Embed Size (px)
Citation preview

1
/CN/Chapter 2
/CT/Practical Anatomy and Physiology of the Ocular Motor System
/CA/Scott D. Z. Eggers
[BOX:
Abbreviations
CN cranial nerve
DLPC dorsolateral prefrontal cortex
DLPN dorsolateral pontine nuclei
EBNs excitatory burst neurons
FEF frontal eye fields
FOR fastigial oculomotor region
IBNs inhibitory burst neurons
INC interstitial nucleus of Cajal
INO internuclear ophthalmoplegia
LGN lateral geniculate nucleus
LLBN long-lead burst neurons
MLF medial longitudinal fasciculus
MST medial superior temporal visual cortex
MVN medial vestibular nucleus
NPH nucleus propositus hypoglossi
NRTP nucleus reticularis tegmenti pontis
OMNs ocular motor neurons

2
OPNs omnipause neurons
OTR ocular tilt reaction
PEF parietal eye fields
PPRF paramedian pontine reticular formation
riMLF rostral interstitial nucleus of the MLF
RIP raphe interpositus nucleus
SC superior colliculus
SCC semicircular canal
SEF supplementary eye fields
VOR vestibulo-ocular reflex
END BOX]
/H1/ Functional Classes of Eye Movements
The common goal of all eye movements is to facilitate a clear and stable view of the
environment (Leigh & Zee, 2006). Lateral-eyed animals like rabbits have a large field of view to
survey the environment and avoid predators. But the brain cannot manage the data processing
requirements of a visual system with high resolution across the entire visual field. Thus, lateral-
eyed animals generally have the trade-off of poor visual acuity. Birds of prey have evolved a
visual compromise by restricting their visual field in return for superior visual acuity within that
narrow field. Other frontal-eyed animals like humans have developed a small area of very high
spatial resolution at the center of the retina (the fovea) while maintaining lower resolution in the
periphery. This “foveal compromise” (Wong, 2008) solves the problem of information overload

3
but also requires that the image of an object of interest fall on the fovea for maximal visual
acuity.
Mechanisms have evolved to complement this foveal compromise strategy by ensuring
that images of interest are brought to and maintained on the foveae of both eyes. Image stability
on the retina must be maintained despite object or head motion, because image “slip” across the
retina or movement away from the fovea leads to blur or degrades visual acuity. Thus, one
category of eye movements helps hold target images steady on the retina. These include (1)
active processes of the visual fixation system to hold the image of stationary objects on the fovea
when the head is still; (2) the vestibular system to hold target images steady on the retina during
brief head movements; and (3) the optokinetic system to hold target images steady on the retina
during sustained head rotation. A second category of eye movements has evolved to direct the
high-resolution fovea to objects of interest. These include (1) the saccadic system to bring a
target image rapidly onto the fovea; (2) the smooth pursuit system to hold the image of a small
moving target on the fovea; and (3) the vergence system to move both eyes in opposite directions
(i.e., convergence or divergence) in order to simultaneously place the target image onto both
foveae regardless of target distance or eccentricity (Table 2–1). To fulfill these visual
requirements, the ocular motor system requires complex anatomy and physiology at every level,
from the extraocular muscles to cortical ocular motor regions.
[Insert Table 2–1 here]
/H2/ Visual Fixation

4
The fixation system holds the image of a stationary target on the fovea while the head is still.
Fixation may be a special type of smooth pursuit (suppressing unwanted drift of the eyes) or due
to an independent fixation system. Rather than simply the absence of visible eye movements,
visual fixation actually consists of constant miniature movements not detectable to the naked
eye, with the illusion of steady fixation. Normal fixation includes (1) microtremor (<0.01 degree,
up to 150 Hz); (2) microsaccades (average 0.1 degrees, 120 Hz); and (3) microdrift (<0.3
deg/sec). The role of these movements in visual fixation is unclear, though they may be
important for preventing peripheral vision fade of stable objects due to habituation of a persistent
stimulus.
/H2/ Vestibular
The vestibulo-ocular reflex holds images of the seen world steady on the retina by producing
compensatory eye movements during brief head rotations or translations. These phylogenetically
old reflexes are generated with a much shorter latency than visually mediated eye movements
and are critical for maintaining stable vision during natural activities such as walking and
running.
/H2/ Optokinetic
Optokinetic eye movements are those generated by movement of a large visual scene and serve
to hold images of the world steady on the retina during sustained head rotation. They supplement
the vestibular-induced eye movements that begin to decline during prolonged rotation.
Optokinetic eye movements consist of a slow phase in the direction of visual scene motion and a
nystagmus quick phase to reset the eye in the opposite direction.

5
/H2/ Saccades
Saccades are rapid, brief, conjugate eye movements that shift the line of sight to bring target
images onto the fovea. They may be volitional (elective and purposeful), reflexive (generated to
novel stimuli occurring unexpectedly in the environment), predictive (in anticipation of or in
search of the appearance of a target at a particular location), memory guided (to the location of a
previous target), to command (generated on cue), or spontaneous (seemingly random in the
absence of any specific task). Saccades are critical for exploring a visual scene and reading,
among other things.
/H2/ Nystagmus Quick Phases
The quick phases of nystagmus generated during vestibular and optokinetic stimulation are a
form of saccades. The evolutionary forerunner of volitional saccades, they serve to reset the eyes
during prolonged rotation and direct gaze toward the oncoming visual scene.
/H2/ Smooth Pursuit
Smooth pursuit allows the image of a small, slowly moving target to be held on the fovea while
the head is still. Smooth pursuit is primarily voluntary, driven by retinal slip from visual motion
and modulated by attention and motivation. The pursuit system is also required to track an object
on a complex moving background and to suppress reflexive vestibular and optokinetic responses
during combined head and eye tracking.
/H2/ Vergence

6
As opposed to versional eye movements, which conjugately move both eyes in the same
direction by the same amount, vergence eye movements move the eyes in opposite directions
(convergence or divergence) so that images of a single object are placed or held simultaneously
on the fovea of each eye. Vergence is provoked by either the retinal blur (loss of image
sharpness) or retinal disparity (image separation when images fall on noncorresponding areas of
each retina) that occur with changes of target image distance during binocular fixation, such as
when shifting gaze from a distant to a very near object.
/H1/ The Final Common Pathway for Eye Movements
/H2/ Orbital Muscle Gross Anatomy
The extraocular muscles reside within the bony confines of the cone-shaped orbit. At the orbital
apex, the four rectus muscles and superior oblique originate from the dense fibrous annulus of
Zinn through which the optic nerve passes. The four rectus muscles course anteriorly through
orbital fat and terminate as tendinous tissue on the sclera. The superior oblique passes through a
ring of connective tissue called the trochlea at the upper nasal portion of the orbital frontal bone
to terminate on the lateral posterior portion of the sclera. The inferior oblique originates in the
inferior nasal orbital wall, laterally crosses the orbital floor, and inserts on the lateral posterior
globe. The globe is suspended and supported in the orbit by a fibrous sac of fascia called Tenon’s
capsule.
/H2/ Extraocular Muscle Actions and Innervation

7
Six extraocular muscles control the movements of each eye: medial rectus, lateral rectus,
superior rectus, inferior rectus, superior oblique, and inferior oblique (Figure 2–1). The medial
rectus, superior rectus, inferior rectus, and inferior oblique are innervated by the oculomotor
nerve (cranial nerve III). The lateral rectus is innervated by the abducens nerve (cranial nerve
VI). The superior oblique is innervated by the trochlear nerve (cranial nerve IV).
Coordinated extraocular muscle action facilitates movement of the eyes in three
directional planes (horizontal, vertical, and torsional) about three axes (craniocaudal, interaural,
and naso-occipital). Nomenclature for these directions is described in Table 2–2. The actions of
each muscle are dependent on the muscle’s origin and terminal insertion, the center of rotation of
the eye, and the optical axis of the eye. Growing evidence also suggests that connective tissue
pulleys in the extraocular muscles contribute to their kinematic properties (Demer, 2006). The
muscles of each eye work in agonist/antagonist pairs. In order to facilitate rotation of the eye,
increased innervation to the agonist results in an equal amount of decreased innervation to the
antagonist (Sherrington’s law of reciprocal innervation). Muscle actions may vary depending on
the position of the globe in the orbit. Each extraocular muscle has a primary action, and all but
the medial and lateral recti also have secondary and tertiary actions. Horizontal eye movements
are controlled by the antagonistic medial rectus and lateral rectus muscles. The primary and only
action of the medial rectus is adduction, and the primary and only action of the lateral rectus is
abduction. Vertical and torsional eye movements are controlled by two antagonist pairs: the
superior and inferior recti and the superior and inferior oblique muscles. The contribution of a
given muscle to vertical eye movement depends on the horizontal position of the eye. When the
eye is in an abducted position, the superior and inferior rectus muscles are the principal elevator
and depressor muscles, respectively. When the eye is in an adducted position, inferior oblique

8
action causes elevation, and superior oblique action causes depression. The superior oblique and
superior rectus muscles are intorters of the eye, and the inferior oblique and inferior rectus are
extorters. The primary, secondary, and tertiary actions of each muscle are shown in Table 2–3.
[Insert Figure 2–1 here]
[Insert Table 2–2 here]
[Insert Table 2–3 here]
In addition to each eye’s antagonistic pairs with opposite directions of action, the
extraocular muscles exist as “yoked” pairs between eyes to generate conjugate eye movements.
The three yoked pairs include (1) the medial rectus in one eye and the lateral rectus in the other
eye, (2) the superior rectus in one eye and the inferior oblique in the other eye, and (3) the
inferior rectus in one eye and the superior oblique in the other eye. “Yoked” muscle pairs receive
equal and simultaneous innervation generated from premotor control systems, stimulating the
cranial nerve nuclei to elicit the conjugate eye movement (Hering’s law of equal innervation).
/H2/ Ocular Motor Nuclei and Nerves
/H3/ Abducens Nerve (Cranial Nerve VI)
Paired abducens nuclei are located in the dorsomedial pons at the floor of the fourth ventricle, in
close proximity to the fascicle of the facial nerve (Figure 2–2). Each nucleus contains abducens
motor neurons that form the abducens nerve and interneurons that decussate at the level of the
nucleus and ascend in the contralateral medial longitudinal fasciculus (MLF) to the oculomotor

9
medial rectus subnucleus to facilitate conjugate horizontal gaze in the direction ipsilateral to the
abducens nucleus of origin. The abducens fascicles arise from the ventral surface of the nucleus,
traverse the brainstem, emerge from the ventral pontomedullary sulcus, and travel as the
abducens nerve in the subarachnoid space where it ascends along the clivus. It pierces the dura,
travels through the cavernous sinus lateral to the internal carotid artery, and enters the superior
orbital fissure to innervate the ipsilateral lateral rectus muscle.
[Insert Figure 2–2 here]
[BOX:
Clinical Correlation: Abducens Nucleus and Nerve Palsies
A lesion of the peripheral abducens nerve or its fascicles within the lower pons will both produce
an isolated ipsilateral lateral rectus muscle weakness. The two may often be distinguished by the
presence of additional central neurological signs in a pontine lesion, such as contralateral arm
and leg weakness from involvement of the corticospinal tract. In either case, the lateral rectus is
weak for all classes of eye movements (saccades, pursuit, vestibular), and thus ipsilateral eye
movements will be slow, of limited range, or absent. However, if the abducens nucleus is intact,
interneurons innervating the contralateral medial rectus subnucleus of III will still produce
adduction upon attempted lateral gaze. If, on the other hand, the lesion affects the abducens
nucleus (including the abducens motor neurons and interneurons), the result is a complete
ipsilesional horizontal gaze paralysis of all eye movement classes, often producing a
contralateral gaze deviation. In this case, the eyes cannot be brought past the midline even by the
vestibulo-ocular reflex.

