Embed Size (px)
Citation preview

Copyright © Physiologia Plantarum 2000PHYSIOLOGIA PLANTARUM 109: 34–43. 2000Printed in Ireland—all rights reser6ed ISSN 0031-9317
Cloning of Arabidopsis and barley cDNAs encoding HAK potassiumtransporters in root and shoot cells
Francisco Rubio, Guillermo E. Santa-Marıa1 and Alonso Rodrıguez-Navarro*
Departamento de Biotecnologıa, Escuela Tecnica Superior de Ingenieros Agronomos, Uni6ersidad Politecnica de Madrid, ES-28040 Madrid,Spain1Present address: Instituto de In6estigaciones Biotecnologicas, Uni6ersidad Nacional de General San Martin, CC 30, San Martin (1650),Buenos Aires, Argentina*Corresponding author, e-mail: arodrigna6ar–[email protected]
Received 27 October 1999; revised 14 December 1999
Systematic reverse transcription-polymerase chain reaction K+:Rb+:Cs+ selectivity of bacterial and eukaryotic Kup-(RT-PCR) isolations of cDNA fragments using specific HAK transporters are coincident with the selectivity data
given in the literature about alkali cation transport in differentprimers for HAK mRNAs have revealed that plant HAK K+
plant tissues, indicating that HAK transporters may be thetransporters are extensively expressed in shoots and roots. Atleast 13 genes encoding this type of transporter have been most representative plant K+ transporters. Phylogenetic anal-identified in Arabidopsis. Apparently, most plant HAK trans- ysis of the 19 plant translated sequences that belong to this
type of transporter shows that there are 4 different groups. Inporters do not show functional expression in trk1 trk2 yeastgroup I and II there are members in which high-affinity K+ ormutants. In one of them, however, a point mutation increased
the Vmax of transport approximately 10-fold without affecting Rb+ transport activity has been demonstrated. In othergroups this has not been proved. However, present informationthe Km or cation selectivity, suggesting that regulatory prob-
lems or targeting to the plasma membrane are the cause of suggests that all HAK transporters may be K+ transporters.failure for functional expression of this clone in yeast. The
Introduction
In all living cells, there are many cellular components whichare negatively charged at physiological pH values, makingthe presence of equivalent amounts of cations an absoluterequirement for electrical neutrality. Among all the cationiccomponents of the cell, K+ is the most abundant, andextensive substitution of other cations for K+ deeply im-pairs cell physiology, except in the case of Na+ in somehalophytic species. In higher plants the consequence of thisfundamental biological K+ requirement is that K+
availability and K+ uptake in non-root cells are not lessimportant than K+ uptake in roots. Therefore, plant K+
nutrition involves a complex array of physical processes,such as diffusion and mass transfer, as well as a keybiological process, which is transport across plasmamembranes.
The types and functions of plasma membrane trans-porters have been long standing subjects of research for
plant physiologists, not only out of basic biological interest,but also because of its practical applications in the searchfor strategies to correct K+ deficiencies and salinity effectsin agriculture. However, because of the difficulties of re-search in many of the different types of plant cells, thekinetics or electrophysiology of K+ uptake have been exten-sively investigated in roots (Kochian and Lucas 1988), andin the guard cells of stomata (Blatt 1992), whereas a lessextensive number of reports refer to mesophyll (Spalding etal. 1991, Kourie and Goldsmith 1992, Spalding et al. 1992,Spalding and Goldsmith 1993) and aleurone cells (Bush etal. 1988). Although the results of this research suggest thata common K+ transporter for all plant cells, equivalent toanimal Na+, K+-ATPases (Vasilets and Schwarz 1993),does not exist, the presence of inward-rectifying K+ chan-nels has been detected in many types of cells, suggesting thatK+ channels may provide important pathways for long-
Abbre6iations – MS, Murashige and Skoog medium; PCR, polymerase chain reaction; RT, reverse transcription.
Physiol. Plant. 109, 200034

term physiological K+ uptake (Schroeder et al. 1994,Maathuis et al. 1997a). This uptake could occur both athigh and low K+ concentrations (Rodrıguez-Navarro2000).
The kinetic and electrophysiological studies on plant cellshave been recently complemented with the molecularcloning of some transporters. Following a reverse geneticapproach, plant cDNAs encoding K+ transporters arecloned first, and their functions in the plant are later de-duced from the transport characteristics displayed by thetransporters when they are expressed in heterologous sys-tems. The functions may also eventually be determined fromthe effects of mutations that eliminate the protein in theplant (Hirsch et al. 1998). A systematic study of this typeduring the last few years has led to the cloning and charac-terisation of several cDNAs encoding inward K+ channelsexpressed in root (Sentenac et al. 1992, Hirsch et al. 1998) orleaf cells (Anderson et al. 1992, Schachtman et al. 1992, Caoet al. 1995, Muller-Rober et al. 1995, Zimmermann et al.1998), and the K+ transporters HKT1 in wheat (Schacht-man and Schroeder 1994, Rubio et al. 1995), HAK1 inbarley (Santa-Marıa et al. 1997) and KT/KUP in Arabidop-sis (Quintero and Blatt 1997, Fu and Luan 1998, Kim et al.1998). The HAK and KT/KUP transporters belong to thesame family and, although the barley HAK1 transporter isexpressed only in roots (Santa-Marıa et al. 1997) and thefunction of the Arabidopsis KT/KUP transporters has beendiscussed much more in relation to root K+ uptake than tothe uptake in other cells, both northern blot analyses (Fuand Luan 1998, Kim et al. 1998) and cDNA cloning (seeGenBank accession number W43757) have demonstratedthat the KT/KUP transporters are expressed in many partsof the plant. Consistent with this notion, rice cDNAs encod-ing transporters of the KT/HAK/KUP type have beenobtained from panicles and etiolated shoots (GenBank ac-cession numbers S1836, C73883, C72058, C71725).
The expression of KT/HAK/KUP transporters in suchdifferent parts of the plant suggests the possibility thattransporters of this family are present in all plant cells,together with K+ channels. The most interesting questionwith relation to this possibility is that these transporters arerelated to the Kup transporter of Escherichia coli (Bosse-meyer et al. 1989, Schleyer and Bakker 1993) and speciallyto the fungal HAK1 transporters described in Schwan-niomyces occidentalis (Banuelos et al. 1995) and Neurosporacrassa (Haro et al. 1999). The fungal HAK transporters areprobably K+-H+ symporters (Haro et al. 1999), exhibitinga notable concentrative capacity (more than 107), which areexpressed when the cells are K+ starved (Banuelos et al.1995, Haro et al. 1999). These characteristics fulfil therequirements of the ‘active’ K+ uptake that roots need fora competent K+ uptake at low K+ (Schroeder and Fang1991, Kochian and Lucas 1993, Maathuis and Sanders 1993,1994, Walker et al. 1996, Maathuis et al. 1997b), but areapparently unnecessary or even inappropriate, because theygive rise to energy wasting, for cells facing millimolar K+
concentrations. Although this consideration may lead to theconclusion that transporters of the HAK family (from nowon we will use the generic name of HAK for the eukaryoticmembers of the bacterial and eukaryotic Kup-HAK type of
transporters) cannot be universal plant K+ transportersbecause in many plant cells the concentration of K+ in theapoplast is millimolar (Lowen and Satter 1989, Long andWidders 1990, Muhling and Sattelmacher 1997), other factsmust also be taken into account. The first is, obviously, thatnot all the transporters in the Kup-HAK family must behigh-affinity K+-H+ symporters, but more importantly, insome cases non-root plant cells do in fact take up K+ frommicromolar concentrations (Reed and Bonner 1974, Blatt1985), following kinetics very similar to those exhibited byroot cortical and epidermal cells (Smith and Epstein 1964,Osmond and Laties 1968). All this supports that in facthigh-affinity K+ transporters belonging to the HAK familymay be ubiquitous in the plant, coexisting with inwardK+-channels as it has been demonstrated in Elodea densaleaves (Bellando et al. 1995).
