thielaviopsis basicola. Applied Microbiology and Biotechnology 29: 44-47 Giesbert, S., Lepping, H-B., Tenberge, K.B., and Tudzynski, P. (1998) The xylanolytic system of Claviceps purpurea: cytological evidence for secretion of xylanases in infected rye tissue and molecular characterisation of two xylanase genes. Phytopathology 88: 1020-1030 Gimeno, C. J., Ljungdahl, P.O., Styles, C.A., and Flink, G.R. (1992) Unipolar cell divisions in the yeast S. cervisiae lead to filamentous growth: regulation by starvation and RAS. Cell 68: 1077-1090 Gold, S.E., Garćia-Pedrajas, M.D., and Martínz-Espinoza, A.D. (2001) New (and used) approaches to the study of fungal pathogenicity. Annual Review of Phytopathology 39: 337-365 Gómes, I., Gómes, J., Steiner, W., and Esterbauer, H. (1992) Production of cellulase and xylanase by a wild strain of Trichoderma viride. Applied Microbiology and Biotechnology 36: 701-707 Gómez-Gómez, E., Roncero, M.I.G., Di Pietro, A., and Hera, C. (2001) Molecular characterisation of a novel endo-1,4-β-xylanase gene from the vascular wilt fungus Fusarium oxysporum. Current Genetics 40: 268-275 Gómez-Gómez, E., Roldán- Ruíz, M.C., Di Pietro, A., Roncero, M.I.G., and Hera, C. (2002) Role in pathogenesis of two endo-β-1,4 xylanase genes from vascular wilt fungus Fusarium oxysporum. Fungal Genetics and Biology 35: 213-222 Gonzalez-Candellas, L., and Kolattukudy, P.E. (1992) Isolation and analysis of a novel pectate lyase gene from the phytopathogenic fungus Fusarium solani. f.sp. pisi (Nectria haematococca, mating population VI). Journal of Bacteriology 174: 6343-6349

- 77 -
Görlach, J.M., Knaap, van D., and Walton, J.D. (1998) Cloning and targeted gene disruption of MLG1, a gene encoding two of three extracellular mixed–linked glucanases of Chochliobolus carbonum. Applied and Environmental Microbiology 64: 385-391 Gouka, R.J., Gerk, C., Hooykas, P.J.J., Bundock, P., Musters, W., Verrps, C.T.,and de Groot, M.J.A. (1999) Transformation of Aspergillus awamori by Agrobacterium tumefaciens-mediated homologous recombination. Nature Biotechnology 17 598-601 Greiner, S., Krausgrill, S., and Rausch, T. (1998) Cloning of a tobacco apoplasmic invertase inhibitor. Proof of function of the recombinant protein and expression analysis during plant development. Plant Physiology 116: 733-742 Hadfield, K.A., and Bennet, A. B. (1998) Polygalacturonases: many genes in search of a function. Plant Physiology 117: 337-343 Hamer, J.E, and Holden, D.W. (1997) Linking approaches in the study of fungal pathogenesis. Fungal Genetics and Biology 21: 11-16 Hanania, U., and Avni, A. (1997) High affinity binding site for ethylene-inducing xylanase elicitor on Nicotiana tabacum membranes. The Plant Journal 12: 113-120 Hanif, M., Pardo, A.J., Grofer, M., and Raudaskoski, M. (2002) T-DNA transfer and integration in the ectomycorrhizal fungus Suillus bovinus using hygromycin B as selectable marker. Current Genetics 41: 183-188 Henrissat, B. (1991) A classification of glycosyl hydrolases based on amino acid sequence similarities. Biochemical Journal 280: 309-316 Henrissat, B., and Bairoch, A. (1993) New families in the classification of glycosylhydrolases based on amino acid sequence similarities. Biochemical Journal 293: 781-788 Henrissat, B., Callebaut, I., Fabrega, S., Lehn, P., Mornon, J.-P., and Davies, G. (1995) Conserved catalytic machinery and the prediction of a common fold for several families of glycosyl hydrolases. Proceedings of the National Academy of Sciences USA 92: 7090-7094 Hilu, H.M., and Hooker, A.L. (1964) Host-pathogen relationship of Helminthosporiumturcicum in resistant and susceptible corn seedlings. Phytopathology 54: 570-575 Hilu, H.M., and Hooker, A.L. (1965) Localized infection by Helminthosporium turcicum on corn leaves. Phytopathology 55: 189-192 Holden, F.R., and Walton, J.D. (1992) Xylanase from the fungal maize pathogen Cochliobolus carbonum. Physiological and Molecular Plant pathology 40: 39-47

