Embed Size (px)
Citation preview

COMO AGEM OS INSETICIDAS
NOS INSETOS Dr. Ricardo Soares Matias
Em geral considera-se que o mecanismo de ação dos inseticidas atua sobre os
insetos do mesmo modo como atua sobre os animais superiores.
Os insetos possuem um organismo menos complicado que os mamíferos; não
possuem pulmões, fígado e sistema cardiovascular; possuem apenas um tipo de
nervo bioquímico, enquanto os mamíferos possuem três. Ainda conseguem uma
alta resistência a anoxia (falta de oxigênio no cérebro).
A classificação dos inseticidas de acordo com os sintomas produzidos nos insetos
é difícil de ser obtida. A natureza do inseto, o modo de penetração e a quantidade
de inseticida consumido pelo organismo, altera bastante a resposta da ação
inseticida.
De um modo geral pode se estabelecer que os inseticidas orgânicos sintéticos
atuam sobre o sistema nervoso do inseto e os sintomas apresentados obedecem
a quatro estágios: excitação, convulsão, paralisia e morte.

A maioria dos inseticidas atuam sobre o sistema nervoso. Tal fato deve-se a uma
sensibilidade especial, pois mesmo com o rompimento temporário do sistema
nervoso, os danos são irreversíveis.
A célula nervosa é denominada Neurônio e apresenta um filamento longo,
Axônio,através do qual o impulso nervoso é transmitido de neurônio a neurônio
até atingir um músculo, glândula ou a pele.
O ponto de contato entre neurônios é a sinapse. Para que o impulso nervoso seja
transmitido e determine uma reação é necessário que esse impulso passe através
de várias sinapses.
Nestas sinapses há uma determinada substância química que permite esta
transmissão, chamadas de neurotransmissor como a acetilcolina.
Podemos dizer que a transmissão do impulso nervoso passa por dois processos
diferentes. Um elétrico que é através do axônio e um químico através da sinapse.
A transmissão química do impulso nervoso se dá na sinapse que é o ponto de
encontro de um neurônio emissor e um neurônio receptor que tem estes nomes
por causa do direção do impulso nervoso.
Até este ponto a transmissão se dá por impulso elétrico. A partir deste momento,
na sinapse, a fim de passar de um neurônio para outro, entram em atividades
neuroreceptores químicos que permitem o deslocamento do impulso nervoso de
um neurônio para outro.
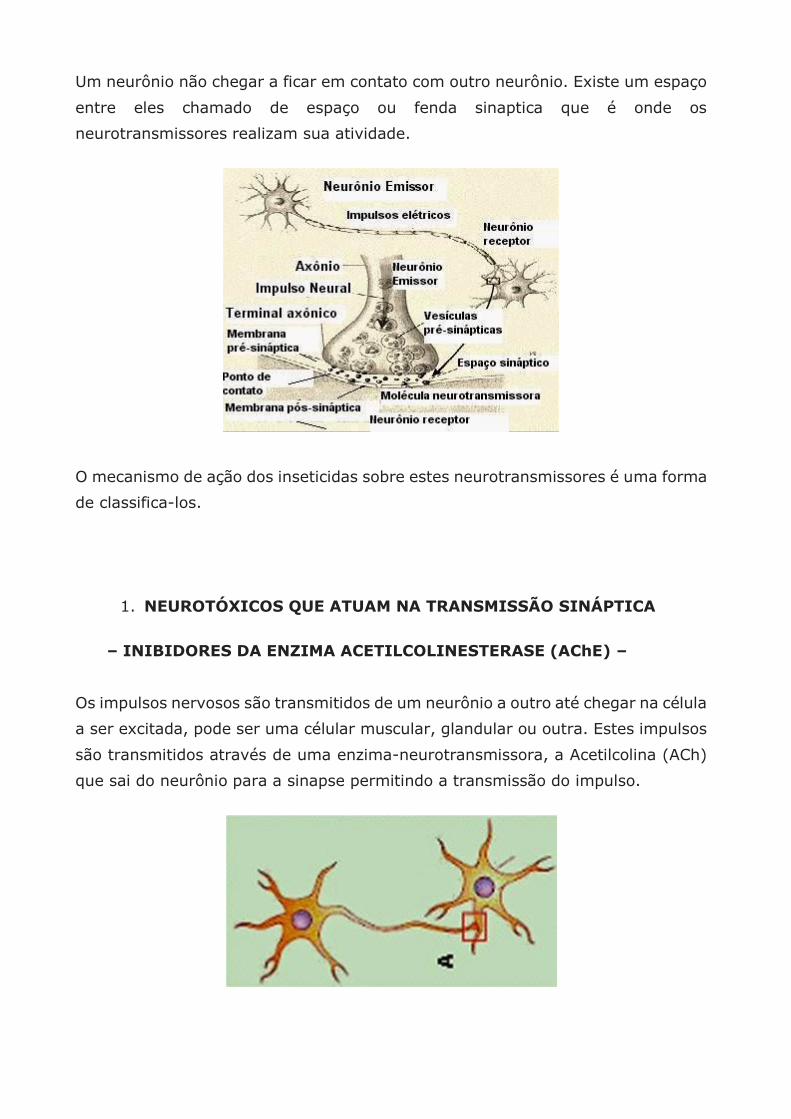
Um neurônio não chegar a ficar em contato com outro neurônio. Existe um espaço
entre eles chamado de espaço ou fenda sinaptica que é onde os
neurotransmissores realizam sua atividade.
O mecanismo de ação dos inseticidas sobre estes neurotransmissores é uma forma
de classifica-los.
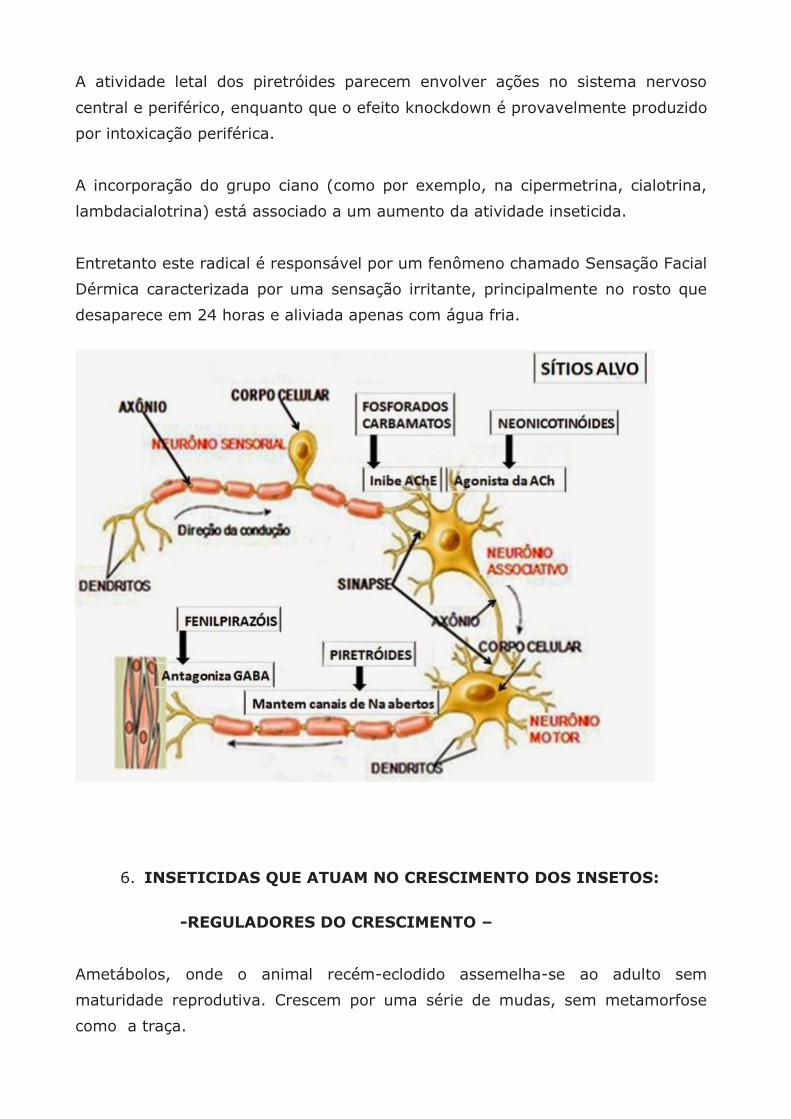
1. NEUROTÓXICOS QUE ATUAM NA TRANSMISSÃO SINÁPTICA
– INIBIDORES DA ENZIMA ACETILCOLINESTERASE (AChE) –
Os impulsos nervosos são transmitidos de um neurônio a outro até chegar na célula
a ser excitada, pode ser uma célular muscular, glandular ou outra. Estes impulsos
são transmitidos através de uma enzima-neurotransmissora, a Acetilcolina (ACh)
que sai do neurônio para a sinapse permitindo a transmissão do impulso.
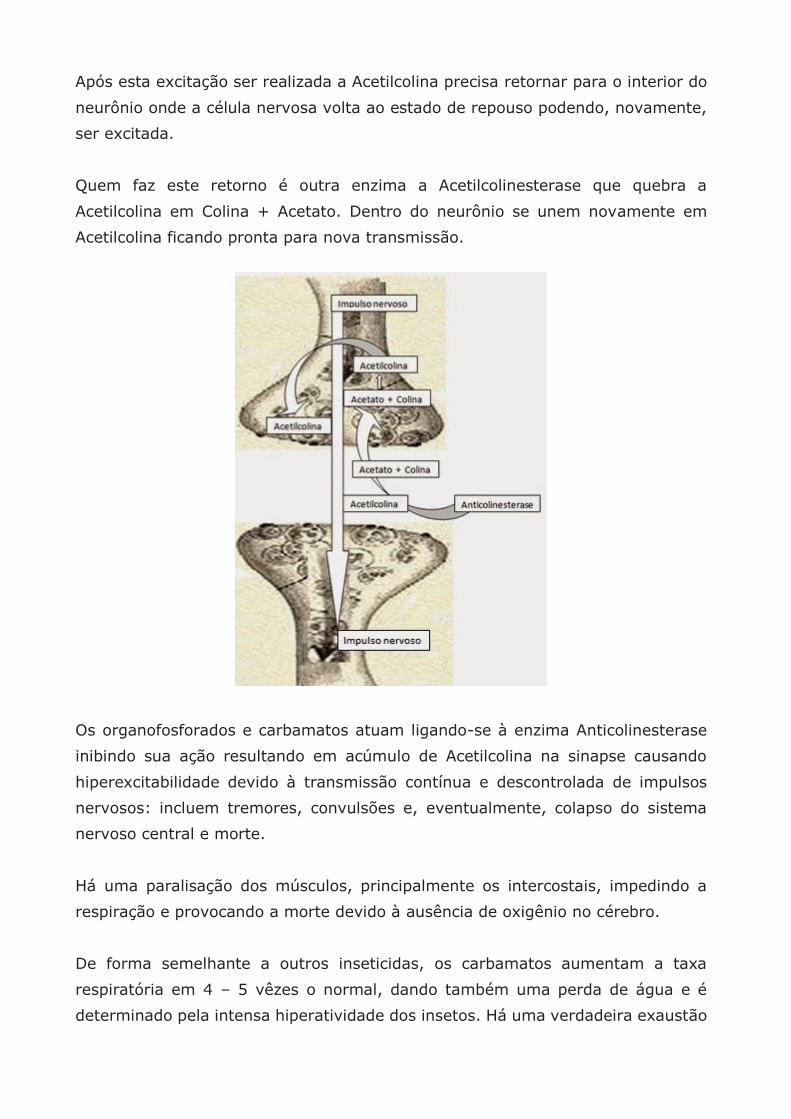
Após esta excitação ser realizada a Acetilcolina precisa retornar para o interior do
neurônio onde a célula nervosa volta ao estado de repouso podendo, novamente,
ser excitada.
Quem faz este retorno é outra enzima a Acetilcolinesterase que quebra a
Acetilcolina em Colina + Acetato. Dentro do neurônio se unem novamente em
Acetilcolina ficando pronta para nova transmissão.
Os organofosforados e carbamatos atuam ligando-se à enzima Anticolinesterase
inibindo sua ação resultando em acúmulo de Acetilcolina na sinapse causando
hiperexcitabilidade devido à transmissão contínua e descontrolada de impulsos
nervosos: incluem tremores, convulsões e, eventualmente, colapso do sistema
nervoso central e morte.
Há uma paralisação dos músculos, principalmente os intercostais, impedindo a
respiração e provocando a morte devido à ausência de oxigênio no cérebro.
De forma semelhante a outros inseticidas, os carbamatos aumentam a taxa
respiratória em 4 – 5 vêzes o normal, dando também uma perda de água e é
determinado pela intensa hiperatividade dos insetos. Há uma verdadeira exaustão

