Embed Size (px)
Citation preview

Cloning and Characterization of the Prostate-SpecificMembrane Antigen PromoterDavid Good,1,2 Paul Schwarzenberger,1,2,3 James A. Eastham,8 Robert E. Rhoads,7 Jay D. Hunt,2,5
Michael Collins,1,2,3 Mark Batzer,1,2,6 Chris Theodossiou,1,2,3 Jay K. Kolls,1,2,3,4 and Sidney R. Grimes7,9*1Gene Therapy Program, School of Medicine, New Orleans, Louisiana 701122Stanley S. Scott Cancer Center, School of Medicine, New Orleans, Louisiana 701123Department of Medicine, School of Medicine, New Orleans, Louisiana 701124Department of Pediatrics, School of Medicine, New Orleans, Louisiana 701125Department of Biochemistry and Molecular Biology, School of Medicine, New Orleans, Louisiana 701126Department of Pathology, School of Medicine, New Orleans, Louisiana 701127Department of Biochemistry and Molecular Biology, School of Medicine, Louisiana State UniversityMedical Center, Shreveport, Louisiana 711308Department of Urology, School of Medicine, Louisiana State University Medical Center, Shreveport,Louisiana 711309Veterans Administration Medical Center, Shreveport, Louisiana 71101
Abstract Prostate-specific membrane antigen (PSMA) is a protein that is expressed predominantly in normalprostate epithelial cells and in most adenocarcinomas of the prostate (Cap) and in virtually all Cap metastases. In thisarticle we describe the cloning of a 2-kb human genomic DNA fragment containing the 58 upstream untranslated regionof the PSMA gene and present evidence that it provides promoter activity. When the DNA fragment was cloned intotransient expression vectors to examine promoter activity, the vectors were functional in promoting expression in severalprostate and nonprostate cell lines in transient transfection assays. A 614-bp fragment derived from the 38 end of the 2-kbfragment may represent the minimal PSMA promoter as determined by deletion mutagenesis. The 2-kb fragmentcompared with the 614-bp fragment provided higher expression levels when using prostate-derived cell lines (DU 145and LNCaP). The increased transcription using the 2-kb fragment was not as great in non-prostate cell lines. Little or notranscription over basal levels was seen with a 232-bp promoter fragment. When the concentration of dihydrotestoster-one was depleted or supplemented in the growth medium, no significant effect was seen on PSMA-promoted transientexpression in LNCaP cells, a prostate cell line. J. Cell. Biochem. 75:395–405, 1999. Published 1999 Wiley-Liss, Inc.†
Key words: prostate cancer; prostate-specific; prostate-specific membrane antigen; prostate-specific membrane antigengene; regulation of gene expression; regulation of gene transcription; promoter; androgen
Prostate-specific membrane antigen (PSMA)is a 750-amino acid type II transmembraneglycoprotein with an apparent molecular weightof approximately 100 kDa [Israeli et al., 1993].PSMA has been shown to be a unique folatehydrolase-carboxypeptidase that can releaseglutamate with either g- or a-linkage [Heston,1997]. It has been shown to have homology to
the brain protein NAALADase, and it exhibitsin vitro neuropeptidase activity [Troyer et al.,1995; Carter et al., 1996]. A 140-amino acidextracellular region of PSMA has significanthomology to the transferrin receptor [Troyer etal., 1995; Carter et al., 1996].
PSMA was originally defined by the monoclo-nal antibody 7E11.C5 and has been shown to behighly prostate specific [Horoszewicz et al.,1987]. In histological studies using monoclonalantibodies that recognize PSMA, the proteinwas shown to have specificity for prostate epi-thelial tissue [Horoszewicz et al., 1987]. Weakexpression was seen in salivary ducts and theproximal small intestine, as well as in smoothmuscle [Murphy, 1995; Troyer et al., 1995]. The
Grant sponsor: National Institutes of Health; Grant num-ber: GM20818; Grant number: HD29381; Grant sponsor:Veterans Administration.*Correspondence to: Sidney Grimes, Research Service (151),VA Medical Center, 510 E. Stoner Ave., Shreveport, LA71101-4295. E-mail: [email protected] 19 January 1999; Accepted 1 February 1999
Journal of Cellular Biochemistry 74:395–405 (1999)
Published 1999 Wiley-Liss, Inc. †This article is a U.S. Govern-ment work and, as such, is in the public domain in the United Statesof America.

protein has been detected in a subset of proxi-mal renal tubules and intense staining wasobserved in endothelial cells of capillary vesselsin peritumoral and endotumoral areas of cer-tain malignancies [Silver et al., 1997].
PSMAmRNAhas been detected in the LNCaPprostate cancer cell line, but not in DU-145 orPC-3 cell lines in Northern blot analyses [Is-raeli et al., 1993]. Ribonuclease protection re-vealed a significant level of PSMA mRNA onlyin prostate. Much lower steady-state levels weredetected in brain and salivery gland [Israeli etal., 1994b]. Nevertheless, the precise functionof PSMA in the prostate, in prostate cancer, andin these various other tissues remains un-known.
The PSMA gene has been cloned [Israeli etal., 1993] and has been shown to be located onchromosome 11 [Leek et al., 1995; Rinker-Schaeffer et al., 1995]. Two molecular forms ofthe protein, designated PSMAand PSMA8, havebeen identified [Su et al., 1995]. PSMA8, whichis derived from an alternatively spliced tran-script that lacks the transmembrane portion ofPSMA, is located in the cytoplasm and its levelis typically higher in normal cells [Su et al.,1995]. Recently, a 1.2-kb portion of the 58 pro-moter region of the PSMA gene was reported tobe functional in driving expression of a reportergene [O’Keefe et al., 1998].
Cancer of the prostate (CaP) is the mostfrequently diagnosed malignancy in men. It isestimated that there will be more than 184,000newly diagnosed cases of CaP and 39,000 deathsin 1998. This disease constitutes a major healthproblem at both its early and advanced stages,with no cure available for the latter. PSMApromises to be useful as a diagnostic target forprostatic cancer detection [Abdel-Nabi et al.,1990; Wynant et al., 1991; Babaian et al., 1994;Murphy et al., 1995; Haseman et al., 1996].Western blot assays of serum derived from pros-tatic cancer patients using the 7E11.C5 anti-body have demonstrated increased levels of7E11.C5 reactive PSMA antigen compared tonormal subjects [Murphy et al., 1995, 1997]. Inaddition to the successful use of these antibod-ies for prostate cancer detection, there is apotential for their use in treatment of prostatecancer.
