Embed Size (px)
Citation preview

ee,g,=on Progress in Neurobiology Vol. 53, pp. 431 to 450, 1997
© 1997 Elsevier Science Ltd. All rights reserved Printed in Great Britain
0301-0082/97/$32.00
Plh S0301-0082(97)00042-7
COGNITIVE PLANNING IN HUMANS: NEUROPSYCHOLOGICAL, NEUROANATOMICAL AND
NEUROPHARMACOLOGICAL PERSPECTIVES
ADRIAN M. OWEN MRC Applied Psychology Unit. 15 Chaucer Road, Cambridge CB2 8EF, U.K.
(Received 9 April 1997)
Abstract--In recent years, considerable progress has been made in understanding the cognitive and neu- roanatomical basis of high-level planning behaviour through a combination of neuropsychological, neuropharmacological and functional neuroimaging approaches. In this article, early evidence suggesting a relationship between planning impairments and damage to the frontal lobe is reviewed and several contemporary studies of planning behaviour in patients with circumscribed frontal lobe excisions are described in detail. These neuropsychological investigations, together with recent functional neuroima- ging studies of normal control subjects, have identified a specific area within the mid-dorsolateral frontal cortex of humans which appears to be critically involved in the cognitive processes that mediate efficient planning. The functions of this region, both in cognitive planning and in related functions such as work- ing memory, are then discussed in the context of a general theoretical framework for understanding the functional organization of "executive" processes within the human lateral frontal cortex.
In the final sections, the relationship between the planning deficits observed after intrinsic frontal lobe damage and those exhibited by patients with neuropathology of primarily sub-cortical origin, such as Parkinson's disease, is discussed. A central model for much of this work has been the concept of cortico- striatal circuitry which emphasizes the relationship between the neocortex and the striatum. The com- bined evidence from comparative studies in patients and from functional neuroimaging studies on Par- kinson's disease suggests that altered cortico-striatal interactions may disrupt normal planning function at a number of levels, possibly consequent upon intrinsic striatal pathology on the one hand and the par- tial loss of (frontal) cortical input to the basal ganglia on the other. © 1997 Elsevier Science Ltd
C O N T E N T S
I. Introduction 2. Neuroanatomical considerations: planning and the frontal lobes 3. Neuropsychological considerations: planning versus working memory 4. Beyond the frontal lobe: cognitive planning and the basal ganglia 5. Conclusions: towards an integrated account of cognitive planning in humans
Acknowledgements References
CBF fMRI GPi HD MR1
ABBREVIATIONS cerebral blood flow MSA functional magnetic resonance imaging PD internal segment of the giobus pallidus PET Huntington's disease rCBF magnetic resonance imaging SPECT
SRO
multiple system atrophy Parkinson's disease positron emission tomography regional cerebral blood flow single photon emission tomography SteeI-Richardson-Olszewski Syndrome
431 432 439 441 447 448 448
I. I N T R O D U C T I O N
The ability to plan, or " th ink ahead" , is a central c o m p o n e n t of many aspects of complex behav iour and is a basic requirement o f many cognitive and mo to r tasks. High-level, or "cogni t ive" , p lann ing may be defined as the ability to organize cognitive behaviour in t ime and space and is necessary in situ- at ions where a goal must be achieved th rough a series of in termediate steps each of which does not necessarily lead directly towards tha t goal.
Cognit ive p lanning is required, therefore, when sequences of responses must be generated and scheduled and when novel courses of act ion must be formula ted and carried out.
The frontal lobes have long been thought to play an impor tan t role in p lann ing behaviour; Fo r example, Harlow, 1868, argued tha t frontal lobe lesions in h u m a n s result in a loss of "p l ann ing skill", whilst much later, Bianchi, 1922, described a loss in the ability to "co-ord ina te the different el- ements of a complex act ivi ty" in monkeys with large
431

432 A.M. Owen
frontal lesions. More contemporary accounts have characterized the role of the frontal cortex in plan- ning behaviour using various, similarly descriptive, terms; e.g., "as a general system for sequencing or guiding behaviour towards the attainment of an im- mediate or distant goal" (Jouandet and Gazzaniga, 1979), or as crucial for the "planning of future actions" (Damasio, 1985; for review, see Shallice, 1988). Until recently, however, the assumed relation- ship between cognitive planning and the frontal lobes lacked solid empirical support and was based largely on anecdotal reports of disorganized beha- viour in patients with relatively non-specific brain injury, or on the behaviour of monkeys with large excisions of the frontal cortex. Moreover, planning difficulties are not unique to patients with circum- scribed frontal lobe damage. For example, "frontal- like" planning deficits have been regularly described in patients with mild Parkinson's disease (Morris et al., 1988; Owen et al., 1992, 1995a), and other basal ganglia disorders, suggesting that an equivalence between the prefrontal cortex and planning function cannot be assumed.
In recent years, however, improved methods of neuropsychological assessment combined with a the- ory-driven approach to task design has led to great advances in our understanding of the fundamental mechanisms which mediate higher-order cognitive processes such as planning. Moreover, the func- tional architecture subserving such processes has been investigated more directly in functional neuroi- maging studies using techniques such as positron emission tomography (PET) and functional mag- netic resonance imaging (fMRI). This article will seek to summarize the progress that has been made recently in understanding the cognitive and neural basis of planning ability through the integration of cognitive, neuropsychological, neurobiological and, most recently, functional neuroimaging approaches.
table and were given a specific goal (to cork a bot- tle). This goal could only be reached by combining a number of discrete steps into a comprehensive plan of action, and then executing each step in the correct order. Fifty patients with unilateral or bilateral fron- tal lobe damage of mixed aetiology exhibited a marked deficit on the task relative to a group of 25 patients with more posterior lesions. In addition, many of the frontal lobe patients reported difficul- ties with planning and structuring their everyday ac- tivities.
Whilst the tasks developed by Klosowska (1976), and earlier by Porteus and Kepner (1944), certainly appear to require cognitive planning, they also have strong visuo-spatial components which may have independently contributed to the deficits described. To overcome this difficulty, Shallice and McCarthy (described in Shallice, 1982), developed the "Tower of London" test, a series of problems thought to depend more heavily on planning than on spatial processing abilities. In this test, subjects are required to move coloured beads between three vertical rods of different lengths in order to match a goal arrangement displayed on three similar rods (see Fig. 1). The difficulty of the problem can be manipu- lated by varying the starting position of the initial arrangement with respect to the goal arrangement.
The Tower of London task clearly requires "for- ward thinking" or planning, since an early, incor- rect, move can render the problem virtually unsolvable unless all previous steps are retraced, and reversed, in order to correct the inappropriate move. Accordingly, successful performance typically involves a number of stages; the overall situation is considered by assessing the inital and goal states with reference to differences in the positions and overall configuration of the balls, a series of sub- goals is defined, a sequence of moves is generated to attain these sub-goals, this sequence is refined and revised according to the results of mental rehearsal
2. NEUROANATOMICAL CONSIDERATIONS: PLANNING AND THE FRONTAL LOBES
In 1935, Wilder Penfield and Joseph Evans, neu- rosurgeons at the Montreal Neurological Institute, described three cases of patients who had sustained extensive neurosurgical excisions of the frontal lobes (Penfield and Evans, 1935). Of particular interest was one young woman who, following surgery, exhibited a marked failure to organize and plan her daily activities. For example, she was unable to plan and prepare an entire family meal, but was, never- theless, perfectly capable of cooking the individual dishes. Subsequently, such disabilites have usually been accounted for in terms of deficits in the cogni- tive processes involved in planning, although rather few studies have addressed this issue directly. An early investigation by Porteus and Kepner (1944), established that, following prefrontal leucotomy, patients were impaired at maze learning, a deficit attributed to a loss of "planning skill". A more direct approach was taken by Klosowska (1976) who developed a novel task that specifically required the development of a plan for successful perform- ance. Subjects were shown a number of objects on a
Fig. 1. A three-move problem from the original Tower of London planning task (adapted from Shallice, 1982). Subjects are required to rearrange the balls in the fore- ground set to match the positions of those in the rear set. In each set, the left peg can hold three balls, the middle peg can hold two balls and the right peg can hold just one
ball.

Cognitive Planning in Humans 433
and, finally, the correct solution is executed. In the original study, reported in Shallice (1982), patients with left anterior cortical pathology were shown to be impaired in the number of moves required to complete the Tower of London problems. This find- ing could not be explained in terms of visuo-spatial factors since the results were unchanged when per- formance was covaried against performance on the spatially demanding Block Design sub-test of the Wechsler Adult Intelligence Scale (WAIS).
Owen et al. (1990), followed up this important finding using a coraputerised version of the Tower of London task from the Cambridge Neuropsychologica] Test Automated Battery (CANTAB), presented on a touch-sensitive screen.*
In the computerized task, the subjects are pre- sented with two se~Ls of three coloured "balls" (i.e. circles), one in the top half of the screen and the other in the bottom half (see Fig. 2). The three balls are distributed in three "pockets" (or "socks"), which can hold one, two, or three balls. On each trial, a red ball, a blue ball and a green ball are placed in predetermined positions in both the upper and the lower pockets of each of the two displays. The subjects are told that the balls in the top half of the screen can not be rearranged, but any ball in the bottom half of the. screen can be moved between pockets by touching it with the index finger of the right hand, and then by touching one of the empty positions in one of the other pockets. Once touched, a tone sounds and the selected ball begins to flash, indicating that it is ready to be moved. When an empty pocket is touched, the selected ball moves, automatically, from its original position to the new one. At any time, the subject can cancel his/her selection of a ball by touching it a second time. Two types of moves are not allowed: a) placing a ball high in a pocket when there is no other ball beneath to support it and b) trying to remove a ball while there is another sitting above it in the same pocket. When such moves are attempted, there is no re- sponse from the computer.
The position of the balls in the top half of the screen is varied with each problem such that a sol- ution can be reached in exactly two, three, four or five moves. The subjects are told to examine the pos- ition of the balls in each problem and to attempt to find the solution that requires the number of moves indicated at the side of the screen. They are encour- aged not to make a first move until they are confi- dent that they can execute the entire sequence needed to solve the problem, thereby maximizing the planning demands of the task. The computer records the number of moves made by the subject to rearrange the balls., which provides the main index of performance, although selection and movement latencies for each move are also recorded.
In general, the two and three-move Tower of London problems require very little cognitive plan- ning, and can be solved using a simple, visual matching-to-sample strategy. This strategy, whereby each ball in the bottom half of the screen is moved
*CeNeS Ltd, 4 Flint Lane, Ely Road, Waterbeach, Cambridge CB5 9QZ.
Fig. 2. The computerized Tower of London planning task from the Cambridge Neuropsychological Test Automated Battery (CANTAB). Subjects are required to move the balls around in the bottom half of the screen to match the goal arrangement in the top half of the screen. As in the original non-computerized task, in each set, the left pocket can hold three balls, the middle pocket can hold two balls and the right pocket can hold just one ball. The example
shown is a five-move problem.
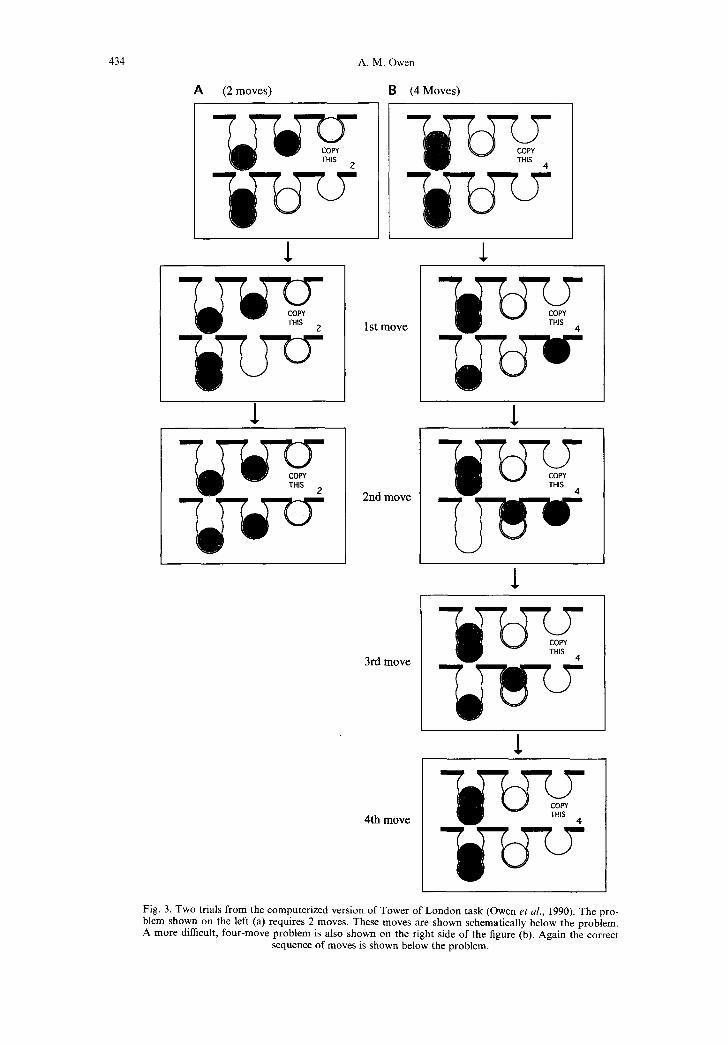
directly to its goal position (i.e. the position corre- sponding to the same coloured ball in the top half of the screen), is illustrated on the left side of Fig. 3 (see Fig. 3a). In contrast, the more difficult four and five-move problems can not be solved using a visual matching-to-sample strategy. In fact, in many cases these problems require the subject to make visually counter-intuitive moves (i.e. to move a ball away from its final destination), in order to execute the appropriate solution, which involves a considerable amount of "thinking ahead" or planning. This more complex task requirement is illustrated on the right side of Fig. 3 (Fig. 3b).
For each test problem, a "yoked" control con- dition is also given to provide a baseline measure of motor initiation and execution times. On each trial of this control condition, the subject is required to follow a sequence of single moves executed by the computer in the top half of the screen by moving the corresponding ball in the lower arrangement. Thus, initially the two arrangements differ by just one ball. Once the subject has made the appropriate move, the top arrangement changes again so that the subject has to make another single move. The test is "yoked" to the main test in the sense that in each trial, the movement of the balls is an exact replication of those moved by the subject in the cor- responding planning trial. By subtracting the re- sponse time from each move in this control condition from that of the corresponding move in the planning condition estimates of initial and sub- sequent "thinking time" are generated. Initial think- ing time refers to the average (per move) time between the presentation of the planning problem and the first touch, minus corresponding motor in- itiation time from the yoked control condition. Subsequent thinking time refers to the time between the selection of the first ball and the completion of the problem minus the average (per move) motor ex- ecution time derived from the corresponding control problem.
434
A (2 m o v e s )
A. M. Owen
8 (4 M o v e s )
1 st m o v e
1
2nd m o v e
l
3rd m o v e
5-
4 th m o v e
Fig. 3. Two trials from the computerized version of Tower of London task (Owen et al., 1990). The pro- blem shown on the left (a) requires 2 moves. These moves are shown schematically below the problem. A more difficult, four-move problem is also shown on the right side of the figure (b). Again the correct
sequence of moves is shown below the problem.