do organismo sendo primeiramente consumidos os carbohidratos e posteriormente
as gorduras
Nos organofosforados, a ligação é bem mais forte e praticamente irreversível
enquanto que nos carbamatos ela é reversível, podendo haver uma recuperação
do inseto.
Esta reversibilidade pode ocorrer pela capacidade que os insetos têm de
permanecer um certo tempo em anóxia, período em que a reversão do efeito
inseticida pode vir a ocorrer pela detoxificação rápida.
2. INSETICIDAS QUE ATUAM NOS RECEPTORES DE ACETILCOLINA
– AGONISTAS (ANÁLOGOS) DA ACETILCOLINA (ACh)-
É o grupo dos neonicotinóides descobertos a partir da molécula de nicotina
(Imidaclopride, Acetamipride, Nitempiram, Tiametoxam, Clotianidina,
Dinotefurano e Tiaclopride). Os neonicotinóides imitam o efeito da Acetilcolina e
competem com ela. Esta ligação é persistente pois os neonicotinóides são
insensíveis à ação da acetilcolinesterase (AChE). A acetilcolinesterase degrada
moléculas de acetilcolina mas não consegue degradar as moléculas de
neonicotinóides.
Desta forma mantem ativação permanente da acetilcolina
causando hiperexcitabilidade do sistema nervoso central devido à transmissão
contínua e descontrolada de impulsos nervosos. Embora atuem de modo
totalmente distinto dos organofosforados e carbamatos, os sintomas resultantes
da intoxicação são semelhantes e incluem tremores, convulsões e, eventualmente,
colapso do sistema nervoso central e morte.