Previous observations revealed an increasein PSMA expression in prostate tumors of pa-tients after chemical or surgical androgen abla-
tion [Wright et al., 1996], suggesting that andro-gens may be involved in downregulatingtranscription by the PSMA promoter. There is adiminished steady-state level of PSMA mRNAin LNCaP cells treated with physiological con-centrations of dihydrotestosterone. In these ex-periments, PSMA mRNA levels were highest incharcoal-stripped medium [Israeli et al., 1994b].However, the promoter has neither been de-fined completely nor examined for regulationby androgens. Because of the potential value ofthe PSMA gene, we felt that it was important toclone the PSMA promoter. Furthermore, it wasimportant to demonstrate that the cloned pro-moter fragment was functional in promotingtranscription of a reporter gene and to deter-mine whether the promoter was responsive tochanges in the concentration of dihydrotestos-terone in transient transfection assays.
MATERIALS AND METHODSCloning of PSMA cDNA
Total cellular RNAwas extracted from LNCaPcells (ATCC, Rockville, MD) by a single-stepmethod [Tjoa et al., 1997]. The primer pairCAR2 (58-AGC CAC GCC ACG CTC TTG-38)and CAR4 (58-TCT TTC TGAGTGACATAC-38),specific for human PSMA, was used to preparecDNA from 5 µg of total RNA, using avianmyeloblastosis virus reverse transcriptase intwo separate reactions following the specifica-tions of the manufacturer (Promega, Madison,WI). The cDNA from these reactions was ampli-fied in two separate polymerase chain reaction(PCR) reactions using the primer pair CAR1(58-TGC AGG GCT GAT AAG CGA G-38) andCAR 2 or CAR3 (58-TCA TCC AAT TGG ATACTA TG-38) and CAR4 (Roche Diagnostic Sys-tems, Branchburg, NJ). PCR was conducted for30 cycles heating 94°C for 30 s, 56°C for 30 s,and 72°C for 1 min. The PCR amplification withCAR1 and CAR2 resulted in a 1,472-bp productspanning nucleotides 134–1,605 (based on Gen-Bank sequence M99487); PCR amplificationwith CAR3 and CAR4 resulted in a 1,427-bpproduct spanning nucleotides 1143–2569. Theseproducts contained a 463-bp overlap. A 2,436-bpPCR product spanning the entire PSMA cDNAwas created using the products of CAR1 andCAR2 amplification (1) and the products ofCAR3 and CAR4 amplification (1) as templatetogether with primers CAR1 and CAR4. The
396 Good et al.

2.4-kb product was TAcloned into pCR2.1 (Invit-rogen, Carlsbad, CA).
Human BAC Library Screening and Analysis
PSMA cDNA served as a PCR template, andits published sequence was used to design PCRoligonucleotide primers. Primer pairs weretested so that PCR amplification from bothhuman genomic DNA and PSMA cDNA yieldedproducts with the same predicted size, ensur-ing amplification of a segment without introns.A primer pair satisfying these requirements (58TAC CAC ATT TAG CAG GAA CA 38); (58 ATGAGT CTT ATT TGG GTA GG 38) amplified thefragment between nucleotides 500 and 614 ofthe published PSMAcDNA. These primers wereused to screen a commercially available pooledhuman bacterial artificial chromosomal (BAC)library (Research Genetics, Huntsville, AL) byPCR. The pooled library is 9-fold redundant, soscreening with a single PCR amplificationshould result in the identification of multipleclones. Three putative positive BAC clones wereidentified. The one (BAL 519 M3) exhibiting thestrongest PCR amplification was chosen for fur-ther studies.
Total BAC DNA and PSMA cDNA were di-gested with Bgl1 and analyzed by Southernblot using random primed cDNA as a probe todetect BAC restriction fragments that contain58 flanking regulatory sequences for the PSMAgene. Southern blots were exposed to KodakXAR-5 film and processed as previously de-scribed [Zhau et al., 1994].
Subcloning the PSMA Promoter
BAC Clone 96012 519 M3 was partially di-gested with Sau3AI and fragments of 10–12 kbwere excised from a 0.7% low melting pointagarose gel (SeaPlaque GTG, FMC BioProd-ucts, Rockland, ME) and subcloned into BamHI-digested and dephosphorylated plBluescriptSK1 vector (Stratagene, La Jolla, CA). The liga-tions were introduced into host Escherichia colicells plated and screened by colony hybridiza-tion, using the PCR product from the 58 UTR ofthe PSMA gene as a probe. A portion of the 58untranslated region (58 UTR) of PSMA wasamplified using the primer pair 58 CAG AGCTAA GAG CTG AAG CAT AGG C 38 and 58 CAGTAG TCA CAT TTA GTG GTG GCG 38. Thermo-cycler settings were 95°C for 5 min, followed by35 cycles at 95°C for 30 s, 55°C for 30 s, and
72°C for 1 min. Separation by electrophoresison a 2% agarose gel provided a fragment of theexpected size (640 bp). This PCR fragment wasexcised, purified, and [32P]-labeled with ran-dom hexamer primers (Stratagene) to serve asprobe for colony hybridization. Three positiveclones were identified. Subsequent restrictionendonuclease cleavage with EcoRI exhibitedsimilar restriction patterns, suggesting that theclones contained identical sequences. Southernblot analysis using the 58 UTR of PSMA as aprobe showed positive hybridization signalsfrom two EcoRI fragments of 2.1 kb and 1.0 kb.These fragments were excised, purified, andsubcloned into EcoRI digested pBluescript SK1.
Sequence Analysis
After restriction digestion of the 2.1-kb in-sert, overlapping fragments were subcloned intopUC19 and sequenced (McConnell Research,San Diego, CA). The sequence of the promoterfragment derived from the BAC library hasaccession number AF044684. The 2.1-kb EcoRIfragment was also subcloned and sequencedfrom an independently screened human PACclone PAC-280-17L (Genome Systems, St. Louis,MO) (Accession numberAF061571). Both cloneswere used for expression studies with similarresults.
Tissue Culture and Transient TransfectionAnalysis
All cell lines except ALVA-31 [Loop et al.,1993] and PPC-1 [Brothman et al., 1989] wereobtained from ATTC and were grown at 37°C in5% CO2. Media was supplemented with 10%fetal bovine serum (FBS). Tissue culture re-agents were obtained from Gibco (Gaithers-burg, MD). The following media was used in therespective cell lines: RPMI for LNCaP, EMEMfor DU45, and Dulbecco’s modified Eagle’s me-dium (DMEM) for 911 and HeLa cells. The PZline was grown in completely defined Keratino-cyte-SFM medium (serum-free medium, Gibco).Within 24 h of passage, cells were transfectedwith LipofectAMINE (Gibco) in 6-well plates.Cells were transfected when 90% confluent. Allcells were analyzed for reporter gene expres-sion 48 h after completion of the transfectionprocedure following the recommendations ofthe manufacturer for lacZ (Galactolite, Tropix,Medford, MA) and luciferase gene (Promega)expression, on a Perkin-Elmer luminometer (2-s
Characterization of the PSMA Promoter 397

delay time and 5-s integration time). Proteinconcentration of each sample was determinedby a Bichinoic acid/Cu-based detection system(Pierce, Rockford, IL) and all measurementsare expressed as relative light units (RLU) permg protein.