- 78 -
Holz, G., and Knox-Davies, P.S. (1986) Possible involvement of apoplast sugar in endo-pectin-trans-eliminase synthesis and onion bulb rot caused by Fusarium oxysporum f.sp. cepae. Physiological and Molecular Plant Pathology 28: 403-410 Howell, H.E. (1975) Correlation of virulence with secretion in vitro of three wall degrading enzymes in isolates of Scerotinia fructigena obtained after mutagen treatment. Journal of General Microbiology 90: 32-40 Huisman, M.M.H., Schols, H.A., and Voragen, AG.J. (2000) Glucuronoarabinoxylan from maize kernel cell walls are more complex than those from Sorghum kernel cell walls. Carbohydrate Polymers 43: 269-279 Hynes, M.J. (1996) Genetic transformation of filamentous fungi. Journal of Genetics 79: 297-311 Ishii, S. (1988) Factors influencing protoplast viability of suspension cultured rice cells during isolation process. Plant Physiology 88: 26-29 Jarvis, M.C., Forsyth, W., and Duncan, H.J. (1988) A survey of pectic content of non lignified monocot cell walls. Plant Physiology 88: 309-314 John, M., Röhrig, Schmidt, J., Walden, R., and Schell, J. (1997) Cell signalling by oligosaccharides. Trends in Plant Science 2: 111-115 Kahmann, R., and Basse, C. (1999) REMI (restriction enzyme mediated integration) and its impact on the isolation of pathogenicity genes in fungi attacking plants. European Journal of Plant pathology 105: 221-229 Kalkkinen, N., and Tilgmann, C. (1988) A gas-pulsed liquid phase sequencer constructed from Beckman 890D instrument by using Applied Biosystems delivery and cartridge blocks. Journal of Protein Chemistry 262: 242-243 Keen, N.T. (2000) A century of plant pathology: a retrospective view on understanding host-parasite interactions. Annual Review of Phytopathology 38: 31-41 Keen, N.T., and Yang, C.-H. (1999) Functional genomics: Plant-microbe interactions gingerly puts a foot in the water. Physiological and Molecular Plant Pathology 55: 313-315 Kiellszewski, M. J., and Lamport, D.T.A. (1994) Extensin: repetitive motifs, functional sites, post translational modification codes, and phylogeny. The Plant Journal 5: 157-172 Knox-Davies, P.S. (1974) Penetration of maize leaves by Helminthosporium turcicum. Phytopathology 64: 1468-1470 Knox-Davies, P.S., and Dickson, J.G. (1960) Cytology of Helminthosporium turcicum andits ascigerous stage, Trichometasphaeria turcica. American Journal of Botany 47: 328-339 Kulkarni, N., Shendye, A., and Rao, M. (1999) Molecular and biotechnological aspects of xylanases. FEMS Microbiology Reviews 23: 411-456

- 79 -