As a first effort to clarify the identity and types of theHAK plant K+ transporters, we have made systematicisolations of cDNA fragments from roots and shoots ofdifferent plants. In some cases the full-length cDNAs wereobtained and expressed in yeast. As previously found, theactivity of these transporters in yeast was absent or very low(Quintero and Blatt 1997, Fu and Luan 1998, Kim et al.1998), but in some of them the activity can be enlarged bypoint mutations. To relate the cloned transporters withthose mediating plant transport processes, we have studiedthe capacities of the transporters to discriminate amongK+, Rb+ and Cs+, and compared them with those foundin different plant cells. We also present a phylogenetic studyof all the HAK transporters known at this moment.
Materials and methods
Yeast and bacterial strains and media
The K+ uptake deficient yeast strain WD3 (MAT a ade2ura3 trp1 trk1?::LEU2 trk2?::HIS3) (Haro et al. 1999) wasused for the expression of plant cDNAs. Yeast strains wereroutinely grown on SD media (Sherman 1991) supplementedwith 100 mM K+. For growth experiments at low K+, theammonium-free AP medium (Rodrıguez-Navarro andRamos 1984) was used. Plasmids were propagated in E. coliDH5a.
Plant cDNA isolations
Arabidopsis thaliana (Columbia ecotype) seedlings weregrown in solid Murashige and Skoog (MS) medium for 2weeks and then K+ starved by transferring the seedlings foran additional week to a modified MS medium, in whichammonium was substituted for K+. Barley (Hordeum 6ul-gare cv. Albacete) seedlings were grown in the dark for 7days in 10 mM MES buffer brought to pH 5.5 withCa(OH)2. Total RNA from Arabidopsis or barley roots orshoots was reversed transcribed by using an anchored oligo-dT primer and avian myeloblastosis virus transcriptase(Amersham Pharmacia, Rainham, UK). The reverse tran-scription (RT) products were amplified by the polymerasechain reaction (PCR) with the Expanded high-fidelity PCR
Physiol. Plant. 109, 2000 35

system (Boehringer Mannheim, Mannheim, Germany) us-ing the degenerate sense primer 5%-GAYAAYGG-NGANGGNGGNACNTTYGC-3% and the degenerateantisense primer 5%-AANTGNCCNARRTCNGCRAA-3%deduced from the conserved regions DNG(D/E)GGTFAand FADLGHF, respectively, present in HvHAK1 andHvHAK2 (Santa-Marıa et al. 1997). A full-length cDNAwas obtained for AtHAK5 and HvHAK2 by using the5%/3% rapid amplification of cDNA ends kit (BoehringerMannheim) following the manufacturer’s instructions.PCR products were cloned into the PCR2.1 vector usingthe TA cloning kit (Invitrogen, Carlsbard, USA). Full-length cDNAs were cloned into plasmids pYPGE15(Brunelli and Pall 1993) or pDR195 (Rentsch et al. 1995) forexpression in yeast.
Nucleic acid gel blot hybridisations
DNA gel blot hybridisations were carried out by theSouthern technique as described elsewhere (Sambrook etal. 1989). Probes were prepared by labelling cDNA frag-ments obtained from PCR with a-32P-dATP by the ran-dom priming method. Membranes (Hybond-N; AmershamPharmacia) were hybridised at 42°C in the presence of50% formamide for 24 h, washed twice in 2× SSC (1×SSC is 0.15 M NaCl, 0.015 M sodium citrate) plus 0.1%sodium dodecyl sulphate (SDS) at 42°C for 30 min andexposed at −72°C for 10 days to Curix RP-2 (Agfa,Mortsel, Belgium) films.
For RNA gel blot hybridisations, Arabidopsis totalRNA was isolated from roots or shoots of plants grownin regular MS medium and then maintained in the samemedium or exposed to the K+-free medium describedabove. Total RNA (30 mg per lane) was separated byelectrophoresis on a formaldehyde-1.1% agarose gel andtransferred to a nylon membrane (Hybond-N; AmershamPharmacia). Membranes were hybridised overnight at42°C in the presence of 50% formamide with probes pre-pared as described above. Membranes were washed twicein 2× SSC plus 0.1% SDS at 42°C, then twice in 0.2×SSC plus 0.1% SDS at 65°C, and exposed as describedabove.
Cation uptake experiments
Yeast cells were grown overnight at 28°C in AP mediumsupplemented with 30 mM K+ and then K+ starved inK+-free AP medium for 6 h. Cells were suspended inuptake buffer containing 2% glucose and 10 mM MESbrought to pH 6.0 with Ca(OH)2. For Rb+ uptake experi-ments, 86Rb+ was added at time 0 to obtain a molaractivity of 40–1500 GBq mol−1 and then samples weretaken at different times, filtered through 0.8 mm Milliporemembranes and washed with a 10 mM RbCl solution.Radioactivity retained in the filters was determined by scin-tillation counting and the Rb+ accumulated in the cells wascalculated from the cellular and external radioactivity ofeach experiment. For Cs+ uptake experiments, Cs+ wasadded at time 0 and cell samples were taken at different
times, filtered through 0.8 mm Millipore membranes andwashed with a 20 mM MgCl2 solution. Filters were incu-bated overnight in a 0.1 M HCl solution and the cellularCs+ was determined by flame emission spectrophotometry.The reported initial rates of uptake are the means of at least3 independent experiments. Error bars denote standarddeviations.
Athak5-1 and H7hak1-1 cDNAs isolation
To isolate spontaneous mutants of the AtHAK5 transporterthat conferred capacity to grow at micromolar K+ concen-trations, 5 plates (150 mm diameter) containing AP mediumsupplemented with 100 mM K+ were inoculated with 108
WD3(AtHAK5) cells per plate and incubated for 4 days at28°C.
To construct a HvHAK1 mutant in which a histidinesubstituted for the Leu764, which corresponds to the con-served Leu776 mutated in Athak5-1, a mutated partial-length cDNA of H6HAK1 was obtained by PCRamplification using an internal sense primer and a 39-merantisense primer starting 6 bases beyond the stop codon.This primer extended over the codon corresponding toLeu764, containing two base substitutions in order tochange the leucine codon (CTG) into a histidine codon(CAT). The mutated fragment was substituted for the corre-sponding fragment in the original clone.
Protein alignments and phylogenetic tree generation
Protein sequence alignments and phylogenetic trees wereobtained by using the Clustal X program (Thompson 1997).