Statistical Analysis
Means of data were calculated by analysis ofvariance with Fisher’s follow-up testing (Stat-view, San Diego, CA). Statistical significance isassumed at P , 0.05. Alignment and compari-son of nucleotide sequences and evaluation oftranscription factor binding sites were per-formed with the MacVector computer program.In addition, the web-based software programTRANSFAC (http://agave.humgen.upen.edu/utess/tess32) was used to help identify transcriptionfactor binding sites.
Construction of Reporter Plasmids
Fragments of various sizes (232, 614, and1,915 bp) containing the upstream portions ofthe PSMA gene and 185 bp of transcribed se-quence were cloned into mammalian gene ex-pression vectors containing a luciferase re-porter gene (the pGL3 series from Promega).These primers are listed below and the loca-tions of the three upstream primers are under-lined in Figure 1: PSM 167, 58 TTACTGGGT-GATCCCACGTTTTAC 38 (used to produce the1,915-bp fragment); PSM 1468, 58 GAT GGGGTT TCA CCA TGT TGG38 (used to producethe 614-bp fragment); and PSM 1860, 58 GCAAGA GCT GGA CAT TTT CCA AG38 (used toproduce the 232-bp fragment). The same lowerprimer (PSM 1 70, 58 GCG GGG TAA AGTCTCT CTC AAT C 38) was used with each of thethree upper primers.
PCR amplification reactions yielded frag-ments of the PSMA promoter with the pre-dicted sizes. PCR conditions were followed asrecommended by the manufacturer of the re-agents (Promega). Thermocycle settings weredenaturation at 95°C for 5 min, followed by 35cycles at 95°C for 30 s, 55°C for 30 s, and 72°Cfor 1 min, followed by heating at 72°C for 10min.
The three resulting fragments were sepa-rated by gel electrophoresis and purified andinitially TA cloned into pCR2.1 (Invitrogen).Clones were excised from pCR2.1 with KpnIand XhoI. The three fragments were moved
into the KpnI and XhoI sites of the polylinker inpGL3-Enhanced (pGL3EV) and in pGL3-Basic(pGL3BV), both of which contain a luciferasereporter gene. The resulting mammalian ex-pression vectors were named EV300, EV600,EV2000, and BV2000. The corresponding pro-moterless constructs were pGL3-Control (desig-nated EV-Control) and pGL3-Basic (BV-Con-trol). The constructs labeled EV (derived frompGL3-Enhanced) differ from those designatedBV (derived from pGL3-Basic) by the presenceof an SV40 enhancer downstream of the re-porter in the enhanced vector. These constructsare represented diagrammatically in Figure 2.
In one experiment, EcoRI-HindIII linkerswere added to the ends of the EcoRI fragmentderived from the PAC clone, the sample wasdigested with HindIII, and the entire fragmentwas cloned into the HindIII site within thepolylinker region of pGL3-Basic and pGL3-Enhanced. Thus, the promoter used containedthe entire EcoRI sequence shown in Figure 1.This promoter construct contained 128 bp ofsequence downstream from the mRNA.
RESULTSCloning and Sequencing the 58 Flanking Region of
the PSMA Gene
In this study two genomic DNA libraries werescreened (a BAC library and a PAC library),and three clones from each library containingthe PSMA gene sequence were identified. A2.1-kb EcoRI restriction fragment located imme-diately upstream of the PSMA gene was identi-fied from one of the clones from each libraryand was subcloned. The sequences of the pro-moter fragment derived from the BAC libraryand the fragment derived from the PAC libraryare depicted in Figure 1, and the positions ofthe EcoRI restriction sites are shown. Differ-ences between the two sequences are markedas shaded nucleotides in Figure 1. Nucleotide86 in the PAC derivative is A rather than T.Nucleotide 642 in the PAC derivative is A ratherthan G. In the PAC sequence, the G at nucleo-tide 687 is omitted. These minor differencesmay be the result of cloning artifacts or sequenc-ing errors, but more likely they are due topolymorphism within the human PSMA gene.An arrow indicates the transcription start siteat position 1898 based on the start site of thepublished PSMA cDNA sequence. We have notidentified an associated TATA box. Although
398 Good et al.

there are multiple potential TATA boxes in thepromoter, they are all located further up-stream.
The Cloned PSMA Promoter Is Not Derived Fromthe PSMA Pseudogene
Work performed by Leek et al. [1995] indi-cates the existence of a PSMA pseudogene,which was mapped in close proximity to the
authentic PSMA gene on chromosome 11. Todetermine whether the isolated BAC clone con-tained the pseudogene or the authentic ge-nomic PSMA sequence, Southern blot analysisusing radiolabeled PSMA cDNA as a probe wasperformed on BglI-digested BAC DNAand BglI-digested PSMA cDNA. The digested BAC ge-nomic DNAsamples produced hybridizing bandsconsiderably larger than fragments obtained
Fig. 1. Sequence of the prostate-specific membrane antigen(PSMA) promoter. The cloned promoter derived from the BAClibrary (Genbank accession number AF044684) and from thePAC library (Genbank accession number AF061571) are identi-cal, except for single base changes at positions 85, 642, and 687(highlighted) (see under Results, for description). The mRNAstart site based on the published cDNA sequence is indicated by
an arrow at position 1898. The positions of the three polymerasechain reaction (PCR) primers used to produce the promoterdeletion constructs are double underlined at positions starting at167, 1468, and 1859. The lower PCR primer is single under-lined at 2061. The EcoRI sites are underlined at positions 1 and2020 and the ATG start codon is underlined at position 2159.
Characterization of the PSMA Promoter 399

from the PSMA cDNA digest (data not shown),because of introns present within the authenticPSMA gene. These results indicate that theBAC clone contains a copy of the authenticPSMA gene, and not the pseudogene.
Transcriptional Analysis of the PSMA Promoter
Experiments were conducted to examine tran-scription driven by the PSMA promoter. In thefirst experiment, four human cell lines weretransfected by expression vectors using thePSMA promoter derived from the BAC library.The cell lines used include two prostate lines(DU 145, a human prostate cancer line andLNCaP, a human PSMA-expressing prostatecancer line) and two non-prostate lines (HeLa,a human cervical cancer line, and 911, a humanretinoblastoma line). Transcription of a lucifer-ase reporter gene construct was driven by the2-kb clone PSMA promoter fragment (BV2000).This vector and the other expression vectorsused in this experiment are represented dia-grammatically in Figure 2.