Kulmburg, P., Mathieu, M., Dowzer, C., Kelly, J., and Felenbok, B. (1993) Specific binding sites in the alcR and alcA promoters of the ethanol regulon for the creA repressor mediating carbon catabolite repression in Aspergillus nidulans. Molecular Microbiology 7: 847-857 Kunik, T., Tzfira, T., Kapulnik, Y., Gafini, Y., Dingwall, C., and Citovisky, V. (2001) Genetic transformation of HeLa cells by Agrobacterium. Proceedings of the National Academy of Sciences, USA 98: 1871-1876 Laemmli, U.K. (1970) Cleavage of structural proteins during the assembly of bacteriophage T4. Nature 227: 680-685 Lotan, T., and Fluhr, R. (1990) Xylanase, a novel elicitor of pathogenesis-related proteins in tobacco, uses a non-ethylene pathway for induction. Plant Physiology 93: 811-817 Malonek, S., and Meinhardt, F. (2001) Agrobacterium tumefaciens-mediated genetic transformation of the phytopathogenic Ascomycete, Calonectra morgani. Current Genetics 40: 152-155 Margolles-Clark, E., Ilmen, M., and Penttilä, M. (1997) Expression patterns of ten hemicellulase genes of the filamentous fungus Trichoderma reesei on various carbon sources. Journal of Biotechnology 57: 167-179 McLauchlan, W.R., Garcia-Conesa, M.T., Williamson, G., Martinus, R., Ravestein, P., and Maat, J. (1999) A novel class of protein from wheat which inhibits xylanases. Biochemical Journal 338: 441-446 McNeil, M., Darvill, A.G., and Fry, S.C. (1984) Structure and function of the primary walls of plants. Annual Review of Biochemistry 53: 625-663 Mehdy, M.C. (1994) Active oxygen species in plant defence against pathogens. Plant Physiology 105: 467-472 Mikosch, T.S.P., Lavvrijssen, B., Sonnenberg, A.S.M., and Griensvenl, J.-L.D. (2001) Transformation of the cultivated mushroom Agaricus bisporus (Lange) using T-DNA from Agrobacterium tumefaciens. Current Genetics 39: 35-39 Miller, G. L. (1959) Use of dinitrosalicylic acid reagent for determination of reducing sugar. Analytical Chemistry 3: 426-428 Mitchell, T.K., Thon, M.R., Jeong, J.-S. Brown, D., Deng, J., and Dean, R.A. (2003) The rice blast pathosystem as a case study for the development of new tools and raw materials for genome analysis of fungal pathogens. New Phytologist 159: 53-61 Mugo, S., and Hoisington, D. (2001) Biotechnology for the improvement of maize for resource poor farmers: the CIMMYT approach. In: Mandefro, N., Tanner, D., and Twumasi-Afriyie, S (ed.), Enhancing the contribution of maize to food security in Ethiopia. Proceedings of the second National workshop of Ethiopia. 12-16 November 2001. EARO and CIMMYT. Addis Ababa, Ethiopia, pp 203-213

- 80 -
Mullins, E.D., Chen, X., Romaine, P., Raina, R., Geiser, D.M., and Kang, S. (2001) Agrobaterium tumefaciens-mediated transformation of Fusarium oxysporum: an efficient tool for insertional mutagenesis and gene transfer. Phytopathology 91: 173-180 Mullins, E.D., and Kang, S. (2001) Transformation: a tool for studying fungal pathogens of plants. Cellular and Molecular Life Sciences 58: 2043-2052 Murphy, J.M., and Walton, J. D. (1996) Three extracellular proteases from Cochliobolus carbonum: cloning and targeted disruption of ALP1. Molecular Plant-Microbe Interactions 9: 290-297 Nkonya, E., Xavery, E.P, Akonaay, H., Mwangi, W., Anandajasekeram, P.,Verkuijl, H., Martella, D and Moshi, A, (1998). Adoption of maize production technologies in Northern Tanzania. CIMMYT, The United Republic of Tanzania and Southern African Center for cooperation in Agricultural Research (SACCAR). 