3. INSETICIDAS QUE ATUAM NOS RECEPTORES GABA
ANTAGONISTAS DE CANAIS DE CLORO MEDIADOS POR GABA
(FENILPIRAZÓIS EX. FIPRONIL)
Sabe-se que antagonizam a ação do neurotransmissor inibitório, GABA (ácido
gama amino butírico) impedindo que após a transmissão de um impulso nervoso,
se desencadeie o processo de inibição restabelecendo o repouso do SNC.
Em uma situação normal o GABA permite um aumento na permeabilidade da
membrana aos íons cloro para dentro da célula nervosa, o que desencadeia o
mecanismo de retorno ao repouso do sistema nervoso após ser excitado.
Os fenilpirazóis afetam este mecanismo, impedindo a entrada dos íons Cl– no
neurônio e assim antagonizando (= revertendo) o efeito “calmante” de GABA.
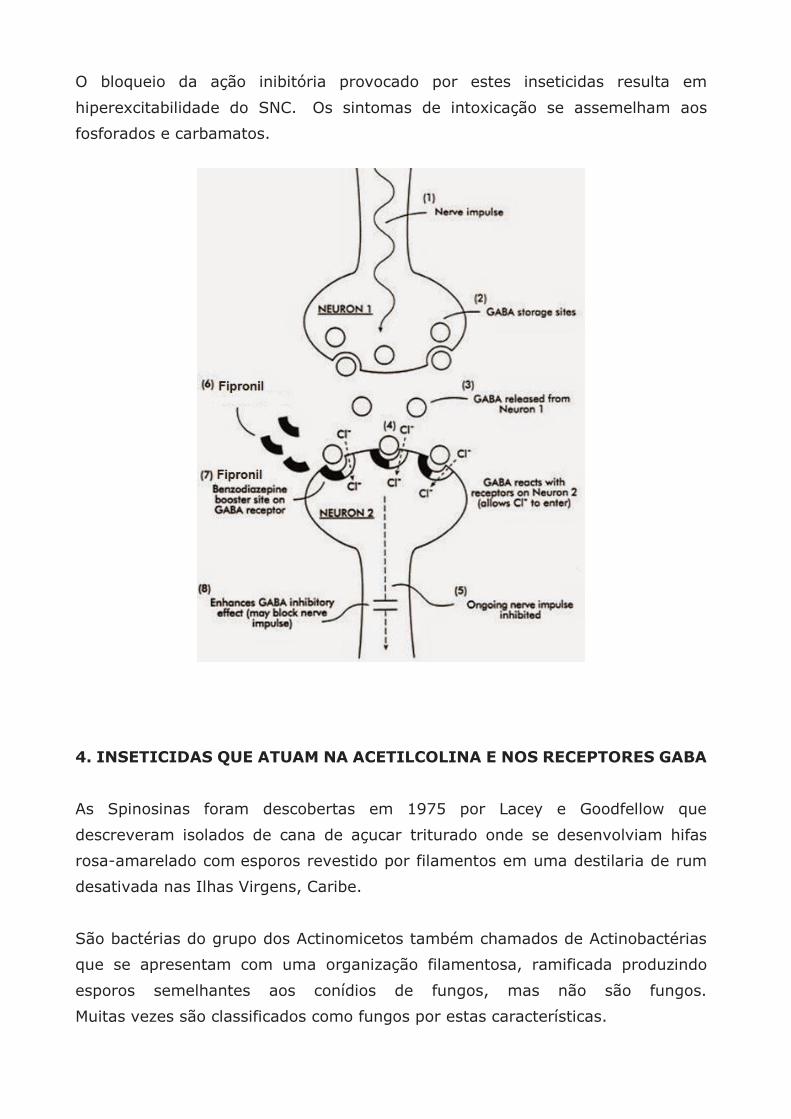
O bloqueio da ação inibitória provocado por estes inseticidas resulta em
hiperexcitabilidade do SNC. Os sintomas de intoxicação se assemelham aos
fosforados e carbamatos.
4. INSETICIDAS QUE ATUAM NA ACETILCOLINA E NOS RECEPTORES GABA
As Spinosinas foram descobertas em 1975 por Lacey e Goodfellow que
descreveram isolados de cana de açucar triturado onde se desenvolviam hifas
rosa-amarelado com esporos revestido por filamentos em uma destilaria de rum
desativada nas Ilhas Virgens, Caribe.
São bactérias do grupo dos Actinomicetos também chamados de Actinobactérias
que se apresentam com uma organização filamentosa, ramificada produzindo
esporos semelhantes aos conídios de fungos, mas não são fungos.
Muitas vezes são classificados como fungos por estas características.
Spinosad é um inseticida baseado na fermentação da bactéria Saccharopolyspora
spinosa. Têm um novo modo de ação baseado na segmentação das ligações nos
receptores de acetilcolina diferente das atividades dos outros inseticidas, pois
causam rompimento da neurotransmissão de acetilcolina.
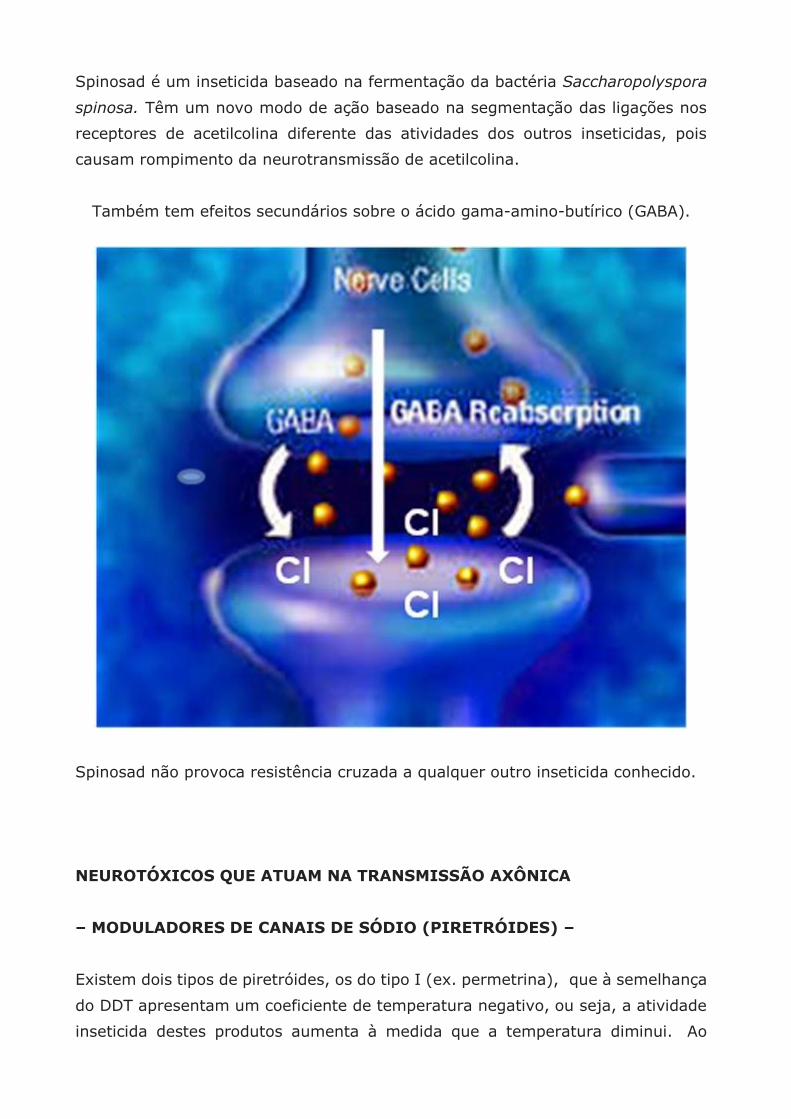
Também tem efeitos secundários sobre o ácido gama-amino-butírico (GABA).
Spinosad não provoca resistência cruzada a qualquer outro inseticida conhecido.
NEUROTÓXICOS QUE ATUAM NA TRANSMISSÃO AXÔNICA
– MODULADORES DE CANAIS DE SÓDIO (PIRETRÓIDES) –
Existem dois tipos de piretróides, os do tipo I (ex. permetrina), que à semelhança
do DDT apresentam um coeficiente de temperatura negativo, ou seja, a atividade
inseticida destes produtos aumenta à medida que a temperatura diminui. Ao

contrário, os piretróides do tipo II (ex. Cipermetrina, deltametrina, etc.) tem um
coeficiente de temperatura positivo.
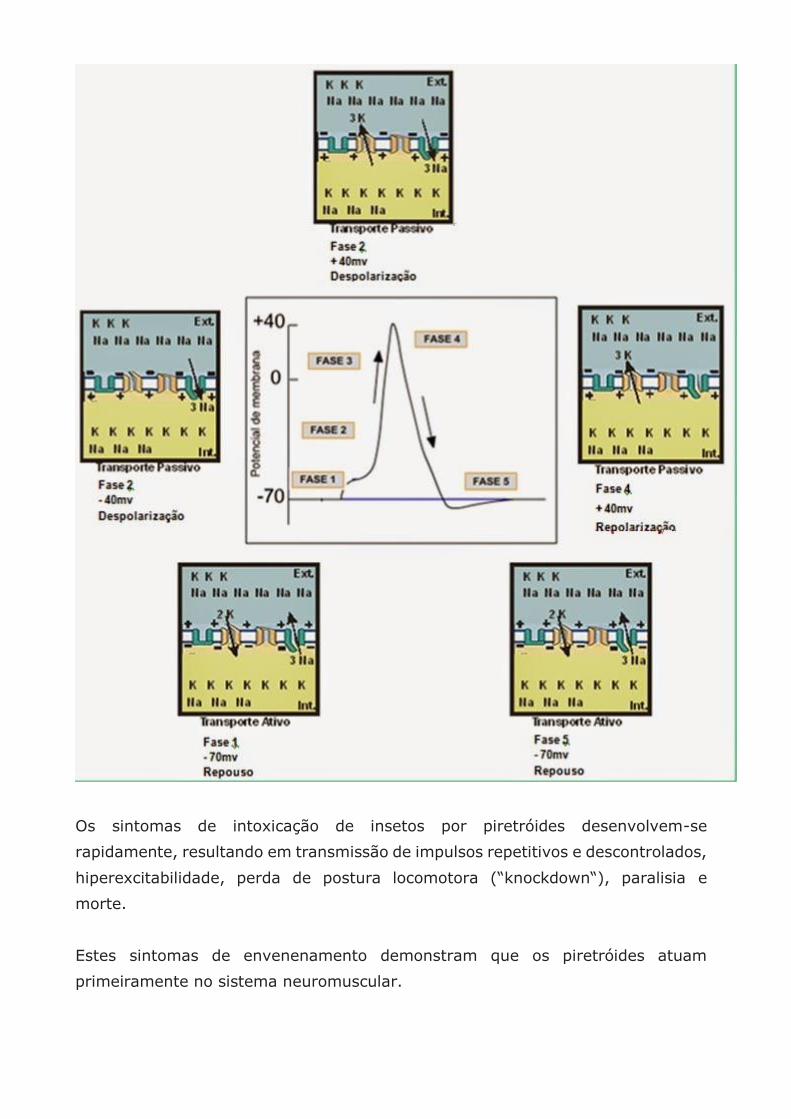
A célula nervosa é controlada eletricamente e sua polarização e despolarização
está na dependência da entrada e saída de Na e K. Quando a célula é excitada este
impulso nervoso se transmite pelas alterações, dentro e fora do axônio, das cargas
negativas e positivas.
Os piretróides (tipo I e II) mantem uma entrada permanente de Na na célula
deixando-a sempre excitada, não entrando em repouso sendo mais intenso no
tipo II.
BLOQUEADORES DE CANAIS DE SÓDIO (oxadiazinas)
As oxadiazinas representam um novo grupo, cujo primeiro e único representante
comercial atualmente é o indoxacarb. O indoxacarb é considerado um pró-
inseticida pois precisa ser bioativado por enzimas específicas no trato
gastrointestinal do inseto para gerar o metabólito ativo com potente atividade.
EVENTOS AXÔNICOS NA CONDUÇÃO DO ESTÍMULO EXCITATÓRIO
A membrana do axônio é permeável a K+ quando em repouso e impermeável a
Na+, portanto a membrana do axônio permanece polarizada no estado de repouso
com potencial próximo ao potencial de equilíbrio do K+ (-50 a -70 mV).
Mediante estímulo, os canais de K da membrana se fecham e os de Na se abrem
permitindo um fluxo de Na para o interior da célula despolarizando-a até atingir
um potencial próximo ao potencial de equilíbrio do Na.
Quando esse “potencial de ação” é atingido no desencadeamento de um estímulo,
os canais de Na se fecham novamente e os K vão lentamente se abrindo até ser
restabelecido o potencial elétrico de repouso (membrana polarizada).
A bomba de Na-K transporta o excesso de Na do interior para o exterior do axônio,
e K do exterior para o interior do mesmo restabelecendo o equilíbrio químico da
célula no estado de repouso.