Accession numbers for the reported sequences
AtHAK5: AF129478; AtHAK6: AF129482; AtHAK7:AF129481; AtHAK8: AF129483; HvHAK1B: AF129480;HvHAK2: AF129479; HvHAK3: AF130340; HvHAK4:AF129484; OsHAK4: AF129485
Results
Several genes of the HAK family are expressed in roots andshoots
We made systematic RT-PCR isolations of plant cDNAfragments belonging to the HAK gene family. Using thespecific primers described in Materials and methods, weisolated 6 different fragments from Arabidopsis. Two ofthem were only isolated from roots and the other 4 wereisolated from roots and shoots. As indicated in Table 1,two isolations (AtKT1/AtKUP1 and AtKT2/AtKUP2) co-incided with cDNAs already isolated, two (AtHAK5 andAtHAK6) coincided with genomic sequences in the data-base and two (AtHAK7, and AtHAK8) were new isola-tions. A comparison of the translated sequences of thenew isolations with already known sequences of the HAKfamily of transporters showed that one of the isolations(AtHAK5) could encode a transporter from roots highly
Physiol. Plant. 109, 200036

similar to the HvHAK1 transporter from barley roots.Another isolation (AtHAK8) could encode a transportersimilar to AtKT2/AtKUP2, and the translation of the thirdone (AtHAK7) showed similarity to the translation of anArabidopsis genomic sequence (CAA20566, see Fig. 3).
Following a similar approach, we isolated 3 new frag-ments from barley to be added to the previously describedH6HAK1 and H6HAK2 cDNAs (Santa-Marıa et al. 1997).The translation of one of them, isolated only from roots,showed more than 90% similarity to HvHAK1 and wasnamed HvHAK1B. Another one was isolated from bothshoots and roots and was named H6HAK3. The third,H6HAK4, was isolated from shoots only.
Southern blot hybridisations demonstrated that the iso-lated cDNA fragments strongly hybridised to fragments ofgenomic DNA of the corresponding species (not shown).
Northern blot hybridisations for the transcripts corre-sponding to the most frequently isolated ArabidopsiscDNAs, AtHAK5, AtHAK6 and AtHAK8, detected only theAtHAK5 transcript. In K+-starved Arabidopsis plants, theAtHAK5 transcript was expressed in both roots and shoots,and roots showed higher levels of expression than shoots. InArabidopsis plants grown in the presence of K+, theAtHAK5 transcript was detected in roots but not in shoots.Remarkably, the roots of K+-grown plants exhibited highercontents of AtHAK5 mRNA than those from K+-starvedplants (Fig. 1).
Identification of DNA sequences encoding plant HAKtransporters
The non-redundant nucleotide sequence databases and ESTsequences in the GenBank were screened for translatedsequences homologous to the so-far described HAK trans-porters, and several entries were found corresponding toArabidopsis genomic DNAs or cDNAs and rice cDNAs. InArabidopsis, 5 novel genomic DNA sequences (accessionnumbers AAC24049, CAA20566, CAA16604, AAD21693and AC004238) and two genomic DNA sequences corre-sponding to the cDNAs of AtHAK5 and AtHAK6 describedin the present work (accession numbers CAB40777 and
Fig. 1. Northern blot analysis of AtHAK5 transcript. Arabidopsistotal RNA was isolated from roots (R) or shoots (S) of plantsgrown in regular medium and then maintained in the same medium(+K+) or exposed to a K+-free medium (−K+). Upper panel,detection of AtHAK5 mRNA by hybridising with a radiolabelledAtHAK5 probe. Lower panel, ethidium bromide staining of the gelcontaining the RNA used for the northern analysis.
AAC18809, respectively) were identified. In Table 2, werecorded all the transporters or putative transporters of theHAK family in Arabidopsis. In rice, 4 cDNAs were iden-tified (accession numbers S1836, C73883, C72058, C71725).One of the rice cDNAs, C71725, was sequenced and foundto be a non full-length cDNA.
Phylogenetic analysis of the HAK family of transporters
The novel HAK transporters described in this paper showthe structural features characteristic of the bacterial andeukaryotic Kup-HAK transporters (Rodrıguez-Navarro2000). Twelve transmembrane fragments can be predicted inall of them and variable-length C- and N-termini (Fig. 2).Using different fragments of the translated sequences of theHAK DNAs or cDNAs, different phylogenetic trees wereproduced, which, in general, were very similar. Among thesephylogenetic trees, the one produced with the fragments thatwe amplified in the RT-PCR experiments was as reliable asthe others. This tree is shown in Fig. 3, and reveals aconsiderable divergence among plant HAK transporters.Four clusters of plant transporters can be distinguished(marked as I, II, III and IV in Fig. 3). Cluster I includes thehigh-affinity transporters HvHAK1A from barley (Santa-Marıa et al. 1997) and AtHAK5 from Arabidopsis (de-scribed in the present work), 3 sequences from Arabidopsisand two from barley. Cluster II includes the high-affinitytransporter KT1/KUP1 from Arabidopsis (Quintero andBlatt 1997, Fu and Luan 1998, Kim et al. 1998), theKT2/KUP2 transporter from Arabidopsis, 4 sequences fromArabidopsis and two from barley. Cluster III contains 3genomic sequences from Arabidopsis and cluster IV holdsthe translated sequence of an incomplete cDNA clone fromrice. A partial sequence in the database from a rice cDNA(accession number C72058) was not included in the treeshown in Fig. 3 because the available sequence does notcover the entire fragment used to construct the tree. How-ever, the analysis of the available sequence indicates that itbelongs to cluster II.
Table 1. cDNA fragments isolated from Arabidopsis root andshoot mRNAs by RT-PCR. Total RNA isolated from roots orshoots of plants grown in regular medium and then exposed to aK+-free medium was reverse transcribed and amplified by PCR,using primers designed from conserved amino acid stretches ofHAK transporters. The sequences of 51 cDNA fragments amplifiedby this procedure showed that they could encode this type oftransporters. Accession numbers are given for sequences alreadypresent in the Genbank. 1 Genomic sequences in data base, seecDNA accession number in Materials and methods.
CDNA Accession no.Number ofOrganisolationsroot/shoot
AtHAK5 Roots 8/0 CAB407771
AtHAK6 Roots/shoots 8/2 AAC188091
2/0AtHAK7 Roots The present workRoots/shootsAtHAK8 10/5 The present work
AtKT1/AtKUP1 Roots/shoots 3/3 AF012656AtKT2/AtKUP2 Roots/shoots 2/8 AF012657
Physiol. Plant. 109, 2000 37
Table 2. KT/KUP/HAK transporters or putative transporters of this family encoded by cDNA or genomic sequences in Arabidopsis.1 Because the sequences of KUP2, KUP3 and KUP4 have not been published it is not clear that they exactly correspond to KT2, KT3 andKT4. 2 Corresponds to AtHAK1 in reference (Rodrıguez-Navarro 2000).
Name Genomic sequencecDNA sequence Transporter or cDNA referenceaccession no. accession no.