56 P. Nojiri, H., Sugimori, M., Yamane, H., Nishimura, Y., Yamada, A., Shibuya, N., Kodama, O., Murofushi, N., and Ohmori, T. (1996) Involvement of jasmonic acid inelicitor-induced phytoalexin production in suspension cultured rice cells. Plant Physiology 110: 387-392 Otte, O., and Barz, W. (1996) The elicitor-induced oxidative burst in cultured chickpea cells drives the rapid insolubilisation of two cell wall structural proteins. Planta 200: 238-246 Peck, S., Nuhse, T.S., Hess, D., Iglesias, A., Meins, F., and Boller, T. (2001) Directed proteomics identifies a plant specific protein rapidly phosphorylated in response to bacterial and fungal elicitors. Plant Cell 13: 1467-1475 Perkins, J.M., and Pedersen, W.L. (1987) Disease development and yield losses associated with northern leaf blight on corn. Plant Disease 71: 940-943 Poutanen, K., and Puls, J. (1988) Characteristics of Trichoderma reesei β-xylosidase and its use in hydrolysis of solubilized xylans. Applied Microbiology and Biotechnology 28: 425-432 Prade,R.A., Ayoubi, P., Krishnan, S., Macwana, S., and Russell, H. (2001) Accumulation of stress and inducer-dependent plant cell wall degrading enzymes during asexual development in Aspergillus nidulans. Genetics 157: 957-967 Ransom, R.F., and Walton, J.D. (1997) Purification and characterisation of extracellular β-xylosidase and α-arabinosidase from plant pathogenic fungus Cochliobolus carbonum. Carbohydrate Research 297: 357-364 Rho, H-S., Kang, S., and Lee, Y-H. (2001) Agrobacterium tumefaciens-mediated transformation of the plant pathogenic fungus Magnaporthe grisea. Molecules and Cells 12: 407-411 Riggle, P.J., and Kumamoto, C. (1998) Genetic analysis in fungi using restriction-enzyme-mediated -integration . Current Opinion in Microbiology 1: 395-399

- 81 -
Robinson, D. G . (1991) What is a plant cell ? The last word. Plant Cell 3: 1145-1146 Ronne, H. (1995). Glucose repression in fungi. Trends in Genetics 11: 12-17 Rose, J.K.C., and Bennet, A. B. (1999) Co-operative disassembly of the cellulose-xyloglucan network of plant cell walls: parallels between cell expansion and fruit ripening.. Trends in Plant Sciences 4: 176-183 Rose, J.K.C., Ham, K-S., Darvill, A.G., and Albersheim, P. (2002) Molecular cloning and characterisation of glucanase inhibitor proteins: coevolution of a counterdefence mechanism by plant pathogens. Plant Cell 14: 1329-1345 Rubinstein, A.L., Marquez, J., Suarez-Cervera, M., and Bedinger, P.A. (1995a) Extensin-like glycoproteins in the maize pollen tube wall. Plant Cell 7: 2211-2211 Rubinstein, A.L., Broadwater, A.H., Lowrey, K.B., and Bedinger, P.A. (1995b) Pex1, a pollen-specific gene with an extensin-like domain. Proceedings of the National Academy of Sciences, USA 92: 3086-3090 Ryan, C.A. (1990) Protease inhibitors in plants: Genes for improving defence againstinsects and pathogens. Annual Review of Phytopathology 28: 425-449 Ryan, C.A., and Farmer, E.E. (1991) Oligosaccharide signals in plants: a current assessment. Annual Review of Plant Physiology and Plant Molecular Biology 42: 651-674 Raymundo, A.D., and Hooker, A.L. (1981) Measuring the relationship between northern leaf blight and yield losses. Plant Disease 65: 325-327 Sambrook,, J., Fritsch, E.F., and Maniatis, E.A. (1989) Molecular cloning. A laboratory manual. Second edition. Cold Spring Harbour Press. Cold Spring Harbour. NY Schnabelrauch, L.S., Keilliszewski, M., Upham, M., B.L., Alizedeh, H., and Lamport, D.T.A. (1996) Isolation of pI 4.6 extension peroxidase from tomato cell suspension culture and identification of Val-Tyr-Lys as putative intermolecular cross-link site. The Plant Journal 9: 477-489 Schooveneveld-Bergmans, M.E.F., Hopman, A.M.C.P., Beldman, G., and Voragen, A.G.J. (1998) Extraction and partial characterisation of feruloylated glucuronoarabinoxylamns from wheat bran. Carbohydrate Polymers 35: 39-47 Schulte, R., and Bonas, U. (1992) A Xanthomonas pathogenicity locus is induced by sucrose and sulphur containing amino acids. Plant Cell 4: 79-86 Schäffer, H.J., Leykam, J., and Walton, J.D. (1994) Cloning and targeted gene disruption of EXG1, encoding exo-β 1,3-glucanase, in the phytopathogenic fungus Cochliobolus carbonum. Applied and Environmental Microbiology 60: 594-598