O impulso transmitido se propaga ao longo do axônio até atingir a extremidade do
mesmo, onde a transmissão passa de elétrica a química.
EVENTOS SINÁPTICOS NA CONDUÇÃO DO ESTÍMULO EXCITATÓRIO
Sinapse é a fenda que separa duas células nervosas intercomunicantes.
A transmissão do impulso nervoso deve atravessá-la para que se propague, o que
é conseguido através da liberação de neurotransmissores na membrana pré-
sináptica.
Esses neurotransmissores migram pela fenda sináptica e atingem receptores
específicos na membrana pós-sináptica onde desencadeiam novos potenciais de
ação.
Canais de Ca++ na membrana pré-sináptica modulam a liberação das vesículas
com neurotransmissor na sinapse.
Principais neurotransmissores em insetos, de importância para o controle químico
deles:
. Acetilcolina: neurotransmissor excitatório, presente no sistema nervoso central
de insetos.
. Ácido gama aminobutírico (GABA): neurotransmissor inibitório, presente no
sistema
nervoso central de insetos e junções neuromusculares.
ESTÍMULO INIBITÓRIO:
Estímulo inibitório leva a uma hiperpolarização da membrana do axônio através
do fluxo de íons Cl- para o interior da membrana do axônio via canais de Cl.
Efeito inibitório bloqueia o excitatório e o neurotransmissor envolvido em sinápses
inibitórias é o GABA.

Os neurotransmissores se difundem pela fenda Sináptica ligando-se em receptores
específicos na Membrana Pós-Sináptica abrindo os Canais permitindo a entrada e
saída de íons Na e K causando uma despolarização neste ponto que vai se
deslocando por todo o neurônio até o próximo Botão Terminal, transmitindo assim,
a mensagem através de novas ondas elétricas.
Os íons Na e K estão se deslocando de um local mais concentrado para um menos
concentrado. Neste instante ocorre uma Despolarização da membrana pela maior
entrada de Na e maior saída de K. Com isto o interior fica (+) e o exterior fica (-).

Inicialmente as forças que geram a entrada de Na para a célula são maiores que
as forças que tendem a liberar K para o exterior da célula e posteriormente vão
mudando conforme o Potencial de Membrana vai aumentando. Os canais de Na se
abrem permitindo a entrada dele (-40mv). Logo depois os canais de K se abrem
(-10mv). Quando chega em + 40mv os canais de Na se fecham e inicia o processo
de Repolarização. Continua saindo K da célula e retorna o Potencial de Repouso
até os canais de K se fecharem e voltar a -70mv.
Os sintomas de intoxicação de insetos por piretróides desenvolvem-se
rapidamente, resultando em transmissão de impulsos repetitivos e descontrolados,
hiperexcitabilidade, perda de postura locomotora (“knockdown“), paralisia e
morte.
Estes sintomas de envenenamento demonstram que os piretróides atuam
primeiramente no sistema neuromuscular.
A atividade letal dos piretróides parecem envolver ações no sistema nervoso
central e periférico, enquanto que o efeito knockdown é provavelmente produzido
por intoxicação periférica.
A incorporação do grupo ciano (como por exemplo, na cipermetrina, cialotrina,
lambdacialotrina) está associado a um aumento da atividade inseticida.
Entretanto este radical é responsável por um fenômeno chamado Sensação Facial
Dérmica caracterizada por uma sensação irritante, principalmente no rosto que
desaparece em 24 horas e aliviada apenas com água fria.
6. INSETICIDAS QUE ATUAM NO CRESCIMENTO DOS INSETOS:
-REGULADORES DO CRESCIMENTO –
Ametábolos, onde o animal recém-eclodido assemelha-se ao adulto sem
maturidade reprodutiva. Crescem por uma série de mudas, sem metamorfose
como a traça.
Os Hemimetábolos (metamorfose incompleta) a as ninfas vivem na água e têm
morfologia diferente (estruturas bastante distintas) dos adultos (e.g. brânquias,
aparelho bucal). Há exemplos nas ordens de insetos aquáticos (Odonata,
Ephemeroptera e Plecoptera)..
Paurometábolos (metamorfose gradual): as ninfas e os adultos não diferem quanto
a forma e estilo de vida, mas há mudanças graduais e inconspícuas no tamanho,
desenvolvimento dos ocelos e, finalmente, formação de asas e genitália. Há
exemplos em Orthoptera, Hemiptera, Homoptera.
Holometábolos (metamorfose completa): os jovens diferem radicalmente dos
adultos quanto a forma geral, alimentação e tipo de vida. Há vários estágios: ovo,
larva, pupa e adulto (imago). Há exemplos em Lepidoptera, Strepsiptera,
Coleoptera, Hymenoptera, Diptera etc.
Um fato interessante é que os mesmos hormônios regulam tanto a metamorfose
completa como a incompleta.