KT1/KUP1/KUP1 Quintero and Blatt 1997, Kim et al. 1998,AF012656/AF029876/AF033118 AAC16965/AAC31846Fu and Luan 1998
KT2/KUP21 AF012657 AC002336 Quintero and Blatt 1997, Kim et al. 1998KT3/KUP31 AF012658 Quintero and Blatt 1997, Kim et al. 1998KT4/KUP41 AF012659 Quintero and Blatt 1997, Kim et al. 1998HAK52 The present workAF129478 CAB40777HAK6 AF129482 AAC18809 The present workHAK7 AF129481 The present work
The present workHAK8 AF129483AAD21693AC004238CAA16604CAA20566AAC24049
Only some plant HAK transporters are functional in yeast
Two plant transporters belonging to the HAK group,HvHAK1 (Santa-Marıa et al. 1997) and AtKUP1(Fu andLuan 1998, Kim et al. 1998), mediate high-affinity uptake,but nothing is known about other transporters of thisgroup. We were able to obtain two full-length cDNAs, onefrom Arabidopsis, AtHAK5, and another from barley,
H6HAK2. These cDNAs were inserted in yeast expressionvectors under the control of strong constitutive yeast pro-moters. The resulting plasmids were then transformed into thetrk1 trk2 mutant of Saccharomyces cere6isiae. Detailedgrowth tests of the transformants in many different conditionsshowed that the plant cDNAs did not have an appreciableeffect on the growth of the transformants at K+ concentra-tions below 5 mM.
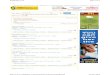
Fig. 2. A model of bacterial and eukaryotic Kup-HAK transporters. The model shows the conserved amino acids in bacterial, fungal andplant Kup-HAK transporters, and those conserved only in plant transporters. Only complete sequences were used. The length of the Nterminus preceding the first transmembrane fragment shows great variability in Kup-HAK transporters, and the length of the N terminusshown in the figure is representative of several plant transporters.
Physiol. Plant. 109, 200038
Appreciable growth of any yeast strain requires a mini-mum K+ uptake of approximately 0.5–1.0 nmol mg−1
min−1, and in a trk1 trk2 mutant transformed with anempty plasmid, the intrinsic (low-affinity) transporter medi-ates this rate of uptake at 2–5 mM external K+. Therefore,for a heterologous transporter to produce a significant en-hancement of the growth rate of a trk1 trk2 mutant, itshould mediate an uptake of at least 0.5 nmol mg−1 min−1
at 2–5 mM K+. At higher K+ concentrations higher rateswould be necessary because the trk1 trk2 mutant increasesits intrinsic uptake rate proportionally to the K+ content ofthe medium, as a consequence of the high-Vmax, high-Km
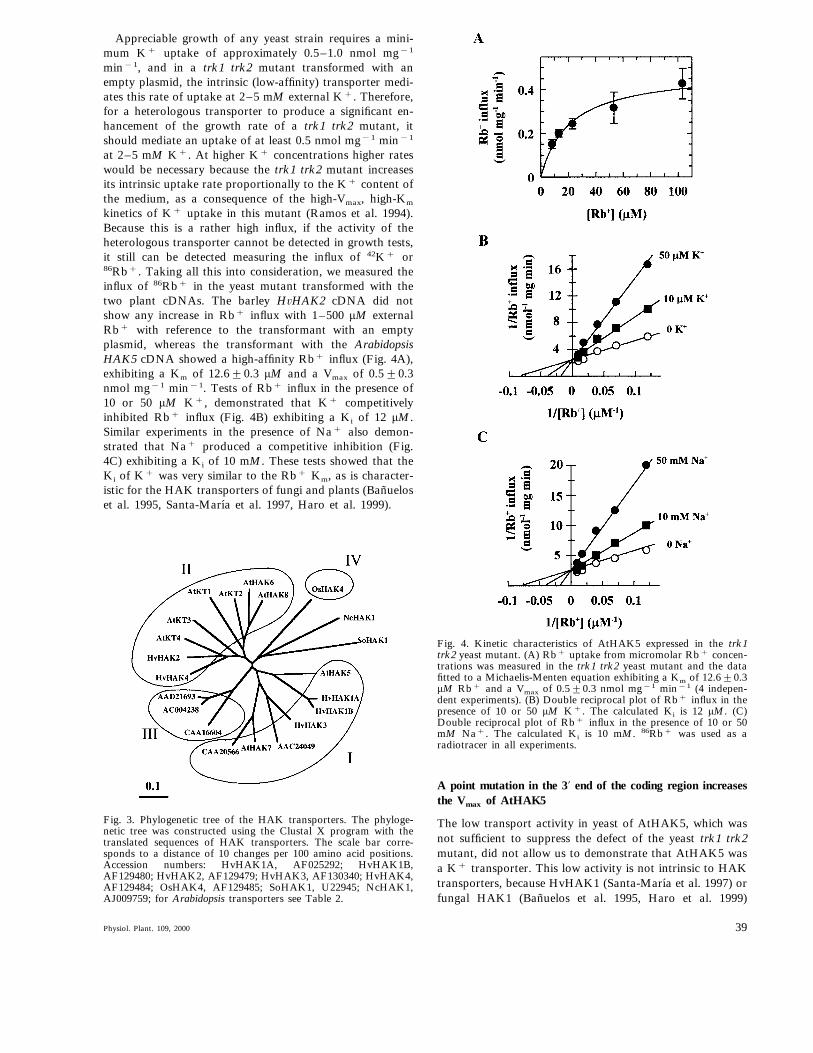
kinetics of K+ uptake in this mutant (Ramos et al. 1994).Because this is a rather high influx, if the activity of theheterologous transporter cannot be detected in growth tests,it still can be detected measuring the influx of 42K+ or86Rb+. Taking all this into consideration, we measured theinflux of 86Rb+ in the yeast mutant transformed with thetwo plant cDNAs. The barley H6HAK2 cDNA did notshow any increase in Rb+ influx with 1–500 mM externalRb+ with reference to the transformant with an emptyplasmid, whereas the transformant with the ArabidopsisHAK5 cDNA showed a high-affinity Rb+ influx (Fig. 4A),exhibiting a Km of 12.690.3 mM and a Vmax of 0.590.3nmol mg−1 min−1. Tests of Rb+ influx in the presence of10 or 50 mM K+, demonstrated that K+ competitivelyinhibited Rb+ influx (Fig. 4B) exhibiting a Ki of 12 mM.Similar experiments in the presence of Na+ also demon-strated that Na+ produced a competitive inhibition (Fig.4C) exhibiting a Ki of 10 mM. These tests showed that theKi of K+ was very similar to the Rb+ Km, as is character-istic for the HAK transporters of fungi and plants (Banueloset al. 1995, Santa-Marıa et al. 1997, Haro et al. 1999).
Fig. 4. Kinetic characteristics of AtHAK5 expressed in the trk1trk2 yeast mutant. (A) Rb+ uptake from micromolar Rb+ concen-trations was measured in the trk1 trk2 yeast mutant and the datafitted to a Michaelis-Menten equation exhibiting a Km of 12.690.3mM Rb+ and a Vmax of 0.590.3 nmol mg−1 min−1 (4 indepen-dent experiments). (B) Double reciprocal plot of Rb+ influx in thepresence of 10 or 50 mM K+. The calculated Ki is 12 mM. (C)Double reciprocal plot of Rb+ influx in the presence of 10 or 50mM Na+. The calculated Ki is 10 mM. 86Rb+ was used as aradiotracer in all experiments.