A significant level of transcription was pro-vided by the 2-kb PSMA fragment in all celllines tested (Fig. 3A, column 1 (BV2000)). Inthe LNCaP, DU 145, and HeLa cell lines, thePSMA promoter in an expression vector withan SV40 enhancer sequence (column 2 (EV2000)marked with an asterisk) provided a signifi-cantly higher level of transcription comparedwith the PSMA promoter alone in BV2000 (a 2logfold increase in the LNCaP cell line and 1logfold increase in DU 145 cell line) (Fig. 3A,
BV2000 vs EV2000). Reporter gene transcrip-tion provided by these two constructs were rela-tively low compared with transcription pro-vided by a cytomegalovirus (CMV) promoterconstruct that served as a positive control withvery high promoter activity in this assay (Fig.3A, column 3 (CMV)). Similar results were ob-tained in two subsequent experiments.
In another experiment (Figure 3B), severalprostate cell lines (PPC-1, ALVA, PC3, DU 145,PZ, and LNCaP), as well as the non-prostateHeLa cell line, were transfected with luciferaseexpression vectors to examine activity of thePSMA promoter derived from the PAC clone.Five different firefly luciferase expression vec-tors were used in this experiment. Three ofthese were controls (pGL3-Control with an SV40promoter and enhancer, pGL3-Basic with nopromoter or enhancer, and pGL3-Enhanced withno promoter but with an SV40 enhancer), andtwo were test expression vectors (PSMA-pro-moted PSMA-pGL3-Basic, and PSMA-promotedPSMA-pGL3-Enhanced). All cell lines were co-transfected with a Renilla luciferase reporterconstruct as a transfection efficiency controlvector using the dual luciferase assay system.In this experiment, the value of the promoter-less vector was not subtracted, but the datawere normalized by dividing firefly luciferaseactivity by Renilla luciferase activity.
The 2-kb PSMA promoter was active in driv-ing transcription of the luciferase gene in allprostate cell lines tested, as well as in HeLacells (Fig. 3B). Transcription was seen in all celllines with all the expression vectors. The lowtranscriptional activity provided by the promot-erless pGL3-Basic vector represents basal tran-scription of the luciferase gene in the absence ofa functional promoter. The PSMA-promotedPSMA-pGL3-Basic test vector consistently pro-vided much higher transcription levels than didthe promoterless pGL3-Basic control vector. ThePSMA-pGL3-Basic vector promoted transcrip-tion at a higher rate than the control pGL3-Enhanced vector in ALVA, PC3, and LNCaPprostate cell lines. By contrast, PSMA-pGL3-Basic provided similar levels of activity in PPC-1and DU 145 cell lines.
The PSMA promoted PSMA-pGL3-Enhancedvector provided significantly higher transcrip-tion levels than pGL3-Enhanced (without thePSMA promoter) in all cell lines tested exceptthe PZ prostate cell line. The large error in this
Fig. 2. Promoter and control constructs. The luciferase re-porter gene under control of the 2-kb prostate-specific mem-brane antigen (PSMA) promoter sequence is designated BV2000.The same construct with an SV40 enhancer sequence at the 38
end of PSMA promoter is named EV2000. Analogous deletionconstructs with the SV40 enhancer sequence consisting of the614-bp and 232-bp 38 PSMA promoter fragments respectivelyare referred to as EV600 and EV300. The promoterless controlEV-Control is not shown. The CMV lac-Z construct was used forcotransfection to normalize transfection efficiency as describedunder Materials and Methods.
400 Good et al.
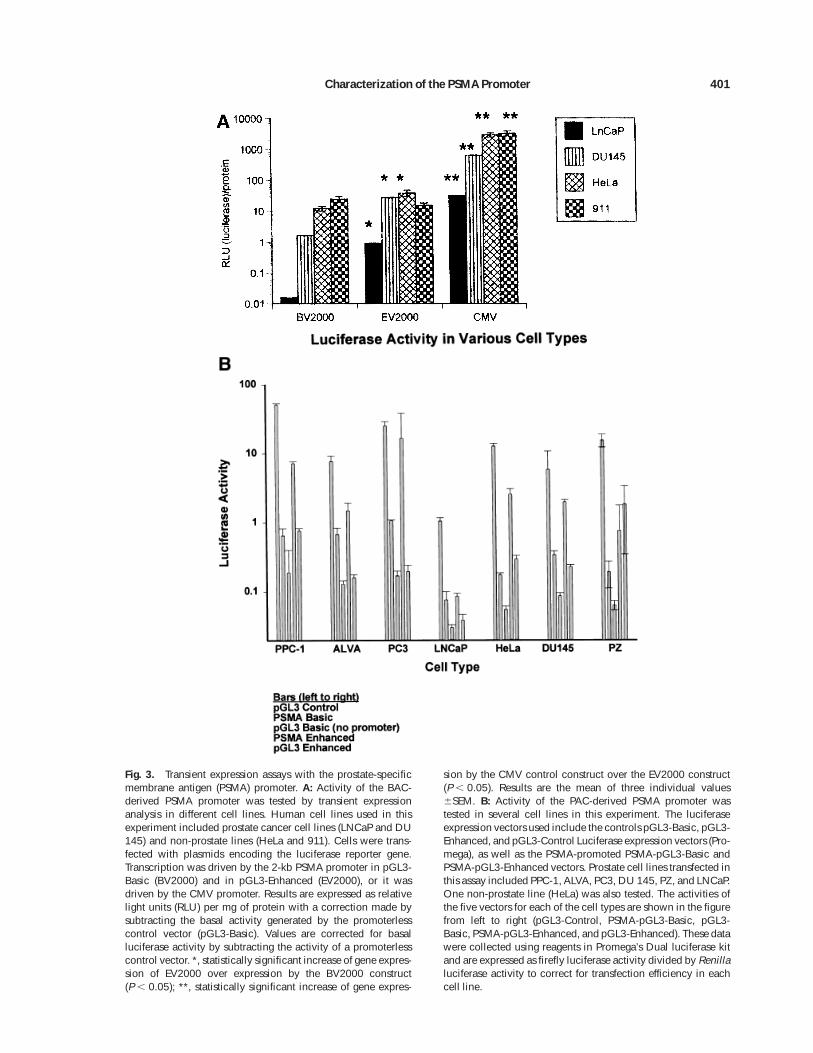
Fig. 3. Transient expression assays with the prostate-specificmembrane antigen (PSMA) promoter. A: Activity of the BAC-derived PSMA promoter was tested by transient expressionanalysis in different cell lines. Human cell lines used in thisexperiment included prostate cancer cell lines (LNCaP and DU145) and non-prostate lines (HeLa and 911). Cells were trans-fected with plasmids encoding the luciferase reporter gene.Transcription was driven by the 2-kb PSMA promoter in pGL3-Basic (BV2000) and in pGL3-Enhanced (EV2000), or it wasdriven by the CMV promoter. Results are expressed as relativelight units (RLU) per mg of protein with a correction made bysubtracting the basal activity generated by the promoterlesscontrol vector (pGL3-Basic). Values are corrected for basalluciferase activity by subtracting the activity of a promoterlesscontrol vector. *, statistically significant increase of gene expres-sion of EV2000 over expression by the BV2000 construct(P , 0.05); **, statistically significant increase of gene expres-
sion by the CMV control construct over the EV2000 construct(P , 0.05). Results are the mean of three individual values6SEM. B: Activity of the PAC-derived PSMA promoter wastested in several cell lines in this experiment. The luciferaseexpression vectors used include the controls pGL3-Basic, pGL3-Enhanced, and pGL3-Control Luciferase expression vectors (Pro-mega), as well as the PSMA-promoted PSMA-pGL3-Basic andPSMA-pGL3-Enhanced vectors. Prostate cell lines transfected inthis assay included PPC-1, ALVA, PC3, DU 145, PZ, and LNCaP.One non-prostate line (HeLa) was also tested. The activities ofthe five vectors for each of the cell types are shown in the figurefrom left to right (pGL3-Control, PSMA-pGL3-Basic, pGL3-Basic, PSMA-pGL3-Enhanced, and pGL3-Enhanced). These datawere collected using reagents in Promega’s Dual luciferase kitand are expressed as firefly luciferase activity divided by Renillaluciferase activity to correct for transfection efficiency in eachcell line.