Cognitive Planning in Humans 435
Fig. 4. Twelve slices from a single MRI (one subject), to illustrate the location and extent of the neuro- surgical excision in a representative case. All slices shown are in the horizontal plane and are progress- ively more superior (top left to bottom right). The patient shown sustained a left-sided removal which included the frontopolar region together with the anterior portion of the orbital surface. On the right side of the figure a diagram based on the neurosurgeon's drawing at the time of the operation is pre-
sented. The blackened area defines the estimated lesion site.
Owen et al. (19c~0) assessed performance on this task in 26 neurosurgical patients with unilateral or bilateral frontal lobe excisions and later (Owen et al. , 1995b), in a group of 20 patients with unilateral temporal lobe exci,;ions and a group of 11 patients in whom the more selective, amygdalo-hippocam- pectomy had been performed. An example of a representative frontal lobe excision, examined using high-resolution magnetic resonance imaging (MRI), is given in Fig. 4, together with the neurosurgeon's drawing at the time of the operation.
Compared to age- and IQ-matched controls, the frontal lobe group required more moves to com- plete the problems and produced fewer perfect sol- utions (Fig. 5). Initial "thinking", or "planning" time was unimpaired in these patients although the amount of time s:pent thinking on line (i.e. sub- sequent to the first move) was significantly pro- longed. This pattern of impairment appears to be relatively specific at the cortical level since no defi- cits were observed in the two groups of neurosur- gical patients with damage to the medial temporal lobe region (Owen et al., 1995b).
In a follow-up study (Owen e t al. , 1995a), the Tower of London task was modified to examine the relationship between thinking (planning) time, pro- blem difficulty and solution accuracy in the group of patients with fronlal lobe excisions. Subjects were required to study each of the original Tower of London problems and then to decide how many moves would be required to reach an ideal solution (i.e. with the minimum number of moves), without
actually moving any of the balls. Because this modi- fication required subjects to evaluate and solve the problems in full, without executing any of the necessary sub-goals (i.e. moving the balls), it was no longer possible to compromise "initial planning time" (i.e. the time before a response was made) in favour of "on-line" consideration of the problem during the execution of the solution (i.e. "sub- sequent thinking time"). This modification served to encourage subjects to plan the solution in full, before they initiated a response. The effects of this alteration were clear from the performance of the control subjects in this study compared to that of those studied previously by Owen et al. (1990) using the earlier version of this task. Thus, difficulty- dependent, linear increases in thinking time, which were (during the more challenging four and five move problems), approximately twice as long as those reported previously (e.g., Owen e t al. , 1990), were observed (see Fig. 6). In the frontal lobe patients, the results of the previous study were essentially confirmed; that is, that, compared to the matched control group, the frontal lobe patients were significantly impaired in terms of solution ac- curacy, whilst solution latency (or "initial thinking time"), was relatively preserved (Fig. 6). One might have expected to see prolonged thinking times in the frontal lobe group given those patients' profound difficulty with solving these problems and the fact that prolonged "subsequent thinking" times on the earlier version of this task were reported previously (Owen et al. , 1990). In the previous study however,

436 A.M. Owen
Proportion of problems
s o l v e d perfect ly
1.00-
0.90
0.80
0.70
0.60
0 .50
0 .40
0.30
O n
$ Frontal
T ~ + A H
C, on t~
2 M~ve, 3 . o ~ , M~es s Moves Diff iculty
Fig. 5. Performance data from the Tower of London planning task (Owen et al., 1990). The proportion of problems solved in the minimum number of moves (perfect solutions), as a function of task difficulty for the frontal lobe patients (frontal), the combined temporal lobe and amygdalo-hippocampectomy
patients (temporal + AH), and for the matched group of normal controls. Bars represent S.E.M.
prolonged subsequent thinking time in frontal lobe patients was assumed to reflect the additional time required to revise and refine a solution following an inadequately planned, or impulsive, attempt to solve the problem. Because performance on the modified Tower of London task used in the later study was measured by a single response, the results further suggest that the behaviour of frontal lobe patients in tests that require forward thinking or planning is indeed impulsive; that is, these patients initiate a re- sponse, or make the first move, before they have successfully generated an appropriate solution to the problem, a view consistent with the conclusions of other investigators (e.g., Stuss and Benson, 1984).
The results of these studies demonstrate, there- fore, a significant association between cognitive planning and the frontal cortex in humans. In spite of this evidence, a number of important neuroanato-
mical and neuropsychological issues remain unre- solved. Most importantly, the frontal cortex is not a homogenous region of the brain (see Fig. 7), but comprises several architectonic areas that differ in terms of their connections with other cortical and subcortical areas (Pandya and Barnes, 1987). Relative to the enormous amount of information that is available about the structural and functional organization of the monkey brain, very little is known of the connections between specific cortical areas in humans. In spite of this, a recent reparcella- tion and comparative cytoarchitectonic analysis of the human and macaque frontal cortex has revealed a remarkable degree of topographic and architec- tural similarity between the two species in this region (Petrides and Pandya, 1994; Fig. 7).
In patient studies, however, it is not possible to establish which areas of the frontal cortex are
Z.6"
2.4"
2.2"
8 '~ 2.0-
~ 1.8-
.~ 1.6-
~ 1,4-
1 .2 -
1 .0 -
O.0
50.
-" Fronta l
Control 40 •
e t 20"
10
I I I I I 0
move Z moves 3 moves 4 moves 5 moves
Difficulty
-- Frontal Control
I I I I I
1 move Z moves 3 moves 4 moves 5 moves
Difficulty
Fig. 6. Performance data from the modified Tower of London planning task for frontal lobe patients and a matched group of healthy control subjects (Owen et al., 1995a). On the left, the number of re- sponses needed to identify correctly the number of moves required for the ideal solution at each level of difficulty (planning accuracy), is shown. On the right, the response latency, for correct responses only, at each level of difficulty is shown (planning latency), for the two groups. In both cases, bars represent
S.E.M.

Cognitive Planning in Humans 437
involved in a given cognitive process with any degree of anatomical precision since the excisions are rarely confined to specific cytoarchitectonic areas (e.g., see Fig. 4). In recent years, functional neuroimaging techniques such as single photon emission tomography (SPECT), PET and fMRI, have provided a unique opportunity for assessing the relationship between patterns of cortical and subcortical activation and different aspects of cogni- tive processing in healthy control volunteers. The most widely used blood flow activation techniques use regional cerebrai[ blood flow (rCBF) as an indir- ect index of neuronal (synaptic) activity. Several recent SPECT studies of normal subjects have demonstrated increased cerebral blood flow in the frontal cortex during versions of the Tower of London task (Morris et al. , 1993; Rezai et al. , 1993), further implicating this region in some of the higher- level processes involved in planning. However, the spatial resolution of neuroimaging techniques such as SPECT is inadequate for investigating functional specialization within the human frontal cortex, given the large number of distinct cytoarchitectonic areas within this region (Fig. 7). More recent studies have measured rCBF using PET, which has improved spatial resolution, combined with struc- tural MRI, to achieve a higher degree of precision with respect to neuroanatomical localization. Using PET, rCBF is measured by determining the spatial distribution of a positron-emitting tracer, 150, throughout the brain, during a 60-120 sec time win- dow. Typically, the subject performs the task of
Fig. 7. Schematic drawing of the lateral surface of the macaque brain (a) and the human brain (b), to indicate the location of the dorsolateral frontal cortex (areas 9, 46 and 9/46), and the ventrolLateral frontal cortex (areas 45, 47, 12). Adapted from Petrides and Pandya (1994). sp, sulcus principalis; ifs, inferio:r frontal sulcus; mrs, middle frontal sulcus; sfs, superior frontal sulcus. Numbers refer to cytoarchitectonic area,; as defined by Petrides and Pandya (1994), and based on the original analysis by Brodmann
(1905, 1908, 1909).
interest (e.g., a planning task), in one scan or set of scans and a "control" task requiring many, but not all, of the same motoric, perceptual and cognitive components during another scan or set of scans. The imaging data are then reconstructed, smoothed and normalized for global CBF, which may vary between different scans. The data are then usually transformed into a standardized stereotaxic coordi- nate system based on the three-dimensional atlas of Talairach and Tournoux (1988). The reconstructed, normalized and transformed CBF images are then averaged across all subjects included in a particular study and subtraction images are generated. These images represent the difference between the rCBF during the (planning) task of interest and that during the "control" task. Statistical parametric maps (Friston et al. , 1991), or t-maps (Worsley e t al. , 1993), are then generated and the stereotaxic coordinates of local maxima are calculated within the standardized stereotaxic system.
Owen et al. (1996a), used PET combined with MRI to investigate the role played by specific fron- tal lobe regions in cognitive planning. Planning was examined directly in two functional (PET) scans which required that subjects solve either simple or difficult Tower of London problems. Blood flow during these conditions was compared to that during a control condition which involved identical stimuli and responses but required minimal plan- ning. Thus, by subtracting between the two con- ditions, the blood flow changes related to the planning component of performance could be iso- lated.
When activation in the control condition was sub- tracted from that during the difficult planning con- dition, a significant rCBF change was observed in the left mid-dorsolateral frontal cortex (Fig. 8). In the human brain, the region generally referred to as the dorsolateral frontal cortex comprises mainly cytoarchitectonic areas 9 and 46 which occupy the mid-part of the superior and middle frontal gyri, a considerable proportion of this cortex lying within the depths of the middle frontal sulcus (see Fig. 7).
The significant focal blood flow change in the left dorsolateral frontal cortex during the difficult plan- ning problems was located within area 9, in the lower part of the middle frontal gyrus, just below the intermediate frontal sulcus (see Fig. 8). This change was only statistically significant in the left frontal cortex, although an area of increased blood flow, which failed to reach significance by standard criteria, was observed slightly more anteriorly in the opposite hemiphere. It is important to acknowledge, however, that the apparent emphasis on the left frontal cortex may simply reflect the subtraction method employed rather than a lateralization of function p e r se. Thus, in the present study, both the difficult planning condition and the control con- dition make considerable demands on visuospatial processes which are widely assumed to be preferen- tially mediated by the right hemisphere (Milner, 1971, 1974). The neural correlates of these processes may therefore be "subtracted out" when the two conditions are compared directly. This interpret- ation is entirely consistent with the finding that a change in rCBF, albeit non-significant, was also