10
END BOX]
/H3/ Trochlear Nerve (Cranial Nerve IV)
Paired trochlear nuclei lie very close to the dorsomedial surface of the midbrain just below the
inferior colliculus. The fascicles emerge from the nuclei and briefly course dorsally before
exiting the dorsal midbrain. The trochlear nerves are the only cranial nerves to emerge from the
dorsal brainstem surface. After emerging, the nerves decussate within the anterior medullary
velum and wrap around the surface of the midbrain to travel ventrally within the subarachnoid
space toward the cavernous sinus. In the cavernous sinus, the trochlear nerve is located in the
lateral dural wall, inferior to the oculomotor nerve. From the cavernous sinus, the nerve passes
into the superior orbital fissure and ultimately innervates the superior oblique muscle
contralateral to the nucleus of origin.
/H3/ Oculomotor Nerve (Cranial Nerve III)
Paired oculomotor nuclei are located in the dorsal midbrain ventral to the periaqueductal gray
matter at the level of the superior colliculus. Each nuclear complex includes individual subnuclei
innervating the ipsilateral inferior rectus, medial rectus, and inferior oblique; a superior rectus
subnucleus that innervates the contralateral superior rectus muscle; and an Edinger-Westphal
nucleus supplying preganglionic parasympathetic output to the iris sphincter and ciliary muscles.
A single midline caudal central subnucleus provides innervation to both levator palpebrae
superioris muscles.
Third-nerve fascicles originate from the ventral surface of each nucleus and traverse the
midbrain, passing through the red nucleus and in close proximity to the cerebral peduncles

11
before emerging as ventral rootlets in the interpeduncular fossa. The rootlets converge into a
third-nerve trunk that continues ventrally through the subarachnoid space toward the cavernous
sinus, passing between the superior cerebellar artery and the posterior cerebral artery. In the
cavernous sinus, the third nerve is located in the dural sinus wall, just lateral to the pituitary
gland. From the cavernous sinus, the third nerve enters the superior orbital fissure. Just prior to
entry, the nerve anatomically divides into superior and inferior divisions. The superior division
innervates the superior rectus and the levator palpebrae superioris, and the inferior division
innervates the inferior and medial recti, the inferior oblique, and the iris sphincter and ciliary
muscles. Prior to innervating the ciliary and sphincter muscles, parasympathetic third-nerve
fibers synapse in the ciliary ganglion within the orbit.
/H2/ Internuclear Connections
The medial longitudinal fasciculus (MLF) is a paramedian pathway that lies in the dorsal
brainstem and carries ocular motor and vestibular signals between the medulla and midbrain. As
noted above, the MLF carries signals from the abducens nucleus to the contralateral medial
rectus portion of the oculomotor nucleus. These signals allow conjugate horizontal eye
movements with synchronous contraction of the ipsilateral lateral rectus and contralateral medial
rectus muscles (Figure 2–3). The MLF also carries signals for vertical gaze from the vestibular
nuclei in the medulla to the midbrain vertical gaze control centers important for vertical smooth
pursuit and vestibular eye movements.
[Insert Figure 2–3 here]

12
[BOX:
Clinical Correlation: Internuclear Ophthalmoplegia
Unilateral inactivation of the MLF results in ipsilaterally slowed or absent adduction with
abducting nystagmus in the contralateral eye during attempted contralateral gaze (internuclear
ophthalmoplegia, INO), in combination with a skew deviation with ipsilateral hypertropia.
Bilateral MLF inactivation results in bilateral impairment of adduction with bilateral dissociated
abducting nystagmus (bilateral INO), impaired vertical smooth pursuit, and reduced vertical
VOR gain.
END BOX]
[BOX:
Clinical Correlation: One-and-a-Half Syndrome
Occasionally a lesion affects the abducens nucleus on one side and the ipsilateral MLF
containing interneurons from the contralateral abducens nucleus that have already crossed and
are destined for the ipsilateral oculomotor nucleus. The result is a complete conjugate horizontal
gaze paralysis toward the side of the lesion as described earlier, plus an INO for gaze in the
opposite direction. Thus, the patient loses all conjugate gaze ipsilesionally (the “one”) and can
only abduct the contralateral eye with attempted contralateral gaze (the “half”).
END BOX]
/H1/ Central Vestibular Structures and Pathways
The basic three-neuron arc of the vestibulo-ocular reflex (VOR) consists of the vestibular
ganglion and nerve, vestibular nuclei, and ocular motor nuclei (Figure 2–4). The VOR serves to

13
maintain stable gaze direction by compensating for head movement. The drive for the VOR is
vestibular rather than visual and thus can operate at a much shorter latency than could occur if
visual information had to reach the visual cortex and then be relayed to the brainstem. However,
the cerebellum has important connections that fine tune the VOR to changing visual
requirements.
[Insert Figure 2–4 here]
/H2/ Vestibular Nuclei
Four nuclei form the vestibular nuclear complex on each side in the dorsal medulla: The rostral
portions (medial and superior vestibular nuclei) predominantly receive input from the SCCs,
with their central projections most important for generating the rotational (angular) VOR. The
caudal portions (lateral and inferior nuclei) mainly receive projections from the otolith organs,
affecting the linear (translational) VOR, vertical ocular alignment, and vestibulospinal reflexes.
/H2/ Cerebellum
The cerebellum receives input from the vestibular nuclei as well as directly from vestibular nerve
afferents that bypass the vestibular nuclei. These inputs travel in the juxtarestiform body of the
inferior cerebellar peduncle to the ipsilateral vestibulocerebellum (flocculonodular lobe,
consisting of the flocculus, adjacent paraflocculus, nodulus, and ventral uvula). Additionally, the
cerebellum receives visual and ocular motor signals from regions such as the pontine nuclei,
nucleus reticularis tegmenti pontis, paramedian tract, and inferior olivary nuclei. Thus, although
not part of the three-neuron arc of the VOR, the cerebellum is critical for adapting the gain (ratio

14
of eye movement to head movement) and direction of the VOR to new visual requirements, as
occurs in disease states like unilateral vestibular loss or even when changing spectacle
prescriptions (Versino, Hurko, & Zee, 1996; Walker & Zee, 1999).
/H2/ Central Organization of the Vestibulo-ocular Reflex
The function of the VOR is to generate compensatory conjugate eye movements in the opposite
direction of brief head movement and thereby maintain a stable angle of gaze and clear vision.
The VOR can be subdivided into canal-ocular reflexes, with input from the semicircular canals
driving the angular VOR (aVOR), and otolith-ocular reflexes driving the translational VOR
(tVOR). An additional otolith-mediated VOR, ocular counterrolling, occurs in response to
change in the static orientation of the head with respect to gravity in the roll (frontal) plane; a
small change in the static torsion (counterrolling) of the eyes occurs in the opposite direction
with sustained head tilt.
/H3/ Canal-ocular Reflexes
Stimulation or inhibition of a single SCC leads to slow-phase eye movements that rotate the eye
in a plane parallel to the canal. Thus, the affected canal can be inferred from the pattern of
nystagmus, such as the mixed vertical-torsional nystagmus of benign paroxysmal positioning
vertigo (BPPV) occurring from inappropriate stimulation of the posterior SCC. Central
mechanisms are used to suppress the effect of persistent vestibular imbalance and thereby
compensate for vestibular lesions. The brisk nystagmus seen in acute peripheral vestibular
lesions will gradually decrease in intensity as the central vestibular system compensates for the

15
imbalance. In addition, visual fixation may significantly suppress the spontaneous nystagmus of
an acute vestibular lesion, necessitating the use of examination techniques that eliminate fixation.
To generate the aVOR, the vestibular nuclei send excitatory and inhibitory signals to
specific ocular motor nuclei in order to activate yoked pairs of extraocular muscles (and inhibit
their antagonists) (Fife, 2010). This reflex leads to activation of specific muscles that move the
eyes in the same plane but in the opposite direction as the semicircular canal being stimulated,
regardless of the initial position of the eye in the orbit. The horizontal aVOR is the simplest and
most commonly tested. Activation of the lateral semicircular canal by ipsilateral head rotation
leads to vestibular nerve excitation synapsing on the vestibular nuclei. From there, second-order
excitatory projections course rostromedially and then cross the midline to synapse on the
contralateral abducens nucleus. As with generation of other conjugate eye movements, abducens
motor neurons and interneurons are activated within the abducens nucleus, leading to activation
of the ipsilateral lateral rectus (contralateral to the vestibular stimulus) and via the MLF to the
oculomotor nucleus, activation of the opposite medial rectus (ipsilateral to the vestibular
stimulus). Some neurons also connect directly from the vestibular nuclei to the ipsilateral medial
rectus subnucleus in the ascending tract of Deiters, but its functional significance is uncertain.
For each extraocular muscle activated, its antagonist must be inhibited to permit rotation of the
eye in the orbit. Thus, for each excitatory pathway, the vestibular nuclei send inhibitory
projections to antagonist muscles (for the horizontal aVOR this is the ipsilateral lateral rectus and
contralateral medial rectus). In addition, during head rotation, the semicircular canal being
inhibited (such as the right horizontal canal during leftward head rotation) reduces its tonic firing
rate, thereby facilitating relaxation of the antagonist muscles. The eye movements elicited by
vestibular stimulation constitute the vestibular slow phase eye movements. However, sustained

16
vestibular stimulation in an awake person leads to nystagmus quick phases opposite the slow
phase direction (toward the side of vestibular stimulation). The nystagmus direction is named
based on quick phase direction.
The vertical semicircular canal projections are more complicated but follow the same
principles. Each anterior (superior) SCC sits in the same plane as the contralateral posterior SCC.
Thus, in natural states of movement, one canal is activated while its contralateral functional pair
is inhibited. The anterior semicircular canal is oriented along the same axis of rotation as that
produced by activation of the ipsilateral vertical recti and contralateral obliques. Stimulation of
the anterior canal leads to excitation of the ipsilateral superior rectus and contralateral inferior
oblique via brainstem connections between the vestibular nuclei and the two oculomotor
subnuclei. Activation of the posterior canal produces excitation of the contralateral inferior
rectus and ipsilateral superior oblique. Thus, with the eyes in the straight ahead position,
stimulation of an individual vertical SCC will produce a combination of vertical and torsional
eye movement based upon the actions of the muscles activated. For example, excitation of the
right posterior SCC produces vestibular slow phases that are downward and with torsion where
the top poles of the eyes roll toward the subject’s left (extorsion of the left eye and intorsion of
the right eye), with nystagmus quick phases that are upward and with oppositely directed torsion.
Because of the fact that the trochlear nucleus and superior rectus subnucleus of III innervate
contralateral muscles, vertical excitatory projections from the vestibular nuclei to ocular motor
nuclei cross the midline, but vertical inhibitory connections do not.
During dynamic head roll in the frontal plane (tilting the head alternately to the right and
left shoulder), the dynamic torsional VOR generates compensatory slow-phase torsional eye
movements in the opposite direction predominantly by the vertical semicircular canals. Rolling

17
to the right stimulates the right anterior and posterior canals while inhibiting the left anterior and
posterior canals. Provided that brainstem saccade networks are intact, torsional nystagmus quick
phases occur opposite the slow phases. The gain of the dynamic torsional VOR is between 0.4
and 0.7, lower than that of the horizontal or vertical VOR.
/H3/ Otolith-ocular Reflexes
The otolith-ocular reflexes refer to compensatory eye movements evoked by stimulation of the
utricle or saccule. The two types include (1) the translational VOR (tVOR) in response to linear
head acceleration, and (2) ocular counterrolling (or static torsional VOR) in response to static
head tilt in the roll plane.
The central otolith projections for the tVOR are less studied than for the aVOR but must
take into account target distance and eccentricity. It appears that the horizontal tVOR may arise
from stimulation of the lateral portion of the utricle during ipsilateral linear head translation, with
polysynaptic connections to the lateral vestibular nuclei then projecting (possibly via the
cerebellum) contralaterally to the abducens nucleus and driving oppositely directed conjugate
slow phase eye movements.
Projections from the medial portion of the utricle may be more important for signaling
head tilt and generating compensatory counterrolling using vertical torsional eye muscles. The
medial portion of the utricle would be excited by sustained ipsilateral head tilt, synapsing on the
lateral vestibular nucleus, with connections via the MLF to the midbrain ocular motor nuclei,
producing counterrolling of the eyes in the opposite direction via excitation of the ipsilateral
superior oblique and superior rectus and contralateral inferior oblique and inferior rectus.

18
The brain is evidently able to use contextual cues to resolve the ambiguity of whether the
head is being linearly translated or tilted relative to gravity despite the fact that the shear forces
on the utricular macula would be the same for each. This may be based upon the frequency of
linear acceleration input, with low-frequency input interpreted as tilt and high-frequency input as
translation. The brainstem velocity storage mechanism may also contribute by computing an
internal estimate of gravity by integrating angular head velocity signals from the SCCs.
[BOX:
Clinical Correlation: Ocular Tilt Reaction
A lesion anywhere along the otolith ocular pathway between the utricle and vertical/torsional
ocular motor nuclei in the midbrain (including in the MLF) can lead to the ocular tilt reaction
(OTR) (Brodsky, Donahue, Vaphiades, & Brandt, 2006). For example, a lesion of the left utricle
or its peripheral or central connections disrupts the normally symmetric utricular input and leads
to a shift in the patient’s internal estimate of absolute vertical (gravity) in the roll plane to the
left. Thus, the brain erroneously registers that the head is tilted to the right with respect to
gravity. This results in an OTR to the left with the pathologic triad of (1) head tilt to the left to
realign the head’s vertical axis with the perceived but incorrect vertical gravitational axis, (2)
torsional ocular counterrolling of the top pole of the eyes to the left to realign the eyes’ vertical
meridian with the perceived but incorrect vertical gravitational axis, and (3) skew deviation (the
left eye depresses and right eye elevates) to realign the eyes’ horizontal meridian with the
perceived but incorrect internal estimate of the earth-horizontal (Figure 2–5). Because central
utricular pathways cross at the pontomedullary junction to enter the MLF, a lesion of the

19
utricular nerve or medulla will cause an ipsilesional OTR, while a lesion in the pons or midbrain
after the projections have crossed will cause a contralesional OTR.
END BOX]
[Insert Figure 2–5 here]
/H3/ Velocity Storage Mechanism
The velocity storage mechanism is a central phenomenon by which the raw rotational vestibular
signal from the cristae ampullaris is prolonged or perseverated in order to improve the ability of
the aVOR to transduce the low-frequency components of sustained head rotation. The result is
that the time constant (the time for an exponential function to decay to 37% of its initial value) of
the aVOR is improved from 6 to 7 s (based on the physical properties of the cupula) to about 15
to 20 s (based on the nystagmus response to sustained rotation). Optokinetic after-nystagmus is
attributed to the vestibular velocity storage mechanism. The velocity storage mechanism may
also be important for helping distinguishing tilt from translation. The vestibular commissure
connects the two vestibular nuclear complexes and appears to be important for velocity storage,
as if it is sectioned, velocity storage is abolished.
[BOX:
Clinical Correlation: Periodic Alternating Nystagmus
Acquired periodic alternating nystagmus (PAN) is a spontaneous horizontal jerk nystagmus
present in straight-ahead gaze that reverses directions about every 2 min. It reflects instability of
the velocity storage mechanism (Furman, Wall, & Pang, 1990; Leigh, Robinson, & Zee, 1981).