ENDOCRINOLOGIA DA METAMORFOSE:
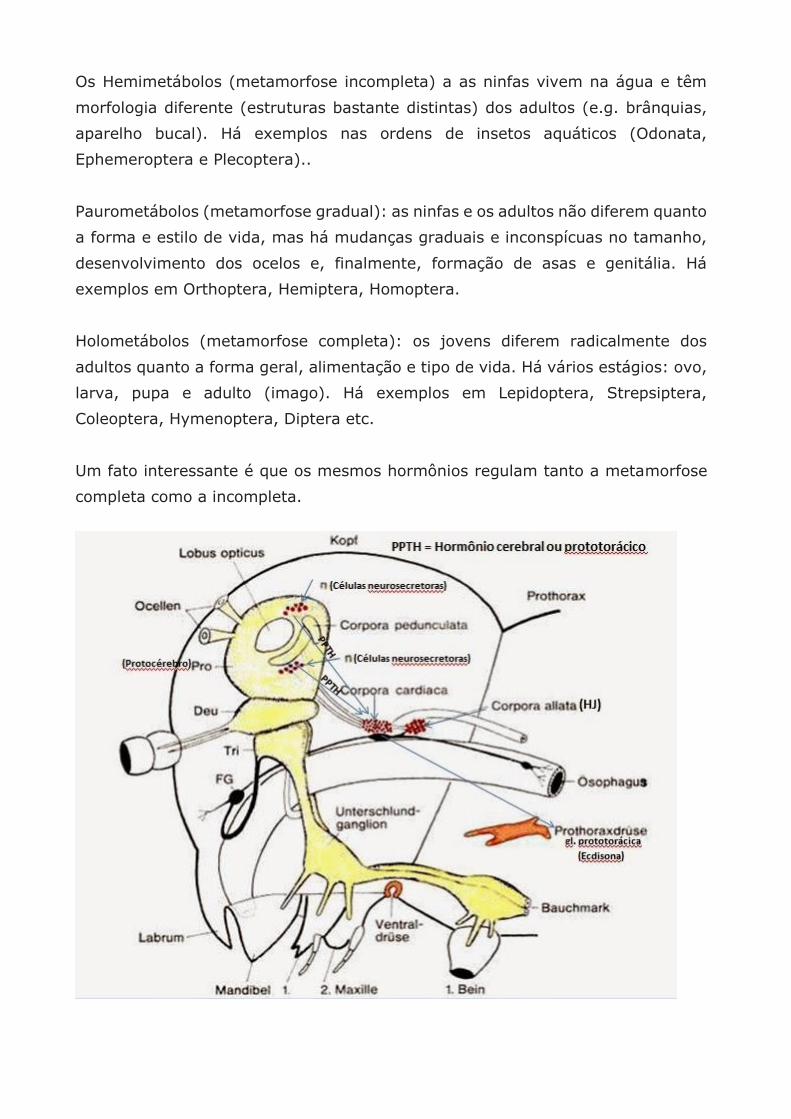
As mudanças que ocorrem da fase imatura à adulta dos insetos, são controladas
por hormônios secretados pelo sistema neuroendócrino do PROTOCÉREBRO que
liberam um hormônio denominado hormônio do cérebro, que transmite
neuromensagens às glândulas endócrinas e a outros tecidos.
O hormônio cerebral ou hormônio PROTOTORÁXICO (PPTH) secretado pelas
células neurosecretoras do protocérebro é transportado pelos microtúbulos dos
axônios até a CORPORA CARDIACA.
Aí é estocado e posteriormente liberado na hemolinfa estimulando a glândula
prototoráxica, localizada no tórax do inseto, a sintetizar e liberar a ECDISONA
(=ECDISTERONA) ou hormônio do crescimento e da muda causando neles uma
diferenciação durante o processo da muda.
Outro importante hormônio no controle muda/ metamorfose é o hormônio juvenil
(HJ) cujo estímulo para a sua síntese varia de espécie para espécie, às vezes em
função de fatores ambientais. Algumas espécies são influenciadas por fatores
sazonais como temperatura e fotoperíodo.
O JH é produzido na CORPORA ALLATA (logo atrás do cérebro) e age sobre a
epiderme determinando que a nova cutícula mantenha características juvenis. Em
todos os estágios em que o HJ está presente ocorre apenas o crescimento do
animal.
Quando, no último estágio, o nível do hormônio juvenil diminui acentuadamente,
o animal muda, sofre metamorfose tornando-se adulto (hemimetábolo) ou passa
para a fase de pupa (holometábolo). Este hormônio volta a ser secretado durante
o período de reprodução.
Durante a fase de desenvolvimento larval o HJ está em níveis altos
e ECDISONA baixo, para manter as características juvenis. Em determinado
momento o nível de HJ começa a diminuir e de ecdisona aumenta inibindo a ação
do HJ e está no momento de mudar de instar.
No último instar larval é onde ocorrem as mudanças mais críticas – pupa e adulto.
Na fase de pupa o HJ vai diminuindo até desaparecer por completo do sangue do

inseto, momento de atingir a fase de imago (Holometábolo). Nos hemimetábolos
ele desaparece entre a fase de última ninfa e adulto.
Durante a fase de reprodução o HJ volta a ser produzido.
A forma adulta dos insetos recebe o nome de imago. Chama-se larva a forma
jovem muito diferente do imago. A ninfa é a forma jovem dos insetos
hemimetábolos (um pouco parecida com imago). Pupa é a forma intermediária
entre larva e o imago nos holometábolos (do grego holo, ‘todo’, ‘tudo’, e metabole,
‘mudança’).
A muda, nos insetos, é desencadeada pelo hormônio ecdisona, cuja produção é
estimulada pelos hormônios cerebrais.
Existe, entretanto, um outro hormônio — hormônio juvenil — que impede a
transformação da larva em pupa, ou desta em imago.
O esteróide ecdisona é um hormônio de desenvolvimento encontrado nos
artrópodes.
O processo de muda é regulado pela variação das concentrações da ecdisona na
larva. O aumento dos níveis deste hormônio leva ao início da muda.
Quando a larva entra no estágio de muda, ela pára de se alimentar e o fluido de
muda, que contém várias enzimas proteolíticas necessárias para a mesma, entra
no espaço ecdisial.
Neste ponto, as células epidérmicas entram em uma fase de aumento na síntese
de proteínas (expressão do gene) e uma nova cutícula é produzida. Um decréscimo
na concentração de ecdisona desencadeia uma fase subseqüente, que inclui a
ativação das enzimas no fluido de muda, digestão da procutícula e reabsorção do
fluido de muda.

Na seqüência, há mais um decréscimo na concentração de ecdisona e outros
hormônios, necessários para completar o processo de muda, são liberados,
reiniciando-se, então, a alimentação.
O crescimento e o desenvolvimento adequado dos insetos dependem da
concentração de ecdisona em vários estágios de diferenciação.
Para que a metamorfose ocorra é necessário que a taxa de hormônio juvenil na
hemolinfa seja muito pequena ou nula.
Caso contrário, o animal realiza a muda, mas passa apenas de uma fase da larva
para outra fase de larva.

HORMÔNIOS JUVENIS ANÁLOGOS (HJA) OU JUVENÓIDES
(exs. Fenoxicarb, hidroprene, methoprene, pyriproxyfen)
São agonistas de HJ atuando da mesma forma, fazendo com que os insetos
permaneçam na fase larval, embora já estejam prontos para passarem à fase
adulta, impedindo-os de se reproduzir e se desenvolver levando-os à morte.
Os juvenóides podem apresentar também efeitos ovicida e esterilizante. Se um
juvenóide for aplicado numa época em que ele deveria estar ausente o resultado
é o aparecimento de um inseto com características juvenis e de adulto incapaz de
se alimentar, copular e se reproduzir, morrendo rapidamente.
Alguns dos juvenóides afetam somente determinados estágios do desenvolvimento
de poucas espécies, outros são específicos para algumas famílias enquanto outros

apresentam atividade contra uma grande variedade de espécies. Os efeitos
obtidos com um grupo de insetos freqüentemente não é aplicável para outro grupo.
Talvez a principal limitação no uso prático de Juvenóides para o controle de insetos
é a necessidade de coordenar a aplicação destes compostos, que normalmente não
são muito persistentes, com a fase sensível do ciclo vital destes insetos, a estes
compostos.
ANTAGONISTAS DE HJ
São chamados de Precocenos, grupo de compostos naturais e sintéticos que
tem ação contrária aos HJ por isto são denominados anti juvenóides ou
antagonistas dos HJ.
Causam metamorfose prematura em diversas espécies de insetos, levando à
formação de adultos estéreis fazendo com que o inseto passe para a fase adulta
precocemente. Ex: allatostatina, gonadotropina e butóxido de piperonila.
AGONISTAS DA ECDISONA (hormônio do crescimento e da muda)
Recentemente, surgiram os inseticidas do grupo das Diacilhidrazinas como o
metoxifeno e tebufenozida (lepidópteros) que atuam como agonistas de
ecdisteróides provocando aceleração no processo da ecdise (troca de pele)
resultando em insetos deformados. O efeito desta classe de inseticidas é a
interrupção do processo de muda dos insetos alvos, resultando em sua destruição.
INIBIDORES DA SÍNTESE DE QUITINA
A quitina é constituída por uma cadeia longa de N-acetilglicosamina que ocorre
naturalmente em diversos organismos, sendo o principal componente do
exosqueleto dos artrópodes atuando como componente estrutural de suporte
celular e de superfície do corpo (cutícula, epiderme, traquéia e epitélio intestinal).
O metabolismo de quitina se divide em processos de síntese e degradação
catalisadas por enzimas específicas.