438 A.M. Owen
observed in the right mid-dorsolateral frontal cortex during the difficult planning condition, and the fact that planning deficits have been reported in patients with both left and right frontal lobe excisions (Shallice, 1982; Owen et al., 1990, 1995a). Likewise, in a recent PET study employing a modified version of the Tower of London task, similar to that used by Owen et al. (1995a), to study patients, bilateral, but predominantly right, frontal activation was observed (Rogers et al. , 1994; Baker et al., 1996). In that study, subjects were required to imagine and compute the number of required moves mentally, rather than execute the solution itself. This task, which undoubtedly places more emphasis on visual imagery than the task used in the study by Owen et al. (1996a), presumably also places greater demands on right hemisphere regions (Milner, 1971, 1974).
No prefrontal blood flow changes were observed during the simple, three-move planning task in the study by Owen et al. (1996a), although significant areas of activation were observed, in right premotor cortex and bilaterally, in posterior regions of the parietal and occipital lobes. This pattern of rCBF
change is consistent with the fact that these simple three-move problems could be solved using a visual matching-to-sample strategy (unlike the more com- plex four- and five-move planning problems), which, presumably, places a significant load on posterior visual and attentional systems and requires little complex planning or "executive" input from an- terior regions of the frontal lobes. The fact that sig- nificant blood flow changes were observed in areas of premotor cortex during the simple (and the diffi- cult) Tower of London scans, may reflect the more basic processes of "motor planning", as opposed to "cognitive planning", which are an integral com- ponent of any complex task of this type.
In summary, functional neuroimaging studies using PET and MRI have been able to confirm and extend previous investigations in patients by identi- fying, more precisely, the frontal cortical area that appears to be critical for performance on the Tower of London planning task; namely, the mid-dorsolat- eral frontal region. The combined evidence from different functional neuroimaging studies (e.g., Rogers et al. , 1994; Baker et al., 1996; Owen et al. ,
Fig. 8. Difficult (4 and 5 moves) Planning minus Difiicult Control. The average PET subtraction images are shown superimposed upon the corresponding averaged MRI scan for the twelve normal control sub- jects included in the study. The schematic (top left), illustrates the approximate position of the two coro- nal slices shown in the lower half of the figure. Subtraction of two conditions yielded the focal changes in blood flow shown as a t-statistic image, whose range is coded by the colour scale placed to the left of the figure. The sagittal section (top right) at x = - 39 (left hemisphere), illustrates the significant rCBF increase observed in the mid-dorsolateral frontal cortex. This rCBF increase is also visible on the coronal section at y = + 18 along with three additional areas of significant blood flow change in the left presup- plementary motor area, in the right superior premotor cortex and in the left caudate nucleus (see text). The more anterior coronal section at y = + 35 (bottom left), illustrates the rCBF increase in the right mid-dorsolateral frontal cortex, which just missed statistical significance, along with the most anterior
edge of the significant left mid-dorsolateral frontal rCBF change.

Cognitive Planning in Humans 439
1996a), together with previous investigations in patients (e.g., Owen et al., 1990, 1995a), suggests that, within the dorsolateral frontal region, neither hemisphere plays a disproportionate role, at least in the type of high-level planning that is required in the Tower of London task. It is important to emphasize however, that none of the results dis- cussed above suggest that this region of the dorso- lateral frontal cortex is either wholly, or uniquely, involved in mediating cognitive planning processes. In the monkey it has been shown that specific regions of the lateral frontal cortex are reciprocally connected with multiple posterior cortical and sub- cortical regions, which undoubtedly reflects close functional relationships between anatomically dis- tant areas. For example, the ventrolateral frontal region receives visual information directly from the inferotemporal cortex (Kuypers et al., 1965; Jones and Powell, 1970; Chavis and Pandya, 1976; Barbas, 1988; Ungerleider et al., 1989), whilst more posterior parietal cortical areas project preferentially to more dorsal regions within and below the sulcus principa- lis (Schwartz and Goldman-Rakic, 1984; Cavada and Goldman-Rakic, 1989; Petrides and Pandya, 1994). The mid-dorsolaterai region of frontal cortex (areas 9 and 46), which appears to be most critical for cognitive planning is closely connected with the ventrolateral frontal cortex (Barbas and Pandya, 1989; Watanabe-Sawaguchi et al., 1991), and, at the same time, with the limbic region of the medial tem- poral lobe (Adey and Meyer, 1952; Nauta, 1964; Goldman-Rakic et al., 1984). In addition, descend- ing cortico-striatal inputs to the caudate and the pu- tamen project back to discrete frontal lobe regions, including the mi,d-dorsolateral frontal cortex (Middleton and St:rick, 1994, 1995), via various thalamus nuclei, closing the so-called "cortico-stria- tal loops". These neuroanatomical data concur fully with the results of the neuroimaging studies described above. Thus, a number of non pre-frontal cortical and sub-cortical regions were also activated by the planning task used by Owen et al. (1996a), including the caudate nucleus (see Fig. 8), the pre- supplementary motor area, the anterior premotor cortex and the posterior parietal cortex. The avail- able anatomical and functional neuroimaging data suggest, therefore, that whilst the mid-dorsolateral frontal cortex plays a critical role in complex plan- ning behaviour, it does so through close functional interactions with multiple cortical and sub-cortical regions.
3. NEUROPSYCHOLOGICAL CONSIDERATIONS: PLANNING VERSUS
WORKING MEMORY
Due to the inherent complexity of most tasks used to study planning behaviour, the precise cogni- tive components that combine to produce an effi- cient plan of action have not been clearly defined experimentally. The Tower of London task requires an active search of possible solutions and the gener- ation of an appropriate response sequence to solve each of the problems. In addition, the chosen sol- ution must be temporarily stored and transposed
into an appropriate sequence of motor movements, which must be carefully monitored during its ex- ecution. Many of these task requirements place a significant load on active working memory. The term "working memory" was introduced into the experimental psychology literature by Baddeley (1986) to replace the existing concept of a passive short-term memory store and to emphasize, within a single model, both the temporary storage and the "on-line" manipulation of information that occurs during a wide variety of cognitive activities. The re- lationship between planning and various aspects of working memory function has been investigated recently in a large scale, factor analytical study in normal control volunteers (Robbins et al., 1997). In all, 341 subjects were assessed on the computerised Tower of London task and on two tests which tap different aspects of spatial short-term, or working memory. In one test, based on the Corsi Block Tapping Task (described by Milner, 1971), the sub- jects were required to remember sequences of boxes which changed colour on the computer screen (Robbins et al., 1997; for description see Owen et al., 1990). The longest remembered sequence or "spatial span" provided a simple index of the sub- jects ability to retain, transpose and execute a defined sequence of spatial moves, all of which are component processes in the Tower of London task. In another self-ordered "spatial search" test, sub- jects were required to search among a number of boxes on the screen, looking for coloured "tokens" which were hidden by the computer (Robbins et al., 1997; for description see Owen et al., 1990, 1995b, 1996c). Once a token had been found in a box, that box would never again be used to hide a token. Thus, the subject's task was to search for the tokens, trying to remember and avoid those locations where one had previously been found. This task is some- what more complex than the spatial span task, requiring the subject to constantly maintain and update an ongoing representation of the current "state of affairs", which is, again, a central require- ment of the Tower of London planning task. Subjects who perform well on this task tend to do so by adopting a search strategy which involves retracing a systematic "route" and "editing" or "monitoring" those locations along the route where tokens have been found previously. This searching strategy can be captured by an index which is demonstrably uncontaminated by overall mnemonic performance, and yet which correlates highly with such performance (for discussion, see Owen et al., 1990, 1996c). Factor analysis of performance in 341 subjects (Robbins et al., 1997), demonstrated that aspects of performance on all three of these tests (Tower of London, Spatial Span and Self-Ordered Spatial Search), loaded, to varying degrees, on a single factor. Thus, measures of both accuracy and subsequent thinking time on the Tower of London test, error score and strategy score on the spatial search test and spatial span score were all inter-re- lated. This finding clearly suggests that both short- term spatial memory (spatial span), and "strategic" or "organizational" aspects of spatial working mem- ory are important determinants of normal perform- ance on the Tower of London planning task.