Fig. 3. Phylogenetic tree of the HAK transporters. The phyloge-netic tree was constructed using the Clustal X program with thetranslated sequences of HAK transporters. The scale bar corre-sponds to a distance of 10 changes per 100 amino acid positions.Accession numbers: HvHAK1A, AF025292; HvHAK1B,AF129480; HvHAK2, AF129479; HvHAK3, AF130340; HvHAK4,AF129484; OsHAK4, AF129485; SoHAK1, U22945; NcHAK1,AJ009759; for Arabidopsis transporters see Table 2.
A point mutation in the 3% end of the coding region increasesthe Vmax of AtHAK5
The low transport activity in yeast of AtHAK5, which wasnot sufficient to suppress the defect of the yeast trk1 trk2mutant, did not allow us to demonstrate that AtHAK5 wasa K+ transporter. This low activity is not intrinsic to HAKtransporters, because HvHAK1 (Santa-Marıa et al. 1997) orfungal HAK1 (Banuelos et al. 1995, Haro et al. 1999)
Physiol. Plant. 109, 2000 39
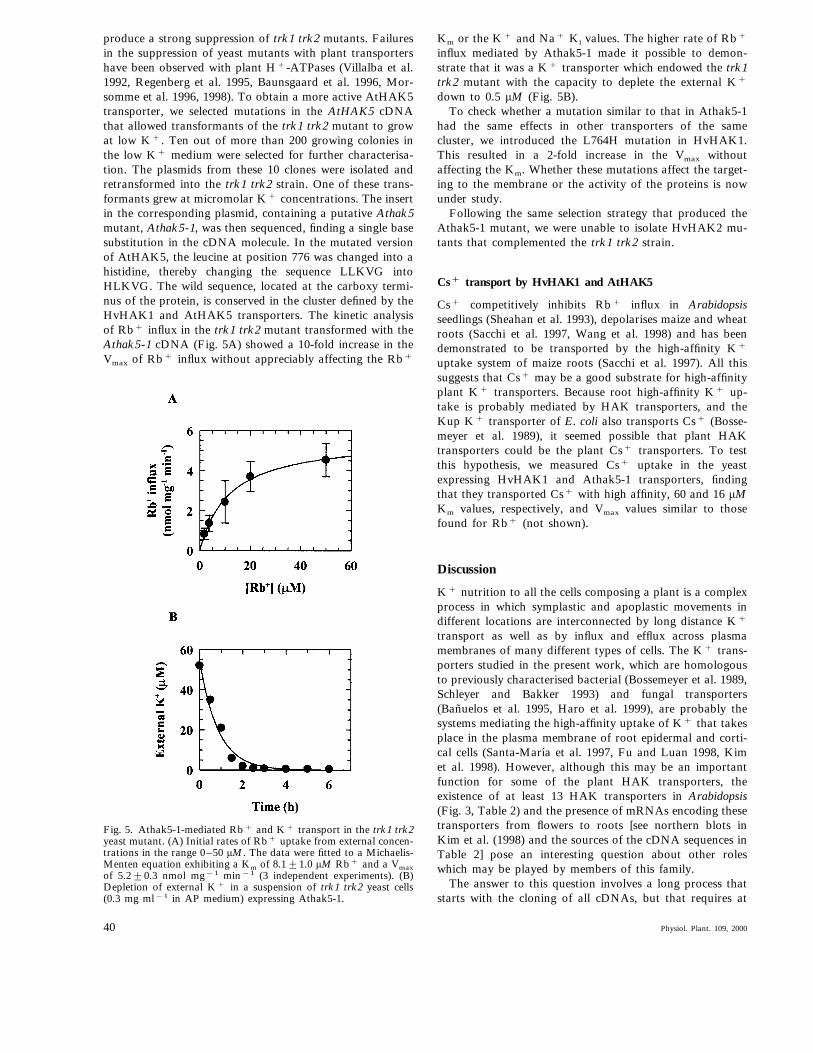
produce a strong suppression of trk1 trk2 mutants. Failuresin the suppression of yeast mutants with plant transportershave been observed with plant H+-ATPases (Villalba et al.1992, Regenberg et al. 1995, Baunsgaard et al. 1996, Mor-somme et al. 1996, 1998). To obtain a more active AtHAK5transporter, we selected mutations in the AtHAK5 cDNAthat allowed transformants of the trk1 trk2 mutant to growat low K+. Ten out of more than 200 growing colonies inthe low K+ medium were selected for further characterisa-tion. The plasmids from these 10 clones were isolated andretransformed into the trk1 trk2 strain. One of these trans-formants grew at micromolar K+ concentrations. The insertin the corresponding plasmid, containing a putative Athak5mutant, Athak5-1, was then sequenced, finding a single basesubstitution in the cDNA molecule. In the mutated versionof AtHAK5, the leucine at position 776 was changed into ahistidine, thereby changing the sequence LLKVG intoHLKVG. The wild sequence, located at the carboxy termi-nus of the protein, is conserved in the cluster defined by theHvHAK1 and AtHAK5 transporters. The kinetic analysisof Rb+ influx in the trk1 trk2 mutant transformed with theAthak5-1 cDNA (Fig. 5A) showed a 10-fold increase in theVmax of Rb+ influx without appreciably affecting the Rb+
Km or the K+ and Na+ Ki values. The higher rate of Rb+
influx mediated by Athak5-1 made it possible to demon-strate that it was a K+ transporter which endowed the trk1trk2 mutant with the capacity to deplete the external K+
down to 0.5 mM (Fig. 5B).To check whether a mutation similar to that in Athak5-1
had the same effects in other transporters of the samecluster, we introduced the L764H mutation in HvHAK1.This resulted in a 2-fold increase in the Vmax withoutaffecting the Km. Whether these mutations affect the target-ing to the membrane or the activity of the proteins is nowunder study.
Following the same selection strategy that produced theAthak5-1 mutant, we were unable to isolate HvHAK2 mu-tants that complemented the trk1 trk2 strain.
Cs+ transport by HvHAK1 and AtHAK5
Cs+ competitively inhibits Rb+ influx in Arabidopsisseedlings (Sheahan et al. 1993), depolarises maize and wheatroots (Sacchi et al. 1997, Wang et al. 1998) and has beendemonstrated to be transported by the high-affinity K+
uptake system of maize roots (Sacchi et al. 1997). All thissuggests that Cs+ may be a good substrate for high-affinityplant K+ transporters. Because root high-affinity K+ up-take is probably mediated by HAK transporters, and theKup K+ transporter of E. coli also transports Cs+ (Bosse-meyer et al. 1989), it seemed possible that plant HAKtransporters could be the plant Cs+ transporters. To testthis hypothesis, we measured Cs+ uptake in the yeastexpressing HvHAK1 and Athak5-1 transporters, findingthat they transported Cs+ with high affinity, 60 and 16 mMKm values, respectively, and Vmax values similar to thosefound for Rb+ (not shown).