As enzimas Sintases da Quitina (CS) catalizam (síntese) a formação de quitina
através da transferência de N-acetilglicosamina para o crescimento da cadeia de
quitina.
Estas enzimas estão localizadas no retículo endoplasmático das células das
membranas do Complexo de Golgi, na Membrana Plasmática e nas vesículas
intracelulares que são levadas para a superfície celular.
As enzimas responsáveis pela degradação da quitina são as Quitinases sendo
cruciais para o desenvolvimento pós-embrionário, especialmente durante a muda
larval e pupação.
O metabolismo de quitina vem sendo considerado um excelente alvo para o
controle seletivo de pragas. A inibição ou desregulação das enzimas chaves são
importantes objetos para o desenvolvimento de inseticidas.
As BENZOILFENILURÉIAS (ex. diflubenzuron, nuvaluron, teflubenzuron,
hexaflumuron, penfluron, triflumuron) são importantes inibidores da síntese de
quitina.
É outro tipo de regulador de crescimento denominado não hormonal que impedem
o transporte da N-acetilglucosamina (um açúcar produzido pelo organismo
responsável pela formação da quitina) que interfere no metabolismo de
ecdisteróides inibindo a formação do exoesqueleto quitinoso matando pela
incapacidade de gerar uma nova cutícula e o inseto não consegue se libertar do
antigo exoesqueleto.
A ausência desta cutícula ou sua formação inadequada faz com que o inseto
desseque pela perda de umidade ou entre em um processo de inanição.
Existe ainda um outro grupo de não hormonais que são
as AMINOTRIAZINAS ou TRIASINAS, afeta o metabolismo da epiderme sendo
um inibidor do processo de esclerotização (i.e. endurecimento) da cutícula.
É um potente inibidor do endurecimento da quitina sendo específico para dípteros,
tendo como exemplo a Ciromazina (ex. moscas, mosquitos).

INSETICIDAS INIBIDORES DA SÍNTESE DE ATP
AMIDINOHIDRAZONAS (ex. Hidramethilona)
A maioria dos inseticidas possui mecanismo de ação relacionado à transmissão de
impulsos nervosos. A Hidrometilona possui um mecanismo diferente de ação, mas
interferindo, indiretamente, na transmissão do impulso nervoso.
A Hidrametilona é um veneno de ação estomacal e portanto precisa ser ingerido
para que sua ação se manifeste. Desta forma atua tanto nas formas imaturas
(ninfas) como na forma adulta.
Os animais, incluindo os insetos, precisam de energia para o funcionamento de
suas funções vitais. Esta energia é produzida através de uma serie de reações
bioquímicas dentro de unidades celulares denominadas mitocôndrias na forma de
ATP.
Para a formação de ATP são necessárias reações químicas chamadas fosforilação
oxidativa que ocorre na cadeia transportadora de elétrons que libera elétrons para
a formação do ATP
.
A fosforilação oxidativa é o processo de síntese de ATP a partir da energia liberada
pelo transporte de elétrons na cadeia respiratória. Durante o fluxo de elétrons há
liberação de energia livre suficiente para a síntese de ATP.

As condições para que ocorra a fosforilação oxidativa são um bombeamento de
prótons pela cadeia respiratória, criando um fluxo da matriz mitocondrial para o
citosol (liquido que preenche o citoplasma das células).
A energia derivada do transporte de elétrons na cadeia respiratória é convertida
numa força utilizada principalmente para bombear prótons da matriz mitocondrial
para o citosol.
A membrana interna da mitocôndria é impermeável aos prótons mas estes
precisam retornar para iniciar a fosforilação oxidativa.
Por isto cria-se um gradiente duplo na membrana mitocondrial interna de pH e
eletrostático através do qual os prótons conseguem retornar para o interior da
mitocôndria através da enzima ATPase determinando síntese de mais ATP.
O não retorno de protons para o interior da mitocondria cria uma instabilidade na
célula e perda de energia quebrando a cadeia respiratória.
A maior quantidade de ATP é formada na terceira etapa da respiração que é a
cadeia respiratória onde são produzidas 32 moléculas de ATP para cada uma de
glicose
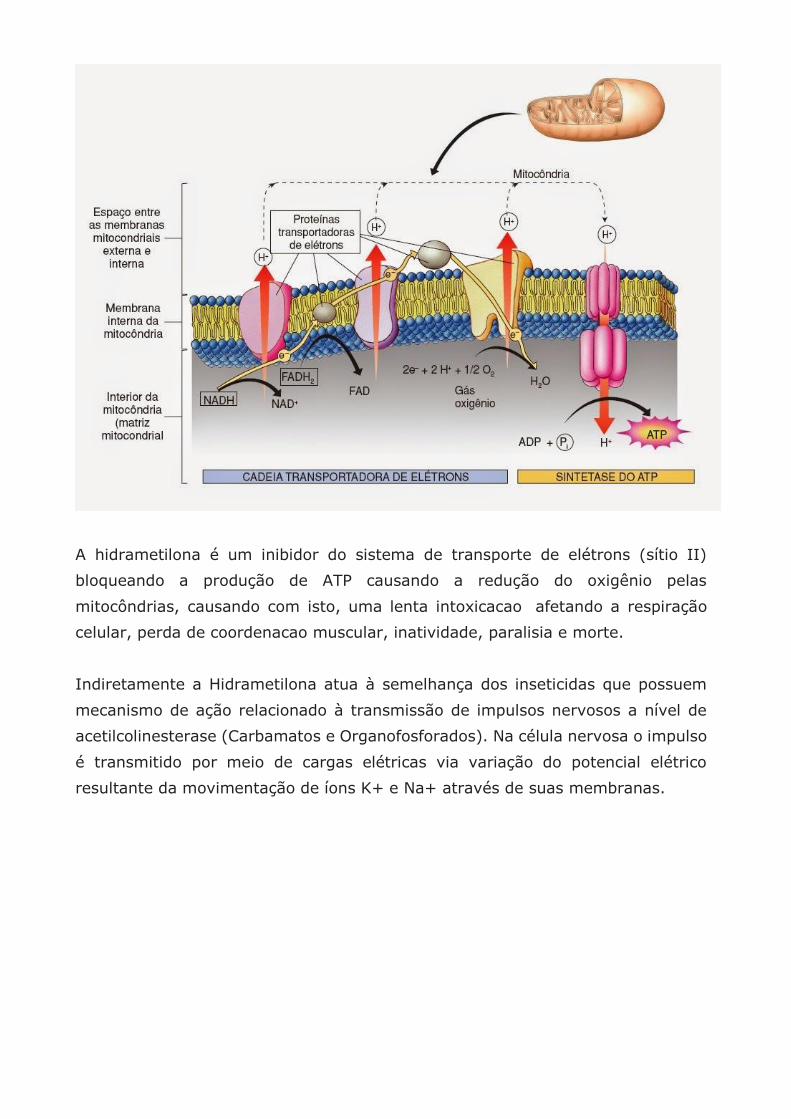
A hidrametilona é um inibidor do sistema de transporte de elétrons (sítio II)
bloqueando a produção de ATP causando a redução do oxigênio pelas
mitocôndrias, causando com isto, uma lenta intoxicacao afetando a respiração
celular, perda de coordenacao muscular, inatividade, paralisia e morte.
Indiretamente a Hidrametilona atua à semelhança dos inseticidas que possuem
mecanismo de ação relacionado à transmissão de impulsos nervosos a nível de
acetilcolinesterase (Carbamatos e Organofosforados). Na célula nervosa o impulso
é transmitido por meio de cargas elétricas via variação do potencial elétrico
resultante da movimentação de íons K+ e Na+ através de suas membranas.
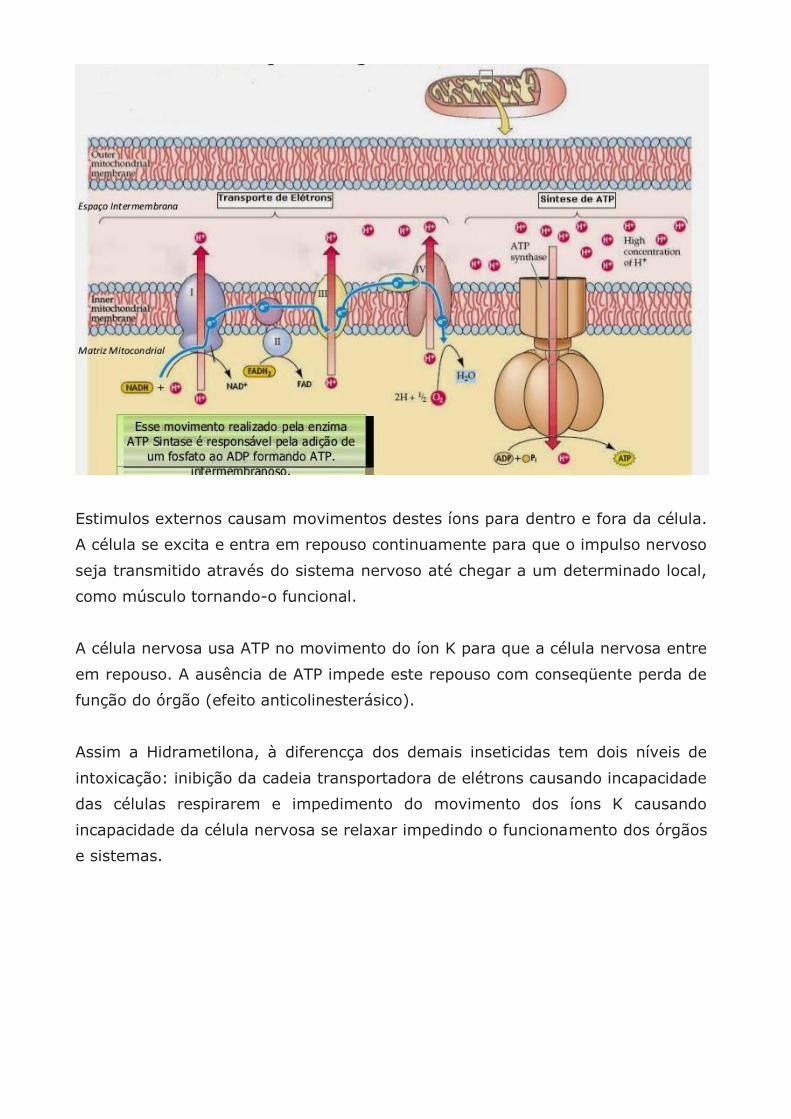
Estimulos externos causam movimentos destes íons para dentro e fora da célula.
A célula se excita e entra em repouso continuamente para que o impulso nervoso
seja transmitido através do sistema nervoso até chegar a um determinado local,
como músculo tornando-o funcional.
A célula nervosa usa ATP no movimento do íon K para que a célula nervosa entre
em repouso. A ausência de ATP impede este repouso com conseqüente perda de
função do órgão (efeito anticolinesterásico).
Assim a Hidrametilona, à diferencça dos demais inseticidas tem dois níveis de
intoxicação: inibição da cadeia transportadora de elétrons causando incapacidade
das células respirarem e impedimento do movimento dos íons K causando
incapacidade da célula nervosa se relaxar impedindo o funcionamento dos órgãos
e sistemas.