20
Normally, GABAergic inhibitory inputs from the cerebellar nodulus and ventral uvula help
control vestibular rotational responses. In the setting of a nodulus or ventral uvula lesion, the
velocity-storage mechanism becomes unstable, and short-term vestibular adaptation leads to
sustained horizontal nystagmus that reverses directions every 2 min as PAN. Although in pure
form PAN is only present in darkness, it may be present during attempted visual fixation if the
adjacent flocculus and paraflocculus are also involved (because of the floccular role in VOR
suppression).
END BOX]
/H1/ Control of Saccadic Eye Movements
In order to properly execute a saccadic eye movement to bring an image detected in the visual
periphery to the fovea (for the purpose of visual search, reading, or to view a specific target), the
brain must simultaneously carry out several complex tasks. The location of a visual stimulus is
represented on the surface of the visual cortex, with different parts of this two-dimensional
cortical map corresponding to different locations on the retina. However, the final effectors of
the ocular motor system, the ocular motor neurons (OMNs), encode the characteristics for
saccades in terms of their temporal discharge, with the size of a saccade proportional to the total
number of discharge spikes. Furthermore, the OMNs cause the extraocular muscles to move the
eyes with respect to the head, not to the environment. Thus, the brain must transform the visual
stimulus that is two-dimensionally “place-coded” in terms of the location of active neurons in the
visual cortex, into a saccadic command to the OMNs that is “temporally coded” in terms of
discharge frequency and duration, further taking into account the three-axis nature of eye
rotation. Once the trajectory for the saccade is determined, this vector must be separated into

21
horizontal and vertical components to stimulate specific premotor burst neurons for oblique
saccades. To ensure accuracy, the desired size of the saccade must take into account overcoming
the elastic inertia of the extraocular orbital tissues, as well as whether the gaze change will
consist of combined head and eye movements or eye movements alone (Sparks, 2002).
/H2/ Brainstem Control of Saccades
Two main types of neurons are important in the brainstem network for generating the premotor
commands for saccades, burst neurons and omnipause neurons (Scudder, Kaneko, & Fuchs,
2002).
/H3/ Excitatory Burst Neurons
Brainstem excitatory burst neurons (EBNs, sometimes referred to as short- or medium-lead or
premotor burst neurons) carry the immediate supranuclear premotor saccadic command and
project monosynaptically to OMNs. They begin firing 8 to 12 ms before a saccade and fire
throughout the duration of the saccade. They are silent during fixation and slow eye movements.
The discharge characteristics of EBNs are tightly correlated with saccade properties when the
head is in a fixed position during the saccade. For example, the number of spikes in the burst
discharge is correlated to the size of the saccade, the duration of the burst discharge is correlated
with the duration of the saccade, and the peak frequency of the burst discharge is correlated with
the peak velocity of the saccade. These relationships between neuronal discharge and saccade
properties may be uncoupled when the head is not fixed during the saccade because small head
movements also contribute to gaze changes and stabilization.

22
Excitatory burst neurons for horizontal saccades are located in the paramedian pontine
reticular formation (PPRF) in the pons just rostral to the abducens nucleus. EBNs for vertical and
torsional saccades lie in the rostral interstitial nucleus of the medial longitudinal fasciculus
(riMLF) rostral to the oculomotor nucleus and ventral to the periaqueductal gray in the
mesencephalic reticular formation (Bhidayasiri, Plant, & Leigh, 2000). For horizontal saccades,
premotor burst signals project to the ipsilateral abducens nucleus, contacting both abducens
motor neurons and internuclear neurons, to generate a conjugate ipsilateral saccade (Figure 2–6).
For vertical saccades, EBNs for upward and downward saccades are intermingled in the riMLF.
Upward EBNs project bilaterally to elevator OMNs, while downward EBNs project only
ipsilaterally to depressor OMNs. EBNs discharge most vigorously for saccades that rotate the
eyes in a plane parallel to that of a pair of reciprocally acting vertical semicircular canals, which
creates a torsional component. For example, EBNs in the right riMLF discharge to extort the
right eye and intort the left eye. Therefore, ipsilateral riMLF lesions abolish ipsilesional torsional
quick phases of nystagmus and impair downward more than upward saccades.
[Insert Figure 2–6 here]
[BOX:
Clinical Correlation: Supranuclear Saccadic Palsies
Disorders affecting excitatory burst neurons will impair saccades (rendering them slow or
absent) but may leave smooth pursuit and the vestibulo-ocular reflex intact. Progressive
conditions like spinocerebellar ataxia type 2 or a pontine tumor may gradually affect saccades,
whereas acute saccadic palsy may occur with an infarction or demyelinating plaque in the EBNs,

23
such as the PPRF. Conditions may preferentially affect pontine horizontal or midbrain vertical
EBNs. Progressive supranuclear palsy affects the riMLF early in its course, leading to slowing of
vertical saccades. Niemann-Pick type C, on the other hand, leads to slowing of horizontal
saccades, causing oblique saccades to follow a curved trajectory with a slower horizontal
component. Rarely after cardiopulmonary bypass, patients can awaken with permanent loss of all
saccades and quick phases in all directions, with other functional classes of eye movements
preserved, apparently from a lesion of the brainstem saccadic generating system (Baloh, Furman,
& Yee, 1985; Bhidayasiri et al., 2001; Eggers, Moster, & Cranmer, 2008).
END BOX]
/H3/ Inhibitory Burst Neurons
In addition to the EBNs described above, inhibitory burst neurons (IBNs) are another type of
premotor burst neuron that project monosynaptically to inhibit antagonist OMNs and their
extraocular muscles during a saccade. The IBNs are located caudal to the abducens nucleus in
the medullary reticular formation for horizontal eye movements and are intermingled with
neurons in the interstitial nucleus of Cajal (INC) for vertical eye movements. For horizontal eye
movements, IBNs project to the contralateral abducens nucleus to inhibit it during ipsilateral
saccades, in addition to inhibiting the contralateral EBNs and IBNs. IBNs receive excitatory
input from the contralateral superior colliculus and inhibitory inputs from omnipause neurons
and contralateral IBNs.
/H3/ Omnipause Neurons

24
In order to maintain stable fixation, EBNs require constant inhibition except when a saccade is
called for. This inhibition is mediated by tonically discharging glycinergic omnipause neurons
(OPNs) located in the nucleus raphe interpositus (RIP), medial to the abducens nerve fascicles.
OPN firing ceases just prior to saccades in any direction and resumes immediately at saccade
end. Microstimulation of OPNs in the middle of a saccade will stall the saccade midflight. The
mechanism by which omnipause neurons are inhibited to allow a saccade to occur appears
complicated. The initial inactivation of the OPNs may result from activity in trigger-latch long-
lead burst neurons (LLBNs) in the rostral pons and midbrain, from the “fixation zone” of the
superior colliculus, and from the frontal eye fields, supplementary eye fields, and fastigial
nucleus of the cerebellum.
/H3/ Long-Lead Burst Neurons
Long-lead burst neurons (LLBNs) exhibit activity 40 to 100 ms prior to saccade onset. They are
located throughout the midbrain and pontine reticular formations and likely consist of several
types: relay LLBNs, trigger-latch LLBNs, and precerebellar LLBNs. Relay LLBNs may form a
connection between the superior colliculus and excitatory burst neurons, synchronizing the onset
and end of saccades. The role of trigger-latch LLBNs is unclear, but they may function to inhibit
omnipause neurons and to hold omnipause neurons off for the duration of the saccade.
Precerebellar LLBNs receive input from the superior colliculus and project to the nucleus
reticularis tegmenti pontis (NRTP) which, in turn, projects primarily to the cerebellar saccadic
areas (the oculomotor vermis and the fastigial oculomotor region) via the middle cerebellar
peduncle.

25
[BOX:
Clinical Correlation: Opsoclonus and Ocular Flutter
A striking eye movement disorder occurs in which patients make involuntary bursts of high-
frequency conjugate oscillations of the eyes, each consisting of a series of back-to-back saccades
that lack an intersaccadic interval. When confined to the horizontal plane it is termed ocular
flutter, and when it also includes vertical and torsional movements it is termed opsoclonus
(Wong, 2007). Patients complain of oscillopsia and vertigo. Ataxia is a commonly
accompanying feature. Diseases causing this include brainstem encephalitis and paraneoplastic
syndromes (neuroblastoma in infants, breast or small cell lung cancer in adults). The
pathophysiological basis for opsoclonus and flutter remains unclear. Since saccadic in origin,
initially it was thought that dysfunction of the OPNs was responsible, but neuropathological
evidence for this is lacking. Abnormal feedback through a cerebellar loop also seems
increasingly unlikely. A hypothesis based on brainstem models suggests the membrane
properties of IBNs may control the frequency and amplitude of these saccadic oscillations
(Ramat, Leigh, Zee, & Optican, 2007).
END BOX]
/H2/ Superior Colliculus
The superior colliculus (SC) is a multilayered structure that lies in the midbrain tectum as the
upper portion of the quadrigeminal plate. It is the primary source of commands to the brainstem
immediate premotor structures for generating saccadic eye movements. It receives signals from
many cortical and subcortical areas and sends output, at least indirectly, to all of the premotor
areas involved with controlling eye and head movements, including EBNs, OMNs, LLBNs, and

26
the vestibular nuclei. Inputs descend to the SC from frontal and parietal eye fields (directly and
through the basal ganglia), as well as from visual cortex containing retinotopically coded
information regarding target location. The SC contains a “motor map” where information about
saccade direction and amplitude is represented as a “place code.” The location of a SC neuron,
not its discharge characteristics, determines the direction and amplitude of the saccade for which
it encodes. This map is two dimensional, and downstream modifications (possibly in the NRTP)
must convert collicular commands to three-dimensional displacement vectors for eye and head
movements.
The rostral pole of this motor map seems important for maintaining steady fixation
(suppressing saccades), and this “fixation zone” sends tonic excitatory projections directly to
OPNs. The more caudal portions are important for target selection (size and direction of
movement) and initiation of eye and eye-head gaze shifts. Deeper layers of the caudal SC are
important for coordinated movements of the head and eyes, including projections as the
tectospinal tract. Discrete lesions of the SC are rare but may cause increased latency and slowing
of saccades, although redundant pathways from the frontal eye fields prevent loss of saccade
generation altogether.
/H2/ Cortical Control of Saccades
Lesional and stimulation experiments in animals as well as functional neuroimaging studies in
humans show that widespread areas of the frontal and parietal cortex are important for saccadic
control. These cortical structures are integral for attention, motivation, target selection, and
programming of eye movements. Rather than a top-down arrangement, these regions probably
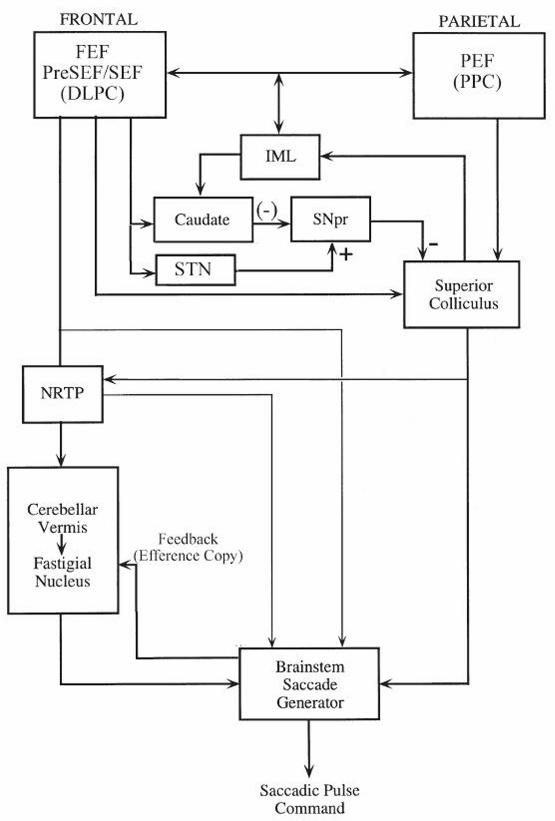
form a vast network with many reciprocal connections (Figure 2–7). The frontal regions include