440 A.M. Owen
It seems likely that this functional relationship between cognitive planning and aspects of working memory performance reflects some degree of com- monality in the neural circuitry that mediates these different processes. Like cognitive planning, in recent years, considerable evidence has accumulated to suggest that many aspects of working memory involve the lateral surface of the frontal lobe. This evidence comes from the study of patients with exci- sions of frontal cortex (Petrides and Milner, 1982; Owen et al., 1990, 1995a, 1996c; see Petrides, 1989 for review), from lesion and electrophysiologicai recording work in non-human primates (see Goldman-Rakic, 1987 and Petrides, 1994 for reviews), and more recently, from functional neuroi- maging studies in humans (e.g., Jonides et al., 1993; Petrides et al., 1993a,b; McCarthy et al., 1994; Courtney et al., 1996; Gold et al., 1996; Goldberg et al., 1996; Owen et al., 1996a,b; Smith et al., 1995, 1996; Sweeney et al., 1996; for review, see Owen, 1997). On the basis of this, and related evidence, a general theoretical framework regarding the role of the different regions of the lateral frontal cortex in working memory processing and its relationship to other high-level or "executive" functions such as cognitive planning has recently been described (Petrides, 1994). According to this view, there are two executive processing systems within the lateral frontal cortex which mediate different aspects of working memory through reciprocal connections to modality specific posterior cortical association areas. The first stage of interaction between these posterior association areas and frontal regions occurs primar- ily within the ventrolateral frontal cortex (i.e. cytoarchitectonic areas 45 and 47). Thus, these areas (see Fig. 7) are concerned primarily with the active organization of sequences of responses based on conscious, explicit retrieval of information from short-term memory. By contrast, the mid-dorsolat- eral frontal cortex (dorsal area 46 and area 9) is assumed to constitute a second level of interaction of executive processes with memory and is recruited only when the active manipulation and monitoring of information within working memory is required (Fig. 7). This two-stage model of lateral frontal cor- tical function, by which two anatomically and cytoarchitectonically distinct regions of the frontal lobe are linked with different aspects of executive processing, describes how information is both retained and manipulated within working memory to optimize performance on a variety of tasks. In particular, the functional characteristics of the ven- trolateral and dorsolateral frontal processing sys- tems relate very closely to those processes assumed to be involved in the tests of spatial span and spatial working memory described above, which emphasize the short-term retention and execution of sequences of spatial responses on the one hand, and active, "on-line" manipulation of spatial information on the other. A recent functional imaging study com- bining PET and MRI has verified that this is, indeed, the case (Owen et al., 1996b). When normal volunteer subjects performed a modified version of the spatial span task, a significant region of increased cerebral blood flow was observed in ven- trolateral frontal cortex (area 47) in the right hemi-
sphere. No significant changes were observed in more dorsolateral areas of frontal cortex, even when subjects were required to learn and reproduce a "supra-span" sequence of eight boxes. In contrast, when subjects performed a modified version of the spatial search task, significant changes in CBF were clearly observed in the right mid-dorsolateral frontal cortex (areas 46 and 9). Clearly, the emphasis on "strategy" in this task implicates the dorsolateral frontal executive processing system according to the model proposed by Petrides (1994). However, like the spatial span task, this test also places a signifi- cant load on short-term memory for spatial infor- mation. Perhaps unsurprisingly, therefore, the same area of the ventrolaterai frontal cortex (area 47), was also activated in this spatial search task. These results have been confirmed and extended using a number of spatial and non-spatial working memory paradigms (for review, see Owen, 1997).
The results of these functional neuroimaging stu- dies (Owen et al., 1996a,b), combined with the nor- mal control factor analytic study described above (Robbins et al., 1997), clearly suggest that cognitive planning is functionally and structurally related to aspects of working memory. On this basis, one might predict that the Tower of London planning deficits in patients with frontal lobe damage would be accompanied by impairments on tests of working memory function. Neurosurgical patients with fron- tal lobe damage who are impaired on the Tower of London task are also significantly impaired on the spatial searching task described above, making more returns to boxes in which a token has previously been found, even at the simplest levels of task diffi- culty (Owen et al., 1990, 1995a, 1996c). In addition, these patients are less efficient in the use of the re- petitive searching strategy which is known to improve performance on this task, confirming that at least some of their impairment in spatial working memory arises secondarily from a more fundamental deficit in the use of organizational strategies. Although this task is also sensitive to deficits in patients with temporal lobectomy or the more selec- tive amygdalo-hippocampectomy (Owen et al., 1995a, 1996c), unlike the frontal lobe patients, these groups utilize a normal and effective searching strat- egy. Patients with frontal lobe damage are not impaired according to the basic spatial span measure (Owen et al., 1990), although deficits are observed when one considers the number of trials required to reach maximum span (T. W. Robbins et al., unpublished observations). This task is not sen- sitive to unilateral temporal lobe damage, or amyg- dalo-hippocampectomy (Owen et al., 1995b).
In the light of these observations the specific fron- tal lobe impairment in cognitive planning can be more clearly understood. As discussed previously, successful planning on the Tower of London task requires working memory, both for the storage and execution of a correct sequence and also in the search processes required in any analytical problem of this type, by which possible solutions are tried and tested, and then accepted or rejected. Given that frontal lobe patients are specifically impaired in "strategic" aspects of performance on the spatial working memory test, the inappropriate use of or-

Cognitive Planning in Humans 441
S P
. . . . j
qlDOqlDqI! ACH NE DA 5-HT
Fig. 9. A schematic diagram illustrating the intimate anatomical relationship that exists between the frontal cortex and the basal ganglia (adapted from Alexander et al., 1986). Widespread topographically organized cortical projections converge upon the striatum, which is connected to the other basal ganglia structures via "direct" and "indirect" projection systems. The inhibitory direct pathway projects mono- synaptically onto the internal segment of the globus pallidus (Gpi) and the substantia nigra pars reticu- lata (SNr), which are the main output nuclei of the basal ganglia. The indirect pathway projects through the external ,;egment of the globus pallidus (Gpe), and the sub-thalamic nucleus (STN), to the Gpi/SNr complex. The basal ganglia project back to discrete frontal lobe regions via various thalamic nuclei, clos- ing the so called fronto-striatal "loops". The activity of the basal ganglia thalamocortical circuitry is modulated by dopaminergic projections, arising in the substantia nigra pars compacts (SNc). In PD, the most significant structural damage is in the dopamine producing cells of the SNc leading to the degener- ation of the nigro-striatal dopamine system and loss of dopamine in the striatum. PD is also character° ised by degeneration of several other subcortico-cortical projection systems illustrated here. Most notably, the cholinergic (ACH), noradrenergic, (NE), dopaminergic (DA), and serotonergic (5-HT), cor- tical projections from the substantia innominata (SI), the locus coeruleus (LC), the ventral tegmental area (VTA) and the raph~ nucleus, respectively, are affected. Abbreviations: VA-dc, Ventral Anterior, denso-cellula:~ region; VA-mc, Ventral Anterior, magno-cellular region; VA-pc, Ventral Anterior, parvo-
cellular region; MD, Dorso-medial.
ganizational strategies to assess the problems may also explain the deficit observed in the Tower of London planning task.
The results of these behavioural studies suggest an association, therefore, between aspects of cogni- tive planning and working memory. Thus, in normal control subjects, strong inter-relationships are observed between different tasks which emphasize either planning or working memory (Robbins et al., 1997), whilst in frontal lobe patients, deficits are observed in both types of task (Owen et al., 1990, 1995a). Moreover, the results of recent functional neuroimaging studies suggest that planning and cer- tain aspects of working memory function activate the same cortical region within the mid-dorsolateral frontal cortex (Owen et al., 1996a,b). Thus, both functionally and neuroanatomically, it appears to be the second order executive system described by Petrides (1994), that is most closely related to cogni- tive planning; that :is, area 9/46 of the mid-dorsolat- eral frontal cortex, which is recruited when the task being performed requires the active manipulation of
information within working memory. Whether this strong association between both the cognitive basis of, and the neural mediation of, planning and work- ing memory processes represents a single mechanism that is common to both types of process, or an emergent property of the interaction between specialized cognitive sub-systems remains to be determined.
4. BEYOND THE FRONTAL LOBE: COGNITIVE PLANNING AND THE BASAL
GANGLIA
There is now overwhelming evidence that patients with neurodegenerative disorders that affect the basal ganglia nuclei, including Parkinson's disease (PD), Huntington's disease (HD), progressive supra- nuclear palsy (or the Steele-Richardson-Olszewski syndrome, SRO), and multiple systems atrophy (MSA), exhibit diverse patterns of cognitive impair- ment that can include deficits of cognitive planning (Robbins et al., 1992, 1994; Lange et al., 1992;

442 A.M. Owen
0.80
o. ~ o.6o.
E ~ o.4o.
~ 11.2o
o.oo
NMED PO NED PD (rnl~) MED PD (se~,~e) Coat~oi.,I
2Mo~s 3 x ~ m 4 t4oves S M ~
D i f f i cu l t y
1 5 -
1
a)
2~k.. 3 ~ , , k . s~k.. Di f f icu l t~
Fig. 10. Performance data from the Tower of London planning task (Owen et al., 1992). On the left, the proportion of problems solved in the minimum number of moves (perfect solutions), as a function of task difficulty are shown for the non-medicated PD patients (NMED PD), the medicated PD patients with mild clinical symptoms (MED PD (mild)), the medicated PD patients with severe clinical symptoms (MED PD (severe)), and the matched controls. On the right, the average initial thinking times (planning time), prior to the first move as a function of task difficulty are shown. Bars represent one standard error of the difference between the means (calculated according to the formula provided by Cochran and
Cox, 1957).
Owen et al. , 1992, 1995a; Owen and Robbins, 1993; Lawrence e t aL, 1996). A central model for much of this work has been the concept of cortico-striatai loops (Alexander et al. , 1986), which emphasizes the functional inter-relationships between the neocortex and the striatum. According to this model, the wide- spread topographically organized cortical projec- tions which converge upon the striatum, project back, via pallidal, nigral and thalamic structures, to discrete frontal regions (Fig. 9).
Thus, different sectors of the caudate nucleus pro- ject to specific premotor regions such as the sup- plementary motor area or to discrete regions within the dorsolateral and orbitofrontal cortex which are implicated in higher cognitive functions. Furthermore, it is known that damage to different regions of the caudate nucleus produces cognitive deficits which resemble the effects of damage to their corresponding targets of projection within the pre- frontal cortex (Divac e t al. , 1967).
Whilst planning deficits have been reported in MSA (Owen and Robbins, 1993; Robbins e t al. , 1992, 1994), SRO (Owen and Robbins, 1993; Robbins e t al. , 1994), and HD (Lawrence et al. , 1996), the majority of studies in this area have focused on PD, an emphasis that undoubtedly reflects the greater patient availability in this group and the fact that the underlying neuropathology is relatively well established. Taylor et al. (1986), suggested that the principal cognitive deficits in patients with PD occur in tasks that involve "self- directed behavioural planning", although in a later study no deficits were found using a three disk planning problem similar in design to the Tower of London test (Saint-Cyr et al. , 1988). Morris et al. (1988), did find deficits in terms of initial thinking time in a group of 12 medicated patients with mild or moderately severe clinical symptoms using a computerized version of the Tower of London task.
One important consideration in comparing different studies is that PD is a progressive neurodegenera- tive disease and, therefore, like the motor symp- toms, cognitive deficits may be qualitatively and quantitatively different at various stages of the dis- ease process. In addition, medication may play a role given that several studies have now shown that levodopa (L-DOPA) can accentuate or ameliorate different aspects of cognitive function in PD (e.g., Bowen e t al. , 1975; Gotham et al. , 1988; Lange e t al. , 1992). To address some of these issues, Owen et al. (1992), compared three groups of patients at different stages of PD on the same computerized Tower of London test of planning used to assess performance in patients with frontal lobe damage (Owen e t al. , 1990). In general the results lend sup- port to the notion that deterioration of cognitive planning in these patients progresses in parallel with the degeneration of motor functions that characterizes this disorder. Thus, no deficits were observed in de novo patients with mild clinical (i.e. motoric) symptoms. Medicated patients who were still in the early stages of PD also had no deficits in solution accuracy, but were impaired in terms of their initial thinking time, a result which concurs fully with the earlier report by Morris e t al. (1988). Finally, medicated patients with severe clinical symptoms were slower than controls to initiate sol- utions to the planning problems (Fig. 10), and, like the frontal lobe patients, were significantly less accurate at solving the problems.
The fact that patients with mild PD were not impaired in terms of accuracy of solutions contrasts markedly with the severe impairment observed in these patients on other "frontal lobe" tests which, for example, emphasize sorting or attentional set shifting abilities (Owen e t al. , 1992; for similar pat- terns of results, see Lees and Smith, 1983; Downes e t al., 1989; Owen et al. , 1993a,b), and may suggest