Characterization of the PSMA Promoter 401
PZ data set does not allow us to see a significantdifference in PSMA-pGL3-Enhanced versuspGL3-Enhanced. The data shown in Figure 3Bconfirm that the PSMA promoter drives expres-sion in both prostate and non-prostate cell linesin transient expression assays.
We conclude that the cloned PSMA promot-ers derived from both the BAC and PAC clonesare functional in six prostate and two nonpros-tate cell lines. When the PSMA promoter isplaced in a vector with an SV40 enhancer se-quence (PSMA-pGL3-Enhanced), the transcrip-tional activity is elevated compared with tran-scription using vector without an SV40enhancer sequence (PSMA-pGL3-Basic).
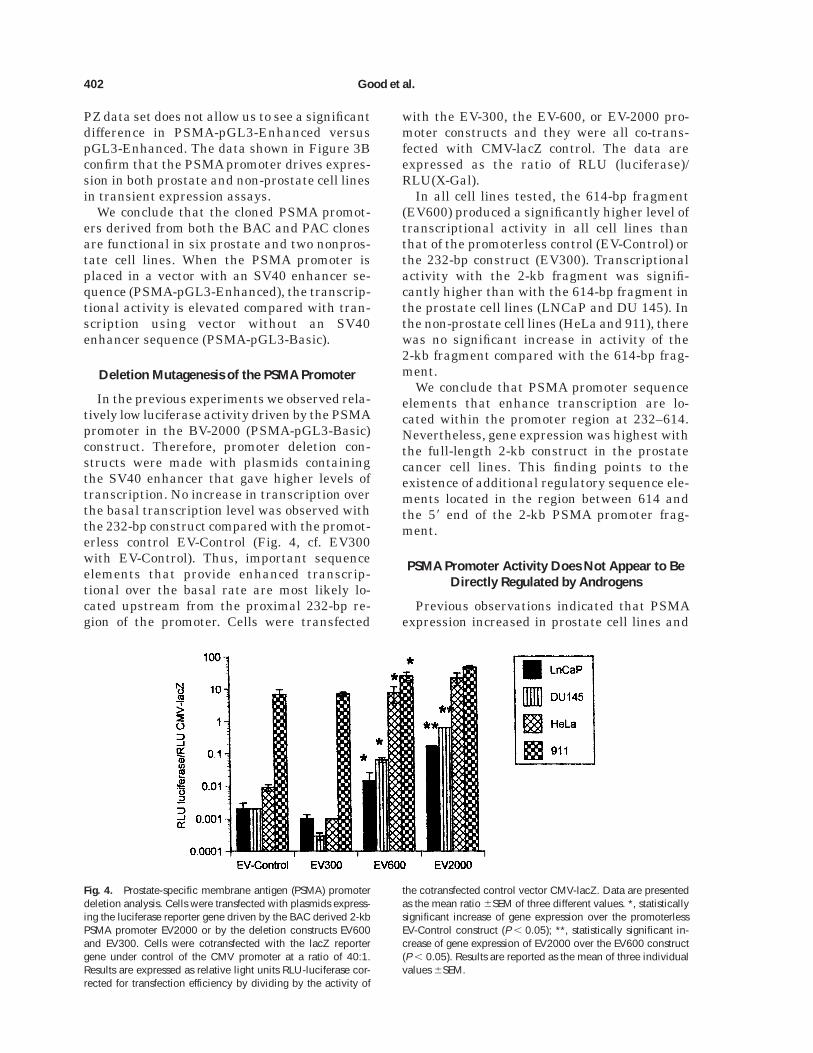
Deletion Mutagenesis of the PSMA Promoter
In the previous experiments we observed rela-tively low luciferase activity driven by the PSMApromoter in the BV-2000 (PSMA-pGL3-Basic)construct. Therefore, promoter deletion con-structs were made with plasmids containingthe SV40 enhancer that gave higher levels oftranscription. No increase in transcription overthe basal transcription level was observed withthe 232-bp construct compared with the promot-erless control EV-Control (Fig. 4, cf. EV300with EV-Control). Thus, important sequenceelements that provide enhanced transcrip-tional over the basal rate are most likely lo-cated upstream from the proximal 232-bp re-gion of the promoter. Cells were transfected
with the EV-300, the EV-600, or EV-2000 pro-moter constructs and they were all co-trans-fected with CMV-lacZ control. The data areexpressed as the ratio of RLU (luciferase)/RLU(X-Gal).
In all cell lines tested, the 614-bp fragment(EV600) produced a significantly higher level oftranscriptional activity in all cell lines thanthat of the promoterless control (EV-Control) orthe 232-bp construct (EV300). Transcriptionalactivity with the 2-kb fragment was signifi-cantly higher than with the 614-bp fragment inthe prostate cell lines (LNCaP and DU 145). Inthe non-prostate cell lines (HeLa and 911), therewas no significant increase in activity of the2-kb fragment compared with the 614-bp frag-ment.
We conclude that PSMA promoter sequenceelements that enhance transcription are lo-cated within the promoter region at 232–614.Nevertheless, gene expression was highest withthe full-length 2-kb construct in the prostatecancer cell lines. This finding points to theexistence of additional regulatory sequence ele-ments located in the region between 614 andthe 58 end of the 2-kb PSMA promoter frag-ment.