Discussion
K+ nutrition to all the cells composing a plant is a complexprocess in which symplastic and apoplastic movements indifferent locations are interconnected by long distance K+
transport as well as by influx and efflux across plasmamembranes of many different types of cells. The K+ trans-porters studied in the present work, which are homologousto previously characterised bacterial (Bossemeyer et al. 1989,Schleyer and Bakker 1993) and fungal transporters(Banuelos et al. 1995, Haro et al. 1999), are probably thesystems mediating the high-affinity uptake of K+ that takesplace in the plasma membrane of root epidermal and corti-cal cells (Santa-Marıa et al. 1997, Fu and Luan 1998, Kimet al. 1998). However, although this may be an importantfunction for some of the plant HAK transporters, theexistence of at least 13 HAK transporters in Arabidopsis(Fig. 3, Table 2) and the presence of mRNAs encoding thesetransporters from flowers to roots [see northern blots inKim et al. (1998) and the sources of the cDNA sequences inTable 2] pose an interesting question about other roleswhich may be played by members of this family.
The answer to this question involves a long process thatstarts with the cloning of all cDNAs, but that requires at
Fig. 5. Athak5-1-mediated Rb+ and K+ transport in the trk1 trk2yeast mutant. (A) Initial rates of Rb+ uptake from external concen-trations in the range 0–50 mM. The data were fitted to a Michaelis-Menten equation exhibiting a Km of 8.191.0 mM Rb+ and a Vmaxof 5.290.3 nmol mg−1 min−1 (3 independent experiments). (B)Depletion of external K+ in a suspension of trk1 trk2 yeast cells(0.3 mg ml−1 in AP medium) expressing Athak5-1.
Physiol. Plant. 109, 200040

least the determination of the kinetics and cation selectivityof the encoded transporter in a convenient system, and thecellular and tissue location of the protein. This will takeseveral years of research. However, the fact that all of theHAK transporters characterised up to now from bacteria toplants show K+ uptake activity suggests that most of themembers of the family are K+ transporters. The existenceof plant cDNAs encoding HAK transporters, which do notshow K+ transport activity in S. cere6isiae, does not neces-sarily mean that they are not K+ transporters, and theirfailure to do so may be explained in two ways: either theregulation of the transporters is not recognised by yeast orthe protein is not targeted to the plasma membrane. Forboth explanations there is extensive information regardingthe expression in yeast of the plant H+-ATPase, in whichpoint mutations make the plant enzyme active in yeast(Villalba et al. 1992, Regenberg et al. 1995, Baunsgaard etal. 1996, Morsomme et al. 1996, 1998), as we found withAtHAK5.
Regarding the kinetic characteristics of the HAK trans-porters, high- or low-affinity, several predictions can bemade with the information available at this moment. Fivefungal and plant HAK transporters, SoHAK1 (Banuelos etal. 1995), NcHAK1 (Haro et al. 1999), HvHAK1 (Santa-Marıa et al. 1997), AtKT1/KUP1 (Fu and Luan 1998, Kimet al. 1998) and AtHAK5 (the present work) proved to behigh-affinity K+ transporters. These results suggest thatHAK transporters might be typically high-affinity K+
transporters that have evolved for the acquisition of K+
against large transmembrane gradients (Haro et al. 1999).There are reports, however, proposing that AtKT2/KUP2 isa low-affinity K+ transporter (Quintero and Blatt 1997),and that AtKT1/KUP1 is a low- and a high-affinity K+
transporter (Fu and Luan 1998, Kim et al. 1998). Due totechnical reasons (Rodrıguez-Navarro 2000) and thedifficulties to characterise low-affinity K+-transporters inyeast trk1 trk2 mutants (Santa-Marıa et al. 1997), HAK-me-diated low-affinity K+-uptake needs to be confirmed byfurther investigations.
Apart from the role of the HAK transporters in roots(Santa-Marıa et al. 1997), the existence of transcripts encod-ing high-affinity HAK K+-transporters not only in rootsbut also in the upper part of the plant (Fu and Luan 1998,Kim et al. 1998) (Fig. 1) suggests that they may mediate themicromolar K+ uptake characterised in non-root tissues(Smith and Epstein 1964, Osmond and Laties 1968, Reedand Bonner 1974, Blatt 1985, Bellando et al. 1995). Thisnotion is further supported by the observations that micro-molar K+ uptake in both root (Sacchi et al. 1997) andnon-root cells (Smith and Epstein 1964) exhibits competitiveinhibition by micromolar Cs+, and that bacterial and eu-karyotic Kup-HAK K+ transporters are the only onesdescribed up to now with the capacity to transport Cs+
(Bossemeyer et al. 1989 and the present work). The highnumber of genes encoding HAK transporters (Fig. 3, Table2) and the easy detection of HAK transcripts in non-rootcells suggest that this type of transporter may be present inmost plant cells. This notion is in contrast with the commonfeeling that ‘active’ K+ uptake in most non-root cells isunnecessary, because K+ concentration in the shoot apo-
plast is millimolar (Bowling 1987, Lowen and Satter 1989,Long and Widders 1990, Muhling and Sattelmacher 1997),high enough to allow the function of inward K+-channels.However, the notion that K+ channels can mediate K+
uptake in all type of plant cells, except in root cells whenexternal K+ is low, is not clear in the literature. Forexample, in mesophyll cells, inward-rectifying K+-channelshave been described in oat (Kourie and Goldsmith 1992),but they may not exist in all species (Li and Assman 1993,Bei and Luan 1998). Even if they do exist, the variations inthe membrane potential in mesophyll cells in response tolight and dark (Spalding et al. 1992, Spalding and Gold-smith 1993) may leave Em positive to EK+, when theexternal K+ is below 1 or 2 mM, making the function of aK+ channel impossible, as has been demonstrated in E.densa (Bellando et al. 1995).
There are many examples of families of plant genes thatencode isoforms of the same enzyme (Hrabak et al. 1996,Chou and Gasser 1997, Milioni and Hatzopoulos 1997). Thecase most related to the HAK family of K+ transportersdescribed herein is the family of the H+-ATPases (Harperet al. 1994, Arango et al. 1998), which are functionallycoupled to K+ transporters both in fungi (Rodrıguez-Navarro et al. 1986) and plants (Maathuis and Sanders1993). The numbers of isoforms of the H+-ATPase inArabidopsis (Harper et al. 1994) and in Nicotianaplumbaginifolia (Arango et al. 1998), 10 and 9, respectively,are not very different from the 13 genes that encode HAKtransporters in Arabidopsis (Fig. 3, Table 2). The highnumber of HAK genes present in Arabidopsis, and possiblyalso in other plants, suggests that HAK transporters playimportant roles. The phylogenetic tree of the HAK trans-porters (Fig. 3) shows the existence of pairs of transportersfrom Arabidopsis and barley (AtHAK5/HvHAK1,AAC24049/HvHAK3, AtKT4/HvHAK2), which possiblyevolved from a common ancestor at the moment of separa-tion of monocot and dicot species. Conservation of thesetransporters indicates that they may play similar essentialfunctions in Arabidopsis and barley.