Cognitive Planning in Humans 443
2 . 6 -
2.4"
2.2- c
'~= 2.0-
~ 1.8-
¢ 1.6- O
'~ 1.4- ~ '
1.2-
1.0-
0,0
2.6"
¢ NMP Control
I I I I I
1 m o v e 2' m o v e s 3 moves 4 moves S m o v e s
Difficulty
2.4-
2 . 2 " t - O
'~ 2.0" "6 tn 0 1.8"
O~ 1.6" .g ¢1 1.4-
1.2-
1 , 0 "
o.o"1
¢ MP-mild ----0---- Control
I I I I I
1 move 2 moves 3 moves 4 moves 5 moves
Difficulty
2.6"
2.4"
2 . 2 ' c O
= 2.0"
i ~ 1,8"
in ~ 1.6"
1.4"
1.2'
1 . 0 '
o.o
~. Mposevere - - -0 - - - Contro l / 1
I I I I I
1 move 2 moves 3 moves 4 moves 5 moves
Difficulty
Fig. 11. Performance data from the modified Tower of London planning task for the non-medicated PD patients (NMP), the medicated PD patients with mild clinical symp- toms (MP-mild), the medicated PD patients with severe clinical symptoms (MP-severe), and their respective matched control groups (Owen et al., 1995a). The number of responses needed to identify correctly the number of moves required for the ideal solution at each level of diffi- culty (planning accuracy), is shown. Bars represent S.E.M.
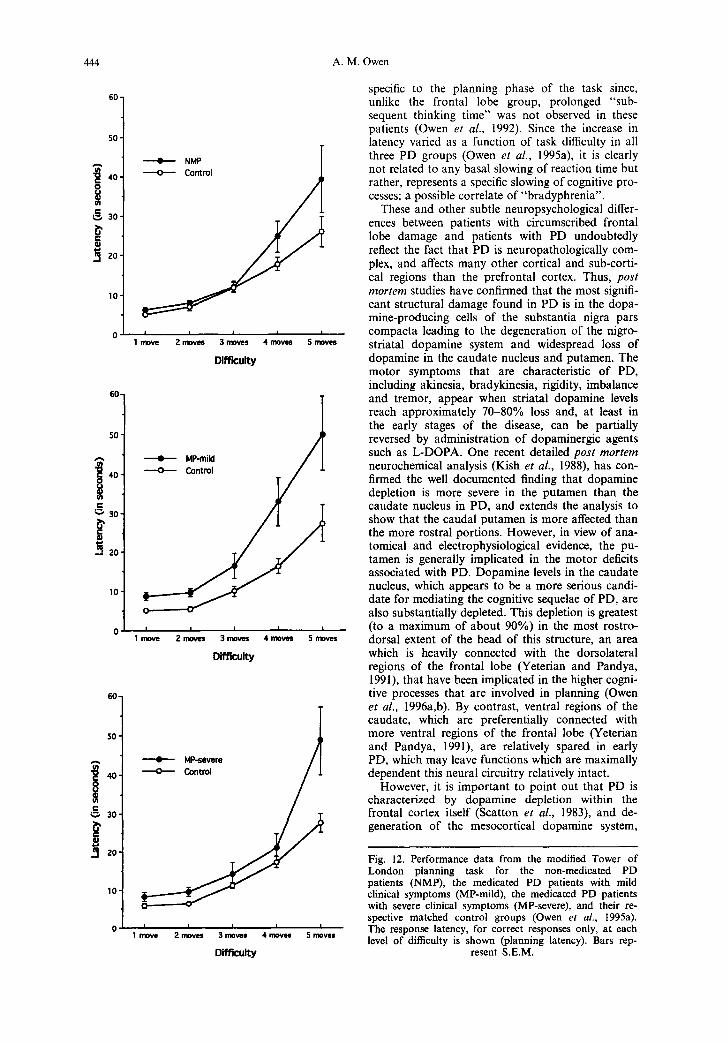
a limited anatomical focus for the cognitive deficits occurring early in the course of PD. An alternative possibility, is that planning may be affected in early PD, but that deficits remain undetected, if the task employed is insufficiently challenging for these patients. This possibility was confirmed in a follow- up study using the operationally more complex ver- sion of the Tower of London task which requires that subjects respond with a single overt response based on the number of moves needed to solve the problem (Owen e t al. , 1995a). Thus, in that study, non-medicated PD patients with mild clinical symp- toms were significantly impaired on this task in terms of both accuracy and latency of performance (Figs 11 and 12).
The results of the follow-up study by Owen e t al. (1995a), also provided new information about the effects of L-DOPA medication in PD and the possi- bility that some aspects of impaired planning per- formance may be more readily ameliorated by L- DOPA medication than others. For example, like the non-medicated PD patients, a group of medi- cated patients with a similar degree of clinical dis- ability were impaired in terms of planning latency, but were completely unimpaired in terms of plan- ning accuracy (Figs 11 and 12).
On the other hand, medicated patients with severe clinical symptoms were impaired in terms of both accuracy and latency measures suggesting that the beneficial effects of L-DOPA medication on cogni- tive performance may decline in more elderly patients or in the later stages of the disease.
In general, the results of these investigations are in agreement with previous studies which have suggested that, like patients with frontal lobe damage, parkinsonian patients are impaired on tasks that involve cognitive planning (Taylor et al. , 1986; Morris et al., 1988), although it is not clear whether this deficit is truly "frontal" in behavioural or neural terms. For example, like frontal lobe patients, in PD, impaired accuracy on the original computerized Tower of London task correlates sig- nificantly with impaired performance on the self- ordered test of spatial working memory described (Owen et al. , 1992). Qualitatively however, the two groups differed in terms of their performance on the spatial working memory task. Thus, whilst the fron- tal lobe group were impaired on the measure of task "strategy", the PD group performed normally in this respect. It seems likely, therefore, that planning ability may be adversely affected by spatial working memory deficits in both frontal lobe patients and in patients with PD, although the precise cognitive mechanisms responsible may not be identical. Behaviourally, the PD and frontal lobe patients also differed in the extent to which their planning beha- viour was characterized by prolonged initial "think- ing", or planning time. Thus, both medicated PD patients with mild clinical symptoms (Morris et al. , 1988; Owen et al., 1992), and non-medicated patients (Owen et al. , 1995a), have been shown to be significantly slower in initiating their solutions to Tower of London problems, although this pattern has not been observed in patients with frontal lobe damage (Owen et al., 1990, 1995a). Moreover, slow- ness of thinking in the PD group, appears to be
444 A.M. Owen
6 0 -
SO
4 0 0
C ~, 30
C
~ zo
1 0
60-
50
i 4 0 -
._c 30-
20-
60-
50-
A
' ~ 40"
C ~ , 30"
ID
2 0 '
lO.
• NMP l
I | I I f
1 move Z moves 3 moves 4 moves 5 m o v e s
Difficulty
MP-mild Control
I I I I I 1move 2 moves 3 m o v u 4 moves 5 moves
Difficulty
¢ MP-severe
(3--- - - J
I I I 1 move 2 moves 3 moves 4 m o v e s
I 5 m o v e s
Difficulty
specific to the planning phase of the task since, unlike the frontal lobe group, prolonged "sub- sequent thinking time" was not observed in these patients (Owen et al., 1992). Since the increase in latency varied as a function of task difficulty in all three PD groups (Owen et al., 1995a), it is clearly not related to any basal slowing of reaction time but rather, represents a specific slowing of cognitive pro- cesses; a possible correlate of "bradyphrenia".
These and other subtle neuropsychological differ- ences between patients with circumscribed frontal lobe damage and patients with PD undoubtedly reflect the fact that PD is neuropathologically com- plex, and affects many other cortical and sub-corti- cal regions than the prefrontal cortex. Thus, post mor tem studies have confirmed that the most signifi- cant structural damage found in PD is in the dopa- mine-producing cells of the substantia nigra pars compacta leading to the degeneration of the nigro- striatal dopamine system and widespread loss of dopamine in the caudate nucleus and putamen. The motor symptoms that are characteristic of PD, including akinesia, bradykinesia, rigidity, imbalance and tremor, appear when striatal dopamine levels reach approximately 70-80% loss and, at least in the early stages of the disease, can be partially reversed by administration of dopaminergic agents such as L-DOPA. One recent detailed post mor tem neurochemical analysis (Kish et al., 1988), has con- firmed the well documented finding that dopamine depletion is more severe in the putamen than the caudate nucleus in PD, and extends the analysis to show that the caudal putamen is more affected than the more rostral portions. However, in view of ana- tomical and electrophysiological evidence, the pu- tamen is generally implicated in the motor deficits associated with PD. Dopamine levels in the caudate nucleus, which appears to be a more serious candi- date for mediating the cognitive sequelae of PD, are also substantially depleted. This depletion is greatest (to a maximum of about 90%) in the most rostro- dorsal extent of the head of this structure, an area which is heavily connected with the dorsolateral regions of the frontal lobe (Yeterian and Pandya, 1991), that have been implicated in the higher cogni- tive processes that are involved in planning (Owen et al., 1996a,b). By contrast, ventral regions of the caudate, which are preferentially connected with more ventral regions of the frontal lobe (Yeterian and Pandya, 1991), are relatively spared in early PD, which may leave functions which are maximally dependent this neural circuitry relatively intact.
However, it is important to point out that PD is characterized by dopamine depletion within the frontal cortex itself (Scatton et al., 1983), and de- generation of the mesocortical dopamine system,
Fig. 12. Performance data from the modified Tower of London planning task for the non-medicated PD patients (NMP), the medicated PD patients with mild clinical symptoms (MP-mild), the medicated PD patients with severe clinical symptoms (MP-severe), and their re- spective matched control groups (Owen et aL, 1995a). The response latency, for correct responses only, at each level of difficulty is shown (planning latency). Bars rep-
resent S.E.M.

Cognitive Planning in Humans 445
Difficult Planning minus Vlsuomotor Control
to
l e f t mid domola taml frontal cortex Right mid dorlmlitarld frontal cortax
~ f f l ~ l m ~ In normal lzNI
-2. -2
-IO -lO
-12 -1~' PO P=~nm ;~,,-mc~hed Commls PO P,die~s AOe-nwl~.xl Oontm~
Group
Fig. 13. Difference in mean blood flow (ml/100 g/min) for PD patients and for control subjects in the mid-dorsolateral frontal cortex. Mean normalized blood flow values measured in ml/100 g/min were extracted for each scanning condition, from the six patients, and separately, from the six control sub- jects, in both left and right hemispheres, using 5 mm diameter regions of interest centred around the stereotaxic coordinates of the highest peak of activation identified for each group, within each subtrac-
tion.
which projects to the frontal lobes and other cortical areas, may also play a significant role in the occur- rence of frontal-like planning deficits in PD. However, this system is known to be less severely affected (50% depletion) than the nigrostriatal dopa- mine system in PD (Agid et al., 1987) and possibly at a later stage of the disease process. In short, two possible mechanisms exist whereby dopamine de- pletion might affect frontal lobe cognitive functions in PD; via disruption of one or more of the corticos- triato-cortical circuits at the level of the striatum or via direct depletion of dopamine in regions of the prefrontal cortex innervated by the ventral tegmen- tal area. In addition, however, the presence of a range of non-doparainergic forms of pathology in PD, including noradreuergic, serotonergic and cholinergic deafferentation of the cortex (Agid et aL, 1987), suggests that some of the cognitive deficits observed may arise: from one or more of these alternative forms of pathology. Finally, cortical Lewy bodies may also be implicated in PD (Byrne et al., 1989; G i b b e t al., 1989), although these are most evident in the later stages of the disease and cannot, therefore, account for the profound deficits observed in non-medicated patients with mild clinical symp- toms.
Slowed "thinking" in patients with PD, which was not seen in patients with frontal lobe damage has been interpreted in a number of ways (Rogers, 1986; Morris et al., 1988, 1990; Sagar et al., 1988; Watts et al., 1988; for discussion, see Owen et al., 1992), although little is known about the neural sub- strates responsible. The lack of effect of L-DOPA and of frontal lobe damage on thinking times (Owen et aL, 1995a), suggests that neither the dopa- minergic mechanisms in the striatum nor the pre- frontal cortex itself, mediate this aspect of planning ability. It seems likely, therefore, that slowed think- ing in PD may arise from one or more of the alternative non-dopaminergic forms of pathology described above.
On the other hand, the fact that accuracy of plan- ning appears to be selectively ameliorated by dopa- minergic therapy in patients with early PD (Owen et aL, 1995a), suggests that the "frontal-like" aspects of performance are indeed related to the central dopaminergic deficit in these patients. A critical question, however, is whether these effects are mediated primarily through cortical or sub-cortical dopaminergic changes; that is, whether they reflect striatal dopamine depletion and its likely affect on the normal flow of information through frontal- striatal circuitry or whether they are the result of disease related depletion of dopamine within the frontal cortex itself.
In a recent study, PET was used to examine whether blood flow in the prefrontal cortex and in the striatum may be differentially affected in PD, during tests of planning and working memory (Owen et al., 1996c). Regional cerebral blood flow was measured in six patients with moderate PD and in six age-matched controls while subjects performed easy and difficult versions of a modified Tower of London planning task and a control condition that involved identical visual stimuli and motor re- sponses (Owen et al., 1996d). In the six age-matched control subjects, significant increases in rCBF were observed in the dorsolateral, ventrolateral and pre- motor areas of the left frontal lobe, in the ventral frontal, premotor, posterior parietal and prestriate cortices of the right hemisphere and in the striate cortex at the midline. In PD patients, significant changes in rCBF were also observed in dorsolateral, ventrolateral and premotor regions of the right fron- tal lobe when blood flow in the visuomotor control conditon was subtracted from that in the difficult planning condition. An increase was also observed in the left mid-dorsolateral frontal region (--46.9, 23.56, 27.0, t = 2.80), although this failed to reach significance according to conventional statistical cri- teria. Significant changes were also seen in the right posterior parietal cortex and in two regions of the