PSMA Promoter Activity Does Not Appear to BeDirectly Regulated by Androgens
Previous observations indicated that PSMAexpression increased in prostate cell lines and
Fig. 4. Prostate-specific membrane antigen (PSMA) promoterdeletion analysis. Cells were transfected with plasmids express-ing the luciferase reporter gene driven by the BAC derived 2-kbPSMA promoter EV2000 or by the deletion constructs EV600and EV300. Cells were cotransfected with the lacZ reportergene under control of the CMV promoter at a ratio of 40:1.Results are expressed as relative light units RLU-luciferase cor-rected for transfection efficiency by dividing by the activity of
the cotransfected control vector CMV-lacZ. Data are presentedas the mean ratio 6SEM of three different values. *, statisticallysignificant increase of gene expression over the promoterlessEV-Control construct (P , 0.05); **, statistically significant in-crease of gene expression of EV2000 over the EV600 construct(P , 0.05). Results are reported as the mean of three individualvalues 6SEM.
402 Good et al.
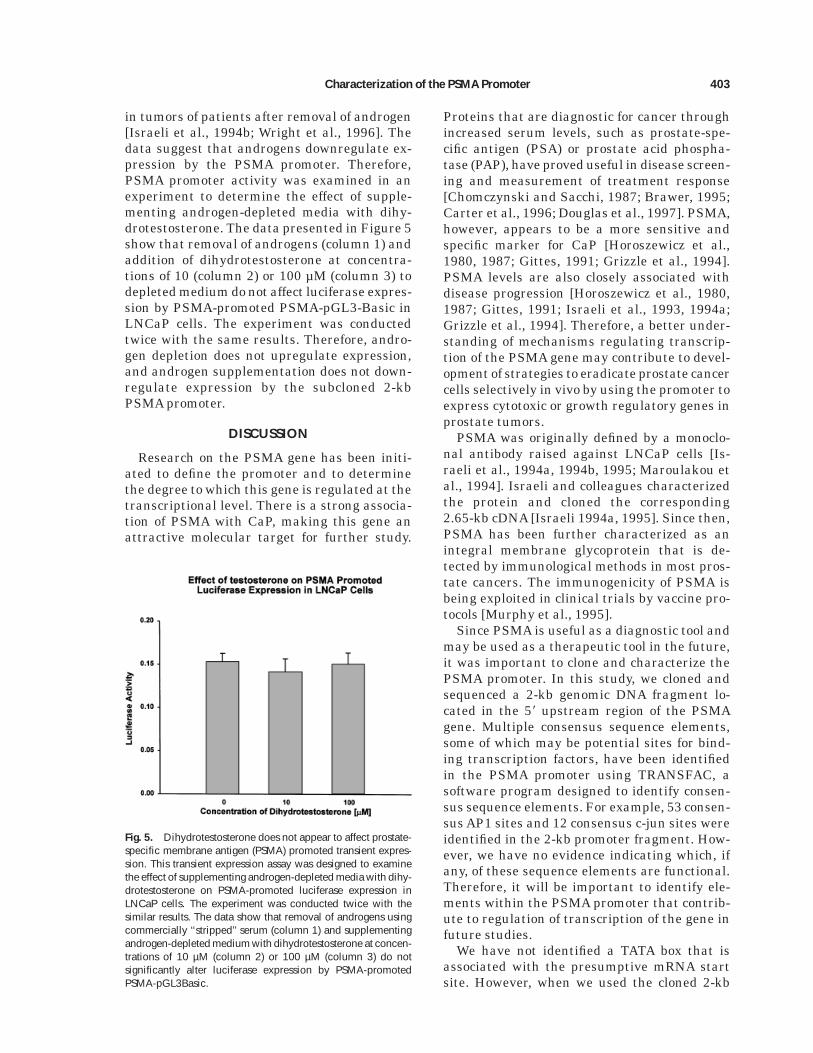
in tumors of patients after removal of androgen[Israeli et al., 1994b; Wright et al., 1996]. Thedata suggest that androgens downregulate ex-pression by the PSMA promoter. Therefore,PSMA promoter activity was examined in anexperiment to determine the effect of supple-menting androgen-depleted media with dihy-drotestosterone. The data presented in Figure 5show that removal of androgens (column 1) andaddition of dihydrotestosterone at concentra-tions of 10 (column 2) or 100 µM (column 3) todepleted medium do not affect luciferase expres-sion by PSMA-promoted PSMA-pGL3-Basic inLNCaP cells. The experiment was conductedtwice with the same results. Therefore, andro-gen depletion does not upregulate expression,and androgen supplementation does not down-regulate expression by the subcloned 2-kbPSMA promoter.
DISCUSSION
Research on the PSMA gene has been initi-ated to define the promoter and to determinethe degree to which this gene is regulated at thetranscriptional level. There is a strong associa-tion of PSMA with CaP, making this gene anattractive molecular target for further study.
Proteins that are diagnostic for cancer throughincreased serum levels, such as prostate-spe-cific antigen (PSA) or prostate acid phospha-tase (PAP), have proved useful in disease screen-ing and measurement of treatment response[Chomczynski and Sacchi, 1987; Brawer, 1995;Carter et al., 1996; Douglas et al., 1997]. PSMA,however, appears to be a more sensitive andspecific marker for CaP [Horoszewicz et al.,1980, 1987; Gittes, 1991; Grizzle et al., 1994].PSMA levels are also closely associated withdisease progression [Horoszewicz et al., 1980,1987; Gittes, 1991; Israeli et al., 1993, 1994a;Grizzle et al., 1994]. Therefore, a better under-standing of mechanisms regulating transcrip-tion of the PSMA gene may contribute to devel-opment of strategies to eradicate prostate cancercells selectively in vivo by using the promoter toexpress cytotoxic or growth regulatory genes inprostate tumors.
PSMA was originally defined by a monoclo-nal antibody raised against LNCaP cells [Is-raeli et al., 1994a, 1994b, 1995; Maroulakou etal., 1994]. Israeli and colleagues characterizedthe protein and cloned the corresponding2.65-kb cDNA [Israeli 1994a, 1995]. Since then,PSMA has been further characterized as anintegral membrane glycoprotein that is de-tected by immunological methods in most pros-tate cancers. The immunogenicity of PSMA isbeing exploited in clinical trials by vaccine pro-tocols [Murphy et al., 1995].
Since PSMA is useful as a diagnostic tool andmay be used as a therapeutic tool in the future,it was important to clone and characterize thePSMA promoter. In this study, we cloned andsequenced a 2-kb genomic DNA fragment lo-cated in the 58 upstream region of the PSMAgene. Multiple consensus sequence elements,some of which may be potential sites for bind-ing transcription factors, have been identifiedin the PSMA promoter using TRANSFAC, asoftware program designed to identify consen-sus sequence elements. For example, 53 consen-sus AP1 sites and 12 consensus c-jun sites wereidentified in the 2-kb promoter fragment. How-ever, we have no evidence indicating which, ifany, of these sequence elements are functional.Therefore, it will be important to identify ele-ments within the PSMA promoter that contrib-ute to regulation of transcription of the gene infuture studies.