27
the frontal eye fields (FEFs), supplementary eye fields (SEFs), dorsolateral prefrontal cortex
(DLPC), and cingulate cortex. The primary parietal region is the parietal eye fields (PEFs) within
the posterior parietal cortex (Pierrot-Deseilligny, Milea, & Muri, 2004).
[Insert Figure 2–7 here]
The FEF dispatch contralateral voluntary and visually guided saccades to targets. They
project to the ipsilateral SC both directly and indirectly through the basal ganglia (caudate and
substantia nigra pars reticulata). FEF also project directly to the contralateral NRTP and
omnipause neurons of the pontine tegmentum. The SEF lie anterior to the supplementary motor
cortex. They are important for programming saccades as part of a learned or complex behavior.
DLPC facilitates memory-guided saccades, antisaccades, and advanced planning of
environmental scanning using memory of target location. PEF receives input from secondary
visual areas and the thalamic pulvinar nucleus. Projections go to the FEF and to the SC directly.
The PEF are important for directing visual attention in extrapersonal space and initiating visually
guided reflexive saccades. Inhibitory projections from the basal ganglia to SC inhibit extraneous
reflexive saccades during attempted fixation and facilitate volitional saccades in the context of
remembered and learned behavior.
[BOX:
Clinical Correlation: Cortical Saccadic Abnormalities
Frontal lobe lesions produce various saccade abnormalities based on the location of the lesion.
Unilateral FEF lesions increase the reaction time of saccades, impair contralateral anticipatory

28
saccades, and impair ability to inhibit inappropriate saccades to visual stimuli, as well as impair
pursuit and optokinetic following toward the side of the lesion (Thurtell, Tomsak, & Leigh,
2007). SEF lesions impair memory-guided saccades after gaze shifts and affect ability to make a
remembered sequence of saccades to an array of visible targets. DLPC lesions produce
inaccurate contralateral memory-guided saccades and impair predictive saccades and
antisaccades.
Larger acute destructive hemispheric lesions such as infarctions, especially right posterior
lesions, may cause ipsilateral sustained horizontal conjugate gaze deviation (where patients may
“look away from the hemiparesis” as opposed to gaze directed toward the hemiparesis in an
acute pontine lesion). In a hemispheric lesion, it is usually possible to drive the eyes across the
midline with head rotation or caloric stimulation. If able to make contralateral saccades, patients
with parietal lobe lesions may have contralateral inattention (with or without homonymous
hemianopia), ipsilateral gaze preference, increased latency for visually guided saccades, and
impaired smooth pursuit ipsilaterally. Intermittent horizontal conjugate gaze deviation suggests
seizures from the contralateral hemisphere.
END BOX]
[BOX:
Clinical Correlation: Ocular Motor Apraxia and Balint’s Syndrome
Acute bilateral frontal or frontoparietal lesions such as infarctions may cause a striking disorder
called acquired ocular motor apraxia, characterized by loss of voluntary control of saccades and
pursuit but preservation of reflexive eye movements such as slow and quick phases of the VOR.

29
Patients struggle to make saccades to command and to follow a pointer, often using blinks and
head movements to facilitate gaze shifts.
When ocular motor apraxia is accompanied by disturbances of peripheral visual attention
(simultagnosia) and inaccurate arm pointing (optic ataxia), the term Balint’s syndrome is applied.
Lesions are typically bilateral parieto-occipital. Voluntary saccades may be made more easily
than in response to visual stimuli, reflecting defects in the visual guidance of saccades and
impaired visual search. Smooth pursuit is also impaired.
END BOX]
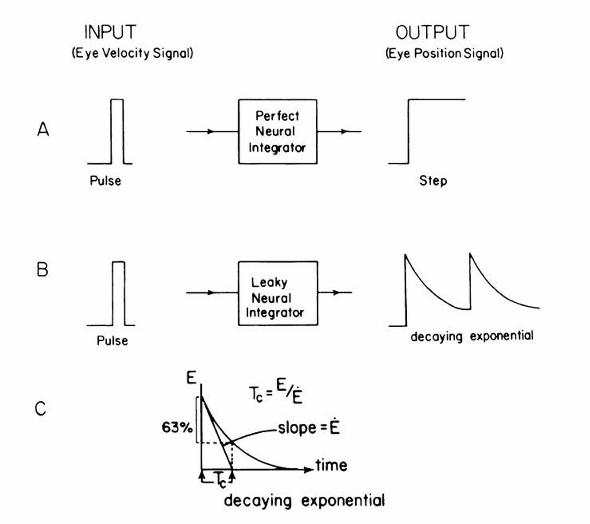
/H1/ Gaze Holding and the Neural Integrator
Once a visual target is acquired, the eyes must be held steady in an eccentric position to maintain
fixation. To counteract the orbital elastic restoring forces that would tend to pull the eyes back to
central position, tonic contraction of the extraocular muscles is achieved by an increase in the
sustained rate of discharge of the OMNs. This gaze-holding function is achieved by networks of
neurons that mathematically integrate saccadic velocity “pulse” signals into position “step”
commands, collectively referred to as the neural integrator (Figure 2–8).
[Insert Figure 2–8 here]
The basic scheme for the neural integrator begins with the EBN. A pulse discharge from
the EBN projects a velocity command signal to the OMN to cause phasic contraction of the
extraocular muscle and overcome viscous drag of the orbit to generate a saccadic eye movement.
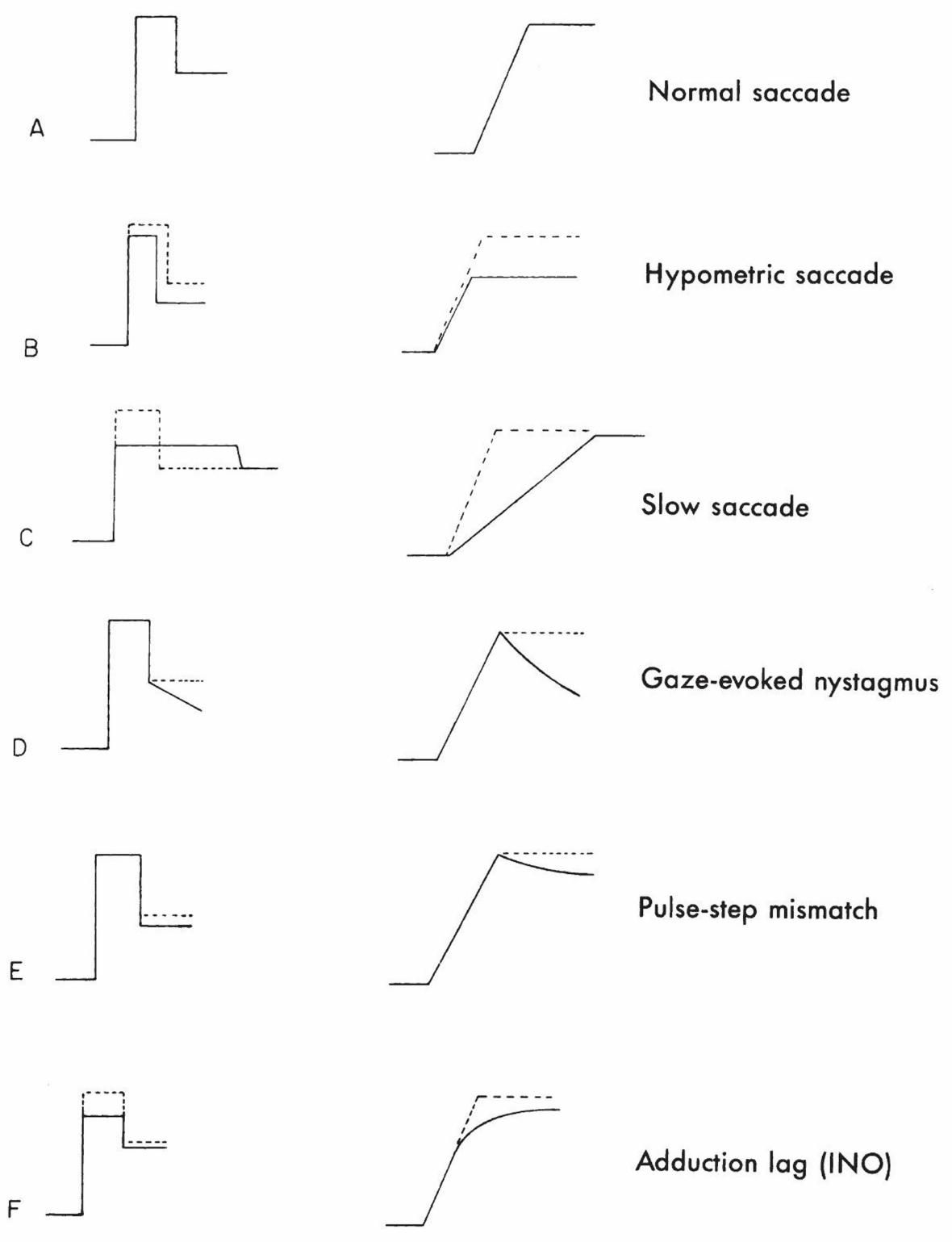
Abnormalities of the pulse can result in hypometric or slow saccades (Figure 2–9). The same

30
pulse signal from EBN is also sent through the neural integrator to generate a step of innervation,
that is, a position command to the OMN that changes the tonic contraction of the extraocular
muscle appropriate to hold the eye in the new position. If the performance of the neural
integrator is perfect, the eye will be held perfectly on the eccentric target.
[Insert Figure 2–9 here]
[BOX:
Clinical Correlation: Gaze-Evoked Nystagmus
Gaze-evoked nystagmus reflects dysfunction of the neural integrator. In this situation, attempts
to hold the eyes in an eccentric position lead to the eyes exponentially drifting back toward the
center because of inability to counteract the elastic restoring forces of the orbit. The neural
integrator is said to be “leaky” in this case. When a leaky integrator causes centripetal drift of the
eyes, corrective saccades are required to bring the eye back to the desired eccentric position, thus
producing gaze-evoked nystagmus. The rate of centripetal drift reflects the time constant of the
neural integrator—the faster the drift, the worse the integrator. Dysfunction can occur in a wide
variety of structural and functional disorders affecting the neural integrator or the cerebellar
structures that modulate it.
END BOX]
The neural substrate for gaze holding consists of cell groups throughout the brainstem and
cerebellum. The nucleus propositus hypoglossi and adjacent medial vestibular nucleus (NPH-
MVN) in the dorsal rostral medulla play a key role in integration of horizontal eye movements.

31
The NPH-MVN receives inputs from every structure that projects to the abducens nucleus and
encodes position signals to abducens neurons and interneurons, mainly those innervating fatigue-
resistant extraocular muscle fibers capable of sustained contraction. The interstitial nucleus of
Cajal (INC), located just caudal to the riMLF in the mesencephalic reticular formation, is the
primary structure responsible for integration of vertical and torsional eye movements (and also
appears important for eye-head coordination in the roll plane). It receives inputs from the riMLF
and from the vestibular nuclei via the MLF. The primary output from the INC decussates in the
posterior commissure to project position commands to the contralateral INC and CN III and IV
nuclei. The cerebellum, particularly the paraflocculi (tonsils) and flocculi (collectively part of the
“vestibulocerebellum”), appears critical for improving the performance of an inherently leaky
neural integrator. For example, the NPH-MVN has connections with the vestibulocerebellum
that likely serve as part of a positive feedback loop to OMNs, helping to increase the gain of the
neural integrator. In the setting of conditions such as cerebellar degeneration or Chiari
malformation (where the tonsils are compressed), gaze-evoked nystagmus commonly develops.
/H1/ Smooth Pursuit
The smooth pursuit system probably evolved to keep the fovea pointed at a stationary target
ahead as we navigate through our environment. Minimizing foveal “slip” of a visual object of
interest improves vision, while the “optic flow” of images across the rest of our retina as we walk
provides information about the three-dimensional layout of our environment and our direction of
movement within it. Subsequently, this system could be harnessed to pursue a small object
moving across a complex background (without inducing a perception of motion of self or the