- 82 -
Scott-Craig, J.S., Pannacione, D.G., Cervone, F., and Walton, J. D. (1990) Endo-polygalacturonase is not required for pathogenicity of Cochliobolus carbonum on maize. Plant Cell 2: 1191-1200 Senior, D.J., Mayers, P.R., and Saddler, J.N. (1989) Production and purification of xylanases. In: Lewis, N.G., and Paice, M.G. (ed.) Plant Cell Wall Polymers Biogenesis and Biodegradation. ACS symposium Series 399, pp 644-653 Sequeira, L. (1993) Research in plant-microbe interactions: making it relevant. In: Nester, E.W., and Verma, D.P.S., (ed.) Advances in Molecular Genetics of Plant-Microbe Interactions. Proceedings of the 6th International Conference on Plant Growth Substances, Seattle, Washington, U.S.A, pp 3-14 Sharon, A., Fuchs, Y., and Anderson, J.D. (1993) The elicitation of ethylene biosynthesis by Trichoderma xylanase is not related to the cell wall degradation activity of the enzyme. Plant Physiology 102: 1325-1329 Shedletzky, E., Shumel, M., Trainin, T., Kalman, S., and Delmer, D. (1992) Cell Wall structure in cells adapted to growth on the cellulose synthesis inhibitor 2,6-dichlorobenzonitrile. A comparison between two dicotyledonous plants and graminaceous monocots. Plant Physiology 100: 120-130 Sheppard, P.O., Grant, F.J., Oort, P.J., Sprecher, C.A., Foster, D.C., Hagen, F.S., Upshall, A., Mcknight, G., L and O´Hara, P.J. (1994) The use of conserved cellulase family-specific sequences to clone cellulose homologous cDNAs from Fusarium oxysporum. Gene 150: 163-167 Shibuya, N., and Nakane, R. (1984) Pectic polysaccharides of rice endosperm cell walls. Phytochemistry 23: 1425-1429 Shibuya, N., and Minami, E. (2001). Oligosaccharide signalling for defence response in plant. Physiological and Molecular Plant Pathology 59: 223-233 Showalter, A.M. (1993). Structure and function of plant cell wall proteins. Plant Cell 5:9-23 Shurtleff, M.C. (1980) Compendium of corn diseases. Second edition. The American Phytopathological Society, St Paul Minnesota Slomezynski, D., Nakas, J.P., and Tanenbaum, S.W. (1995) Production and characterisation of laccase from Botrytis cinerea. Applied and Environmental Microbiology 61: 907-912 Soanes, D.M., Skinner, W., Keon, J., Hargreaves, J., and Talbolt, N.J. (2002) Genomics of phytopathogenic fungi and development of bioinformatic resources. Molecular Plant-Microbe Interactions 15: 421-427 Sommer-Knudsen, J., Bacic, A., and Clarke, A.E. (1998) Hydroxyproline-rich glycoproteins. Phytochemistry 47: 483-497

- 83 -
Solomon, P.S., Tan, K.-C., and Oliver, R.P. (2003) The nutrient supply of pathogenic fungi; a fertile field for study. Molecular Plant Pathology 4: 203-210 Southerton, S.G., Osbourn, A.E., Dow, J.M., and Daniels, M.J. (1993) Two xylanases from Gaeumannomyces graminis with identical N-terminal amino acid sequence. Physiological and Molecular Plant Pathology 42: 97-107 Sposato, J.P., Ahn, J-H., and Walton, J.D. (1995) Characterisation and disruption of a gene in the maize pathogen Cochliobolus carbonum encoding a cellulase lacking a cellulase binding domain and hinge region. Molecular Plant-Microbe Interactions 8: 602-609 Srivastava, B.R., and Srivastava, A.K. (1993) Characterisation of bacterial xylanase resistant to catabolic repression by glucose and xylose. Biotechnology Letters 