We have not identified a TATA box that isassociated with the presumptive mRNA startsite. However, when we used the cloned 2-kb
Fig. 5. Dihydrotestosterone does not appear to affect prostate-specific membrane antigen (PSMA) promoted transient expres-sion. This transient expression assay was designed to examinethe effect of supplementing androgen-depleted media with dihy-drotestosterone on PSMA-promoted luciferase expression inLNCaP cells. The experiment was conducted twice with thesimilar results. The data show that removal of androgens usingcommercially ‘‘stripped’’ serum (column 1) and supplementingandrogen-depleted medium with dihydrotestosterone at concen-trations of 10 µM (column 2) or 100 µM (column 3) do notsignificantly alter luciferase expression by PSMA-promotedPSMA-pGL3Basic.
Characterization of the PSMA Promoter 403

EcoRI fragment as a probe in Northern blotassays, PSMA mRNA of the expected lengthwas detected in total cellular RNA derived fromthe LNCaP cell line but not from DU145 andPC-3 cell line (data not shown). This confirmsthe published pattern of PSMA steady-statemRNA levels in these cell lines [Israeli et al.,1993]. These data also indicate that a portion ofthe mRNA leader region is located within the2-kb EcoRI fragment.
This study demonstrates that the cloned 2-kbpromoter is functional in a number of cell linesby transient transfection assays. Furthermore,the 2-kb PSMA promoter activity increased to agreater degree compared with the 614-bp pro-moter in the presence of an enhancer providedby pGL3-Enhanced in prostate cell lines versusnon-prostate cell lines (Fig. 4). Although theenhanced activity appears to be specific forprostate cell lines, there is nevertheless a rela-tively high level of expression in non-prostatecell lines. Identification of specific promoterelements that provide enhanced transcrip-tional activity in prostate cells requires furtherinvestigation. It was recently reported that a1.2-kb promoter fragment of the PSMA genewas functional in driving expression in prostatecell lines but not in breast-derived cell lines[O’Keefe et al., 1998]. We have not examinedbreast-derived cell lines, but our study showsthat both 614-bp and 2-kb promoter fragmentsare functional in driving expression in severalprostate and at least two non-prostate cell lines.
Previous observations revealed that PSMAexpression increased in prostate tumors of pa-tients after chemical or surgical androgen abla-tion [Wright et al., 1996]. The results of thosestudies suggested that androgens may be in-volved in downregulating transcription by thePSMA promoter. Therefore, we tested the effectof androgen concentration on PSMA promoteractivity in transient expression assays in theLNCaP cell line. We were surprised to find littleor no effect of androgen deprivation or andro-gen supplementation of ‘‘androgen stripped’’ se-rum upon PSMA-promoted transient expres-sion (Fig. 5). It is possible that transcriptionalcontrol elements responsive to androgens arelocated upstream from the cloned 2-kb pro-moter fragment used in this study. If the PSMApromoter is to be used as a tool for gene therapy,upregulation of the promoter when androgenconcentration is lowered may be advantageousover promoters such as the PSA promoter that
are known to be activated by androgens. Uponandrogen ablation, the cloned PSMA promotershould remain active, whereas activities of an-drogen-dependent promoters are reduced. Con-sequently, if PSMA promoter activity is foundto increase in the absence of androgen, thepromoter is potentially even more useful.
In summary, the results of the experimentspresented in this article provide a rationale forcontinued research on the PSMA promoter.There is a need to identify specific sequenceelements that enhance transcription in pros-tate epithelial cells and in prostate cancer cellsand that silence transcription in most non-prostate cells. In the transient expression as-says described in this study, the PSMA pro-moter does not appear to provide enhancedprostate-specific transcription. It is possible thatenhancers and other elements that lead to en-hanced expression in prostate epithelial cellsand in prostate cancer are located farther up-stream or downstream from the 2-kb fragmentexamined in this study. Likewise, androgen-responsive elements within the promoter havenot been identified. Therefore, future experi-ments will be directed toward the identificationof these elements.
ACKNOWLEDGMENTS
The authors thank Dr. Julie Brown of NewOrleans for the critical review of the manu-script. The authors also thank Mr. Everett G.Robert for technical and secretarial assistanceduring the early stages of the project. Thisresearch was supported by the Elsa U. PardeeFoundation (to P.S.), by grants GM20818 (toR.E.R.) and HD29381 (to S.G.) from the Na-tional Institutes of Health, and by the VeteransAdministration (to S.G.).
REFERENCES
Abdel-Nabi HH, Ortman-Nabi JA, See W, Lee J, Ireton R,Boileau M, Unger MW, Halverson C. 1990. Clinical expe-rience with intra lymphatic administration of 111In-labelled monoclonal antibody PAY 276 for the detection ofpelvic nodal metastases in prostatic carcinoma. Eur JNucl Med 16:149–156.
Babaian RJ, Sayer J, Podotoff DA, Steelhammer LC, Bhad-kamkar VA, Gulfo JV. 1994. Radioimmunoscintigraphyof pelvic lymph nodes with 111indium-labeled monoclo-nal antibody CYT-356. J Urol 152(6 Pt 1):1952–1955.
Brawer MK. 1995. How to use prostate-specific antigen inthe early detection or screening for prostatic carcinoma.CA Cancer J Clin 45:148–164.
404 Good et al.

Brothman AR, Lesho LJ, Somers KD, Wright GL, Jr., Mer-chant DJ. 1989. Phenotypic and cytogenetic characteriza-tion of a cell line derived from primary prostatic carci-noma. Int J Cancer 44:898–903.
Carter RE, Feldman AR, Coyle JT. 1996. Prostate-specificmembrane antigen is a hydrolase with substrate andpharmacologic characteristics of a neuropeptidase. ProcNatl Acad Sci USA 93:749–753.
Chomczynski P, Sacchi N. 1987. Single-step method of RNAisolation by acid guanidinium thiocyanate-phenol-chloro-form extraction. Anal Biochem 162:156–159.
Douglas TH, Morgan TO, McLeod DG, Moul JW, MurphyGP, Barren R, 3rd, Sesterhenn IA, Mostofi FK. 1997.Comparison of serum prostate specific membrane anti-gen, prostate specific antigen, and free prostate specificantigen levels in radical prostatectomy patients. Cancer80:107–114.
Gittes RF. 1991. Carcinoma of the prostate [see comments].N Engl J Med 324:236–245.
Grizzle WE, Myers RB, Arnold MM, Srivastava S. 1994.Evaluation of biomarkers in breast and prostate cancer. JCell Biochem Suppl 19:259–266.