32
stationary world) as well as assist with visual fixation (holding the image of a stationary object
on the fovea while the observer is stationary).
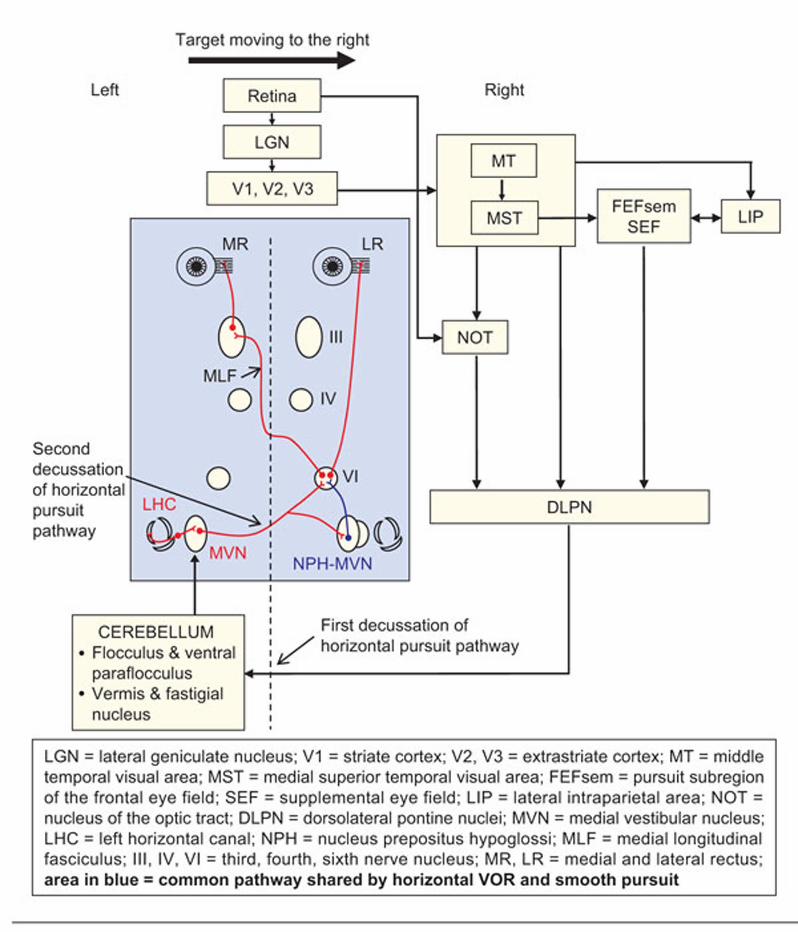
Signals encoding speed and direction of retinal image motion pass via the lateral
geniculate nucleus (LGN) to striate and extrastriate (middle temporal and medial superior
temporal, MST) cortex and posterior parietal cortex (i.e., PEF) (Figure 2–10). The MST seems to
contain visual tracking neurons that encode representations of object motion in world-centered
coordinates, being sensitive to retinal slip, slow eye movements, and slow head movements.
From there, further projections to the FEF and SEF may contribute to predictive aspects of
pursuit, utilizing some of the same corticofugal pathways as the saccadic system. The nucleus of
the optic tract and accessory optic system in the midbrain pretectum receives retinal slip
information directly from the retina and may help initiate pursuit.
[Insert Figure 2–10 here]
The main pursuit projections descend from parieto-temporo-occipital cortex to the pons,
particularly the DLPN and NRTP, encoding various visual and ocular motor signals including
eye velocity. The DLPN projects contralaterally to the cerebellar flocculus and paraflocculus via
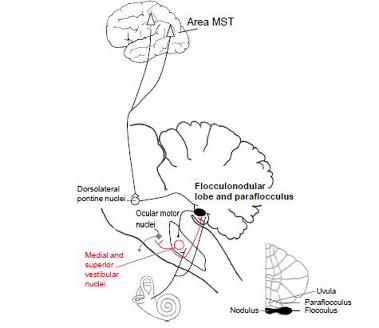
the middle cerebellar peduncle, whereas the NRTP projects to the dorsal vermis and then to the
caudal fastigial nucleus. From these cerebellar structures, fibers then reach the superior and
medial vestibular nuclei, which then (for horizontal pursuit) project back across the midline to
the abducens nucleus (completing a double decussation). The paraflocculus appears important
for controlling smooth pursuit and the flocculus for calibrating the VOR, while the vermis and
fastigial nucleus are critical for pursuit initiation (Krauzlis, 2004; Thier & Ilg, 2005).

33
[BOX:
Clinical Correlation: Smooth Pursuit Abnormalities
Pathways serving smooth pursuit are widespread throughout the cerebral hemispheres,
brainstem, and cerebellum, so pursuit abnormalities commonly accompany lesions throughout
the central nervous system. Reduced pursuit gain (the ratio of eye velocity to target velocity)
manifests as “choppy” or “saccadic” pursuit, as small saccades are made to catch up with the
moving target as the eyes fall behind. One must recognize that pursuit gain declines normally
with advancing age, especially at high velocities, but also becomes impaired in Parkinson
disease, progressive supranuclear palsy, cerebellar disorders, and large cerebral lesions. While
diffuse disorders cause omnidirectional pursuit abnormalities, large unilateral cerebral lesions
(especially of the parieto-occipital cortex and underlying white matter but also of the MST visual
cortex and the FEF) cause pursuit tracking deficits predominantly to the side of the lesion,
independent of homonymous hemianopia or visual neglect. This asymmetric pursuit can be
demonstrated with an “optokinetic” tape or drum. Directional pursuit abnormalities may also be
encountered with unilateral lesions of the descending pursuit pathways, including the midbrain
tegmentum, DLPN, and cerebellum. Because of the double decussation of the pursuit pathway,
lesions of the pontocerebellar projections and vestibular nuclei may impair either ipsilateral or
contralateral smooth pursuit. Unless the VOR is also impaired by a disease process, smooth
pursuit deficits are generally accompanied by impairment of VOR suppression during combined
eye-head tracking.
Isolated defects of vertical pursuit are less common. Bilateral INO from MLF lesions impairs
vertical pursuit (and the vertical VOR), as the MLF transmits pursuit and vestibular signals from

34
the vestibular nuclei to CN III and IV serving vertical eye movements. An unusual disturbance
can occur with cavernous angiomas of the middle cerebellar peduncle, where vertical pursuit is
accompanied by torsional nystagmus, suggesting that pursuit signals processed through the
cerebellum may be encoded in the same reference planes as the semicircular canals.
END BOX]
/H1/ Cerebellar Influences on Gaze
The cerebellum optimizes or refines eye movements so that they are calibrated to improve
accuracy and ensure clearest vision. In order to perform this role, the cerebellum receives both
sensory and motor information regarding the eye movement and must compare the predicted eye
movement based on the command with the desired eye movement and generate a signal to
decrease the error between predicted and desired to get the eyes accurately on target. Three
primary regions of the cerebellum are involved with ocular motility: (1) the flocculus and
paraflocculus; (2) the dorsal vermis (oculomotor vermis) and caudal fastigial nucleus; and (3) the
nodulus and ventral uvula.
As part of the vestibulocerebellum, the paired flocculi lie adjacent to the paraflocculi,
ventral to the inferior cerebellar peduncle and next to the vestibulocochlear nerve (CN VIII)
(Figure 2–11). The flocculi and paraflocculi receive mossy fiber input from the vestibular nuclei,
NPH, NRTP, DLPN, and paramedian tract (PMT) cell group and climbing fiber input from the
contralateral inferior olivary nucleus. The main output from the floccular and parafloccular
Purkinje cells is to the ipsilateral superior and medial vestibular nuclei. Studies suggest that the
flocculus is more important for calibrating the VOR, while the paraflocculus mainly contributes
to smooth pursuit. The flocculus and paraflocculus also appear to contribute inhibitory influence

35
to counteract the inherent tonic upward VOR bias otherwise favoring the anterior over posterior
SCC pathways, as lesions of these cerebellar structures often leads to upward slow phases and
downbeat nystagmus as would be seen with unopposed anterior canal activation.
[Insert Figure 2–11 here]
Lobules VI and VII of the dorsal vermis (Figure 2–12) receive mossy fiber inputs from
the PPRF, NRTP, DLPN, NPH, and vestibular nuclei as well as climbing fiber input from the
contralateral inferior olivary nucleus. Projections coming from the NRTP relay information
necessary for planning saccades, whereas those from the DLPN are more important for smooth
pursuit. Dorsal vermis Purkinje cells discharge prior to contralateral saccades as well as encode
target velocity during pursuit and combined eye-head tracking. Dorsal vermis Purkinje cells
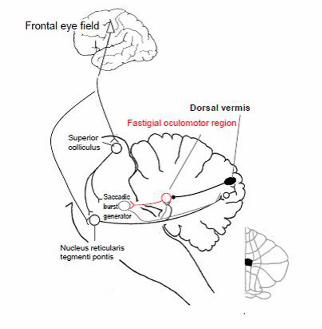
project to the caudal part of the deep cerebellar fastigial nucleus, coined the fastigial oculomotor
region (FOR), which also receives collaterals from the same climbing and mossy fiber inputs
destined for the dorsal vermis. The main projection from the fastigial nucleus crosses through the
other fastigial nucleus and enters the uncinate fasciculus in the border of the superior cerebellar
peduncle to reach the premotor burst neurons and OPNs in the brainstem. Fastigial neurons
discharge prior to and during contralateral saccades (facilitating them) and discharge late for
ipsilateral saccades (perhaps serving as a stop signal to end a saccade on target).
[Insert Figure 2–12 here]

36
The nodulus, which is the midline portion of the flocculonodular lobe, and the adjacent
ventral uvula control the velocity-storage mechanism of the VOR. This central vestibular
mechanism functions to enhance the responsiveness of the VOR to low-frequency stimuli such
as sustained rotation. Without a velocity-storage mechanism, the discharge from vestibular
nuclei driving the VOR would quickly decay during sustained rotation, based on the mechanical
properties of cupular deflection in the semicircular canals. Velocity storage prolongs the raw
vestibular signal so that the angular VOR can better transduce the low-frequency components of
head rotation. The neural substrate for velocity storage may be the medial vestibular nuclei and
their interconnections, since sectioning the vestibular commissure abolishes velocity storage.
Purkinje cells of the nodulus send GABAergic projections to the vestibular nuclei to control the
velocity-storage mechanism of the VOR.
[BOX:
Clinical Correlation: Cerebellar Syndromes
Three principal ocular motor syndromes may occur in the setting of cerebellar disease depending
on the location of dysfunction, with overlap being common due to more widespread cerebellar
dysfunction (Versino et al., 1996).
Lesions of the Flocculus and Paraflocculus
Focal dysfunction of the flocculi and paraflocculi, such as may be seen in Chiari malformation,
produces several abnormalities. Gaze-evoked nystagmus results from loss of the cerebellar
contribution to the fidelity of the brainstem neural integrator. Downbeat nystagmus and rebound
nystagmus are other common sequelae. Smooth pursuit is impaired (choppy or saccadic), as is

37
the ability to suppress the VOR during combined eye-head tracking or during caloric irrigation
by fixating on a stationary target. Finally, patients cannot adapt the VOR to changing visual
needs, such as new spectacle correction.
Lesions of the Dorsal Vermis and Fastigial Nucleus
Dorsal vermis and fastigial nucleus lesions cause saccadic dysmetria (inaccurate saccades),
typically hypometria if the vermis alone is involved and hypermetria if the fastigial nuclei are
involved. Unilateral lesions of the dorsal vermis would cause ipsilateral hypometria and mild
contralateral hypermetria of saccades. A unilateral lesion of the fastigial nucleus would cause
ipsilateral hypermetria and contralateral hypometria of saccades as well as cause the eyes to
tonically deviate ipsilaterally (ipsipulsion). While such asymmetric dysfunction is rarely seen
clinically since the fastigial projections cross within the opposite nucleus, the same functional
effect occurs in Wallenberg syndrome (below). Because those crossed projections ascend in the
contralateral uncinate fasciculus of the superior cerebellar peduncle, a lesion in the uncinate
fasciculus can cause contrapulsion of saccades. Finally, dorsal vermis and fastigial lesions can
impair smooth pursuit initiation.
Lesions of the Nodulus and Ventral Uvula
Nodulus and ventral uvula lesions lead to loss of the GABAergic inhibition of the velocity-
storage mechanism, thus maximizing the velocity-storage effect in circumstances that would
usually reduce it, such as pitching the head forward during post-rotatory nystagmus. The most
clinically important result of a nodulus lesion is periodic alternating nystagmus (PAN), where the
velocity-storage mechanism becomes unstable, and short-term vestibular adaptation leads to

38
sustained horizontal nystagmus that reverses directions every 2 min. Although in pure form PAN
is only present in darkness, it may be present during attempted visual fixation if the adjacent
flocculus and paraflocculus are also involved (because of its role in VOR suppression).
END BOX]
[BOX:
Clinical Correlation: Wallenberg Syndrome
Infarction of the dorsolateral medulla (generally from occlusion of the ipsilateral vertebral or
posterior inferior cerebellar arteries or vertebral dissection) causes Wallenberg syndrome,
consisting of ipsilateral impairment of facial pain and temperature sensation, Horner’s syndrome,
limb ataxia, and bulbar disturbances leading to dysarthria and dysphagia. Contralaterally, pain
and temperature sensation are impaired in the trunk and limbs. Patients commonly report vertigo
and a variety of unusual sensations of body and environmental tilt, including the room being
tilted on its side or upside down. Lateropulsion, a compelling sensation of being pulled to one
side (in this case toward the side of the lesion), is a common complaint.
Several ocular motor abnormalities are characteristic of Wallenberg syndrome and may be the
main or sole manifestation (Baloh, Yee, & Honrubia, 1981; Brazis, 1992). Lateropulsion of
saccades develops because the lesion affects the inferior cerebellar peduncle carrying climbing
fibers from the inferior olivary nucleus to the dorsal cerebellar vermis, leading to the functional
equivalent of a fastigial nucleus lesion. Lateropulsion of the eyes is easy to detect at the bedside.
If the patient is asked to fixate straight ahead and gently close the eyes, the eyes will conjugately
deviate toward the side of the lesion, requiring a refixation saccade back to the straight-ahead

39
position after opening the eyes again. Horizontal saccades are hypermetric toward the side of the
lesion (ipsipulsion) and hypometric contralateral to the lesion. Vertical saccades take an oblique
trajectory, with an inappropriate horizontal component toward the side of the lesion (requiring a
horizontal corrective saccade once the vertical saccade is complete).
The lesion’s involvement of the vestibular nuclei can produce spontaneous nystagmus (often
mixed horizontal-torsional), with slow phases usually directed toward the side of the lesion.
Smooth pursuit is impaired for targets moving away from the side of the lesion. The ocular tilt
reaction also occurs because of imbalance of the otolith-ocular pathway from the vestibular
nucleus lesion. The result is skew deviation with the ipsilateral eye lower (with corresponding
vertical diplopia), cylcodeviation where the top poles of the eyes roll toward the side of the
lesion, and ipsilateral head tilt.
END BOX]

40
/H1/References
Baloh, R. W., Furman, J., & Yee, R. D. (1985). Eye movements in patients with absent voluntary
horizontal gaze. Annals of Neurology, 17(3), 283–286.
Baloh, R. W., Yee, R. D., & Honrubia, V. (1981). Eye movements in patients with Wallenberg's
syndrome. Annals of the New York Academy of Sciences, 374, 600–613.
Benarroch, E. E. (2006). Basic neurosciences with clinical applications. Philadelphia, PA:
Butterworth Heinemann/Elsevier.
Bhidayasiri, R., Plant, G. T., & Leigh, R. J. (2000). A hypothetical scheme for the brainstem
control of vertical gaze. Neurology, 54(10), 1985–1993.
Bhidayasiri, R., Riley, D. E., Somers, J. T., Lerner, A. J., Buttner-Ennever, J. A., & Leigh, R. J.
(2001). Pathophysiology of slow vertical saccades in progressive supranuclear palsy.
Neurology, 57(11), 2070–2077.
Brazis, P. W. (1992). Ocular motor abnormalities in Wallenberg’s lateral medullary syndrome.
Mayo Clinic Proceedings, 67(4), 365–368.
Brodsky, M. C., Donahue, S. P., Vaphiades, M., & Brandt, T. (2006). Skew deviation revisited.
Survey of Ophthalmology, 51(2), 105–128.
Demer, J. L. (2006). Current concepts of mechanical and neural factors in ocular motility.
Current Opinion in Neurology, 19(1), 4–13.
Eggers, S. D., Moster, M. L., & Cranmer, K. (2008). Selective saccadic palsy after cardiac
surgery. Neurology, 70(4), 318–320. doi:10.1212/01.wnl.0000287139.01789.97
Fife, T. D. (2010). Overview of anatomy and physiology of the vestibular system. In S. D. Z.
Eggers & D. S. Zee (Eds.), Vertigo and imbalance: Clinical neurophysiology of the
vestibular system (pp. 575). Amsterdam, the Netherlands: Elsevier.