Acknowledgements – We thank J. Dubcovsky, University of Cali-fornia at Davis, for the Southern analyses of the barley genes. Wealso wish to thank NIAR/STAFF, STAFF Institute, Japan, forproviding the rice clones C72058 and C71725. This work wassupported by the European Commission DG XII BiotechnologyProgramme, contract number BIO4CT960775, and by grant PB97-0560 from Direccion General de Ensenanza Superior, Ministerio deEducacion y Cultura. F. R. is a postdoctoral fellow funded byMinisterio de Educacion y Cultura, Spain, and G. E. S.-M. wasrecipient of a fellowship from Consejo Nacional de InvestigacionesCientıficas y Tecnicas of Argentina.
ReferencesAnderson JA, Huprikar SS, Kochian LV, Lucas WJ, Gaber RF
(1992) Functional expression of a probable Arabidopsis thalianapotassium channel in Saccharomyces cere6isiae. Proc Natl AcadSci USA 89: 3736–3740
Arango M, Oufattole M, Moriau L, Boutry M (1998) Expressionanalysis of eight plasma membrane H+-ATPase genes of Nico-tiana plumbaginifolia. In: Tester M, Morris C, Davies J (eds)11th International Workshop on Plant Membrane Biology.Cambridge, UK. The Society for Experimental Biology andJournal of Experimental Botany. Springer-Verlag, Berlin, p 18
Physiol. Plant. 109, 2000 41

Banuelos MA, Klein RD, Alexander SJ, Rodrıguez-Navarro A(1995) A potassium transporter of the yeast Schwanniomycesoccidentalis homologous to the Kup system of Escherichia colihas a high concentrative capacity. EMBO J 14: 3021–3027
Baunsgaard L, Venema K, Axelsen KB, Villalba JM, Welling A,Wollenweber B, Palmgren MG (1996) Modified plant plasmamembrane H+-ATPase with improved transport coupling effi-ciency identified by selection in yeast. Plant J 10: 451–458
Bei Q, Luan S (1998) Functional expression and characterization ofa plant K+ channel gene in a plant cell model. Plant J 13:857–865
Bellando M, Marre MT, Sacco S, Talaricio A, Venegoni A, MarreE (1995) Transmembrane potential-mediated coupling betweenH+ pump operation and K+ fluxes in Elodea densa leaveshyperpolarized by fusicoccin, light or acid load. Plant CellEnviron 18: 963–976
Blatt MR (1985) Extracellular potassium activity in attached leavesand its relation to stomatal function. J Exp Bot 36: 240–251
Blatt MR (1992) K+ channels of stomatal guard cells: Characteris-tics of the inward rectifier and its control by pH. J Gen Physiol99: 615–644
Bossemeyer D, Borchard A, Dosch DC, Helmer GL, Epstein W,Booth IR, Bakker EP (1989) Specific cesium transport via theEscherichia coli Kup (TrkD) K+ uptake system. J Bacteriol 171:2219–2221
Bowling DJF (1987) Measurement of the apoplastic activity of K+
and Cl− in the leaf epidermis of Cammelina communis inrelation to stomatal activity. J Exp Bot 193: 1351–1355
Brunelli JP, Pall ML (1993) A series of yeast/Escherichia coli lexpression vectors designed for directional cloning of cDNAsand cre/lox-mediated plasmid excision. Yeast 9: 1309–1318
Bush DS, Hedrich R, Schroeder JI, Jones RL (1988) Channel-medi-ated K+ flux in barley aleurone protoplasts. Planta 176: 368–377
Cao Y, Ward JM, Kelly WB, Ichida AM, Gaber RF, Anderson JA,Uozumi N, Schroeder JI, Crawford NM (1995) Multiple genes,tissue specificity, and expression-dependent modulation con-tribute to the functional diversity of potassium channels inArabidopsis thaliana. Plant Physiol 109: 1093–1106
Chou IT, Gasser CS (1997) Characterization of the cyclophilin genefamily of Arabidopsis thaliana and phylogenetic analysis ofknown cyclophilin proteins. Plant Mol Biol 35: 873–892
Fu H-H, Luan S (1998) AtKUP1: A dual affinity K+ transporterfrom Arabidopsis. Plant Cell 10: 63–73
Haro R, Sainz L, Rubio F, Rodrıguez-Navarro A (1999) Cloning oftwo genes encoding potassium transporters in Neurospora crassaand expression of the corresponding cDNAs in Saccharomycescere6isiae. Mol Microbiol 31: 511–520
Harper JF, Manney L, Sussman MR (1994) The plasma membraneH+-ATPase gene family in Arabidopsis : Genomic sequence ofAHA10 which is expressed primarily in developing seeds. MolGen Genet 244: 572–587
Hirsch RE, Lewis BD, Spalding EP, Sussman MR (1998) A role forthe AKT1 potassium channel in plant nutrition. Science 280:918–921
Hrabak EM, Dickmann LJ, Satterlee JS, Sussman MR (1996)Characterization of eight new members of the calmodulin-likedomain protein kinase gene family from Arabidopsis thaliana.Plant Mol Biol 31: 405–412
Kim EJ, Kwak JM, Uozumi N, Schroeder JI (1998) AtKUP1: AnArabidopsis gene encoding high-affinity potassium transport ac-tivity. Plant Cell 10: 51–62
Kochian LV, Lucas WJ (1988) Potassium transport in roots. AdvBot Res 15: 93–178
Kochian LV, Lucas WJ (1993) Can K+ channels do it all? PlantCell 5: 720–721
Kourie J, Goldsmith MHM (1992) K+ channels are responsible foran inwardly rectifying current in the plasma membrane ofmesophyll protoplasts of A6ena sati6a. Plant Physiol 98: 1087–1097
Li W, Assman SA (1993) Characterization of a G-protein-regulatedoutward K+ current in mesophyll cells of Vicia faba L. ProcNatl Acad Sci USA 90: 262–266
Long JM, Widders IE (1990) Quantification of apoplasticpotassium content by elution analysis of leaf lamina tissue frompea (Pisum sati6um L. cv. Argenteum). Plant Physiol 94: 1040–1047
Lowen CZ, Satter RL (1989) Light-promoted changes in apoplasticK+ activity in the Samanea saman pulvinus, monitored withliquid membrane microelectrodes. Planta 179: 421–427
Maathuis FJM, Sanders D (1993) Energization of potassium uptakein Arabidopsis thaliana. Planta 191: 302–307
Maathuis FJM, Sanders D (1994) Mechanism of high-affinitypotassium uptake in roots of Arabidopsis thaliana. Proc NatlAcad Sci USA 91: 9272–9276
Maathuis FJM, Ichida AM, Sanders D, Schroeder JI (1997a) Rolesof higher plant K+ channels. Plant Physiol 114: 1141–1149
Maathuis FJM, Sanders D, Gradmann D (1997b) Kinetics ofhigh-affinity K+ uptake in plants, derived from K+-inducedchanges in current-voltage relationships. Planta 203: 229–236
Milioni D, Hatzopoulos P (1997) Genomic organization of hsp90gene family in Arabidopsis. Plant Mol Biol 35: 955–961