446 A. M. Owen
left prestriate cortex in these patients (Owen et al., 1996d).
When the patient and the control groups were compared directly, no significant difference in acti- vation pattern was observed in the prefrontal cortex. Since previous studies in normal control subjects (Baker et al., 1996; Owen et al., 1996a), had specifi- cally implicated the mid-dorsolateral frontal region in planning, an exploratory qualitative analysis was conducted in this area. Mean normalized blood flow values (ml/100 g/min) were extracted for both left and right hemispheres, using 5 mm diameter regions of interest around the highest peak of activation identified for each subject group, within each sub- traction (Fig. 13). Similar rCBF changes were observed in the mid-dorsolateral region of the fron- tal cortex in the control subjects and in the patients with PD.
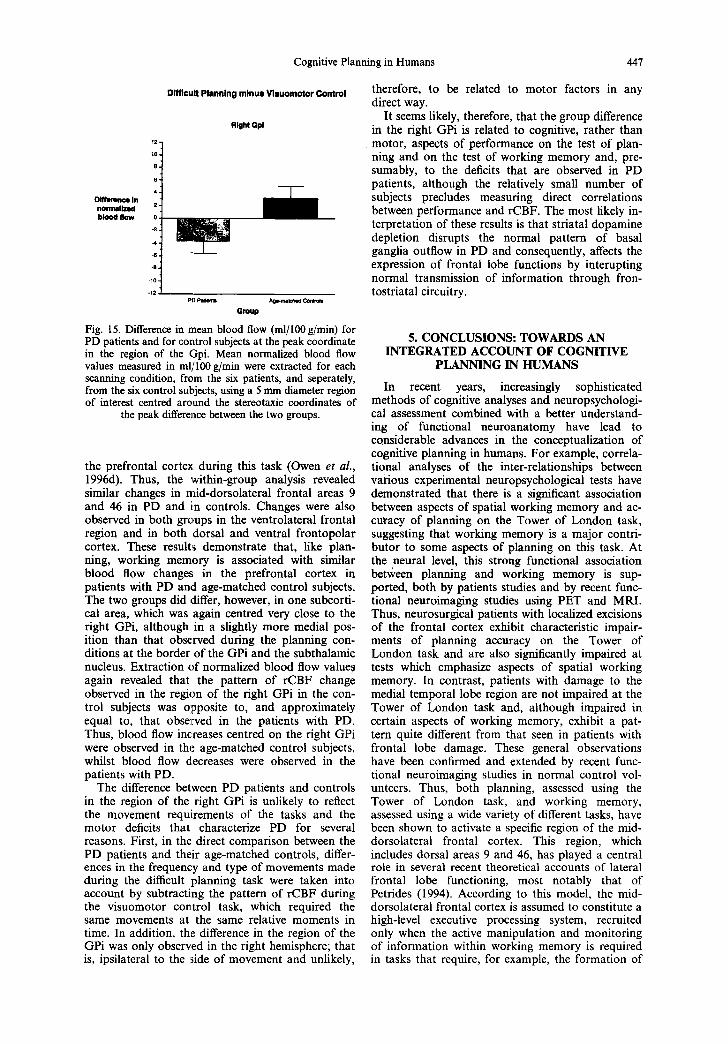
The groups did differ, however, in one subcortical area centred on the right internal segment of the globus pallidus (GPi) (Fig. 14). Thus, when com- pared directly, there was a significant difference between the PD patients and the control subjects in
the rCBF change observed in this region during the planning task.
The GPi constitutes the main basal ganglia out- flow nucleus by which descending cortico-striatal inputs project back to discrete frontal regions, including the mid-dorsolateral frontal cortex (Middleton and Strick, 1994, 1995), via the thala- mus, closing the so-called "cortico-striatal loops". Extraction of mean normalized rCBF values in this region, revealed that, relative to the visuomotor con- trol task, the difficult planning task was associated with an increase in controls, but a decrease in PD patients (Fig. 15). The same rCBF pattern (increase in controls, reduction in PD) was not observed in any other cortical or subcortical area examined, including the mid-dorsolateral frontal cortex, which is known to be involved in these cognitive tasks.
This result is strengthened by the results of a par- allel study in which the same six patients and age- matched controls were compared on a test of spatial working memory known to activate both dorsal and ventral frontal lobe regions (Owen et al., 1996a). When the patients and the controls were compared directly, no significant differences were observed in
Fig. 14. The averaged PET subtraction images are shown superimposed upon the corresponding aver- aged MRI scan of all 12 subjects participating in the study. Direct comparisons between the six patients and the six control subjects yielded the focal difference in blood flow shown as a t-statistic image, whose range is coded by the colour scale placed to the right of the figure. In the coronal section, the y coordi- nate represents the position relative to the anterior commissure (positive = anterior) and has been cho- sen to illustrate the statistically significant difference in the region of the right GPi, between the control subjects and the PD patients when the difficult planning condition was compared to the visuomotor con- trol condition. The same peak is also shown in the horizontal plane (z = - 3). The schematic (bottom right), illustrates the approximate position of the two MRI slices shown. On the left of the figure, identi- cal slices are shown from a high resolution MRI of a single (normal control) subject, to assist anatomical
localization of the significant blood flow change to the Gpi.
Cognitive Planning in Humans 447
Diff icul t P lanning m|nua V lsuomoto r Control
RUght a~
Dlffenmoe In normlnzed blood flow
P[) ~ Age-ffUlld~d C4:nVOM Group
Fig. 15. Difference in mean blood flow (ml/100 g/min) for PD patients and for control subjects at the peak coordinate in the region of the Gpi. Mean normalized blood flow values measured in ml~rl00 g/min were extracted for each scanning condition, from the six patients, and seperately, from the six control subjects, using a 5 mm diameter region of interest centred around the stereotaxic coordinates of
the peak difference between the two groups.
the prefrontal cortex during this task (Owen et al., 1996d). Thus, the within-group analysis revealed similar changes in mid-dorsolateral frontal areas 9 and 46 in PD and i:n controls. Changes were also observed in both groups in the ventrolateral frontal region and in both dorsal and ventral frontopolar cortex. These results demonstrate that, like plan- ning, working memory is associated with similar blood flow changes in the prefrontal cortex in patients with PD and age-matched eontrol subjects. The two groups did ,:lifter, however, in one subcorti- cal area, which was again centred very close to the right GPi, although in a slightly more medial pos- ition than that observed during the planning con- ditions at the border of the GPi and the subthalamic nucleus. Extraction of normalized blood flow values again revealed that the pattern of rCBF change observed in the region of the right GPi in the con- trol subjects was opposite to, and approximately equal to, that observed in the patients with PD. Thus, blood flow increases centred on the right GPi were observed in the age-matched control subjects, whilst blood flow decreases were observed in the patients with PD.
The difference between PD patients and controls in the region of the right GPi is unlikely to reflect the movement requirements of the tasks and the motor deficits that characterize PD for several reasons. First, in the direct comparison between the PD patients and their age-matched controls, differ- ences in the frequency and type of movements made during the difficult planning task were taken into account by subtracting the pattern of rCBF during the visuomotor control task, which required the same movements at the same relative moments in time. In addition, the difference in the region of the GPi was only observed in the right hemisphere; that is, ipsilateral to the side of movement and unlikely,
therefore, to be related to motor factors in any direct way.
It seems likely, therefore, that the group difference in the right GPi is related to cognitive, rather than motor, aspects of performance on the test of plan- ning and on the test of working memory and, pre- sumably, to the deficits that are observed in PD patients, although the relatively small number of subjects precludes measuring direct correlations between performance and rCBF. The most likely in- terpretation of these results is that striatal dopamine depletion disrupts the normal pattern of basal ganglia outflow in PD and consequently, affects the expression of frontal lobe functions by interupting normal transmission of information through fron- tostriatal circuitry.
5. CONCLUSIONS: TOWARDS AN INTEGRATED ACCOUNT OF COGNITIVE
PLANNING IN HUMANS
In recent years, increasingly sophisticated methods of cognitive analyses and neuropsychologi- cal assessment combined with a better understand- ing of functional neuroanatomy have lead to considerable advances in the conceptualization of cognitive planning in humans. For example, correla- tional analyses of the inter-relationships between various experimental neuropsychological tests have dehaonstrated that there is a significant association between aspects of spatial working memory and ac- curacy of planning on the Tower of London task, suggesting that working memory is a major contri- butor to some aspects of planning on this task. At the neural level, this strong functional association between planning and working memory is sup- ported, both by patients studies and by recent func- tional neuroimaging studies using PET and MRI. Thus, neurosurgical patients with localized excisions of the frontal cortex exhibit characteristic impair- ments of planning accuracy on the Tower of London task and are also significantly impaired at tests which emphasize aspects of spatial working memory. In contrast, patients with damage to the medial temporal lobe region are not impaired at the Tower of London task and, although impaired in certain aspects of working memory, exhibit a pat- tern quite different from that seen in patients with frontal lobe damage. These general observations have been confirmed and extended by recent func- tional neuroimaging studies in normal control vol- unteers. Thus, both planning, assessed using the Tower of London task, and working memory, assessed using a wide variety of different tasks, have been shown to activate a specific region of the mid- dorsolateral frontal cortex. This region, which includes dorsal areas 9 and 46, has played a central role in several recent theoretical accounts of lateral frontal lobe functioning, most notably that of Petrides (1994). According to this model, the mid- dorsolateral frontal cortex is assumed to constitute a high-level executive processing system, recruited only when the active manipulation and monitoring of information within working memory is required in tasks that require, for example, the formation of