41
Furman, J. M., Wall, C., 3rd, & Pang, D. L. (1990). Vestibular function in periodic alternating
nystagmus. Brain, 113(Pt 5), 1425–1439.
Krauzlis, R. J. (2004). Recasting the smooth pursuit eye movement system. Journal of
Neurophysiology, 91(2), 591–603. doi:10.1152/jn.00801.2003
Leigh, R. J., Robinson, D. A., & Zee, D. S. (1981). A hypothetical explanation for periodic
alternating nystagmus: Instability in the optokinetic-vestibular system. Annals of the New
York Academy of Sciences, 374, 619–635.
Leigh, R. J., & Zee, D. S. (2006). The neurology of eye movements. New York, NY: Oxford
University Press.
Pierrot-Deseilligny, C., Milea, D., & Muri, R. M. (2004). Eye movement control by the cerebral
cortex. Current Opinion in Neurology, 17(1), 17–25.
Ramat, S., Leigh, R. J., Zee, D. S., & Optican, L. M. (2007). What clinical disorders tell us about
the neural control of saccadic eye movements. Brain, 130(Pt 1), 10–35.
Scudder, C. A., Kaneko, C. S., & Fuchs, A. F. (2002). The brainstem burst generator for saccadic
eye movements: A modern synthesis. Experimental Brain Research, 142(4), 439–462.
Sparks, D. L. (2002). The brainstem control of saccadic eye movements. Nature Reviews
Neuroscience, 3(12), 952–964.
Thier, P., & Ilg, U. J. (2005). The neural basis of smooth-pursuit eye movements. Current
Opinion in Neurobiology, 15(6), 645–652. doi:10.1016/j.conb.2005.10.013
Thurtell, M. J., Tomsak, R. L., & Leigh, R. J. (2007). Disorders of saccades. Current Neurology
and Neuroscience Reports, 7(5), 407–416.
Versino, M., Hurko, O., & Zee, D. S. (1996). Disorders of binocular control of eye movements in
patients with cerebellar dysfunction. Brain, 119(Pt 6), 1933–1950.

42
Walker, M. F., & Zee, D. S. (1999). Directional abnormalities of vestibular and optokinetic
responses in cerebellar disease. Annals of the New York Academy of Sciences, 871, 205–
220.
Wong, A. (2007). An update on opsoclonus. Current Opinion in Neurology, 20(1), 25–31.
doi:10.1097/WCO.0b013e3280126b51
Wong, A. M. F. (2008). Eye movement disorders. New York, NY: Oxford University Press.

43
Figure Legends
Figure 2–1. Orbital anatomy. A. The relative position of the five muscles just behind the eyeball.
SR, superior rectus; SO, superior oblique; LR, lateral rectus; MR, medial rectus; IR, inferior
rectus; ON, optic nerve. The plane of this section is shown in (C). B. Top view with the medial
rectus and lateral rectus (shown in the shaded area), which are responsible for horizontal eye
movements. Only a small segment of the superior rectus is shown in order to show the optic
nerve beneath it. C. Similar view with the superior oblique, showing the pulling direction and
axis of rotation. The axis of rotation and pulling direction are similar for the inferior oblique. D.
Axis of rotation of the superior rectus (shaded region) and the inferior rectus located directly
beneath it. The superior and inferior recti mainly move the eye vertically. From Handbook of
Balance Function Testing, Jacobson et al., 1997. Reprinted with permission of Delmar Learning,
a division of Thomson Learning: www.thomsonrights.com.
Figure 2–2. Brainstem structures involved in eye movements. A parasagittal section of the
monkey brainstem shows the location of key structures responsible for saccadic eye movements.
Excitatory burst neurons (EBN) for horizontal saccades lie in the parmedian pontine reticular
formation (PPRF). Inhibitory burst neurons (IBN) for horizontal saccades lie in the medullary
reticular formation (MedRF). EBN for vertical and torsional saccades lie in the rostral interstitial
nucleus of the medial longitudinal fasciculus (riMLF). Some vertical IBN may reside in or close
to the interstitial nucleus of Cajal (INC). EBN and IBN project to ocular motor neurons lying in
the abducens nucleus (VI), trochlear nucleus (IV), and oculomotor nucleus (III). Omnipause
neurons (OPN, indicated by an asterisk) lie in the nucleus raphe interpositus in the midline of the
pons between the rootlets of the abducens nerve (CN VI) and influence the activity of EBN and
IBN. The mesencephalic reticular formation (MRF) may help keep the OPN inhibited until a

44
saccade is complete and the eye is on target. CG: central grey; MB: mammillary body; CN III:
rootlets of the oculomotor nerve; CN IV: trochlear nerve; CN VII: genu of facial nerve; ND:
nucleus of Darkschewitsch; NRTP: nucleus reticularis tegmenti pontis; NPH: nucleus prepositus
hypoglossi: PC: posterior commissure; TR: tractus retroflexus. Reproduced from Ramat et al
(2007) with permission from Dr. Jean Büttner-Ennever and Oxford University Press.
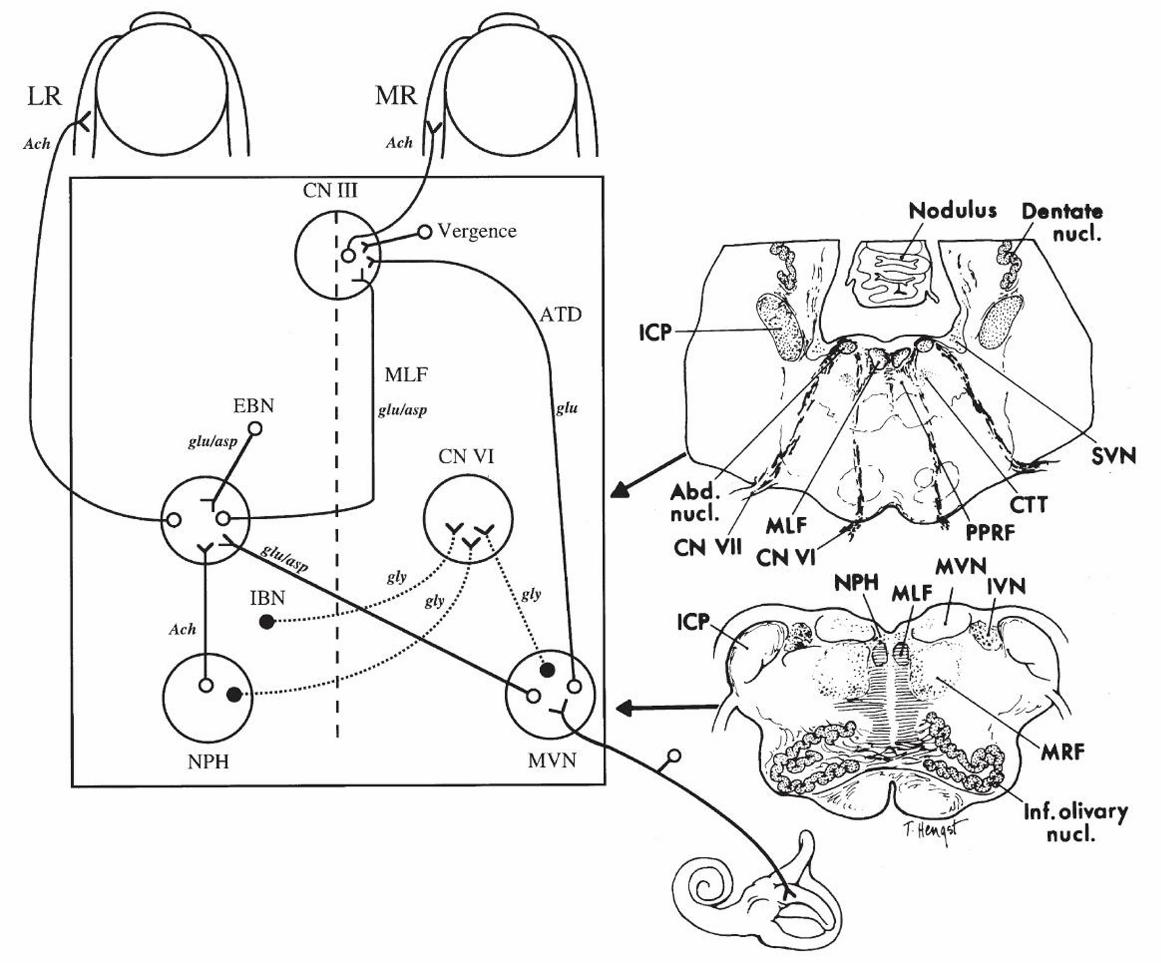
Figure 2–3. Anatomic scheme for the synthesis of signals for horizontal eye movements. The
abducens nucleus (CN VI) contains abducens motor neurons that innervate the ipsilateral lateral
rectus muscle (LR) and abducens internuclear neurons that send an ascending projection up the
contralateral medial longitudinal fasciculus (MLF) to reach the medial rectus (MR) motor
neurons in the contralateral oculomotor nucleus (CN III). From the horizontal semicircular canal,
primary vestibular nerve afferents project mainly to the medial vestibular nucleus (MVN), where
they synapse and then send an excitatory connection to the contralateral abducens nucleus and an
inhibitory projection to the ipsilateral abducens nucleus. Saccadic inputs reach the abducens
nucleus from the ipsilateral excitatory burst neurons (EBN) and contralateral inhibitory burst
neurons (IBN). The neural integrator within the nucleus prepositus hypoglossi (NPH) and
adjacent MVN send eye position information to the abducens nucleus. The medial rectus motor
neurons in CN III also receive commands for vergence eye movements. Putative
neurotransmitters for each pathway are shown: Ach: acetylcholine; asp: aspartate; glu: glutamate;
gly: glycine. The anatomic sections on the right correspond to the level of the arrow heads on the
schematic on the left. Abd. nucl.: abducens nucleus; CN VI: abducens nerve; CN VII: facial
nerve; CTT: central tegmental tract; ICP: inferior cerebellar peduncle; IVN: inferior vestibular
nucleus; Inf. olivary nucl.: inferior olivary nucleus; MRF: medullary reticular formation; SVN:

45
superior vestibular nucleus. Reproduced from Leigh and Zee (2006) with permission from
Oxford University Press.
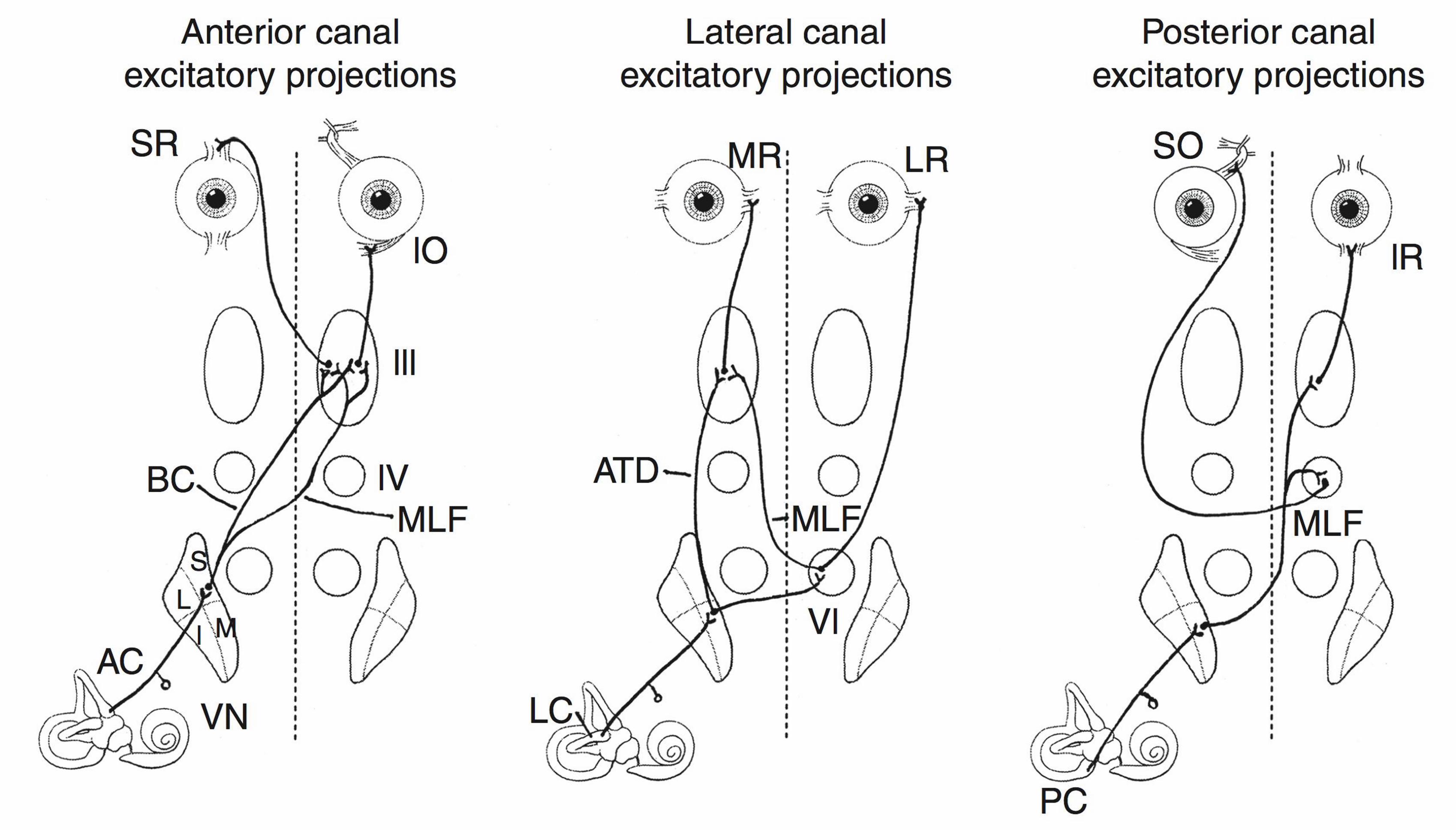
Figure 2–4. Excitatory projections from individual semicircular canals on the right side to the
extraocular muscles. SO: superior oblique; IO: inferior oblique; IR: inferior rectus; LR: lateral
rectus; SR: superior rectus; MR: medial rectus; AC: anterior canal; PC: posterior canal; LC:
lateral canal; MLF: medial longitudinal fasciculus; ATD: ascending tract of Deiters; BC:
brachium conjunctivum; VN: vestibular nuclei (S = superior; I = inferior; L = lateral; M =
medial); III: oculomotor nucleus; IV: trochlear nucleus; VI: abducens nucleus. Reproduced from
Fife (2010) with permission from Elsevier.
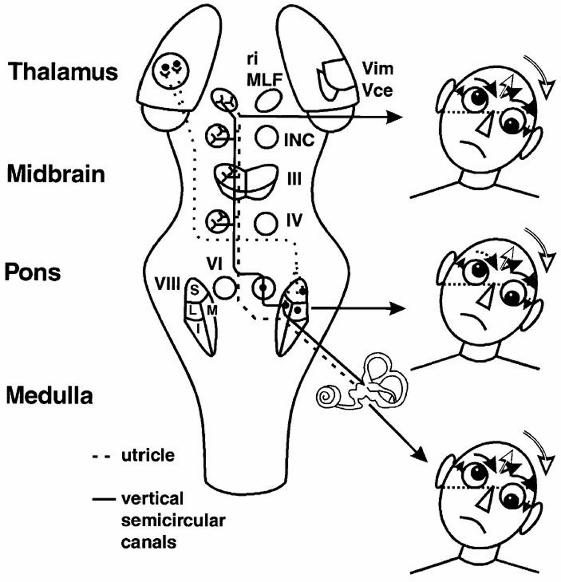
Figure 2–5. Graviceptive pathways from the otoliths and vertical semicircular canals mediating
the vestibular reactions in the roll plane. The projections from the otoliths and the vertical
semicircular canals to the ocular motor nuclei (trochlear nucleus IV, oculomotor nucleus III,
abducens nucleus VI), and the supranuclear centers of the interstitial nucleus of Cajal (INC) and
the rostral interstitial nucleus of the medial longitudinal fasciculus (riMLF), are shown. They
subserve the vestibulo-ocular reflex (VOR) in three planes. The VOR is part of a more complex
vestibular reaction that also involves vestibulospinal connections via the medial and lateral
vestibulospinal tracts for head and body posture control. Ocular tilt reaction is depicted on the
right in relation to the level of the lesion (i.e., ipsiversive with peripheral and pontomedullary
lesions and contraversive with pontomesencephalic lesions). Reproduced from Brodsky et al
(2006) with permission from Elsevier.
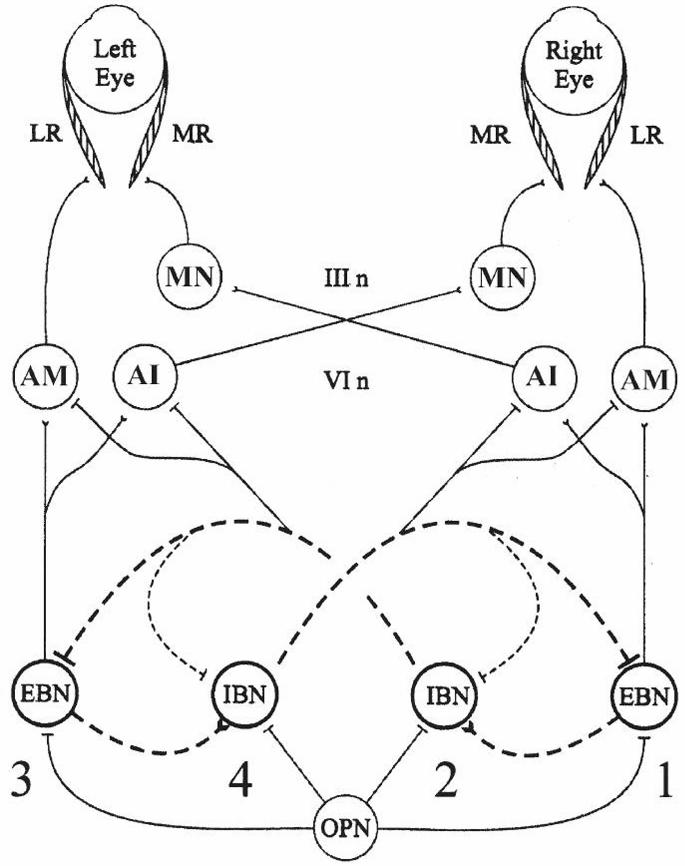
Figure 2–6. A brainstem neural network model for generating horizontal saccades. Projections
with flat ending are inhibitory; the others are excitatory. Saccades require reciprocal innervation
to the medial rectus (MR) and lateral rectus (LR) of both eyes. The LR is driven by the ipsilateral

46
abducens nucleus (VI n) motoneurons (AM). The VI n also contains abducens internuclear
neurons (AI) that send axons to the contralateral oculomotor nucleus (III n), which drive the MR
of the other eye. Excitatory burst neurons (EBNs) provide the saccadic drive to ipsilateral AM
and AI. EBNs also project to inhibitory burst neurons (IBNs). IBNs provide inhibition to the
contralateral AM and AI. Thus, an EBN/IBN pair provides reciprocal innervation. IBNs also
provide inhibition to the contralateral EBNs and IBNs. A consequence of this cross-coupling is
that the EBN/IBN pairs form a short-latency, positive feedback loop. When omnipause neurons
(OPNs) are active, they prevent this loop from oscillating. At the beginning of a saccade, OPNs
cease discharge, allowing one set of EBNs (1) to start firing and activate ipsilateral IBNs (2).
During IBN (2) firing, contralateral EBNs (3) receive a hyperpolarizing input that keeps them
silent. At the end of the saccade, when the IBNs (2) cease firing, the EBNs (3) start to discharge
because of rebound depolarization, which stimulates ipsilateral IBNs (4), which in turn, inhibit
original EBNs (1) that fired. Thus, the EBN/IBN pairs tend to spontaneously oscillate whenever
the OPNs are inhibited and there is no specified saccadic command. Reproduced from Leigh and
Zee (2006) with permission from Oxford University Press.
Figure 2–7. Higher-level control of the saccadic pulse generator. Shown here are the major
structures that project to the brainstem saccade generator (premotor burst neurons in PPRF and
riMLF). Also shown are projections from cortical eye fields to superior colliculus. DLPC,
dorsolateral prefrontal cortex; FEF, frontal eye fields; IML, intramedullary lamina of thalamus;
NRTP, nucleus reticularis tegmenti pontis; PEF, parietal eye field; PPC, posterior parietal
cortex; SEF, supplementary eye field; SNpr, substantia nigra pars reticulata; STN, subthalamic
nucleus. −, inhibition; +, excitation. Reproduced from Leigh and Zee (2006) with permission
from Oxford University Press.

47
Figure 2–8. Neural integrator physiology. For saccades, a pulse of innervation is the input to the
neural integrator that generates a step. If the system is perfect, the pulse (an eye velocity
command) becomes a step (eye position command), as seen in A. If the neural integrator is leaky
or imperfect B., the eye position signal decays with time. In this case, the eye will drift toward
the midline until a corrective saccade repositions the eye on target, creating gaze-evoked
nystagmus. C. The centripetal drift of the eyes that occurs with a leaky integrator can be
described by its time constant (Tc), given by the time at which the eye has drifted 63% of the
way back to the midline. From The Neurology of Eye Movements, by R. J. Leigh. and D. S. Zee,
2006, p. 246, Figure 5–4. Reprinted by permission of Oxford University Press, Inc.
Figure 2–9. Disorders of the saccadic pulse and step. Innervation patterns are shown on the left,
eye movements on the right. Dashed lines indicate the normal response. A. Normal saccade. B.
Hypometric saccade: pulse amplitude (width × height) is too small but pulse and step are
matched appropriately. C. Slow saccade: decreased pulse height with normal pulse amplitude
and normal pulse-step match. D. Gaze-evoked nystagmus: normal pulse, poorly sustained step.
A. Pulse-step mismatch (glissade): step is relatively smaller than pulse. F. Pulse-step mismatch
due to internuclear ophthalmoplegia (INO: the step is larger than the pulse, and so the eye drifts
onward after the initial rapid movement. Reproduced from Leigh and Zee (2006) with
permission from Oxford University Press.
Figure 2–10. Smooth pursuit pathways. Shaded area represents common pathway shared by
horizontal VOR and smooth pursuit. LGN, lateral geniculate nucleus; V1, striate cortex; V2 and
V3, extrastriate cortex; MT, middle temporal visual area; MST, medial superior temporal visual
area; FEFsem, pursuit subregion of the frontal eye field; SEF, supplemental eye field; LIP,
lateral intraparietal area; NOT, nucleus of the optic tract; DLPN, dorsolateral pontine nuclei;

48
MVN, medial vestibular nucleus; LHC, left horizontal canal; NPH, nucleus prepositus
hypoglossi; MLF, medial longitudinal fasciculus; III, oculomotor nucleus; IV, trochlear nucleus;
VI, abducens nucleus; MR, medial rectus; LR, lateral rectus. Reproduced from Wong (2008) with
permission from Oxford University Press.
Figure 2–11. Role of the vestibulocerebellum (flocculonodular lobe and paraflocculus) in the
control of eye movements. The main role of the flocculonodular lobe, adjacent uvula and ventral
paraflocculus (vestibulocerebellum) is the control of certain eye movements. The nodulus and
uvula receive vestibular inputs both directly from the labyrinth and via the vestibular nuclei;
inputs from the frontal eye fields and the middle superior temporal (MST) area via the
dorsolateral pontine nuclei; and inputs from the ocular motor control network via the nucleus
prepositus hypoglossi and the nuclei of the paramedian tracts (not shown). The flocculus and
paraflocculus project to the medial and superior vestibular nuclei predominantly for control of
the horizontal and vertical vestibulo-ocular reflexes and smooth pursuit. Reprinted with
permission from Dr. Eduardo Benarroch (Benarroch, 2006) and Mayo Foundation for Medical
Education and Research.
Figure 2–12. Role of the dorsal vermis and fastigial oculomotor region for control of saccades.
The posterior or dorsal vermis, via the fastigial oculomotor region, controls the amplitude and
direction of saccadic eye movements. These structures receive inputs from the paramedian
pontine reticular formation, which contains the excitatory burst neurons for horizontal saccades,
and from the nucleus reticularis tegmenti pontis, which relays saccadic signals from the frontal
eye fields and superior colliculus. The fastigial nucleus projects to the saccadic burst generator of
the brainstem via the uncinate fasciculus to control the amplitude of ipsilateral and contralateral