SULFLURAMIDA
A sulfluramida mata insetos pelo rompimento do fluxo normal de prótons usados
por suas células de armazenamento de energia (mitocôndrias) para criar o ATP
(Adenosina Trifosfato). A sulfluramida afeta, portanto, o processo de fosforilação
oxidativa (respiração aeróbia) agindo sobre as mitocôndrias e interrompendo a
produção de ATP.
A perda de produção de ATP é letal para qualquer ser vivo.
REGULAGEM HÍDRICA
Um exemplo é o ácido bórico, inseticida mineral, se apresentando na forma de gel
ou pó seco. Ele age a nível celular causando interrupção na regulagem hídrica
causando desidratação o que resulta em mais ingestão de isca para compensar a
perda líquida. Na forma de pó seco ele é abrasivo ao exoesqueleto.
SEMIOQUÍMICOS é o nome dado às substâncias químicas responsáveis pela
comunicação entre os seres vivos e também às toxinas liberadas por estes
organismos.
INFOQUÍMICOS se refere às substâncias químicas na interação entre organismos
provocando reações comportamentais ou fisiológicas que dependendo da resposta
podem ser benéficas ou não ao receptor e/ou emissor.

ALELOQUÍMICOS são substâncias químicas empregadas na comunicação entre
espécies diferentes (interespecíficas):
1) Alomônios são substancias químicas produzidas por uma espécie que quando
entra em contato com outra espécie diferente provoca uma reação benéfica à
espécie emissora.
Também são alomônios as substancias produzidas por diversos animais que agem
como repelentes contra seus predadores. As operárias de espécies de formigas
produzem um jato de substância repelente a animais intrusos.
Alomônios que servem para subjugar presas ocorre em alguns percevejos
cujassubstâncias liberadas ao serem consumidas por formigas ficam narcotizadas
constituindo-se em presas fáceis para o percevejo.
O besouro Myrmecaphodius excavaticollis (=Martinezia dutertrei) caminha
livremente em colônia de Solenopsis ritcheri até obter alimento das operárias,
graças à aquisição rápida de hidrocarbonetos das formigas durante seus contatos
passando a ser reconhecido como companheiro. Transferidos para outra colônia
perde este odor e adquire desta nova colônia.
Certas espécies de baratas produzem uma cetona que faz parte do sistema de
alarme de formigas para seu uso na defesa.
As aranhas boleadeiras (Mastophora spp) produzem um alomônio semelhante ao
feromônio sexual de duas espécies de Noctuidae para atrair os machos e lançam
uma bola pegajosa para prender estas mariposas.
2) Cairomônios são substancias químicas que emitidas por um organismo de uma
espécie provoca uma resposta em um organismo de outra espécie que se beneficia
deste estimulo (favorecem a espécie receptora).

Alguns parasitas e predadores de insetos muitas vezes localizam suas presas por
meio de substancias voláteis emitidas por elas.
O odor do hospedeiro é utilizado por quase todos os insetos hematófagos
(beneficiado) no processo de localizacão do hospedeiro (prejudicado).
Durante o metabolismo dos vertebrados, um grande número de substâncias
químicas voláteis (CO2, acetonas, etc.) e não voláteis (ácido lático, ácidos graxos,
etc.) é emitido durante a expiracão e da pele. Provavelmente cada inseto
hematófago se especializou em detectar estas substâncias para localizar o
hospedeiro, como ocorre em insetos ornitófílicos, antropofílicos, etc.
A resposta de mosquitos aumenta quando o hospedeiro está suando ou suado.
O CO2 é o principal cairomônio liberado pelos hospedeiros e usado em armadilhas
para capturar culicídeos, simulídeos, tabanídeos, triatomíneos, muscídeos e
califorídeos.
O ácido lático iberado em todo o corpo de vertebrados atrai mosquitos e
principalmente quando em combinacão com CO2.
Fezes de galinha ou de coelhos e extrato de racão de coelho estimulam a
oviposicão de flebotomíneos pela emanacão de voláteis.
Infusão da grama e de esterco atraem mosquitos por voláteis emanados.
Provavelmente bactérias em água contaminada eliminem voláteis semelhantes o
que atrai mosquitos.
Os voláteis da carne fresca em decomposicão atraem C. hominivorax e Chrysomia
bezziana.
Água com ácido cáprico induziram a oviposicão de A. aegypti e Culex spp da
mesma forma que culturas de Pseudomonas aeruginosa.
Levedura de Sacharomyces cerevisae em sacarose atrairam barbeiros tanto
quanto o odor de um hospedeiro (rato). A atracão foi significativamente diminuída
com a retirada da fonte de CO2 da cultura de levedo.

3) Sinomônios
Favorecem ambas espécies.
FEROMÔNIOS
São substâncias excretadas por organismos vivos e detectadas por outros
indivíduos da mesma espécie (intraespecíficos), produzindo mudanças de
comportamento específicas. Exs. Periplanona-B, feromônio sexual da
barata Periplaneta americana.
Os feromonios(do grego, pherein = levar, carregar + hormon = estimular, excitar)
tiveram esta designacão por não poderem ser chamados de hormônios uma vez
que atuam fora do organismo quer as produziu.
De um modo geral, nos insetos, são produzidos por glândulas situadas nos dois
últimos segmentos abdominais. Seu conteúdo é desprendido evertendo-as e
expondo seu conteúdo ao ar.
Uma vez que são produzidos em quantidades muito pequenas as espécies tem
que ter uma grande capacidade de detectá-las. Por exemplo espécies de mariposas
são capazes de perceber a presença da fêmea a 11 km de distância.
Feromônios são substâncias químicas secretadas por um indivíduo e que permitem
a sua comunicação com outros indivíduos da mesma espécie.
A mensagem química transmitida pelos feromônios tem por objetivo estimular
determinado comportamento, que pode ser de alarme, agregação, contribuição na
produção de alimentos, defesa, ataque, acasalamento, etc.