448 A.M. Owen
sequences of actions or "plans". The extent to which this proposed functional and structural as- sociation between planning and working memory represents the operation of a single common mech- anism, or alternatively, emergent properties of inter- actions between specialized cognitive sub-systems remains to be determined.
Whilst the findings discussed above clearly suggest a critical role for the mid-dorsolateral frontal cortex in cognitive planning, it is now quite clear that this executive influence is mediated through close func- tional connections with multiple cortical and sub- cortical regions. In view of the intimate circuitry linking the striatum to the prefrontal cortex, via par- allel, segregated cortico-striatal loops that feed back via the pallidum and the thalamus to frontal cortex (Alexander et aL, 1986), it seems likely that both cortical and subcortical components of this distribu- ted system are required for efficient planning to occur. This suggestion is supported fully, by the results of recent functional neuroimaging studies which have shown specific areas of activation in the basal ganglia during high-level planning and, par- ticularly, by recent comparative neuropsychological studies of patients with circumscribed frontal lobe excisions and patients with PD. Much of this work has sought to verify the "frontal" nature of cogni- tive deficits in PD and, on the whole, experimental results have supported such a model. However, that is not to say that cognitive deficits arising from striatal dysfunction are identical to those seen fol- lowing damage to associated frontal regions. In fact, when task demands are subjected to careful process analysis, subtle but important differences emerge between frontal lobe patients and patients with PD suggesting that the study of these different disorders may enable us to dissociate different aspects o f task performance which may depend upon different el- ements within functional neural loops. Accuracy and efficiency of planning for example, appears to be mediated by mechanisms within the fronto-stria- tal system whilst speed of thinking may be modu- lated by non-dopaminergic neurotransmitter systems of subcortical origin that innervate either or both the cortex and the caudate nucleus. In normal sub- jects these facets o f performance are in balance, pro- ducing optimal trade-off between speed and accuracy. However, this balance appears to be dis- rupted in patients with PD or frontal lobe damage, though in rather different ways, possibly reflecting altered cortico-striatal interactions consequent upon striatal dopamine depletion or the partial loss of cortical input to the striatum, respectively.
In future, increasingly sophisticated functional imaging techniques such as PET and f M R I will sup- plement such comparisons between groups o f patients with different pathology of the cortico-stria- tal circuitry and provide a mechanism by which the neural underpinnings of some of the deficits described can be more clearly defined. For example, a recent activation study using PET has sought to examine how blood flow in the frontal cortex and in the basal ganglia may be affected in PD, during two tests of planning and working memory that are known to recruit fronto-striatal circuitry. The results clearly suggest that "frontal-l ike" deficits in PD are
the result, not of frontal lobe abnormalities per se, but rather of dopamine depletion at the level of the basal ganglia which disrupts the normal trans- mission of information through fronto-striatal cir- cuitry. Such investigations, when combined with information derived from cognitive psychology, neu- ropsychology and neurobiology can only improve our understanding of the relationship between brain and behaviour and help to further refine the way we think about planning, working memory and other complex cognitive processes in terms of the precise mechanisms involved.
Acknowledgements--During the preparation of this manu- script the author was supported by a Pinsent-Darwin Research Scholarship at the University of Cambridge. The author would like to thank Drs T. W. Robbins, B. J Sahakian, M. Petrides, and J. Doyon who collaborated on much of the work presented in this review.
R E F E R E N C E S
Adey, W. R. and Meyer, M. (1952) An experimental study of hip- pocampal afferent pathways from prefrontal and cingulate areas in the monkey. J. Anat. 86, 58-75.
Agid, Y., Ruberg, M., Dubois, B. and Pillon, B. (1987) Anatomoclinical and biochemical concepts of subcortical demen- tia. In: Cognitive Neurochemistry, pp. 248-271. Eds S. M. Stahl, S. D. Iversen and E. C. Goodman~ Oxford University Press: Oxford.
Alexander, G. E., Delong, M. R. and Strick, P. L. 0986) Parallel organisation of functionally segregated circuits linking basal ganglia and cortex. A. Rev. Neurosci. 9, 357-381.
Baddeley, A. D. (1986) Working Memory. Oxford University Press, New York.
Baker, S. C., Rogers, R. D., Owen, A. M., Frith, C. D., Dolan, R. J., Frackowiak, R. S. J. and Robbins, T. W. (1996) Neural sys- tems engaged in planning: A PET study of the Tower of London Task. Neuropsychologia 34(6), 515-526.
Barbas, H. (1988) Anatomic organization of basoventral and med- iodorsal visual recipient prefrontal regions in the rhesus monkey. J. Comp. Neurol. 276, 313-342.
Barbas, H. and Pandya, D. N. 0989) Architecture and intrinsic connections of prefrontal cortex in rhesus monkey. J. Comp. Neurol. 286, 353-375.
Bianchi, L. (1922) The Mechanism of the Brain and the Function of the Frontal Lobes. Livingstone, Edinburgh.
Bowen, F. P., Kamienny, M. A., Burns, M. M. and Yahr, M. D. (1975) Parkinsonism: Effects of Levodopa treatment on con- cept Formation. Neurology 25, 701--704.
Brodmann, K. (1905) Beitraege zur histologischen Lokalisation der Grosshirnrinde. III Mineilung. Die Rinenfelder der neideren Alien. J. Psychol. Neurol. 4, 177-226.
Brodmann, K. (1908) Beitraege zur histologiscben Lokalisation der Grosshirnrinde. VI Mitteilung. Die Cortexgliederung des Menscen. J. Psychol. Neurol. 10, 231-246.
Brodmann, K. (1909) Vergleichende Lokalisation der Grosshirnrinde in ihren Prinzipien Dargestellt auf Grand des Zellenbaues. Barth, Leipzig.
Byrne, E. J., Lennox, G., Lowe, J. and Godwin-Austin, R. B. (1989) Diffuse Lewy body disease: clinical features in 15 cases. J. Neurol. Neurosurg. Psychiat. 52, 709-717.
Cavada, C. and Goldman-Rakic, P. S. 0989) Posterior parietal cortex in rhesus monkey: II. Evidence for segregated corticoeor- tical networks linking sensory and limbic areas with the frontal lobe. J. Comp. Neurol. 287, 422-445.
Chavis, D. A. and Pandya, D. N. (1976) Further observations on cortieo-frontal connections in the rhesus monkey. Brain Res. ll7, 369-386.
Cochran, W. G. and Cox, G. M. (1957) Experimental Designs. Wiley, New York.
Courtney, S. M., Ungerlieder, L. G., Keil, K. and Haxby, J. V. (1996) Object and spatial visual working memory activate

Cognitive Planning in Humans 449
separate neural systems in human cortex. Cerebral Cortex 6, 39- 49.
Damasio, A. R. (1985) The frontal lobes. In: Clinical Neuropsychology, 2nd Edn. Eds K. M. Heilman and E. Valenstein. Oxford University Press, New York.
Downes, J. J., Roberts, A. C., Sahakian, B. J., Evenden, J. L., Morris, R. G. and Robbins, T. W. (1989) Impaired extra-dimen- sional shift performance in medicated and unmedicated Parkinson's disease: evidence for a specific attentional dysfunc- tion. Neuropsychologia 27, 1329-1343.
Divac, I., Rosvold, H. E. and Szwarcbart, M. K. (1967) Behavioural effects of selective ablation of the caudate nucleus. J. Comp. Physiol. Psychol. 63, 184-190.
Friston, K. J., Frith, C. I)., Liddle, P. F. and Frackowiak, R. S. J. (1991) Comparing functional (PET) images: The assessment of significant change. J. Cerebr. Blood Flow Metab. 11, 690-699.
Gibb, W. R. G., Luthert, P. J., Janota, I. and Lantos, P. L. (1989) Cortical Lewy body detaentia: clinical features and classification. J. Neurol. Neurosurg. Psychiat. 52, 185-192.
Gold, J. M., Berman, K. F., Randolph, C., Goldberg, T. E. and Weinberger, D. R. (1996) PET validation of a novel prefrontal task: Delayed response alternation. Neuropsychology 10, 3-10.
Goldberg, T. E., Berman, K. F., Randolph, C., Gold, J. M. and Weinberger, D. R. (1996) Isolating the mnemonic component in spatial delayed response:: a controlled PET ~sO-labelled water re- gional cerebral blood flow study in normal humans. Neuroimage 3, 69-78.
Goldman-Rakic, P. S., Selemon, L. D. and SchwartZ, M. L. (1984) Dual pathways connecting the dorsolateral prefrontal cortex with the hippocampal formation and parahippocampal cortex in the rhesus monkey. Neuroscience 12, 719-743.
Goldman-Rakic, P. S. (1987) Circuitry of primate prefrontal cortex and the regulation of behavior by representational memory. In: Handbook of Physiology, Sec 1. The Nervous System, Vol. 5, pp. 373-417. Eds F. Plum and V. Mountcastle. American Physiological Society: Bethesda, MD.
Gotham, A. M., Brown, R. G. and Marsden, C. D. (1988) "Frontal" cognitive functions in patients with Parkinson's dis- ease "on" and "off" levodopa. Brain I l l , 299-321.
Harlow, J. M. (1868) Recovery from the passage of an iron bar through the head. Publ. Mass. Med. Soc. (_rloston) 2, 327-346.
Jones, E. G. and Powell, T. P. S. (1970) An anatt,~ical study of converging sensory pathways within the cerebral cortex of the monkey. Brain 93, 793-820.
Jonides, J., Smith, E. E., Koeppe, R. A., Awh, E., Minoshima, S. and Mintun, M. A. (1993) Spatial working memory in humans as revealed by PET. Nature 363, 623-625.
Jouandet, M. and Gazzaniga, M. S. (1979) The frontal lobes. In Handbook of Behavio~ral Neurobiology. Vol. 2. Ed. M. S. Gazzaniga, Plenum Pre~:s, New York.
Kish, S. J., Shannak, K. and Hornykiewicz, O. (1988) Uneven pat- terns of dopamine loss in the striatum of patients with idiopathic Parkinson's disease. Pathophysiologic and clinical implications. New EngL J. Med. 318, 876-880.
Klosowska, D. (1976) Relation between ability to program actions and location of brain damage. Polish Psychol. Bull. 7, 245-255.
Kuypers, 1t. G. J. M., Szwarcbart, M. K., Mishkin, M. and Rosvold, H. E. (1965) Occipito-temporal corticocortical connec- tions in the rhesus monkey. Exp. Neurol. 11, 245-262.
Lange, K. W., Robbins, T. W., Marsden, C. D., James, M., Owen, A. M. and Paul, G. M. (1992) L-DOPA withdrawal in Parkinson's disease selectively impairs cognitive performance in tests sensitive to frontal lobe dysfunction. Psychopharmacology 107, 394--404.
Lawrence, A. D., Sahakian, B. J., Hodges, J. R., Rosser, A. E., Lange, K. W. and Robbins, T. W. (1996) Executive and mnemo- nic functions in early Huntington's disease. Brain 119, 1633- 1645.
Lees, A. J. and Smith, E. (1983) Cognitive deficits in the early stages of Parkinson's di,;ease. Brain 106, 257-270.
McCarthy, G., Blumire, A. M., Puce, A., Nobre, A. C., B1och, G., Hyder, F., Goldman-Rakic, P. and Shulman, R. G. (1994) Functional magnetic resonance imaging of human prefrontal cortex activation during a spatial working memory task. Proc. Natl. Acad. Sci. U.S.A. 91, 8690-8694.
Middleton, F. A. and Strick, P. L. (1994) Anatomical evidence for cerebellar and basal ganglia involvement in higher cognitive function. Science 266, 458-461.
Middleton, F. A. and Strick, P. L. (1995) Anatomical basis for basal ganglia involvement in working memory. Soc. Neurosci. Abstr. 272.1, 1995.
Milner, B. (1971) Interhemispberic differences and psychological processes. Br. Med. Bull. 27, 272-277.
Milner, B. (1974) In: The Neurosciences: Third Study Program, pp. 75-89. Eds F. O. Schmitt and F. G. Worden. MIT Press: Cambridge, MA.
Morris, R. G., Dowries, J. J., Evenden, J. L., Sahakian, B. J., Heald, A. and Robbins, T. W. (1988) Planning and spatial work- ing memory in Parkinson's disease. J. Neurol. Neurosurg. Psychiat. 51, 757-766.
Morris, R. G., Downes, J. J., Robbins and T. W. (1990) The nature of the dysexecutive syndrome in Parkinson's disease. In: Lines of Thinking, pp. 247-258. Eds K. J. Gilhooly, M. T. G. Kean¢, R. H. Logic and G. Erdos. Wiley and Sons Ltd: Cbichester.
Morris, R. G., Ahmed, S., Syed, G. M. and Toone, B. K. (1993) Neural correlates of planning ability: frontal lobe activation during the Tower of London test. Neuropsychologia 31, 1367- 1378.
Nanta, W. J. H. (1964) Some efferent connections of the prefrontal cortex in the monkey. In: The Frontal Granular Cortex and Behavior, pp. 397-409. Eds J. M. Warren and K. Akert. McGraw-Hill: New York.
Owen, A. M. (1997)The functional organization of working mem- ory processes within human lateral frontal cortex: The contri- bution of functional neuroimaging. European Journal of Neuroscience 9, 1329-1339.
Owen, A. M., Downes, J. D., Sahakian, B. J., Polkey, C. E. and Robbins, T. W. (1990) Planning and spatial working memory following frontal lobe lesions in man. Neuropsychologia 28, 1021-1034.
Owen, A. M., James, M., Leigh, P. N., Summers, B. A., Quinn, N. P., Marsden, C. D. and Robbins, T. W. (1992) Fronto-striatal cognitive deficits at different stages of Parkinson's disease. Brain 115, 1727-1751.
Owen, A. M., Roberts, A. C., Hodges, J. R., Summers, B. A., Polkey, C. E. and Robbins, T. W. (1993a) Contrasting mechan- isms of impaired attentional set-shifting in patients with frontal lobe damage or Parkinson's disease. Brain 116, 1159-1179.
Owen, A. M., Beksinska, M., James, M., Leigh, P. N., Summers, B. A., Marsden, C. D., Quinn, N. P., Sahakian, B. J. and Robbins, T. W. (1993b) Visuo-spatial memory deficits at differ- ent stages of Parkinson's disease. Neuropsychologia 31(7), 627- 644.
Owen, A. M. and Robbins, T. W. (1993) Comparative Neuropsychology of Parkinsonian Syndromes. In: Mental Dysfunction in Parkinson's Disease, pp. 221-242. Eds E. Ch. Wolters and P. Scheltens. Vrije Universiteit: Amsterdam.
Owen, A. M., Sahakian, B. J., Hodges, J. R., Summers, B. A., Polkey, C. E. and Robbins, T. W. (1995a) Dopamine-dependeat fronto-striatal planning deficits in early Parkinson's disease. Neuropsychology 9, 126-140.
Owen, A. M., Sahakian, B. J., Semple, J., Polkey, C. E. and Robbins, T. W. (1995b) Visuo-spatial short term recognition memory and learning after temporal lobe excisions, frontal lobe excisions or amygdalo-hippocampectomy in man. Neuropsychologia 33, 1-24.
Owen, A. M., Doyon, J., Petrides, M. and Evans, A. C. (1996a) Planning and spatial working memory examined with positron emission tomography (PET). Eur. J. Neurosci. 8, 353-364.
Owen, A. M., Evans, A. C. and Petrides, M. (1996b) Evidence for a two-stage model of spatial working memory processing within the lateral frontal cortex: a positron emission tomography study. Cerebral Cortex 6, 31-38.
Owen, A. M., Morris, R. G., Sabakian, B. J., Polkey, C. E. and Robbins, T. W. (1996c) Double dissociations of memory and executive functions in working memory tasks following frontal lobe excisions, temporal lobe excisions or amygdalo-hippocam- pectomy in man. Brain 119, 1597-1615.
Owen, A. M., Doyon, J., Dagher, A. and Evans, A. C. (1996d) Abnormal Basal Ganglia Outflow in Parkinson's Disease Identified with PET: Implications for Higher Cortical Functions.