49
saccades. Reprinted with permission from Dr. Eduardo Benarroch (Benarroch, 2006) and Mayo
Foundation for Medical Education and Research.
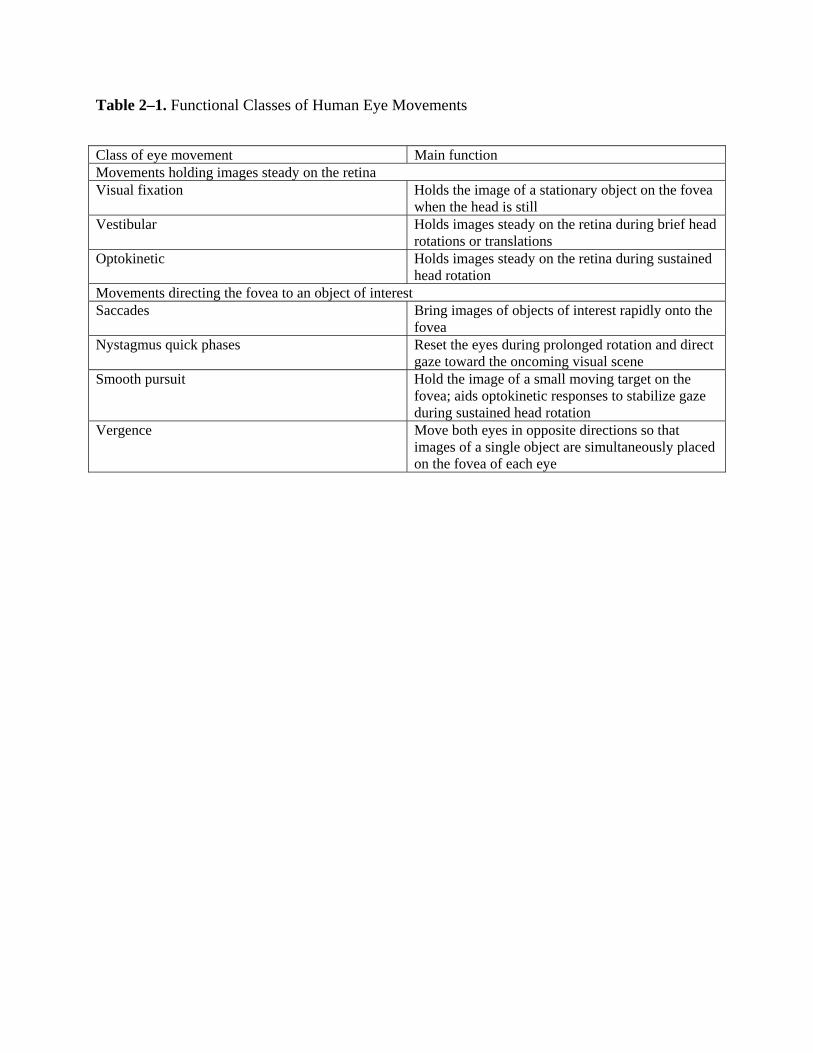
Table 2–1. Functional Classes of Human Eye Movements
Class of eye movement Main function Movements holding images steady on the retina Visual fixation Holds the image of a stationary object on the fovea
when the head is still Vestibular Holds images steady on the retina during brief head
rotations or translations Optokinetic Holds images steady on the retina during sustained
head rotation Movements directing the fovea to an object of interest Saccades Bring images of objects of interest rapidly onto the
fovea Nystagmus quick phases Reset the eyes during prolonged rotation and direct
gaze toward the oncoming visual scene Smooth pursuit Hold the image of a small moving target on the
fovea; aids optokinetic responses to stabilize gaze during sustained head rotation
Vergence Move both eyes in opposite directions so that images of a single object are simultaneously placed on the fovea of each eye
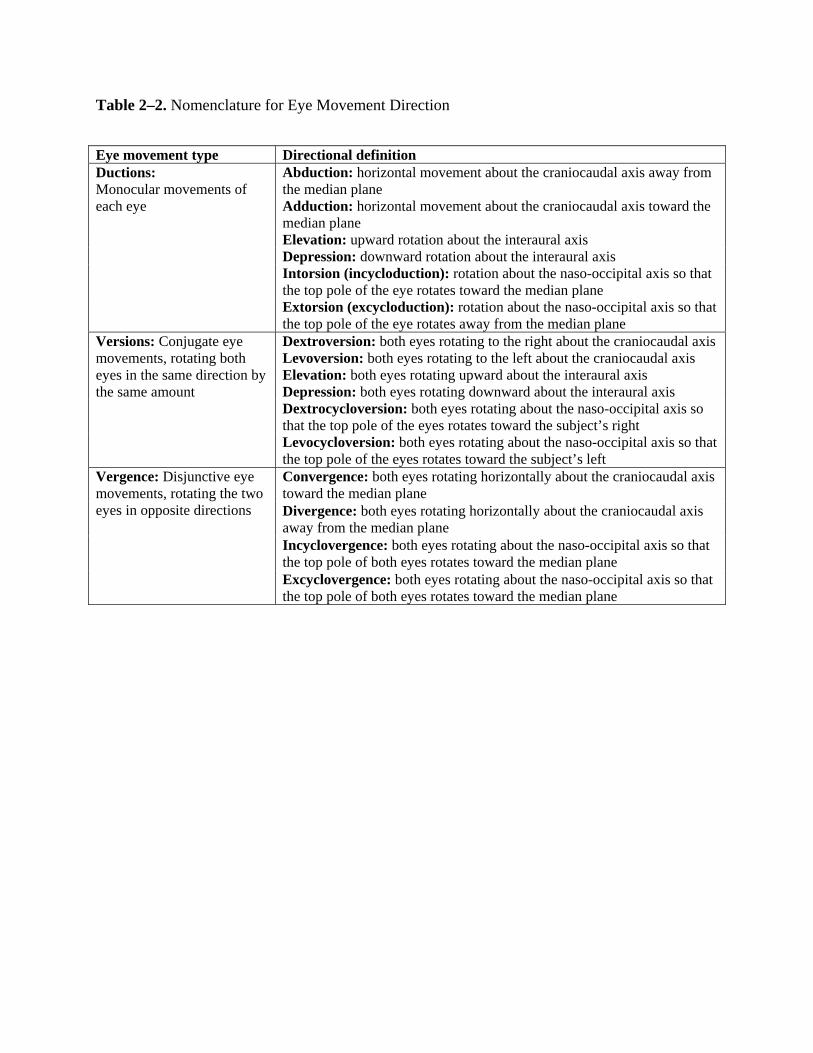
Table 2–2. Nomenclature for Eye Movement Direction
Eye movement type Directional definition Ductions: Monocular movements of each eye
Abduction: horizontal movement about the craniocaudal axis away from the median plane Adduction: horizontal movement about the craniocaudal axis toward the median plane Elevation: upward rotation about the interaural axis Depression: downward rotation about the interaural axis Intorsion (incycloduction): rotation about the naso-occipital axis so that the top pole of the eye rotates toward the median plane Extorsion (excycloduction): rotation about the naso-occipital axis so that the top pole of the eye rotates away from the median plane
Versions: Conjugate eye movements, rotating both eyes in the same direction by the same amount
Dextroversion: both eyes rotating to the right about the craniocaudal axis Levoversion: both eyes rotating to the left about the craniocaudal axis Elevation: both eyes rotating upward about the interaural axis Depression: both eyes rotating downward about the interaural axis Dextrocycloversion: both eyes rotating about the naso-occipital axis so that the top pole of the eyes rotates toward the subject’s right Levocycloversion: both eyes rotating about the naso-occipital axis so that the top pole of the eyes rotates toward the subject’s left
Vergence: Disjunctive eye movements, rotating the two eyes in opposite directions
Convergence: both eyes rotating horizontally about the craniocaudal axis toward the median plane Divergence: both eyes rotating horizontally about the craniocaudal axis away from the median plane Incyclovergence: both eyes rotating about the naso-occipital axis so that the top pole of both eyes rotates toward the median plane Excyclovergence: both eyes rotating about the naso-occipital axis so that the top pole of both eyes rotates toward the median plane
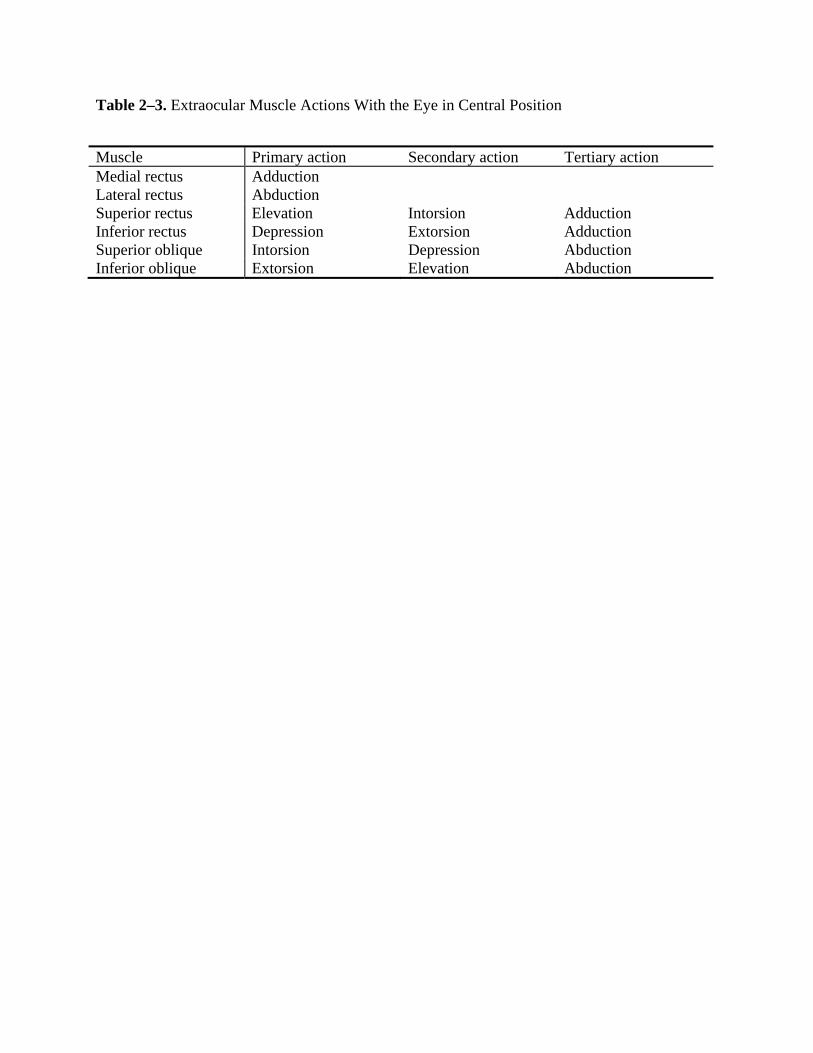
Table 2–3. Extraocular Muscle Actions With the Eye in Central Position
Muscle Primary action Secondary action Tertiary action Medial rectus Adduction Lateral rectus Abduction Superior rectus Elevation Intorsion Adduction Inferior rectus Depression Extorsion Adduction Superior oblique Intorsion Depression Abduction Inferior oblique Extorsion Elevation Abduction
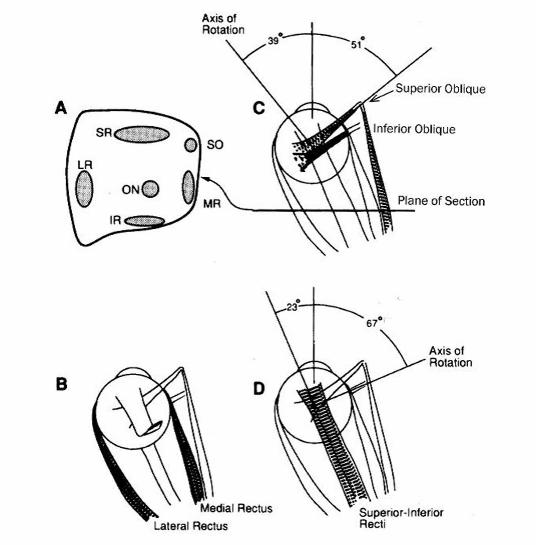
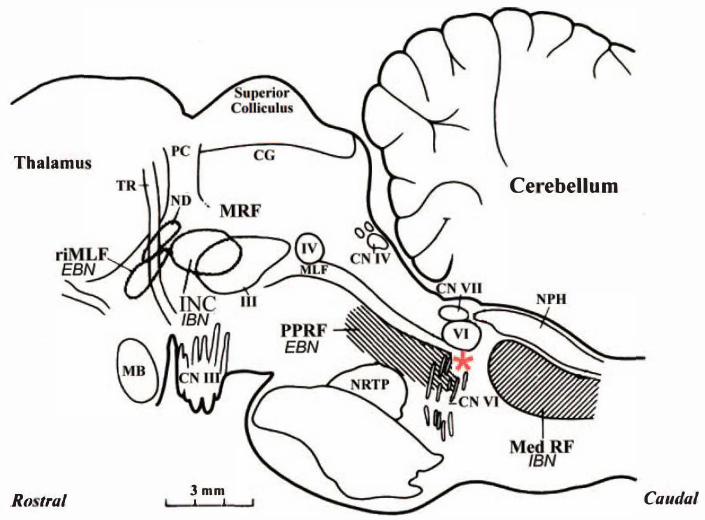
Tbalamu
Ro tra/ 3mm ' '
Ce ebellum
Caudal