Haseman MK, Reed NL, Rosenthal SA. 1996. Monoclonalantibody imaging of occult prostate cancer in patientswith elevated prostate-specific antigen. Positron emis-sion tomography and biopsy correlation. Clin Nucl Med21:704–713.
Heston WD. 1997. Characterization and glutamyl prefer-ring carboxypeptidase function of prostate specific mem-brane antigen: a novel folate hydrolase. Urology 49(3Asuppl):104–112.
Horoszewicz JS, Leong SS, Chu TM, Waisman ZL, Fried-man M, Papsidero L, Kim U, Chai LS, Kakati S, Arya SK,Sandberg AA. 1980. The LNCaP cell line—a new modelfor studies on human prostate carcinoma. Prog Clin BiolRes 37:115–132.
Horoszewicz JS, Kawinski E, Murphy GP. 1987. Monoclo-nal antibodies to a new antigenic marker in epithelialprostatic cells and serum of prostatic cancer patients.Anticancer Res 7:927–935.
Israeli RS, Powell CT, Fair WR, Heston WD. 1993. Molecu-lar cloning of a complementary DNA encoding a prostatespecific membrane antigen. Cancer Res 53:227–230.
Israeli RS, Miller WH Jr, Su SL, Powell CT, Fair WR,Samadi DS, Huryk RF, DeBlasio A, Edwards ET, WiseGJ, et al. 1994a. Sensitive nested reverse transcriptionpolymerase chain reaction detection of circulating pros-tatic tumor cells: comparison of prostate-specific mem-brane antigen and prostate-specific antigen-based as-says. Cancer Res 54:6306–6310.
Israeli RS, Powell CT, Fair WR, Heston WD. 1994b. Expres-sion of the prostate specific membrane antigen. CancerRes 54:1807–1811.
Israeli RS, Miller WH Jr, Su SL, Samadi DS, Powell CT,Heston WD, Wise GJ, Fair WR. 1995. Sensitive detectionof prostatic hematogenous tumor cell dissemination us-ing prostate specific antigen and prostate specific mem-brane-derived primers in the polymerase chain reaction.J Urol 153(3 Pt 1):573–577.
Leek J, Lench N, Maraj B, Bailey A, Carr IM, Andersen S,Cross J, Whelan P, MacLennan KA, Meredith DM, et al.1995. Prostate-specific membrane antigen: evidence forthe existence of a second related human gene. Br JCancer 72:583–588.
Loop SM, Rozanski TA, Ostenson RC. 1993. Human pri-mary prostate tumor cell line, ALVA-31: a new model forstudying the hormonal regulation of prostate tumor cellgrowth. Prostate 22:93–108.
Maroulakou IG, Anver M, Garrett L, Green JE. 1994.Prostate and mammary adenocarcinoma in transgenicmice carrying a rat C3(1) simian virus 40 large tumorantigen fusion gene. Proc Natl Acad Sci USA 91:11236–11240.
Murphy GP. 1995. Radioscintiscanning of prostate cancer.Cancer 75:1819–1822.
Murphy G, Ragde H, Hall MC, Zhao LS, Troncoso P, PollackA, Chung LW. 1995. Comparison of prostate specific mem-brane antigen, and prostate specific antigen levels inprostatic cancer patients. Anticancer Res 15:1473–1479.
Murphy GP, Maguire RT, Rogers B, Partin AW, Nelp WB,Troychak MJ, Ragde H, Kenny GM, Barren RJ, 3rd,Bowes VA, Gregorakis AK, Holmes EH, Boynton AL.1997. Comparison of serum PSMA, PSA levels with re-sults of Cytogen-356 ProstaScint scanning in prostaticcancer patients. Prostate 33:281–285.
O’Keefe DS, Su SL, Bacich DJ, Horiguchi Y, Luo Y, PowellCT, Zandvliet D, Russell PJ, Molloy PL, Nowak NJ,Shows TB, Mullins C, Vonder Haar RA, Fair WR, HestonWDW. 1998. Mapping, genomic organization and pro-moter analysis of the human prostate-specific membraneantigen gene. Biochim Biophys Acta 1443:113–127.
Rinker-Schaeffer CW, Hawkins AL, Su SL, Israeli RS, Grif-fin CA, Isaacs JT, Heston WD. 1995. Localization andphysical mapping of the prostate-specific membrane anti-gen (PSM) gene to human chromosome 11. Genomics30:105–108.
Silver DA, Pellicer I, Fair WR, Heston WDW, Cordon-CardoC. 1997. Prostate-specific membrane antigen expressionin normal and malignant human tissues. Clin CancerRes 3:81–85.
Su SL, Huang IP, Fair WR, Powell CT, Heston WD. 1995.Alternatively spliced variants of prostate-specific mem-brane antigen RNA: ratio of expression as a potentialmeasurement of progression. Cancer Res 55:1441–1443.
Tjoa BA, Erickson SJ, Bowes VA, Ragde H, Kenny GM,Cobb OE, Ireton RC, Troychak MJ, Boynton AL, MurphyGP. 1997. Follow-up evaluation of prostate cancer pa-tients infused with autologous dendritic cells pulsed withPSMA peptides. Prostate 32:272–278.
Troyer JK, Beckett ML, Wright GL. 1995. Detection andcharacterization of the prostate-specific membrane anti-gen (PSMA) in tissue extracts and body fluids. Int JCancer 62:552–558.
Wright GL Jr, Grob BM, Haley C, Grossman K, Newhall K,Petrylak D, Troyer J, Konchuba A, Schellhammer PF,Moriarty R. 1996. Upregulation of prostate-specific mem-brane antigen after androgen-deprivation therapy. Urol-ogy 48:326–334.
Wynant GE, Murphy GP, Horoszewicz JS, Neal CE, CollierBD, Mitchell E, Purnell G, Tyson I, Heal A, Abdel-Nabi H,et al. 1991. Immunoscintigraphy of prostatic cancer: pre-liminary results with 111In-labeled monoclonal antibody7E11-C5.3 (CYT-356). Prostate 18:229–241.
Zhau HE, Pisters LL, Kenny G, Barren R, 3rd, Erickson S,Tjoa B, Boynton A, Holmes E, Gilbaugh J, Douglas T.1994. Biomarkers associated with prostate cancer pro-gression. J Cell Biochem Suppl 19:208–216.
Characterization of the PSMA Promoter 405


















![Cloning reaction - PNAS · technique should also lead to the cloning of antigen-binding ... TGCAGSAGTCWGG)in whichS =CorG, M=AorC, R = A or G, and W= A or T] or VK1BACK [5'-d(GA-](https://img.pdfslide.net/doc/110x75/5b927f2709d3f232708bea67/cloning-reaction-technique-should-also-lead-to-the-cloning-of-antigen-binding.jpg)