15: 847-852 Stahl, D. J., and Schaeffer, W. (1992) Cutinase is not required for fungal pathogenesis on pea. Plant Cell 4: 621-629 Stebbins, G. L. (1992) Comparative aspects of plant morphogenesis: a cellular, molecular, and evolutionary approach. American Journal of Botany 79: 589-598 Stiefel,V., Ruiz-Avila, L., Raz, R., Valles, M.P., Gomez, J., Pages,M, Martinez-Izquierdo, J.A., Ludevid, M.D., Langdale, J.A.Nelson, T., and Puigdomenech, P. (1990) Expression of a maize wall hydroxyproline-rich glycoprotein gene in early leaf and root vascular differentiation. Plant Cell 2: 785-793 Strauss, J., Mach, R.L., Zeilinger, S., Hartler, G., Georg, Stöffler, G., Wolschek, M., and Kubicek, C.P. (1995) Cre1, the carbon catabolite repressor protein from Trichoderma reesei. FEBS Letters 376: 103-107 Sunna, A., and Antranikian, G. (1997) Xylanolytic enzymes from fungi and bacteria. Critical Reviews in Biotechnology 17: 39-67 Sweigard, J. A., Chumly, F. G., and Valent, B. (1992) Disruption of Magnaporthe grisea cutinase gene. Molecular and General Genetics 232: 183-190 Swofford, D.L. (2000) PAUP∗ . Phylogenetic analysis using Parsimony and other methods. Version 4. Sinauer Associates, Sunderland, Massachsetts Syvanen, M. (1994) Horizontal gene transfer: evidence and possible consequences. Annual Review of Genetics 28: 237-261 Tahir, T.A., Berrin, J-G., Flatman, R., Russel, A., and Roepstorff, P. (2002) Specific characterisation of substrate and inhibitor binding sites for a glycosyl hydrolase family 11 xylanase from Aspergillus niger. The Journal of Biological Chemistry 277: 44035-44043 Tans-Kersten, J., Guan, Y., and Allen, C. (1998) Ralstonia solanacearum pectin Methyle sterase is required for growth on methylated pectin, but not for bacterial wilt virulence. Applied and Environmental Microbiology 64: 4918-4923

- 84 -
Thompson, J.D., Higgins, D.G., and Gibson, T.J. (1994) CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, positions-specific gap penalties and weight matrix choice. Nucleic Acid Research 22: 4673-4680 Thomson, J.A. (1993) Molecular biology of xylan degradation. FEMS Microbiology Reviews 104: 65-82 Thon, M.R., Nuckles, E.M., and Vaillancourt, L.J. (2000) Restriction-enzyme-mediated-integration used to produce pathogenicity mutants of Colletothrichum graminicola. Molecular Plant-Microbe Interactions 13: 1356-1365 Tilahun, T., Ayana, G., Abebe, F., and Wegary, D (2001) Maize pathology research in Ethiopia: a review. In: Mandefro, N., Tanner, D., and Twumasi-Afriyie, S., (ed.), Enhancing the contribution of maize to food security in Ethiopia. Proceedings of the second National workshop of Ethiopia. 12-16 November 2001, EARO and CIMMYT, Addis Ababa, Ethiopia, pp 97-105 Tonukari, N.J., Scott-Craig, J.S., and Walton, J.D. (2000) The Cochliobolus carbonum SNF1 gene is required for cell-wall degrading enzyme expression and virulence on maize. Plant Cell 12: 237-247 Tonukari, N.J., Scott-Craig, J.S., and Walton, J.D. (2002) Influence of carbon source on the expression of Cochliobolus carbonum xylan degrading enzyme genes. African Journal of Biotechnology 1: 64-66 Treitel, M.A., Kuchen, S., and Carlson, M. (1998) Snf1 protein kinase regulates posphorylation of the Mig1 repressor in Saccharomyces cervisae. Molecular and Cellular Biology 18: 6273-6280 Uffen, R.L. (1997) Xylan degradation: a glimpse at microbial diversity. Journal of Industrial Microbiology and Biotechnology 19: 1-6 Van Hoof, A., Leykam, J. Shaeffer, H.J., and Walton, J.D. (1991) A