Morsomme P, d’Exaerde AdK, Meester SD, Thines D, Goffeau A,Boutry M (1996) Single point mutation in various domains of aplant plasma membrane H+-ATPase expressed in Saccha-romyces cere6isiae increase H+-pumping and permit yeastgrowth at low pH. EMBO J 15: 5513–5526
Morsomme P, Dambly S, Maudoux O, Boutry M (1998) Singlepoint mutations distributed in 10 soluble and membrane regionsof the Nicotiana plumbaginifolia plasma membrane PMA2 H+-ATPase activate the enzyme and modify the structure of theC-terminal region. J Biol Chem 273: 34837–34842
Muhling KH, Sattelmacher B (1997) Determination of apoplasticK+ in intact leaves by ratio imaging of PBFI fluorescence. JExp Bot 48: 1609–1614
Muller-Rober B, Ellenberg N, Willmitzer L, Busch H, Becker D,Dietrich P, Hoth S, Hedrich R (1995) Cloning and electrophysi-ological analysis of KST1, an inward rectifying K+ channelexpressed in potato guard cells. EMBO J 14: 2409–2416
Osmond CB, Laties GG (1968) Interpretation of the dual isothermfor ion absorption in beet tissue. Plant Physiol 43: 747–755
Quintero J, Blatt MR (1997) A new family of K+ transportersfrom Arabidopsis that are conserved across phyla. FEBS Lett415: 206–211
Ramos J, Alijo R, Haro R, Rodrıguez-Navarro A (1994) TRK2 isnot a low-affinity potassium transporter in Saccharomyces cere-6isiae. J Bacteriol 1176: 249–252
Reed NR, Bonner BA (1974) The effect of abscisic acid on theuptake of potassium and chloride in A6ena coleoptile sections.Planta 116: 173–185
Regenberg B, Villalba JM, Lanfermeijer FC, Palmgren MG (1995)C-terminal deletion analysis of plant plasma membrane H+-ATPase: Yeast as a model system for solute transport across theplasma membrane. Plant Cell 7: 1655–1666
Rentsch D, Laloi M, Rouhara I, Schmelzer E, Delrot S, FrommerWB (1995) NTr1 encodes a high affinity oligopeptide transporterin Arabidopsis. FEBS Lett 370: 264–268
Rodrıguez-Navarro A, Ramos J (1984) Dual system for potassiumtransport in Saccharomyces cere6isiae. J Bacteriol 159: 940–945
Rodrıguez-Navarro A, Blatt MR, Slayman CL (1986) A potassium-proton symport in Neurospora crassa. J Gen Physiol 87: 649–674
Rodrıguez-Navarro A (2000) Potassium transport in fungi andplants. Biochim Biophys Acta (in press)
Rubio F, Gassmann W, Schroeder JI (1995) Sodium-driven potas-sium uptake by the plant potassium transporter HKT1 andmutations conferring salt tolerance. Science 270: 1660–1663
Sacchi GA, Espen L, Nocito F, Cocucci M (1997) Cs+ uptake insubapical maize root segments: Mechanism and effects on H+
release, transmembrane electrical potential and cell pH. PlantCell Physiol 38: 282–289
Sambrook J, Fritsch EF, Maniatis T (1989) Molecular Cloning: ALaboratory Manual. ISBN 87969-3099-6. Cold Spring HarborLaboratory Press, New York, NY
Santa-Marıa GE, Rubio F, Dubcovsky J, Rodrıguez-Navarro A(1997) The HAK1 gene of barley is a member of a large genefamily and encodes a high-affinity potassium transporter. PlantCell 9: 2281–2289
Schachtman DP, Schroeder JI, Lucas WJ, Anderson JA, Gaber RF(1992) Expression of an inward-rectifying potassium channel bythe Arabidopsis KAT1 cDNA. Science 258: 1654–1658
Schachtman DP, Schroeder JI (1994) Structure and transport mech-anism of a high-affinity potassium uptake transporter fromhigher plants. Nature 370: 655–658
Physiol. Plant. 109, 200042

Schleyer M, Bakker EP (1993) Nucleotide sequence and 3%-enddeletion studies indicate that the K+-uptake protein Kup fromEscherichia coli is composed of a hydrophobic core linked to alarge and partially essential hydrophilic C terminus. J Bacteriol175: 6925–6931
Schroeder JI, Fang HH (1991) Inward-rectifying K+ channels inguard cells provide a mechanism for low-affinity K+ uptake.Proc Natl Acad Sci USA 88: 11583–11587
Schroeder JI, Ward JM, Gassmann W (1994) Perspectives on thephysiology and structure of inward-rectifying K+ channels inhigher plants: Biophysical implications for K+ uptake. AnnuRev Biophys Biomol Struct 23: 441–471
Sentenac H, Bonneaud N, Minet M, Lacroute F, Salmon J-M,Gaymard F, Grignon C (1992) Cloning and expression in yeastof a plant potassium ion transport system. Science 256: 663–665
Sheahan JJ, Ribeiro-Neto L, Sussman MR (1993) Cesium-insensi-tive mutants of Arabidopsis thaliana. Plant J 3: 647–656
Sherman F (1991) Getting started with yeast. Methods Enzymol194: 3–21
Smith RC, Epstein E (1964) Ion absorption by shoot tissue: Kinet-ics of potassium and rubidium absorption by corn leaf tissue.Plant Physiol 39: 992–996
Spalding EP, Slayman CL, Goldsmith MHM, Bertl A (1991) Potas-sium channels in the plasma membrane of Arabidopsis meso-phyll cells. Plant Physiol 96S: 35
Spalding EP, Slayman CL, Goldsmith MHM, Gradmann D, BertlA (1992) Ion channels in Arabidopsis plasma membrane. Trans-
port characteristics and involvement in light-induced voltagechanges. Plant Physiol 99: 96–102
Spalding EP, Goldsmith MHM (1993) Activation of K+ channelsin the plasma membrane of Arabidopsis by ATP producedphotosynthetically. Plant Cell 5: 477–484
Thompson JD (1997) The CLUSTAL–X windows interface: Flex-ible strategies for multiple sequence alignment aided by qualityanalysis tools. Nucleic Acids Res 25: 4876–4882
Vasilets LA, Schwarz W (1993) Structure-function relationships ofcation binding in the Na+/K+-ATPase. Biochim Biophys Acta1154: 201–222
Villalba JM, Palmgren MG, Berberian GE, Ferguson C, Serrano R(1992) Functional expression of plant plasma membrane H+-ATPase in yeast endoplasmic reticulum. J Biol Chem 267:12341–12349
Walker DJ, Leigh RA, Miller AJ (1996) Potassium homeostasisin vacuolate plant cells. Proc Natl Acad Sci USA 93: 10510–10514
Wang T-B, Gassmann W, Rubio F, Schroeder JI, Glass ADM(1998) Rapid up-regulation of HKT1, a high-affinity potassiumtransporter gene, in roots of barley and wheat following with-drawal of potassium. Plant Physiol 118: 651–659
Zimmermann S, Talke I, Ehrhardt T, Nast G, Muller-Rober B(1998) Characterization of SKT1, an inward rectifying potas-sium channel from potato, by heterologous expression in insectcells. Plant Physiol 116: 879–890
Edited by J. K. Schjo*rring
Physiol. Plant. 109, 2000 43