O termo “feromônio” pode ser usado para indicar tanto uma substância em
particular, como uma mistura de substâncias. Eles foram descobertos em 1950.
Em 1959, o pesquisador alemão Butenandt conseguiu isolar e identificar o
primeiro feromônio conhecido como bombicol ((10,12)-hexadecadien-1-ol), que é
o feromônio da mariposa do bicho-da-seda Bombyx mori. Ele precisou matar 500
mil fêmeas desse inseto para obter apenas 1 mg da substância ativa.
Os insetos são os que mais liberam esse tipo de composto químico, mas eles não
são os únicos; os mamíferos (como camundongos, preás, porcos, cães e até o ser
humano) também realizam essa comunicação olfativa.
O uso de feromônios na agricultura sempre foi uma prática regularmente usada
desde sua descoberta e assim é até hoje tanto é que na ANVISA, Agência Nacional
de Vigilância Sanitária, estão registrados 28 feromônios para o controle de insetos-
praga na agricultura. Os produtos comerciais feromonais disponíveis no mercado
atendem 28 das 49 culturas do país.
Infelizmente o uso como domissanitários não segue a mesma prioridade levando
o usuários à utilização de compostos químicos.

1)Sexual
Os feromônios mais estudados são os sexuais, excretados pela fêmea e em muitos
casos também pelo macho, utilizados para atrair o parceiro para a cópula e assim
preservar a espécie, através da procriação.
Um exemplo de atraente sexual excretado pelas fêmeas da mosca domestica é o
cis-9-tricoseno, mostrado a seguir. Seu isômero na forma trans não apresenta a
propriedade de agir como feromônio.
Alguns insetos utilizam como feromônio de acasalamento o ácido tetradec-3,5-
dienoico:
Observe que esse feromônio possui os hidrogênios dos carbonos 3 e 4 da dupla
ligação na forma trans, um em cada lado do plano; enquanto que os hidrogênios
dos carbonos 5 e 6 estão na forma cis. A troca de posição dos grupos ligados a

uma dessas insaturações corresponderia a outra substância com propriedades
diferentes e que não seria reconhecida pelo inseto.
2) Agregação
São substâncias químicas eliminadas pelos insetos com o intuito de mante-los
próximos para manutenção das sociedades de insetos (abelha), colonização de
novos habitat e agregação antes do acasalamento.
Esta agregação também ocorre com insetos que não apresentam este
comportamento social mas que realizam agregação como baratas e o cascudinho
da cama de frangos. Este é um comportamento que poderia ser utilizado na
captura através de armadilhas.
3) Dispersão
Afastar o potencial inimigo como o gambá que esguicha a partir de glândulas
circum-anais (localizadas ao redor do ânus) jatos de substâncias de odor pútrido
contendo feromônios que afastam seus inimigos.
Também servem para manutenção de umespaço mínimo parasobrevivência
(formiga) e para antiagregação(moscas-das-frutas).
4)Alarme
para sinalizar perigo e ameaça,provocando a fuga (ex.: pulgão), agressão contra
outro inseto (abelha) ou inibição de agressão(formiga). Esses feromônios têm um
raio de ação médio de 10 cm por um período médio de 10 min.
5)Território
Delimita territórios, minimizando ou até evitando encontros indesejáveis e
agressivos.

6)Trilha
Direcionam até o alimento. Avistado uma fonte de alimento as formigas liberam
os feromônios para que todas sigam uma mesma trilha sem se dispersarem até o
alimento, seria como um feromônio de orientação.

INSETICIDAS BOTÂNICOS
Na primeira metade do século 20, o Brasil foi um grande produtor e exportador
de inseticidas botânicos, como piretro, rotenona e nicotina, que apresentam maior
segurança em seu uso agrícola e de menor impacto ambiental.
Os inseticidas botânicos foram muitos populares e importantes entre as décadas
de 30 e 40 para o controle de pragas. A importação de materiais botânicos ou
derivados para uso como inseticidas representou um empreendimento comercial
considerável.
Como sempre acontece, com raras exceções novos inseticidas, sejam, naturais ou
sintéticos sempre iniciam sua aplicação na área agrícola e depois é que se
direcionam para a saúde pública. Com os inseticidas botânicos não foi diferente.
Entretanto alguns como o óleo de nim, óleo de citronela, d-limoneno, linalol e o
quassim já tem sido usado no combate aos sinantrópicos.
Atualmente pouco se tem ofertas de inseticidas botânicos para ser usado nos
programas de gerenciamento. Isto tudo mostra que ainda temos muito a criar no
que se refere a inseticidas não orgânicos. Por exemplo, acima de 6.700 toneladas
de raízes de Derris elliptica foram importadas para os Estados Unidos do sudeste
da Ásia em 1947, mas diminuiu para 1.500 toneladas em 1963.
Isto reflete a extensão pela qual os inseticidas botânicos foram substituídos pelos
inseticidas químicos sintéticos, também conhecidos como agrotóxicos, dentre eles,

os organoclorados, organofosforados, carbamatos e piretróides, produzidos pelos
países industrializados nas décadas de 50 e 60.
O volume de agrotóxicos usados no mundo chegou a ultrapassar 20.000 toneladas
de ingredientes ativos na década de 90, quando as importações de piretro para
os Estados Unidos totalizaram apenas umas 350 toneladas. Hoje, os inseticidas
botânicos constituem apenas 1% do mercado mundial de inseticidas.
Afortunadamente, o interesse em desenvolver e usar produtos botânicos para o
manejo de pragas está novamente aumentando nos últimos anos, estimando-se
um crescimento anual na ordem de 10 a 15%.
Inseticidas botânicos podem causar a morte do inseto por intoxicação, mas, às
vezes, são repelentes ou fagodeterrentes. Sua ação tóxica se faz no sistema
nervoso central dos insetos interferindo na transmissão sináptica (à semelhança
dos organofosforados e carbamatos) ou axônica (à semelhança dos piretródes) do
impulso nervoso. Da mesma forma que é tóxica para os insetos também é para os
seres humanos. Outros atuam na muda, na síntese de ATP, análogos dos
reguladores de crescimento, maturação de ovos.
Alguns exemplos são os óleos de citros extraídos da casca de laranjas e outras
frutas cítricas e refinados para originar os compostos d-limoneno e linalol atuando
sobre pulga, piolhos, ácaros e carrapatos e mosca doméstica.
O extrato de alho é usado na agricultura uma vez que seu efeito se dá por
repelência. Pela sua ação sistêmica o extrato é absorvido pelas plantas e seu
sistema radicular mudando o odor natural da planta enganando os insetos praga.
A azadirachtina atua no sistema hormonal dos insetos.
Outras substâncias que têm sido empregadas são, por exemplo, os terpenos
presentes nos óleos extraídos da resina de pinheiro, o nerol extraído do óleo
essencial do capim limão, e algumas substâncias obtidas de plantas utilizadas
como condimento alimentar, como o eugenol do cravo da índia, o mentol da
hortelã, a piperina da pimenta-do-reino.
Algumas plantas produzem substâncias aromáticas que tem apresentado
bioatividade como o mentrasto que é repelente a insetos ou agem como

reguladores de crescimento e a erva-de-santa-maria ou mastruz cujos frutos tem
compostos bioativos contra insetos de grãos armazenados.
Óleo da citronela com repelente a insetos, rotenona extraída do timbó atua na
mitocôndria impedindo a produção de ATP, o quassim extraído da planta chamada
no nordeste de amarga pelo seu gosto extremamente amargo, chegando a ser dito
que ele é 50 vezes mais amargo que o quinino, sendo ativo contra larvas do
pernilongo atuando na inibição do desenvolvimento da cutícula.
A cevadina e a veratridina são compostos derivados das sementes maduras da
sabadila que é um lírio perene cujo mecanismo de ação imita os piretróides por
atuarem no impedimento do impulso nervoso através do axônio. À semelhança dos
piretróides é um dos inseticidas botânicos menos tóxicos para mamíferos sendo
irritante para a pele e mucosas dos humanos.
Os preparados inseticidas a base de rianodina são derivados dos talos e raízes
de Ryania speciosa Vahl (Flacourtiaceae). A rianodina é mais estável do que a
piretrina erotenona e, portanto, tem atividade residual mais longa, proporcionando
até duas semanas de controle após a aplicação, mas é mais seletivo.
A rianodina tem um modo de ação bastante particular, por ligar-se
irreversivelmente os canais de cálcio das fibras musculares, que permanecem
abertos, deixando os íons Ca++ livres para inundar o interior das fibras, induzindo
a contração dos músculos esqueléticos e causando paralisia muscular. A
alimentação, o movimento e a reprodução cessam gradualmente após contato com
essa substância.