single β-1,3-glucanase secreted by the maize pathogen Cochliobolus carbonum acts by an exolytic mechanism. Physiological and Molecular Plant Pathology 39: 259-267 Van Kan, J.A.L., vant Klooster, J., Wagemakers, C.A.M., Dees, D.C.T., and vander Vult-Bergmans, C.J.B. (1997) Cutinase A of Botrytis cinerea is expressed but not essential during penetration of Gerbera and Tomato. Molecular Plant-Microbe Interactions 10: 30-38 Van Peij, N.N.M.E., Gielkenes, M.M.C., and De Vries, R.P., Visser, J., and De Graaff,L-H. (1998) The transcriptional activator XlnR regulates both xylanolytic and endoglucanase gene expression in Aspergillus niger. Applied and Environmental Microbiology 64: 3615-3619 Varner, J.E., and Lin, L.S. (1989) Plant cell wall architecture. Cell 56:231-239 Viikari, L., Kantelinen, A., Sundquist, J., Linko, M. (1994) Xylanase in bleaching: from an idea to industry. FEMS Microbiology Reviews 13: 335-350

- 85 -
Waksman, G. (1988) Molecular cloning of genes expressed specifically during induction of cell wall degrading enzymes from Sclerotinia sclerotiorum and preliminary identification of a fungal β-galactosidase encoding gene by expression in Escherichia coli. Current Genetics 14: 91-93 Waksman, G. (1989) Molecular cloning of β-glucosidase–encoding gene from Sclerotinia sclerotiorum by expression in E. coli. Current Genetics 15: 295-297 Walker-Simmsons, M., Jin, D., West, C.A., Hadwiger, l., Ryan, C.A. (1984) Comparison of proteinase inhibitor inducing activities and phytoalexin elicitor activities of a pure fungal endopolygalacturonase, pectic fragments of chitosans. Plant Physiology 76: 833-836 Walton, J.D., and Cervone, F. (1990) Endopolygalacturonase from the maize pathogen Cochliobolus carbonum. Physiological and Molecular Plant Pathology 36: 351-359 Wang, H., and Jones, R.W. (1995) Cloning, characterisation and functional expression of an endoglucanase-encoding gene from the phytopathogenic fungus Macrophomina phaseolina. Gene 158: 125-128 Warren, R.A.J. (1996) Microbial hydrolysis of polysaccharides. Annual Review of Microbiology 50: 183-212 Weselake, R.J., Macgregor, A.W., and Hill, R.D. (1983) An endogeneous α-amylase inhibitor in barley kernels. Plant Physiology 72: 809-812 Wong, K.K.Y., Tan, L.U.L., and Saddler, J.N. (1986) Functional interactions among three xylanases from Trichoderma harzianum. Enzyme and Microbial Technology 8: 617-622 Wong, K.K.Y.,Tan, L.U.L., and Saddler, J.N. (1988) Multiplicity of β-1,4-xylanase in microorganisms: functions and applications. Microbiological Reviews 52.305-317 Wu, S.-C., Kauffmann, S., Darvill, A.G., and Albersheim, P. (1995) Purification, cloning and characterisation of two xylanases from Magnaporthe grisea, the rice blast fungus. Molecular Plant-Microbe Interactions 8: 506-514 Wu, S.-C., Ham, K-S., Darvill, A.G., and Albersheim, P. (1997) Deletion of two endo-β-1,4-xylanase genes reveals additional isozymes secreted by rice blast fungus. Molecular Plant-Microbe Interactions 10: 700-708 Yelton, M.M., Hamer, J.E., and Timberlake, W.E. (1984) Transformation of Aspergillus nidulans by using a trpC plasmid. Proceedings of the National Academy of Sciences, USA 81: 1470-1474 York, W.S., Darvill, A.G., McNeil, M., Stevenson, T.T., and Albersheim, P. (1985) Isolation and characterisation of plant cell walls and cell wall components. Methods in Yuan, S., Wu, Y., and Cosgrove, D.J. (2001) A fungal endoglucanase with plant cell wall extension activity. Plant Physiology 127: 324-333


