450 A M. Owen
Society For Neuroscience 26th Annual Meeting, Washington D.C., U.S.A~
Pandya, D. N. and Barnes, C. L. (1987) Architecture and connec- tions of the frontal lobe. Ill: The Frontal Lobes Revisitedr Edr E Perecman. The IRBN Press: New York.
Penfield, W. and Evans, J. (1935) The frontal lobe in man: a clini- cal study of maximum removals. Brain 58, 115-133.
Petrides, M. and Milner, B. (1982) Deficits on subject-ordered tasks after frontal and temporal lobe lesions in man. Neuropsychologia 20, 249-262.
Petrides, M. (1989) Frontal lobes and memory. In: Handbook of Neuropsychology, Vol. 9, pp. 59-82. Eds F. Boller and J. Grafman. Elsevier: Amsterdam.
Petrides, M., Alivisatos, B., Evans, A. C. and Meyer, E. (1993a) Dissociation of human mid-dorsolateral from posterior dorsolat- eral frontal cortex in memory processing. Proc. Natl. Acad. Sci. U.S.A. 90, 873-877.
Petrides, M., Alivisatos, B., Evans, A. C. and Meyer, E. (1993b) Functional activation of the human frontal cortex during the performance of verbal working memory tasks. Proc. Natl. Acad. Sci. U.S.A. 90, 878-882.
Petrides, M. (1994) Frontal lobes and working memory: evidence from investigations of the effects of cortical excisions in nonhu- man primates. In Handbook of Neuropsvchology, Vol. 9, pp. 59 82. Eds F. Boiler and J. Grafman. Elsevier: Amsterdam.
Petrides, M. and Pandya, D. N. (1994) Comparative architectonic analysis of the human and the macaque frontal cortex. In: Handbook of Neuropsychology, Vol. 9, pp. 17--58. Eds F. Boiler and J. Grafman. Elsevier: Amsterdam.
Porteus, S. D. and Kepner, R. D. (1944) Mental changes after bi- lateral prefrontal lobotomy. Genetic Psychology Monographs 29, 3-115.
Rezai, K., Andreasen, N. C., Allinger, R., Cohen, G., Swayze, V. and O'Leary, D. S. (1993) The neuropsychology of the pre- frontal cortex. Archs Neurol. 50, 636-642.
Robbins, T. W., James, M., Lange, K., Owen, A. M., Quinn, N. and M arsden, C. D. (1992) Cognitive performance in patients with Multiple System Atrophy. Brain 115, 271-291.
Robbins, T. W., James, M., Owen, A. M., Lange, K. W., Lees, A J., Leigh, P. N., Marsden, C. D., Quinn, N, P. and Summers, B. A. (1994) Cognitive deficits in Progressive Supranuclear Palsy, Parkinson's disease and Multiple System Atrophy in tests sensi- tive to frontal lobe dysfunction../. Neurol. Neurosurg. Psychiat. 57, 79--88.
Robbins, T, W., James, M., Owen, A. M., Sahakian, B. J.~ Mclnnes, L. and Rabbitt, P., A study of performance on tests from the CANTAB battery sensitive to frontal lobe dysfunction in a large sample of normal volunteers: implications for theories of executive function and cognitive ageing. (In press).
Rogers, D. (1986) Bradyphrenia in Parkinson's disease: A historical review. Psychol. Med. 16, 257 265.
Rogers, R. D., Baker, S. C., Owen, A. M., Frith, C. D., Dolan, R. J., Frackowiak, R. S. J. and Robbins, T. W. (1994) Frontal and Parietal Activation in a Test of Planning: A PET Study of the Tower of London Task. Society For Neuroscience 24th Annual Meeting, Miami, U.S.A.
Sagar, H. J., Sullivan, E. V., Gabrieli, J. D. E., Corkin, S. and Growdon, J. H. (1988) Temporal ordering and short-term mem- ory deficits in Parkinson's disease. Brain 111, 525-539.
Saint-Cyr. J. A., Taylor, A. E. and Lange, A. E. (1988l Procedural learning and neostriatat dysfunction in man. Brain 111,941-959.
Scatton, B., Javoy-agid, F., Rouqier, L., Dubois, B. and Agid, Y. (1983) Reduction of cortical dopamine, noradrenalin, seroto- nin and their metabolites in Parkinson's disease. Brain Res. 275, 321-328.
Schwartz, M. L. and Goldman-Rakic, P. S. (1984) Callosal and intrahemispheric connectivity of the prefrontal association cortex in rhesus monkey: relation between intraparietal and principal sulcal cortex. J. Comp. Neurol. 226, 403 420.
Shallice, T. (1982) Specific impairments of planning. Phil. Trans R. Soe. Lond. B 298, 199-209.
Shallice, T. (1988) From Neuropsychology to Mental Structure. Cambridge University Press, Cambridge.
Smith, E. E., Jonides, J. J., Koeppe, R. A., Awh, E., Schumacher, E. H. and Minoshima, S. (1995) Spatial versus object working memory: PET investigations. J. Cogn. Neurosci. 7(3), 337-356.
Smith, E. E., Jonides, J. J. and Koeppe, R. A. (1996) Dissociating verbal and spatial working memory using PET. Cerebral Cortex 6, 11-20.
Stuss, D. T. and Benson, D. F. (1984) Neuropsyehological studies of the frontal lobes. Psychol. Bull. 95, 3-28.
Sweeney, J. A., Minutun, M. A., Kwee, S., Wiseman, M. B., Brown, D. L., Rosenberg, D. R. and Carl, J. R. (1996) Positron emission tomography study of voluntary saccadic eye move- ments and spatial working memory. J. NeurophysioL 75, 454- 468.
Talairach, J. and Tournoux, P. (1988) Co-planar Stereotactic Atlas of the Human Brain: 3-Dimensional Proportional System: An Approach to Cerebral Imaging. Georg Thieme Verlag, Stuttgart, New York.
Taylor, A. E., Saint-Cyr, J. A. and Lang, A. E. (1986) Frontal lobe dysfunction in Parkinson's disease. Brain 109, 845-883.
Ungerleider, L. G., Gaffan, D. and Pelak, V. S. (1989) Projections from inferior temporal cortex to prefrontal cortex via the unicate fascicle in rhesus monkeys. Exp. Brain Res. 76, 473-484.
Watanabe-Sawaguchi, K., Kubota, K. and Arikuni, T. (1991) Cytoarchitecture and intrafrontal connections of the frontal cor- tex of the brain of the hamadryas baboon (Papio hamadryas). J. Comp. Neurol. 311, 108-133.
Watts, F. N., Macleod, A. K. and Morris, L. (1988) Associations between phenomenal objective aspects of concentration pro- blems in depressed patients. Br. J. Psychol. 79, 241-250.
Worsley, K. J., Evans, A. C., Marrett, S. and Neelin, P. (1993) Detecting and estimating the regions of activation in CBF acti- vation studies in human brain. In: Qualification of Brain Function: Tracer Kinetics and Image Analysis in Brain PET, pp. 535 548. Eds K. Uemura, N. Lassen, T. Jones and I. Kanno. Excerpta Medica: London.
Yeterian, E. H. and Pandya, D. N. (1991) Prefrontostriatal connec- tions in relation to cortical architectonic organisation in rhesus monkeys. J. Comp. Neurol 312, 43-67.






![Neuropsychological and neuroanatomical phenotype in 17 ... · formed in cystinosis patients [11, 22–24]. Cerebral atrophy, small cerebellum with decreased cerebellar cellularity,](https://img.pdfslide.net/doc/110x75/60fa32792c7a313aa472986a/neuropsychological-and-neuroanatomical-phenotype-in-17-formed-in-cystinosis.jpg)






















